KAJIAN MORFOLOGI DAN GENETIK MIMI
(XIPHOSURA, LIMULIDAE) SEBAGAI DASAR
KONSERVASI DAN PENGELOLAAN DI PULAU JAWA
LUSITA MEILANA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul “Kajian Morfologi dan Genetik Mimi (Xiphosura, Limulidae) Sebagai Dasar Konservasi dan Pengelolaan di Pulau Jawa” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
LUSITA MEILANA.Kajian Morfologi dan Genetik Mimi (Xiphosura, Limulidae) Sebagai Dasar Konservasi dan Pengelolaan di Pulau Jawa.Dibimbing oleh YUSLI WARDIATNO, NURLISA ALIAS BUTET, dan MAJARIANA KRISANTI.
Mimi atau belangkas merupakan hewan dari famili Limulidae yang dikenal sebagai living fossils dan di Indonesia merupakan salah satu sumberdaya genetik yang dilindungi oleh SK Menteri Kehutanan No. 12/Kpts-II/1987 dan Peraturan Pemerintah RI No. 7/1999. Keberadaan ketiga spesies Asia yaitu Tachypleus gigas (Muller 1785), Tachypleus tridentatus (Leach 1819), dan Carcinoscorpius rotundicauda (Lattreille 1802) telah dinyatakan near threatened (2010), threatened (2014) dan data deficient (2015) oleh IUCN. Kajian mengenai mimi di Indonesia sangat jarang dilakukan dan belum diketahui dengan jelas status populasinya.
Penelitian dilaksanakan pada bulan Oktober 2014 sampai Juni 2015. Penelitian menggunakan metode random sampling, mimi yang tertangkap jaring dibawa ke daratan dan diukur secara hidup-hidup, pengukuran karakteristik morfologi berupa karakter morfometrik yang meliputi panjang (total, telson, badan, prosoma, median ringe, depan occeli, marginal spine (I, II, III, IV, V, VI), ophistoma), tebal ventral messel, lebar maksimum prosoma, jarak antar (mata majemuk, auriculata spine, marginal proses, sudut posterior, sudut anal), lebar dan tinggi pertengahan telson, diameter (capit chelicera, capit pedipalpi, capit kaki jalan (I, II, III, IV)). Sedangkan analisis molekuler meliputi beberapa tahapan diantaranya yaitu isolasi dan ekstraksi DNA, elektroforesis DNA, amplifikasi dan visualisasi fragmen DNA, sekuensing DNA, dan PCR-RFLP.
Penelitian ini bertujuan untuk 1) mengkaji karakteristik morfologi mimi dari pesisir Ujung Kulon Banten, Segara Menyan Subang, Semarang, Demak, Rembang, Pantai Kenjeran Surabaya, dan Campur Rejo Gresik, 2) mengidentifikasi mimi berdasarkan marka gen COI dan mengetahui haplotipe dengan menggunakan metode PCR-RFLP di Pulau Jawa (Banten, Segara Menyan Subang, Semarang, Demak, Rembang, dan Surabaya) sebagai informasi dalam penentuan strategi pengelolaan dan konservasi mimi di pesisir Pulau Jawa. Penelitian dilaksanakan pada bulan Oktober 2014 sampai Juli 2015 di lokasi sampling dan Laboratorium Biologi Molekuler MSP IPB FPIK.
Secara morfologi, C. rotundicauda Subang memiliki ukuran tubuh lebih kecil, sedangkan Rembang memiliki ukuran rata-rata lebar maksimal prosoma paling besar. T. gigas Surabaya memiliki ukuran tubuh lebih besar. T. tridentatus Subang memiliki ukuran tubuh lebih kecil, Gresik memiliki ukuran tubuh lebih besar. Ukuran tubuh mimi secara berurutan dari yang paling besar T. gigas> T. tridentatus > C. rotundicauda. Subang merupakan lokasi terbanyak ditemukannya mimi (58 individu), serta memiliki ketiga spesies. Sedangkan Rembang merupakan daerah terbanyak ditemukannya spesies C. rotundicauda (21 individu).
spesies mimi. Penggunaan enzim restriksi EcoR1 dapat memberikan hasil pemotongan pita DNA hasil PCR-RFLP.
Terjadi ketidaksinkronan antara hasil molekuler dengan morfologi. Secara morfologi teridentifikasi tiga spesies sedangkan secara molekuler T. tridentatus identik dengan T. gigas dan C. rotundicauda perlu dilakukan analisis ulang. Berdasarkan hasil RFLP diperoleh Demak dapat ditentukan sebagai daerah gen stok untuk C. rotundicauda, Subang sebagai gen stok T. gigas, dan Semarang untuk T. tridentatus karena memiliki keanekaragaman haplotipe lebih tinggi dibandingkan dengan lokasi lain.
SUMMARY
LUSITA MEILANA. Assessment of Horseshoe Crabs (Xiphosura, Limulidae) Morphology and Genetic as Basis for Conservation and Management in the Java Island Coast. Supervised byYUSLI WARDIATNO, NURLISA ALIAS BUTET, and MAJARIANA KRISANTI.
Horseshoe crabs are aquatic biota from the Limulidae family known as living fossils and in Indonesia as one of the genetic resources that are protected by the Forestry Ministerial Decree No. 12/Kpts-II/1987 and Government Regulation No. 7/1999. The existence of three Asian species i.e. Tachypleus gigas (Muller 1785), Tachypleus tridentatus (Leach 1819), and Carcinoscorpius rotundicauda (Lattreille 1802) have been declared as near threatened (2010), threatened (2014) and data deficient (2015) by IUCN. The study of Indonesian’s Horseshoe crabs is few and unclear status of the population.
The research was conducted in October 2014 until June 2015. The study was used random sampling methods, horseshoe crabs that caught by nets brought ashore and alive measured. The measurement of the morphological characteristics such as the length (total, telson, body, prosoma, median ringe, front occeli, marginal spine (I, II, III, IV, V, VI), ophistoma), the thick of ventral messel, the maximum width of prosoma, the distance between (a compound eye, auriculata spine, marginal process, posterior edges, corners anal), width and height of the mid-telson, diameter (chelicera claw, claw pedipalpi, claw foot path (I, II, III, IV)). While, the several stages of molecular analysis including the DNA isolation and extraction, DNA electrophoresis, amplification and visualization of DNA fragments, DNA sequencing, and PCR-RFLP.
The research were aimed at assessing the horseshoe crabs morphological characters (Ujung Kulon Banten, Segara Menyan Subang, Semarang, Demak, Rembang, Kenjeran Surabaya, and Campur Rejo Gresik Coast) and identification based on molecular marker of COI gene (Ujung Kulon Banten, Segara Menyan Subang, Semarang, Demak, Rembang, and Kenjeran Surabaya). The research was conducted on October 2014 until July 2015 at the field and Laboratory of Molecular Biology MSP FPIK IPB.
Morphologically, C. rotundicauda Subang had a smaller body size, while Rembang had a largest prosoma maximum width. T. gigas Surabaya had bigger body size. T. tridentatus Subang had a smaller body size, Gresik had bigger body size. Horseshoe crabs body size in order from greatest to the small T. gigas> T. tridentatus> C. rotundicauda. Subang was the highest location of the horseshoe crabs discovery (58) and had three species, Demak also. Rembang was the largest area of the C. rotundicauda discovery (21).
Molecular identification of T. gigas from this study have shown taxonomy certainty. Identification of the T. tridentatus and C. rotundicauda did not succeed by using CO1 gene. T. tridentatus was identical with T. gigas, while C. rotundicauda need to be re-analyzed. PCR-RFLP using primer F1 and F2 could applied to three species of horseshoe crabs. The use of restriction enzymes EcoR1 could give results DNA ribbon cutting.
rotundicauda need to be re-analyzed. Based on the RFLP results obtained that Demak could be defined as a region gene stock for C. rotundicauda, Subang as the gene stock of T. gigas, and Semarang for T. tridentatus because they had a higher haplotype diversity compared to other location.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pengelolaan Sumberdaya Perairan
KAJIAN MORFOLOGI DAN GENETIK MIMI
(XIPHOSURA, LIMULIDAE) SEBAGAI DASAR
KONSERVASI DAN PENGELOLAAN DI PULAU JAWA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
PRAKATA
Puji syukur kehadirat Allah SWT berkat rahmat dan karunia-Nya, penulis dapat menyelesaikan tesis yang berjudul “Kajian Morfologi dan Genetik Mimi (Xiphosura, Limulidae) Sebagai Dasar Konservasi dan Pengelolaan di Pulau Jawa”. Tesis ini merupakan hasil penelitian yang dilaksanakan pada bulan Oktober 2014 sampai Juni 2015 dan berlokasi di pesisir Pulau Jawa. Terima kasih penulis ucapkan kepada :
1 Institut Pertanian Bogor yang telah memberikan kesempatan studi kepada penulis.
2 Dr Ir Yusli Wardiatno, MSc; Dr Ir Nurlisa A Butet, MSc; dan Dr Majariana Krisanti, SPi MSi selaku dosen pembimbing yang telah banyak memberikan bimbingan, saran, dan arahan selama pelaksanaan penelitian dan penyusunan tesis.
3 Dr Ir Isdradjad Setyobudiandi, MSc selaku penguji tamu dan Dr Ir Sigid Hariyadi, MSc selaku ketua program studi Pengelolaan Sumberdaya Perairan atas saran dan masukan yang berarti.
4 Keluarga penulis, Bapak Sutiyo, Ibu Dwi Setiya Wati, kakak Novita Sastrawati, serta keluarga besar penulis di Lampung, Jakarta, dan Palembang yang telah memberikan motivasi moril serta materil untuk menyelesaikan studi.
5 Dr Ir Yusli Wardiatno, MSc dan PPLH yang telah memberikan bantuan dana penelitian.
6 Laboratorium Biologi Mikro 1, Laboratorium Biologi Makro, Laboratorium Biologi Molekuler Departemen Manajemen Sumberdaya Perairan, dan Laboratorium terpadu FPIK, Institut Pertanian Bogor.
7 Lukman Guam Hakim, Agus Alim Hakim, Febi Ayu Pramithasari, Yuyun Qonita, Khoirul Marom, Rizal Zakaria, Mudhofar Susanto, dan para nelayan atas bantuan saat penelitian di lapangan. Rana, Rinrin, dan teman-teman MSP 47 lainnya yang tidak mungkin disebutkan satu persatu.
8 Kak Panji, kak Wahyu, bang Reiza, mbak Yanti, mbak Mega, bang Samsul, mas Arif, mbk Fajrin, mbak Lita, mbak Yustin, mbak Lela, kk Findra, yang telah banyak membantu dalam menyelesaikan penelitian.
9 Teman-teman Fast track MSP 2014 yang telah membantu penulis dalam menyelesaikan dan penyusunan tesis.
10 Teman-teman SDP 2013 yang telah memberikan semangat dan dukungan kepada penulis.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vii
1. PENDAHULUAN
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 4
Manfaat Penelitian 4
2. KEANEKARAGAMAN MORFOLOGI MIMI YANG DITEMUKAN DI PESISIR PULAU JAWA
Abstract 5
Pendahuluan 5
Metode 7
Hasil dan Pembahasan 11
Simpulan dan Saran 22
Daftar Pustaka 23
3. IDENTIFIKASI MIMI YANG TERTANGKAP DI PERAIRAN PESISIR PULAU JAWA BERDASARKAN MARKA GEN CO1
Abstract 26
Pendahuluan 26
Metode 28
Hasil dan Pembahasan 30
Simpulan dan Saran 38
Daftar Pustaka 38
4. PEMBAHASAN UMUM 41
5. SIMPULAN DAN SARAN 44
DAFTAR PUSTAKA 44
LAMPIRAN 49
DAFTAR TABEL
1. Rincian jumlah mimi yang tertangkap selama penelitian 12 2. Perbandingan karakter morfometrik Carcinoscorpius rotundicauda pada
lokasi Semarang, Demak, Rembang, Gresik, Surabaya, dan Subang 15 3. Konektivitas antar lokasi Carcinoscorpius rotundicauda berdasarkan 27
karakter yang telah dirasiokan 16
4. Perbandingan karakter morfometrik Tachypleus gigas pada lokasi Semarang, Demak, Surabaya, Subang, dan Banten 17 5. Konektivitas antar lokasi Tachypleus gigas berdasarkan 27 karakter yang
telah dirasiokan 18
6. Perbandingan karakter morfometrik Tachypleus tridentatus pada lokasi
Semarang, Gresik, Subang, dan Banten 19
7. Konektivitas antar lokasi Tachypleus tridentatus berdasarkan 27 karakter
yang telah dirasiokan 21
8. Matriks jarak genetik fragmen gen CO1 pada tiga spesies mimi (Carcinoscorpius rotundicauda, Tachypleus tridentatus, dan T. gigas) dari famili Limulidae dari lokasi Banten, Subang, Semarang, Demak,
Rembang, dan Surabaya. 32
9. Keragaman haplotipe pada tiga spesies mimi (Carcinoscorpius rotundicauda, Tachypleus tridentatus, dan T. gigas)dari Banten, Subang,
Semarang, Demak, Rembang, dan Surabaya 33
10. Haplotipe ketiga spesies mimi (Carcinoscorpius rotundicauda, Tachypleus tridentatus, dan T. gigas) 35
DAFTAR GAMBAR
1. Mimi yang menjadi bycatch nelayan rajungan dan mimi merusak jaring
nelayan di Segara Menyam Subang 3
2. Diagram alir perumusan masalah kajian morfologi dan genetik mimi (Xiphosura, Limulidae) sebagai dasar konservasi dan pengelolaan di
Pulau Jawa 4
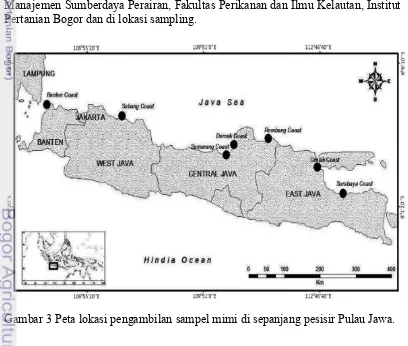
3. Peta lokasi pengambilan sampel mimi di sepanjang pesisir Pulau Jawa 7 4. Ketiga spesies mimi secara berurutan dari kiri ke kanan Tachypleus
tridentatus, T. gigas, dan Carcinoscorpius rotundicauda 8 5. Pengambilan sampel mimi dengan menggunakan jaring nelayan 8 6. Karakter morfometrik mimi yang diukur pada penelitian ini 9 7. Perbedaan antara mimi jantan (a) dan betina (b) berdasarkan ciri
morfologi 9
8. Perbedaan telson pada genus Carcinoscorpius (a) dan genus Tachypleus
(b) 10
9. Perbedaan warna pada Tachypleus tridentatus (kiri) dan T. gigas (kanan) 10 10. Duri badan Tachypleus tridentatus (a) dan T. gigas (b) 11 11. Sebaran spesies Tachypleus gigas, T. tridentatus, dan Carcinoscorpius
rotundicauda di pesisir Banten, Subang, Semarang, Demak, Rembang,
12. Variasi geografis ukuran tubuh mimi, diukur pada lebar maksimal
prosoma ke tiga spesies 13
13. Variasi geografis ukuran tubuh mimi, diukur pada panjang Badan ke tiga
spesies 13
14. Konstruksi pohon fenetik spesies Tachypleus gigas, T. tridentatus, dan Carcinoscorpius rotundicauda berdasarkan 14 karakter morfologi 14 15. Dendrogram karakter morfometrik Carcinoscorpius rotundicauda pada
daerah Semarang, Demak, Rembang, Gresik, Surabaya, dan Subang 16 16. Dendrogram karakter morfometrik Tachypleus gigas pada daerah
Semarang, Demak, Subang, Banten, Surabaya 18
17. Dendrogram karakter morfometrik Tachypleus tridentatus pada daerah
Semarang, Banten, Gresik, dan Surabaya 20
18. Peta lokasi pengambilan mimi di Pulau Jawa 28
19. Visualisasi DNA hasil pre-tes produk PCR pada gel agarosa 0,8 %. Kolom kiri sampai kanan: marker 1 kb, 1) Tt SBG, 2) Cr SBG, 3) Tg SMG, 4) Tt SMG, 5) Tg DMK, 6) Cr DMK, 7) Cr RMG, 8) Tg SRB, 9)
Tg SBG, 10) Tg BTN 31
20. Konstruksi pohon filogeni berdasarkan gen CO1 pada tiga spesies mimi (Carcinoscorpius rotundicauda, Tachypleus tridentatus, dan T. gigas)
dari famili Limulidae 33
21. Hasil digesti fragmen mtDNA oleh enzim restriksi EcoR1. 1= Marker, 2-6= Tg Demak, 7-10= Tg Subang, 11-13= Tg Semarang 34 22. Hasil digesti fragmen mtDNA oleh enzim restriksi EcoR1. 1= Marker,
2-8= Cr Demak, 9-13= Cr Subang 34
23. Hasil digesti fragmen mtDNA oleh enzim restriksi EcoR1. 1= Marker, 2-5= Cr Rembang, 6-10= Tt Subang, 11= Tt Semarang, 12= Tg Surabaya,
13= Tg Banten 34
24. Ilustrasi tipe digesti yang terbentuk dari ke-3 spesies mimi (Carcinoscorpius rotundicauda, Tachypleus tridentatus, dan T. gigas) 35
DAFTAR LAMPIRAN
1. Klasifikasi tiga spesies mimi a) Tachypleustridentatus, b) T. gigas, dan c) Carcinoscorpius rotundicauda menurut Sekiguchi (1998) 49
2. Formula menghitung keanekaragaman haplotipe 49
3. Komposisi basa nukleotida mimi (Tachypleus tridentatus, T. gigas, dan
Carcinoscorpius rotundicauda) 50
1
PENDAHULUAN
Latar Belakang
Mimi atau belangkas merupakan hewan dari famili Limulidae yang dikenal sebagai livingfossils dan phylogeneticrelicts (Selander et al. 1970). Hewan ini telah mengalami diversifikasi sejak zaman paleogene (65-23 Mya) (Obst et al. 2012) dan di Indonesia merupakan salah satu sumberdaya genetik yang dilindungi (SK Menteri Kehutanan No. 12/Kpts-II/1987 dan Peraturan Pemerintah RI No. 7/1999) (Mulya 2004; Rubiyanto 2012). Hingga saat ini, empat spesies mimi yang masih ada di dunia, yaitu Limulus polyphemus (Linnaeus 1758) hanya terdapat di pantai Atlantik Amerika Utara (Walls et al. 2002), dan ke-3 lainnya merupakan spesies Asia yaitu Tachypleus gigas (Muller 1785), Tachypleus tridentatus (Leach 18192), dan Carcinoscorpius rotundicauda (Lattreille 1802) (Christianus & Saad 2007; Lee & Morton 2005). Keberadaan ketiga spesies Asia tersebut dinyatakan near threatened oleh IUCN (2010), threatened (2014) dan data deficient (2015). Sedangkan untuk L. polyphemus adalah lower risk/near threatened berdasarkan IUCN (2015). Klasifikasi ketiga spesies dapat dilihat pada Lampiran 1.
Mimi merupakan hewan yang memiliki peranan penting, baik secara ekonomi maupun ekologi. Secara ekonomi, mimi dimanfaatkan sebagai hewan umpan untuk menangkap ikan sembilang (Euristhmus microceps) di Perairan Kuala Tungkal Jambi (Rubiyanto 2012), belut (Anguilla rostrata), siput besar (Ferari & Targett 2003), dan whelk (Busycon carica dan B. canaliculatum) (Novitsky et al. 2002). Ratusan mimi setiap minggunya ditangkap di Sadeli Kechil dan Johor pantai timur Malaysia dan diekspor ke Thailand (Christianus & Saad 2007). T. tridentatus di Hong Kong menjadi makanan dijual di restoran (Shin et al. 2009), sedangkan di Thailand dan Malaysia, ketiga spesies mimi Asia ini dikonsumsi sebagai makanan eksklusif (Christianus & Saad 2007). Peranan mimi di bidang medis yaitu haemolymph dari C. rotundicauda dapat menetralisir Tetrodotoxin (TTX) (Yeo et al. 1996). Sel darah L. polyphemus digunakan oleh industri medis untuk mensterilkan produk medis dan farmasi (Hurton 2003) karena dalam darahnya mengandung Limulus Amebocyte Lysate (LAL) yang dapat mendeteksi endotoksin pada darah manusia begitu juga dengan genus Tachypleus menghasilkan Tachyplesin Amoebocyt Lysate (TAL) yang dapat mendeteksi endotoksin bakteri gram negatif, mendeteksi endotoksin darah manusia, dan menguji obat bahwa bebas dari bakteri patogen sebelum dikonsumsi oleh manusia (Novitsky 1994). Sel darah C. rotundicauda sebagai imun aktif saat terjadi infeksi (Ding et al. 2005) dan menghasilkan Carcinoscorpius Amoebocyt Lysate (CAL) (Novitsky 1994).
Secara ekologi, mimi memiliki peranan dalam penyeimbang rantai makanan dan sebagai sumber protein bagi setidaknya 20 spesies burung pantai yang bermigrasi (Dietl et al. 2000; Harrington 2001; Beekey et al. 2013). Mimi juga berperan sebagai bioturbator dan mengendalikan hewan bentik invertebrata (Smith 2007; John et al. 2012). Selain itu, mimi juga dikonsumsi oleh monyet mangrove (Macaca fascicularis) (Rubiyanto 2012).
2
et al. 2009). Seperti yang terjadi di India, Hong Kong, dan Singapura (Shin et al. 2009; Taylor et al. 2011). Tahun 1990 populasi mimi Tachypleus gigas di Balramgari India tercatat sebanyak 306 individu yang didapat dalam transek sepanjang 200 meter (Chatterji et al. 1992) dan pada tahun 1996 jumlah populasinya menurun menjadi 220 individu. Begitu pula di Hong Kong, pengamatan pada bulan Mei-Desember 2002 tercatat densitas juvenil T. tridentatus di Pak Nai adalah 197 individu/hadan dua lokasi di Hak Pak Nai adalah 155 individu/ha dan 114 individu/ha(Chiu & Morton 2004) dan pada September 2004-Februari 2005 tercatat densitasnya adalah 8 individu/ha, 16 individu/ha, dan 23 individu/ha (Li 2008). Kategori rendahnya kepadatan mimi juga terjadi di salah satu perairan Indonesia yaitu di Perairan Kuala Tungkal, kepadatan mimi C. rotundicauda dan T. gigas terhitung masing-masing 62 individu/ha dan 2 individu/ha (Rubiyanto 2012).
Penelitian terkait mimi telah banyak dilakukan di berbagai negara, namun di Indonesia penelitian mengenai genetik mimi masih sangat jarang dilakukan. Penelitian mimi di Indonesia diantaranya aspek biologi reproduksi (Fachrul 1989; Eidman 1992; Purnomo 1992; Eidman 1997; Mulya 2004; Muslihah 2004), Perkembangan embrio (Santoso 1992; Balasani 1994; Ismurwanti 1994; Rahmalia 1995; Vauziyah 1995), morfometrik (Suparta 1992), dan studi populasi mimi (Rubiyanto 2012).
Genetika molekuler dibutuhkan terkait dengan ketepatan mengidentifikasi spesies dan mendukung hasil identifikasi berdasarkan sifat morfologi. Identifikasi suatu organisme mulai spesies hingga subspesies secara akurat terhadap berbagai spesies yang sulit dibedakan secara morfologi dapat menggunakan DNA barcoding (Tudge 2000). Salah satu kelompok gen yang dapat dijadikan sebagai marka molekuler untuk penentuan spesies adalah gen Cythocrome Oxsydase subunit I (COI) pada DNA mitokondria (Solihin 1994). COI pada mitokondria merupakan gen yang berevolusi cepat dibandingkan gen 12S rRNA dan 16S rRNA. Gen tersebut memiliki variasi yang sedikit sehingga dapat digunakan sebagai DNA barcoding serta sedikit mengalami delesi dan insersi dalam sekuennya (Hebert et al. 2003).
Perumusan Masalah
3 pengelolaannya. Padahal, di Amerika mimi sangat berperan penting dalam bidang kesehatan dan kuliner. Berdasarkan hasil wawancara pada nelayan mimi hanya by catch yang tertangkap dan biasanya mati terbuang saat pengambilan organisme target (sortir) dari jaring yang dilakukan di darat, mimi juga dianggap sebagai hama yang mengganggu dan merusak jaring nelayan khususnya nelayan rajungan (Gambar 1). Selain itu, pada hari raya untuk mainan anak-anak dalam kondisi hidup-hidup di Pondok Bali tempat wisata (hari-hari tertentu untuk mainan).
Baru baru ini, secara ekonomis mimi di Subang di ekspor ke Jepang dengan agen pembeli dari Medan, namun hanya bulan dan masa pemijahan saja, serta hanya mimi yang berkualitas yaitu mimi dengan ukuran besar, dan betina dengan diameter telur besar. Belum ada peraturan yang mengatur tentang penangkapan mimi ataupun bentuk pengelolaan khusus dari pemerintah lokal hingga saat ini.
Gambar 1 Mimi yang menjadi bycatch nelayan rajungan dan mimi merusak jaring
inelayan di Segara Menyam Subang
Mimi di daerah Subang tidak diperjualbelikan di pasar. Ketidaktertarikan masyarakat terhadap mimi yang memiliki sedikit daging dan sulitnya dalam mengelola menjadi bahan makanan. Dipihak lain, menurut para nelayan mimi di daerah ini telah mengalami penurunan akibat degradasi lingkungan, sekitar tahun 1900-an mimi masih sangat melimpah namun pada tahun 2000-an telah mengalami penurunan yang sangat signifikan. Penurunan ini terjadi bukan karena pemanfaatan secara berlebih, karena pada faktanya mimi tidak begitu dimanfaatkan di daerah ini, tapi lebih disebabkan karena degradasi lingkungan akibat kegiatan antropogenik. Kegiatan ini mampu merubah habitat mimi, misal dari tekstur substrat, kualitas perairan ataupun faktor lainnya.
4
Gambar 2 Diagram alir perumusan masalah kajian morfologi dan genetik mimi (Xiphosura, Limulidae) sebagai dasar konservasi dan pengelolaan di Pulau Jawa
Tujuan Penelitian
Penelitian ini bertujuan untuk 1) mengkaji karakteristik morfologi mimi (dari pesisir Ujung Kulon Banten, Segara Menyan Subang, Semarang, Demak, Rembang, Pantai Kenjeran Surabaya, dan Campur Rejo Gresik) dan 2) mengidentifikasi mimi berdasarkan marka gen COI dan mengetahui haplotipe dengan menggunakan metode PCR-RFLP (dari Ujung Kulon Banten, Segara Menyan Subang, Semarang, Demak, Rembang, dan Pantai Kenjeran Surabaya) sebagai informasi dalam penentuan strategi pengelolaan dan konservasi mimi di pesisir Pulau Jawa. Hasil penelitian ini dibagi menjadi dua makalah untuk mempermudah dalam pemaparan pencapaian tujuan diatas.
Manfaat Penelitian
5
2
KEANEKARAGAMAN MORFOLOGI MIMI YANG
DITEMUKAN DI PESISIR PULAU JAWA
Lusita Meilana, Yusli Wardiatno, Nurlisa A Butet, Majariana Krisanti Program Studi Pengelolaan Sumberdaya Perairan, Fakultas Perikanan dan Ilmu
Kelautan, Institut Pertanian Bogor. Email: lusitameilana@yahoo.co.id
ABSTRACT
Horseshoe crabs are distributed widely in Indonesian waters, and there are three common species inhabiting coastal waters of Java Island. The research was aimed at assessing the horseshoe crabs morphological characters (Ujung Kulon Banten, Segara Menyan Subang, Semarang, Demak, Rembang, Kenjeran Surabaya, and Campur Rejo Gresik Coast). A total of 27 parameters in a ratio used to determine connectivity and character identifier. Data were tested using non-parametric analysis. C. rotundicauda Subang had a smaller body size, while Rembang had a largest prosoma maximum width. T. gigas Surabaya had bigger body size. T. tridentatus Subang had a smaller body size, Gresik had bigger body size. Horseshoe crabs body size in order from greatest to the small T. gigas> T. tridentatus> C. rotundicauda. Subang was the highest location of the horseshoe crabs discovery (58) and had three species, Demak also. Rembang was the largest area of the C. rotundicauda discovery (21). The results obtained based on morphological characters that individual was classified as an original individual from each location.
Keywords: horseshoe crabs, Java Island, morphology
PENDAHULUAN
6
Mimi merupakan hewan yang memiliki peranan penting, baik secara ekonomi maupun ekologi. Secara ekonomi, mimi dimanfaatkan sebagai hewan umpan untuk menangkap ikan sembilang (Euristhmus microceps) di Perairan Kuala Tungkal Jambi (Rubiyanto 2012), belut (Anguilla rostrata), siput besar (Ferari & Targett 2003), dan whelk (Busycon carica dan B. canaliculatum) (Novitsky et al. 2002). Ratusan mimi setiap minggunya ditangkap di Sadeli Kechil dan Johor pantai timur Malaysia dan diekspor ke Thailand (Christianus & Saad 2007). T. tridentatus di Hong Kong menjadi makanan dijual di restoran (Shin et al. 2009), sedangkan di Thailand dan Malaysia, ketiga spesies mimi Asia ini dikonsumsi sebagai makanan eksklusif (Christianus & Saad 2007). Peranan mimi di bidang medis yaitu haemolymph dari C. rotundicauda dapat menetralisir Tetrodotoxin (TTX) (Yeo et al. 1996). Sel darah L. polyphemus digunakan oleh industri medis untuk mensterilkan produk medis dan farmasi (Hurton 2003) karena dalam darahnya mengandung Limulus Amebocyte Lysate (LAL) yang dapat mendeteksi endotoksin pada darah manusia begitu juga dengan genus Tachypleus menghasilkan Tachyplesin Amoebocyt Lysate (TAL) yang dapat mendeteksi endotoksin bakteri gram negatif, mendeteksi endotoksin darah manusia, dan menguji obat bahwa bebas dari bakteri patogen sebelum dikonsumsi oleh manusia (Novitsky 1994). Sel darah C. rotundicauda sebagai imun aktif saat terjadi infeksi (Ding et al. 2005) dan menghasilkan Carcinoscorpius Amoebocyt Lysate (CAL) (Novitsky 1994).
Secara ekologi, mimi memiliki peranan dalam penyeimbang rantai makanan dan sebagai sumber protein bagi setidaknya 20 spesies burung pantai yang bermigrasi (Dietl et al. 2000; Harrington 2001; Beekey et al. 2013). Mimi juga berperan sebagai bioturbator dan mengendalikan hewan bentik invertebrata (Smith 2007; John et al. 2012). Selain itu, mimi juga dikonsumsi oleh monyet mangrove (Macaca fascicularis) (Rubiyanto 2012).
Mimi mempunyai risiko kepunahan yang tinggi akibat adanya degradasi habitat, reklamasi, pencemaran, dan perburuan komersial (Mishra 2009), hilangnya habitat dan sumber makanan, perubahan kondisi air, serta peningkatan predasi (Hu et al. 2009). Seperti yang terjadi di India, Hong Kong, dan Singapura (Shin et al. 2009; Taylor et al. 2011). Tahun 1990 populasi mimi Tachypleus gigas di Balramgari India tercatat sebanyak 306 individu yang didapat dalam transek sepanjang 200 meter (Chatterji et al. 1992) dan pada tahun 1996 jumlah populasinya menurun menjadi 220 individu. Begitu pula di Hong Kong, pengamatan pada bulan Mei-Desember 2002 tercatat densitas juvenil T. tridentatus di Pak Nai adalah 197 individu/hadan dua lokasi di Hak Pak Nai adalah 155 individu/ha dan 114 individu/ha(Chiu & Morton 2004) dan pada September 2004-Februari 2005 tercatat densitasnya adalah 8 individu/ha, 16 individu/ha, dan 23 individu/ha (Li 2008). Kategori rendahnya kepadatan mimi juga terjadi di salah satu perairan Indonesia yaitu di Perairan Kuala Tungkal, kepadatan mimi C. rotundicauda dan T. gigas terhitung masing-masing 62 individu/ha dan 2 individu/ha (Rubiyanto 2012).
7 pada masing-masing habitat. Berdasarkan penelitian-penelitian terkait mimi yang telah dilakukan diperoleh kesimpulan bahwa belum adanya kejelasan mengenai status populasi mimi yang ada di Indonesia, sehingga perlu dilakukan penelitian awal yang mendasar untuk mengawali kajian populasi yaitu dengan melihat karakter mimi yang ada di Indonesia. Oleh karena itu penelitian ini akan dilakukan pengukuran terhadap beberapa karakter mimi pada lokasi yang berbeda yaitu Ujung Kulon Banten, Segara Menyan Subang, Semarang, Demak, Rembang, Pantai Kenjeran Surabaya, dan Campur Rejo Gresik untuk melihat variasi karakter morfologi mimi pada suatu populasi.
METODE
Waktu dan Lokasi Penelitian
Penelitian dilaksanakan pada bulan Oktober 2014 sampai Juni 2015 dengan pengambilan sampel pada enam lokasi yaitu Ujung Kulon Banten, Segara Menyan Subang, Semarang, Demak, Rembang, Pantai Kenjeran Surabaya, dan Campur Rejo Gresik sebagi perwakilan sepanjang pantai Pulau Jawa (Gambar 3). Analisis morfometrik dilaksanakan di Laboratorium Biologi Makro 1 Departemen Manajemen Sumberdaya Perairan, Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor dan di lokasi sampling.
8
Alat dan Bahan
Alat dan bahan yang diperlukan dalam penelitian ini yaitu kapal nelayan dan jaring udang, penggaris, caliper, sheet, timbangan digital, gunting, kamera, meteran, dan ketiga spesies mimi yang ditunjukkan oleh Gambar 4.
Gambar 4 Ketiga spesies mimi secara berurutan dari kiri ke kanan Tachypleus tridentatus, T. gigas, dan Carcinoscorpius rotundicauda
Cara Kerja
Penelitian menggunakan metode random sampling, contoh mimi diambil dengan menggunakan jaring yang dijalankan oleh kapal nelayan atau langsung diambil dengan tangan (Gambar 5). Mimi yang tertangkap jaring dibawa ke daratan dan diukur secara hidup-hidup, pengukuran karakteristik morfologi berupa karakter morfometrik yang meliputi panjang (total, telson, badan, prosoma, median ringe, depan occeli, marginal spine (I, II, III, IV, V, VI), ophistoma), tebal ventral messel, lebar maksimum prosoma, jarak antar (mata majemuk, auriculata spine, marginal proses, sudut posterior, sudut anal), lebar dan tinggi pertengahan telson, diameter (capit chelicera, capit pedipalpi, capit kaki jalan (I, II, III, IV)). Identifikasi berdasarkan Sekiguchi & Shuster (2009) dalam buku berjudul Biology and Conservation of Horseshoe Crabs yang membedakan mimi dari morfologi telson serta duri pada ophistoma.
9
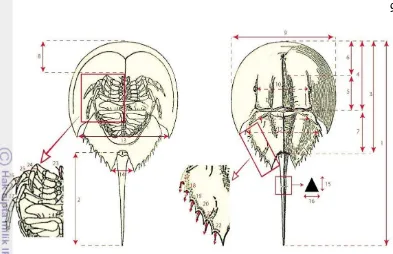
Gambar 6 Karakter morfometrik mimi yang diukur pada penelitian ini Karakter morfometrik mimi yang telah diukur (Gambar 6) diantaranya yaitu XX1 merupakan karakter ukuran tubuh (1. Panjang total (X1), 2. Panjang telson (X2), 3. Panjang badan (X3), 4. Panjang prosoma (X4), 5. Panjang median ridge (X5), 6. Panjang depan occeli (X6), 7. Panjang ophistoma (X7), 8. Tebal ventral messel (X8), 9. Lebar maksimum prosoma (X9), 10. Jarak antar mata majemuk (X10), 11. Jarak antar auriculata spine (X11), 12. Jarak antar marginal process (X12), 13. Jarak antar sudut posterior (X13), 14. Jarak antar sudut anal (X14), 15. Tinggi pertengahan telson (X15), 16. Lebar pertengahan telson) (X16). XX2 merupakan karakter ukuran anggota tubuh (17. Panjang marginal spine 1 (X17), 18. Panjang marginal spine II (X18), 19. Panjang marginal spine III (X19), 20. Panjang marginal spine IV (X20), 21. Panjang marginal spine V (X21), 22. Panjang marginal spine VI (X22), 23. Diameter capit chelicera (X23), 24. Diameter capit pedipalpi (X24), 25. Diameter capit kaki jalan I (X25), 26. Diameter capit kaki jalan II (X26), 27. Diameter capit kaki jalan III (X27), 28. Diameter capit kaki jalan IV (X28)). Setelah dilakukan pengukuran mimi kemudian dilepaskan kembali.
Berikut merupakan ciri yang membedakan antara mimi jantan dan betina yang ditunjukkan oleh Gambar 7:
(a) (b)
10
Secara morfologi mimi dapat diidentifikasi melalui bentuk telson, warna, dan duri ophistoma. Telson pada genus Carcinoscorpius (gambar 8a) tidak berduri sedangkan pada genus Tachypleus (gambar 8b) berduri.
(a) (b)
Gambar 8 Perbedaan telson pada genus Carcinoscorpius (a) dan genus Tachypleus (b)
T. gigas dan T. tridentatus dapat dibedakan dari warna dan duri ophistoma, T. gigas berwarna coklat kehitaman sedangkan T. tridentatus memiliki warna coklat kehijauan (Gambar 9).
11
(a) (b)
Gambar 10 Duri badan Tachypleus tridentatus (a) dan T. gigas (b)
Analisis Data
Analisis data karakter morfologi diantaranya membentuk konstruksi pohon fenetik berdasarkan 14 karakter morfologi dengan aplikasi PAUP. Sedangkan analisis statistik yang digunakan yaitu uji Kruskal-Wallis untuk komparasi karakter morfometrik di semua wilayah dengan aplikasi SPSS 17, uji Mann-Whitneyuntuk mengetahui karakter penciri bagi masing-masing populasi mimi antar populasi dengan SPSS 17, analisis Multinomiallogistic yaitu untuk mengetahui konektivitas antar lokasi dengan SPSS 16. Analisis kluster untuk melihat kelompok-kelompok yang terbentuk berdasarkan karakter morfometrik yang diukur dengan SPSS 16.
HASIL DAN PEMBAHASAN
Hasil
Jumlah Mimi Tertangkap
12
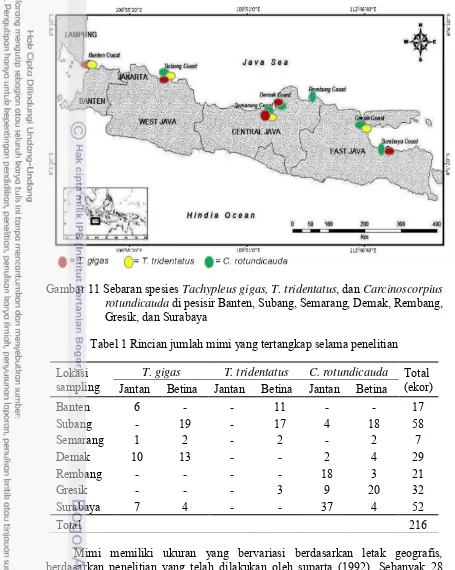
Gambar 11 Sebaran spesies Tachypleus gigas, T. tridentatus, dan Carcinoscorpius rotundicauda di pesisir Banten, Subang, Semarang, Demak, Rembang, Gresik, dan Surabaya
Tabel 1 Rincian jumlah mimi yang tertangkap selama penelitian Lokasi
sampling Jantan Betina Jantan Betina Jantan Betina T. gigas T. tridentatus C. rotundicauda Total (ekor)
Banten 6 - - 11 - - 17
Subang - 19 - 17 4 18 58
Semarang 1 2 - 2 - 2 7
Demak 10 13 - - 2 4 29
Rembang - - - - 18 3 21
Gresik - - - 3 9 20 32
Surabaya 7 4 - - 37 4 52
Total 216
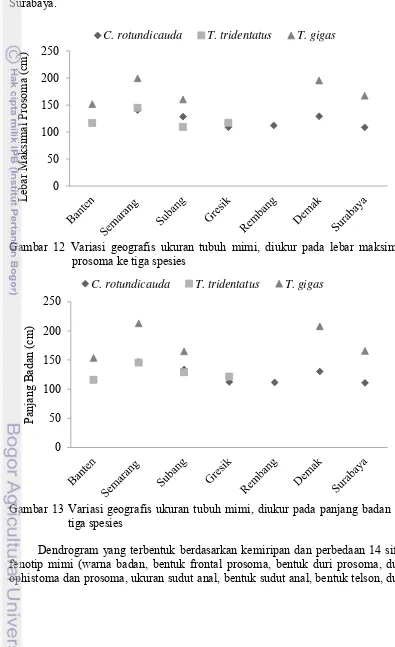
13 gigas yang berasal dari Demak memiliki ukuran hampir sama untuk daerah Banten, Subang, dan Surabaya. T. tridentatus pada daerah Semarang memiliki ukuran lebar maksimal prosoma dan badan paling besar, sedangkan Banten, Subang, dan Gresik memiliki ukuran yang hampir sama. C. rotundicauda Semarang, Subang, dan Demak memiliki ukuran lebih besar dibandingkan dengan Gresik, Rembang, dan Surabaya.
Gambar 12 Variasi geografis ukuran tubuh mimi, diukur pada lebar maksimal prosoma ke tiga spesies
Gambar 13 Variasi geografis ukuran tubuh mimi, diukur pada panjang badan ke tiga spesies
Dendrogram yang terbentuk berdasarkan kemiripan dan perbedaan 14 sifat fenotip mimi (warna badan, bentuk frontal prosoma, bentuk duri prosoma, duri ophistoma dan prosoma, ukuran sudut anal, bentuk sudut anal, bentuk telson, duri
14
marginal, gerigi pada telson, bagian depan operculum, pedipalp 1, pedipalp 2, ukuran duri marginal, dan tebal lapis cangkang) didapatkan informasi bahwa T. gigas dan T. tridentatus memiliki karakter yang hampir sama, sedangkan C. rotundicauda memiliki karakter tersendiri.
Gambar 14 Konstruksi pohon fenetik spesies Tachypleus gigas, T. tridentatus, dan
iCarcinoscorpius rotundicauda berdasarkan 14 karakter morfologi Carcinoscorpius rotundicauda
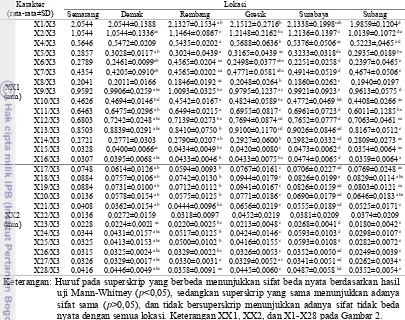
Hasil uji Kruskal Wallis berdasarkan 27 karakter morfometrik C. rotundicauda yang telah dirasiokan secara keseluruhan diperoleh hasil sebanyak 1 karakter yang tidak berbeda nyata (p>0,05) pada lokasi Semarang, Demak, Surabaya, Subang, dan Banten. Namun, pada uji ini tidak dapat diketahui secara spesifik karakter mana yang memiliki beda nyata antar lokasi, sehingga dilakukan uji Mann-Whitney untuk mengetahui karakter yang berbeda nyata (p<0,05) antar lokasi. Berikut merupakan hasil uji Mann-Whitney perbandingan karakter morfometrik yang telah dirasiokan (Tabel 2).
15 Tabel 2 Perbandingan karakter morfometrik Carcinoscorpius rotundicauda pada
lokasi Semarang, Demak, Rembang, Gresik, Surabaya, dan Subang
Karakter
(rata-rata±SD) Semarang Demak Rembang Lokasi Gresik Surabaya Subang
XX1 (mm)
X1/X3 2,0544 2,0544±0,1388 2,1327±0,1534 ab 2,1512±0,2716b 2,1338±0,1998cab 1,9859±0,1204d
X2/X3 1,0544 1,0544±0,1336ae 1,1464±0,0867 e 1,2148±0,2162 bc 1,2136±0,1397 c 1,0139±0,1072 da
X4/X3 0,5646 0,5472±0,0209 0,5435±0,0202 a 0,5688±0,0636 b 0,5376±0,0506 ac 0,5223±,0465 ad
X5/X3 0,2857 0,3028±0,0117 ab 0,3024±0,0439 a 0,3165±0,0439 ac 0,3233±0,0318de 0,2935±0,0189 be
X6/X3 0,2789 0,2461±0,0099eb 0,4565±0,0204 ea 0,2498±0,0377 abc 0,2251±0,0258 d 0,2397±0,0465 e
X7/X3 0,4354 0,4205±0,0910eb 0,4565±0,0202 ea 0,4771±0,0581 abc 0,4914±0,0519 d 0,4674±0,0506 e
X8/X3 0,2041 0,2011±0,0166 0,1846±0,0192 ac 0,2048±0,0264 b 0,1860±0,0262 c 0,1940±0,0197
X9/X3 0,9592 0,9906±0,0259 abe 1,0093±0,0325 be 0,9795±0,1237 ac 0,9921±0,0923 e 0,9613±0,0575 d
X22/X3 0,0136 0,0272±0,0159 0,0318±0,0097 0,0452±0,0219 0,0381±0,0209 0,0374±0,0209
X23/X3 0,0228 0,0224±0,0021 ac 0,0220±0,0025 bc 0,0213±0,0048 c 0,0268±0,0041 d 0,0180±0,0042 e Keterangan: Huruf pada superskrip yang berbeda menunjukkan sifat beda nyata berdasarkan hasil
uji Mann-Whitney (p<0,05), sedangkan superskrip yang sama menunjukkan adanya
sifat sama (p>0,05), dan tidak bersuperskrip menunjukkan adanya sifat tidak beda
nyata dengan semua lokasi. Keterangan XX1, XX2, dan X1-X28 pada Gambar 2.
Analisis Kluster Morfometrik Carcinoscorpius rotundicauda
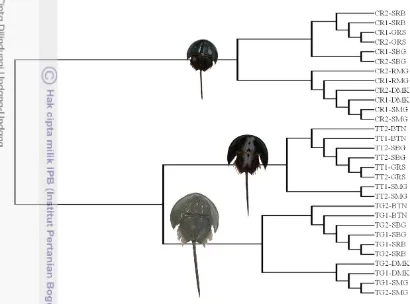
Berikut merupakan dendrogram yang terbentuk dari analisis kluster berdasarkan 27 karakter morfometrik C. rotundicaudayang telah dirasiokan, pada tingkat kesamaan 60% diperoleh hasil bahwa daerah Semarang dan Demak membentuk satu kelompok dengan tingkat kesamaan sebesar 67,24%. Kedua lokasi tersebut memiliki tingkat kesamaan dengan Rembang sebesar 64,68%. Gresik dan Surabaya membentuk satu kelompok dengan tingkat kesamaan sebesar 79,53%. Sedangkan lokasi Subang membentuk kelompok tersendiri.
16
Gambar 15 Dendrogram karakter morfometrik Carcinoscorpius rotundicauda pada daerah Semarang, Demak, Rembang, Gresik, Surabaya, dan Subang
Pengelompokan Carcinoscorpius rotundicauda
Hasil pengklasifikasian C. rotundicauda dengan menggunakan analisis logistik berdasarkan 27 karakter yang telah dirasiokan ditunjukkan oleh Tabel 3. Tabel 3 Konektivitas antar lokasi Carcinoscorpius rotundicauda berdasarkan 27
karakter yang telah dirasiokan
Observed
Predicted
Semarang Demak Rembang Gresik Surabaya Subang PercentCorrect
Semarang 1 100,0%
Demak 0 6 100,0%
Rembang 0 0 21 100,0%
Gresik 0 0 0 28 100,0%
Surabaya 0 0 0 0 38 100,0%
Subang 0 0 0 0 0 21 100,0%
Overall
Percentage 0,9% 5,2% 18,3% 24,3% 33,0% 18,3% 100,0%
17 Tachypleus gigas
Hasil uji Kruskal Wallis berdasarkan 27 karakter morfometrik T. gigas yang telah dirasiokan secara keseluruhan diperoleh hasil sebanyak 13 karakter yang tidak berbeda nyata (p>0,05) pada lokasi Semarang, Demak, Surabaya, Subang, dan Banten. Uji ini tidak dapat diketahui secara spesifik karakter mana yang memiliki beda nyata antar lokasi, sehingga dilakukan uji Mann-Whitney untuk mengetahui karakter yang berbeda nyata (p<0,05) antar lokasi. Berikut merupakan hasil uji Mann-Whitney (Tabel 4).
Tabel 4 Perbandingan karakter morfometrik Tachypleus gigas pada lokasi Semarang, Demak, Surabaya, Subang, dan Banten
Karakter
(rata-rata±SD) Semarang Demak Surabaya Lokasi Subang Banten
XX1 (mm)
X1/X3 1,9974±0,0299 1,9589±0,1597 2,1519±0,3688 1,9685±0,1287 1,9362±0,1920
X2/X3 0,9974±0,0299 0,9561±0,2616a 1,1546±0,2311b 0,9955±0,1438 1,0003±0,2036
X4/X3 0,5751±0,0051 0,5504±0,0636 0,5984±0,0945 0,5590±0,0519 0,5664±0,0080
X5/X3 0,3168±0,0103 0,2973±0,0154b 0,3379±0,0477a 0,3171±0,0232ac 0,3216±0,0276ad
X6/X3 0,2606±0,0019 0,2589±0,0229 0,2715±0,0516 0,2546±0,0220 0,2547±0,0141
X7/X3 0,4249±0,0051bd 0,3554±0,1144b 0,4922±0,0757c 0,4479±0,0233ad 0,4420±0,0199de
X8/X3 0,2137±0,0076 0,2065±0,0144 0,2149±0,0345 0,2072±0,0109 0,2071±0,0053
X9/X3 0,9370±0,0353 0,9440±0,1330 1,0267±0,1536 0,9721±0,0736 0,9898±0,0307
X10/X3 0,5142±0,0136 0,4913±0,0300 0,5430±0,1123 0,4954±0,0233 0,4974±0,0283
X11/X3 0,5329±0,0007 0,5303±0,0230 0,5972±0,1318 0,5399±0,0313 0,5620±0,0477
X12/X3 0,6152±0,0189 0,6047±0,0386a 0,6876±0,1374b 0,6134±0,0384 0,6508±0,0437db
X13/X3 0,7862±0,0142 0,7767±0,0704a 0,8594±0,1460 0,8184±0,0368b 0,8191±0,0358
X14/X3 0,2606±0,0019 0,2537±0,0316a 0,2892±0,0624 0,2777±0,0143b 0,2798±0,0200
X15/X3 0,0429±0,0035abd 0,0421±0,0070b 0,0581±0,0123ce 0,0471±0,0043d 0,0508±0,0034ed
X16/X3 0,0381±0,0011abd 0,0388±0,0055b 0,0516±0,0115ce 0,0432±0,0053d 0,0456±0,0017ed
XX2 (mm)
X17/X3 0,1106±0,0254 0,1092±0,0189 0,1200±0,0341 0,1201±0,0093 0,1269±0,0145
X18/X3 0,1085±0,0486 0,1160±0,0263a 0,1291±0,0292 0,1366±0,0152b 0,1349±0,0198
X19/X3 0,0990±0,0418ab 0,1023±0,0385b 0,1247±0,0512 0,1447±0,0128c 0,1504±0,0295
X20/X3 0,0308±0,0239abc 0,0780±0,0560b 0,1114±0,0637cd 0,1436±0,0118d 0,1519±0,0251ade
X21/X3 0,0378±0,0207ab 0,0718±0,0578b 0,1068±0,0651 0,1456±0,0158cd 0,1475±0,0184ad
X22/X3 0,0308±0,0239abc 0,0700±0,053b 0,1388±0,0134bc 0,1388±0,0134d 0,1322±0,0152cde
X23/X3 0,0249±0,0019 0,0233±0,0026ac 0,0274±0,0062b 0,0226±0,0042c 0,0247±0,0043
X24/X3 0,0357±0,0001abc 0,0443±0,0100b 0,0569±0,0178cd 0,0529±0,0059d 0,0584±0,0033ade
X25/X3 0,0250±0,0133ab 0,0425±0,0103b 0,0561±0,0167cd 0,0522±0,0057d 0,0568±0,0027ade
X26/X3 0,0337±0,0017 0,0328±0,0021ac 0,0376±0,0064bd 0,0317±0,0032c 0,0350±0,0010d
X27/X3 0,0337±0,0018 0,0329±0,0020abc 0,0358±0,0066bd 0,0314±0,0030c 0,0356±0,0015d
X28/X3 0,0456±0,0080 0,0440±0,0028ad 0,0517±0,0114b 0,0388±0,0044c 0,0388±0,0023cd
Keterangan: Huruf pada superskrip yang berbeda menunjukkan sifat beda nyata berdasarkan hasil
uji Mann-Whitney (p<0,05), sedangkan superskrip yang sama menunjukkan adanya
sifat sama (p>0,05), dan tidak bersuperskrip menunjukkan adanya sifat tidak beda nyata
dengan semua lokasi. Keterangan XX1, XX2, dan X1-X28 pada Gambar 2.
Sebanyak 14 karakter morfometrik T. gigas dari ke-5 lokasi tersebut memiliki hubungan sifat beda nyata (p<0,05) antar lokasi. Sebanyak 15 karakter tubuh, T. gigas Surabaya memiliki ukuran tubuh (X1-X2 dan X4-X16) lebih besar dan 5 anggota tubuh (panjang marginal spine VI (X22), diameter capit chelicera (X23), diameter capit kaki jalan II (X26), diameter capit kaki jalan III (X27), diameter capit kaki jalan IV (X28)) yang relatif lebih besar dibandingkan daerah Semarang, Demak, Subang, dan Banten (memiliki 6 anggota tubuh yaitu X17-X21 dan X24-X25), sedangkan T. gigas Semarang dan Demak memiliki ukuran relatif sama, begitu juga dengan Subang dan Banten.
Analisis Kluster Morfometrik Tachypleus gigas
18
kesamaan 60% diperoleh hasil bahwa daerah Semarang dan Demak membentuk satu kelompok dengan tingkat kesamaan sebesar 66,11%. Begitu juga dengan Subang dan Banten memiliki tingkat kesamaan sebesar 79,66%. Sedangkan Surabaya membentuk populasi tersendiri.
Gambar 16 Dendrogram karakter morfometrik Tachypleus gigas pada daerah Semarang, Demak, Subang, Banten, Surabaya
T. gigas Surabaya membentuk kelompok terpisah berdasarkan karakter morfometrik karena pada daerah ini memiliki ukuran tubuh dan anggota tubuh yang relatif lebih besar dibandingkan daerah lainnya. T. gigas Semarang dan Demak memiliki ukuran tubuh dan anggota tubuh relatif sama sehingga keduanya membentuk satu kelompok, begitu juga dengan Subang dan Banten.
Pengelompokan Tachypleus gigas
Hasil pengklasifikasian T. gigas dengan menggunakan analisis logistik berdasarkan 27 karakter yang telah dirasiokan pada Table 5.
Tabel 5 Konektivitas antar lokasi Tachypleus gigas berdasarkan 27 karakter yang telah dirasiokan
Observed Predicted
Semarang Demak Surabaya Subang Banten PercentCorrect
Semarang 2 100,0%
Demak 0 21 100,0%
Surabaya 0 0 11 100,0%
Subang 0 0 0 18 100,0%
Banten 0 0 0 0 5 100,0%
Overall
19
Berdasarkan tabel Pseudo R-Square diperoleh R2 sebesar 93,8%. Hal ini berarti variable independen dapat menjelaskan perbedaan morfometrik antar tiap daerah dan 6,2% dijelaskan oleh variabel di luar model. Berdasarkan tabel classification, model memiliki kemampuan untuk membedakan antar kelompok populasi sebesar 100% yaitu individu diklasifikasikan sebagai individu asli yang berasal dari Semarang (n=2), Demak (n=21), Surabaya (n=11), Subang (18), dan Banten (n=5). Percent correct dapat mencapai hingga 100% diduga karena jumlah individu yang ditemukan sangat sedikit disetiap daerahnya.
Tachypleus tridentatus
Hasil uji Kruskal Wallis berdasarkan 27 karakter morfometrik T. tridentatus yang telah dirasiokan secara keseluruhan diperoleh hasil sebanyak 12 karakter yang tidak berbeda nyata (p>0,05) pada lokasi Semarang, Gresik, Subang, dan Banten. Namun, pada uji ini tidak dapat diketahui secara spesifik karakter mana yang memiliki beda nyata antar lokasi, sehingga dilakukan uji Mann-Whitney untuk mengetahui karakter yang berbeda nyata (p<0,05) antar lokasi. Berikut merupakan hasil uji Mann-Whitney (Tabel 6).
Tabel 6 Perbandingan karakter morfometrik Tachypleus tridentatus pada lokasi
iSemarang, Gresik, Subang, dan Banten
Karakter
(rata-rata±SD) Semarang Gresik Lokasi Subang Banten
XX1 (mm)
X1/X3 2,1407±0,0455 2,0521±0,0538 2,0852±0,2747 2,0755±0,1365
X2/X3 1,1407±0,0455 1,1715±0,0498ac 1,0479±0,1007b 1,1588±0,1370c
X4/X3 0,5770±0,0391 0,5538±0,0017ab 0,5382±0,0357b 0,5840±0,0139c
X5/X3 0,3210±0,0175 0,3144±0,0385a 0,3144±0,0385b 0,3191±0,0120c
X6/X3 0,2560±0,0216 0,2237±0,0498 0,2605±0,0278 0,2627±0,0143
X7/X3 0,4230±0,0391 0,4444±0,0173a 0,3750±0,1003b 0,4655±0,0258
X8/X3 0,2243±0,0117 0,2215±0,0204 0,2192±0,0161 0,2161±0,0128
X9/X3 1,0017±0,0634 0,9566±0,0272a 0,8699±0,2946b 1,0117±0,0443
X10/X3 0,5275±0,0214 0,4910±0,0089ab 0,4962±0,0216b 0,5186±0,0180c
X11/X3 0,5716±0,0139 0,5931±0,0356 0,5604±0,0333 0,5808±0,0172
X12/X3 0,6535±0,0076 0,7158±0,0625a 0,6571±0,0415b 0,6991±0,0442
X13/X3 0,8276±0,0355 0,8292±0,0210a 0,8135±0,0428b 0,8892±0,0594
X14/X3 0,3124±0,0033 0,2697±0,0729 0,3042±0,0216 0,3162±0,0169
X15/X3 0,0582±0,0087 0,0564±0,0091 0,0517±0,0052 0,0554±0,0047
X16/X3 0,0515±0,0086 0,0564±0,0091a 0,0444±0,0060b 0,0482±0,0065
XX2 (mm)
X17/X3 0,1237±0,0004 0,1263±0,0253a 0,1091±0,0164b 0,1419±0,0310
X18/X3 0,1385±0,0125 0,1341±0,0319a 0,1283±0,0226b 0,1571±0,0406
X19/X3 0,1477±0,0006 0,1636±0,0073ac 0,1430±0,0187b 0,1660±0,0376c
X20/X3 0,1508±0,0050 0,1532±0,0174a 0,1449±0,0173b 0,1728±0,0342
X21/X3 0,1346±0,0070ab 0,1549±0,0091 0,1483±0,0126b 0,1682±0,0348c
X22/X3 0,1385±0,0125 0,1410±0,0173 0,1423±0,0104 0,1494±0,0360
X23/X3 0,0256±0,0037 0,0215±0,0034ab 0,0186±0,0049b 0,0263±0,0057c
X24/X3 0,0393±0,0015ab 0,0353±0,0018b 0,0275±0,0059c 0,0353±0,0097bd
X25/X3 0,0378±0,0004ab 0,0390±0,0048b 0,0289±0,0050c 0,0366±0,0025bd
X26/X3 0,0389±0,0012ab 0,0390±0,0048b 0,0285±0,0052c 0,0362±0,0037bd
X27/X3 0,0343±0,0013ac 0,0332±0,0025 0,0277±0,0055b 0,0358±0,0014c
X28/X3 0,0420±0,0031 0,0440±0,0009a 0,0377±0,0054b 0,0410±0,0038
20
Sebanyak 15 karakter morfometrik T. tridentatus dari ke-4 lokasi memiliki hubungan sifat beda nyata (p<0,05) antar lokasi, Subang memiliki ukuran tubuh dan anggota tubuh yang relatif lebih kecil dibandingkan daerah Semarang, Gresik, dan Banten. T. tridentatus Semarang memiliki 5 ukuran tubuh (X1, X5, X8, X10, X15) dan 1 anggota tubuh (X24) lebih besar. Sedangkan T. tridentatus Gresik memiliki 4 ukuran tubuh (X2, X11, X12, X16) dan 3 anggota tubuh (X25, X26, X28) lebih besar. Banten memiliki 6 ukuran tubuh (X4, X6, X7, X9, X13, X14) dan 8 ukuran anggota tubuh (X17-X23, X27) lebih besar.
Analisis Kluster Morfometrik Tachypleus tridentatus
Gambar 17 merupakan dendrogram yang terbentuk dari analisis kluster berdasarkan 27 karakter morfometrik T. tridentatus yang telah dirasiokan, pada tingkat kesamaan 60% diperoleh hasil bahwa daerah Semarang, Banten, Gresik, dan Subang membentuk kelompok tersendiri.
Gambar 17 Dendrogram karakter morfometrik Tachypleus tridentatus pada daerah
iSemarang, Banten, Gresik, dan Surabaya
T. tridentatus Subang membentuk kelompok terpisah berdasarkan karakter morfometrik karena pada daerah ini memiliki ukuran tubuh dan anggota tubuh yang relatif lebih kecil dibandingkan daerah lainnya. T. tridentatus Semarang dan Banten memiliki ukuran tubuh dan anggota tubuh relatif sama sehingga keduanya membentuk satu kelompok, sedangkan Gresik memiliki ukuran tubuh dan anggota tubuh lebih besar dari Subang, Semarang, dan Banten. Namun tingkat kesamaan karakter Gresik lebih dekat dengan Semarang dan Banten.
Pengelompokan Tachypleus tridentatus
21 dan 12.2% dijelaskan oleh variable di luar model. Berdasarkan tabel classification, model memiliki kemampuan untuk membedakan antar kelompok populasi sebesar 100% yaitu individu diklasifikasikan sebagai individu asli yang berasal dari Semarang (n=2), Gresik (n=3), Subang (n=17), dan Banten (n=10). Percent correct dapat mencapai hingga 100% diduga karena jumlah individu yang ditemukan sangat sedikit disetiap daerahnya.
Tabel 7 Konektivitas antar lokasi Tachypleus tridentatus berdasarkan 27 karakter yang telah dirasiokan
Observed Predicted
Semarang Gresik Subang Banten Percent Correct
Semarang 2 100,0%
Gresik 0 3 100,0%
Subang 0 0 17 100,0%
Banten 0 0 0 10 100,0%
Overall Percentage 6,3% 9,4% 53,1% 31,3% 100,0%
Pembahasan
Sebanyak 7 lokasi penelitian diperoleh jumlah jantan dan betina masing-masing spesies berbeda. Terjadi ketidakseimbangan antara spesies jantan dan betina pada T. gigas, betina lebih banyak ditemukan. T. tridentatus jantan tidak ditemukan sama sekali pada semua lokasi, sedangkan C. rotundicauda jantan lebih banyak ditemukan, khususnya pada daerah Surabaya dari 41 total C. rotundicauda hanya ditemukan 4 betina. Menurut Johnson & Brockmann (2010) perbedaan perbandingan komposisi jantan dan betina disebabkan adanya masa dimana mimi jantan yang tidak memiliki pasangan berkumpul di pantai dan bertindak sebagai satelit untuk menunggu giliran memijah.
C. rotundicauda adalah spesies yang paling sering ditemukan hampir di semua lokasi, kecuali pada daerah Banten. Umumnya perairan Banten memiliki kedalaman tidak lebih dari 13 meter, dan berlumpur yang bercampur dengan pasir. C. rotundicauda hidup di air payau dengan substrat berlumpur, biasanya pada daerah mangrove (Li 2008; Shin & Cheung 2009; Cartwright-Taylor et al. 2011; Rubiyanto 2012). Menurut penelitian yang telah dilakukan oleh Rubiyanto (2012) di Perairan Kuala Tungkal mimi ini memiliki kepadatan lebih tinggi (62 ind/100m2,
n=246) dibandingkan dengan T. gigas (2 ind/100m2, n=8). Daerah Singapura telah ditemukan sebanyak 172 individu C. rotundicauda lebih banyak dibandingkan dengan T. gigas sebanyak 6 individu pada daerah dengan substrat berlumpur (Carthwright-Taylor et al. 2011). Menurut penelitian yang telah dilakukan oleh Mishra (2009) C. rotundicauda banyak ditemukan pada perairan dengan salinitas rendah. Pola sebaran C. rotundicauda adalah acak (Rubiyanto 2012), hal ini menunjukkan kemampuan toleransi suatu individu terhadap ketersediaan sumberdaya lingkungan (Cartwright-Taylor et al. 2009).
22
adanya reklamasi pantai dan degradasi habitat telah menghilang sejak tahun 1990 (Shin et al. 2009). Akibat adanya sarana rekreasi dan pembangunan pelabuhan mimi di Singapura tidak terdapat aktivitas pemijahan di tepi pantai (Cartwright et al. 2011).
Daerah Rembang C. rotundicauda memiliki ukuran lebar maksimal prosoma paling besar dibandingkan dengan daerah lain. Hal ini dikarenakan pada daerah Rembang merupakan daerah mangrove dengan substrat berlumpur tebal. Mangrove dengan substrat berlumpur merupakan habitat dengan sumber makanan yang disukai oleh mimi jenis C. rotundicauda. Lingkungan dengan sumber makanan yang banyak mengandung Ca (bivalvia, polycaeta, moluska) akan membantu pertumbuhan karapas dengan baik. Ukuran tubuh mimi dipengaruhi oleh kepadatan populasi, ketersediaan pangan, dan pengaruh kondisi lingkungan (Chatterji et al. 2000; Zadeh et al. 2011). Penelitian yang telah dilakukan oleh Chatterji et al. (2000) diperoleh hasil bahwa pertambahan panjang dan lebar karapas dipengaruhi oleh ketersediaan pakan dan kepadatan populasi. Mimi memiliki ukuran karapas yang berbeda pada habitat yang berbeda seperti karapas populasi mimi yang ada di Malaysia memiliki karapas yang lebih besar dibandingkan dengan karapas yang ada di India dan Thailand (Chatterji 1999; Srijaya et al. 2010).
C. rotundicauda dan T. tridentatus pada daerah Subang memiliki rata-rata ukuran tubuh dan anggota tubuh paling kecil jika dibandingkan dengan dearah lainnya. Perairan Mayangan Subang berhadapan langsung dengan Laut Jawa disebelah utara sehingga suhu dan salinitas sangat dipengaruhi oleh dinamika perairan Laut Jawa. Rataan suhu yang bervariasi antara 27,5-28,7oC dengan
salinitas 30-33 ppt, kedalaman 20 m. Perairan Mayangan Subang memiliki kekeruhan tinggi akibat pengaruh dari karakteristik Laut Jawa. Pada penelitian ini, Subang merupakan lokasi dengan jumlah tangkapan mimi terbanyak, dalam sehari sebanyak 45 ekor mimi telah tertangkap. Sebanyak 378 ekor mimi tertangkap selama 10 bulan Maret-Desember 2003 penelitian yang telah dilakukan oleh Muslihah (2004) di Mayangan Subang. Kepadatan populasi, umur, ketersediaan pangan, dan pengaruh kondisi lingkungan mempunyai hubungan yang signifikan dengan perubahan dimensi tubuh mimi (Chatterji et al. 2000; Zadeh et al. 2011). Di Balramgari, Orissa pertambahan panjang dan lebar karapak T. gigas dipengaruhi oleh kepadatan dan ketersediaan pakannya. Makanan mimi tersedia dibanyak tempat, jenis makanan mimi diantaranya ikan mati, polycaeta, bivalvia, moluska, serta alga (Chatterji et al. 2000).
SIMPULAN DAN SARAN
Simpulan
23 yaitu telson bergerigi pada genus Tachypleus dan telson halus pada genus Carcinoscorpius. T. tridentatus terdapat duri tajam pada bagian ophistoma dan prosoma, sedangkan tidakuntuk T. gigas.
Subang merupakan lokasi terbanyak ditemukannya mimi, serta memiliki ketiga spesies, sedangkan Rembang merupakan daerah terbanyak ditemukannya spesies C. rotundicauda. Tidak terdapat konektivitas berdasarkan karakter morfologi melalui perhitungan Multinomiallogistic.
Saran
Pengambilan sampel sebaiknya dilakukan pada bulan Agustus-Desember, karena pada bulan Januari-Juli mimi sulit untuk ditemukan, hal ini berkaitan dengan musim pemijahan. Pengukuran dilakukan di lapangan namun dibatasi jumlah perharinya untuk menghindari kesalahan paralaks. Peluang penelitian yang dapat dilakukan selanjutnya yaitu mengenai kajian daerah spawning, feeding, nursery grounds mimi di Indonesia, kualitas darah mimi serta pemanfaatannya.
DAFTAR PUSTAKA
Beekey MA, Mattei JH, Pierce BJ. 2013. Horseshoe crab eggs: A rare resource for predators in Long Island Sound. Journal of Experimental Marine Biology and Ecology. 439(1):152-159.
Cartwrigt-Taylor L, Lee J, & Hsu CC. 2009. Population structure and breeding pattern of the mangrove horseshoe crab Carcinoscorpius rotundicauda in Singapore. Aquatic Biology. 8:61-69
Cartwrigt-Taylor L, Bing YV, Chi HC, Tee LS. 2011. Distribution and abundance of horseshoe crabs Tachypleus gigas and Carcinoscorpius rotundicauda around the main island Singapore. Aquatic Biology. 13:127-136.
Chatterji A, Vijayakumar R, Parulekar AH. 1992. Spawning migration of the horseshoe crab, Tachypleus gigas (Muller), in relation to lunar cycle. Asian Fisheries Science. 5:123-128.
Chatterji, A. 1999. New record of the sympatric distribution of two Asian species of the horseshoe crab. Indian Journal of Marine Science. 77(6):43-48. Chatterji A, Parulekar AH, Vijayakumar R. 2000. Morphometric characteristics in
the horseshoe crab Tachypleus gigas (Artropoda: Merostomata). Indian Journal of Marine Science. 29:333-335.
Christianus A, Saad CR. 2007. Horseshoe crabs in Malaysia and the world. Fishery Mail. 16:8-9.
Chiu HMC, Morton B. 2004. The behavior of juvenil horseshoe crabs, Tachypleus tridentatus (Xiphosura), on nursery beach at Shin Hau Wan, Hong Kong. Hydrobiologia. 523:29-35.
24
Ding JL, Tan KC, Thangamani S, Kusuma N, Seow WK, Bui THH, Wang J, Ho B. 2005. Spatial and temporal coordination of expression of immune response genes during Pseudomonas infection of horseshoe crab, Carcinoscorpius rotundicauda. Nature Publishing Group. 6(7):557-574.
Ferari KM, Targett NM. 2003. Chemical attractants in horseshoe crab, Limulus polyphemus, eggs: the potential for an artificial bait. Journal of Chemical Ecology. 29:477-496.
Hu M, Wang Y, Chen Y, Cheung SG, Shin PKS, Li Q. 2009. Summer distribution and abundance of juvenile Chinese horseshoe crabs Tachypleus tridentatus along an intertidal zone in Southern China. Aquatic Biologi. 7:107-112. Hurton L. 2003. Reducing Post-Bleeding Mortality of Horseshoe Crabs (Limulus
polyphemus) Used in The Biomedical Industry [Tesis]. Virginia (US): Virginia Polytechnic Institute and State University.
Harrington BA. 2001. Red Knot (Calidris canutus). In The Birds of North America, No. 563 (A. Poole and F. Gill, eds.). The Birds of North America, Inc., Philadelphia, Pennsylvania.
[IUCN] International Union for Conservation of Nature. 2015. Red list of threatened species. www. Iuc-nredlist.org. 18 Mei 2015. Pukul. 18.27 WIB. John BA, Kamaruzzaman BY, Jalal KCA, Zaleha K. 2012. Feeding Ecology and
Food Preferences of Carcinoscorpius rotundicauda Collected from the Pahang Nesting Grounds. Sains Malaysiana. 41(7):855-861.
Johnson SL, Brockmann HJ. 2010. Costs of multiple mates: an experimental study in horseshoe crabs. Animal Behaviour. 80:773-782.
Lee CN, Morton B. 2005. Experimentally derived estimates of growht by juvenile Tachypleus tridentatus and Carcinoscorpius rotundicauda (Xiphosura) from nursery beaches in Hong Kong. Jurnal Marine Biology Ecology. 318:39-49. Li HY. 2008. The conservation of horseshoe crabs in Hong Kong [Tesis]. Hong
Kong (HK): City Universitas of Hong Kong.
Muslihah. 2004. Beberapa aspek biologi reproduksi mimi bulan Tachypleusgigas di Perairan Mayangan, Kabupaten Subang, Jawa Barat [Skripsi]. Bogor (ID): Institut Pertanian Bogor.
Mishra JK. 2009. Horseshoe crabs, their eco-biological status along the northeast coast of India and the necessity for ecological conservation. In: Tanacredi JT et al. (eds). 2009. Biology and conservation of horseshoecrabs. Springer Science & Business Media: 89-96.
Mulya MB. 2014. Pelestarian, pemanfaatan sumberdaya genetika mimi ranti (Carcinosscorpius rotundicauda, L) dan mimi bulan (Tachypleus gigas, M). © 2004 Digitized by USU digital library.
Novitsky TJ, Thomas, Dawson, Michael E, Paus, Erik J. 2002. Artificial bait. United States Patent: 639-1295.
Novitsky TJ. 1994. Limulus amebocyte lysate (LAL) detection of endotoxin in human blood. Journal of endotoxin research. 1(4):253-263.
Obst M, Faurby S, Bussarawit S, Funch P. 2012. Molecular phylogeny of extant horseshoe crabs (Xiphosura, Limulidae) indicates Paleogene diversification of Asian species. Molecular Phylogenetics and Evolution. 62(1):21-26. Rubuyanto E. 2012. Studi populasi mimi (Xiphosura) di perairan Kuala Tungkal,
25 Selander RK, Yang SY, Lewontin RC, Johnson WE. 1970. Genetic variation in the horseshoe crab (Limulus polyphemus), a phylogenetic ‘‘relic’’. Evolution. 24(2):402-414
Shin P, Li HY, Cheung SG. 2009. Horseshoe crabs in Hong Kong: Current population status and human exploitation. In: Tanacredi JT et al. (eds). 2009. Biology and conservation of horseshoe crabs. Springer Science & Business Media: 347-360.
Smith DR. 2007. Effect of horseshoe crab spawning density on nest disturbance and exhumation of eggs: a simulation study. Estuaries. 30:287-295.
Sekiguchi, Shuster. 2009. Limits on the global distribution of horseshoe crabs (Limulacea). In: Tanacredi JT et al. (eds). 2009. Biology and conservation of horseshoe crabs. Springer Science & Business Media: 347-360.
Suparta. 1992. Keragaman sifat-sifat morfometrik mimi, Tachypleus gigas (MULLER) & Carcinoscorpius rotundicouda (LATREILLE) di perairan pantai Kabupaten Pandeglang, Jawa Barat dan perairan pantai Kabupaten Rembang, Jawa Tengah [skripsi]. Bogor (ID): Institut Pertanian Bogor. Srijaya TC, Pradeep PJ, Mitun S, Hasan A, Shaharom F, Chatterji A. 2010. A new
record on the morphometric variation in the population of horseshoe crab (Carcinoscorpius rotundicauda, Latreille) obtained from two different ecological habitats of Peninsular Malaysia. Nature. 8:204-211.
Taylor LC, Lee J, Hsu CC. 2011. Population structure and breeding pattern of the mangrove horseshoe crab Carcinoscorpius rotundicauda in Singapore. Aquatic Biology. (8):61–69.
Vauziyah C. 1995. Perkembangan embrio mimi bulan Tachypleus gigas (Muller) dari perairan Teluk Banten pada berbagai salinitas media [Skripsi]. Bogor (ID): Institut Pertanian Bogor.
Walls EL, Berkson J, Smith SA. 2002. The horseshoe crab, Limulus polyphemus: 200 million years of existence, 100 years of study. Review Fisheries Sciences. 10:39-73.
Webster M. 2007. A cambrian peak in morphological variation within trilobite species. Science. 317(5837):499-502.
Yeo DSA, Ding JL, Ho B. 1996. Neuroblastoma cell culture assay shows that Carcinoscorpius rotundicauda haemolymph neutralizes tetrodotoxin. Pergamon. 34(9):1054-1057.
26
3
IDENTIFIKASI MIMI YANG TERTANGKAP DI
PERAIRAN PESISIR PULAU JAWA BERDASARKAN
MARKA GEN CO1
Lusita Meilana, Yusli Wardiatno, Nurlisa A Butet, Majariana Krisanti Program Studi Pengelolaan Sumberdaya Perairan, Fakultas Perikanan dan Ilmu
Kelautan, Institut Pertanian Bogor. Email: lusitameilana@yahoo.co.id
ABSTRACT
Horseshoe crabs are distributed widely in Indonesian waters, and there are three common species inhabiting some coastal waters of Java Island. The research were aimed at establishing phylogenetic relationship among Indonesian horseshoe crab of Java Island and identification based on molecular marker of COI gene and PCR-RFLP in combination with morphological characters. The sample collections were conducted at six sites, i.e. Ujung Kulon, Subang, Semarang, Demak, Rembang, and Kenjeran. The results showed that there were 67 specific nucleotide sites, becoming species identifier of three spesies horseshoe crabs with outgroup species. Molecular identification of T. gigas from this study has shown taxonomy certainty. However, identification of T. tridentatus and C. rotundicauda did not succeed using CO1 gene. PCR-RFLP method revealed that Demak could be defined as a region of C. rotundicauda gene pool, while Subang for T. gigas, and Semarang for T. tridentatus. Those are due to higher haplotype diversity compared to other locations. Keywords: haplotype, horseshoe crabs, Java Island, phylogeny