PENGARUH TIGA JENIS MANGSA TERHADAP BIOLOGI
KEPIK PREDATOR Sycanus annulicornis DOHRN
(HEMIPTERA: REDUVIIDAE)
WINA PURWANINGRUM
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
SURAT PERNYATAAN
Dengan ini saya menyatakan bahwa tesis yang berjudul "Pengaruh Tiga Jenis Mangsa terhadap Biologi Kepik Predator Sycanus annulicornis Dohrn. (Hemiptera: Reduviidae)" adalah karya saya sendiri dan belum pernah diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Bogor, Oktober 2006
ABSTRAK
WINA PURWANINGRUM. Pengaruh Tiga Jenis Mangsa terhadap Biologi Kepik Predator Sycanus annulicornis Dohrn (Hemiptera: Reduviidae). Dibimbing oleh DADAN HINDAYANA, AUNU RAUF, dan SUGENG SANTOSO.
Penelitian ini bertujuan mempelajari pengaruh (1) tiga jenis mangsa terhadap biologi predator S. annulicornis, (2) rasio betina dan jantan terhadap fekunditas dan fertilitas telur predator, dan (3) status mangsa yang diberikan terhadap perilaku predator.
Penelitian dilaksanakan di Laboratorium Ekologi, Departemen Proteksi Tanaman, Fakultas Pertanian, Institut Pertanian Bogor, sejak bulan Juli 2005 sampai dengan Juni 2006. Predator S. annulicornis didapatkan dari lahan pertanian Kebun Percobaan IPB Sawah Baru dan Situ Gede, Darmaga Bogor. Mangsa yang digunakan adalah Corcyra cephalonica Stainton (Lepidoptera: Pyralidae), Tenebrio molitor Linnaeus (Coleoptera: Tenebrionidae) dan, Spodoptera litura Fabricius (Lepidoptera: Noctuidae) dalam status hidup dan mati.
Larva dan imago C. cephalonica didapatkan dari gudang beras di daerah Cianjur dan sekitarnya, larva T. molitor didapatkan dari pasar lokal yang ada di sekitar Bogor dan larva S. litura didapatkan dari pertanaman kedelai di sekitar Kampus IPB Darmaga. Ketiga jenis mangsa ini kemudian diperbanyak di laboratorium.
ABSTRACT
WINA PURWANINGRUM. Effects of Three Different Preys on Biology of Predator Bug Sycanus annulicornis Dohrn (Hemiptera: Reduviidae). Supervisors: DADAN HINDAYANA, AUNU RAUF, and SUGENG SANTOSO.
The objective of the research was to study the effects of (1) three different spesies of preys on the biology of predator S. annulicornis, (2) ratio of female to male on the fecundity and fertility of the predator, and (3) prey status on the predator behaviors.
The research was held at Ecology Laboratory, Departemen Proteksi Tanaman, Fakultas Pertanian, IPB, since July 2005 until June, 2006. Predator S. annulicornis was collected from agricultural field of Kebun Percobaan IPB Sawah Baru and Situ Gede, Darmaga Bogor. Preys used were Corcyra cephalonica Stainton (Lepidoptera: Pyralidae), Tenebrio molitor Linnaeus (Coleoptera: Tenebrionidae), and Spodoptera litura Fabricius (Lepidoptera: Noctuidae), both as dead and alive larvae.
Larvae and adult of C. cephalonica was collected from rice warehouse at Cianjur and surroundings, larvae T. molitor from local market at Bogor, while larvae S. litura from soybean fields at Kampus IPB Darmaga. Those preys then were reared in the laboratory.
© Hak cipta milik Institut Pertanian Bogor, tahun 2006
Hak cipta dilindungi
Dilarang mengutip dan memperbanyak tanpa izin tertulis dari Institut Pertanian Bogor, sebagian atau seluruhnya dalam bentuk apa pun,
PENGARUH TIGA JENIS MANGSA TERHADAP BIOLOGI
KEPIK PREDATOR Sycanus annulicornis DOHRN
(HEMIPTERA: REDUVIIDAE)
WINA PURWANINGRUM
Tesis
sebagai salah satu syarat untuk memperoleh gelar
Magister Sains pada
Program Studi Entomologi dan Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
BOGOR
Judul penelitian : Pengaruh Tiga Jenis Mangsa terhadap Biologi Kepik Predator Sycanus annulicornis Dohrn. (Hemiptera: Reduviidae)
Nama : Wina Purwaningrum
NIM : A451040041
Program Studi : Entomologi/Fitopatologi
Disetujui Komisi Pembimbing
Dr. Ir. Dadan Hindayana Ketua
Prof. Dr. Ir. Aunu Rauf, M.Sc. Anggota
Dr. Ir. Sugeng Santoso, M.Agr. Anggota
Diketahui Ketua Program Studi
Entomologi dan Fitopatologi
Dr. Ir. Sri Hendrastuti Hidayat, M.Sc.
Dekan Sekolah Pascasarjana
Prof. Dr.Ir. Khairil Anwar Notodiputro, MS.
PRAKATA
Dengan Rahmat Allah SWT, penulis dapat menyelesaikan tesis ini yang berjudul “Pengaruh Tiga Jenis Mangsa terhadap Biologi Kepik Predator Sycanus annulicornis Dohrn. (Hemiptera: Reduviidae).”
Penulis mengucapkan terimakasih kepada Dr. Ir. Dadan Hindayana, Prof. Dr. Ir. Aunu Rauf, M.Sc., dan Dr. Ir. Sugeng Santoso, M.Agr. selaku pembimbing, atas saran dan bimbingan yang diberikan selama penelitian hingga penulisan tesis. Penulis juga mengucapkan terima kasih kepada Kementrian Riset dan Teknologi atas bantuan dana yang diberikan selama penelitian karena penelitian ini merupakan bagian dari Riset Unggulan Terpadu 2005-2006.
Ungkapan terima kasih juga disampaikan kepada ayah, ibu, para sahabat dan adik-adik tercinta, atas segala doa dan kasih sayangnya. Semoga hasil penelitian ini bermanfaat dan berguna bagi perkembangan dan kemajuan pengetahuan khususnya di bidang entomologi.
Bogor, Oktober 2006
RIWAYAT HIDUP
Penulis dilahirkan di Cianjur pada tanggal 1 September 1980, sebagai anak tunggal pasangan Bapak E. Juanda dan Ibu Tuti Saadah.
Tahun 1998 penulis lulus dari Sekolah Menengah Atas Negeri I Cianjur. Pada tahun yang sama penulis diterima di IPB melalui jalur Undangan Seleksi Masuk IPB (USMI). Selama perkuliahan penulis aktif dalam kegiatan himpunan profesi (HIMASITA), menjadi asisten mata kuliah Hama dan Penyakit Tanaman Perkebunan dan Vertebrata Hama.
Tahun 2003 penulis pernah bekerja sebagai staf pengamat hama dan penyakit tanaman di PT. Sang Hyang Seri (PERSERO), dan dari tahun 2004 sampai dengan sekarang penulis bekerja sebagai staf di Direktorat Pengembangan Institusi dan Usaha Penunjang IPB.
DAFTAR ISI
Halaman
DAFTAR TABEL ... x
DAFTAR GAMBAR ... xi
DAFTAR LAMPIRAN ... xii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 2
TINJAUAN PUSTAKA ... 3
Karakteristik Predator ... 3
Perilaku Pemangsaan Predator... 4
Tahapan Penemuan Mangsa... 5
Predator Famili Reduviidae ... 6
Predator Sycanus annulicornis ... 7
Nutrisi Predator ... 8
Tingkat Konsumsi Rata-rata dan Efisiensi ... 11
BAHAN DAN METODE ... 13
Tempat dan Waktu Penelitian ... 13
Persiapan Penelitian ... 13
Perbanyakan Mangsa ... 13
Pemeliharaan S. annulicornis ... 15
Perlakuan Penelitian ... 16
Studi Pengaruh Tiga Jenis Mangsa terhadap Biologi S. annulicornis ... 16
Studi Efisiensi Penyerapan Makanan Predator S. annulicornis .... 17
Studi Rasio Betina dan Jantan terhadap Karakteristik Reproduksi Predator S. annulicornis ... 18
Studi Status Mangsa terhadap Perilaku Pemangsaan ... Predator ... 19
Rancangan Percobaan ... 19
HASIL DAN PEMBAHASAN ... 20
Hasil Penelitian ... 20
Morfologi Predator S. annulicornis... 21
Pengaruh Tiga Jenis Mangsa dan Status Mangsa terhadap Biologi S. annulicornis ... 21
Pengaruh Tiga Jenis Mangsa dan Status Mangsa terhadap Efisiensi Penyerapan Makanan Predator S. annulicornis... 23
Pengaruh Rasio Betina dan Jantan terhadap Karakteristik Reproduksi Predator S. annulicornis... 24
Pengaruh Status Mangsa terhadap Perilaku Pemangsaan Predator ... 26
DAFTAR TABEL
Halaman
1. Jumlah dan tahapan instar mangsa yang diberikan pada setiap tahapan instar predator S. annulicornis untuk perlakuan pemberian
mangsa dalam status mati dan hidup... 18 2. Persentase kematian S. annulicornis pada tiga jenis mangsa
dengan status mangsa hidup dan status mangsa mati... 21 3. Pengaruh status mangsa terhadap stadium nimfa (hari) dan lama
hidup imago (hari) predator S. annulicornis... 22 4. Pengaruh tiga jenis mangsa terhadap stadium nimfa (hari) dan
lama hidup imago (hari) predator S. annulicornis... 23 5. Pengaruh status mangsa terhadap total konsumsi selama stadium
nimfa, bobot tubuh predator dan ECI... 23 6. Pengaruh tiga jenis mangsa terhadap total konsumsi selama
stadium nimfa, bobot tubuh predator dan ECI ... 24 7. Praoviposisi, oviposisi, frekuensi peneluran, keperidian, rata-rata
jumlah telur yang menetas, persen penetasan telur, dan stadium telur predator S. annulicornis pada empat rasio yang berbeda
dengan mangsa C. cephalonica yang diberikan dalam status mati... 25 8. Masa pencarian (menit), masa penanganan (menit), masa
pengisapan (menit), dan jumlah mangsa yang dimangsa (ekor) oleh nimfa predator S. annulicornis dengan mangsa C. cephalonica
DAFTAR GAMBAR
Halaman
1. Perbanyakan C. cephalonica ... 13
2. Perbanyakan T. molitor ... 14
3. Perbanyakan S. litura ... 15
4. Perbanyakan S. annulicornis ... 16
5. Perkembangan instar nimfa predator S. annulicornis... 20
6. Paket telur ... 26
7. Predator S. annulicornis yang sedang memangsa C. cephalonica... 28
8. Konsumsi harian selama fase nimfa predator S. annulicornis pada mangsa C. cephalonica, T. molitor, dan S. litura dengan status hidup dan mati... 31
DAFTAR LAMPIRAN
Halaman 1 Hasil analisis proksimat ... 42 2 Hasil Uji Tukey pengaruh tiga jenis mangsa terhadap stadium instar
nimfa dan lama hidup imago ... 43 3 Hasil Uji-t pengaruh status mangsa terhadap stadium instar
nimfa dan lama hidup imago ... 46 4 Hasil Uji Tukey pengaruh tiga jenis mangsa terhadap total konsumsi, bobot
tubuh predator dan ECI ... 49 5 Hasil Uji -t pengaruh status mangsa terhadap total konsumsi, bobot tubuh
PENDAHULUAN
Latar Belakang
Sycanus annulicornis Dohrn (Hemiptera: Reduviidae) merupakan kepik predator yang hidup pada berbagai agroekosistem. Di Indonesia Sycanus sp. pernah dilaporkan menjadi predator utama hama pengisap daun, Helopeltis antonii Sign (Hemiptera: Miridae) pada tanaman teh (Kalshoven 1981). Kepik predator ini memiliki kisaran mangsa yang luas terutama dari Ordo Lepidoptera (Manley 1982). Di Malaysia, Sycanus sp. memangsa Phaenacantha saccharicida Karsch (Hemiptera: Colobathrisidae) (Sweet 2000). Di Thailand, Sycanus collaris Fabricius dilaporkan menyerang hama daun lengkeng Oxyodes scrobiculata Fabricius (Lepidoptera: Noctuidae) (Chooyoung et al. 2004).
Pada beberapa percobaan di laboratorium, kepik predator ini diketahui merupakan predator generalis, memiliki kemampuan adaptasi dan pemangsaan cukup baik dengan kemampuan memangsa mencapai rata-rata 4,7 ekor perhari (De Clercq 2000). Mangsa-mangsa yang pernah dicobakan untuk pemeliharaan di laboratorium adalah Spodoptera litura Fabricius (Lepidoptera: Noctuidae), Crocidolomia pavonana Fabricius (Lepidoptera: Pyralidae) dan Corcyra cephalonica Stainton (Lepidoptera: Pyralidae) (Cahyadi 2004).
Mencermati adanya potensi tersebut, maka diperlukan suatu metode yang tepat untuk pembiakan predator tersebut. Hasil pembiakan massal di laboratorium dapat digunakan untuk upaya pelepasan massal predator di lapangan. Penggunaan predator di lapangan untuk menekan populasi hama sesuai dengan konsep pengendalian hayati (PH) yaitu musuh alami berperan untuk memelihara kepadatan populasi organisme lain pada suatu tingkat populasi rata-rata yang lebih rendah daripada tingkat populasi rata-rata tanpa pengendalian hayati tersebut (Bosch et al. 1982).
dan nutrisi alternatif. Nutrisi essensial adalah nutrisi yang dapat mendukung pertumbuhan dan perkembangan organisme pradewasa dan reproduksi imago, sedangkan nutrisi alternatif adalah nutrisi yang hanya dapat menyokong atau bertahan suatu organisme (Hagen 1987).
Berkaitan dengan nutrisi yang diperlukan oleh predator, perlu dilakukan penelitian mengenai keberadaan mangsa yang sesuai untuk pertumbuhan dan perkembangan predator. Selain itu, kemudahan dalam pembiakan di laboratorium juga merupakan hal yang harus dipelajari. Memodifikasi status mangsa yang diberikan misalnya dalam status mati, adalah salah satu hal untuk memudahkan penyimpanan dan penyediaan mangsa secara kontinyu. Modifikasi mangsa dengan status mangsa mati akan memberi keuntungan diantaranya tidak perlu menyediakan pakan mangsa, menghemat tempat penyimpanan, memudahkan dalam pemeliharaan dan dapat menyediakan mangsa yang seragam setiap saat.
Hal yang dikhawatirkan dan penting sekali dipertimbangkan adalah kemungkinan berpengaruh terhadap perilaku predator pada saat pelepasan di lapangan, ketika predator tersebut harus memangsa mangsa yang hidup.
Tujuan Penelitian
TINJAUAN PUSTAKA
Karakteristik Predator
Entomofaga merupakan istilah dari bahasa Yunani (Entomon= insect: phagein= to eat) yaitu kelompok organisme yang menjadikan serangga sebagai makanannya untuk tumbuh dan berkembang. Yang termasuk salah satu serangga entomofaga yang berperan sebagai pengendali serangga hama adalah serangga predator (Bosch et al. 1982). Predator memiliki karakteristik sebagai berikut: 1) secara ekologi, predator memiliki fungsi penting karena dapat menyebabkan mortalitas tinggi terhadap mangsanya, 2) membinasakan atau melumpuhkan mangsa sebelum mengkonsumsi, 3) mengejar dan membunuh mangsa, 4) membunuh mangsa hanya untuk dirinya sendiri dengan cara mengunyah dan menghisap cairan tubuh mangsanya secara cepat, 5) membutuhkan mangsa dalam jumlah banyak selama hidupnya, 6) secara relatif memiliki ukuran tubuh lebih besar dari mangsanya, 7) memiliki kemampuan daya jelajah yang luas, 8) mampu mengatasi dan mengelabui mangsanya yang akif, 9) organ tubuh berkembang baik sehingga dapat bergerak cepat atau terbang jauh serta memiliki embelan yang memungkinkan melakukan fungsi yang lebih dari yang dimiliki serangga pada golongan lainnya, 10) beberapa memproduksi racun, 11) ada heteroeisme, 12) umumnya generalis, 13) beberapa mampu beradaptasi pada lingkungan yang ekstrim (Bosch et al. 1982: Taylor 1984; New 1991). Secara umum serangga yang berperan sebagai predator dapat ditemukan baik pada fase nimfa atau larva maupun fase imago, tetapi ada juga beberapa serangga predator yang hanya fase larva saja yang beperan sebagai predator, sebagai contoh adalah lalat Syrphidae yang memangsa kutu daun (Bosch et al. 1982).
Perilaku Pemangsaan Predator
Secara umum tahapan kejadian predator dalam menemukan mangsanya meliputi 1) pencarian dan pengarahan terhadap suatu mangsa, 2) pengejaran, 3) penangkapan dan 4) makan (New 1991). Predator memiliki metode yang bervariasi dalam menemukan mangsanya, mengejar mangsa dan atau diam (menunggu) tetapi menjerat mangsa dengan umpan ( New 1991 ).
Beberapa strategi predator saat menangkap mangsamencakup:
1. Diam-Menunggu, serangga predator laba-laba menangkap mangsa dengan cara menjerat menggunakan jaringnya. Serangga tersebut memiliki perilaku diam, menunggu dan memakan mangsa yang hanya terjerat dalam jaringnya. Serangga tipe ini memiliki tungkai yang panjang untuk menjaga jarak dari mangsa yang berbahaya, dan membuat jaring ditempat yang terdapat banyak mangsa. Perilaku diam dan menunggu akan menghasilkan periode puasa yang panjang, dan predator tersebut biasanya memiliki kapasistas yang luar biasa dalam menahan lapar.
2. Menyergap, predator diam pada suatu bunga, dan menunggu kehadiran serangga mangsa yang sedang mencari pollen/ nektar. Selama diam di dalam bunga predator tersebut melakukan kripsis, apabila serangga mangsa tersebut sudah mendekat, maka secara tiba-tiba predator tersebut menyergap, menangkap dan memangsanya.
3. Insinuasi, serangga predator secara perlahan berusaha menenangkan mangsanya (mangsa yang aktif) dengan menggunakan tungkai, setelah mangsa cukup tenang, maka predator menghisap cairan mangsanya.
5. Predator terbang (Ordo Odonata dan Ordo Diptera), memiliki daya penglihatan yang baik untuk memburu, predator tersebut memiliki perilaku diam di tempat yang agak tinggi, mengintai mangsanya dan bergerak hanya pada saat menangkap mangsa dengan demikian dapat mengefisienkan energi.
6. Kleptoparasitisme, predator yang mendapat mangsa dengan cara mencuri dari serangga lain.
Tahapan Penemuan Mangsa oleh Predator
Predator memiliki beberapa tahapan dalam menemukan mangsanya. Faktor kimia dan faktor fisik merupakan hal utama yang menuntun predator dalam menemukan mangsanya. Hagen (1987) dan New (1991), menjelaskan tahapan penemuan mangsa oleh predator yang terdiri dari penemuan habitat mangsa, penemuan mangsa, penerimaan mangsa dan kesesuaian mangsa.
a. Penemuan Habitat Mangsa
Kemoreseptor (olfaksi) adalah indera utama yang mengarahkan predator menuju habitat yang sesuai yang terdapat banyak mangsa, kairomon yang volatil biasanya berfungsi sebagai atraktan jarak jauh bagi predator dari semua ordo. Selain kimia, tuntunan fisik seperti warna, bentuk, tekstur habitat dapat membantu predator dalam menemukan habitat mangsa.
b. Penemuan Mangsa
predator menemukan mangsanya. Sebagai contoh, larva Aphidoletes aphidiomyza (Homoptera: Aphididae) dapat menemukan kutu daun melalui aroma embun madu yang merupakan ekskresi dari kutu daun tersebut, kemudian imago Syrphidae meletakkan telur pada tempat yang banyak mengandung embun madu.
c. Penerimaan Mangsa
Ukuran, bentuk, pergerakan, suara, dan faktor kimia internal-eksternal dari mangsa merupakan stimulan bagi serangga predator untuk menerima mangsanya. Beberapa serangga predator melalui antenanya dapat menemukan mangsa kemudian mencicipi mangsa untuk memeriksa kesesuaian mangsa tersebut. Apabila mangsa tersebut sesuai maka predator akan melanjutkan pemangsaan tetapi apabila tidak sesuai maka predator akan menolak mangsa tersebut.
d. Kesesuaian Mangsa
Apabila suatu mangsa telah diterima oleh predator, maka predator akan meneruskan memakan mangsa tersebut sebagai sarana untuk mendukung pertumbuhan dan perkembangan serta reproduksi predator, namun apabila mangsa tersebut tidak sesuai maka reaksi yang terjadi akan berbeda untuk setiap predator, 1) predator langsung memuntahkan mangsa tersebut, 2) predator seketika mengalami kematian disebabkan kandungan racun pada mangsa, 3) predator tetap hidup namun dengan pertumbuhan dan perkembangan yang sangat lambat, dan apabila berhasil mencapai imago maka lama hidupnya akan singkat dan memiliki fekunditas dan fertilitas yang sangat rendah.
Predator Famili Reduviidae
anggota lebih dari 1000 spesies. Serangga tersebut mempunyai kelimpahan yang tinggi sehingga dapat ditemukan di setiap daerah (Matheson 1951; Richards & Davies 1977).
Kepik Reduviidae biasa hidup disekitar semak-semak, tanaman herba, dan daun-daunan (Richards & Davies 1977). Mangsa utama dari serangga ini adalah larva Lepidoptera, lundi, kutu tanaman, dan hama-hama lainnya (Matheson 1951). Beberapa dari serangga ini juga dapat menjadi vektor penyakit, sebagai contoh yaitu penyakit chagas yang ditularkan oleh Rhodnius prolixus Stal. (Hemiptera: Reduviidae) di Amerika (Richards & Davies 1977).
Famili Reduviidae dapat dikenali dengan bentuk yang khas, yaitu antena seringkali memiliki segmen dengan total 40 ruas. Rostrum meruncing terdiri dari 3 segmen, tibia tungkai depan memiliki fosula hal ini berkaitan dengan daya adhesi, sedangkan bagian tarsus terdiri dari tiga segmen (Richards & Davies 1977). Kepik predator melumpuhkan mangsanya dengan mengeluarkan saliva yang beracun yang dapat menyebabkan paralisis pada mangsanya (Matheson 1951; Gillott 1995).
Predator Sycanus annulicornis
Sebagai salah satu contoh dari Famili Reduviidae adalah Sycanus annulicornis yang digolongkan dalam Ordo Hemiptera: Famili Reduviidae (Kalshoven 1981). S. annulicornis merupakan kepik predator yang sangat potensial. Kepik ini dapat hidup pada berbagai agroekosistem, baik pada agroekosistem tanaman pangan, sayuran maupun perkebunan dengan kisaran mangsa yang luas, terutama dari Ordo Lepidoptera (Manley 1982).
predator dan parasitoid telur (Gillot 1995). Lama stadia telur yaitu antara 14-16 hari (Cahyadi 2004).
Nimfa instar I berwarna jingga dengan ukuran panjang tubuh ±1,73 mm. Nimfa yang baru menetas biasanya berkumpul disekitar paket telur dengan memakan sisa-sisa telur. Lama stadia nimfa instar I yaitu 11 hari. Nimfa instar II berwarna jingga dengan ukuran ± 4,26 mm. Lama stadia nimfa instar II yaitu 8 hari. Nimfa instar III berwarna jingga dan pada seluruh tungkai terdapat bercak yang berwarna hitam. Ukuran panjang tubuh ± 8,5 mm. Lama stadia nimfa instar III yaitu 8 hari. Nimfa instar IV berwarna jingga kecoklatan dengan warna hitam pada tungkai dan toraks. Ukuran panjang tubuh ± 12-15 mm. Lama stadia nimfa instar IV yaitu 12 hari. Nimfa instar V berwarna jingga tua kecoklatan dengan warna hitam pada bagian toraks, abdomen dan keseluruhan tungkai. Ukuran panjang tubuh ± 14-15 mm. Lama stadium yaitu 19 hari (Cahyadi 2004).
Imago S. annulicornis berwarna hitam dengan corak berwarna jingga pada sayap bagian depan. Imago berukuran antara 19-25 mm, dengan lama hidup yaitu 24 hari (Cahyadi 2004).
Proses ganti kulit pada predator S. annulicornis ditandai dengan nimfa yang akan berganti kulit tidak beraktivitas, berwarna jingga pucat, kemudian memulai proses ganti kulit dengan menyobek eksuvia toraks bagian dorsal lalu ke bagian kepala dan instar baru akan keluar dengan cara kepala keluar terlebih dahulu diikuti tungkai bagian depan. Nimfa yang baru berganti kulit berwarna jingga pucat kemudian berubah warnanya menjadi lebih tua, sedangkan imago yang baru terbentuk berwarna coklat kehitaman (Cahyadi 2004).
Nutrisi Predator
dapat dijadikan sebagai kisaran mangsanya. Kesesuaian mangsa dapat dievaluasi dengan cara mempelajari pertumbuhan, perkembangan, daya tahan, dan fekunditas predator (Hagen 1987; Toft & Wise 1999).
Sebagai contoh yaitu kepik Podisus maculiventris Say (Heteroptera: Pentatomidae) memerlukan mangsa yang berukuran kecil untuk perkembangan selama fase nimfa, tetapi serangga tersebut memerlukan mangsa dengan ukuran yang lebih besar untuk reproduksi dari fase imago. Contoh lainnya yaitu predator kumbang kubah merupakan predator utama dari kutu daun, namun tidak semua spesies kutu daun sesuai untuk perkembangan kumbang kubah tersebut. Beberapa kutu daun bersifat toksik, langsung mematikan predator tersebut atau menghambat pertumbuhan dan perkembangan predator (New 1991; Dixon 2000).
Dipandang dari segi kualitas, makanan untuk predator dikategorikan menjadi nutrisi essensial dan nutrisi alternatif. Nutrisi essensial adalah sumber makanan yang mengandung nutrisi yang dapat mendukung pertumbuhan dan perkembangan organisme pradewasa dan reproduksi imago sedangkan nutrisi alternatif adalah sumber makanan yang mengandung nutrisi yang hanya dapat menyokong atau bertahan suatu organisme (Hagen 1987). Nutrisi yang dibutuhkan serangga menurut (Hagen 1987; Chapman 1998) yaitu :
1). Asam amino : tersedia dalam bentuk protein dan secara struktur membentuk enzim, setiap serangga membutuhkan kadar protein yang berbeda. Enzim berfungsi sebagai media transport dan penyimpanan dan sebagai molekul reseptor. Sebagai contoh, Tyrosine penting untuk serangga dalam proses sklerotisasi.
2). Karbohidrat : tidak termasuk ke dalam kategori essensial untuk serangga pada umumnya, lebih umum diperlukan sebagai sumber energi. Karbohidrat dapat disintesis dari asam amino.
4). Vitamin : dibutuhkan untuk mendukung berjalannya fungsi tubuh, vitamin juga dibutuhkan untuk membentuk jaringan tubuh. Sebagai contoh β–carotene (provitamin A) berguna sebagai komponen pigmen penglihatan, α-tocopherol (Vitamin E) penting untuk reproduksi, fertilitas dan perkembangan embrio. 5). Mineral: dibutuhkan untuk pertumbuhan dan perkembangan tetapi dibutuhkan
dalam jumlah sedikit.
6). Purines dan pyrimidines : DNA dan RNA adalah molekul yang membawa dan memediasi kode genetik.
7). Air: penting untuk serangga secara umum.
Terdapat perbedaan kebutuhan komposisi jenis nutrisi pada setiap fase predator, sebagai contoh penelitian yang dilakukan oleh Niijima (1993 dalam Thompson & Hagen 1999) melaporkan bahwa larva Chrysopa pallens Wesm. (Neuroptera: Chrysopidae) membutuhkan asam amino yang lebih tinggi bila dibandingkan dengan fase imagonya. Contoh lainnya yaitu Chrysopa nigricornis Burro. (Neuroptera: Chrysopidae) hanya membutuhkan kutu daun untuk keberhasilan kopulasi, sedangkan Chrysopa quadripunctata Burm. (Neuroptera: Chrysopidae) selain membutuhkan aphid juga membutuhkan gula dan air yang berlebih Roussett (1984 dalam Thompson & Hagen 1999).
Serangga gudang seperti Corcyra cephalonica (Lepidoptera: Pyralidae) dan Tenebrio molitor (Coleoptera: Tenebrionidae) merupakan serangga gudang yang diindikasikan memiliki kadar protein yang cukup tinggi, sedangkan Spodoptera litura (Lepidoptera: Noctuidae) merupakan serangga pemakan tanaman yang diindikasikan memiliki kadar protein yang rendah. Perbedaan bioekologi serangga tersebut dapat menyebabkan perbedaan komposisi nutrisi dalam tubuhnya.
mempunyai metamorfosis sempurna (holometabola), sehingga mengalami stadia telur, larva, pupa dan imago.
Kerusakan komoditas pertanian akibat serangan C. cephalonica pada stadium larva berupa penyusutan kuantitas maupun kualitas. Penyusutan kuantitas terjadi akibat aktivitas makan sehingga bobot bahan makanan menjadi berkurang. Penyusutan kualitas terjadi karena kontaminasi bahan makanan dengan kotoran larva, serangan ngengat beras ini pada stadium larva dapat menyebabkan penggumpalan benang sutera pada komoditas serta komoditas menjadi berlubang (Rosadi 2001).
Serangga S. litura digolongkan dalam Ordo Lepidoptera: Famili Noctuidae (Matheson 1951; Kalshoven 1981). Serangga ini lebih dikenal sebagai ulat grayak dan merupakan hama kosmopolit yang menyerang banyak tanaman budidaya seperti tembakau, kedelai, kangkung dan talas. S. litura tersebar di Asia, kepulauan Pasifik dan Australia (Kalshoven 1981). Di benua Asia hama ini tersebar di India, Pakistan, Srilangka, Birma, Filipina, dan Taiwan (Grist dan Lever 1969). S. litura mempunyai metamorfosis sempurna (holometabola), sehingga mengalami stadia telur, larva, pupa dan imago.
Serangga T. molitor merupakan salah satu contoh dari Ordo Coleoptera: Famili Tenebrionidae, serangga ini dikenal sebagai ”Ulat Hongkong”, dan merupakan hama gudang yang bersifat kosmopolit. Terdapat kurang lebih 100 spesies yang berasosiasi dengan produk pasca panen seperti tepung, terigu dan produk pasca panen lainnya (Munro 1966). Spesies dari genus ini hidup di kotoran binatang, sisa binatang yang telah mati, cendawan, kulit kayu, akar tanaman muda, humus, dan daun yang telah gugur (Clausen 1940; Toerngadi et al. 1974; Richards & Davies 1977; Kalshoven 1981).
Tingkat Konsumsi Rata-Rata dan Efisiensi
menjadi komponen tubuh disebut dengan Efisiensi Penyerapan Makanan atau Efficiency of Conversion of Ingested food (ECI). Nilai ECI merupakan suatu parameter efisiensi penyerapan makanan oleh predator yang mampu diubah menjadi komponen tubuh. Semakin tinggi nilai ECI maka semakin efisien penyerapan kandungan nutrisi mangsa oleh predator (Slansky & Rodriguez 1987).
Serangga Heteroptera merupakan kelompok yang dikenal efisien dalam pemanfaatan mangsanya dan memiliki kisaran nilai ECI 21,5%-53%. Nilai ECI umumnya bervariasi dari setiap spesies dan tahapan instar serangga, sebagai contoh, nilai ECI predator Podisus maculiventris instar 2-5 yang memangsa larva lepidoptera berturut-turut adalah 37,4%; 50,9%; 54,7% dan 45,2%. Predator Blepharidopterus angulatus Fallen. (Hemiptera: Miridae) yang memangsa kutu daun, memiliki nilai ECI dengan kisaran 39%-47% untuk instar 1-4, dan 28% untuk imago (Hagen 1987).
Tempat dan Waktu Penelitian
Penelitian dilakukan di Laboratorium Ekologi Serangga, Departemen Proteksi Tanaman, Institut Pertanian Bogor. Penelitian ini dilaksanakan sejak bulan Juni 2005 sampai dengan Juni 2006.
Persiapan Penelitian Perbanyakan Mangsa
Penelitian ini menggunakan tiga jenis mangsa, yaitu C. cephalonica, T. molitor, dan S. litura. Serangga mangsa yang diperoleh dari habitat aslinya selanjutnya diperbanyak di laboratorium.
Larva dan imago C. cephalonica diperoleh dari daerah Cianjur. Larva C. cephalonica diperbanyak di dalam kotak plastik berisi beras menir steril. Imago yang diperoleh dari lapang ditempatkan di dalam kurungan dari karton dan berbentuk silinder dengan diameter 10,5 cm dan tinggi 12,5 cm. Bagian atas dan bawah silinder diberi saringan kawat untuk ventilasi. Pada bagian bawah diletakkan kertas berwarna gelap untuk menampung telur yang diletakkan. Telur yang terkumpul disebarkan secara merata pada kotak plastik yang berisi beras menir steril dan dibiarkan menetas sampai menjadi larva (Gambar 1).
a b
Larva T. molitor diperoleh dari pasar lokal di sekitar wilayah Bogor. Larva T. molitor banyak dijual sebagai pakan burung. Perbanyakan T. molitor dilakukan di dalam kotak plastik berisi pur jagung yang merupakan pakan imago dan sebagai media untuk bertelur. Setelah imago melakukan kopulasi dan menghasilkan telur, imago dipindah ke tempat biakan lainnya (Gambar 2).
a b
Gambar 2 Perbanyakan T. molitor (a) pur jagung sebagai pakan dan tempat pembiakan larva T. molitor, (b) larva T. molitor yang berhasil menjadi imago
Larva S. litura berasal dari pertanaman kedelai yang ada disekitar Kampus IPB Dramaga, yaitu Bubulak dan Situgede. Larva S. litura yang diperoleh, dipelihara di dalam kotak plastik berventilasi.
Setiap hari larva diberi pakan daun tanaman kedelai segar. Tanaman kedelai tersebut ditanam dalam polibag dengan media tanah dan pupuk kandang dengan perbandingan 1:5. Tanaman diberi pupuk NPK (16:16:16) dengan bobot 2,5 g per polibag.
b a
c
Gambar 3 Perbanyakan S. litura (a) daun kedelai sebagai pakan dan tempat pembiakan larva S. litura, (b) serbuk gergaji sebagai tempat berpupa, (c) kurungan plastik sebagai tempat imago berkopulasi
Pemeliharaan S. annulicornis
Predator S. annulicornis diperoleh dari daerah Bubulak, Situgede dan Sawah Baru, Kabupaten Bogor. Setiap pasang imago yang diperoleh dari lapang ditempatkan di dalam wadah plastik yang berdiameter 6,5 cm. Pasangan imago tersebut terus dipelihara hingga berkopulasi dan meletakkan telur. Telur yang diletakkan oleh imago dipindahkan ke wadah lain agar tidak terganggu oleh imago.
Nimfa instar pertama tidak diberi mangsa selama tiga hari, tetapi nimfa tersebut hanya diberi kapas lembab. Selama fase nimfa instar pertama, tidak dilakukan pemisahan per individu predator karena nimfa instar pertama masih bergerombol dan memakan sisa-sisa paket telur. Setelah memasuki instar kedua,
a b
[image:30.595.140.475.123.429.2]d c
Gambar 4 Perbanyakan S. annulicornis (a) wadah plastik tempat imago berkopulasi, (b) paket telur, (c) telur yang baru menetas, (d) wadah plastik tempat pemeliharaan nimfa
Perlakuan Penelitian
Studi Pengaruh Tiga Jenis Mangsa terhadap Biologi S. annulicornis
Studi ini dilakukan untuk melihat pengaruh tiga jenis mangsa terhadap S. annulicornis. Mangsa yang diberikan adalah larva C. cephalonica, T. molitor, dan S. litura. Dalam studi ini, mangsa yang diberikan dalam 2 status, yaitu status mati (larva dibekukan dalam lemari pendingin) dan status hidup.
Jumlah dan tahapan instar mangsa yang diberikan pada setiap tahapan
instar predator terdapat pada Tabel 1. Untuk mematikan mangsa, larva C. cephalonica dan S. litura dibekukan selama 1 jam sedangkan larva T. molitor
Untuk mengetahui banyaknya kebutuhan konsumsi harian predator, pada setiap pergantian mangsa dilakukan penimbangan bobot tubuh mangsa sebelum dan setelah dimangsa. Penimbangan bobot tubuh predator dilakukan saat predator berganti kulit menjadi imago. Parameter yang diamati adalah mortalitas nimfa dan imago, lama stadium tiap instar nimfa, lama hidup imago, jumlah biomassa (cairan) yang diisap oleh predator selama fase nimfa dan imago, dan bobot tubuh imago. Masing-masing perlakuan diulang 20 kali.
Studi Efisiensi Penyerapan Makanan Predator S. annulicornis
Besarnya cairan tubuh mangsa yang diserap dan kemudian diubah menjadi komponen tubuh disebut dengan Efisiensi Penyerapan Makanan atau Efficiency of Conversion of Ingested Food (ECI). Nilai ECI merupakan suatu parameter efisiensi penyerapan makanan oleh predator yang mampu diubah menjadi komponen tubuh. Semakin tinggi nilai ECI maka semakin efisien penyerapan kandungan nutrisi mangsa oleh predator.
E.C.I = Pertambahan bobot predator = Bobot akhir - Bobot awal x100% Bobot mangsa yang diisap Bobot mangsa yang diisap
x100%
Tabel 1 Jumlah dan tahapan instar mangsa yang diberikan pada setiap tahapan instar predator S. annulicornis untuk perlakuan pemberian mangsa dalam status mati dan hidup
Jenis mangsa Tahapan instar predator
Jumlah mangsa yang diberikan
(ekor)
Tahapan instar mangsa
C. cephalonica I 15 II
II 2 II III 2 III IV 3 IV V 3 IV Imago 3 IV
T. molitor I 15 II
II 2 II III 2 II IV 2 III V 2 III Imago 2 III
S. litura I 15 II
II 2 II III 2 II IV 2 III V 2 III Imago 2 III
Studi Rasio Betina dan Jantan terhadap Karakteristik Reproduksi Predator
S. annulicornis
Studi ini dilakukan dengan dasar 1) pengamatan di lapangan imago jantan lebih sering ditemukan dibanding imago betina, 2) pengamatan di laboratorium menunjukkan fertilitas telur predator yang bervariasi. Dalam studi ini mangsa yang digunakan adalah mangsa yang sesuai untuk predator yang didasarkan hasil Studi “Pengaruh Tiga Jenis Mangsa terhadap Biologi Predator S. annulicornis”.
Parameter yang diamati adalah lama praoviposisi, lama oviposisi, frekuensi peneluran, keperidian, lama stadium telur, dan persen penetasan telur.
Studi Status Mangsa terhadap Perilaku Predator
Studi ini dilakukan untuk melihat pengaruh pemberian mangsa dalam status mati terhadap sifat-sifat alami predator yaitu agresifitas yang dicerminkan dengan : 1) masa pencarian, yaitu waktu yang dibutuhkan untuk mencari dan menemukan mangsa, 2) masa penanganan, yaitu total waktu yang diperlukan untuk menangani dan membunuh mangsa, 3) masa pengisapan yaitu waktu yang dibutuhkan untuk mengkonsumsi satu ekor mangsa, dan 4) jumlah mangsa yang dikonsumsi per hari. Predator yang digunakan adalah keturunan kedua dari predator yang diperoleh dari lapangan. Dalam studi ini mangsa yang digunakan adalah mangsa yang sesuai untuk predator yang didasarkan pada hasil Studi “Pengaruh Tiga Jenis Mangsa terhadap Biologi S. annulicornis “ dan diberikan dalam status hidup.
Selama fase nimfa instar pertama tidak dilakukan pemisahan serangga uji. Setelah instar kedua serangga uji dipisahkan satu per satu ke dalam wadah plastik berdiameter 4 cm. Pengamatan dilakukan selama 4 jam. Perlakuan diulang 5 kali.
Rancangan Percobaan
HASIL DAN PEMBAHASAN Hasil Penelitian
Morfologi Predator S. annulicornis
Stadium nimfa yaitu masa sejak nimfa keluar dari telur hingga menjadi imago. Sebagian besar nimfa yang diberi tiga jenis mangsa dengan status yang berbeda dapat mencapai imago. Predator S. annulicornis mengalami lima tahapan instar nimfa yaitu nimfa instar I hingga nimfa instar V (Gambar 5).
a b
c
d
[image:34.595.137.499.268.511.2]e f
Gambar 5 Perkembangan instar nimfa predator S. annulicornis (a) nimfa instar 1, (b) nimfa instar 2, (c) nimfa instar 3, (d) nimfa instar 4, (e) nimfa instar 5 dan (f) imago
S. annulicornis berwarna hitam atau lebih gelap dibanding nimfa yang berwarna jingga kehitaman dan memiliki corak berwarna jingga pada sayap bagian depan.
Pengaruh Tiga Jenis Mangsa dan Status Mangsa terhadap Biologi
S. annulicornis.
Jenis mangsa dengan kandungan kualitas nutrisi yang berbeda dan dengan status yang berbeda dapat mempengaruhi mortalitas nimfa predator, stadium instar nimfa dan lama hidup imago.
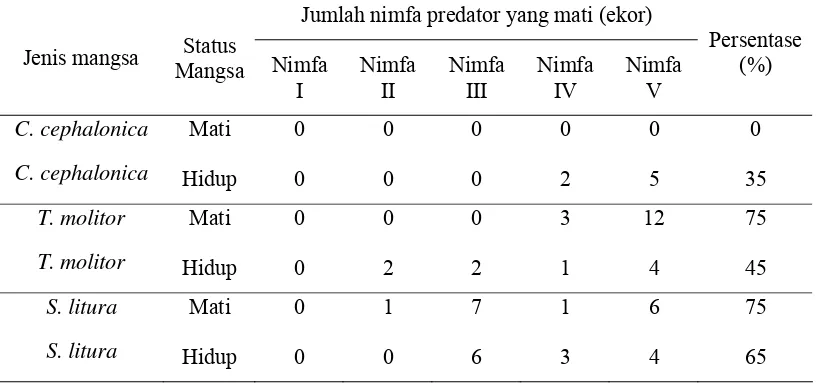
Tiga jenis mangsa yang berbeda yaitu C. cephalonica, T. molitor dan S. litura dengan status hidup dan mati memberikan pengaruh nyata terhadap mortalitas nimfa predator.
Tabel 2 Persentase kematian predator S. annulicornis pada tiga jenis mangsa dengan status mangsa hidup dan status mangsa mati
Jumlah nimfa predator yang mati (ekor)
Jenis mangsa Status
Mangsa Nimfa I
Nimfa II
Nimfa III
Nimfa IV
Nimfa V
Persentase (%)
C. cephalonica Mati 0 0 0 0 0 0
C. cephalonica Hidup 0 0 0 2 5 35
T. molitor Mati 0 0 0 3 12 75
T. molitor Hidup 0 2 2 1 4 45
S. litura Mati 0 1 7 1 6 75
S. litura Hidup 0 0 6 3 4 65
[image:35.595.109.517.399.591.2]hidup atau status mati tidak terlalu berbeda, (50%) untuk mangsa dengan status mati dan (48,34%) untuk mangsa dengan status hidup.
[image:36.595.111.516.364.443.2]Faktor status mangsa tidak memberikan pengaruh nyata terhadap total stadium instar nimfa dan lama hidup imago (Tabel 3). Pemberian mangsa dengan status mati terhadap predator, memiliki total stadium nimfa (54,23±7,61) dan total stadium nimfa dengan mangsa pada status mangsa hidup yaitu (54,41±0,45). Pemberian mangsa dengan status hidup menghasilkan lama hidup imago yaitu (26,07±15,14), sedangkan pemberian mangsa dengan status mati menghasilkan lama hidup imago yaitu (26,87± 9,37) (Tabel 3).
Tabel 3 Pengaruh status mangsa terhadap stadium nimfa (hari) dan lama hidup imago (hari) predator S. annulicornis
Stadium Nimfa Instar ke-
Status Mangsa
I II III IV V
Total
stadium
nimfa
Lama hidup
imago
Hidup 7 7,52±1,60a 10,26±3,71a 12,81±5,78a 16,82±4,94a 54,41±0,45a 26,07±15,14a
Mati 9,7 9,34±4,75b 8,40±3,08b 12,23±5,84a 14,56±3,68a 54,23±7,61a 26,87± 9,37a
Rataan dalam kolom yang diikuti oleh huruf yang sama tidak berbeda nyata (Uji t, α = 0,05)
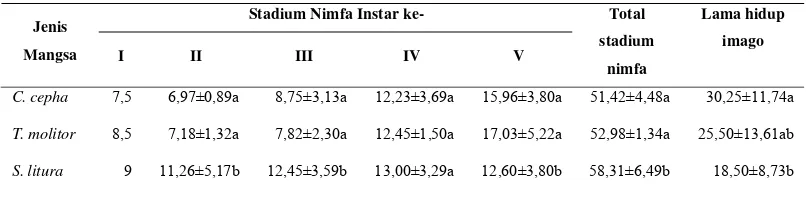
Tabel 4 Pengaruh tiga jenis mangsa terhadap stadium nimfa (hari) dan lama hidup imago (hari) predator S. annulicornis
Stadium Nimfa Instar ke- Jenis
Mangsa I II III IV V
Total
stadium
nimfa
Lama hidup
imago
C. cepha 7,5 6,97±0,89a 8,75±3,13a 12,23±3,69a 15,96±3,80a 51,42±4,48a 30,25±11,74a
T. molitor 8,5 7,18±1,32a 7,82±2,30a 12,45±1,50a 17,03±5,22a 52,98±1,34a 25,50±13,61ab
S. litura 9 11,26±5,17b 12,45±3,59b 13,00±3,29a 12,60±3,80b 58,31±6,49b 18,50±8,73b
Rataan dalam kolom yang diikuti oleh huruf yang sama tidak berbeda nyata (Uji Tukey, α = 0,05)
Pengaruh Tiga Jenis Mangsa dan Status Mangsa terhadap Efisiensi Penyerapan Makanan S. annulicornis
Status mangsa hidup dan mati tidak memberikan pengaruh nyata terhadap total konsumsi predator selama fase nimfa, bobot tubuh dan ECI. Walaupun demikian, status mangsa mati menghasilkan total konsumsi selama fase nimfa predator yang lebih tinggi (0,50g) dibanding dengan total konsumsi selama fase nimfa predator dengan status mangsa hidup (0,48 g) (Tabel 5).
Tabel 5 Pengaruh status mangsa terhadap total konsumsi selama stadium nimfa, bobot tubuh predator dan ECI
Parameter Status Mangsa
Total Konsumsi (g) Bobot Tubuh (g) ECI (%)
Hidup 0,48±0,13a 0,10±0,03a 25,15±4,72a
Mati 0,50±0,09a 0,12±0,00a 27,89±4,03a
Rataan dalam kolom yang diikuti oleh huruf yang sama tidak berbeda nyata (Uji t, α = 0,05)
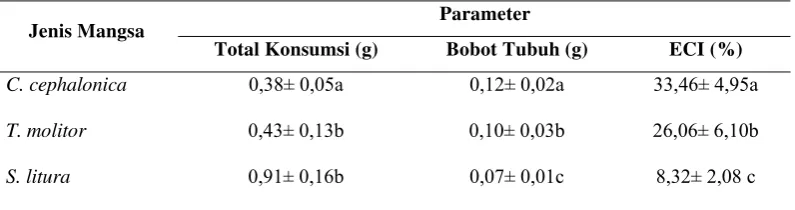
[image:37.595.108.514.489.571.2]Tabel 6 Pengaruh tiga jenis mangsa terhadap total konsumsi selama stadium nimfa, bobot tubuh predator dan ECI
Parameter Jenis Mangsa
Total Konsumsi (g) Bobot Tubuh (g) ECI (%)
C. cephalonica 0,38± 0,05a 0,12± 0,02a 33,46± 4,95a
T. molitor 0,43± 0,13b 0,10± 0,03b 26,06± 6,10b
S. litura 0,91± 0,16b 0,07± 0,01c 8,32± 2,08 c
Rataan dalam kolom yang diikuti oleh huruf yang sama tidak berbeda nyata (Uji Tukey, α = 0,05)
Tiga jenis mangsa yang berbeda memberikan pengaruh nyata terhadap total konsumsi predator selama fase nimfa, bobot tubuh dan ECI. Total konsumsi tertinggi adalah predator dengan mangsa S. litura (0,91g), sedangkan total konsumsi terrendah adalah predator dengan mangsa C. cephalonica (0,38g). Bobot tubuh predator tertinggi adalah predator dengan mangsa C. cephalonica (0,12 g), sedangkan bobot tubuh terrendah adalah predator dengan mangsa S. litura (0,07 g). Nilai ECI tertinggi ditunjukan pada predator dengan mangsa C. cephalonica (33,46%), sedangkan nilai ECI terrendah ditunjukan pada predator dengan mangsa S. litura (8,32%) (Tabel 6).
Pengaruh Rasio Betina dan Jantan terhadap Karakteristik Reproduksi Predator S. annulicornis
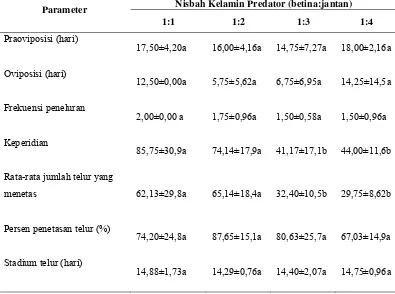
Tabel 7 Praoviposisi, oviposisi, frekuensi peneluran, keperidian, rata-rata jumlah telur yang menetas, persen penetasan telur, dan stadium telur predator S. annulicornis pada empat rasio yang berbeda dengan mangsa C. cephalonica yang diberikan dalam status mati
Nisbah Kelamin Predator (betina:jantan) Parameter
1:1 1:2 1:3 1:4
Praoviposisi (hari)
17,50±4,20a 16,00±4,16a 14,75±7,27a 18,00±2,16a
Oviposisi (hari)
12,50±0,00a 5,75±5,62a 6,75±6,95a 14,25±14,5a
Frekuensi peneluran
2,00±0,00 a 1,75±0,96a 1,50±0,58a 1,50±0,96a
Keperidian
85,75±30,9a 74,14±17,9a 41,17±17,1b 44,00±11,6b
Rata-rata jumlah telur yang
menetas 62,13±29,8a 65,14±18,4a 32,40±10,5b 29,75±8,62b
Persen penetasan telur (%)
74,20±24,8a 87,65±15,1a 80,63±25,7a 67,03±14,9a
Stadium telur (hari)
14,88±1,73a 14,29±0,76a 14,40±2,07a 14,75±0,96a
Rataan dalam baris yang diikuti oleh huruf yang sama tidak berbeda nyata (Uji BNT, α = 0,05)
Hasil penelitian menunjukkan kisaran masa praoviposisi yaitu dari 14 hari sampai dengan 18 hari. Masa oviposisi terpanjang terjadi pada rasio 1:4 (14,25±14,59) sedangkan masa oviposisi terpendek pada rasio 1:2 (5,75±5,62). Selama masa hidupnya predator meletakkan telur antara 1,5 kali sampai dengan 2 kali. Secara statistik, perbedaan masa praoviposisi, masa oviposisi dan frekuensi peneluran pada ke empat nisbah kelamin tidak signifikan (p>0,05).
jumlah telur yang menetas pada ke empat nisbah kelamin adalah signifikan (p<0,05). Persen penetasan telur predator tertinggi adalah pada nisbah kelamin 1:2 (87.65 ± 15.12) dan yang terrendah adalah pada nisbah kelamin 1:4 (67.03 ± 14.96). Semua rasio betina dan jantan menunjukkan rata-rata stadium telur yaitu 14 hari. Secara umum predator meletakkan telur pada permukaan tutup wadah yang terdapat kain kasa.
Gambar 6 Paket telur S. annulicornis
Telur berbentuk jorong, berwarna coklat, dan diletakkan pada paket yang tersusun dalam beberapa baris (Gambar 6). Imago betina meletakkan telur pada kain kasa karena permukaannya yang agak kasar sehingga dapat mencegah telur jatuh.
Pengaruh Status Mangsa terhadap Perilaku Pemangsaan Predator
Dalam studi ini adalah melihat pengaruh dari pemberian mangsa C. cephalonica dalam status mati terhadap perilaku predator keturunannya,
Tabel 8 Masa pencarian (menit), masa penanganan (menit), masa pengisapan (menit), dan jumlah mangsa yang dimangsa (ekor) oleh nimfa predator S. annulicornis dengan mangsa C. cephalonica dengan status hidup
Tahapan instar predator Aktivitas
I II III IV V
Penemuan (H)a
20,44±9,58a 22,96±12,94a 17,43±5,19a 10,86±0,48a 10,95±5,74a
Penemuan (M)a
19,67±10,58a 24,68±15,49a 18,59±3,63a 10,72±0,41a 9,62±2,56b
Penanganan (H)a
5,55±3,78a 6,20±3,64a 3,16±2,57a 2,33±0,39a 2,39±0,52a
Penanganan (M)a
5,89±2,98a 5,04±3,39a 2,84±2,08a 1,97±0,56a 1,82±0,63b
Pengisapan (H)a
163,33±26,46a 119,82±13,72a 55,94±12,94a 38,24±1,37a 44,11±8,93a
Pengisapan (M)a
165,56±17,40a 109,04±9,74a 51,69±7,26b 37,57±2,98a 44,35±9,35a
Jml mangsa (H)a
7,78±0,97a 1,33±0,47a 1,71±0,06a 2,41±0,10a 2,73±0,03a
Jml mangsa (M)a
7,78±0,67a 1,54±0,50a 1,79±0,19a 2,49±0,12a 2,78±0,10a
Rataan dalam satu kolom dan aktivitas, yang diikuti oleh huruf yang sama tidak berbeda nyata (Uji -T, α = 0,05)
a
(H) = keturunan dari induk yang diberi mangsa C. cephalonica status hidup (M) = keturunan dari induk yang diberi mangsa C. cephalonica status mati
Masa pencarian dapat diamati saat predator dilepaskan hingga menemukan mangsa. Predator biasanya tidak langsung dapat menangkap mangsa namun berkeliling dulu di sekitar mangsa.
[image:41.595.109.516.167.480.2]predator untuk menemukan mangsanya dengan mengenali senyawa kimia yang dikeluarkan oleh tubuh mangsa dan fesesnya.
[image:42.595.166.460.272.496.2]Masa pengisapan dapat diamati sejak predator menusukkan stiletnya sampai selesai mengisap cairan tubuh mangsa. Apabila cairan tubuh mangsa telah diisap, larva tampak mengempis dan menjadi keriput (Gambar 7). Setelah selesai mengisap mangsa predator melakukan gerakan mengusap-usap antena dengan menggunakan tungkai depan.
Gambar 7 Predator S. annulicornis yang sedang memangsa larva C. cephalonica
Jumlah mangsa yang dimangsa dihitung dengan menghitung jumlah mangsa yang dikonsumsi oleh predator per hari. Pada tahapan pemberian mangsa kepada predator nimfa instar I, tidak dilakukan pemisahan predator karena perilaku predator nimfa instar I masih bergerombol, berukuran kecil, dan tubuhnya masih menempel pada cangkang telurnya.
secara statistika tidak memberikan pengaruh nyata terhadap agresifitas predator keturunannya atau tidak berpengaruh buruk terhadap perilaku pemangsaan oleh predator keturunannya.
Pembahasan
Tiga jenis mangsa dan dengan status mangsa yang berbeda memberikan pengaruh terhadap mortalitas nimfa, stadium nimfa, lama hidup imago, total konsumsi selama fase nimfa, bobot tubuh imago, serta nilai ECI. Dua faktor tersebut, yaitu perbedaan mangsa dan status mangsa merupakan hal utama yang harus diperhatikan untuk mendapatkan hasil yang diharapkan pada saat melakukan pembiakan massal.
Faktor pertama yaitu perbedaan jenis mangsa, perbedaan jenis mangsa menyebabkan persentase mortalitas yang berbeda pada fase nimfa predator, mortalitas yang terjadi disebabkan karena tiga jenis mangsa tersebut memiliki komposisi dan kisaran nutrisi yang berbeda. Di antaranya adalah perbedaan kadar protein C. cephalonica (12,39%), T. molitor (16,13%), dan S. litura (6,89%) (Lampiran 1), dan kemungkinan terdapatnya zat toksik dalam tubuh mangsa yang dapat berpengaruh terhadap stadium instar nimfa dan lama hidup imago. Semakin pendek stadium instar nimfa dan semakin panjang lama hidup imago, maka mangsa tersebut sesuai atau memiliki kualitas yang tinggi dalam mendukung pertumbuhan dan perkembangan selama fase pradewasa dan reproduksi imago predator.
Mangsa C. cephalonica memberikan pengaruh yang baik terhadap predator yang dicerminkan dengan persentase mortalitas yang rendah, total stadium nimfa yang pendek dan lama hidup imago yang panjang. Sementara mangsa S. litura memberikan pengaruh yang buruk terhadap predator karena menyebabkan mortalitas yang tinggi, total stadium yang panjang dan lama hidup imago yang pendek.
mori menghasilkan stadium nimfa yang paling singkat (18,68±1,02 hari) dibanding mangsa pupa M. domestica (19,03±1,40 hari). Dalam kasus tersebut, larva B. mori lebih sesuai untuk predator P. distinctus.
De Clercq (2002) melaporkan bahwa lama stadium instar nimfa P. maculiventris yang diberi mangsa Nezara viridula Linnaeus (Hemiptera:
Pentatomidae) lebih panjang dibandingkan yang diberi mangsa S. littoralis Torr. (Lepidoptera: Noctuidae), meskipun demikian uji mortalitas menunjukkan bahwa pemberian mangsa N. viridula tersebut menghasilkan mortalitas yang rendah pada predator sehingga pemberian mangsa tersebut masih tetap direkomendasikan.
Predator S. annulicornis memiliki nilai total konsumsi yang berbeda pada ketiga jenis mangsa. Larva S. litura merupakan mangsa yang paling banyak dikonsumsi oleh predator dan sebaliknya larva C. cephalonica yang paling sedikit dikonsumsi oleh predator. Hasil penelitian mengenai total konsumsi berbanding terbalik dengan data bobot tubuh, bobot tubuh predator yang tertinggi pada predator yang diberi mangsa C. cephalonica, dan justru sebaliknya, bobot tubuh terrendah pada predator yang diberi mangsa S. litura. Data ini menunjukkan bahwa nilai total konsumsi tidak berhubungan dengan bobot tubuh. Hal ini mengandung arti bahwa predator yang memiliki nilai total konsumsi yang paling tinggi belum tentu memiliki bobot tubuh yang tinggi. Bobot tubuh predator yang tinggi sangat diharapkan karena erat kaitannya dengan kebugaran, sedangkan kebugaran untuk serangga predator dewasa menentukan produksi telur, jumlah telur, ukuran, kualitas, kemampuan dispersal dan lama hidup.
0.000 0.010 0.020 0.030 0.040 0.050 0.060
1 6 11 16 21 26 31 36 41 46 51 56 61 66 71
Hari R at a -r at a B io m as sa yan g d ih isap ( g )
Merah = mangsa status mati Biru = mangsa status hidup
C. cephalonica
1 2 3
4 5
0.000 010 020 030 040 050 060
1 6 11 16 21 26 31 36 41 46 51 56 61 66
Hari R a ta -r a ta B io m as sa ya n g di hi s a p ( g) 0.
Merah = mangsa status mati Biru = mangsa status hidup
T. molitor 0.
5
0. 4
1 2 3
0. 0. 0. 0.000 0.010 020 0.030 0.040 0.050 0.060
1 6 11 16 21 26 31 36 41 46 51 56 61 66
Hari R a ta-rat a Bi o m as sa ya n g di hi s a p ( g) S. litura
Merah = mangsa status mati Biru = mangsa status hidup
[image:45.595.124.501.114.691.2]5 4 3 2 1 0.
Pemberian mangsa C. cephalonica menunjukkan nilai total konsumsi yang rendah tetapi mendapatkan bobot tubuh yang paling tinggi. McGregor et al. (1999) melaporkan bahwa pemberian mangsa Trialeurodes vaporariorum Westwood (Homoptera: Aleyrodidae) lebih sesuai bagi predator Dicyphus hesperus Knight (Heteroptera: Miridae) dibandingkan Tetranychus urticae Koch (Acari: Tetranychidae). Hal tersebut ditunjukkan oleh stadium nimfa yang pendek (24,5±0,3 hari) dengan mangsa T. vaporariorum dan (26,3±0,4 hari) dengan mangsa T. urticae dan juga ukuran tibia yang lebih panjang pada pemberian mangsa T. vaporariorum (1,95±0,02 mm betina: 1,91±0,02 mm jantan), dan (1,84±0,01 mm betina: 1,79±0,05 mm jantan ) dengan mangsa T. urticae. Selain itu, predator juga lebih memilih tanaman tomat yang terinfestasi lebih banyak populasi T. vaporariorum dibandingkan T. urticae.
Secara umum, predator memiliki kisaran mangsa yang luas di alam namun hanya beberapa mangsa saja yang sesuai untuk pertumbuhan dan perkembangan hidupnya. Hal ini dikarenakan mangsa yang sesuai tersebut memiliki komposisi nutrisi yang cocok bagi pertumbuhan dan perkembangan hidup predator (Evans et al. 1999).
Tinggi dan rendahnya total konsumsi yang diserap oleh suatu predator pada jenis mangsa tertentu erat kaitannya dengan kualitas nutrisi yang terkandung pada tubuh mangsa. Hal ini merupakan bentuk dari respon fisiologi yang dimiliki serangga predator tersebut yaitu serangga predator akan meningkatkan konsumsi rata-rata sebagai respon apabila makanannya memiliki kandungan nutrisi yang rendah atau tidak berkualitas ( Slansky & Rodriguez 1987).
Data penelitian menunjukkan bahwa predator memiliki total konsumsi yang tinggi dengan mangsa larva S. litura tetapi menghasilkan mortalitas yang tinggi, dan sebaliknya predator memiliki total konsumsi yang rendah dengan mangsa larva C. cephalonica dan menghasilkan mortalitas yang rendah.
tersebut secara perlahan-lahan menyebabkan serangga memiliki kemampuan beradaptasi, dalam hal ini adaptasi secara fisiologi (respon fisiologi) terhadap konsumsi makanan, pemanfaatan makanan dan alokasi makanan dengan tujuan serangga tersebut mampu bertahan di alam dan memiliki kriteria kebugaran yang diharapkan. Contoh lain dari respon fisiologi serangga adalah berjemur lebih lama dan atau berkerumun membentuk kelompok pada saat terjadinya penurunan suhu ( Slansky & Rodriguez 1987).
Kualitas dari suatu predator ditentukan oleh tingkat konsumsi rata-rata suatu makanan dan tingkat efisiensi. Besarnya cairan tubuh mangsa yang diserap dan kemudian diubah menjadi komponen tubuh disebut dengan Efisiensi Penyerapan Makanan atau Efficiency of Conversion of Ingestedfood (ECI). Nilai ECI merupakan suatu parameter efisiensi penyerapan makanan oleh predator yang mampu diubah menjadi komponen tubuh. Semakin tinggi nilai ECI maka semakin efisien penyerapan kandungan nutrisi mangsa oleh predator (Slansky & Rodriguez 1987).
Nilai ECI tertinggi yaitu pada predator dengan mangsa C. cephalonica dan nilai ECI terrendah yaitu dengan mangsa S. litura. Nilai ECI yang tinggi merupakan parameter pendukung lainnya selain persentase mortalitas yang rendah, stadium nimfa yang pendek dan lama hidup imago yang panjang pada predator S. annulicornis dengan mangsa C. Cephalonica. Data-data tersebut menunjukkan bahwa dari tiga jenis mangsa yang diberikan pada predator, mangsa C. cephalonica adalah yang paling sesuai untuk predator S. annulicornis.
Pengaruh yang berbeda antara status mangsa hidup dan mati disebabkan antara lain, mangsa dalam status hidup memiliki cara mempertahankan diri dari musuhnya, diantaranya larva C. cephalonica yang menghasilkan sutera yang dapat menghambat predator. Selain itu perilaku mangsa juga berpengaruh terhadap preferensi predator. Sebagai contoh De Clercq & Degheele (1994) melaporkan bahwa predator Podisus maculiventris Say (Hemiptera: Pentatomidae) dan P. sagitta Fabricius (Hemiptera: Pentatomidae) lebih memilih memangsa Spodoptera exigua Hubner (Lepidoptera: Noctuidae) dibandingkan memangsa S. littoralis dan Mamestra brassicae Linnaeus (Lepidoptera: Noictuidae) karena S. exigua tidak agresif pada saat diserang berbeda dengan S. littoralis dan M. brassicae yang sangat agresif pada saat melakukan perlawanan. Hal ini menyebabkan predator cenderung memangsa S. exigua. Mangsa yang agresif mampu meloloskan diri dari tangkapan predator dengan melakukan perlawanan fisik sehingga dapat melarikan diri.
Predator P. maculiventaris juga lebih memilih mangsa yang relatif diam dan tidak melakukan perlawanan. De Clercq & Degheele (1994) melaporkan bahwa P. maculiventris lebih memilih memangsa pupa S. exigua. dibandingkan larva S. exigua. Predator ini juga kurang memilih imago ngengat S. litura karena ngengat ini aktif terbang.
Larva S. litura dengan status mati, membusuk, menghasilkan bau yang menyengat dan berlendir diindikasikan mengandung toksin. Penyebab terjadinya kebusukan dari suatu organisme yaitu terdapatnya mikroorganisme pembusuk, kegiatan enzim dan reaksi kimia, sedangkan pertumbuhan dan perkembangan yang berlebihan dari mikroorganisme pembusuk dapat menghasilkan toksin (Desroiser 1988).
semakin tinggi kepadatan predator memberikan pengaruh yang negatif terhadap fekunditas dan fertilitas telur yaitu terjadinya kanibalisme (Gambar 9), hal ini disebabkan karena terjadinya persaingan imago jantan untuk berkopulasi dengan betina sehingga mengganggu proses terjadinya kopulasi.
[image:49.595.179.449.212.403.2]
Gambar 9 Kanibalisme predator S. annulicornis
Hal ini didukung oleh penelitian Saharayaj (2002) yang mengungkapkan bahwa mortalitas predator Rhynocoris marginatus Fabricius (Hemiptera: Reduviidae) pada kepadatan 25-50 adalah sekitar 10%, sedangkan mortalitas pada kepadatan 75-100 adalah 32,5%. Selain itu penelitian yang dilakukan oleh De Clercq & Degheele (1994) menunjukkan bahwa mortalitas imago P. maculiventaris (Hemiptera: Pentatomidae) meningkat seiring dengan bertambahnya kepadatan populasi predator. Dengan semakin padatnya populasi maka semakin meningkat terjadinya kanibalisme antar individu predator sehingga mengganggu proses kopulasi, fertilisasi, dan perbanyakan keturunan.
menyebabkan sifat kanibalisme suatu predator (Riechert 1981 dalam Hindayana 2001). Kanibalisme merupakan salah bentuk kompetisi yang ekstrim hingga menyebabkan kematian. Terbatasnya sumber daya merupakan faktor penting terjadinya kanibalisme, dalam penelitian ini sumber daya yang terbatas adalah predator betina. Terjadinya saling membunuh antara predator jantan adalah untuk mendapatkan predator betina agar bisa meneruskan keturunannya.
Hal yang pokok dari studi pengaruh rasio betina dan jantan adalah 1) melihat rata-rata jumlah telur yang diletakkan/betina (keperidian) dan 2) melihat rata-rata jumlah telur yang menetas. Rasio 1:2 memiliki rata-rata jumlah telur yang diletakkan/betina lebih rendah dibanding dari rasio 1:1, tetapi memiliki rata-rata jumlah telur yang menetas lebih tinggi dibanding rasio 1:1. Hal ini menunjukkan bahwa rasio 1:2 adalah paling sesuai untuk predator S. annulicornis untuk mendapatkan fekunditas dan fertilitas telur yang tinggi.
Perilaku pemangsaan oleh predator S. annulicornis diawali dengan masa pencarian yaitu waktu yang dibutuhkan untuk mencari dan menemukan mangsa, kemudian masa penanganan yaitu total waktu yang diperlukan untuk menangani dan membunuh mangsa, masa pengisapan yaitu waktu yang dibutuhkan untuk mengkonsumsi satu ekor mangsa, dan jumlah mangsa yang dimangsa yaitu jumlah mangsa yang dikonsumsi per hari.
Semakin muda instar nimfa maka memiliki kemampuan yang rendah untuk menangkap mangsa (New 1991) dan sebaliknya semakin berkembangnya instar nimfa predator maka aktivitas predator yang dicerminkan dengan masa pencarian, masa penanganan, masa pengisapan adalah semakin singkat (Bellow & Fisher 1999). Hal tersebut disebabkan fotoreseptor (indera penglihatan) dan kemoreseptor (senyawa kimia) predator semakin berkembang dalam mendeteksi keberadaan mangsa.
Senyawa kimia terdiri dari dua tipe, yaitu alomon dan kairomon. Alomon adalah senyawa kimia yang menguntungkan bagi penghasil, sedangkan kairomon merupakan senyawa kimia yang merugikan penghasil. Kairomon membantu memudahkan predator untuk menemukan mangsa.
KESIMPULAN DAN SARAN
Kesimpulan
Larva C. cephalonica adalah mangsa yang paling sesuai untuk pembiakan massal predator S. annulicornis di laboratorium. Rasio 1:2 (betina: jantan) merupakan rasio yang paling sesuai untuk menghasilkan fekunditas dan fertilitas yang tinggi.
Status mangsa mati tidak berpengaruh terhadap perilaku pemangsaan keturunan pertama predator tersebut.
Pemberian mangsa C. cephalonica dengan status mati dan rasio 1:2 adalah paling sesuai untuk pembiakan massal predator S. annulicornis di laboratorium.
Saran
DAFTAR PUSTAKA
Bellows TS, Fisher FW. 1999. Biological Control: Principles and Applications of Biological Control. Academic Press.
Bosch RV, Messenger PS, Gutierrez AP. 1982. An Introduction to Biological Control. New York dan London: Plenum Press.
Cahyadi AT. 2004. Biologi Sycanus annulicornis (Hemiptera: Reduviidae) pada tiga jenis mangsa [skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor.
Chapman RF. 1998. The Insects Structure and Function. Melbourne: Cambridge University Press.
Chooyoung C, Charnnarong D, Nisit B, Chainarat S, Somchai S. 2004. Biological control of insect pest of fruit trees in the northern region of Thailand: Longan Leaf Eating Semilooper, Oxyodes scrobiculata (Fabricius) and its natural enemies. Journal of Entomological Science 7(1): 47.
Clausen CP. 1940. Entomophagous Insects. New York dan London. McGraw-Hill Book Company.
De Clercq P. 2000. Predaceous stink bugs (Pentatomidae: Asopinae). Di dalam: Schaefer CW, Panizzi AR, editor. Heteroptera of Economic Importance. Boca Raton: CRC Press.
De Clercq P, Degheele D. 1994. Laboratory measurement of predation by Podisus maculiventris and P. sagitta (Hemiptera: Pentatomidae) on beet armyworm (Lepidoptera: Noctuidae). Journal Economic Entomology 87(1): 76-83.
De Clercq P, Wyckhuys K, De Oliveira HN, Klapwijk J. 2002. Predation by Podisus maculiventris on different life stages of Nezara viridula. Florida Entomologist 85(1): 197-202.
Desroiser NW. 1988. Teknologi Pengawetan Pangan. Muchji Muljohardjo, penerjemah. Jakarta: Universitas Indonesia. Terjemahan dari: The Technology of Food Preservation.
Dixon AFG. 2000. Insect Predator-Prey Dynamics Ladybird Beetles & Biological Control. Cambridge University Press.
Hagen KS. 1987. Nutritional ecology of terrestrial insect predators. Di dalam: Slansky F, Rodriguez JG, editor. Nutritional Ecology of Insects, Mites, Spiders, and Related Invertebrates. Kanada: John Wiley & Sons. hlm 533-577.
Harahap MS, Atjung. 1969. Hama-Hama Dalam Gudang dari Bahan Tanaman Hasil Pertanian. Jakarta: Karantina Tumbuh-Tumbuhan, Departemen Pertanian.
Hindayana D. 2001. Resource exploitation by Episyrphus balteatus DeGeer (Diptera: Syrphidae) and intraguild predation [Disertasi]. Germany: Von dem Fachbereich Gartenbau der Universitat Hannover.
Kalshoven LGE. 1981. The Pests of Crops in Indonesia. Laan PA van der, penerjemah. Jakarta: Ichtiar Baru-van Hoeve. Terjemahan dari: De Plagen van de Cultuurgewassen in Indonesie.
Lacerda MC, Ferreira AM, Zanuncio TV, Zanuncio JC, Bernardino AS, Espindula MC. 2004. Development and reproduction of Podisus distinctus (Heteroptera: Pentatomidae) fed on larva of Bombyx mori (Lepidoptera: Bombycidae). Braz. J. Biol 64(2): 237-242.
Manley GV. 1982. Biology and life history of the rice field predator Andrallus spinidens F. (Hemiptera: Pentatomidae). Entomological News 93 (1): 19-24. Matheson R. 1951. Entomology for Introductory Courses. Edisi ke-2. New
York: Comstock Publishing Company.
McGregor RR, Gillespie DR, Quiring DMJ, Foisy MRJ. 1999. Potential use of Dicyphus Hesperus Knight (Heteroptera: Miridae) for biological control of pests of greenhouse tomatoes. Biological Control 16: 104-110.
Munro JW. 1966. Pests of Stored Products. London: Hutchinson of London. New TR. 1991. Insects As Predators. New South Wales: New South Wales
University Press.
Richards OW, Davies RG. 1977. IMMS’ General Textbook of Entomology. Edisi ke-10. London: Chapman & Hall.
Rosadi A. 2001. Pengaruh komposisi beberapa jenis pakan terhadap siklus hidup dan daya produksi telur Corcyra cephalonica Stainton (Lepidoptera; Pyralidae) [skripsi]. Bogor: Fakultas Pertanian, Institut Pertanian Bogor. Saharayaj K. 2002. Small scale laboratory rearing of reduviid predator,
Slansky F, Rodriguez JG. 1987. Nutritional ecology of insect, mites, spiders, and related invertebrates: an overview. Di dalam: Slansky F, Rodriguez JG, editor. Nutritional Ecology of Insects, Mites, Spiders, and Related Invertebrates. Kanada: John Wiley & Sons. hlm 1-70.
Sweet MH II 2000. Seed and Chinch Bugs (Lygaeoidea). Di dalam: Schaefer CW, Panizzi AR, editor. Heteroptera of Economic Importance. Boca Raton: CRC Press.
Taylor RJ. 1984. Predation. New York London: Chapman and Hall.
Thompson SN, Hagen KS. 1999. Nutrition of entomophagous insects and other arthropods. Di dalam: Bellows TS, Fisher TW, editor. Handbook of Biological Control Principles and Application of Biological Control. California: Academic Press. hlm 594-637.
Toerngadi A, Suroto, Manuwoto S, Utomo, Nurhayati. 1974. Penuntun Praktikum Ilmu Hama Tumbuhan Umum. Bogor: IPB.
Toft S, Wise DH. 1999. Behavioral and ecophysiological responses of a generalist predator to single- and mixed-species diets of different quality. Oecologia 119(2): 198-207.
Lampiran 1 Hasil Analisis Proksimat
Protein (%) Mangsa Status
mangsa
Kadar air
(%) B.bsh B.krg
Lampiran 2 Hasil - Uji Tukey Pengaruh Tiga Jenis Mangsa terhadap Stadium Instar Nimfa dan Lama Hidup Imago
Instar 2
Statistix 8.0 7/27/2006, 10:14:53 PM
Analysis of Variance Table for IN2
Source DF SS MS F P
P 2 455.902 227.951 35.50 0.0000 S 1 105.791 105.791 16.47 0.0001 P*S 2 289.889 144.945 22.57 0.0000 Error 111 712.809 6.422
Total 116
Note: SS are marginal (type III) sums of squares
Grand Mean 8.4720 CV 29.91
Statistix 8.0 7/27/2006, 10:31:33 PM
Tukey HSD All-Pairwise Comparisons Test of IN2 for P
P Mean Homogeneous Groups
L 11.261 A M 7.181 B C 6.975 B
Alpha 0.05 Critical Q Value 3.360
Error term used: Error, 111 DF
There are 2 groups (A and B) in which the means are not significantly different from one another
Instar 3
Statistix 8.0 7/27/2006, 10:41:14 PM
Analysis of Variance Table for IN
Source DF SS MS F P
P 2 346.501 173.251 21.50 0.0000 S 1 45.228 45.228 5.61 0.0198 P*S 2 49.652 24.826 3.08 0.0505 Error 96 773.633 8.059
Total 101
Note: SS are marginal (type III) sums of squares
Statistix 8.0 7/27/2006, 10:42:58 PM
Tukey HSD All-Pairwise Comparisons Test of IN for P
P Mean Homogeneous Groups
L 12.452 A C 8.750 B M 7.819 B
Alpha 0.05 Critical Q Value 3.367
Error term used: Error, 96 DF
There are 2 groups (A and B) in which the means are not significantly different from one another.
Instar 4
Statistix 8.0 7/27/2006, 10:53:01 PM
Analysis of Variance Table for INS
Source DF SS MS F P
P 2 8.334 4.1670 0.49 0.6163 S 1 13.676 13.6757 1.60 0.2097 P*S 2 67.004 33.5021 3.91 0.0237 Error 85 727.610 8.5601
Total 90
Note: SS are marginal (type III) sums of squares Grand Mean 12.559 CV 23.30
Tukey HSD All-Pairwise Comparisons Test of INS for P
P Mean Homogeneous Groups
L 13.000 A M 12.445 A C 12.231 A
Alpha 0.05 Critical Q Value 3.374
Error term used: Error, 85 DF
There are no significant pairwise differences among the means.
Instar 5
Statistix 8.0 7/27/2006, 11:00:22 PM
Analysis of Variance Table for INS
Source DF SS MS F P
Total 57
Note: SS are marginal (type III) sums of squares
Grand Mean 15.199 CV 26.28
Statistix 8.0 7/27/2006, 11:01:45 PM
Tukey HSD All-Pairwise Comparisons Test of INS for P
P Mean Homogeneous Groups
M 17.033 A C 15.963 A L 12.600 B
Alpha 0.05 Critical Q Value 3.413
Error term used: Error, 52 DF
There are 2 groups (A and B) in which the means are not significantly different from one another.
Imago
Statistix 8.0 7/29/2006, 2:53:45 PM
Analysis of Variance Table for IMAGO
Source DF SS MS F P
P 2 1084.64 542.321 4.02 0.0239 S 1 114.51 114.512 0.85 0.3613 P*S 2 625.92 312.959 2.32 0.1086 Error 51 6881.84 134.938
Total 56
Note: SS are marginal (type III) sums of squares Grand Mean 24.751 CV 46.93
Statistix 8.