Keanekaragaman Hymenoptera Parasitoid Di Perkebunan Kelapa Sawit Ptpn VIII Cindali, Bogor
Teks penuh
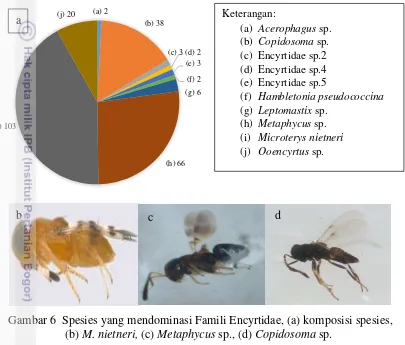
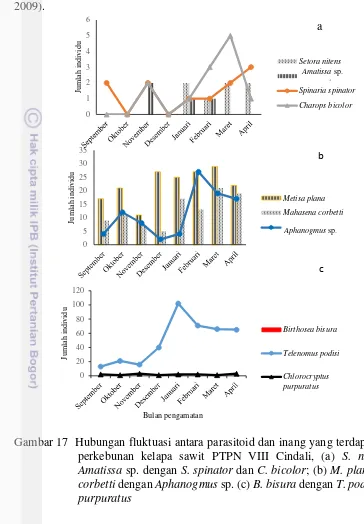
Gambar




Dokumen terkait
Cocopet yang ditemukan pada bunga jantan kelapa sawit di kebun Cimulang terdiri 3 spesies yang termasuk dalam tiga famili, yaitu Chelisochidae, Forficulidae, dan
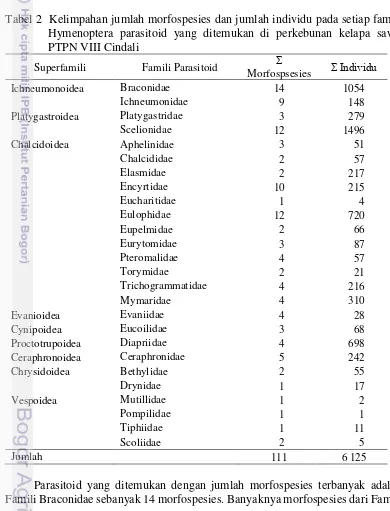
Kelimpahan dan keanekaragaman morfospesies parasitoid yang paling dominan berasal dari Ordo Hymenoptera, baik pada pertanaman kelapa sawit maupun padi sawah dengan
Famili, jumlah spesies, dan kelimpahan relatif Hymenoptera parasitoid pada tanaman sayuran dan vegetasi non-crop di lanskap persawahan Alahan Panjang dan Kayu
Meskipun pada skala lanskap tidak ada perbedaan dalam kekayaan spesies dan kelimpahan individu parasitoid pada enam tipe lanskap yang berbeda, kelimpahan Hymenoptera
Komposisi Hymenoptera parasitoid dari jumlah famili dan individu pada fase pertumbuhan tanaman vegetatif lebih rendah dibandingkan fase generatif dari kedua cara
Tujuan penelitian mengidentifikasi Hymenoptera parasitoid yang berasosiasi dengan serangga hama dan mengukur keanekaragaman serta dominansi Hymenoptera parasitoid di
Lanskap Nyalindung dengan struktur yang kompleks dan terdiri atas ekosistem padi, palawija, dan sayuran mempunyai jumlah individu, spesies, dan famili Hymenoptera yang lebih
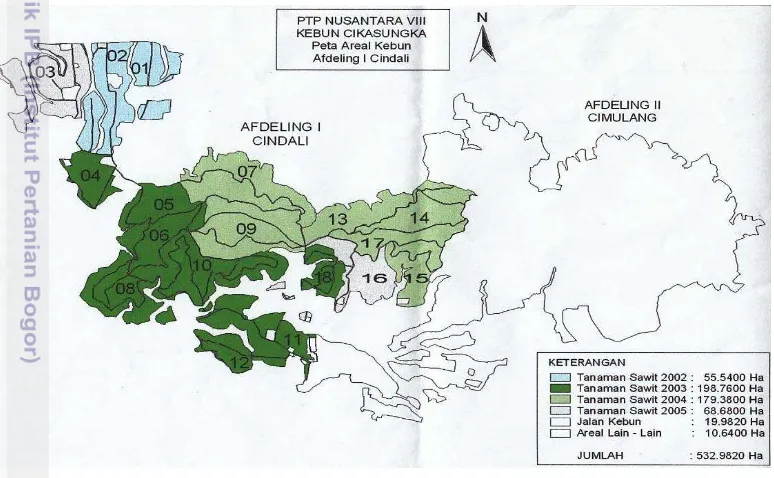
Tujuan dari penelitian ini adalah untuk mengetahui jenis dan persentase parasitisasi parasitoid yang memarasit hama pemakan daun di kebun kelapa sawit PTPN VIII, Cindali,
