PEMANFAATAN
Chromolaena odorata
SEBAGAI PAKAN
TERNAK POTENSIAL DENGAN BERBAGAI MACAM
METODE PENGOLAHAN
YELLY MAGDALENA MULIK
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Pemanfaatan
Chromolaena odorata sebagai Pakan Ternak Potensial dengan berbagai Macam Metode Pengolahan adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2016
Yelly Magdalena Mulik
RINGKASAN
YELLY MAGDALENA MULIK. Pemanfaatan Chromolaena odorata sebagai Pakan Ternak Potensial dengan berbagai Macam Metode Pengolahan. Dibimbing oleh MUHAMMAD RIDLA, IWAN PRIHANTORO dan MARTHEN LUTHER MULLIK.
Chromolaena odorata merupakan salah satu gulma invasif yang keberadaannya dapat menurunkan kualitas dan kuantitas padang penggembalaan. Kendati demikian, gulma ini memiliki potensi sebagai pakan ternak karena memiliki kandungan protein yang berkisar 18-36%. Terbatasnya pemanfaatan C. odorata sebagai pakan ternak karena adanya kandungan metabolit sekunder sebagai faktor pembatas seperti tanin dan trypsin inhibitor. Keberadaan metabolit sekunder tanin dalam pakan dapat menurunkan palatabilitas, konsumsi, kecernaan dan juga penyerapan nutrien. Bahkan dalam jumlah tertentu dapat menimbulkan kematian bagi ternak yang mengkonsumsinya. Trypsin inhibitor dalam pakan juga dapat menurunkan daya cerna protein. Kendala tersebut dapat diatasi dengan proses pengolahan baik secara fisik, kimia, maupun biologis. Penelitian ini terdiri atas dua tahapan, tahap pertama bertujuan untuk mengkaji kualitas nutrisi, kecernaan dan kandungan metabolit sekunder C. odorata dengan pengolahan fisik, kimia dan biologis.
Hasil penelitian menunjukkan bahwa perlakuan biologis (fermentasi) mampu mempertahankan kualitas nutrisi, kecernaan serta menurunkan kandungan tanin dan trypsin inhibitor C. odorata. Tahap kedua bertujuan untuk mengevaluasi pengaruh penambahan (tepung putak sebagai sumber karbohidrat) dan isi rumen (sebagai sumber enzim) pada level berbeda terhadap kualitas nutrisi dan fermentatif serta kandungan tanin dan trypsin inhibitor silase C. odorata. Hasil penelitian menunjukkan bahwa perlakuan silase dengan penambahan tepung putak
10% dan isi rumen 10% memiliki kualitas nutrisi dan fermentatif terbaik serta mampu menurunkan tanin dan trypsin inhibitor pada silase C. odorata.
SUMMARY
YELLY MAGDALENA MULIK. Utilization Chromolaena odorata as a Potential feedstuff using various Processing Method. Supervised by MUHAMMAD RIDLA, IWAN PRIHANTORO and MARTHEN LUTHER MULLIK
Chromolaena odorata is one of the invasive weeds on native pastures in in dryland areas. Its presence can reduce quality and quantity of edable forages on rangelands. Nevertheless, this weed has potential for livestock feed because it has a protein content ranging from 18 to 36%. The limited use of C. odorata as fodder for its content of secondary metabolites as limiting factors such as tannins and
trypsin inhibitors. The presence of secondary metabolite compounds such as tannins and anti-trypsin in feed materials can reduce palatability, intake, digestibility and nutrient absorption. Even with a certain amount can cause death of animals that ingest them. These obstacles can be solved by using physical, chemical, and biological processing methods of the feed materials. This study consisted of two stages. The first stage aimed at assessing nutrient composition, digestibility and content of secondary metabolites of C. odorata which was subjected to physical, chemical and biological processing.
The results showed that biological treatment (fermentation) in C. odorata
was able to maintain nutrient quality, digestibility and lower the concentration of tannin and trypsin inhibitor. The second stage was designed to evaluate the effect of adding various levels of Corypha (putak) powder and rumen content on nutrient fermentation, nutritional quality and concentration of tannins and trypsin inhibitors in C. odorata. The results showed the best treatment was the addition of 10% putak and 10% rumen contentasit can maintain nutrient quality and reduces concentration of tannins and trypsin inhibitors in C. odorata silage.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Nutrisi dan Pakan
PEMANFAATAN
Chromolaena odorata
SEBAGAI PAKAN
TERNAK POTENSIAL DENGAN BERBAGAI MACAM
METODE PENGOLAHAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
PRAKATA
Puji dan syukur penulis panjatkan ke hadirat Tuhan Yang Maha Kuasa karena atas hikmat-Nya sehingga karya ilmiah ini dapat diselesaikan. Penelitian ini dilaksanakan sejak Tahun 2014 dengan judul Pemanfaatan Chromolaena odorata sebagai Pakan Ternak Potensial dengan berbagai Macam Metode Pengolahan. Bagian dari tesis ini telah diterbitkan pada Jurnal Ilmu Ternak dan Veteriner (JITV) volume 21(1) Tahun 2016 dengan judul “Anaerobic fermentation effectively reduces concentration of total tannins in siam weed (Chromolaena odorata)”.
Terima kasih penulis ucapkan kepada kemenristekdikti yang telah memberikan beasiswa calon dosen kepada penulis. Terima kasih yang tulus penulis sampaikan kepada Bapak Dr Ir Muhammad Ridla, MAgr, Bapak Dr Iwan Prihantoro, SPt MSi dan Bapak Ir Marthen Luther Mullik, PGDipAgrSt Ph.D selaku komisi pembimbing yang telah banyak memberikan saran, waktu dan pikiran serta dengan sabar dan iklas telah membimbing penulis selama mengikuti pendidikan magister. Penulis juga mengucapkan terima kasih kepada Dr Anuraga Jayanegara, SPt MSc selaku dosen penguji pada ujian sidang tesis. Ucapan terima kasih pun penulis haturkan kepada Ketua program studi INP Prof Dr Ir Yuli Retnani, MSc dan sekretaris program studi INP Dr Ir Lilis Khotijah, MS atas masukan terhadap penulisan karya ilmiah ini dan selama proses pendidikan. Terimakasih juga penulis ucapkan kepada staf program studi INP (Mas Supri dan Bu Ade) atas bantuan administrasinya, Ibu Dian Anggraeny, Pak Sofyan dan tim lab PBMT atas bantuan dan kebaikan hati menerima penulis sebagai anggota Laboratorium Nutrisi Ternak Perah, dan Laboratorium Ilmu dan Teknologi Pakan. Teman-teman Pasca-INP 2013 dan 2012, teristimewa Ujie, Rikardo, Mbak Novi, Mas Hilmi, Fina, dan teman-teman yang tidak disebutkan namanya terima kasih atas motivasi dan semangat yang diberikan selama penulis mengikuti pendidikan magister. Terima kasih juga kepada K Willy, kru warung bambu, dan kru GAMANUSRATIM atas motivasi, dan dukungan serta seluruh pihak yang telah berkontribusi bagi penulis. Ungkapan terimakasih yang sebesar-besarnya penulis haturkan kepada orang tua (Papa dan Mama, Daddy dan Mama), oma, adik-adik tersayang serta seluruh keluarga besar atas doa, dukungan dan motivasinya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juni 2016
DAFTAR ISI
DAFTAR TABEL vii
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN vii
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Hipotesis 2
Manfaat Penelitian 3
2 UJI EFEKTIVITAS BERBAGAI MACAM PENGOLAHAN TERHADAP KONSENTRASI TOTAL TANIN, TRYPSIN INHIBITOR DAN KECERNAAN PAKAN Chromolaena odorata SECARA In vitro 3
ABSTRAK 3
ABSTRACT 4
PENDAHULUAN 4
METODE 5
Waktu dan Tempat Penelitian 5
Bahan dan Persiapannya 6
Perlakuan dan Rancangan Percobaan 6
Peubah yang Diamati 7
HASIL DAN PEMBAHASAN 8
Kandungan Metabolit Sekunder 8
Komposisi Nutrient C. odorata setelah perlakuan 11
Kualitas Fermentasi Rumen secara invitro 13
Nilai Kecernaan InvitroC. odorata 14
SIMPULAN 16
3 KUALITAS SILASE C. odorata DENGAN DAN TANPA PENAMBAHAN
ISI RUMEN SAPI 16
ABSTRAK 16
ABSTRACT 17
PENDAHULUAN 18
METODE 19
Waktu dan Tempat Penelitian 19
Materi 19
Pembuatan Tepung Putak 20
Pembuatan Silase 20
Pemanenan Silase 20
Peubah yang Diamati 20
HASIL DAN PEMBAHASAN 21
Kualitas Fisik Silase 21
Kandungan Nutrisi Silase C. odorata 23
Kandungan Metabolit Sekunder 25
Kualitas Fermentasi Rumen secara invitro 26
Nilai Kecernaan In vitro Silase C. odorata 27
SIMPULAN 29
4 PEMBAHASAN UMUM 29
5 SIMPULAN UMUM 33
DAFTAR PUSTAKA 34
LAMPIRAN 40
DAFTAR TABEL
1 Komposisi kimia daun Chromolaena odorata 6
2 Kandungan metabolit sekunder C. odorata akibat berbagai perlakuan 8 3 Kandungan nutrien C. odorata akibat berbagai perlakuan 12 4 Kualitas fermentatif invitroC. odorata pada berbagai perlakuan 13 5 Nilai kecernaan in vitroC. odorata pada berbagai perlakuan 15
6 Kualitas fisik silase C. odorata 22
7 Suhu dan pH silase C. odorata 23
8 Komposisi nutrien C. odorata sebelum dan sesudah ensilase 24 9 Kandungan metabolit sekunder silase C. odorata 25
10 Kualitas fermentasi rumen silase C. odorata 27
11 Nilai kecernaan in vitro silase C. odorata 28
12 Matriks korelasi komposisi kimia C. odorata dan kecernaan in vitro 30 13 Matriks korelasi komposisi kimia dan kecernaan in vitro silase C. odorata 32
DAFTAR GAMBAR
1 Warna silase C. odorata 21
2 Chromolaena odorata 57
3 Proses pembuatan tepung putak (Coryphaelata robx) 57
4 Tahapan pembuatan silase Chromolaena odorata 57
DAFTAR LAMPIRAN
1 Kuisioner skoring kualitas fisik silase 41
9 Uji lanjut duncan BO daun C. odorata akibat berbagai perlakuan 44 10 Hasil sidik ragam PK daun C. odorata akibat berbagai perlakuan 44 11 Uji lanjut duncan PK daun C. odorata akibat berbagai perlakuan 44 12 Hasil sidik ragam LK daun C. odorata akibat berbagai perlakuan 44 13 Uji lanjut duncan LK daun C. odorata akibat berbagai perlakuan 45 14 Hasil sidik ragam SK daun C. odorata akibat berbagai perlakuan 45 15 Uji lanjut duncan SK daun C. odorata akibat berbagai perlakuan 45 16 Hasil sidik ragam BETN daun C. odorata akibat berbagai perlakuan 45 17 Uji lanjut duncan BETN daun C. odorata akibat berbagai perlakuan 46 18 Hasil sidik ragam NH3 daun C. odorata akibat berbagai perlakuan 46
19 Hasil sidik ragam VFA daun C. odorata akibat berbagai perlakuan 46 20 Hasil sidik ragam KCBK daun C. odorata akibat berbagai perlakuan 46 21 Uji lanjut duncan KCBK daun C. odorata akibat berbagai perlakuan 47 22 Hasil sidik ragam KCBO daun C. odorata akibat berbagai perlakuan 47 23 Uji lanjut duncan KCBO daun C. odorata akibat berbagai perlakuan 47 24 Hasil sidik ragam keberadaan jamur silase C. odorata 47
25 Hasil sidik ragam suhu silase C. odorata 48
26 Uji lanjut interaksi perlakuan PA dan hari terhadap suhu silase C.
odorata 48
27 Hasil sidik ragam pH silase C. odorata 48
28 Uji lanjut interaksi perlakuan PA dan hari terhadap pH silase C. odorata 49
29 Hasil sidik ragam BKsilase C. odorata 49
30 Uji lanjut duncan perlakuan PA terhadap BKsilase C. odorata 49
31 Hasil sidik ragam BOsilase C. odorata 49
32 Uji lanjut duncan perlakuan PA terhadap BO silase C. odorata 50
33 Hasil sidik ragam PKsilase C. odorata 50
34 Uji lanjut interaksi perlakuan PA dan hari terhadap PK silase C. odorata 50
35 Hasil sidik ragam LKsilase C. odorata 50
36 Uji lanjut interaksi perlakuan PA dan hari terhadap LK silase C. odorata 51
37 Hasil sidik ragam SKsilase C. odorata 51
38 Uji lanjut interaksi perlakuan PA dan hari terhadap SK silase C. odorata 51 39 Hasil sidik ragam BETNsilase C. odorata 52 40 Uji lanjut perlakuan PA terhadap BETN silase C. odorata 52
42 Uji lanjut Duncan perlakuan PA terhadap tanin silase C. odorata 52 43 Hasil sidik ragam trypsin inhibitorsilase C. odorata 53 44 Uji lanjut interaksi perlakuan PA dan hari terhadap trypsin inhibitor
silase C. odorata 53
45 Hasil sidik ragam NH3silase C. odorata 53
46 Uji lanjut perlakuan PA terhadap NH3 silase C. odorata 54
47 Hasil sidik ragam VFA silase C. odorata 54
48 Uji lanjut perlakuan PA terhadap VFA silase C. odorata 54 49 Hasil sidik ragam pH rumensilase C. odorata 54 50 Uji lanjut interaksi perlakuan PA dan hari terhadap pH rumen silase C.
odorata 55
51 Hasil sidik ragam KCBKsilase C. odorata 55
52 Uji lanjut interaksi perlakuan PA dan hari terhadap KCBK silase C.
odorata 55
53 Hasil sidik ragam KCBO silase C. odorata 56
54 Uji lanjut interaksi perlakuan PA dan hari terhadap KCBO silase C.
odorata 56
55 Hasil sidik ragam KCPKsilase C. odorata 56
56 Uji lanjut interaksi perlakuan PA dan hari terhadap KCPK silase C.
1
PENDAHULUAN
Latar Belakang
Chromolaena odorata merupakan jenis semak perdu berkayu tahunan yang dianggap sebagai salah satu jenis gulma yang paling invasif di dunia (Hawaii Department of Agriculture 2016; United States Department of Agriculture 2016). Hal ini dikarenakan tanaman C. odorata memiliki sifat pertumbuhan yang sangat cepat sehingga dalam waktu singkat dapat menutupi area tempat tumbuhnya. Tanaman C. odorata memiliki 2 sisi yang berbeda. Di satu sisi, C. odorata
merupakan gulma yang sangat invasif karena dapat menjadi competitor tanaman lain, dapat menurunkan kualitas padang penggembalaan khususnya terkait dengan kapasitas tampung dan keragaman tanaman yang tumbuh di padang penggembalaan serta memiliki kandungan metabolit sekunder yang dapat bersifat anti nutrisi. Di sisi lain C. odorata memiliki berbagai potensi yang bermanfaat misalnya dapat digunakan sebagai pakan ternak karena kandungan proteinnya yang tinggi yaitu sekitar 18-36% (Basha et al. 2012; Hai et al. 2013; Mullik 2002). Beberapa kajian melaporkan bahwa C.odorata dapat dimanfaatkan sebagai pakan domba hingga level 20% dari total bahan kering, kelinci hingga 30%, broiler finisher hingga level 3% (Apori et al. 2001; Bamikole et al. 2004; Bonsu
et al. 2013). Rangkaian penelitian Hai et al. (2012; 2013; 2014) melaporkan bahwa anak kambing yang induknya diberi tepung daun C. odorata 50 gram selama masa bunting memiliki preferensi makan dan konsumsi terhadap tepung C. odorata yang lebih baik dibanding anak kambing yang induknya tidak diberi tepung C. odorata. Mullik et al.(2014)dan Bira et al. (2015) melaporkan bahwa
C.odorata bentuk pellet dan mash dapat digunakan sebagai pakan ternak sapi umur 2 tahun hingga level 40%. Bila pemanfaatannya lebih dari 40% dikuatirkan akan memberikan dampak negatif bagi ternak, sebab pada level pemberian sebesar 40% menyebabkan semua parameter konsumsi, daya cerna dan fermentasi rumen mulai memperlihatkan tren menurun meskipun belum nyata.
Metabolit sekunder yang terkandung dalam C. odorata antara lain tanin,
trypsin inhibitor, saponin, alkaloid, steroid, terpenoids, dan flavonoid (Akinmoladun et al. 2010; Onkaramurthy et al. 2013). Keberadaan metabolit sekunder selain sebagai faktor pembatas, kuatnya bau dan rasa yang dimiliki oleh
C. odorata pun dapat menurunkan palatabilitas (Hai et al. 2012). Tanin dan
trypsin inhibitor merupakan metabolit sekunder yang paling banyak jumlahnya yaitu masing-masing 0.55% dan 22.37 mg g-1 (Ikhimioya et al. 2007). Perbedaan konsentrasi tanin dikarenakan perbedaan metode analisis, perbedaan ekotipe, musim, zona ekologi dan umur tanaman (Mueller-Harvey 2006).
2
terhadap tanin asal bahan pakan yang berbeda pun bervariasi. Perbedaan toleransi tanin tersebut tergantung pada sumber, jenis, level dan sifat kimia dari masing-masing bahan pakan (Makkar dan singh 1991; Mueller-Harvey 2006; Patra dan Saxena 2011). Trypsin inhibitor merupakan senyawa yang memiliki kemampuan menghambat aktivitas enzim proteolitik karena pembentukan ikatan kompleks antara enzim proteolitik dan senyawa anti tripsin sehingga tidak mampu memecah protein dan menyebabkan daya cerna protein menurun. Bila anti tripsin terakumulasi dalam saluran pencernaan, senyawa anti tripsin akan menghambat kerja enzim tripsin dan kimotripsin. Keberadaan anti tripsin dalam pakan, bukan saja menekan konsumsi namun juga menekan pertumbuhan mencit yang diberi biji tepary (Osman etal. 2003)
Berdasarkan uraian di atas, maka diperlukan upaya untuk mengeliminasi kandungan tanin dan trypsin inhibitor C. odorata sehingga pemanfaatan C. odorata sebagai pakan ternak menjadi optimal. Upaya menurunkan kandungan tanin dan trypsin inhibitor dapat dilakukan dengan proses mekanik, kimia, dan proses biologis (Roger et al. 2015). Kajian sebelumnya oleh Habiba (2002) menunjukkan bahwa prosesing menggunakan panas dan masak menurunkan tanin dan trypsin inhibitor secara signifikan karena kedua senyawa ini rentan terhadap panas, tetapi Wina et al. (2000) melaporkan adanya peningkatan tanin dalam daun
Calliandra calothyrsus yang dikeringkan secara aerobik. Selain itu, penggunaan basa, asam dan fermentasi dapat pula menurunkan kedua senyawa tersebut (Norton 2000; Roger et al. 2015). Adanya ketidak konsistenan metode pengolahan terhadap tanin dan trypsin inhibitor berdampak terhadap kandungan nutrien dan kecernaan dari tanaman C.odorata. Sebagaimana diketahui bahwa kedua metabolit sekunder tersebut mengikat protein sehingga ketersediaan protein menurun dan mampu menurunkan kecernaan. Dengan demikian, perlu adanya kajian mengenai pengaruh dari metode pengolahan yang diterapkan terhadap tanin dan trypsin inhibitor serta ketersediaan nutrien dan kecernaan dari tanaman
C.odorata.
Tujuan Penelitian
Tujuan dari penelitian ini adalah:
1. Mengevaluasi metode pengolahan terbaik dalam menurunkan kandungan tanin dan trypsininhibitor serta pengaruhnya terhadap komposisi nutrien dan kecernaan in vitroC.odorata
2. Menguji pengaruh fermentasi dengan penambahan putak dan isi rumen sebagai aditif pada level berbeda terhadap kualitas silase C. odorata,
kandungan tanin dan trypsin inhibitor, komposisi nutrien dan kualitas kecernaan in vitro silase C.odorata.
Hipotesis
Hipotesis dari penelitian ini adalah:
3 2. Ada salah satu perlakuan penambahan putak dan isi rumen yang memiliki kualitas silase terbaik, menurunkan tanin dan trypsin inhibitor, mempertahankan komposisi nutrien dan kualitas kecernaan in vitro silase C. odorata.
Manfaat Penelitian
Hasil dari penelitian ini diharapkan dapat dijadikan sebagai landasan pengetahuan dalam pemanfaatan semak bunga putih sebagai pakan ternak ruminansia serta sebagai jembatan untuk penelitian-penelitian selanjutnya.
2
UJI EFEKTIVITAS BERBAGAI MACAM PENGOLAHAN
TERHADAP KONSENTRASI TOTAL TANIN,
TRYPSIN
INHIBITOR
DAN KECERNAAN PAKAN
Chromolaena
odorata
SECARA
In vitro
ABSTRAK
C. odorata merupakan sumber pakan alternatif potensial, namun penggunaannya terkendala oleh kandungan berbagai senyawa metabolik sekunder dalam jaringan tumbuhan ini. Kelompok senyawa tersebut di antaranya adalah tanin dan trypsin inhibitor. Penelitian ini bertujuan untuk mengevaluasi pengaruh berbagai metode perlakuan terhadap konsentrasi total tanin, trypsin inhibitor,daya cerna bahan kering dan bahan organik di in vitro, dan konsentrasi produk fermentasi. Rancangan Acak Lengkap (8x3) digunakan untuk menguji perbedaan 8 jenis perlakuan yaitu daun C. odorata segar sebagai kontrol (Segar), dijemur selama 3x24 jam (Jemur), dioven pada suhu 60oC selama 24 jam (Oven), direbus dalam air selama 5 menit (Rebus), direndam dalam air biasa selama 4 jam (RenAir), direndam dalam NaOH selama 4 jam (RenNaOH), direndam dalam HCl selam 4 jam (RenHCl), atau difermentasi secara anaerobik selama 21 hari (Fermentasi). Peubah yang diukur adalah konsentrasi total tanin, aktivitas trypsin inhibitor, kandungan nutrien, konsentrasi NH3 dan VFA, serta kecernaan bahan
kering (KCBK) dan bahan organik (KCBO). Hasil penelitian menunjukkan bahwa perlakuan jemur, rebus, rendam air, dan fermentasi secara nyata menurunkan total tanin sekitar 4.45–61.9% dibanding kontrol. Penurunan terbesar (61.9%) ditunjukkan oleh perlakuan fermentasi. Sebaliknya, penggunaan panas tinggi (dioven) atau bahan kimia (HCl dan NaOH) tidak menurunkan konsentrasi tanin. Nilai kecernaan (KCBK dan KCBO) dan konsentrasi produk fermentasi rumen (NH3 dan VFA) tidak nyata dipengaruhi oleh perlakuan. Disimpulkan bahwa
metode fermentasi anaerobik dapat digunakan sebagai strategi efektif untuk menurunkan konsentrasi tanin, trypsin inhibitor dalam tumbuhan semak bunga putih (Chromolaena odorata) tanpa mengurangi nilai nutrisinya sebagai bahan pakan.
4
ABSTRACT
C. odorata is a potential feed source but its usage is hampered by presence of various secondary metabolic compounds in the tissues of the plant. Two groups of the active compunds are tannins and trypsin inhibitors. This experiment aimed to evaluate various treatment methods on total tannin concentration and trypsin inhibitor, in vitro digestibility of dry- and organic matter. An 8 x 3 completely randomized experimental design was employed to test 8 different treatments. The treatments were: freshly-chopped chromolaena leaves as control (Segar), sun-dried (3x24 hours) C. odorata leaves (Jemur), oven-dried (60oC for 24 hours) C. odorata leaves (Oven), water-boiled (5 minutes) C. odorata leaves (Rebus), water-soaked (4 hours) C. odorata leaves (RendAir), NaOH-soaked (4 hours) C. odorata leaves (RenNaOH), HCl-soaked (4 hours) C. odorata leaves (RenHCl), and anaerobically-fermented (21 days) C. odorata leaves (Fermentasi). Variable measured were concentration of total tannins, activity of trypsin inhibitor, in vitro
digestibility of dry matter (IVDMD) and organic matter (IVOMD), and concentration of ammonia (NH3) and total volatile fatty acids (VFAs). The results
showed that application of low heat (Jemur), hot water (Rebus), soaking in water (RenAir), and anaerobic fermentation techniques reduced a significant amount of total tannin by 4,45 – 61,9% compared to control. The highest suppression (61.9%) was achieved by fermentasi treatment. In contrast, medium heat application (oven dried at 60oC) and chemical treatments (HCl or NaOH) had no effect. In vitro fermentation data showed that boiling in water or soaking in NaOH reduced IVDMD and IVOMD significantly. All processing methods tested in this present experiment have no effect on NH3 and VFA concentration. It is concluded
that low heat drying or boiling in water or anaerobic fermentation can be used as an effective processing method to reduce total tannin concentration in
Chromolaena odorata. On the other hand, soaking or boiling in water or soaking in NaOH solution will reduce rate of in vitro digestion.
Keywords: C. odorata, digestibility, NH3, tannins, trypsin inhibitor, VFA.
PENDAHULUAN
Semak bunga putih (Chromolaena odorata) merupakan gulma padang penggembalaan di wilayah Indonesia Timur dengan produksi biomasa yang sangat tinggi (mencapai 70 ton BK ha-1 thn-1) dengan kandungan protein kasar berkisar 21-36% (Mullik 2002), sehingga mampu menampung ternak sapi dengan bobot 200 kg sebanyak 31 ekor tahun-1. Kendala pemanfaatan C.odorata yaitu adanya berbagai senyawa metabolik sekunder yang bersifat anti nutrisi. Metabolit sekunder tersebut antara lain tanin, anti tripsin, haemaglutinin, saponin, oxalate, asam pitat, alkaloid, steroid, terpoids, flavonoid (Akinmoladun et al. 2010; Onkaramurthy et al. 2013). Keberadaan senyawa-senyawa metabolik sekunder tersebut menyebabkan palatabilitas C. odorata rendah (Hai et al. 2012) karena bau
5
odorata menjadi palatabel bagi ternak. Namun, yang menjadi pertimbangan utama dalam pengolahan awal adalah perlakuan yang diberikan harus menurunkan/menghilangkan senyawa anti nutrisi dalam bahan pakan tetapi tetap mempertahankan nilai nutrisi bahan pakan tersebut, serta tidak membahayakan kesehatan ternak dan lingkungan.
Mengingat tanin dan trypsin inhibitor merupakan senyawa anti nutrisi dominan dalam C. odorata (Onkaramurthy et al. 2013) maka penurunan atau penghilangan kandungan tanin dalam C. odorata kemungkinan besar akan meningkatkan palatabilitas dan nilai manfaatnya sebagai bahan pakan. Pakan ternak yang berkandungan tanin tinggi dapat menurunkan konsumsi, menurunkan daya cerna, dan menurunkan pertambahan bobot badan harian bila diberi sebagai pakan tunggal (Wina 2010), mengurangi degradasi protein dalam rumen sebagai akibat berikatannya tanin dan protein (Patra dan Saxena 2011), serta menimbulkan efek toksik bagi mikroba rumen (Bhatta et al. 2009). Selain itu, tanin pun dapat melukai saluran pencernaan sehingga mengganggu fungsi saluran pencernaan (Makkar 2003) karena terjadi penghambatan aktivitas enzim pencernaan termasuk protease, lipase dan glikosidase (Hagerman 1992). Trypsininhibitor menghambat aktivitas enzim proteolitik sehingga daya cerna protein akan menurun. Bila anti tripsin terakumulasi dalam saluran pencernaan, senyawa anti tripsin akan menghambat kerja enzim tripsin dan kimotripsin. Keberadaan anti tripsin dalam pakan, bukan saja menekan konsumsi namun juga menekan pertumbuhan mencit yang diberi biji tepary (Osman etal. 2003)
Teknik pengolahan yang digunakan untuk menurunkan atau meminimalisir efek negatif dari tanin melalui proses fisik, kimia, dan biologis (Roger et al.
2015). Teknik pengolahan secara fisik (pencacahan, penggilingan, pembuatan pelet) atau kimia (pemanasan, perendaman dengan air atau bahan kimia yang bersifat asam atau basa) atau biologis (fermentasi mikroba) telah menjadi cara pengolahan umum yang dipakai secara luas sehingga dapat menjadi pilihan untuk mengurangi kandungan tanin dalam C. odorata. Namun, berbagai hasil riset menunjukkan bahwa respon tanin terhadap perlakuan tidak konsisten antar bahan pakan. Sebagai contoh, pelayuan dan pengeringan terbukti secara nyata menurunkan konsentrasi tanin pada daun singkong (Hue et al. 2010), tetapi Wina
et al. (2000) melaporkan adanya peningkatan tanin dalam daun Calliandra calothyrsus yang dikeringkan secara aerobik. Disebabkan oleh adanya inkonsistensi respon tanin terhadap pengolahan, maka tidak dapat dipastikan teknik pengolahan apa yang paling efektif untuk menurunkan konsentrasi tanin dalam C. odorata sebagai upaya mengolahnya menjadi pakan ternak. Atas dasar tersebut, maka dalam penelitian ini diuji efektivitas teknik pengeringan matahari, pengovenan, perebusan, perendaman dalam air atau asam (HCl) atau basa (NaOH), dan fermentasi terhadap kandungan tanin, kandungan nutrisi dan kecernaan in vitro dari C. odorata.
METODE
Waktu dan Tempat Penelitian
6
Peternakan IPB. Evaluasi trypsin inhibitor dan kecernaan in vitro dilaksanakan di Laboratorium Nutrisi dan Ternak Perah, Fakultas Peternakan IPB.
Bahan dan Persiapannya
Bahan yang digunakan adalah daun C. odorata yang diperoleh dari Kupang, Nusa Tenggara Timur. Daun tanaman C. odorata dipanen pada bulan Agustus 2014, dipangkas dengan panjang sekitar ±50 cm dari ujung/pucuk tanaman. Hasil pangkasan dibawa ke Bogor, selanjutnya daun Chromolaena odorata dipisahkan dari bagian batang untuk digunakan sebagai bahan penelitian. Komposisi kimia daun C. odorata ditampilkan pada Tabel 1.
Tabel 1 Komposisi kimia daun Chromolaena odorata*
Komposisi kimia Persentase (% BK)
Bahan Kering 26.61
Abu 9.08
Protein Kasar 17.54
Serat Kasar 12.87
Lemak Kasar 2.57
Beta-N 57.94
Neutral Detergent Fiber 69.29
Hemiselulosa 16.35
Acid Detergent Fiber 52.95
Selulosa 30.16
Lignin 22.55
Silika 0.24
*Hasil analisis Laboratorium Ilmu dan Teknologi Pakan, Fakultas Peternakan IPB 2014
Perlakuan dan Rancangan Percobaan
Penelitian ini terbagi atas dua tahap. Tahap pertama terdiri dari 8 perlakuan untuk mengetahui pengaruh perlakuan terhadap kandungan total tanin dan trypsin inhibitor. Desain penelitian sebagai berikut:
Segar = C. odorata segar
Jemur = C. odorata dijemur selama 3 x 24 jam Oven = C. odorata dioven 60oC selama 24 jam Rebus = C. odorata direbus selama 5 menit
RenAir = C. odorata direndam dalam air selama 4 jam
RenNaOH = C. odorata direndam dalam NaOH 0.1N 10% selama 4 jam RenHCl = C. odorata direndam dalam HCl 0.1 N 10% selama 4 jam Fermentasi = C. odorata difermentasi tanpa bahan aditif selama 21 hari
7 dan dikeringkan lalu digiling halus kemudian digunakan untuk analisis. Bahan untuk pelakuan RendAir, diperoleh dengan cara merendam 200 g daun segar dalam 1 L air biasa pada suhu ruangan selama 4 jam. Perlakuan RenNaOH dan RenHCl, prosedurnya sama seperti perlakuan RenAir, hanya tidak digunakan air biasa tetapi digunakan larutan NaOH atau HCl 0,1N 10%. Perlakuan fermentasi, daun C. odorata segar dicincang, dimasukan dalam toples ukuran 1 liter, dipadatkan, ditutup rapat (kedap udara) selama 21 hari. Setelah proses fermentasi, toples dibuka, bahan dikeluarkan dan dianginkan, selanjutnya dikeringkan, digiling halus dan dibawa ke laboratorium untuk analisis.
Penelitian tahap kedua difokuskan untuk mengetahui kualitas fermentatif secara in vitro dari tumbuhan C. odorata. Semua perlakuan sama seperti pada tahap pertama di atas, tetapi ada satu perlakuan yang dihilangkan yaitu perlakuan Segar. Dengan demikian, tinggal 7 perlakuan yaitu Jemur, Oven, Rebus, RenAir, RenNaOH, RenHCl, dan Fermentasi. Alasan dikeluarkannya perlakuan segar karena semua proses menggunakan bahan yang telah jemur kering, dengan demikian, tidak ada material segar yang digunakan.
Rancangan percobaan yang digunakan dalam kedua tahap penelitian adalah Rancangan Acak Lengkap (RAL) dengan model matematis sebagai berikut:
Xij= μ + τi+ εij
Keterangan:
Xij = nilai pengamatan perlakuan ke-i, ulangan ke-j
μ = rataan umum
τi = pengaruh perlakuan ke-i
εij = eror perlakuan ke-i, ulangan ke-j
Pada penelitian tahap pertama digunakan RAL dengan konfigurasi 8 perlakuan dan 3 ulangan. Sementara itu, pada penelitian tahap kedua digunakan RAL 7 perlakuan dan 3 ulangan. Data yang diperoleh dianalisis dengan ANOVA (Analysis of Variance) untuk mengetahui pengaruh perlakuan terhadap peubah yang diamati. Jika perlakuan berpengaruh nyata maka dilakukan uji lanjut Duncan (α 0.05). Proses analisis data menggunakan SPSS versi 16.0.
Peubah yang Diamati
Peubah yang diamati dalam penelitian ini adalah:
Komposisi nutrien. Analisis komposisi nutrien C. odorata meliputi: kandungan bahan kering (BK), bahan organik (BO), protein kasar (PK), lemak kasar (LK), dan serat kasar (SK) C. odorata menggunakan metode AOAC (2005).
Kandungan metabolit sekunder. Pengukuran kandungan metabolit sekunder meliputi: a) Kandungan total tanin, diuji menggunakan metode titrimetric (Atanassova dan Christova-Bagdassarian 2009); b) kandungan trypsin inhibitor menggunakan metode Kakade et al (1974).
Kualitas fermentasi rumen secara in vitro, meliputi: a) konsentrasi
ammonia (NH3) diukur menggunakan metode Difusi Conway (Conway dan
8
Nilai kecernaan in vitro C. odorata, meliputi: a) kecernaan bahan kering
(KCBK); b) kecernaan bahan organik (KCBO) secara in vitro menggunakan metode Tilley dan Terry (1963).
HASIL DAN PEMBAHASAN
Kandungan Metabolit Sekunder
Tanin merupakan senyawa fenolik yang memiliki kemampuan menghambat aktivitas mikroba rumen dengan cara menonaktifkan enzim, disamping dapat mengakibatkan keracunan bagi mikroba (Makkar dan Singh 1991). Tanin dapat dieliminasi atau diturunkan dengan pemanasan, pengeringan,
perendaman menggunakan air, asam, alkali, oxidising, urea, polyvinyl-pyrrolidine (PVP), PEG, dan ferric salts (Norton 2000).
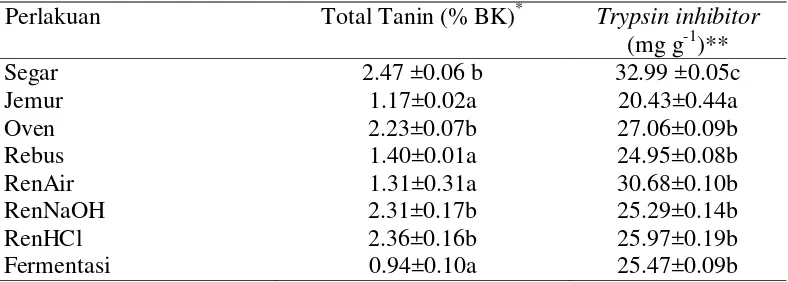
Data konsentrasi total tanin dalam C. odorata yang telah diberikan berbagai perlakuan (Tabel 2) menunjukkan bahwa semua perlakuan menurunkan kandungan tanin C. odorata tetapi dengan derajat penurunan yang berbeda. Tingkat penurunan terbesar yakni 62% diperlihatkan oleh perlakuan fermentasi (0.94%) dibanding dengan kontrol (2.47%). Penurunan yang cukup tinggi (43-53%) juga akibatkan oleh perlakuan jemur (1.17%), rebus (1.40%), dan rendam air (1.31%), sedangkan tingkat penurunan terkecil (4-9%) didapati pada perlakuan Oven (2.23%), RenNaOH (2.31%), dan RenHCl (2.36%). Konsentrasi total tanin yang dideteksi dalam penelitian ini berada dalam rentangan nilai 1.3%-17.2% yang dilaporkan oleh Gemeda dan Hassen (2015) pada berbagai jenis hijauan semak tropis di Afrika Selatan.
Tabel 2 Kandungan metabolit sekunder C. odorata akibat berbagai perlakuan Perlakuan Total Tanin (% BK)* Trypsin inhibitor
(mg g-1)**
Huruf kecil yang berbeda pada kolom yang sama menunjukkan berbeda nyata pada taraf uji 5% (uji selang berganda Duncan); *Hasil analisis Laboratorium Ilmu dan Teknologi Pakan, Fakultas Peternakan IPB (2014) ** Hasil analisis Laboratorium Nutrisi Ternak Perah, Fakultas Peternakan IPB (2015)
9 menurunkan konsentrasi total tanin. Penjelasan pasti tentang akibat perlakuan terhadap penurunan konsentrasi tanin tidak memungkinkan untuk dilakukan karena jenis tanin yang ada dalam C.odorata belum diketahui dan tidak juga dikarakterisasi dalam penelitian ini. Namun, diduga bahwa penurunan tanin akibat fermentasi dalam penelitian ini dikarenakan adanya aktivitas kimia dari enzim-enzim yang diproduksi oleh berbagai mikrorganisme fermentatif sehingga merusak ikatan tanin-enzim dan protein-tanin, akibatnya tanin terbebaskan dan larut dalam cairan residu fermentasi (Taylor and Duodu 2014). Selain itu, kemungkinan enzim-enzim mikroba menghidrolisis lebih lanjut sebagian tanin, terutama yang larut menjadi senyawa lain selama proses fermentasi anaerobik berlangsung.
Penurunan tanin yang sangat nyata pada perlakuan rebus dan rendam air kemungkinan juga berhubungan dengan larutnya senyawa tanin mudah larut (jenis
gallotannins) dalam air sehingga tidak terdeteksi dalam sampel karena yang digunakan hanya sampel padat saja. Diakui, bahwa kelemahan dalam penelitian ini adalah tidak dilakukan identifikasi jenis tanin sehingga tidak diketahui berapa proporsi tanin yang mudah larut dibanding dengan tiga kelompok lainnya (ellagitannins, complex tannins dan condensed tannins). Larutnya tanin dalam air dapat dipahami karena sebagian besar tanin jenis gallotannins memiliki ikatan residu polyol yang berasal dari D-glucose (Khanbabaee dan van Ree 2001). Menurut kedua peneliti tersebut (Khanbabaee dan van Ree 2001), gallotannins jenis 2,3,4,6-tetra-O-galloyl-D-glucopyranose dan 1,2,3,4,6-penta-O-galloyl-β -D-glucopyranose merupakan senyawa intermediate kunci yang berperan penting dalam biosintesis hampir semua senyawa phenolik dalam tanaman. Dengan adanya air, maka akan menjadi media yang baik bagi reaksi-reaksi kimia yang berhubungan dengan hidrolisis tanin.
Pemberian panas rendah (penjemuran di bawah sinar matahari ) juga sangat nyata menurunkan konsentrasi tanin (53%) dalam C.odorata. Tren ini jauh berbeda dengan pemberian panas tinggi (dioven pada suhu 60oC) di mana terjadi penurunan hanya sebesar 9% sehingga tidak berbeda nyata dengan kontrol. Penurunan kadar tanin yang kecil (3.4%) juga dilaporkan oleh Rakić et al. (2004) ketika kulit pohon oak dioven pada suhu 60oC. Menurut Djordjević (1995) reaksi penting yang terjadi terhadap tanin akibat pemanasan adalah reaksi hidrolisis, oksidasi, polimerisasi, interaksi dari komposisi dan reaksi dekomposisi.
Berbedanya reaksi tanin terhadap suhu, mungkin dapat dijelaskan oleh pendapat Hagerman (2002) bahwa reaksi terhadap tanin sangat dipengaruhi oleh suhu di mana suhu tinggi cenderung meningkatkan konsentrasi tanin sebab pada suhu tinggi, terjadi kondensasi sehingga membentuk ikatan yang kompleks dengan senyawa lain. Sebaliknya, pengeringan secara lambat pada suhu rendah seperti pada perlakuan jemur mungkin memberikan kesempatan terjadinya hidrolisis dan dekomposisi tanin (Makkar dan Becker 1996) karena memiliki ikatan-ikatan hirophobic, maka akan membentuk ikatan ionik kuat pada suhu tinggi (Haslam 1989). Inilah yang menjelaskan mengapa terjadi penurunan kadar tanin yang tidak nyata pada perlakuan oven.
10
(1993) pada asam tanat. Hal ini kemungkinan berkaitan dengan ionisasi pada gugus-gugus hidroksil phenol pada tanin (Hagerman 2002) sehingga membuatnya kurang mengalami hidrolisis.
Trypsin inhibitor merupakan salah satu senyawa protein yang bersifat menghambat kerja enzim trypsin dalam menghidrolisis protein. Trypsin inhibitor
dalam pakan akan mengurangi konsumsi, gangguan pencernaan protein, hipertropi pankreatik dan menghambat pertumbuhan (Soetrisno dan Suryana 1991). Trypsin inhibitor memiliki dua kelompok utama, yaitu Kunitz Trypsin Inhibitor (KTI) dan
Browman-Birk inhibitor (BBI). KTI memiliki berat molekul 21 kD dengan dua ikatan disulfida, sedangkan BBI mempunyai berat molekul 8 kD yang mengandung cystein lebih tinggi dengan membentuk 7 ikatan disulfida. Perbedaan struktur antara kedua jenis inhibitor ini menyebabkan perbedaan sifat stabilitasnya terhadap panas. KTI tidak stabil terhadap panas, sedangkan BBI sangat stabil terhadap pemanasan karena strukturnya lebih kompleks dan kokoh dibanding KTI (Kanetro et al. 2014).
Data konsentrasi trypsin inhibitor dalam C.odorata yang telah diberikan berbagai perlakuan (Tabel 2) menunjukkan bahwa semua perlakuan menurunkan
trypsin inhibitor dalam C. odorata tetapi dengan derajat penurunan yang berbeda. Tingkat penurunan terbesar yakni 38.07% diperlihatkan oleh perlakuan jemur (20.43 mg g-1) dibanding dengan kontrol (32.99 mg g-1). Penurunan yang cukup tinggi (17.98-24.37%) juga akibatkan oleh perlakuan oven (27.06 mg g-1), RenHCl (25.97 mg g-1), fermentasi (25.47 mg g-1), RenNaOH (25.29 mg g-1), dan Rebus (24.95%), sedangkan tingkat penurunan terkecil 7% didapati pada perlakuan rendam air (30.68 mg g-1). Konsentrasi trypsin inhibitor semua perlakuan dalam penelitian ini lebih tinggi dari yang dilaporkan Ikhimioya et al.
(2007) sebesar 22.37 mg g-1, kecuali perlakuan jemur dengan konsentrasi trypsin inhibitor (20.43 mg g-1).
Hasil analisis sidik ragam menunjukkan bahwa perlakuan jemur sangat nyata (p<0.001) menurunkan aktivitas trypsin inhibitor, diikuti perlakuan rebus, RenNaOH, Fermentasi, RenHCl, dan perlakuan oven, sedangkan perlakuan rendam air tidak nyata berbeda menurunkan aktivitas trypsin inhibitor dibanding kontrol. Penurunan aktivitas trypsin inhibitor pada perlakuan panas (jemur dan oven) senada dengan yang dilaporkan Vidal-Valverde et al. (1994) bahwa proses perlakuan panas dapat menonaktifkan aktivitas trypsin inhibitor pada lentils. Hal ini dikarenakan salah satu sifat dari trypsin inhibitor yaitu tidak stabil pada panas. Panas dapat merubah struktur trypsin inhibitor atau interaksinya dengan konstituen lain. Pemanasan menyebabkan adanya kelompok thiol bebas yang dapat menukar ikatan disulfide sehingga membantu dalam inaktivasi inhibitor (Vidal-Valverde et al. 1994). Selain itu, konsentrasi protein yang cukup tinggi dapat meningkatkan kemungkinan interaksi nonkovalen protein-protein, yang menyebabkan hilangnya aktivitas penghambatan melalui denaturasi dan atau pengendapan (DiPietro dan Liener 1989). Menurunnya aktivitas trypsin inhibitor
pada perlakuan rebus diduga bahwa trypsin inhibitor lepas selama proses perebusan dan juga adanya panas yang dapat menginaktivasi trypsin inhibitor
kemungkinan karena terjadi perubahan konfigurasi kimia akibat pemanasan. Penurunan aktivitas trypsin inhibitor pada perlakuan kimia (RenNaOH dan RenHCl) berhubungan dengan perubahan suasana asam atau basa. McDonald et al.
11 asam dan basa yang berbeda-beda. Pada larutan asam molekul protein akan bermuatan positif dan pada larutan basa protein akan bermuatan negatif. Pada kisaran pH tertentu, jumlah muatan positif dan negatif pada protein dapat sama jumlahnya dan saling menetralkan, sehingga protein akan menggumpal dan mengendap. Perubahan suasana asam atau basa mengakibatkan ikatan kompleks enzim-inhibitor menjadi lemah dan hidrolisis berjalan lambat sehingga aktivitas inhibitor berkurang (Kanetro et al. 2014).
Perlakuan fermentasi menurunkan trypsin inhibitor karena diduga protease yang dihasilkan dari jaringan tanaman atau sel mikroba selama fermentasi dapat memotong molekul protein menjadi peptida dan asam-asam amino. Terpotongnya molekul protein mengakibatkan trypsin inhibitor terdegradasi atau termodifikasi sehingga kehilangan aktivitasnya dalam mengikat trypsin (Chen et al. 2013).
Tidak adanya perbedaan aktivitas trypsin inhibitor pada perlakuan rendam air terhadap kontrol senada dengan laporan Liu dan Markakis (1987) bahwa perendaman biji kedelai menggunakan air dengan suhu 22oC selama 24 jam tidak signifikan menurunkan aktivitas trypsin inhibitor. Selanjutnya, Trugo et al. (1990) juga melaporkan hal yang sama ketika black beans direndam dalam air selama 16 jam tidak menurunkan aktivitas trypsin inhibitor. Kondisi ini menunjukkan bahwa
trypsin inhibitor tidak reaktif terhadap air pada suhu lingkungan. Komposisi Nutrien C. odorata setelah perlakuan
Kandungan nutrien C.odorata dengan berbagai perlakuan ditampilkan pada Tabel 3. Ada tiga variabel nutrisi penting penentu nilai biologis bahan pakan dari C.odorata yang dipengaruhi oleh perlakuan yaitu kandungan bahan organik, protein kasar, dan serat kasar. Data pada Tabel 3 memperlihatkan bahwa kadar bahan organik tertinggi terdapat pada perlakuan RenHCl (92.2%) dan yang terendah pada perlakuan fermentasi (87.9%). Bahan organik pada umumnya terdiri dari komponen hidrogen sehingga peluang tanin untuk berikatan dengan bahan organik lain pun sangat tinggi seperti selulosa dan hemiselulosa (McSweeney et al. 2001). Dengan adanya perlakuan RenHCl pada daun
C.odorata maka ikatan antara tanin dan protein maupun ikatan tanin dan bahan organik lainnya terlepas. Sebab asam memiliki kemampuan dalam menghidrolisis tanin yang berakibat terhadap lepasnya bahan organik. Namun, hasil ini tidak sejalan dengan hasil pada Tabel 2, di mana kemampuan HCl dalam menurunkan tanin C.odorata lebih rendah dibanding kemampuan fermentasi dalam menurunkan total tanin. Hal ini dikarenakan pada perlakuan RenHCl bahan organik yang terlepas tidak dirombak lagi, sebaliknya pada perlakuan fermentasi walaupun kemampuan mengurangi tanin tinggi yang berakibat terhadap tingginya bahan organik yang terlepas, namun bahan organik tersebut dimanfaatkan kembali oleh mikroba selama proses fermentasi berlangsung sehingga ketersediaan bahan organik menjadi berkurang. Penambahan bahan aditif berupa karbohidrat mudah larut sangat disarankan dalam proses fermentasi guna meningkatkan nilai nutrisi dan mengurangi kehilangan bahan kering (McDonald et al. 1991).
12
kasar C.odorata tertinggi pada perlakuan fermentasi sebesar 29.0% dan yang rendah pada perlakuan segar sebesar 17.4%. Perlakuan fermentasi meningkatkan protein kasar C.odorata karena kemampuannya yang tinggi dalam menurunkan tanin yang berakibat terhadap banyaknya protein yang terlepas. Selain itu, karena adanya aktivitas kimia dari organisme fermentasi dan rusaknya tanin-enzim dan protein-tanin sehingga lepas sebagai tanin bebas (Taylor and Duodu 2014). Kandungan protein kasar pada perlakuan rebus (19.6%) walaupun lebih tinggi dari segar, namun lebih rendah dari perlakuan lainnya. Hal ini kemungkinan berhubungan dengan larutnya senyawa tanin yang larut dalam air yang masih berikatan dengan protein. Dengan adanya perebusan, terjadi juga proses pemanasan sehingga diduga protein dari C. odorata mengalami denaturasi.
Tabel 3 Kandungan nutrien C. odorata akibat berbagai perlakuan*
Perlakuan
Segar 26.6±0.02b 90.8±0.01c 17.4±0.02f 3.0±0.11e 12.7±0.03d 57.8±0.02a
Jemur 83.9±0.01a 86.6±0.0f 25.8±0.02b 4.8±0.04d 10.8±0.03c 45.2±0.01d
Oven 86.3 ±0.0a 88.8±0.01d 25.0±0.02c 6.7±0.02c 9.9±0.02b 47.1±0.02b
Rebus 24.5±0.00b 91.4±0.01b 19.6±0.02e 10.6±0.0a 13.5±0.01e 47.7±0.04b
RenAir 27.1±0.79b 91.4±0.01b 22.8±0.03c 10.9±0.0a 14.5±0.02f 43.3±0.05e
RenNaOH 24.7±0.12b 86.7±0.0f 19.9±0.04de 6.2±0.08c 13.0±0.04d 47.4±0.03b
RenHCl 18.3±0.00c 92.2±0.01a 20.2±0.03d 10.7±0.0a 15.1±0.03g 46.2±0.03c
Fermentasi 24.3±0.01b 87.9±0.01e 29.0±0.01a 7.5±0.00b 8.6±0.06a 42.9±0.04e
Huruf yang berbeda pada kolom yang sama menunjukkan perakuan berbeda nyata (P<0.05).*Hasil analisis Laboratorium Ilmu dan Teknologi Pakan Fakultas Peternakan IPB (2014)
Lemak kasar C.odorata segar sebesar 3%, setelah adanya perlakuan komposisi lemak kasar meningkat. Peningkatan ini diduga disebabkan oleh berkurangnya kandungan air akibat adanya perlakuan misalnya pengeringan sehingga proporsi lemak meningkat (Suharyanto 2007). Meningkatnya kadar lemak C.odorata juga diduga karena lemak yang berikatan dengan protein (lipoprotein) terpisah sehingga dapat meningkatkan kandungan lemak (Almatsier 2004).
Serat kasar terendah terdapat pada perlakuan fermentasi yaitu sebesar 8.6%, sedangkan serat kasar tertinggi terdapat pada perlakuan rendam air (14.5%). Penurunan serat kasar pada perlakuan fermentasi dikarenakan adanya aktifitas bakteri pencerna serat selama proses fermentasi berlangsung. Diantara spesies selulolitik ada yang berfungsi mencerna serat, berupa selulosa, hemiselulosa dan pati (Kana Hau et al. 2005). Kemampuan perlakuan rendam air dalam menurunkan serat kasar sangat rendah. Dapat dipahami bahwa serat terdiri atas selulosa, hemiselulosa dan lignin. Selulosa dan hemiselulosa umumnya akan didegradasi dengan bantuan enzim selulose dan hemiselulose, sedangkan air tidak memiliki kemampuan dalam mendegradasi selulosa dan hemiselulosa.
13 mudah larut belum terdegradasi sedangkan pada perlakuan fermentasi diduga senyawa karbohidrat yang mudah larut telah terdegradasi oleh mikroba untuk kelangsungan hidup selama proses fermentasi. Penambahan bahan aditif berupa karbohidrat mudah larut sangat disarankan dalam proses fermentasi (McDonald et al. 1991).
Kualitas Fermentasi Rumen secara invitro
Konsentrasi NH3 merupakan indikator adanya perombakan protein yang
masuk dalam rumen. Produk NH3 yang dihasilkan dimanfaatkan oleh beberapa
mikroba rumen untuk sintesis protein mikroba serta untuk pertumbuhannya. Arora (1995) menyatakan bahwa mikroba rumen memanfaatkan kembali NH3 yang
terbentuk untuk membangun sel tubuhnya.
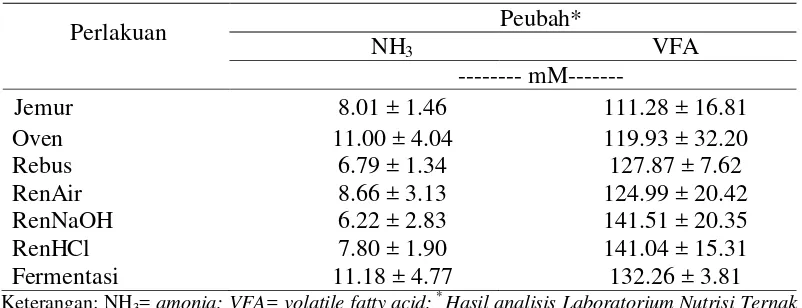
Konsentrasi NH3 C. odorata akibat diberikan berbagai perlakuan
memperlihatkan pengaruh tidak nyata (Tabel 4). Bahan pakan yang diproteksi tanin dapat menurunkan produksi NH3 (Subrata et al. 2005). Konsentrasi tertinggi
terdapat pada perlakuan fermentasi (11.18 mM), dan yang terendah terdapat pada perlakuan RenNaOH (6.22 mM). Konsentrasi ini cukup untuk memenuhi kebutuhan sintesis protein mikroba rumen yang berkisar antara 6 hingga 21 mM (McDonald et al. 2002). Hasil penelitian ini senada dengan yang dilaporkan Gemeda dan Hassen (2015) bahwa konsentrasi NH3 bahan pakan yang
mengandung tanin berkisar antara 3.88 mM hingga 21.30 mM.
Tabel 4 Kualitas fermentatif invitroC. odorata pada berbagai perlakuan
Perlakuan Peubah*
Keterangan: NH3= amonia; VFA= volatile fatty acid; * Hasil analisis Laboratorium Nutrisi Ternak
Perah Fakultas Peternakan IPB (2014)
Tingginya konsentrasi NH3 pada perlakuan fermentasi (11.18 mM)
menunjukkan bahwa terjadi perombakan protein dalam jumlah yang besar dibanding perlakuan lain. Meningkatnya konsentrasi NH3 dikarenakan protein
yang tersedia mudah terdegradasi akibat berkurangnya tanin sehingga kehilangan kemampuan dalam melindungi protein (Tabel 2). Selain itu, tingginya energi pakan serta tingginya pertumbuhan mikroba rumen pun berdampak terhadap meningkatnya konsentrasi NH3. Fermentasi protein menghasilkan produk akhir
NH3 yang sangat penting untuk sintesis protein di dalam rumen. Amonia dalam
rumen sebagian dimanfaatkan oleh mikroba untuk sintesis protein mikroba (Arora 1995). Perlakuan RenNaOH memiliki konsentrasi NH3 terendah (6.22 mM) dan
14
masih tinggi sehingga masih memiliki kemampuan sebagai senyawa pelindung protein selama proses fermentasi protein berlangsung. Gemeda dan Hassen (2015) melaporkan bahwa produksi NH3 bervariasi antar setiap hijauan, di mana hijauan
dengan kandungan tanin rendah dan tinggi memiliki produksi NH3 yang sama. Hal
ini menunjukkan bahwa produksi NH3 tidak semata-mata dipengaruhi oleh
kandungan tanin tetapi juga dipengaruhi oleh faktor lain seperti sifat kimia dari masing-masing bahan pakan (Mueller-Harvey 2006).
Produk VFA merupakan hasil hidrolisa polisakarida menjadi monosakarida oleh enzim-enzim mikroba rumen, dan selanjutnya difermentasi menjadi VFA serta gas metan (CH4) dan CO2. VFA akan diserap melalui dinding
rumen sedangkan gas CH4 dan CO2 akan hilang melalui eruktasi. Senyawa VFA
terdiri dari dua kelompok yaitu VFA berantai lurus dan berantai cabang. Kelompok VFA berantai lurus terdiri dari asetat, propionat, dan butirat, sedangakan VFA berantai cabang meliputi iso butirat, valerat, dan iso-valerat. Propionat akan digunakan oleh jaringan hati ternak untuk sintesis glukosa melalui jalur glukoneogenesis (Bergman et al. 1971), sedangkan asetat dan butirat merupakan bahan baku untuk sintesis lemak dalam jaringan lemak (Beits dan Nizzi 1997). VFA berantai cabang digunakan oleh mikroba rumen sebagai sumber kerangka karbon (Andries et al.1987), sedangkan amonia menjadi sumber nitrogen (Van Soest 1994) bagi pertumbuhan mikroba rumen.
Konsentrasi VFA tergantung pada jenis ransum yang di konsumsi, penyerapan serta fermentabilitas dari karbohidrat dan pemanfaatan mikroba (McDonald et al. 2002). Konsentrasi VFA in vitro daun C.odorata yang telah diberikan berbagai perlakuan menunjukkan tidak ada perbedaan yang nyata (Tabel 4). Kisaran VFA yang dihasilkan dalam penelitian ini antara 111.28 mM hingga 141.51 mM. Hasil ini bila dibandingkan dengan Gemeda dan Hassen (2015), bahan pakan dengan kandungan tanin lebih tinggi dari C.odorata justru memiliki produksi VFA yang setara. Variasi konsentrasi VFA C.odorata dan tanaman mengandung tanin lainnya seperti yang dilaporkan Gemeda dan Hassen (2015) mungkin dipengaruhi oleh adanya perbedaan kandungan karbohidrat dan protein dari tanaman, adanya peningkatan mikroba dapat pula meningkatkan aktivitas fermentasi sehingga mempengaruhi konsentrasi VFA. Selain itu, variasi konsentrasi VFA dapat diakibatkan oleh perbedaan dari sumber, jenis dan level tanin dan sifat kimia dari masing-masing bahan pakan (Makkar dan singh 1991, Mueller-Harvey 2006, Patra dan Saxena 2011).
Nilai Kecernaan InvitroC. odorata
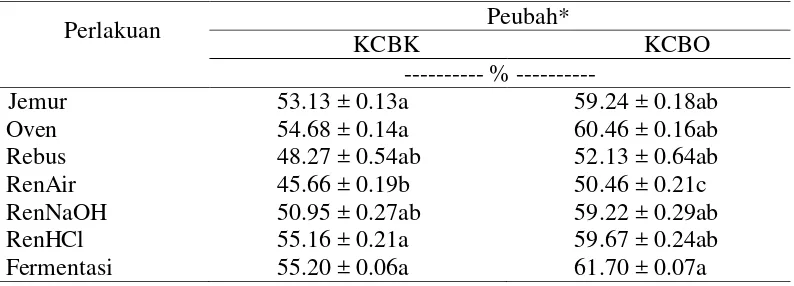
Kecernaan pakan dapat dijadikan sebagai tolok ukur dalam menilai kualitas suatu pakan. Kecernaan memberikan gambaran mengenai jumlah zat makanan yang dapat dicerna. Semakin tinggi kecernaan maka semakin tinggi juga nutrisi yang dapat dimanfaatkan. Hasil pengukuran KCBK dan KCBO C. odorata
dengan berbagai perlakuan selama penelitian disajikan dalam Tabel 5. Dalam penelitian ini terlihat bahwa KCBK dan KCBO dipengaruhi (P<0.05) oleh perlakuan yang diberikan.
Data pada Tabel 5 memperlihatkan bahwa KCBK dan KCBO C. odorata
masing-15 masing 45.66% dan 50.46%). Kecernaan bahan kering berkorelasi dengan kecernaan bahan organik. Bila kecernaan bahan kering tinggi maka kecernaan bahan organik akan meningkat sehingga peluang nutrisi yang dapat dimanfaatkan pun tinggi. Tingginya kecernaan pada perlakuan fermentasi dapat dijelaskan menggunakan data pada Tabel 2 dan Tabel 3 di mana konsentrasi tanin dan serat kasar C.odorata rendah pada perlakuan fermentasi adalah yang paling rendah. Kondisi ini memudahkan mikroba rumen untuk memanfaatkan karbohidrat dan protein serta bahan organik lainnya. Perlakuan rendam air memiliki nilai kecernaan rendah karena perlakuan ini memiliki kemampuan yang rendah dalam menurunkan tanin serta memiliki kandungan serat kasar yang lebih tinggi dibanding perlakuan fermentasi. Tingginya kandungan serat kasar mengurangi aktivitas mikroba rumen dalam mencerna pakan. Menurut Guglielmelli et al. (2011) konsentrasi tanin yang berbeda dalam bahan pakan yang sama akan menunjukkan variasi dalam kecernaan. Perbedaan konsentrasi tanin inilah yang menyebabkan adanya variasi dalam kecernaan pada penelitian ini.
Tabel 5 Nilai kecernaan in vitroC. odorata pada berbagai perlakuan
Perlakuan Peubah*
Keterangan: KCBK= kecernaan bahan kering; KCBO= kecernaan bahan organik; Huruf kecil yang berbeda pada kolom yang sama menunjukkan berbeda nyata pada taraf uji 5% (uji selang berganda Duncan); * Hasil analisis Laboratorium Nutrisi Ternak Perah Fakultas Peternakan IPB (2014)
16
sumber protein pakan. Sumber protein dengan nilai kelarutan yang tinggi, memiliki nilai kecernaan yang tinggi. Selain itu, setiap protein memiliki ketahanan degradasi terhadap mikroba rumen yang berbeda-beda. Diduga, kelarutan protein C. odorata rendah serta ketahanan degradasi protein yang tinggi sehingga nilai kecernaannya rendah. Ketahanan degradasi protein yang tinggi diduga karena adanya tanin yang berfungsi sebagai agen defaunasi.
SIMPULAN
Pemanasan kering suhu rendah, pemanasan basah, dan fermentasi efektif menurunkan kandungan total tanin dan trypsin inhibitor, sedangkan pemanasan kering suhu tinggi dan penggunaan bahan kimia (NaOH dan HCl) tidak memiliki efek. Di lain pihak, perendaman air atau NaOH atau perebusan cenderung menurunkan nilai kecernaan in vitro C. odorata. Konsentrasi ammonia dan asam lemak terbang tidak dipengaruhi oleh semua metode pengolahan yang digunakan dalam penelitian ini.
3
KUALITAS SILASE
C. odorata
DENGAN DAN TANPA
PENAMBAHAN ISI RUMEN SAPI
ABSTRAK
Penelitian tahap pertama menunjukkan bahwa fermentasi merupakan metode terbaik dalam menurunkan kandungan metabolit sekunder C.odorata, namun terjadi penurunan kandungan karbohidrat mudah tercerna sebagai akibat dari belum adanya penambahan bahan aditif berupa karbohidrat mudah larut dan kurang optimal kerja bakteri dalam proses fermentasi C.odorata. Tujuan dari penelitian ini adalah mengkaji pengaruh penambahan zat aditif berupa tepung
putak (sebagai sumber karbohidrat) dan isi rumen (sebagai sumber enzim)dalam pembuatan silase C. odorata terhadap kandungan metabolit sekunder, kualitas fisik silase, kandungan nutrien, kualitas fermentasi rumen dan kecernaan in vitro. Perlakuan yang digunakan terdiri atas 4 macam, yaitu: CO (C. odorata segar), COP (C. odorata segar + tepung putak 10%), COPR5 (C. odorata segar + tepung
putak 10% + isi rumen 5%), dan COPR10 (C. odorata segar + tepung putak 10% + isi rumen 10%). Rancangan percobaan yang digunakan adalah Rancangan Acak Lengkap pola Faktorial 4 x 2. Faktor pertama adalah perlakuan (CO, COP, COPR4, COPR10) dan faktor kedua adalah waktu fermentasi silase (0 hari dan 21 hari). Masing-masing perlakuan diulang sebanyak 4 kali. Peubah yang diukur adalah kualitas fisik silase, kandungan konsentrasi total tanin, aktivitas trypsin inhibitor, kandungan nutrien, konsentrasi NH3 dan VFA, serta kecernaan bahan
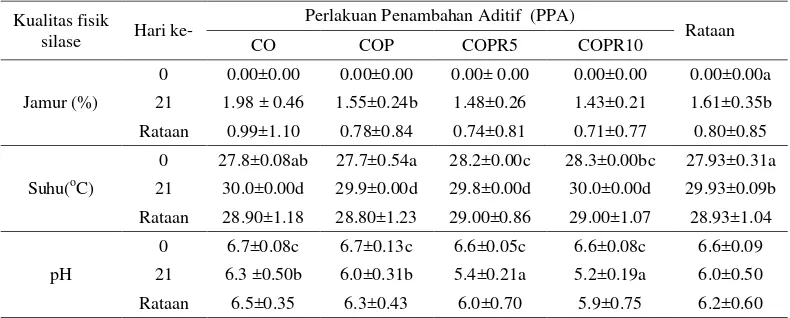
17 dengan penambahan aditif sebesar 5.2 dibanding tanpa aditif yang mencapai pH 6.3. Penurunan tanin dan trypsin inhibitor masing-masing 17.20% dan 15.50%. Kandungan nutrien (BO dan PK) meningkat masing-masing 3.45% dan 41.81% serta terjadi penurunan serat kasar 3% bila dibandingkan silase tanpa aditif. KCBK, KCBO, dan KCPK meningkat pula masing-masing 10.79%, 8.14%, dan 18.45% dibanding silase tanpa penambahan aditif. Waktu fermentasi silase
C.odorata selama 21 hari menghasilkan kualitas silase terbaik dibanding waktu fermentasi 0 hari.
Kata kunci: isi rumen, tepung putak, silase, tanin, trypsin inhibitor
ABSTRACT
The first stage of the study showed that fermentation was the best method to reduce the content of secondary metabolites in C. odorata. Yet, there was a decrease in readily fermentable carbohydrates content. This might related lack due easily digested as a result of the absence of the addition of the additive in the soluble carbohydrates hence lower the activity of fermentative in the fermentation process.The purpose of this study was to examine the effect of the usage of putak
and rumen content as additives in the making of silage C.odorata on the content of secondary metabolites, physical quality silage, nutrient content, quality and digestibility of rumen fermentation in vitro. The treatments used were: CO (C. odorata fresh), COP (C. odorata fresh + 10% putak), COPR5 (C. odorata fresh + 10% putak + 5% rumen content), and COPR10 (C. odorata fresh + 10% putak + 10% rumen content). The experimental design employed was completely randomized design, with 4 x 2 factorial pattern. The first factor was the treatment (CO, COP, COPR5, COPR10) and the second factor was the silage fermentation time (0 days and 21 days). Each treatment was repeated 4 times. The parameters measured were physical quality silage, concentration of total tannin, trypsin inhibitor activity, nutrient content, concentrations of NH3 and VFA, and in vitro
dry matter digestibility (IVDMD), in vitro organic matter digestibility (IVOMD) and in vitro crude protein digestibility (IVCPD). The results showed that silage with additive rate of 10% putak and 10% rumen content produced better silage quality than silage without additives. The percentage of fungi was only 1.42% in silage with additive compared to 1.97% without additives. pH of silage with addition of additives was 5.2, and without additives was 6.3. Tannin and
trypsin inhibitors were reduced by 17.20% and 15.50% respectively. Organic matter and crude protein content increased byr 3.45% and 41.81% respectively. Crude fiber decline by 3% compared to silage without additives. The IVDMD, IVOMD, and IVCPD were also increased by 10.79%, 8.14% and 18.45% respectively. Fermentation for 21 days produced the best quality of silage compared to unfermentation ones (0 day).
18
PENDAHULUAN
Penelitian pertama menunjukkan bahwa dari delapan perlakuan yang diuji (segar, jemur, oven, rebus, rendam air, rendam NaOH, rendam HCl, dan fermentasi), semuanya mampu menurunkan metabolit sekunder C.odorata, di mana perlakuan fermentasi merupakan metode yang paling efektif dalam menurunkan kandungan tanin dan trypsin inhibitor C. odorata. Data-data tersebut menjelaskan bahwa teknik fermentasi dapat digunakan untuk menurunkan efek negatif dari senyawa anti nutrisi dalam Chromolaena odorata. Namun, timbul pertanyaan lain yaitu apakah pemberian sumber karbohidrat seperti halnya dalam proses pembuatan silage akan meningkatkan lagi nilai manfaat Chromolaena odorata? Pertanyaan ini timbul karena dua alasan. Pertama, dalam penelitian pertama tidak ditambahkan aditif. Kedua, kandungan protein kasar C. odorata
cukup tinggi sehingga pembuatan fermentasi mungkin akan berakhir dengan pembusukan karena protein cenderung meningkatkan pH sehingga aktivitas mikroba asam laktat akan terhambat.
Putak merupakan salah satu bahan sumber karbohidrat lokal yang sudah umum dikenal masyarakat di Pulau Timor, Nusa Tenggara Timur. Putak diperoleh dari bagian tengah (isi) batang pohon gewang (Corypha elata robx) (Hilakore et al. 2013). Tepung putak mengandung bahan organik 95.17%, protein kasar 9.79%, serat kasar 5.39%, lemak kasar 0.84%, dan BETN 79.15%. Tepung putak dapat dimanfaatkan sebagai bahan sumber karbohidrat yang mudah dan murah dalam proses ensilase.
Penambahan enzim pun sangat diperlukan sehingga memaksimal kerja bakteri selama ensilase. Isi rumen sapi merupakan salah satu limbah rumah potong hewan yang belum termanfaatkan. Isi rumen adalah bahan pakan yang terdapat dalam rumen yang belum menjadi feses, namun sudah dikeluarkan dari dalam rumen karena ternak dipotong. Kandungan nutrisi isi rumen tidak berbeda dengan kandungan nutrisi bahan baku sebagai akibat dari belum tercernanya bahan pakan secara sempurna dan mengandung mikroba yang berpotensi memperbaiki kualitas pakan. Isi rumen dapat berfungsi sebagai sumber enzim menggantikan enzim komersial untuk mengatasi kualitas pakan yang rendah (Budiansyah et al. 2010) sehingga dapat dimanfaatkan sebagai sumber mikroba dalam proses ensilase. Soepranianondo (2005) melaporkan bahwa protein kasar isi rumen sapi 9.13%, dan serat kasar 34.68%. Tujuan penggunaan isi rumen adalah meningkatkan populasi bakteri agar proses fermentasi dapat berjalan dengan cepat dan memacu terbentuknya suasana asam.
Penambahan aditif putak dan isi rumen bertujuan untuk mencukupi kebutuhan karbohidrat mudah larut dan memaksimalkan kerja bakteri selama proses fermentasi sehingga memacu terbentuknya suasana asam. Liukae (2007) melaporkan bahwa penambahan putak sebagai aditif dalam silase C.odorata
sebanyak 10% sangat optimal mempertahankan kandungan energi dilihat dari kandungan bahan organik dan kandungan proteinnya. Kajian mengenai penambahan isi rumen sapi dalam proses pembuatan silase C. odorata belum pernah dilakukan sehingga perlu dilakukan uji level isi rumen terbaik dalam silase
19 nutrisi dan fermentatif serta kandungan tanin dan trypsin inhibitor silase C. odorata.
METODE
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan dalam dua tahapan, yaitu tahap pembuatan silase dan tahapan analisis kimia (kualitas fisik, proksimat, tanin, trypsin inhibitor,
dan analisis in vitro). Tahap pembuatan silase dilaksanakan di Kupang-Nusa Tenggara Timur (NTT) dari bulan Maret-Mei 2015 di Laboratorium Mikrobiologi, Fakultas Peternakan UNDANA. Analisis kimia dilaksanakan bulan Juni-Desember 2015. Evaluasi tanin dan komposisi proksimat dilaksanakan di Laboratorium Ilmu dan Teknologi Pakan, Fakultas Peternakan IPB, sedangkan evaluasi trypsin inhibitor dan kecernaan in vitro dilaksanakan di Laboratorium Nutrisi dan Ternak Perah, Fakultas Peternakan IPB.
Materi
Bahan yang digunakan dalam penelitian ini meliputi tanaman C. odorata
yang dipanen bulan Maret 2015 di padang penggembalaan yang ada di NTT dan tepung putak. Isi rumen sapi diperoleh dari rumah potong hewan Kotamadya Kupang-NTT. Cairan rumen yang digunakan dalam analisis in vitro diperoleh dari sapi fistula milik LIPI Cibinong. Alat yang digunakan meliputi silo dari botol plastik kapasitas 1 liter, pH meter, thermometer suhu, serta alat lainnya untuk penentuan nilai nutrien dan kecernaan in vitro.
Perlakuan, Rancangan Percobaan dan Analisis Data
Rancangan percobaan yang digunakan adalah Rancangan Acak Lengkap Faktorial (RALF) dengan konfigurasi 4x2, 4 ulangan. Dimana ada 4 perlakuan penambahan aditif (CO, COP, COPR5, COPR10) dan 2 waktu fermentasi silase yang berbeda (0 hari dan 21 hari). Model matematisnya sebagai berikut:
Yijk= μ + αi +βj+ (αβ)ij + εijk
Keterangan :
Yijk = nilai pengamatan perlakuan ke-i dan perlakuan ke-j pada ulangan ke-k
μ = rataan umum
αi = pengaruh faktor perlakuan ke-i
βj =pengaruh faktor perlakuan ke-j
(αβ)ij = interaksiperlakuan ke-i dan perlakuan ke-j
εijk = eror perlakuan ke-i dan perlakuan ke-j pada ulangan ke-k
Rancangan perlakuan yang dicobakan sebagai berikut:
CO = C. odorata segar,
COP = C. odorata segar + putak 10%,