ANALISIS GENETIK KARAKTER KUANTITATIF DAN
PECAH BUAH PADA TANAMAN TOMAT
(
Lycopersicon esculentum
Mill.)
SRI WAHYUNI
A253110021
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis “Analisis Genetik Karakter Kuantitatif dan Pecah Buah pada Tanaman Tomat (Lycopersicon esculentum Mill.)” adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Februari 2014
Sri Wahyuni
NIM A253110021
RINGKASAN
SRI WAHYUNI. Analisis Genetik Karakter Kuantitatif dan Pecah Buah pada Tanaman Tomat (Lycopersicon esculentum Mill.). Dibimbing oleh MUHAMAD SYUKUR, JOKO RIDHO WITONO dan SYARIFAH IIS AISYAH.
Pecah buah menjadi salah satu kendala pada budidaya tanaman tomat di dataran rendah. Dua puluh lima genotipe tomat telah dikarakterisasi untuk mengevaluasi tingkat toleransi terhadap pecah buah dan keragaman genetiknya. Selanjutnya, sepuluh genotipe dipilih untuk memperoleh informasi penanda seleksi pemilihan bahan tetua. Enam genotipe tomat dipilih sebagai tetua untuk membentuk populasi dialel 6×6 yang digunakan untuk menduga parameter genetik, daya gabung, heterosis dan keragaan hibrida. Penelitian dimulai dari April hingga Agustus 2012 dan Maret hingga Juli 2013 di Laboratorium Pemuliaan Tanaman, Fakultas Pertanian IPB dan Kebun Percobaan Leuwikopo (6o56‟34‟‟S, 106o72‟ 56‟‟E) IPB. Percobaan disusun dalam rancangan kelompok lengkap teracak faktor tunggal dengan 3 ulangan. Ketahanan terhadap pecah buah dievaluasi dengan indeks pecah buah dan kriteria ketahanan. Keragaman genetik dianalisis dengan analisis komponen utama dan analisis gerombol. Penanda/kriteria seleksi dianalisis dengan sidik lintas. Parameter genetik diduga dengan metode Hayman dan daya gabung dengan metode Griffing. Pendugaan nilai heterosis hibrida tomat dianalisis berdasarkan nilai tengah kedua tetuanya dan nilai tengah tetua terbaik. Pendugaan keragaan hibrida terbaik diuji dengan uji Dunnet.
Hasil penelitian menunjukkan indeks pecah buah 25 genotipe tomat berkisar 0.00-20.58. Berdasarkan karakter kuantitatif terdapat beberapa genotipe yang memiliki kriteria yang dapat dijadikan sebagai bahan tetua pada pemuliaan tanaman tomat. Hasil analisis komponen utama dan analisis gerombol menunjukkan genotipe tomat mengelompok menjadi empat kelompok. Genotipe-genotipe tomat IPBT1, IPBT3, IPBT13, IPT33, IPBT60, IPBT64, IPBT73, IPBT78, IPBT83 dan IPBT86 direkomendasikan sebagai bahan tetua persilangan dialel untuk analisis genetik karakter pecah buah.
Karakter pecah buah dipengaruhi oleh ragam aditif dan dominan, akan tetapi ragam dominan lebih berperan dalam mengendalikan karakter pecah buah. Karakter hasil dan komponen hasil, karakter buah dan karakter vegetatif pada tanaman tomat di dataran rendah dipengaruhi ragam aditif dan ragam dominan, akan tetapi pengaruh ragam aditif lebih penting dalam mengendalikan karakter tersebut. Pengaruh tetua betina terjadi pada karakter hasil pertanaman, jumlah buah, bobot per buah, panjang buah, diameter buah, jumlah rongga dan tebal daging buah. Ragam DGU dan DGK berpengaruh nyata pada semua karakter yang diamati. Program perakitan varietas galur murni dapat dikembangkan jika ragam DGU berpengaruh nyata dan genotipe memiliki nilai duga DGU yang baik. Program perakitan varietas hibrida dapat dilakukan jika ragam DGK berpengaruh nyata dan memiliki nilai duga DGK yang baik. Genotipe IPBT78 merupakan tetua dengan DGU terbaik pada karakter hasil per tanaman, bobot per buah, panjang buah dan tebal daging buah sehingga direkomendasikan untuk dikembangkan sebagai varietas galur murni. Kombinasi persilangan IPBT3×IPBT78, IPBT78×IPBT3, IPBT13×IPBT78 dan IPBT64× IPBT78 direkomendasikan untuk diuji lebih lanjut dan dikembangkan sebagai varietas hibrida.
Hibrida hasil persilangan dialel mempunyai karakter melebihi varietas pembanding maupun tidak berbeda nyata dengan pembanding terbaiknya sehingga berpotensi dilakukan pengujian lebih lanjut untuk memperoleh informasi adaptasi dan preferensi konsumen. Beberapa hibrida yang berpotensi untuk dikembangkan lebih lanjut adalah IPBT3×IPBT78, IPBT78×IPBT3, IPBT78 ×IPBT13, IPBT78×IPBT64.
SUMMARY
SRI WAHYUNI. Genetic Analysis of Quantitative and Fruit Cracking Characters in Tomato (Lycopersicon esculentum Mill.). Supervised by MUHAMAD SYUKUR, JOKO RIDHO WITONO and SYARIFAH IIS AISYAH
Tomato fruit cracking had became one of the problems in lowland cultivation. Morphology characterization of twenty five tomato genotypes were carried out to evaluate the fruit cracking tolerance level and the genetic diversity. Ten genotypes were selected to estimate variance and heritabilities of crack index character, yield and component of yield, fruit and vegetative character and then six genotypes were selected to conduct a 6×6 full diallel cross set of tomato to estimate genetic parameter, combining ability and hybrid performance. The experiment was conducted from April until August 2012 and from April until August 2013 at Leuwikopo Field Station (6o56‟34‟‟S, 106o72‟56‟‟E), Bogor Agricultural University. Randomized complete block design was used with three replications. The genetic diversity was analyzed by clustering analysis. Marker character were selected based on heritabilities value and correlation between character. Genetic parameter was analized by Hayman method and combining ability was analyzed by Griffing Method. The difference between the hybrids and varieties comercial was analyzed by Dunnet‟s tested.
Result of resistance evaluation showed that fruit crack index of 25 tomato genotypes range from 0.00 to 20.58. According to quantitative character some genotype can be selected as material breeding. Based on genetic diversity analysis, all genotypes could be divided into four groups. Tomato genotypes IPBT1, IPBT3, IPBT13, IPT33, IPBT60, IPBT64, IPBT73, IPBT 78, IPBT83 dan IPBT86 were recommended as parent of diallel cross to fruit cracking genetic analysis.
The marker selection on tomato breeding program to develop tomato varieties that have high yield and fruit cracking resistant using characters with high heritabilities value, correlated significantly and directly influent to the fruit crack index. Fruit cracking character, yield and components of yield, fruit character and vegetative character include to the high category of heritability. According correlation and path analysis the three components very highly correlated with crack index were fruit crack weight percentage of plant, number of fruit crack percentage of plant and total soluble solids. The three components that very highly correlated with yield per plant were marketable fruit weight, individual fruit weight and plant height.
non-additive gene effects for all of the character were highly significant. The highest significant reciprocal effect occurs for yield per plant, number of fruit, individual fruit weight, fruit length, fruit diameter, number of locule and fruit thickness. The analysis of variance for general combining ability (GCA) and specific combining ability (SCA) were significant for all traits. Pure line varieties breeding programs could be developed if variance for GCA were highly significant and the genotype parent has a good estimated GCA value. Tomato genotype IPBT78 proved to be the best general combiner for yield and yield components. Hybrid varieties breeding programs developed if variance for SCA were highly significant and the genotype parent has a good estimated SCA value. The most recommended cross combinations are IPBT3×IPBT78, IPBT78×IPBT3, IPBT13×IPBT78 and IPBT64×IPBT78.
Hybrids performance show that some character are better than commercial variety or have similar performance to the commercial variety. Therefore, it is highly suggested for further test to obtain information of adaptation and preference of consumers. Tomato genotype IPBT78 proved to be the best general combiner for yield and yield components. The most recommended cross combinations are IPBT3× IPBT78, IPBT78×IPBT3, IPBT13×IPBT78 and IPBT64×IPBT78.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta dilindungi Undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa
mencantumkan atau menyebutkan sumbernya. Pengutipan hanya
untuk kepentingan pendidikan, penelitian, penulisan karya
ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu
masalah; dan pengutipan tersebut tidak merugikan kepentingan
IPB
ANALISIS GENETIK KARAKTER KUANTITATIF DAN
PECAH BUAH PADA TANAMAN TOMAT
(
Lycopersicon esculentum
Mill.)
SRI WAHYUNI
A253110021
Tesis
Sebagai salah satu syarat untuk memperoleh gelar
Magisters Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis :Analisis Genetik Karakter Kuantitatif dan Pecah Buah pada Tanaman Tomat (Lycopersicon esculentum Mill.)
Nama : Sri Wahyuni
NIM : A253110021
Disetujui oleh Komisi Pembimbing
Prof Dr Muhamad Syukur, SP MSi Ketua
Dr Ir Joko Ridho Witono, MSi Anggota
Dr Ir Syarifah Iis Aisyah, MScAgr Anggota
Diketahui oleh
Ketua Program Studi
Pemuliaan dan Bioteknologi Tanaman
Dr Ir Yudiwanti Wahyu EK, MS
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
Tanaman Tomat (Lycopersicon esculentum Mill.) Nama : Sri Wahyuni
NIM : A25311 0021
Disetujui oleh
セGエQBャGエZQGエuャ「ゥョァ@
Ketua
Dr Ir Joko Ridho Witono, MSi Dr Ir Syarifah lis Aisyah, MScAgr
Anggota Anggota
Diketahui oleh
Ketua Program Studi
Pemuliaan dan Bioteknologi Tanaman
Dr Ir Yudiwa i Wahyu EK, MS
PRAKATA
Puji syukur penulis panjatkan ke hadirat Allah SWT atas segala rahmat dan karunia-Nya sehingga penelitian tentang analisis genetik karakter pecah buah dan karakter kuantitatif pada tanaman tomat ini dapat diselesaikan. Tomat merupakan komoditas hortikultura unggulan yang bernilai ekonomis dan strategis di Indonesia. Upaya peningkatan produksi perlu terus dilakukan dalam rangka mendukung program ketahanan pangan. Kajian tentang pola pewarisan karakter pecah buah dan karakter agronomi lainnya sangat membantu dalam rangka perakitan varietas, baik varietas galur murni atau varietas hibrida di dataran rendah.
Pada kesempatan ini penulis menyampaikan penghargaan dan ucapan terima kasih kepada:
1 Kementerian Riset dan Teknologi selaku pemberi beasiswa melalui Program Beasiswa Pascasarjana Kementerian Riset dan Teknologi tahun 2011-2013.
2 Kepala Pusat Konservasi Tumbuhan Kebun Raya Bogor, LIPI atas izin studi yang diberikan.
3 Prof Dr Muhamad Syukur, SP MSi, Dr Ir Joko Ridho Witono, MSi, Dr Ir Syarifah Iis Aisyah, MScAgr dan Dr Rahmi Yunianti, SP, MSi (Almh) selaku komisi pembimbing atas segala bimbingan, arahan, kritik dan masukan selama penelitian hingga penulisan tesis ini.
4 DIKTI yang telah memberikan dana penelitian melalui Hibah Bersaing Tahun 2012 a.n. Dr Rahmi Yunianti, SP, MSi dan Hibah Unggulan Perguruan Tinggi tahun 2013 a.n. Dr Ir Syarifah Iis Aisyah, MScAgr. 5 Prof Dr Surjono Hadi Sutjahjo, MS selaku penguji luar komisi pada
ujian akhir tesis atas saran dan arahan untuk perbaikan tesis.
6 Dr Ir Yudiwanti Wahyu EK, MS selaku dosen penguji perwakilan dari Program Studi Pemuliaan dan Bioteknologi Tanaman pada ujian akhir tesis atas saran dan arahan untuk perbaikan tesis.
7 Seluruh staf pengajar Program Studi Pemuliaan dan Bioteknologi Tanaman atas ilmu dan pengetahuan yang diberikan selama kuliah. 8 Keluarga tercinta, Ibu Sarmi Suhudi, Ibu Hj. Sri Haryanti, Bapak Suhudi
(Alm), Bapak H. Khoerudin, suami Khairul Anwar dan anak Muhammad Ayyasy Anwar, dan seluruh keluarga besar yang telah memberikan dukungan moril maupun materiil kepada penulis dalam menempuh studi dan penelitian.
9 Bapak-Ibu dan rekan-rekan di Pusat Konservasi Tumbuhan Kebun Raya Bogor, LIPI atas dukungannya selama penulis menempuh studi.
10 Bapak Darwa, rekan-rekan PBT angkatan 2011 dan rekan-rekan di Laboratorium Pemuliaan Tanaman Departemen Agronomi dan Hortikultura IPB terutama Ibu Tri Budiyanti, Mas Undang, Marlina Mustafa, Helfi Eka Saputra, Muhammad Ridha Alfarabi Istiqlal, Abdul Hakim, Tiara Yudilastari, Arya Widura R, Estriana Riti, Vitria Rahadi, Nura, Eka MDR dan Faradilla serta keluarga Ibu Anih/Bapak Heri, atas bantuan dan kerjasama selama studi dan penelitian.
DAFTAR ISI
DAFTAR TABEL xii
DAFTAR GAMBAR xiv
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitia 2
Hipotesis Penelitian 2
Kerangka Pemikiran 3
2 TINJAUAN PUSTAKA 4
2.1 Potensi Tanaman Tomat 4
2.2 Kualitas Buah 5
2.3 Pecah Buah pada Tomat 5
2.4 Persilangan Dialel 10
3 KETAHANAN 25 GENOTIPE TOMAT (Lycopersicon esculentum Mill.) TERHADAP PECAH BUAH DAN KERAGAMAN
GENETIKNYA 13
Pendahuluan 14
Bahan dan Metode 15
Hasil dan Pembahasan 23
Simpulan 33
4 PEMILIHAN PENANDA SELEKSI UNTUK PERAKITAN TOMAT (Lycopersicon esculentum Mill.) TOLERAN
TERHADAP PECAH BUAH 34
Pendahuluan 35
Bahan dan Metode 36
Hasil dan Pembahasan 40
Simpulan 48
5 PENDUGAAN DAYA GABUNG DAN PARAMATER GENETIK KARAKTER PECAH BUAH DAN BEBERAPA KARAKTER
AGRONOMI PADA TOMAT (Lycopersicon esculentum Mill.) 48
Pendahuluan 49
Bahan dan Metode 50
Hasil dan Pembahasan 60
Simpulan 80
6 EVALUASI 30 HIBRIDA TOMAT ADAPTIF DATARAN
RENDAH 81
Pendahuluan 82
Bahan dan Metode 82
Hasil dan Pembahasan 86
DAFTAR ISI (lanjutan)
7 PEMBAHASAN UMUM 92
Kondisi Umum Penelitian 92
Pembahasan Umum Hasil Penelitian 93
DAFTAR PUSTAKA 100
DAFTAR TABEL
1 Daftar genotipe tomat yang digunakan dalam penelitian 15
2 Sumber keragaman dan nilai harapan 19
3 Nilai indeks pecah buah pada 25 genotipe tomat dan tingkat
ketahanannya 24
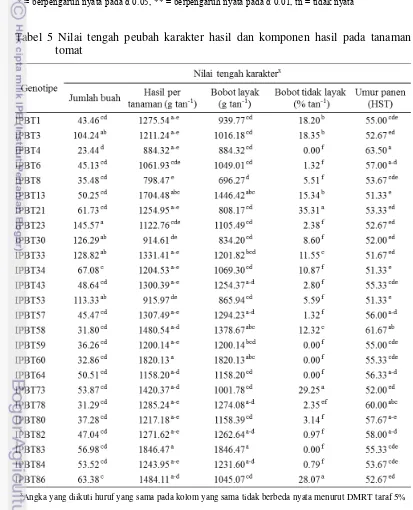
4 Kuadrat tengah karakter indeks pecah buah dan komponen hasil
pada tanaman tomat 25
5 Nilai tengah peubah karakter indeks pecah buah dan komponen
hasil pada tanaman tomat 25
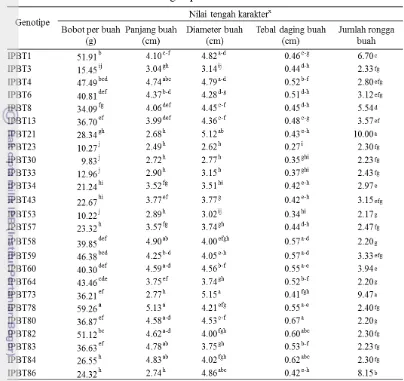
6 Kuadrat tengah karakter buah tomat 26
7 Nilai tengah peubah karakter buah tomat 27
8 Kuadrat tengah karakter vegetatif tanaman tomat 27 9 Nilai tengah peubah karakter vegetatif tanaman tomat 28 10 Nilai akar ciri komponen utama berdasarkan analisis komponen
utama 29
11 Nilai vektor ciri tiga komponen utama 30
12 Sumber keragaman dan nilai harapan 39
13 Kuadrat tengah karakter pecah buah dan komponen hasil pada
tanaman tomat 40
14 Kuadrat tengah karakter buah tomat 40
15 Kuadrat tengah karakter vegetatif tanaman tomat 41 16 Nilai duga komponen ragam dan heritabilitas karakter indeks pecah
buah, komponen hasil, buah dan vegetatif pada tanaman tomat 42 17 Nilai korelasi antar karakter terhadap karakter indeks pecah buah
pada tanaman tomat 43
18 Pengaruh langsung dan tidak langsung masing-masing peubah terhadap indeks pecah buah pada tanaman tomat 44 19 Nilai korelasi antar karakter terhadap karakter hasil per tanaman
tomat 46
20 Pengaruh langsung dan tidak langsung masing-masing peubah
terhadap hasil per tanaman tomat 47
21 Bagan persilangan dialel dengan 6 tetua 51
22 Komponen analisis ragam pada analisis dialel 55 23 Persilangan setengah dialel karakter-karakter agronomi tomat di
dataran rendah 56
24 Komponen analisis ragam untuk daya gabung menggunakan
DAFTAR TABEL (lanjutan)
25 Kuadrat tengah pecah buah dan komponen hasil pada tanaman
tomat 60
26 Kuadrat tengah karakter buah tomat 60
27 Kuadrat tengah karakter vegetatif pada tanaman tomat 60 28 Nilai indeks pecah buah dan kriteria toleransi pada populasi tomat 61 29 Pendugaan parameter genetik karakter pecah buah dan beberapa
komponen hasil tomat menggunakan analisis silang dialel Metode
Hayman 62
30 Pendugaan parameter genetik karakter buah tomat menggunakan
analisis silang dialel Metode Hayman 64
31 Pendugaan parameter genetik karakter vegetatif tomat mengguna-
kan analisis silang dialel Metode Hayman 65
32 Sebaran Vr + Wr pada karakter pecah buah dan komponen hasil
pada tanaman tomat 65
33 Sebaran Vr + Wr pada karakter buah tomat 66 34 Sebaran Vr + Wr pada karakter vegetatif pada tanaman tomat 66 35 Kuadrat tengah DGU, DGK, resiprokal dominan dan nilai
koefisien keragaman karakter indeks pecah buah, hasil dan
komponen hasil pada tanaman tomat 71
36 Kuadrat tengah DGU, DGK, resiprokal dan nilai koefisien
keragaman karakter buah 71
37 Kuadrat tengah DGU, DGK, resiprokal dan nilai koefisien
keragaman karakter vegetatif pada tanaman tomat 72 38 Daya gabung umum karakter indeks pecah buah dan komponen
hasil pada tanaman buah 72
39 Daya gabung umum karakter buah tomat 73
40 Daya gabung umum karakter vegetatif pada tanaman tomat 73 41 Daya gabung khusus indeks pecah buah, hasil dan komponen hasil
pada tanaman tomat 74
42 Daya gabung hhusus karakter buah tomat 75
43 Daya gabung khusus karakter vegetatif pada tanaman tomat 76 44 Heterosis (MP) dan heterobeltiosis (HP) karakter indeks pecah
buah, hasil dan komponen hasil pada tanaman tomat 77 45 Heterosis (MP) dan heterobeltiosis (HP) karakter buah tomat 78 46 Heterosis (MP) dan heterobeltiosis (HP) karakter vegetatif pada
tanaman tomat 79
47 Sumber keragaman dan nilai harapan 85
48 Kuadrat tengah pecah buah karakter indeks pecah buah, hasil dan
komponen hasil pada tanaman tomat 86
49 Kuadrat tengah karakter buah tomat 87
50 Kuadrat tengah karakter vegetatif pada tanaman tomat 87 51 Rata-rata karakter indeks pecah buah, hasil dan komponen hasil
pada hibrida tomat 89
DAFTAR TABEL (lanjutan)
53 Nilai tengah karakter vegetatif hibrida tomat 91
54 Kondisi iklim pada tahun 2012 dan 2013 92
55 Kudrat tengah gabungan karakter indeks pecah buah tahun 2012
dan 2013 94
56 Nilai ragam, daya gabung dan heterosis terbaik hasil kombinasi persilangan tetua tomat karakter indeks pecah, hasil dan
komponen hasil 96
57 Nilai ragam, daya gabung dan heterosis terbaik hasil kombinasi persilangan tetua tomat karakter buah dan vegetatif 97 58 Keragaan hibrida tomat terbaik hasil persilangan dialel pada
karakter indeks pecah buah, hasil dan komponen hasil pada
tanaman tomat 98
59 Keragaan hibrida tomat terbaik hasil persilangan dialel pada karakter
buah tomat 98
60 Keragaan hibrida tomat terbaik hasil persilangan dialel pada
karakter vegetatif tanaman tomat 99
DAFTAR GAMBAR
1 Diagram alur penelitian 3
2 Skoring pecah buah pada tomat 17
3 Letak daun 20
4 Pembagian helai daun 20
5 Letak anak daun terhadap tulang daun utama 20
6 Tipe tandan bunga 20
7 Lapisan absisi pada tangkai buah 21
8 Bentuk buah dalam penampang membujur 21
9 Depresi buah pada ujung tangkai buah 22
10 Bentuk ujung buah 22
11 Jumlah rongga buah 22
12 “Bahu buah hijau” sebelum masak 23
13 Analisis komponen utama 25 genotipe tomat KU I dan KU II 31 14 Analisis komponen utama 25 genotipe tomat KU I dan KU III 31 15 Analisis komponen utama 25 genotipe tomat KU II dan KU III 32 16 Dendrogram hasil analisis gerombol 25 genotipe tomat 33 17 Diagram lintasan beberapa karakter dengan indeks pecah buah
pada genotipe tomat 45
18 Diagram lintasan beberapa karakter dengan hasil per tanaman pada
genotipe tomat 47
19 Genotipe tetua tomat untuk persilangan dialel penuh 50
20 Pecah buah pada tomat 51
DAFTAR GAMBAR (Lanjutan)
22 Hubungan peragam (Wr) dan ragam (Vr) karakter indeks pecah
buah pada tomat 63
23 Hubungan peragam (Wr) dan ragam (Vr) karakter hasil per
tanaman pada tomat 66
24 Hubungan peragam (Wr) dan ragam (Vr) karakter jumlah buah
pada tomat 67
25 Hubungan peragam (Wr) dan ragam (Vr) karakter bobot per buah
pada tomat 67
26 Hubungan peragam (Wr) dan ragam (Vr) karakter panjang buah
pada tomat 67
27 Hubungan peragam (Wr) dan ragam (Vr) karakter diameter buah
pada tomat 68
28 Hubungan peragam (Wr) dan ragam (Vr) karakter jumlah rongga
buah pada tomat 68
29 Hubungan peragam (Wr) dan ragam (Vr) karakter tebal daging
buah pada tomat 68
30 Hubungan peragam (Wr) dan ragam (Vr) karakter padatan total
terlarut pada tomat 69
31 Hubungan peragam (Wr) dan ragam (Vr) karakter tinggi tanaman
pada tomat 69
32 Hubungan peragam (Wr) dan ragam (Vr) karakter panjang daun
pada tomat 69
33 Hubungan peragam (Wr) dan ragam (Vr) karakter lebar daun pada
tomat 70
34 Genotipe tomat hibrida IPBT78×IPBT1 87
35 Genotipe tomat hibrida IPBT78×IPBT13 88
36 Genotipe tomat hibrida IPBT64×IPBT73 88
37 Genotipe tomat hibrida IPBT3×IPBT78 88
38 Kejadian penyakit pada pertanaman tomat 93
1 PENDAHULUAN
Latar Belakang
Tomat (Lycopersicon esculentum Mill.) termasuk komoditas hortikultura unggulan yang bernilai ekonomis dan strategis di Indonesia. Selama periode tahun 2007-2011 produksi tomat terus mengalami peningkatan, yaitu 635 474 ton menjadi 954 046 ton dan menurun pada tahun 2012 menjadi 893 504 ton, sehingga pada tahun 2012 Indonesia mengimpor tomat sebesar 9 857 ton (Ditjenhort 2011). Hal ini menunjukkan bahwa produksi tomat di Indonesia belum mencukupi kebutuhan. Data menunjukkan rata-rata produktivitas tomat nasional sebesar 14.3 ton masih lebih rendah dibandingkan dengan potensi produksi yaitu 45.7–80 ton ha-1 (Kementan 2012). Sentra-sentra produksi tomat pada umumnya di dataran tinggi. Budidaya tomat di dataran rendah mengalami kendala antara lain rendahnya produksi (Dane et al. 1991; Hanson et al. 2002) dan terjadi pecah buah. Pecah buah (fruit cracking) merupakan kelainan fisiologi yang sering ditemukan pada buah tomat. Pecah buah menyebabkan peningkatan kehilangan hasil baik pada tomat yang dikonsumsi segar maupun tomat olahan. Pecah buah dapat menurunkan penampilan pada tomat yang dikonsumsi segar sehingga dapat menurunkan jumlah buah yang dapat dipasarkan, sedangkan pada tomat olahan dapat memungkinkan masuknya patogen pada buah sehingga menyebabkan busuk sekunder. Laporan penelitian di Amerika menyebutkan kehilangan hasil panen tomat akibat pecah buah mencapai 35%. Terdapat dua tipe pecah buah yaitu tipe radial dan tipe konsentrik (Peet 1992; Peet dan Willits 1995; Dorais et al. 2004; Maboko 2006).
Pecah buah terjadi karena perubahan yang cepat pada laju pertumbuhan buah disebabkan faktor genetik dan faktor lingkungan. Faktor lingkungan yang berpengaruh terhadap pecah buah diantaranya adalah hujan, pengairan yang tidak teratur, pemangkasan, suhu tinggi, intensitas cahaya tinggi, kelembaban udara, kandungan kalsium dan boron pada tanah (Hudson 1956; Dorais et al. 2004; Maboko 2006; Liebisch 2009). Faktor lingkungan tersebut sulit dikontrol secara teknis, terlebih dengan kondisi lingkungan Indonesia yang berada di wilayah tropis. Pergeseran wilayah penanaman sayuran, termasuk tomat, ke dataran rendah menyebabkan resiko terjadinya pecah buah semakin besar. Penggunaan varietas unggul tomat yang tahan terhadap pecah buah dipandang sebagai cara yang efektif untuk mengatasi permasalahan pecah buah pada tomat.
tahan diperoleh F1 dan F2 mulai dari kelas rentan sampai tahan, dilaporkan juga bahwa pada sesama varietas tahan mempunyai susunan genetik yang berbeda. Amstrong dan Tompson (1967), melakukan uji dialel terhadap tiga belas varietas tomat. Hasil percobaannya menunjukkan galur tahan mempunyai daya gabung umum tertinggi terhadap pecah buah dan diketahui juga bahwa sifat pecah buah pada tomat dikendalikan oleh gen ganda yang mempunyai sifat dominan sebagian. Pendapat tersebut sesuai dengan hasil penelitian Hernandes dan Nassar (1970) yang menyatakan bahwa sifat ketahanan terhadap pecah buah tomat dikendalikan oleh gen ganda (poligenik) yang bersifat dominansi sebagian, sedangkan laporan AVRDC (1982) menyebutkan bahwa sifat pecah buah pada tomat dikendalikan oleh gen tunggal sederhana.
Program perakitan varietas tomat tahan pecah buah memerlukan tetua donor yang memiliki ketahanan terhadap pecah buah serta informasi tentang kendali genetik untuk menentukan metode introgresi serta metode seleksi yang efektif. Berdasarkan hal tersebut, maka studi tentang pewarisan pecah buah perlu dilakukan guna menentukan strategi program pemuliaan yang efektif dan efisien untuk memperoleh varietas tomat tahan terhadap pecah buah dan berdaya hasil tinggi.
Tujuan Penelitian
Tujuan dari penelitian ini adalah:
1 Memperoleh informasi tentang keragaman genetik dan kemiripan antar genotipe tomat.
2 Memperoleh informasi tingkat ketahanan genotipe tomat terhadap pecah buah. 3 Memperoleh penanda dan/atau kriteria seleksi yang efektif untuk perakitan
tomat tahan terhadap pecah buah.
4 Memperoleh informasi parameter genetik pecah buah pada tomat.
5 Memperoleh informasi daya gabung umum (DGU) dan daya gabung khusus (DGK) beberapa genotipe tomat.
6 Memperoleh informasi keragaan hibrida hasil silang dialel.
Hipotesis Penelitian
Hipotesis yang diajukan dari penelitian ini adalah:
1 Terdapat keragaman genetik dan kemiripan antar genotipe tomat. 2 Terdapat beberapa genotipe tomat yang tahan terhadap pecah buah.
3 Terdapat penanda/kriteria seleksi yang efektif untuk perakitan tomat tahan pecah buah.
4 Ketahanan tomat terhadap pecah buah dikendalikan oleh banyak gen dengan aksi gen aditif dan dominan.
5 Terdapat satu genotipe yang mempunyai daya gabung umum baik dan sepasang genotipe yang mempunyai daya gabung khusus baik untuk ketahanan terhadap pecah buah.
Kerangka Pemikiran
Kegiatan percobaan 1 akan menghasilkan informasi tingkat ketahanan genotipe tomat terhadap pecah buah, keragaman genetik dan kemiripan antar genotipe yang akan digunakan untuk memilih genotipe pada percobaan 2. Kegiatan percobaan 2 akan diperoleh informasi kriteria seleksi yang berkorelasi langsung dengan karakter pecah buah dan hasil untuk mempermudah kegiatan seleksi pada tahap selanjutnya serta bermanfaat dalam menentukan strategi pemuliaan yang efektif. Genotipe tomat terpilih disilangkan menggunakan metode persilangan dialel penuh pada percobaan 3. Kegiatan percobaan 3 akan menghasilkan informasi parameter genetik, daya gabung umum, daya gabung khusus dan heterosis. Kegiatan percobaan 4 yaitu penanaman kombinasi persilangan F1 bersamaan dengan beberapa varietas hibrida komersial dilakukan
untuk memperoleh informasi keragaan daya hasil calon varietas unggul hibrida tahan pecah buah yang adaptif di dataran rendah (Gambar 1).
2 TINJAUAN PUSTAKA
2.1 Potensi Tanaman Tomat
Tanaman tomat tergolong ke dalam famili Solanaceae dan genus Lycopersicon (Díez dan Nuez 2008). Genus Lycopersicon merupakan tanaman herba setahun (annual) atau tanaman semi tahunan yang mempunyai jumlah kromosom 2n = 24 (Tigchelaar 1986). Genus Lycopersicon terdiri dari dua subgenus yang berbuah merah disebut Eulycopersicon dan yang berbuah hijau disebut subgenus Eriopersicon. Subgenus Eulycopersicon meliputi L. esculentum Mill. dan L. pimpinellifolium Jusl Mill. Eriopersicon mempunyai warna hijau yang meliputi L. cheesmanii Riley, L. glandulosum C. H. Muller, L. hirsutum Humb Boupl dan L. peruvianum Mill. Spesies esculentum merupakan spesies penting dan banyak dibudidayakan. Tomat berasal dari Amerika. Tomat dan kerabatnya asli dari wilayah Andean, sekarang daerah tersebut meliputi Bolivia, Chile, Colombia, Equador dan Peru, dari habitat aslinya tomat dibawa ke Eropa setelah penemuan “dunia baru”. Awalnya tomat didomestikasi di Meksiko, kemudian tomat menyebar ke daerah tropik dan subtropik Amerika dan selanjutnya menyebar ke seluruh daerah tropik di “dunia lama” (Opeňa 1990).
Tomat sudah dijadikan sebagai salah satu sayuran yang mendapat prioritas untuk dikembangkan di Indonesia. Beberapa alasan yang melatarbelakangi pentingnya pengembangan tomat di Indonesia adalah tomat merupakan salah satu komoditas sayuran yang mempunyai prospek pasar cerah. Disamping itu tomat merupakan sumber nutrisi dan metabolit sekunder yang sangat penting bagi kesehatan manusia, seperti senyawa folat, vitamin A, C dan E, flavonoid, serat, karotenoid, likopen, vitamin dan mineral penting lainnya (Van der Ploeg dan Heuvelink 2005; Kailaku et al. 2007; Passam et al. 2007; Bhowmik et al. 2012). Tomat memiliki banyak kegunaan, baik sebagai sayuran maupun sebagai bahan baku industri makanan dan minuman. Potensi pasar tomat juga dapat dilihat dari segi harga yang terjangkau oleh seluruh lapisan masyarakat, sehingga membuka peluang yang lebih besar terhadap serapan pasar.
Luas panen tomat di Indonesia kian meningkat dari tahun ke tahun. Pada tahun 2007–2010, luas areal budidaya tomat terus meningkat dari 51 523 hektar menjadi 61 154 hektar. Luasan panen berkorelasi dengan semakin meningkatnya produksi tomat di Indonesia. Badan Pusat Statistik (BPS) menyebutkan total produksi tomat nasional pada tahun 2007–2011 telah meningkat dari 635 474ton menjadi 954 046ton per tahun (Badan Pusat Statistik 2012; Ditjenhort 2012).
2.2 Kualitas Buah
Kualitas buah sangat perlu diperhatikan dalam memenuhi kebutuhan konsumen. Kriteria kualitas hasil tomat sangat beragam bergantung pada konsumen. Kualitas buah pada tomat mencakup ukuran, kandungan bahan-bahan dalam buah, warna buah, penampilan dan lain-lain. Purwati (2007) menjelaskan bahwa kriteria kualitas yang ada pada buah tomat dapat diklasifikasikan dalam tiga kelompok yaitu : 1) kualitas bagian luar, terdiri atas warna kulit, bentuk buah, kekerasan dan ukuran buah, 2) kualitas bagian dalam, terdiri atas jumlah biji, ketebalan daging, dan kandungan lendir, 3) kualitas rasa, terdiri atas rasa manis, rasa asam, kekenyalan dan jumlah air buah.
Hasil penelitian Purwati (2007) melaporkan bahwa masyarakat pada umumnya menyukai buah tomat yang warna kulitnya merah terang (dengan nilai 7–8 hasil pengukuran CBT color chart), kekerasan buah sedang (nilai 110–130 mm 50-1 g 10-1 det pengukuran dengan penetrometer), bentuk buah agak lonjong (sphericity indeks 99–199), ukuran buah agak besar (volume 80–90 cm3), rasa buah manis (gula reduksi 4.25–5.0%), tidak masam (kadar total asam 0.34– 0.37%), banyak mengandung air buah (kandungan air 92–93%), dan buahnya kenyal/renyah.
Warna buah tomat adalah salah satu atribut paling penting dan kompleks kualitas buah. Kompleksitas warna tomat adalah karena adanya sistem pigmen karotenoid yang beragam dengan penampilan dikondisikan oleh jenis dan konsentrasi pigmen, dan tunduk pada regulasi genetik dan lingkungan (Arias et al. 2000; Lopez dan Gomez 2004).
Kualitas buah yang diinginkan untuk konsumsi rumah tangga berbeda dengan standar kualitas untuk industri. Tomat untuk tujuan konsumsi rumah tangga sebagai substitusi buah-buahan dan sebagai pelengkap bumbu masak, sehingga konsumen lebih mengutamakan tomat dengan rasa manis, sedikit asam, renyah dan mempunyai kandungan air sedang. Tomat untuk keperluan industri sebagai tomat olahan memiliki kriteria padatan total terlarut tinggi (+ 4.5 oBrix), pH rendah (+ 4.4), kompak, mudah dikuliti, tahan terhadap retak dan warnanya merah cerah (Villreal 1981). Hal ini sesuai dengan Tigchelaar (1986) yang menyebutkan beberapa parameter kualitas tipe olahan adalah warna, pH, total keasaman, total zat terlarut dan viskositas, sedangkan untuk tipe konsumsi segar adalah kekerasan buah, keseragaman bentuk, warna, ukuran dan bebas dari kerusakan/kelainan fisik seperti pecah buah.
2.3 Pecah Buah pada Tomat
banyak menurunkan produksi buah di lapang apabila dibanding bentuk radial (Thompson dan Kelly 1957). Pashar dan Lambeth (1960) menduga pecah buah terjadi akibat pengembangan jaringan yang tidak terkoordinasi selama pertumbuhan atau hanya merupakan peristiwa akibat pembengkakan saja. Peet (1992) mengusulkan tujuh teori yang menjelaskan terjadinya pecah buah, terutama pada tipe pecah buah radial, yaitu (1) Pengairan yang tidak teratur, terutama dari kondisi sangat kering menjadi sangat basah, (2) Suhu dan cahaya tinggi, (3) Aspek anatomi buah, (4) Pertumbuhan buah yang sangat cepat, (5) Pengaruh genetik, (6) Perbedaan yang tinggi antara suhu siang dan malam dan (7) Kelembaban yang tinggi.
Beberapa faktor yang diduga menjadi faktor penyebab terjadinya pecah buah dirangkum dari berbagai sumber adalah :
2.3.1 Genetik
Genetik merupakan salah satu faktor yang mempengaruhi terjadinya pecah buah pada tomat, dengan demikian perbaikan varietas dengan pemindahan gen tahan dirasa lebih menguntungkan. Beberapa peneliti telah melakukan penelitian- penelitian genetik tentang peristiwa pecah buah, namun belum diperoleh kesatuan pendapat mengenai pola pewarisan tersebut. Menurut Young (1959) pecah buah tipe radial dikendalikan oleh dua pasang gen mayor yaitu cr cr dan lr lr. Gen ketahanan pecah buah ditemukan berasosiasi dengan warna kematangan buah yaitu buah merah muda, jumlah buah per tanaman tinggi, rata-rata jumlah rongga buah rendah, diameter buah kecil dan pola pertumbuhan determinate. Ketahanan pecah buah dikendalikan dua pasang gen dengan sifat rentan dikendalikan oleh pasangan gen resesif (Pashar dan Lambeth 1960). Hasil penelitian Pashar dan Lambeth (1960) telah dibuktikan bahwa sifat ketahanan terhadap pecah buah adalah sifat kuantitatif yang dikendalikan oleh gen mayor dan minor. Persilangan antara dua tetua yang resisten diperoleh F1 dan F2 mulai dari kelas rentan sampai resisten, dilaporkan juga bahwa pada sesama varietas resisten mempunyai susunan genetik yang berbeda.
2.3.2 Air Hujan
Air hujan menyebabkan pecah buah pada tomat, terutama apabila hujan turun setelah panas mengakibatkan buah pecah beberapa jam kemudian. Hal ini disebabkan buah mengalami cekaman pada perikarp setelah buah menyerap air (Dickinson dan McCollum 1964). Kekerasan atau kekuatan perikarp ini tergantung dari varietasnya (Hall 1987). Pecah buah radial dapat terjadi pada interlokular septum karena secara anatomi buah tersebut mempunyai septum yang lemah, di samping septum-septum ini dijumpai adanya pseudo septum. Apabila pseudo septum ini juga dalam keadaan lemah kemudian hujan turun menyebabkan terjadinya pecah buah konsentris pada tomat.
2.3.3 Pengairan yang Tidak Teratur
Pemberian air khususnya dari kondisi yang sangat kering ke kondisi yang sangat basah dapat menyebabkan terjadinya pecah buah. Mekanisme tersebut dapat dijelaskan dengan penelitian Kamimura et al. (1972). Hasil penelitian tersebut menyatakan bahwa kelembaban tanah yang tinggi akan menurunkan kekuatan perenggangan (tensile strength) kulit buah tomat. Bertambahnya kecepatan pertumbuhan buah dalam kondisi seperti ini akan mendorong terjadinya keretakan kecil ini dan akan berkembang menjadi pecah buah. Laporan penelitian Peet and Willits (1995) menyatakan bahwa pemberian air yang berlebihan pada tanaman tomat di rumah kaca dapat meningkatkan pecah buah tipe radial.
2.3.4 Temperatur dan Cahaya yang Tinggi
Peningkatan temperatur secara drastis dapat menyebabkan terjadinya peningkatan tekanan pada kulit buah akibat adanya pulp dalam kulit buah tersebut. Tekanan tersebut akan menurunkan tingkat kekakuan dan kelenturan kulit buah dan mendorong terjadinya pecah buah (Lang dan During 1990). Menurut hasil penelitian Awan (1983) jumlah buah yang mengalami pecah buah lebih banyak terjadi pada penanaman di dalam rumah plastik apabila dibanding dengan penanaman di lapang. Hal ini kemungkinan disebabkan karena tingginya suhu udara yang terjadi di dalam rumah plastik. Max et al. (2009) melaporkan bahwa pengurangan suhu pada rumah kaca sebesar 2.6 dan 3.2 oC (siang) dan 1.2 dan 2.3 oC (malam) dapat meningkatkan insiden pecah buah.
naungan atau terlindungi oleh daun. Kejadian pecah buah mencapai 49% pada buah yang terpapar cahaya langsung dan 20% pada buah yang terlindungi.
2.3.5 Aspek Anatomi Buah
Sifat kulit buah yang berhubungan dengan ketahanan terhadap terjadinya pecah buah pada buah tomat telah dilaporkan oleh Kamimura et al. (1972). Kekuatan perenggangan dan kelenturan kulit buah pada periode breaker stage sampai pink stage sangat diperlukan apabila dikaitkan dengan pecah buah. Kultivar yang resisten terhadap pecah buah paling sedikit mempunyai salah satu diantara sifat di atas. Peet (1992) menyebutkan karakteristik buah yang berasosiasi dengan ketahanan terhadap pecah buah adalah ukuran buah kecil, kulit buah tipis, diameter buah kecil, jumlah buah per tanaman tinggi dan pola pertumbuhan tanaman tomat semideterminate.
2.3.6 Kecepatan Tumbuh Buah
Kecepatan tumbuh buah yang berlebihan dapat memacu terjadinya pecah buah. Pemangkasan atau penjarangan buah yang dilakukan menyebabkan persaingan antar buah dalam memperoleh asimilat akan berkurang, sehingga kecepatan tumbuh buah semakin bertambah. Menurut Bakker dan Janse dalam Peet (1992) pada buah tomat yang menerima asimilat lebih banyak akan mengalami pertumbuhan yang lebih cepat dan buah tersebut lebih rentan terhadap pecah buah.
2.3.7 Kelembapan yang Tinggi
Kelembapan udara yang tinggi dapat meningkatkan kejadian pecah buah tomat dalam rumah kaca, khususnya pada suhu yang tinggi. Tekanan air dan udara dalam buah tomat akan meningkat dengan meningkatnya suhu. Terdapat dua mekanisme dalam mengatasi kondisi ini, pertama dengan pelonggaran kulit buah, kedua dengan mengirim kembali air ke batang atau daun. Kondisi kelembapan udara tinggi menyebabkan mekanisme kedua tidak akan terjadi karena transpirasi rendah. Kondisi seperti ini menyebabkan mekanisme pertama yang akan terjadi. Hal ini akan mendorong terjadinya pecah buah (Byari dalam Peet 1992).
2.3.8 Kekuatan Dinding Sel Buah
Teori-teori di atas secara tidak langsung menyebutkan pecah buah berkaitan erat dengan kekuatan kulit (epidermis) buah. Mekanisme penguatan dan pelenturan kulit buah dapat diperjelas dengan melihat proses yang terjadi pada dinding sel.
dengan serat-serat mikro selulose menyebabkan dinding sel mudah bergeser, sehingga dinding sel akan mudah meregang. Mekanisme fisik dan kimia yang menyebabkan terputusnya ikatan hidrogen ini memudahkan dinding sel bergerak sehingga lebih elastis. Hal ini merupakan faktor yang perlu dipertimbangkan dalam kasus pecah buah pada tomat.
Mekanisme penguatan dinding sel dapat dilihat pada reaksi-reaksi yang terjadi dalam polisakarida pektik. Interkonversi senyawa pektat pada dinding sel tanaman, galaktosa dioksidasi pada C-6 suatu karbinol (-CH2OH) menjadi suatu gugus karboksil (-COOH) yang disebut asam galakturonat. Polimerisasi 1-4 rantai asam galakturonat menjadi asam pektat. Asam pektat dapat berubah menjadi pektin apabila terjadi pergantian gugus (-COOH) dengan (-CH3) yang mengalami proses esterifikasi. Asam pektat pun dapat pula berubah menjadi kalsium pektat dengan penambahan kalsium. Penambahan kalsium pada dinding sel mengakibatkan terjadinya rigiditas dinding sel (Wareing dan Phillips 1981). Jika kondisi kalsium tidak terdapat dalam dinding sel, maka rigiditas akan menurun, hal ini akan memudahkan sel-sel tersebut pecah bila sel mengadakan pembesaran.
2.3.9 Auksin
Auksin adalah salah satu hormon yang tidak terlepas dari proses pertumbuhan dan perkembangan tanaman. Salah satu peran fisiologi auksin adalah pembesaran sel.
Mekanisme ekspansi sel kemungkinan disebabkan oleh longgarnya/ bergesernya dari dinding sel primer. Menurut Albersheim dalam Moore (1979) auksin berfungsi mengaktifkan pompa ion H+ yang terletak dalam plasmalema. Hal ini akan mengakibatkan rendahnya pH pada dinding sel, sehingga ikatan hidrogen antara xyloglucan dengan serat mikroselulose pada dinding sel primer akan terputus. Hal tersebut menyebabkan dinding sel akan mudah mengalami pelonggaran.
IAA (Indol Acetic Acid) adalah auksin endogen yang terdapat dan disintesis dalam tanaman. Senyawa-senyawa lain yang mempunyai aktivitas serupa auksin dan merupakan senyawa sintetik salah satunya adalah NAA atau Naphthalene Acetic Acid (Wattimena 1988).
Menurut Yamamoto et al. (1992), NAA dapat menekan terjadinya pecah buah pada sweet cherry. Aplikasi NAA 24 hari sebelum panen pada konsentrasi 0.5, 1, dan 2 ppm dapat menekan terjadinya pecah buah hingga 13-40 % dari kontrol. Menurut Byers et al. (1990) aplikasi GA4+7 dapat menekan terjadinya pecah buah pada apel dari 56 % menjadi 21 %, dinyatakan pula bahwa kombinasi GA4+7, Daminozide, NAA dan Vapor Gard dapat menekan terjadinya pecah buah dari 93 % menjadi 22 %. Hal ini disebabkan karena Giberelin dapat meningkatkan aktivitas auksin dalam tanaman.
2.3.10 Kalsium
membantu sintesis protein, transfer karbohidrat dan keberadaannya dapat menekan efek racun dari logam berat dalam tanaman (Jones et al. 1991).
Berbeda dengan hara makro lain, proporsi yang tinggi dari total kalsium dalam jaringan tanaman terdapat pada dinding sel (apoplas). Kalsium dalam tanaman terdapat dalam berbagai bentuk yaitu : bentuk terlarut, terikat oleh pektat, oksalat dan fosfat. Bentuk terlarut banyak terdapat dalam vakuola, demikian juga bentuk terikat pektat terdapat dalam dinding sel (lamela tengah dan permukaan luar membran plasma). Dalam kondisi suplai kalsium yang rendah bentuk pektat dapat terjadi sebanyak 50 % dari total kalsium (Marschner 1986).
Pola pergerakan kalsium dalam tanaman berbeda dengan unsur lain. Kalsium merupakan unsur yang tidak mobil dalam phloem, sehingga translokasi kalsium ke bagian-bagian tanaman terjadi melalui xylem dengan adanya aliran transpirasi (Bollard dan Buttler dalam Bollard 1970).
Kandungan kalsium buah tomat menurut Bollard (1970) sebanyak 0.13 % berat kering, lebih rendah apabila dibandingkan dengan kandungan kalsium pada daun (6.08 %). Perbandingan kandungan unsur-unsur nitrogen, fosfor dan kalium antara daun dan buah tidak jauh berbeda. Hal ini menunjukkan bahwa kalsium yang berada dalam daun tidak ditranslokasikan ke dalam buah bersama asimilat seperti terjadi pada unsur-unsur yang lain. Menurut Link (1974) dan Redmond (1975), pemberian kalsium yang efektif untuk peningkatan kualitas buah adalah dengan penyemprotan langsung terhadap buah tersebut.
Menurut Knott (1962) aplikasi kalsium pada tomat melalui penyemprotan daun menggunakan kalsium klorida (CaCl2 36.1 % Ca) dapat dilakukan pada
konsentrasi 6000-12000 ppm kalsium klorida, dan bentuk kalsium nitrat (Ca(NO3)2) 28 % Ca) pada konsentrasi 6000-18000 ppm kalsium nitrat. Menurut
Yamamoto et al. (1992) aplikasi kalsium dalam bentuk kalsium nitrat (Ca(NO3)2)
pada konsentrasi 0.5 % (5000 ppm) 7 hari sebelum panen dapat menekan pecah buah pada sweet cherry. Liebisch et al. ( 2009) melaporkan bahwa aplikasi kalsium (CaCl2, 5 g l1) dan boron (H3BO3, 0.2 g l-1) (CaB) pada dua kultivar tomat yang peka dan tahan terhadap pecah buah dan busuk pangkal buah (blossom end-rot). Aplikasi kalsium dan boron tidak memberikan pengaruh signifikan terhadap hasil buah total, namun perlakuan ini memberikan pengaruh yang berbeda pada kultivar. Aplikasi pada kultivar rentan pecah buah dan busuk pangkal buah tidak layak lebih banyak daripada kultivar tahan selain itu aplikasi kalsium dan boron menurunkan kejadian busuk pangkal buah namun meningkatkan kejadian pecah buah pada waktu yang sama. Pemilihan kultivar tahan tampaknya menjadi prioritas utama untuk mengatasi kendala pecah buah dan busuk pangkal buah.
2.4 Persilangan Dialel
tetua-tetua secara genetik berbeda luas agar terjadi rekombinasi baru yang dituju, yaitu kultivar-kultivar dengan gen-gen yang baik. Cara ini menggantungkan pada interaksi gen agar diperoleh kultivar dengan karakter yang dituju. Salah satu metode persilangan yang masuk dalam kelompok ini adalah metode persilangan dialel (Baihaki 2000).
Metode persilangan dialel digunakan untuk mengidentifikasi tetua-tetua superior dalam jumlah besar atau juga mengidentifikasi dari sejumlah besar genotipe tetua yang mampu memberikan efek heterotik pada hibrida F1-nya (Baihaki 2000; Syukur et al. 2012). Persilangan dialel merupakan persilangan dengan menggunakan seluruh kombinasi persilangan yang mungkin diantara sekelompok tetua, termasuk persilangan sendiri tetua tersebut. Analisis dialel digunakan untuk menganalisis data hasil seluruh kombinasi persilangan tersebut. Analisis dialel akan memberikan informasi mengenai (i) parameter genetik dan besarannya, serta (ii) kemampuan daya gabung umum (general combining ability/GCA) dan daya gabung khusus (specific combining ability/SCA) dari tetua persilangan. GCA adalah keragaan suatu galur dalam kombinasi silang tunggal dengan galur-galur yang lain, sedangkan SCA adalah hasil hibrida suatu galur dengan galur lain (Singh dan Chaudary 1979). Analisis dialel juga bertujuan untuk mengevaluasi dan memilih tetua berdasarkan turunan terbaik dan evaluasi turunan terbaik. Analisis silang dialel diperlukan untuk menduga efek aditif dan dominan dari suatu populasi yang selanjutnya dapat digunakan untuk menduga ragam genetik dan heritabilitas (Baihaki 2000).
Persilangan dialel dapat dibagi menjadi tiga tipe persilangan, yaitu (1) dialel penuh (full diallel), (2) setengah dialel (half diallel), dan (3) dialel parsial (partial diallel) (Singh dan Chaudhary 1979). Dalam pelaksanaannya, analisis ini harus memenuhi beberapa asumsi berikut: (1) segregasi diploid, (2) tidak ada perbedaan antara persilangan resiprokal, (3) tidak ada interaksi antara gen-gen yang tidak satu alel, (4) tidak ada multialelisme, (5) tetua homozigot, (6) gen-gen menyebar secara bebas diantara tetua (Hayman 1954). Keuntungan dari teknik silang dialel adalah (1) secara eksperimental merupakan pendekatan sistematik, (2) secara analitik merupakan evaluasi genetik menyeluruh yang berguna dalam mengidentifikasi persilangan bagi potensi seleksi yang terbaik pada awal generasi. Beberapa pendugaan parameter genetik pada analisis dialel dapat dilakukan tanpa pembentukan populasi F2, BCP1 dan BCP2.
heritabilitas arti sempit. Analisis melalui pendekatan ini dapat dilanjutkan ke populasi F2 (Singh dan Chaudary 1979).
Nilai DGU dan DGK yang dihasilkan dari analisis dialel digunakan sebagai dasar pemilihan tetua dan kombinasi persilangan yang berpotensi untuk dikembangkan selanjutnya. Dirangkum dari Greenleaf (1986) dan Poespodarsono (1988), daya gabung diartikan sebagai kemampuan genotipe untuk memindahkan sifat yang diinginkan kepada keturunannya. DGU digunakan untuk membandingkan penampilan setiap tetua dalam kombinasi persilangan dengan semua tetua lain. Dengan kata lain, DGU menunjukkan kemampuan rata-rata suatu genotipe bila disilangkan dengan sejumlah genotipe lain. Jika penampilan rata-rata keturunannya tinggi dibandingkan tetua atau genotipe lain, berarti DGU-nya tinggi. DGU biasaDGU-nya diDGU-nyatakan sebagai deviasi dari keseluruhan nilai tengah. Daya Gabung Umum dapat bernilai positif atau negatif. Nilai yang positif menunjukkan penampilan tetua berada di atas rata-rata progeni, sedangkan nilai yang negatif menunjukkan sebaliknya. Daya Gabung Khusus digunakan untuk membandingkan penampilan progeni hasil persilangan (F1) secara individual
dengan penampilan rata-rata genotipe-genotipe tetua. Dengan kata lain, DGK dapat diartikan sebagai kemampuan suatu kombinasi persilangan untuk menunjukkan kemampuan keturunan. Jika progeni suatu kombinasi persilangan menunjukkan penampilan tinggi dikatakan DGK-nya tinggi. Nilainya juga positif atau negatif seperti halnya DGU. Daya gabung khusus akan selalu merujuk pada progeni persilangan dan bukan pada tetua.
Pendekatan Griffing adalah metode lain yang digunakan untuk menganalisis hasil persilangan dialel. Pendekatan Griffing menjelaskan daya gabung umum dan daya gabung khusus dari persilangan dialel tersebut. Berdasarkan pendekatan Griffing, terdapat 4 metode analisis silang dialel, yaitu: metode I (full diallel) yaitu persilangan yang terdiri dari tetua, F1 dan resiprokal dengan analisis [n(n-1)/2]. Metode II yaitu persilangan yang terdiri dari tetua, F1 tanpa resiprokal dengan analisis [n(n+1)/2]. Metode III yaitu persilangan yang terdiri dari F1 dan resiprokal dengan analisis n(n-1). Metode IV yaitu persilangan yang terdiri dari hanya F1 tanpa resiprokal, dengan analisis n(n-1)/2 (Griffing 1956).
3 KETAHANAN 25 GENOTIPE TOMAT (Lycopersicon esculentum Mill.) TERHADAP PECAH BUAH DAN KERAGAMAN GENETIKNYA
ABSTRAK
Pecah buah menjadi salah satu kendala pada budidaya tanaman tomat di dataran rendah. Pecah buah dapat menurunkan kuantitas dan kualitas hasil, baik pada tomat konsumsi segar maupun olahan. Penelitian ini bertujuan untuk mengevaluasi ketahanan 25 genotipe tomat terhadap pecah buah dan keragaman genetiknya. Penelitian dimulai dari April hingga Agustus 2012 di Laboratorium Pemuliaan Tanaman, Fakultas Pertanian IPB dan Kebun Percobaan Leuwikopo (6o56’34’’S, 106o72’ 56’’E) IPB. Percobaan disusun dalam rancangan kelompok lengkap teracak faktor tunggal dengan 3 ulangan. Ketahanan terhadap pecah buah dievaluasi dengan indeks pecah buah dan persentase pecah buah. Perbedaan antar genotipe diuji dengan uji F dilanjutkan dengan uji lanjut DMRT. Keragaman genetik dianalisis dengan menggunakan analisis komponen utama dan analisis gerombol. Hasil penelitian menunjukkan indeks pecah buah 25 genotipe tomat berkisar 0.00-20.58. Berdasarkan karakter agronomi terdapat beberapa genotipe yang memiliki kriteria yang dapat dijadikan sebagai bahan tetua pada pemuliaan tanaman tomat. Hasil analisis komponen utama dan analisis gerombol menunjukkan genotipe tomat mengelompok menjadi empat kelompok.
Kata kunci: analisis gerombol, keragaman genetik, sidik lintas
3 GENETIC DIVERSITY AND RESISTANCE OF 25 TOMATO GENOTYPES (Lycopersicon esculentum Mill.)
TO FRUIT CRACKING
ABSTRACT
Tomato fruit cracking had became one of the problems in lowland cultivation. It causes decreasing and subsequent serious economic loses, either both on freshmarket or processing tomato. The objective of this study was to evaluate the resistance level of 25 tomato genotypes to fruit cracking and the genetic diversity. The experiment was conducted from April until August 2012 at Leuwikopo Field Station, Bogor Agricultural University. Randomized complete block design was used with three replications. Fruit crack index was used to evaluate tomato fruit crack resistance. The genetic diversity was analyzed by clustering analysis. Result of resistance evaluation showed that fruit crack index of 25 tomato genotypes range from 0.00 to 20.58. According to agronomic character some genotype can be selected as material breeding. Based on genetic diversity analysis, all genotypes could be divided into four groups.
PENDAHULUAN
Tomat (Lycopersicon esculentum Mill.) termasuk komoditas hortikultura unggulan yang bernilai ekonomis dan strategis di Indonesia. Tomat juga merupakan sumber nutrisi dan metabolit sekunder yang sangat penting bagi kesehatan manusia, seperti senyawa folat, vitamin A, C dan E, flavonoid, serat, karotenoid, likopen, vitamin dan mineral penting lainnya (Van der Ploeg dan Heuvelink 2005; Kailaku et al. 2007; Passam et al. 2007; Bhowmik et al. 2012), sehingga tomat kaya akan manfaat baik sebagai sayuran, bumbu masak, bahan minuman maupun sebagai bahan baku industri makanan, kosmetik dan obat-obatan. Selama periode tahun 2007-2011 produksi tomat terus mengalami peningkatan, yaitu 635 474 ton menjadi 954 046 ton dan menurun pada tahun 2012 menjadi 893 504 ton, sehingga pada tahun 2012 Indonesia mengimpor tomat sebesar 9 857 ton (Ditjenhort 2012). Hal ini menunjukkan bahwa produksi tomat nasional belum dapat mencukupi kebutuhan dalam negeri.
Tomat pada umumnya dibudidayakan di dataran tinggi. Semakin terbatasnya ketersediaan lahan penanaman tomat di dataran tinggi yang disebabkan oleh persaingan dengan komoditas hortikultura lain, juga karena sebagian wilayah tersebut merupakan daerah konservasi, sehingga perlu perluasan areal tanam ke dataran menengah dan rendah. Usaha penanaman tomat di dataran rendah menghadapi kendala berupa penurunan daya hasil, selain itu jumlah varietas tomat dataran rendah yang sudah dilepas jumlahnya terbatas (Purwati 2007). Kendala lain adalah terjadinya pecah buah (fruit cracking). Pecah buah merupakan kelainan fisiologi yang tidak disebabkan oleh infeksi penyakit atau serangga. Pecah buah terjadi karena pertumbuhan buah yang cepat pada kondisi ketersediaan air melimpah dan suhu tinggi, dan menurunnya elastisitas kulit buah terutama ketika kondisi ini diikuti periode stres tanaman (Peet 1992; Dorais et al. 2004; Liebisch et al. 2009; Masarirambi et al. 2009). Pecah buah menyebabkan peningkatan kehilangan hasil baik pada tomat yang dikonsumsi segar maupun tomat olahan (Dorais et al. 2004; Matas et al. 2004; Liebisch et al. 2009; Max dan Horst 2009). Pecah buah dapat menurunkan penampilan sehingga menurunkan jumlah buah yang dapat dipasarkan pada tomat yang dikonsumsi segar, sedangkan pada tomat olahan, pecah buah memungkinkan masuknya patogen pada buah sehingga menyebabkan busuk sekunder (Peet 1992; Simon 2006; Liebisch et al. 2009). Hasil penelitian di Amerika Serikat melaporkan kehilangan hasil panen tomat akibat pecah buah mencapai 35% (Dorais et al. 2004). Kerugian akibat pecah buah dapat diatasi dengan menggunakan kultivar tahan, mencukupi kebutuhan air dan nutrisi tanaman yang seimbang (Liebisch et al. 2009; Masarirambi et al. 2009).
diperlukan untuk menentukan arah dan metode pemuliaannya. Informasi tentang keanekaragaman genetik di dalam dan di antara tanaman spesies yang terkait erat sangat penting untuk pemanfaatan sumber daya genetik dan sangat berguna dalam karakterisasi aksesi individu dan kultivar (Satori et al. 2002).
Tujuan dari penelitian ini adalah untuk memperoleh data ketahanan 25 genotipe tomat terhadap pecah buah, keragaman dan pola hubungan kekerabatan antar genotipe.
BAHAN DAN METODE
Waktu dan Tempat Penelitian
Penelitian dilakukan mulai bulan Maret hingga Agustus 2012 di laboratorium Pemuliaan Tanaman, Fakultas Pertanian IPB dan Kebun Percobaan Leuwikopo, IPB. Lahan berada pada ketinggian 250 m di atas permukaan laut, memiliki tipe tanah latosol.
Materi Genetik
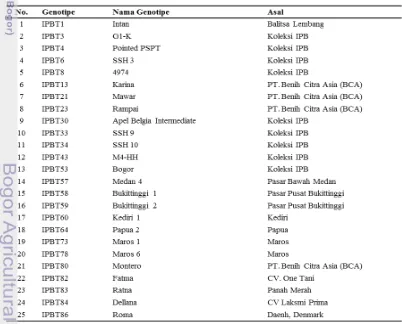
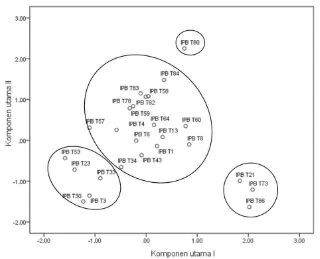
[image:34.595.108.510.418.742.2]Materi genetik yang digunakan adalah 25 genotipe tomat koleksi Bagian Genetika dan Pemuliaan Tanaman Departemen Agronomi dan Hortikultura IPB dan nomor-nomor lokal yang telah digalurkan (Tabel 1).
Tabel 1 Daftar genotipe tomat yang digunakan dalam penelitian
Pelaksanaan Percobaan
Kegiatan penelitian diawali dengan penyemaian benih. Benih disemai sebanyak 1 butir per lubang tray yang berisi media semai steril. Penyiraman dilakukan setiap hari pada pagi dan sore hari. Pemupukan dilakukan setelah bibit berumur 2 minggu setelah semai dengan periode satu minggu sekali menggunakan pupuk NPK (16:16:16) dengan dosis 10 gl-1air dan gandasil 2 gl-1 air diaplikasikan dengan cara disiramkan pada pangkal bibit. Penyemprotan pestisida dilakukan jika terlihat gejala serangan hama dan penyakit pada persemaian. Pengolahan lahan dan pembuatan bedengan dilakukan bersamaan saat kegiatan penyemaian.
Percobaan disusun dalam rancangan kelompok lengkap teracak (RKLT) faktor tunggal dengan 3 ulangan, masing-masing satuan percobaan terdiri atas 20 tanaman. Satu satuan percobaan terdiri atas 20 tanaman yang ditanam pada bedengan berukuran 1 m x 5 m yang ditutup mulsa plastik hitam perak, jarak tanam 50 cm x 50 cm. Bibit dipindah ke lapang setelah berumur ± 4 minggu. Bibit yang dipilih adalah bibit dengan pertumbuhannya tegar, berdaun 3-5 helai, warna daun hijau dan tidak terkena hama penyakit (Pangaribuan et al. 2011). Pemupukan dalam bentuk larutan NPK (16:16:16) 10 g l-1 dilakukan setiap seminggu sekali, masing-masing tanaman 250 ml. Penyemprotan pestisida dilakukan 2 minggu sekali dengan menggunakan fungisida mancozeb 80% atau propineb 2 g l-1, insektisida profenovos dengan dosis 2 ml l-1. Pewiwilan tunas air dilakukan agar tanaman dapat tumbuh optimal. Pengendalian gulma dilakukan secara manual. Kegiatan pemanenan dilakukan pada kondisi buah masak pada skor 4 dan 5 pada standar skala warna kulit buah tomat. Pemanenan dilakukan setiap 5 hari sekali selama 8 kali panen.
Pecah buah
Gambar 2 Skoring pecah buah pada buah tomat A. tipe konsentrik; B dan C tipe radial
1 Persentase bobot buah pecah (% tan-1)
Pengamatan dilakukan pada saat panen. Buah yang dipanen dipisahkan yang pecah kemudian ditimbang, bobot buah pecah dihitung dari 8 kali panen. Persentase bobot buah pecah dihitung dengan rumus:
2 Jumlah buah per tanaman
Pengamatan dilakukan pada saat panen. Jumlah buah dihitung dari 8 kali panen.
3 Panjang buah (cm)
Panjang buah diukur dengan menggunakan jangka sorong dari pangkal hingga ujung buah.
4 Diameter buah (cm)
Diameter buah diukur dengan menggunakan jangka sorong pada bagian tengah buah.
5 Tebal daging buah (cm)
Tebal daging buah diukur dengan menggunakan jangka sorong. Buah dipotong melintang pada bagian tengah buah. Tebal daging buah diukur 3 kali pada bagian yang memiliki tebal daging yang berbeda kemudian diambil rata-ratanya.
0 1 2 3 4
6 Jumlah rongga buah
Buah dipotong melintang pada bagian tengah buah kemudian dihitung jumlah rongganya.
7 Umur berbunga (HST)
Pengamatan dilakukan pada saat tanaman berbunga yaitu dengan mencatat pada hari keberapa pada saat 50% populasi tanaman telah berbunga pada bunga ketiga dari tandan kedua.
8 Umur panen (HST)
Pengamatan dilakukan pada saat 50% populasi tanaman telah dipanen pada kondisi buah masak pada > skor 3 pada standar skala warna kulit buah tomat (Kader 1995).
9 Tinggi tanaman (cm)
Tinggi tanaman diukur setelah panen ke-2 dari permukaan tanah sampai titik tumbuh tertinggi.
10 Diameter batang (cm)
Diameter batang diukur setelah panen ke-2 pada sepertiga tanaman bagian tengah tanaman
11 Panjang daun (cm) dan lebar daun (cm)
Panjang dan lebar daun diukur setelah panen ke-2 pada sepertiga tanaman bagian tengah tanaman
12 Total padatan terlarut (obrix)
Diukur pada buah panen ke-2 dan ke-3 dengan menggunakan alat handrefraktometer.
13 Kekerasan buah (kgcm-1)
Kekerasan buah diukur pada buah panen ke-2 dan ke-3 dengan menggunakan alat penetrometer.
14 Kadar air (%)
Kadar air diukur dengan mengeringkan buah menggunakan oven listrik pada suhu 100oC selama 24 jam. Buah tomat pada panen ke-2 dan ke-3 dipotong menjadi 4-8 bagian kemudian dimasukkan dalam amplop kertas. Buah tomat ditimbang bersama amplop sebagai data berat basah. Setelah dioven ditimbang kembali buah tomat bersama amplop sebagai berat kering. Kadar air dihitung dengan rumus:
Analisis Data
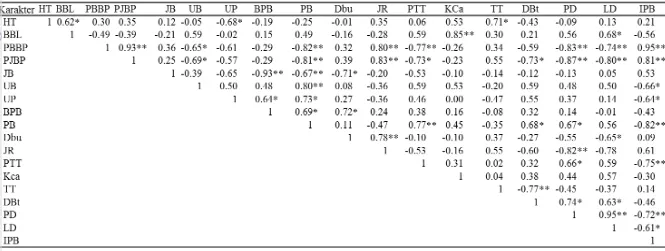
Perbedaan antar genotipe diuji menggunakan uji F pada taraf nyata 5%, bila terdapat perbedaan yang nyata maka untuk mengetahui genotipe yang berpenampilan lebih baik diantara genotipe yang lain dilanjutkan dengan uji lanjut DMRT (Duncan’s Multiple Range Test). Sidik ragam (Tabel 2) yang digunakan dalam penelitian ini berdasarkan Mattjik dan Sumertajaya (2006).
Tabel 2 Sumber keragaman dan nilai harapana
Sumber
keragaman db JK KT F-hitung
Ulangan (r) r-1 JKu Jku/( r-1) KTu/KTe
Genotipe (g) g-1 JKg JKg/(g-1) KTg/KTe
Galat (r-1)(g-1) JKe JKe/(r-1)(g-1)
Total gr -1 JKt
a
r = jumlah ulangan; g = jumlah genotipe; Jku = jumlah kuadrat ulangan; JKg = jumlah kuadrat genotipe; Jke = jumlah kuadrat galat; Ktu = kuadrat tengah ulangan; KTg = kuadrat tengah genotipe; KTe = kuadrat tengah galat
Model linier dalam analisis ragam berdasarkan Gomez dan Gomez (2007) sebagai berikut:
Keterangan: Yij = nilai fenotipe pada perlakuan ke-i dan kelompok ke- j
µ = nilai tengah umum
αi = pengaruh genotipe ke- i (1, 2, 3, …, 25)
βj = pengaruh kelompok ke- j (1, 2, 3)
ɛij = galat percobaan
Perbedaan nyata antar genotipe diuji berdasarkan uji lanjut DMRT pada taraf 5% menggunakan rumus (Steel dan Torrie 1989).
Keterangan: �� � � = nilai tabel Duncan pada taraf nyata α p = Jarak peringkat antar dua perlakuan Keragaman Genetik 25 Genotipe Tomat
Sebanyak 25 genotipe ditanam masing-masing sebanyak 10 tanaman. Peubah yang diamati terdiri atas 27 peubah kualitatif yang dirangkum dari Panduan Pengujian Individual Kebaruan, Keunikan, Keseragaman dan Kestabilan Tomat (PPVT 2007), International Union for the Protection of New Varieties of Plants (UPOV 2011) dan Descriptor for Tomato (Lycopersicon spp.) (IPGRI 1996). Pengamatan juga dilakukan pada 8 peubah kuantitatif yaitu panjang buah, diameter buah, jumlah rongga buah, tinggi tanaman, diameter batang, panjang buku, panjang daun dan lebar daun.
Karakter kualitatif yang diamati meliputi :
1 Pewarnaan antosianin pada hipokotil: (1) tidak ada , (9) ada.
2 Pewarnaan antosianin pada ruas tiga teratas: (1) tidak ada atau sangat lemah, (3) lemah, (5) sedang, (7) kuat, (9) sangat kuat.
3 Letak daun (pada sepertiga tanaman bagian tengah): (3) semi tegak, (5) horizontal, (7) menggantung
Yij = µ+ αi+ βj +ɛij
(3) semi tegak (5) horizontal (7) menggantung
Gambar 3 Letak daun (pada sepertiga tanaman bagian tengah) Gambar diadaptasi dari UPOV (2011)
4 Pembagian helai daun: (1) menyirip, (2) menyirip ganda 5 Tipe daun: (1) Tipe 1, (2) Tipe 2, (3) Tipe 3, (4) Tipe 4
(1) menyirip (2) menyirip ganda Gambar 4 Pembagian helai daun Gambar diadaptasi dari UPOV (2011)
6 Intensitas warna hijau daun: (3) terang, (5) sedang, (7) gelap.
7 Letak anak daun terhadap tulang daun utama: (1) ke atas, (2) mendatar, (3) ke bawah.
Gambar 5 Letak anak daun terhadap tulang daun utama Gambar diadaptasi dari UPOV (2011)
8 Tipe tandan bunga (pada pelepah daun kedua dan ketiga): (1) secara umum uniparous, (2) sebagian uniparous sebagian multiparous, (3) secara umum multiparous.
(1) (2) (3)
9 Cabang pada tandan bunga (bunga pertama pada tandan bunga): (1) tidak ada, (9) ada.
10 Warna bunga: (1) kuning, (2) jingga
11 Lapisan absisi: (1) tidak ada, (9) ada.
Gambar 7 Lapisan absisi pada tangkai buah Gambar diadaptasi dari UPOV (2011)
12 Panjang Pedisel (dari lapisan absisi terhadap calyx): (3) pendek, (5) sedang, (7) panjang.
13 Ukuran buah: (1) sangat kecil, (3) kecil, (5) sedang, (7) besar, (9) sangat besar.
14 Bentuk buah dalam penampang membujur: (1) pipih, (2) agak pipih, (3) bulat, (4) persegi, (5) silinder, (6) bentuk hati, (7) bentuk telur sungsang, (8) bentuk telur, (9) bentuk pear, (10) bentuk pear lancip.
Gambar 8 Bentuk buah dalam penampang membujur Gambar diadaptasi dari UPOV (2011) dan PPVT (2007)
15 Irisan melintang: (1) tidak bulat, (2) bulat.
Gambar 9 Depresi buah pada ujung tangkai buah Gambar diadaptasi dari UPOV (2011)
17 Ukuran lapisan gabus di sekeliling parut tangkai buah: (1) sangat kecil, (3) kecil, (5) sedang, (7) besar, (9) sangat besar.
18 Ukuran parut pada bekas tangkai putik: (1) sangat kecil, (3) kecil, (5) sedang, (7) besar, (9) sangat besar.
19 Bentuk ujung buah: (3) melekuk, (4) melekuk agak datar, (5) datar, (6) datar meruncing, (7) meruncing.
Gambar 10 Bentuk ujung buah. Gambar diadaptasi dari UPOV (2011) dan PPVT (2007)
20 Ukuran bagian tengah buah dalam irisan melintang: (1) sangat kecil, (3) kecil, (5) sedang, (7) besar, (9) sangat besar.
21 Jumlah rongga buah: (1) dua, (2) dua dan tiga, (3) tiga dan empat, (4) lebih dari empat.
(1) (2) (3) (4)
22 “Bahu buah hijau” sebelum masak: (1) tidak ada, (9) ada.
(1) (9)
Gambar 12 “Bahu buah hijau” sebelum masak Gambar diadaptasi dari UPOV (2011)
23 Luas “bahu buah hijau”: (3) kecil, (5) sedang, (7) besar.
24 Intensitas warna hijau buah pada bahu buah: (3) lemah, (5) sedang, (7) kuat. 25 Intensitas warna hijau buah sebelum matang: (3) terang, (5) sedang, (7) gelap. 26 Ribbing at peduncle end: (1) tidak ada, (9) ada.
27 Warna buah masak: (1) kuning, (2) jingga, (3) merah muda, (4) merah. 28 Warna daging buah: (1) kuning, (2) jingga, (3) merah muda, (4) merah.
Keragaman genetik dan pola hubungan kekerabatan dianalisis dengan analisis gerombol (cluster analysis) dan analisis komponen utama (AKU) menggunakan software SPSS versi 19. Informasi hubungan kekerabatan digunakan sebagai dasar dalam rekomendasi tetua yang akan dipilih pada pembentukan populasi studi pewarisan. Penentuan genotipe calon tetua jika terdapat beberapa genotipe terpilih akan dipilih genotipe yang lebih ekstrim, hubungan genetiknya lebih jauh, serta lebih mudah persilangannya.
HASIL DAN PEMBAHASAN
Pecah buah pada tomat
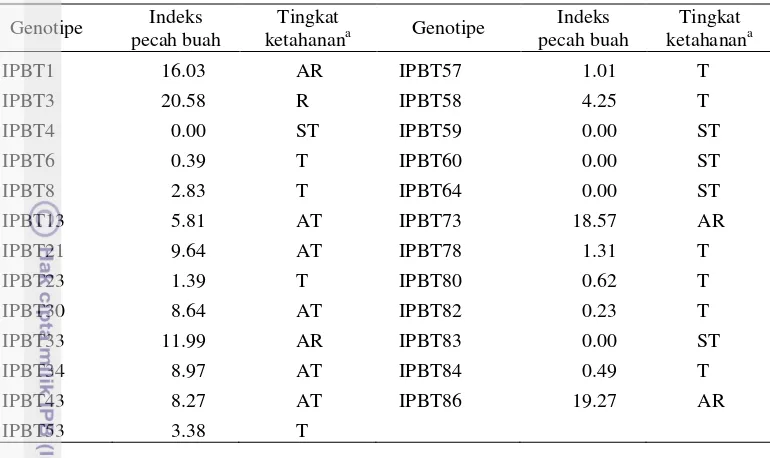
Tabel 3 Nilai indeks pecah buah pada 25 genotipe tomat dan tingkat ketahananya Genotipe Indeks
pecah buah
Tingkat
ketahanana Genotipe
Indeks pecah buah
Tingkat ketahanana
IPBT1 16.03 AR IPBT57 1.01 T
IPBT3 20.58 R IPBT58 4.25 T
IPBT4 0.00 ST IPBT59 0.00 ST
IPBT6 0.39 T IPBT60 0.00 ST
IPBT8 2.83 T IPBT64 0.00 ST
IPBT13 5.81 AT IPBT73 18.57 AR
IPBT21 9.64 AT IPBT78 1.31 T
IPBT23 1.39 T IPBT80 0.62 T
IPBT30 8.64 AT IPBT82 0.23 T
IPBT33 11.99 AR IPBT83 0.00 ST
IPBT34 8.97 AT IPBT84 0.49 T
IPBT43 8.27 AT IPBT86 19.27 AR
IPBT53 3.38 T
a
ST= sangat tahan, T= tahan, AT= agak tahan, AR= agak rentan, R= rentan
Pecah buah pada tomat dapat dibedakan menjadi dua macam berdasarkan bentuknya, yaitu tipe radial dan tipe konsentrik (Dorais et al. 2004; Masarirambi et al. 2009). Beberapa genotipe menunjukkan pecah buah tipe radial yaitu IPBT1, IPBT21, IPBT73 dan IPBT86. Genotipe-genotipe tersebut pada umumnya adalah genotipe yang memiliki jumlah rongga yang banyak (Tabel 8). Genotipe yang lain menujukkan tipe konsentrik. Ada kalanya genotipe dengan jumlah rongga sedikit juga mengalami tipe pecah radial. Belum ada korelasi yang jelas antara bentuk buah dengan tipe pecah buah (Dorais et al. 2004). Beberapa faktor yang diduga menjadi faktor penyebab terjadinya pecah buah adalah genetik (Dorais et al. 2004), hujan dan kelembapan tinggi atau pengairan yang intensif setelah periode kekeringan (Simon 2006), suhu dan cahaya tinggi (Liebisch et al