EVALUASI KERAGAAN FENOTIPE NILAM (
Pogostemon
cablin
Benth.) HASIL MUTASI KROMOSOM DENGAN
KOLKISIN
ULIL AZMI NURLAILI AFIFAH
DEPARTEMEN AGRONOMI DAN HORTIKULTURA FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa skripsi berjudul Evaluasi Keragaan Fenotipe Nilam Pogosthemon cablin Benth. Hasil Mutasi Kromosom dengan Kolkisin adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Maret 2015
RINGKASAN
ULIL AZMI NURLAILI AFIFAH. Evaluasi Keragaan Fenotipe Nilam (Pogostemon cablin Benth) Hasil Mutasi Kromosom dengan Kolkisin. Dibimbing oleh SUDARSONO dan NI MADE ARMINI WIENDI.
Pogostemon cablin Benth. yang dikenal dengan nama nilam di Indonesia adalah tanaman aromatik yang banyak digunakan dalam industri wewangian terutama parfum Penelitian ini bertujuan untuk mempelajari keragaan fenotipe in vitro dan perubahan jumlah kromosom planlet nilam Pogostemon cablin Benth hasil mutasi kromosom dengan kolkisin.
Penelitian ini terdiri dari dua percobaan yang berkelanjutan. Percobaan pertama adalah induksi perakaran pada tunas nilam (Pogostemon cablin Benth.) hasil induksi mutasi dengan kolkisin. Percobaan ini disusun berdasarkan rancangan acak lengkap faktorial dengan dua faktor. Faktor pertama adalah asal eksplan berupa 12 kombinasi kolkisin dan lama perendaman hasil penelitian dari Anne (2012), dan faktor kedua adalah media perakaran dengan dua kombinasi yaitu MS + 0.1 mg/l IAA + 0.1 mg/l BA (media N1) dan MS + 0.5 mg/l IAA + 0.1 mg/l BA media (N2). Percobaan kedua adalah subkultur nilam (Pogostemon cablin Benth.) hasil induksi mutasi dengan kolkisin untuk mendapatkan mutan solid. Planlet pada subkultur kedua (II) berasal dari planlet subkultur pertama (I). Percobaan subkultur ini menggunakan rancangan acak lengkap satu faktor yaitu faktor kombinasi perlakuan kolkisin dan lama perendaman dari percobaan satu.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Pertanian
pada
Departemen Agronomi dan Hortikultura
EVALUASI KERAGAAN FENOTIPE NILAM (
Pogostemon
cablin
Benth.) VARIETAS SIDIKALANG HASIL MUTASI
KROMOSOM DENGAN KOLKISIN
ULIL AZMI NURLAILI AFIFAH
DEPARTEMEN AGRONOMI DAN HORTIKULTURA FAKULTAS PERTANIAN
INSTITUT PERTANIAN BOGOR BOGOR
Judul Skripsi : Evaluasi Fenotipe Nilam (Pogostemon cablin Benth.) Varietas Sidikalang Hasil Mutasi Kromosom dengan Kolkisin.
Nama : Ulil Azmi Nurlaili Afifah NIM : A24090074
Disetujui oleh
Prof Dr Ir Sudarsono, MSc
Pembimbing I Dr Ir Ni Made Armini Wiendi, MS Pembimbing II
Diketahui oleh
Dr Ir Agus Purwito, MScAgr Ketua Departemen
PRAKATA
Puji dan syukur penulis panjatkan kehadirat Tuhan Yang Maha Esa atas limpahan karunia-Nya sehingga karya ilmiah yang berjudul “Evaluasi Keragaan Fenotipe Tanaman Nilam (Pogostemon cablin Benth.) Hasil Mutasi Kromosom dengan Kolkisin” ini dapat terselesaikan.
Penulis mengucapkan terima kasih kepada pihak-pihak yang telah membantu penulis dalam menyelesaikan penulisan skripsi ini. Dosen pembimbing skripsi Bapak Prof. Dr. Ir. Sudarsono, MSc (selaku pembimbing skripsi 1) dan Ibu Dr. Ir. Ni Made Armini Wiendi, MS (selaku pembimbing skripsi 2), yang telah membantu selama penelitian dan telah membimbing hingga karya tulis ini selesai. Ungkapan terima kasih juga disampaikan kepada ayah, ibu, serta seluruh keluarga, dan teman-teman atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat bagi semua pihak yang berkepentingan dan untuk pengembangan ilmu pengetahuan.
Bogor, Maret 2015
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vi
PENDAHULUAN 1
Latar Belakang 1
Tujuan 2
Hipotesis 2
TINJAUAN PUSTAKA 2
Tanaman Nilam 2
Kolkisin dan Poliploidi 3
BAHAN DAN METODE 4
Waktu dan Tempat 4
Alat dan Bahan 4
Metode Penelitian 5
Pelaksanaan Penelitian 7
HASIL DAN PEMBAHASAN 9
Percobaan 1. Pertama adalah induksi perakaran pada tunas nilam (Pogostemon cablin Benth.) 9 Percobaan 2. Subkultur nilam (Pogostemon cablin Benth.) Hasil
Mutasi Kromosom dengan Kolkisin untuk Mendapatkan Mutan Solid
12
KESIMPULAN 25
DAFTAR PUSTAKA 25
LAMPIRAN 28
DAFTAR TABEL
1 Kombinasi perlakuan pada percobaan I induksi perakaran pada
nilam 5
2 Kombinasi perlakuan pada percobaan II subkultur nilam hasil
mutasi kromosom dengan kolkisin 7
3 Rekapitulasi hasil uji F pengaruh kombinasi kolkisin dan media perakaran terhadap pertumbuhan tunas Pogostemon cablin Benth. hasil mutasi kromosom dengan kolkisin
10
4 Pengaruh auksin IAA terhadap jumlah akar nilam Pogostemon cablim Benth.
11 5 Pengaruh kombinasi kolkisin terhadap jumlah akar nilam
Pogostemon cablin Benth. pada media perakaran
11 6 Pengaruh perlakuan kolkisin terhadap jumlah tunas nilam
Pogostemon cablin Benth pada subkultur kedua
12 7 Pengaruh kolkisin terhadap jumlah buku nilam Pogostemon cablin
Benth. pada subkultur kedua 14
8 Pengaruh perlakuan kolkisin terhadap jumlah daun nilam Pogostemon cablin Benth. pada subkultur kedua
15 9 Pengaruh perlakuan kolkisin terhadap jumlah akar nilam
Pogostemon cablin Benth. pada subkultur kedua
16 10 Pengaruh kolkisin terhadap jumlah stomata Pogostemon cablin
Benth. hasil mutasi kromosom dengan kolkisin pada subkultur kedua
17
11 Pengaruh konsentrasi kolkisin terhadap ukuran stomata Pogostemon cablin Benth hasil mutasi kromosom dengan kolkisin pada subkultur kedua
18
12 Pengaruh konsentrasi kolkisin terhadap jumlah kloroplas pada stomata daun nilam Pogostemon cablin Benth. hasil mutasi kromosom dengan kolkisin pada subkultur kedua
20
13 Jumlah kromosom sel planlet nilam hasil mutasi kromosom dengan
DAFTAR GAMBAR
1 Tunas nilam 9 MST: A: kontrol, B: kolkisin 0% lama perendaman 72 jam, C: kolkisin 0,02% lama perendaman 48 jam, D: kolkisin 0,02% lama perendaman 72 jam, E: kolkisin 0,06% lama perendaman 24 jam, F: kolkisin 0,06 % lama perendaman 48 jam, G: kolkisin 0,06% lama perendaman 24 jam
14
2 Stomata pada daun planlet Pogostemon cablin Benth hasil mutasi dengan kolkisin pada subkultur kedua A: kontrol. B:konsentrasi 0.02% lama perendaman 48 jam. C : konsentrasi 0.02% lama perendaman 48 jam D: konsentrasi 0.02% lama perendaman 72 jam. E: konssentrasi 0.02% lama perendaman 24 jam. F: konsentrasi 0.04% lama perendaman 48jam. G: konsentrasi 0.06% lama perendaman 24 jam. Pengamatan pada perbesaran 4x10.
18
3 Jumlah kloroplas Pogostemon scablin Benth. A: kontrol. B:konsentrasi 0.02% lama perendaman 48 jam. C : konsentrasi 0.02% lama perendaman 48 jam D: konsentrasi 0.02% lama perendaman 72 jam. E: konssentrasi 0.02% lama perendaman 24 jam. F: konsentrasi 0.04% lama perendaman 48jam. G: konsentrasi 0.06% lama perendaman 24 jam. Perbesaran 10x10.
19
4 Jumlah kromosom sel Pogostemon cablin Benth hasil mutasi dengan kolkisin setelah subkultur kedua A: kontrol. B:konsentrasi 0.02% lama perendaman 48 jam. C : konsentrasi 0.02% lama perendaman 48 jam D: konsentrasi 0.02% lama perendaman 72 jam. E: konssentrasi 0.02% lama perendaman 24 jam. F: konsentrasi 0.04% lama perendaman 48jam. G: konsentrasi 0.06% lama perendaman 24 jam. Pengamatan dengan perbesaran 4x10.
21
5 Keragaan buku dan percabangan nilam Pogostemon cablin Benth. 9 MST A: buku normal dengan sistem percabangan opposite pada planlet perlakuan kolkisin 0% lama perendaman 72 jam B: buku dengan tiga mata tunas aksilar dan sistem percabangan opposite dan alternate perlakuan kolkisin 0.06% lama perendaman 24 jam
22
6 Planlet kerdil nilam Pogostemon cablin Benth. perlakuan konsentrasi 0.02% lama perendaman 72 jam. 23 7 Kimera tingkat jaringan pada kloroplas dengan perlakuan A:
kolkisin 0,02% lama perendaman 48 jam B: kolkisin 0,02% lama perendaman 72 jam
9 Kimera tingkat jaringan pada kloroplas dengan perlakuan kolkisin
0,06% lama perendaman 24 jam 24
10 Kimera tingkat organ pada kromosom dengan perlakuan A: kolkisin 0,02% lama perendaman 48 jam B: kolkisin 0,02% lama perendaman 72 jam
24
11 Kimera tingkat jaringan pada kromosom dengan perlakuan kolkisin
DAFTAR LAMPIRAN
1 Lampiran 1 Komposisi Media Murashige-Skoog (Gunawan,
1992) 28
2 Sidik ragam pengaruh kolkisin dan media terhadap jumlah akar
nilam pada subkultur satu 29
3 Sidik ragam pengaruh kokisin terhadap jumlah tunas nilam pada
subkultur dua 30
4 Sidik ragam pengaruh kolkisin terhadap jumlah buku nilam pada
subkultur dua 31
5 Sidik ragam pengaruh kolkisin terhadap jumlah daun nilam pada
subkultur dua 32
6 Sidik ragam pengaruh kolkisin terhadap jumlah daun nilam pada
1
PENDAHULUAN
Latar Belakang
Tanaman umumnya mengeluarkan gas CO2 dan sejumlah bahan organik lainnya, terutama yang menghasilkan senyawa dengan aroma khas tertentu yang dikenal dengan senyawa atsiri. Tanaman-tanaman tertentu mengeluarkan minyak atsiri dalam jumlah relatif besar. Minyak atsiri ini tidak dikeluarkan dalam bentuk aroma namun tetap tersimpan dalam tanaman dengan jumlah yang besar (Guenther 1988). Tanaman nilam dikenal sebagai tanaman penghasil minyak atsiri yang bagus. Kebutuhan yang meningkat akan berbagai macam pengharum, pewangi, kosmetik dan obat-obatan mendorong permintaan produksi minyak atsiri semakin meningkat. Saat ini di Indonesia nilam semakin banyak dibudidayakan karena kondisi permintaan yang tinggi.
Minyak nilam merupakan salah satu dari beberapa jenis minyak atsiri yang banyak digunakan dalam industri kosmetika dengan permintaan yang cukup tinggi baik untuk keperluan dalam maupun luar negeri. Minyak atsiri nilam digunakan sebagai bahan baku kosmetik, parfum, antiseptik, dan berbagai produk lainnya. Minyak atsiri sendiri merupakan salah satu hasil proses metabolisme dalam tanaman, yang terbentuk karena reaksi berbagai persenyawaan kimia dengan air (Titik dan Endang 2002).
Indonesia merupakan penghasil minyak atsiri nilam yang potensial, dan sebagai pemasok utama minyak nilam di pasar dunia dengan kontribusi 90% dari total kebutuhan dunia. Di Indonesia, minyak nilam menyumbang devisa lebih dari 50% dari total ekspor minyak atsiri (Nuryani et al., 2005).
Salah satu masalah pada tanaman nilam ialah kadar dan mutu minyak yang masih rendah. Sampai saat ini tanaman nilam tidak ditemukan berbunga, baik di sentra-sentra produksi maupun di daerah pengembangan lainnya. Hal ini menyebabkan usaha untuk mendapatkan keragaman genetik baru dengan sifat yang diharapkan sulit dilakukan.
Pasokan minyak nilam Indonesia cenderung kurang berkualitas dan bermutu. Rendahnya ekspor minyak nilam Indonesia salah satunya disebabkan oleh keterbatasan pasokan bibit nilam yang unggul dengan kualitas dan kuantitas produksi yang tinggi. Oleh karena itu diperlukan teknologi alternatif dalam perbanyakan bibit nilam sehingga dapat dihasilkan bibit dalam jumlah besar dalam waktu singkat yang bermutu baik (Tresnasih 2011).
Tanaman nilam yang dibudidayakan di Indonesia masih terbatas. Nilam hanya memiliki sedikit varietas yang mampu menghasilkan produktivitas tinggi. Usaha meningkatkan keragaman genetik untuk menghasilkan varietas unggul yang diharapkan dapat dilakukan melalui mutasi yang diinduksi secara kimiawi maupun fisik. Salah satunya adalah dengan kolkisin. Senyawa ini dapat menghambat proses pembentukan benang-benang spindel yang mengakibatkan terjadi penggandaan kromosom yang dapat meningkatkan ploidi sel tanaman.
2
(Koornneef. 1991). Secara mendasar tidak terdapat perbedaan antara mutasi yang terjadi secara alami dan mutasi induksi. Keduanya dapat menimbulkan variasi genetik untuk dijadikan dasar seleksi tanaman (Soeranto 2003). Secara relatif, proses mutasi dapat menimbulkan perubahan pada sifat-sifat genetik tanaman baik ke arah positif maupun negatif, dan kemungkinan mutasi yang terjadi dapat juga kembali normal. Mutasi yang terjadi ke arah sifat positif dan terwariskan ke generasi berikutnya merupakan mutasi yang dikehendaki oleh pemulia tanaman pada umumnya (Soeranto 2003).
Keragaman genetik yang ditimbulkan dari induksi mutasi bersifat acak. Dalam mengidentifikasi keragaman genetik hasil induksi mutasi ke arah perubahan yang diinginkan, dapat digunakan teknik seleksi in vitro. Pada teknik in vitro, seleksi ketahanan terhadap cekaman abiotik seperti kekeringan, keracunan Al, pH tanah rendah, dan salinitas dapat digabungkan dalam media kultur in vitro dan digunakan untuk menumbuhkan varian somaklonal yang diperoleh. Tanaman hasil regenerasi jaringan pada kultur in vitro kemungkinan akan mempunyai fenotipe yang toleran terhadap kondisi seleksi (Yunita 2009).
Tanaman nilam hasil mutasi kromosom dengan kolkisin dievaluasi karakter genetik dan karakter fenotipenya untuk menyeleksi galur-galur nilam yang potensial menjadi galur-galur unggul baru yang mampu berproduksi tinggi. Karakter yang timbul dibandingkan dengan karakter unggul yang telah dirilis. Fenotipe nilam hasil mutasi kromosom dengan kolkisin diharapkan memiliki jumlah kromosom yang lebih banyak dan sifat unggul lain yang mampu meningkatkan produksi minyak atsiri nilam.
Tujuan Penelitian
Penelitian ini bertujuan untuk mempelajari keragaan fenotipe in vitro dan perubahan jumlah kromosom planlet nilam Pogostemon cablin Benth. hasil mutasi kromosom dengan kolkisin.
Hipotesis
1. Terdapat paling sedikit satu kombinasi perlakuan kolkisin dan lama perendaman yang memberikan perbedaan fenotipe pada tanaman nilam (Pogostemon cablin Benth.) hasil mutasi kromosom dengan kolkisin. 2. Terdapat paling sedikit satu planlet nilam (Pogostemon cablin Benth.)
hasil mutasi dengan kolkisin yang mengalami meningkatan jumlah kromosom sehingga menimbulkan perubahan karakter morfologi dibandingkan dengan tanaman yang tidak mendapat perlakuan kolkisin.
TINJAUAN PUSTAKA
Tanaman Nilam
3
Sympetale, ordo: Solanales/ Tubiflorae/ Personatae, famili: Lamiaceae/ Labiatae, genus: Pogostemon; dan species: Pogostemon sp. Menurut Ditjenbun (2009), nilam dikenal dengan berbagai nama di beberapa daerah, seperti: dilem (Sumatera–Jawa), rei (Sumba), pisak (Alor), ungapa (Timor). Nama asing dikenal dengan pathcouly. Dikalangan ilmuwan nilam dikenal dengan Pogostemon sp.
Varietas nilam yang dikenal adalah: (a) Pogostemon cablin Benth. Populer dengan nama nilam Aceh. Ciri utamanya adalah daunnya membulat seperti jantung dan dipermukaan bagian bawahnya terdapat bulu-bulu rambut. Sampai umur 3 tahun tanaman ini hampir tidak berbunga. (b) Pogostemon hortensis Backer. Dikenal dengan nama nilam sabun. Ciri-cirinya lembaran daun lebih tipis, tidak berbulu, permukaan daun tampak mengkilat, dan warnanya hijau. (c) Pogostemon heyneanus Benth, sering disebut nilam hutan atau nilam jawa. Ciri-cirinya yaitu ujung daun agak runcing, lembaran daun tipis dengan warna hijau tua dan berbunga lebih cepat (Ditjenbun 2009).
Nilam sidikalang adalah salah satu dari tiga varietas unggul nilam aceh. Varietas ini memiliki produktivitas terna (daun basah) dan kadar minyak paling tinggi dibandingkan dengan varietas lainnya, yaitu varietas tapak tuan dan varietas Lhokseumawe. Nilam sidikalang merupakan tanaman terna aromatis dengan tinggi sekitar 0.3 sampai 0.75 m. (Direktorat Budidaya Tanaman Semusim 2010).
Kolkisin dan Poliploidi
4
yang terus menerus maka akan terbentuk organisme poliploid baik secara generatif maupun secara vegetatif (Eigsti dan Dustin 1957; Poehlman dan Sleper 1995).
Poliploid adalah organisme yang memiliki lebih dari dua set kromosom atau genom dalam sel-sel somatiknya ( Crodwer 1993; Poespodarsono 1988). Banyak tanaman budidaya yang termasuk poliploid alami, diantaranya kacang tanah, tomat, ubi jalar, kapas, tembakau, tebu, nenas, kopi dan teh. Ditinjau dari sudut kepentingan pemuliaan tanaman, keadaan ini merupakan suatu hal yang patut dipelajari dalam usaha meningkatkan sifat tanaman yang diharapkan terutama pada tanaman yang sulit membentuk keragaman genetik secara alami. Pada tanaman talas perlakuan kolkisin terbukti memperlihatkan perubahan pada jumlah kromosom, ukuran stomata dan sel penjaga (Tambon, et al., 1998).
BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan di Laboratorium Kultur Jaringan 2, Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor, Kampus IPB Dramaga Bogor. Analisis sitologi untuk mengetahui jumlah kloroplas dan kromosom dilakukan di Laboratorium Mikro Teknik Departemen Agronomi dan Hortikultura, Fakultas Pertanian, Institut Pertanian Bogor. Penelitian berlangsung dari bulan Maret 2012 sampai Maret 2013. Pada saat perlakuan, suhu ruang inkubasi kultur in vitro adalah 22 0 C, dengan kelembaban relatif (RH) 80%.
Bahan dan Alat
Penelitian ini merupakan penelitian lanjutan dari Anne (2012). Bahan yang digunakan dalam penelitian meliputi bahan tanaman, bahan media dan bahan sterilan. Bahan tanaman untuk percobaan dalam penelitian ini adalah planlet nilam varietas Sidikalang hasil mutasi kromosom dengan kolkisin yang telah dilakukan oleh anne (Anne 2012). Bahan tanaman yang digunakan adalah stek buku tunggal dari planlet nilam hasil mutasi kromosom dengan kolkisin, dari 12 kombinasi perlakuan dosis kolkisin dan lama perendaman. Untuk analisis kloroplas dan kromosom, bagian planlet yang digunakan adalah daun dan akar kultur nilam yang berusia 10 minggu setelah subkultur kedua.
Bahan media terdiri dari media untuk perakaran. Media ini terdiri dari komposisi media MS dengan penambahan zat pengatur tumbuh auksin Indole Acetic Acid yang dikombinasikan dengan sitokinin BA, agar 7 g l-1 dan gula 30 g l-1 dengan pH media 5.9. Perlakuan media untuk induksi akar adalah N1 : 0.1 mg/l IAA dan 0.1 mg/l BA dan N2 : 0.5 mg/l IAA dan 0.1 mg/l BA. Bahan sterilan yang digunakan antara lain NaClO, alkohol 70%, dan air steril.
5
cabinet, pinset, petridish, bunsen, rak kultur, termometer ruangan, gelas piala dan pengaduk kaca. Peralatan yang digunakan untuk uji sitologi terdiri dari mikroskop dan perlengkapannya, silet, cat kuku, pensil dengan ujung berpenghapus, pemanas air, pinset dan gelas kimia.
Metode Penelitian
Penelitian ini terdiri dari dua percobaan yang berkelanjutan. Percobaan pertama adalah induksi perakaran pada tunas nilam (Pogostemon cablin Benth.) hasil induksi mutasi dengan kolkisin, dan percobaan kedua adalah subkultur nilam (Pogostemon cablin Benth.) hasil induksi mutasi dengan kolkisin untuk mendapatkan mutan solid.
Percobaan I. Induksi Perakaran pada Tunas Nilam (Pogostemon cablin
Benth.) Hasil Mutasi dengan Kolkisin
Percobaan ini disusun berdasarkan rancangan acak lengkap faktorial dengan dua faktor. Faktor pertama adalah asal eksplan berupa 12 kombinasi kolkisin dan lama perendaman hasil penelitian dari Anne (2012) dan faktor kedua adalah media perakaran dengan dua kombinasi yaitu MS + 0.1 mg/l IAA + 0.1 mg/l BA (N1) dan MS + 0.5 mg/l IAA + 0.1 mg/l BA (N2).
Tabel 1. Kombinasi perlakuan pada percobaan I induksi perakaran pada nilam Perlakuan Konsentrasi kolkisin
Keterangan : K : Konsentrasi kolkisin
L : Lama perendaman dengan kolkisin N : Konsentrasi auksin N1 : 0.1 mg/l IAA
6
Terdapat 24 kombinasi perlakuan dimana setiap perlakuan diulang sebanyak tiga kali, sehingga diperoleh 72 satuan percobaan. Kombinasi perlakuan pada percoban I dapat dilihat pada tabel 1. Setiap satuan percobaan terdiri dari tiga mata tunas aksilar/ stek buku tunggal sehingga terdapat 216 eksplan sebagai satuan amatan.
Model linier dari percobaan I adalah:
Yijk = µ + αi + βj + (αβ)ij + εijk Keterangan:
Yijk : Nilai pengamatan faktor kombinasi kolkisin dan lama perendaman serta faktor media taraf ke-j dan ulangan ke-k (k = 1,2,3)
µ : Nilai tengah umum
αi : Pengaruh kombinasi kolkisin dan lama perendaman ke-i = 1,2,3,4 βj : Pengaruh auksin IAA dalam media perakaran ke-j, j=1,2,3
(αβ)ij : Pengaruh interaksi kolkisin dan auksin IAA
εijk : Pengaruh galat dari satuan percobaan ke-i, pada ulangan ke-j
Peubah yang diamati adalah jumlah akar, jumlah tunas, jumlah buku dan jumlah daun. Data pengamatan diuji menggunakan uji F pada taraf 5%. Apabila pengaruh perlakuan berbeda nyata, maka dilakukan uji lanjut menggunakan DMRT (Duncan Multiple Range Test) pada taraf 5%.
Percobaan II. Subkultur Nilam (Pogostemon cablin Benth.) Hasil Induksi Mutasi dengan Kolkisin untuk Mendapatkan Mutan Solid
Planlet pada subultur dua berasal dari planlet subkultur satu yang berumur 10 minggu setelah tanam yang ditanam dalam media MS + 0.1 mg/l IAA + 0.1 mg/l BA. Percobaan subkultur ini menggunakan rancangan acak lengkap satu faktor. Faktor yang digunakan adalah faktor kombinasi perlakuan kolkisin dan lama perendaman dari percobaan satu di atas. Terdapat enam kombinasi perlakuan yang digunakan dari percobaan satu yaitu kolkisin 0% selama 72 jam, kolkisin 24 jam selama 48 jam, Kolkisin 0.02% selama 72 jam, kolkisin 0.04% selama 24 jam, kolkisin 0.04% selama 48 jam dan kolkisin 0.06% selama 24 jam. Masing-masing kombinasi perlakuan ini diambil sembilan tunas yang dihasilkan kemudian diulang sebanyak tiga kali sehingga terdapat 162 satuan percobaan. Setiap satuan percobaan terdiri dari tiga tunas sehingga terdapat 486 satuan amatan, seperti tersaji pada Tabel 2.
Model linier yang digunakan sebagai berikut: Yij = μ + αi + εijk
Keterangan :
Yij : Nilai pengamatan pada perlakuan kolkisin ke-i pada ulangan ke j
μ : Nilai tengah umum
7
Peubah yang diamati adalah jumlah akar, jumlah tunas, jumlah buku dan jumlah daun. Data yang diperoleh dianalisis dengan uji F pada taraf 5%. Apabila berbeda nyata dilakukan uji lanjut DMRT (Duncan Multiple Range Test) pada taraf 5%. Planlet yang dihasilkan dari subkultur dua digunakan untuk uji sitologi. Uji sitologi dilakukan pada peubah jumlah stomata, jumlah kloroplas dan jumlah kromosom nilam.
Tabel 2. Kombinasi perlakuan pada percobaan II subkultur nilam hasil mutasi kromosom dengan kolkisin
Pembuatan Media dan Sterilisasi. Media dasar yang digunakan
8
auksin dan sitokinin sesuai perlakuan dan selanjutnya ditambahkan aquades sampai tanda tera (satu liter). Larutan ini kemudian diukur pHnya, pH diukur dan diatur agar sesuai dengan kondisi tumbuh eksplan. Dalam penelitian ini pH yang digunakan adalah 5.9. Pengaturan pH yang diinginkan dilakukan penambahan KOH 1 N jika pH larutan di bawah 5.9 dan dilakukan penambahan HCL 1 N jika pH diatas 5.9. Setelah diatur pHnya, larutan ini kemudian dituang ke dalam panci, selanjutnya ditambahkan agar-agar 7 g/l. Larutan media dipanaskan untuk melarutkan bahan pemadat berupa agar sambil diaduk sampai mendidih, kemudian dituangkan ke dalam botol kultur sebanyak 25 ml/botol (volume botol 200 ml). Selanjutnya botol ditutup plastik dan diikat dengan karet gelang. Media disterilisasi menggunakan autoklaf dengan tekanan 17.5 psi, 1210 C selama 20 menit. Sterilisasi alat seperti pisau, pinset, scalpel, cawan petri, botol kultur kosong, dan botol berisi air steril disterilisasi menggunakan autoklaf dilakukan selama satu jam pada suhu 1210 C dan tekanan 17.5 psi.
Sterilisasi planlet dan Sub Kultur Planlet. Planlet disubkultur pada
media MS + 0.1 mg/l IAA + 0.1 mg/l BA dan MS + 0.5 mg/l IAA + 0.5 mg/l BA pada subkultur pertama. Subkultur kedua planlet nilam disubkultur ke dalam media MS + 0.1 mg/l IAA + 0.1 mg/l BA. Planlet hasil mutasi yang digunakan di dalam penelitian mengalami kontaminasi sehingga perlu disterilisasi terlebih dahulu sebelum disubkultur ke dalam media. Tunas dipotong–potong pada masing–masing buku menghasilkan stek buku tunggal. Stek buku ini kemudian dibilas dengan menggunakan air steril sebanyak dua kali, lalu dimasukkan kedalam larutan NaClO 3% selama 10 menit sambil digoyang-goyang dengan tangan. Planlet yang sudah disterilisasi disubkultur ke dalam media. Pada saat penanaman, semua peralatan yang digunakan disemprot dengan alkohol 70%, sebelum dimasukan ke dalam laminar air flow cabinet. Alat–alat yang digunakan untuk memindahkan eksplan, sebelum digunakan dibakar dahulu sampai panas kemudian didiamkan dahulu sampai dingin. Pada setiap botol ditanam 3 eksplan. Inkubasi kultur dilakukan pada ruangan dengan penyinaran ± 650 lux, dan suhu 220 C. Pengamatan dilakukan selama 10 minggu setelah tanam pada masing-masing subkultur.
Uji Sitologi. Planlet yang diuji sitologi adalah dari hasil subkultur kedua.
Tunas yang digunakan adalah tunas berumur 10 MST yang diambil bagian daun dan akarnya. Percobaan ini terdiri dari analisis stomata, kloroplas dan kromosom tunas nilam yang masing-masing perlakuan diambil 3 sampel. Waktu pengambilan akar untuk uji kromosom adalah pukul 08.30 pagi.
9
perhitungan. Dokumentasi dibuat di bawah mikroskop dengan perbesaran 4x10.
Penghitungan jumlah kromosom menggunakan metode squashing yang telah dimodifikasi seperti dilaporkan oleh (Rodiansah 2007). Pengambilan sampel dilakukan pada pukul 08.30 pagi. Pengambilan material : ujung akar dipotong sepanjang ± 1 cm kemudian dimasukkan ke dalam air untuk menghilangkan kotorannya. Potongan bahan tanaman tersebut kemudian dimasukan ke dalam botol berisi hidroksiquinolin 150.002 M (0.32 g/l aquades), selanjutnya botol tersebut disimpan pada suhu 20oC selama 3–5 jam. Fiksasi sampel : potongan ujung akar dimasukkan ke dalam air bersih kemudian dibuang tudung akarnya dengan pisau silet, potongan tersebut kemudian dimasukkan ke dalam larutan asam asetat 45% selama 10 menit. Selanjutnya potongan diangkat dan dimasukkan ke dalam larutan yang terdiri dari 1 N HCl dan asam asetat 45% dengan perbandingan 3:1 (v/v) pada suhu 80oC selama 3–5 menit. Potongan bahan tanaman tadi diangkat dan dimasukkan ke dalam pewarna Orsein 2%, selanjutnya dipindahkan pada gelas preparat yang ditetesi Orsein 2%. Potongan ujung akar tersebut dipotong lagi sehingga berukuran 1–2 mm dari ujung akar dan sisanya dibuang. Gelas penutup kemudian dipasang, dipukul–pukul perlahan–lahan dengan pangkal pensil berkaret dan dipanaskan sebentar. Selanjutnya gelas penutup ditekan halus dengan jempol dan pinggirnya direkat dengan cat kuku bening dan siap diamati di bawah mikroskop dengan perbesaran 10x10. Setelah terlihat penyebaran kromosom, dilakukan penghitungan jumlah kromosom dan dibuat dokumentasinya di bawah mikroskop.
HASIL DAN PEMBAHASAN
Percobaan I. Induksi Perakaran pada Tunas Nilam (Pogostemon cablin
Benth.) Hasil Induksi Mutasi dengan Kolkisin pada Subkultur Satu
Percobaan ini menggunakan sumber eksplan Pogostemon cablin Benth. hasil mutasi kimia dengan kolkisin sebanyak 12 perlakuan yang ditumbuhkan pada media awal MS ditambah dengan sitokinin 0.5 mg/l BA dan 0.5 mg/l kinetin. Pada media awal ini nilam tidak berakar oleh karena itu ditambahan auksin IAA dalam media untuk menginduksi perakaran nilam. Media perakaran yang digunakan dengan menambahkan auksin IAA dengan dua taraf yaitu 0.1 dan 0.5 mg/l.
10
Tabel 3. Rekapitulasi hasil uji F pengaruh kombinasi kolkisin dan media perakaran terhadap pertumbuhan tunas Pogostemon cablin Benth. hasil mutasi kromosom dengan kolkisin
11
Media perakaran yang ditambahkan tidak berpengaruh nyata terhadap jumlah akar nilam Pogostemon cablin Benth hingga minggu ke-4 setelah dikulturkan. Media perakaran mulai berpengaruh pada minggu ke-6 setelah dikulturkan. Tabel 4 menunjukkan pengaruh media terhadap jumlah akar nilam. Jumlah akar tunas nilam pada media dengan IAA 0.1 mg/l menghasilkan nilai rataan yang lebih tinggi dibandingkan dengan media dengan IAA 0.5 mg/l. Hasil rataan jumlah akar tersebut mengindikasikan bahwa media dengan IAA 0.1 mg/l lebih baik dalam menginduksi perakaran nilam Pogostemon cablin Benth dalam percobaan ini. Nilai sidik ragam dapat dilihat pada Lampiran 2.
Tabel 4. Pengaruh auksin IAA terhadap jumlah akar nilam Pogostemon cablin Benth. tidak berbeda nyata berdasarkan uji DMRT 5%.
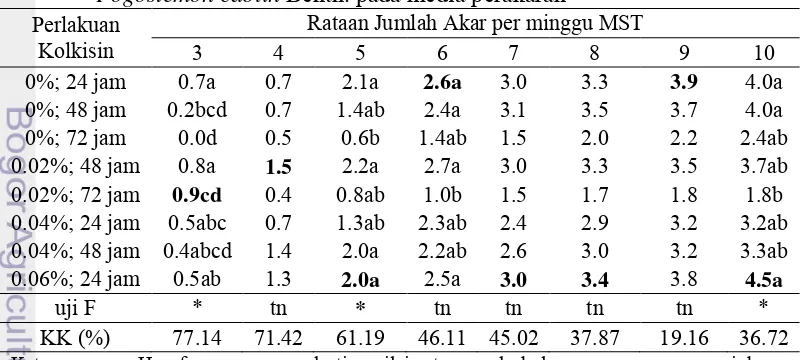
Tabel 5 menyajikan pengaruh perlakuan kolkisin terhadap jumlah akar nilam. Perlakuan 0.06% selama 24 jam memiliki jumlah akar tertinggi namun nilai ini tidak berbeda dengan perlakuan kontrol. Konsentrasi 0.02% selama 72 jam menunjukkan jumlah akar yang paling rendah dibandingkan dengan perlakuan kolkisin yang lain. Hal ini menunjukkan masih terjadi menghambatan pembentukan akar pada nilam yang memperoleh perlakuan kolkisin.
Tabel 5. Pengaruh kombinasi perlakuan kolkisin terhadap jumlah akar nilam Pogostemon cablin Benth. pada media perakaran
12
Rata-rata planlet mulai membentuk akar pada minggu ketiga. Selama pengamatan berlangsung terdapat beberapa planlet yang membentuk kalus terutama pada perlakuan media MS + 0.5 mg/l IAA + 0.1 mg/l BA. Wattimena et al. (1992) menyatakan bahwa Asam Indol Asetat (IAA) auksin diproduksi di pucuk dan ditranslokasikan secara base petal atau polair pangkal batang tanaman untuk menginduksi perakaran. Menurut Mariami (2003), zat pengatur tumbuh sitokinin berperan dalam pembelahan sel dan morfogenesis, sedang auksin berperan dalam mengatur pertumbuhan dan pemanjangan sel. Pemanjangan sel, pembelahan sel, morfogenesis dan pengaturan pertumbuhan merupakan proses yang penting dalam memicu terbentuknya kalus. Tingginya konsentrasi IAA pada media dengan penambahan 0,5 mg/l cenderung memicu terbentuknya kalus pada planlet nilam. Pembentukan kalus ini kurang dikehendaki di dalam produksi bibit.
Kolkisin tidak berpengaruh nyata terhadap pertumbuhan akar tunas nilam minggu 7, 8 dan 9 dan berpengaruh nyata pada akar nilam minggu 10 setelah tanam. Pada akhir pengamatan konsentrasi kolkisin 0.06% selama 24 jam memiliki jumlah akar tertinggi meskipun tidak berbeda nyata dengan kontrol. Jumlah akar terendah adalah perlakuan kolkisin 0.02% selama 72 jam dengan nilai 1.82. Nilai jumlah akar ini dapat mengindikasikan bahwa perlakuan 0.02% selama 72 jam menghambat perakaran tunas nilam. Menurut Anne (2012), tanaman yang mendapat perlakuan perendaman kolkisin tidak dapat membentuk akar, kecuali pada perlakuan konsentrasi kolkisin 0.02% dengan perendaman 24 jam pada umur 4 MST dan perlakuan konsentrasi kolkisin 0.06% dengan perendaman 24 jam pada umur 8 minggu setelah tanam. Pada penelitian ini, tunas yang mendapat perlakuan kolkisin dari penelitian sebelumnya mampu membentuk akar mulai dari minggu 3 setelah tanam. Beberapa perlakuan masih mengalami penghambatan perakaran seperti pada perlakuan 0.02% selama 72 jam.
Percobaan II. Subkultur Nilam (Pogostemon cablin Benth.) Hasil Induksi Mutasi dengan Kolkisin untuk Mendapatkan Mutan Solid Jumlah Tunas
Planlet nilam yang disubkultur pada percobaan II ini berumur 10 minggu setelah dikulturkan dan ditumbuhkan pada media MS + 0.1 mg/l IAA + 0.1 mg/l BA. Planlet yang disubkultur dalam percobaan ini adalah planlet yang mendapat perlakuan kolkisin 0% selama 72 jam; kolkisin 0.02% selama 48 jam; 0.02% selama 72 jam; 0.04% selama 24 jam; 0.04% selama 48 jam dan 0.06% selama 24 jam. Perlakuan lainnya tidak disubkultur karena mengalami kontaminasi.
13
Konsentrasi kolkisin 0.06% dengan lama perendaman 24 jam menghasilkan jumlah tunas yang lebih sedikit dibandingkan kontrol. Hal ini dapat disebabkan karena rusaknya sel-sel pada tanaman. Damayanti dan Mariska (2003) melaporkan perendaman dengan kolkisin dapat mengakibatkan penundaan pertumbuhan akibat jaringan yang rusak dan memerlukan waktu lama untuk tumbuh. Ajijah dan Bermawie (2003) melaporkan perlakuan kolkisin 1% pada tanaman kencur generasi pertama mengakibatkan pembentukan anakan terhambat sehingga memiliki jumlah anakan yang sedikit. Hal ini menunjukan bahwa pemberian kolkisin dapat menghambat terjadinya pertunasan sehingga tanaman kontrol memiliki jumlah tunas yang lebih banyak. Keragaan tunas nilam hasil mutasi dengan kolkisin disajikan dapat dilihat pada Gambar 1. Sidik ragam jumlah tunas nilam dapat dilihat pada lampiran 3.
Tabel 6. Pengaruh perlakuan kolkisin terhadap jumlah tunas nilam Pogostemon cablin Benth pada subkultur dua
Perlakuan Kolkisin
Rataan jumlah tunas minggu ke- (MST)
1 2 3 4 5 6 7 8 9
Pada tunas nilam subkultur kedua berikut menunjukkan, tunas yang mendapat perlakuan kolkisin 0.06% selama 24 jam mengalami penghambatan proliferasi tunas. Perlakuan 0.06% selama 24 jam ini pada minggu 9 setelah tanam menghasilkan jumlah tunas terendah dibandingkan kontrol maupun perlakuan kolkisin yang lain. Jumlah tunas tertinggi terdapat pada perlakuan kontrol namun nilainya tidak berbeda nyata dengan perlakuan 0.02% selama 48 jam; 0,02% selama 72 jam dan 0,04% selama 24 jam. Perlakuan dengan jumlah tunas yang lebih rendah memiliki keragaan batang yang lebih besar dibandingkan dengan dengan tanaman kontrol seperti pada gambar 1.
14
Gambar 1: Tunas nilam 9 MST: A: kontrol. B: kolkisin 0% lama perendaman 72 jam. C: kolkisin 0.02% lama perendaman 48 jam. D: kolkisin 0.02% lama perendaman 72 jam. E: kolkisin 0.06% lama perendaman 24 jam. F: kolkisin 0.06 % lama perendaman 48 jam. G: kolkisin 0.06% lama perendaman 24 jam
Jumlah Buku
Kolkisin memberi pengaruh nyata terhadap jumlah buku nilam Pogostemon cablin Benth pada minggu 3 dan minggu 10 seperti pada tabel 7. Pada minggu pertama setelah tanam perlakuan konsentrasi 0.04% dengan lama perendaman 24 jam menghasilkan tunas dengan jumlah buku paling tinggi diantara perlakuan lain. Pada minggu selanjutnya pada 2-3 minggu setelah tanam jumlah buku tertinggi pada perlakuan kolkisin 0.02% lama perendaman 48 jam. Pengamatan pada 4-10 minggu setelah tanam jumlah buku tertinggi adalah tanaman kontrol dengan kolkisin 0% dan lama perendaman 72 jam. Tabel 7. Pengaruh kolkisin terhadap jumlah buku nilam Pogostemon cablin
Benth pada subkultur dua
Perlakuan Kolkisin
rataan jumlah buku minggu ke- (MST)
1 2 3 5 6 7 8 9 10 buku/tunas
0%; 72 jam 0.7 1.6 2.7ab 5.0 6.0 6.8 7.8 8.7 9.8a
0.02%; 48 jam 1.2 2.0 3.2a 5.1 6.0 6.8 7.6 8.5 9.6ab
0.02%; 72 jam 1.1 1.7 2.7ab 4.7 5.6 6.7 7.5 8.2 8.8a 0.04%; 24 jam 1.3 1.9 2.8ab 4.4 5.3 6.1 6.8 7.6 8.4a
0.04%; 48 jam 1.3 1.9 2.9ab 4.5 5.3 6.2 7.0 7.7 8.3ab 0.06%; 24 jam 0.8 1.3 2.1b 4.0 5.0 5.8 6.7 7.4 8.1b uji F tn tn * tn tn tn tn tn * KK (%) 101.2 81.2 49.4 21.4 15.9 11.6 9.3 8.1 8.8
15
Dari tabel dapat dilihat bahwa pertumbuhan buku tanaman dengan perlakuan kolkisin cenderung lambat dibandingkan dengan tanaman kontrol. Hasil sidik ragam jumlah buku dapat dilihat pada Lampiran 4. Hal ini sesuai yang dilaporkan oleh Poespodarsono (1988) mutasi kromosom dapat mengakibatkan terjadi perubahan sifat pada tanaman. Bertambahnya jumlah kromosom dapat pula mengakibatkan penurunan ukuran sel dan produksi karena adanya penurunan fertilitas akibat poliploidi.
Pada pengamatan morfologi tunas dari buku nilam secara in vitro, buku nilam perlakuan perendaman dengan kolkisin 0.06% selama 24 jam terdapat planlet yang mengalami perubahan fenotipe akibat kimera. Perubahan fenotipe mengindikasikan adanya kimera. Tanaman nilam memiliki sistem percabangan opposite. yaitu terdapat dua daun pada setiap buku tunasnya. Pada Gambar 3 menunjukan terdapat tiga mata tunas aksilar dalam satu buku dengan sistem percabangan opposite (satu buku dua daun) dan alternate (satu buku satu daun) dalam satu planlet yang sama. Kimera disebabkan karena jaringan eksplan mengalami mutasi parsial akibatnya tanaman mengalami malformasi, dan jumlah mata tunas aksilar berbeda baik dengan tunas lain yang berasal dari eksplan yang sama maupun dengan cabang tunas yang berkembang diatasnya. Blakeslee dan Avery (1937) melaporkan bahwa perlakuan kolkisin 0.4% selama satu hari pada biji Datura dapat menginduksi kimera dan mengakibatkan deformasi yang bervariasi pada tanaman akibat perubahan jumlah kromosom. Daun tanaman mengalami malformasi, permukaan daun kasar dan batang yang kerdil.
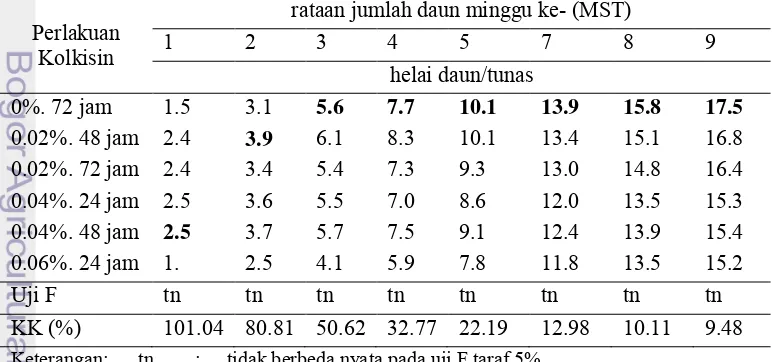
Jumlah Daun
Jumlah daun terus mengalami kenaikan setiap minggu seiring dengan pertambahan dan pertumbuhan tunas. Pada 10 minggu setelah tanam, daun mulai mengalami senescence dan bagian bawah tunas mencoklat. Hal ini disebabkan karena umur tanaman sudah terlalu tua, sehingga harus di subkultur ke dalam media baru.
Tabel 8 Pengaruh perlakuan kolkisin terhadap jumlah daun nilam Pogostemon cablin Benth pada subkultur kedua
Perlakuan Kolkisin
rataan jumlah daun minggu ke- (MST)
16
Hasil uji F pada Tabel 8 menunjukkan bahwa perlakuan kolkisin tidak berpengaruh nyata terhadap jumlah daun nilam Pogostemon cablin Benth. Perlakuan kolkisin 0.04% lama perendaman 48 jam memiliki jumlah daun paling banyak pada minggu pertama setelah tanam, sedangkan untuk minggu 2 setelah tanam nilai jumlah daun paling banyak ada pada perlakuan konsentrasi 0.02% lama perendaman 48 jam. Pada minggu 3 hingga akhir pengamatan jumlah daun tertinggi ada pada tanaman kontrol dengan konsentrasi kolkisin 0% lama perendaman 72 jam. Sidik ragam jumlah daun dapat dilihat pada Lampiran 5.
Permadi et al. (1991) melaporkan bahwa pada tanaman bawang merah. pemberian kolkisin menekan tinggi tanaman dan jumlah daun pada tanaman generasi pertama. Pada penelitian nilam, perlakuan kolkisin juga menghasilkan tunas dengan jumlah daun yang lebih sedikit dibandingkan dengan tunas kontrol.
Jumlah Akar
Planlet mulai berakar pada 3 minggu setelah tanam dan terus bertambah sampai 9 minggu setelah tanam. Pada Tabel 9 berikut menunjukkan pengaruh kolkisin terhadap jumlah akar nilam Pogostemon cablin Benth selama. Menurut Anne (2012) tanaman nilam hasil mutasi dengan kolkisin tidak berakar kecuali pada perlakuan konsentrasi kolkisin 0.02 % dengan perendaman 24 jam pada umur 4 minggu setelah tanam dan perlakuan konsentrasi kolkisin 0.06 % dengan perendaman 24 jam pada umur 8 minggu setelah tanam. Pada penelitian kali ini tanaman berakar karena adanya penambahan auksin Indole Acetic Acid pada media yang digunakan untuk menginduksi perakaran nilam Pogostemon cablin Benth namun perakaran tanaman masih terlihat terhambat. Sidik ragam jumlah akar terdapat pada Lampiran 6.
Tabel 9 Pengaruh perlakuan kolkisin terhadap jumlah akar nilam Pogostemon cablin Benth. subkultur kedua
Perlakuan Kolkisin
Rataan jumlah akar minggu ke- MST
1 2 3 4 5 6 7 8 9
17
dihasilkan oleh nilam yang mendapat perlakuan kolkisin 0.04% selama 48 jam. Pada minggu ke 4-6 jumlah akar tertinggi adalah akibat perlakuan kolkisin 0.06% dan lama perendaman 24 jam. Pada tiga minggu terakhir 7-9 minggu setelah tanam jumlah akar tertinggi ada pada konsentrasi kolkisin 0.02% lama perendaman 48 jam. Menurut Poespodarsono (1988) mutasi kromosom dapat mengakibatkan terjadinya perubahan sifat pada tanaman. Bertambahnya jumlah kromosom dapat pula mengakibatkan penurunan ukuran sel dan produksi karena adanya penurunan fertilitas akibat poliploidi. Beragamnya nilai jumlah akar ini diduga karena terdapat perubahan sifat pada tanaman nilam Pogostemon cablin Benth.
Uji Sitologi
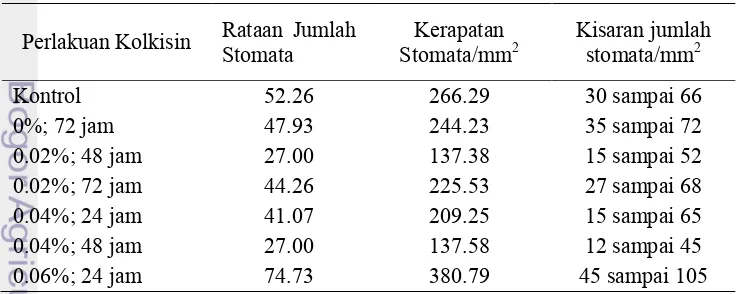
Pengamatan Kerapatan Stomata. Kerapatan stomata dihitung
berdasarkan jumlah stomata per luasan bidang pandang. Pengamatan jumlah stomata dilakukan di bawah mikroskop dengan perbesaran 4x10. Perlakuan konsentrasi kolkisin menyebabkan kerapatan stomata semakin rendah seperti pada Tabel 10 dan Gambar 2 di bawah ini. Tanaman yang mendapat perlakuan kolkisin 0.02 % lama perendaman 48 jam memiliki jumlah stomata yang paling sedikit. Nilai ini hampir sama besarnya dengan perlakuan kolkisin 0.04% lama perendaman 48 jam. Jumlah stomata berhubungan dengan kerapatan stomata. Tanaman dengan jumlah stomata yang banyak memiliki kerapatan stomata yang tinggi, dan sebaliknya. Nilai kerapatan stomata tertinggi ada pada perlakuan kolkisin 0.06% lama perendaman 24 jam setelah itu diikuti oleh nilai rataan kerapatan stomata tanaman kontrol dengan nilai 266.55 mm2. Tanaman dengan perlakuan kolkisin memiliki jumlah stomata yang lebih sedikit dibandingkan dengan tanaman kontrol kecuali pada perlakuan kolkisin 0.06% lama 24 jam. Dari hasil ini dapat disimpulkan bahwa kolkisin mampu mengubah sifat tanaman nilam Pogostemon cablin Benth.
Tabel 10 Pengaruh kolkisin terhadap jumlah stomata Pogostemon cablin Benth. hasil mutasi kromosom dengan kolkisin pada subkultur kedua
Perlakuan Kolkisin Rataan Jumlah Stomata Stomata/mmKerapatan 2 Kisaran jumlah stomata/mm2
Kontrol 52.26 266.29 30 sampai 66
18
menjadi indikator bahwa tanaman mengalami peningkatan jumlah ploidi. Menurut Suryo (1995) tanaman poliploid umumnya memiliki sel-sel yang lebih besar dibandingkan diploidnya. Usman et al. (2008) melaporkan jeruk mandarin Kinnow poliploid memiliki 5.2 stomata/mm2 dan diploidnya 7.5 stomata/mm2.
Gambar 2: Stomata pada daun planlet Pogostemon cablin Benth hasil mutasi dengan kolkisin pada subkultur kedua A: kontrol. B:konsentrasi 0.02% lama perendaman 48 jam. C : konsentrasi 0.02% lama perendaman 48 jam D: konsentrasi 0.02% lama perendaman 72 jam. E: konssentrasi 0.02% lama perendaman 24 jam. F: konsentrasi 0.04% lama perendaman 48jam. G: konsentrasi 0.06% lama perendaman 24 jam. Pengamatan pada perbesaran 4x10.
Ukuran Stomata Daun. Dari Tabel 11 dibawah ini terlihat bahwa
panjang stomata yang paling panjang adalah pada perlakuan kolkisin 0.02% selama 48 jam dan stomata terpendek adalah perlakuan kolkisin 0.04% selama 24 jam. Lebar stomata memiliki nilai rataan yang paling tinggi pada kolkisin 0.02% selama 48 jam dan nilai terendah 0.04% selama 24 jam. Terlihat bahwa terdapat perbedaan ukuran stomata antara kontrol dan perlakuan kolkisin. Perlakuan kolkisin 0.02% selama 48 jam mampu meningkatkan ukuran stomata dan perlakuan, 0.04% selama 24 jam membuat ukuran stomata menjadi lebih kecil dibandingkan dengan kontrol.
19
Tabel 11 Pengaruh konsentrasi kolkisin terhadap ukuran stomata Pogostemon cablin Benth hasil mutasi kromosom dengan kolkisin pada subkultur kedua
Perlakuan kolkisin panjang stomata Rata-rata ukuran stomata µm (µm) lebar stomata (µm)
Kontrol 32035.38 30403.85
0%; 72 jam 29263.50 23369.80
0.02%; 48 jam 34520.49 30333.33
0.02%; 72 jam 32320.37 25513.93
0.04%; 24 jam 28337.87 23586.29
0.04%; 48 jam 31847.54 28026.11
0.06%; 24 jam 29045.87 21729.92
Pengamatan Jumlah Kloroplas. Kloroplas diamati dengan perbesaran
mikroskop 10x10. Kloroplas terletak pada sel-sel penjaga stomata. Setelah menemukan stomata, perhitungan jumlah kloroplas dapat dilakukan dengan perbesaran mikroskop 10x10. Tabel 12 dan Gambar 3 dibawah ini menunjukkan rata-rata jumlah kloroplas Pogostemon cablin Benth. pada perlakuan konsentrasi kolkisin dan lama perendaman tertentu. Hasil dari pengamatan menunjukkan bahwa perlakuan kolkisin dan lama perendaman memiliki rata-rata jumlah kloroplas yang lebih tinggi dibandingkan dengan tanaman kontrol.
20
kloroplas. Omidbaigi et al.(2010) melaporkan bahwa semakin meningkatnya tingkat ploidi tanaman, jumlah kloroplas akan semakin meningkat. Percobaan pada tanaman jahe menunjukkan jumlah kloroplas pada tanaman jahe menunjukkan perbedaan yang nyata antara tanaman diploid, triploid dan tetraploid (Sakhanokho 2009). Kondisi ini sesuai dengan hasil pengamatan jumlah kloroplas Pogostemon cablin Benth. yang semua perlakuannya memiliki jumlah klorolas lebih besar dibandingkan tanaman kontrol.
Tabel 12 Pengaruh kolkisin terhadap jumlah kloroplas pada stomata daun planlet nilam Pogostemon cablin Benth. hasil mutasi kromosom dengan kolkisin pada subkultur kedua
Perlakuan Kolkisin Rataan jumlah kloroplas/stomata Kisaran jumlah kloroplas/stomata
Kontrol 13.13 8 sampai 18
Pengamatan Jumlah Kromosom. Penggunaan kolkisin pada titik
tumbuh akan mencegah pembentukan benang – benang spindel sehingga menyebabkan penggandaan kromosom tanpa pembentukan dinding sel. Perlakuan ini dapat menyebabkan peningkatan jumlah kromosom sebelum terjadi penggandaan. Kromosom dapat terlihat jelas selama tahap – tahap tertentu pembelahan inti. Biasanya kromosom jelas teramati pada tahap metafase (Crowder. 1993).
Berdasarkan hasil uji sitologi terhadap planlet nilam hasil mutasi kromosom dengan kolkisin secara in vitro, bagian akar lebih mudah diamati dibandingkan dengan bagian pucuk. Bagian pucuk terlalu tebal sehingga banyak sel yang menumpuk. Selain itu preparat yang dihasilkan kurang jelas karena bagian pucuk mengandung klorofil. Tabel 13 berikut menunjukkan jumlah kromosom dalam tiap perlakuan kolkisin.
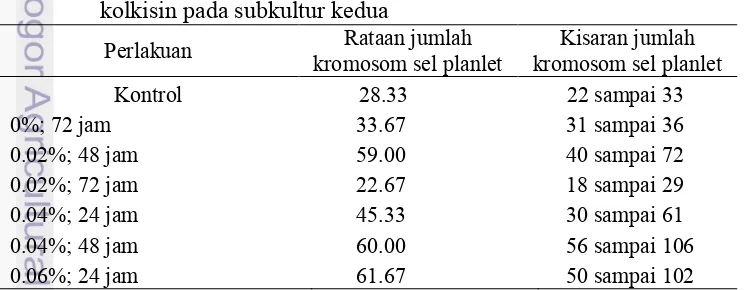
Tabel 13 Jumlah kromosom sel planlet nilam hasil mutasi kromosom dengan kolkisin pada subkultur kedua
21
Gambar 4: Jumlah kromosom sel Pogostemon cablin Benth hasil mutasi dengan kolkisin setelah subkultur kedua A: kontrol. B:konsentrasi 0.02% lama perendaman 48 jam. C : konsentrasi 0.02% lama perendaman 48 jam D: konsentrasi 0.02% lama perendaman 72 jam. E: konssentrasi 0.02% lama perendaman 24 jam. F: konsentrasi 0.04% lama perendaman 48jam. G: konsentrasi 0.06% lama perendaman 24 jam. Pengamatan dengan perbesaran 4x10.
Wu Yu dan Li Ming (2013) melaporkan bahwa jumlah kromosom nilam adalah 2n = 28. Tanaman dengan jumlah kromosom tertinggi pada konsentrasi 0.06% lama perendaman 24 jam dengan rata-rata jumlah kromosom 61.67. Nilai ini hampir tiga kali lipat lebih banyak jika dibandingkan dengan jumlah kromosom kontrol dengan nilai 28.33. Rata-rata jumlah kromosom nilam memiliki nilai yang lebih tinggi dibandingkan dengan jumlah kromosom kontrol kecuali pada konsentrasi 0.02% dengan lama perendaman 72 jam. Perlakuan ini memiliki rataan jumlah kromosom 22.67 yang lebih rendah dari kromosom tanaman kontrol. Variasi jumlah kromosom yang dihasilkan disebabkan karena kolkisin diserap oleh eksplan berbeda. Tidak semua tunas yang dihasilkan terbukti mengalami penambahan jumlah kromosom. Handro et al (1993) mengemukakan bahwa dalam satu agregat kalus yang telah direndam dengan kolkisin 10-3 M selama 72 hari, menunjukan perbedaan karakteristik dari bentuk dan jumlah ploidi sel didalamnya. tergantung pada kekompakan agregatnya. Jumlah kromosom sel nilam Pogostemon cablin Benth. dapat disajikan pada Gambar 4.
Kimera
22
parsial akibatnya tanaman mengalami malformasi. Blakeslee dan Avery (1937) melaporkan bahwa perlakuan kolkisin 0.4% selama satu hari pada biji Datura dapat menginduksi kimera dan mengakibatkan deformasi yang bervariasi pada tanaman akibat perubahan jumlah kromosom, daun mengalami malformasi, permukaan daun kasar dan batang yang kerdil.
Perendaman kolkisin dilakukan pada stek buku tunggal tanaman nilam. Stek buku tunggal memiliki potensi jaringan yang tidak aktif membelah lebih besar dibandingkan dengan jaringan meristematik. Jaringan meristemastik biasanya terdapat pada ujung-ujung bagian tanaman. Hal ini menyebabkan banyak ditemukan adanya kimera pada penelitian ini. Pada sunkultur kedua masih ditemukan adanya kimera baik secara fenotipik maupun secara genotipik.
Gambar 5,6,7,8,9,10 dan 11 dibawah ini menunjukkan kimera yang ditemukan selama penelitian berlangsung. Pada Gambar 5 merupakan kimera yang terjadi dalam sistem pembentukan tunas aksilar nilam Pogostemon cablin Benth. Gambar 6 menunjukkan terbentuknya tunas nilam Pogostemon cablin Benth yang kerdil tanpa akar. Gambar 7 sampai 9 menunjukkan adanya kimera pada jaringan akar planlet nilam Pogostemon cablin Benth yang menghasilkan stomata dengan ukuran stomata dan jumlah kloroplas yang beragam. Gambar 10 dan 11 menunjukkan jumlah kromosom nilam Pogostemon cablin Benth yang berbeda pada 2 sel yang berbeda pada satu organ akar.
23
Gambar 6: Planlet kerdil nilam Pogostemon cablin Benth. perlakuan konsentrasi 0.02% lama perendaman 72 jam.
Gambar 7 : Kimera tingkat jaringan pada kloroplas dengan perlakuan A: kolkisin 0.02% lama perendaman 48 jam B: kolkisin 0.02% lama perendaman 72 jam
Gambar 8 : Kimera tingkat jaringan pada kloroplas dengan perlakuan A : kolkisin 0.04% lama perendaman 24 jam B: kolkisin 0.04% lama perendaman 48 jam
B
B
A
24
Gambar 9 : Kimera tingkat jaringan pada kloroplas dengan perlakuan kolkisin 0.06% lama perendaman 24 jam
Gambar 10 : Kimera tingkat organ pada kromosom dengan perlakuan A: kolkisin 0.02% lama perendaman 48 jam B: kolkisin 0.02% lama perendaman 72 jam
Gambar 11 : Kimera tingkat jaringan pada kromosom dengan perlakuan kolkisin 0.06% lama perendaman 24 jam
25
KESIMPULAN
Tunas nilam Pogostemon cablin Benth. hasil induksi dengan kolkisin pada subkultur pertama menunjukkan perbedaan kemampuan dalam membentuk akar pada media perakaran IAA 0.1 mg/l maupun pada 0.5 mg/l. Pemberian IAA 0.1 mg/l menginduksi perakaran lebih baik dibandingkan konsentrasi IAA 0.5 mg/l. Pada subkultur kedu konsentrasi kolkisin berpengaruh nyata pada peubah jumlah tunas pada minggu 8 dan 9 setelah tanam. Perlakuan kontrol memiliki jumlah tunas tertinggi namun tidak berbeda nyata dengan perlakuan kolkisin 0.02% dan 0.04%. Perlakuan kolkisin dengan nilai jumlah tunas terendah adalah pada konsentrasi 0.06%. Perlakuan kolkisin berpengaruh nyata terhadap jumlah buku pada minggu 3 dan 10 setelah tanam. Konsentrasi kolkisin 0.06% mempunyai nilai paling rendah dibandingkan dengan perlakuan lain. Perlakuan kontrol memiliki jumlah buku tertinggi namun tidak berbeda nyata dengan perlakuan kolkisin 0.02% dan 0.04%. Pada peubah jumlah daun dan jumlah akar kolkisin tidak memberikan pengaruh yang nyata.
Hasil uji sitologi menunjukan bahwa kolkisin dapat menginduksi terjadinya mutasi kromosom pada sel tunas nilam secara in vitro. Pengamatan ukuran stomata menunjukkan konsentrasi kolkisin 0.02% selama 48 jam memiliki ukuran yang paling besar sedangkan untuk ukuran paling kecil adalah perlakuan kolkisin 0.06% selama 24 jam. Pada peubah jumlah kloroplas nilai tertinggi ada pada konsentrasi 0.04% dengan lama perendaman 48 jam dan terendah adalah perlakuan kontrol. Kloroplas dari planlet yang mendapat perlakuan kolkisin memiliki jumlah yang lebih tinggi dari jumlah kontrolnya. Perlakuan yang menghasilkan planlet dengan jumlah kromosom melebihi jumlah diploidnya adalah perlakuan perendaman 0.02% kolkisin selama 48 jam. 0.04% kolkisin selama 24 jam. kolkisin 0.04% selama 48 jam dan kolkisin 0.06% selama 24 jam. Jumlah kromosom pada planlet yang dihasilkan beragam. dari 18 hingga 106 kromosom per sel. Pada subkultur kedua masih ditemukan adanya kimera baik secara fenotipik maupun genotipik.
DAFTAR PUSTAKA
Aisyah, Syarifah Iis. 2006. Mutasi induksi. hal 159-178. S. Sastrosumarjo (Ed). Sitogenetika Tanaman. Departemen Agronomi dan Hortikultura IPB. Bogor. 268 hal.
Ajijah,N. dan N. Bermawie. 2003. Pengaruh Kolkisin Terhadap Pertumbuhan dan Produksi Dua Tipe Kencur (Kaempferia galanga Linn). Buletin Tanaman Rempah dan Obat 16(1):46-55.
Anne, Y.P. 2012. Induksi Mutasi Melalui Penggandaan Kromosom Nilam (Pogostemon cablin Benth.) Varietas Sindikalang dengan Penggunaan Kolkisin secara In Vitro. Skripsi. Institut Pertanian Bogor. 48 hal
26
Crowder, L. V. 1993. Genetika Tumbuhan (Diterjemahkan oleh L. Kustiarti). Gajah Mada University Press. Yogyakarta. 499 hal.
Damayanti, F. dan I. Mariska. 2003. Induksi poliploidi dengan kolkisin pada hibrida F hasil persilangan antar spesies pada tanaman panili asal Ciamis. Berita Biologi 6(4):589-594.
Direktorat Jendral Perkebunan. 2009. Komoditian Tanaman Nilam. http://ditjenbun.litbang.deptan.go.id. (14 desember 2012)
Direktorat Budidaya Tanaman Semusim. 2010. Komoditan Nilam. http://ditjenbun.deptan.go.id/budtansim/. (15 Januari 2013)
Eigsti, O. J. and P, Dustin. 1957. Colchicine in Agriculture. Medicine. Biology and Chemistry. The Lowa State Collage Press. Ames. Lowa. USA. 470p. Gembong Tjitrosoepomo. 2000. Taksonomi Tumbuhan (Spermatophyta).
Yogyakarta: Gadjah Mada University Press. 906 hal.
Guenther, E. 1988. Susunan kimia, sumber dan kegunaan minyak atsiri dalam kehidupan tanaman. 91-99 dalam S.Ketaren. Minyak Atsiri.Direktorat Jendral Pendidikan Tinggi Departemen Pendidikan dan Kebudayaan. Jakarta.
Gunawan, L. W. 1992. Teknik Kultur Jaringan Tanaman. Departemen Pendidikan Tinggi. Pusat Antar Universitas Bioteknologi Institut Pertanian Bogor. Bogor. 165 hal. Induction and identification of polyploidy in basil (Ocimum basilicum L.) medicinal plant by colchicine treatment. International J. of Plant Production 4(2):87- 98.
Poespodarsono, S. 1988. Dasar-dasar Ilmu Pemuliaan Tanaman. PAU - LSI. IPB.Bogor. 169hal.
Poehlman, J. M. and D. A. Sleper. 1995. Breeding Field Crops 4. Iowa State University Press. Ames. 49p.
Sakhanokho, H. F.. K. Rajasekaran. R. Y. Kelley and N. Islam-Faridi. 2009. Induced polyploidy in diploid ginger (Hedycium muluense R. M. Smith) using colchicine and oryzalin. HortSci 44(7):1809-1814.
Soeranto, H. 2003. Peran iptek nuklir dalam pemuliaan tanaman untuk mendukung industri pertanian. Puslitbang Teknologi Isotop dan Radiasi. Badan Tenaga Nuklir Nasional. Jakarta.
Suryo. 1995. Sitogenetika. Gajah Mada University Press. Yogyakarta. 446 hal. Titik, S. dan Endang. S. 2002. Budidaya dan Penyulingan Nilam. Jakarta:
Penebar Swadaya.
Tambon, J.T.. V.T. Sapra and S. Garton. 1998. In Vitro induction of tetraploids in colchicine-treated cocoyam plantlets. Euphytica 104: 191 - 195.
27
Usman, M..B. Fatima. K. A. Gillani.M. S. Khan. and M. M. Khan. 2008. Exploitation of potential target tissue to develop polyploids in Citrus. Pakistan Journal of Botany 40(4):1755-1766.
Yunita, R. 2009. Pemanfaatan variasi somaklonal dan seleksi in vitro dalam perakitan tanaman toleran cekaman abiotik. http://pustaka.litbang.deptan.go.id . [12maret2012]
28
Lampiran 1 Komposisi Media Murashige-Skoog (Gunawan, 1992)
stok Jenis Senyawa
Konsentrasi Larutan Stok
(g/l)
Volume dalam Media
(ml/l)
Konsetrasi Senyawa dalam Media
(ppm)
A NH4NO3 82.500 20 1650.000
B KNO3 95.000 20 1900.000
C KH2PO4 34.000 5 170.000
H3BO3 1.240 6.200
KI 0.166 0.830
NaMoO4.2H2O 0.050 0.250
CoCl.6H2O 0.005 0.025
D CaCl.2H2O 88.000 5 440.000
E MgSO4.7H2O 74.000 5 370.000
MgSO4.4H2O 4.460 22.300
ZnSO4.7H2O 1.720 8.600
CuSO4.5H2O 0.005 0.025
F Na2EDTA.2H2O 3.730 10 37.300
FeSO4.7H2O 2.780 27.800
Myo Myo-Inositol 10.000 10 100.000
Vitamin Thiamin 0.010 10 0.100
Niacin 0.050 0.500
Pyridoxine 0.050 0.500
Glycin 0.200 2.000
29
Lampiran 2 Sidik ragam pengaruh kolkisin dan media terhadap jumlah akar nilam pada Percobaan I
Umur Planlet (MST)
Sumber
Keragaman Db JK KT Hit F- Pr>F R2 KK (%) 3 Media 1 0.1061 0.2497 1.45 0.2393 0.6244 77.14
Kolkisin 7 2.5564 0.3761 4.98 0.0010 Interaksi 7 0.4085 0.8170 1.11 0.3761
4 Media 1 1.1881 1.1881 2.56 0.1212 0.5015 71.42 Kolkisin 7 8.1339 1.1619 2.50 0.0404
Interaksi 7 3.3948 0.6789 1.46 0.2344
5 Media 1 1.8264 1.8264 2.20 0.1507 0.500 61.19 Kolkisin 7 14.043 2.0061 2.41 0.0492
Interaksi 7 4.5765 1.1441 1.38 0.2703
6 Media 1 4.7755 4.7755 5.36 0.0290 0.5380 46.11 Kolkisin 7 13.094 1.8706 2.10 0.0812
Interaksi 7 5.8802 1.4700 1.65 0.1928
7 Media 1 9.5068 9.5068 7.51 0.0117 0.4790 45.02 Kolkisin 7 9.8064 1.4009 1.11 0.3921
Interaksi 7 3.0007 1.0002 0.79 0.5118
8 Media 1 7.5714 7.5714 6.39 0.0188 0.4944 37.87 Kolkisin 7 9.2442 1.3206 1.12 0.3871
Interaksi 7 5.0375 1.6791 1.42 0.2630
9 Media 1 14.706 14.7060 9.64 0.0050 0.5049 39.16 Kolkisin 7 9.8256 1.4036 0.92 0.5095
Interaksi 7 4.1407 1.3802 0.90 0.4542
10 Media 1 8.9326 8.9325 5.94 0.0233 0.5269 36.72 Kolkisin 7 12.250 1.7500 1.16 0.3621
30
Lampiran 3 Sidik ragam pengaruh kokisin terhadap jumlah tunas nilam pada Percobaan II
Umur Planlet (MST)
Sumber
Keragaman db JK KT Hit F- Pr>F R2 KK (%) 1 Kolkisin 6 0.855 0.142 0.45 0.8286 0.1981 126.4
Galat 11 3.459 0.314 Koreksi 17 4.016
2 Kolkisin 6 1.047 0.174 0.47 0.8185 0.2033 78.72 Galat 11 4.106 0.373
Koreksi 17 5.154
3 Kolkisin 6 2.452 0.408 1.48 0.2709 0.4467 43.51 Galat 11 3.038 0.276
Koreksi 17 5.490
4 Kolkisin 6 0.634 0.126 0.38 0.8503 0.1482 36.71 Galat 11 3.644 0.331
Koreksi 17 4.279
5 Kolkisin 6 0.876 0.175 0.42 0.8264 0.1598 33.96 Galat 11 4.607 0.418
Koreksi 17 5.484
6 Kolkisin 6 0.718 0.143 0.47 0.7919 0.1756 24.32 Galat 11 3.372 0.306
Koreksi 17 4.090
7 Kolkisin 6 1.101 0.220 0.93 0.4991 0.2966 18.05 Galat 11 2.612 0.237
Koreksi 17 3.174
8 Kolkisin 6 1.306 0.261 1.76 0.0202 0.4441 12.47 Galat 11 1.635 0.148
Koreksi 17 2.942
9 Kolkisin 6 2.025 0.405 3.87 0.0286 0.6377 9.398 Galat 11 1.150 0.104
Koreksi 17 3.175
10 Kolkisin 6 2.281 0.456 7.26 0.0031 0.7674 6.784 Galat 11 0.691 0.062
31
Lampiran 4 Sidik ragam pengaruh kolkisin terhadap jumlah buku nilam pada Percobaan II
Umur Planlet (MST)
Sumber
Keragaman Db JK KT F-Hit Pr>F R2 KK (%) 1 Kolkisin 6 1.927 0.321 0.30 0.9264 0.138 101.21
Galat 11 11.94 1.085 Koreksi 17 12.56
2 Kolkisin 6 3.630 0.605 0.33 0.9065 0.153 81.298 Galat 11 20.07 1.824
Koreksi 17 23.07
3 Kolkisin 6 8.946 1.491 0.88 0.0386 0.324 49.456 Galat 11 18.59 1.690
Koreksi 17 27.54
4 Kolkisin 6 1.722 0.344 0.25 0.9324 0.101 31.531 Galat 11 15.30 1.391
Koreksi 17 17.13
5 Kolkisin 6 2.202 0.440 0.44 0.8116 0.166 21.435 Galat 11 11.00 1.002
Koreksi 17 13.20
6 Kolkisin 6 2.277 0.455 0.58 0.7175 0.207 15.983 Galat 11 8.690 0.790
Koreksi 17 10.96
7 Kolkisin 6 2.574 0.514 0.92 0.5036 0.294 11.671 Galat 11 6.160 0.560
Koreksi 17 8.735
8 Kolkisin 6 3.161 0.632 1.39 0.3023 0.386 9.3151 Galat 11 5.021 0.456
Koreksi 17 8.183
9 Kolkisin 6 3.427 0.685 1.60 0.2402 0.420 8.1562 Galat 11 4.7219 0.429
Koreksi 17 8.1491
10 Kolkisin 6 6.827 1.365 2.24 0.0123 0.504 8.8045 Galat 11 6.7179 0.610
32
Lampiran 5 Sidik ragam pengaruh kolkisin terhadap jumlah daun nilam pada Percobaan II
Umur Planlet (MST)
Sumber
Keragaman db JK KT F-Hit Pr>F R2 KK (%) 1 Kolkisin 6 7.268 1.211 0.29 0.9273 0.138 101.04
Galat 11 45.29 4.118 Koreksi 17 52.56
2 Kolkisin 6 14.04 2.340 0.34 0.9009 0.156 80.814 Galat 11 75.49 6.863
Koreksi 17 89.53
3 Kolkisin 6 33.70 5.616 0.82 0.5795 0.307 50.624 Galat 11 75.72 6.883
Koreksi 17 109.4
4 Kolkisin 6 7.314 1.462 0.25 0.9306 0.102 32.774 Galat 11 64.12 5.829
Koreksi 17 71.43
5 Kolkisin 6 9.809 1.961 0.47 0.7941 0.174 22.189 Galat 11 46.32 4.211
Koreksi 17 56.13
6 Kolkisin 6 8.196 1.639 0.45 0.8081 0.168 17.448 Galat 11 40.48 3.680
Koreksi 17 48.67
7 Kolkisin 6 9.221 1.844 0.67 0.6562 0.232 12.985 Galat 11 30.39 2.763
Koreksi 17 39.61
8 Kolkisin 6 11.85 2.371 1.11 0.4107 0.334 10.106 Galat 11 23.58 2.144
Koreksi 17 35.44
9 Kolkisin 6 12.89 2.578 1.10 0.4135 0.333 9.4843 Galat 11 25.78 2.344
Koreksi 17 38.68
10 Kolkisin 6 23.55 4.711 1.87 0.1801 0.459 8.9027 Galat 11 27.42 2.522
33
Lampiran 6 Sidik ragam pengaruh kolkisin terhadap jumlah akar nilam pada Percobaan II
Umur Planlet (MST)
Sumber
Keragaman db JK KT F-Hit Pr>F R2 KK (%) 1 Kolkisin 6 0.000 0.000 0.61 0.7177 0.250 313.04
Galat 11 0.000 0.000 Koreksi 17 0.002
2 Kolkisin 6 0.007 0.001 1.05 0.4446 0.3644 332.29 Galat 11 0.013 0.001
Koreksi 17 0.122
3 Kolkisin 6 0.096 0.016 0.63 0.7039 0.255 124.15 Galat 11 0.281 0.025
Koreksi 17 0.344
4 Kolkisin 6 2.358 0.393 2.17 0.1255 0.542 72.36 Galat 11 1.990 0.180
Koreksi 17 2.113
5 Kolkisin 6 3.847 0.641 2.31 0.1088 0.557 43.987 Galat 11 3.056 0.227
Koreksi 17 4.007
6 Kolkisin 6 2.075 0.345 1.21 0.3691 0.398 31.940 Galat 11 3.137 0.285
Koreksi 17 4.665
7 Kolkisin 6 4.296 0.716 1.15 0.3972 0.3853 38.348 Galat 11 6.853 0.623
Koreksi 17 7.336
8 Kolkisin 6 3.194 0.532 0.52 0.7841 0.2200 42.923 Galat 11 11.32 1.029
Koreksi 17 14.51
9 Kolkisin 6 2.725 0.454 0.33 0.9091 0.1513 45.801 Galat 11 15.28 1.389
Koreksi 17 18.00
10 Kolkisin 6 2.467 0.411 0.24 0.9526 0.1168 48.156 Galat 11 18.64 1.690
34
RIWAYAT HIDUP
Penulis lahir pada tanggal 20, Februari 1991 di kota Blitar Jawa Timur dan merupakan anak pertama dari tiga bersaudara. Sejak berusia satu tahun penulis tinggal dan dibesarkan di kota Bojonegoro Jawa Timur. Pada tahun 2006 penulis melanjutkan pendidikannya di SMA Negeri 1 Bojonegoro. Selama pendidikan Sekolah Menengah Atas penulis aktif diprogram Olimpiade Sains Nasional dalam bidang Geo Sience. Penulis menjadi juara 3 dalam Olimpiade Sains Nasional Geo Sience tingkat kabupaten Bojonegoro dan mewakili kabupaten dalam Olimpiade Sains Nasional tingkat provinsi Jawa Timur. Penulis juga aktif dalam regu paduan suara kabupaten Bojonegoro.