1
PRODUKSI GULA PEREDUKSI MELALUI REKAYASA
PROSES PRA-PERLAKUAN BAMBU BETUNG
(Dendrocalamus asper (Schult. f))
WIDYA FATRIASARI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
CIPTA*
Dengan ini saya menyatakan bahwa disertasi berjudul Produksi Gula Pereduksi Melalui Rekayasa Proses Pra-perlakuan Bambu Betung (Dendrocalamus asper (Schult. f)) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, November 2014
Widya Fatriasari
RINGKASAN
WIDYA FATRIASARI. Produksi Gula Pereduksi melalui Rekayasa Proses Pra-perlakuan Bambu Betung (Dendrocalamus asper (Schult.f)). Dibimbing oleh WASRIN SYAFII, NYOMAN J WISTARA, KHASWAR SYAMSU dan BAMBANG PRASETYA.
Lignin dan struktur kristalin selulosa merupakan faktor pembatas utama pada proses konversi bahan berlignoselulosa menjadi gula pereduksi. Dalam rangka meningkatkan aksesibilitas enzim maka perlu dilakukan modifikasi struktur lignoselulosa dengan proses pra-perlakuan. Pra-perlakuan ramah lingkungan yang menarik dikembangkan diantaranya pra-perlakuan biologis dan gelombang mikro dalam medium air. Penggunaan pra-perlakuantunggal (biologis atau gelombang mikro) pada bambu betung dengan hidrolisis asam-gelombang mikro untuk memperbaiki rendemen gula hidrolisis enzimatis belum pernah dilaporkan sebelumnya. Selain itu, kombinasi pra-perlakuan secara biologis-gelombang mikro merupakan alternatif untuk memperbaiki rendemen gula, sehingga proses hidrolisisnya lebih efisien. Hal ini mengingat lebih optimalnya aktivitas degradasi lignin, pelarutan hemiselulosa, pelunakan substrat dan peningkatan porositasnya.
Bambu termasuk dalam kelompok rumput-rumputan yang berpotensi dijadikan alternatif bahan bioenergi. Beberapa hal yang mendasarinya adalah produktivitas biomasanya tinggi, mudah dan cepat diproduksi, ketersediaanya melimpah, tumbuh dengan baik pada unsur hara yang minim dan kandungan holoselulosanya tinggi. Bambu ini tumbuh tersebar terutama di Asia dan bambu di Indonesia menempati tempat ketiga setelah Tiongkok dan India. Bambu betung merupakan salah satu jenis bambu terpenting di Indonesia yang berpotensi dikembangkan. Bambu ini memiliki morfologi dan kandungan komponen kimia cukup baik untuk dikonversi menjadi bioetanol.
Pengaruh pra-perlakuan tunggal (biologis atau gelombang mikro) dan kombinasi biologis-gelombang mikro dalam mengubah struktur selulosa dan lignin bambu betung menjadi fokus dalam penelitian ini. Pengaruh perubahan karakteristik bambu setelah pra-perlakuan tersebut kemudian dihubungkan dengan kinerja hidrolisis enzimatis dan asam-gelombang mikro. Perbandingan peningkatan rendemen gula pereduksinya terhadap kontrol dan antar pra-perlakuan tunggal dan kombinasi juga didiskusikan. Selain itu juga dicoba untuk memperbaiki kinerja hidrolisis asam-gelombang mikro dengan penambahan karbon aktif.
Penelitian ini terdiri dari enam tahap penelitian yaitu penelitian tahap 1, 2, 3 untuk menganalisis perubahan karakteristik lignin dan selulosa selama pra-perlakuan biologis menggunakan jamur pelapuk putih,
informasi kinerja hidrolisis enzimatis dan asam-gelombang mikro dari kondisi pra-perlakuan biologis dan gelombang mikro terpilih serta kombinasi kedua pra-perlakuan tunggal. Parameter kunci untuk mengevaluasi kinerja hidrolisis pada penelitian tahap ini meliputi rendemen gula pereduksi, nisbah hidrolisis dan senyawa coklatnya. Pengaruh penambahan katalis karbon aktif sebagai adsorber inhibitor selama proses hidrolisis asam-gelombang mikrojuga dikaji pada tahap ini.
Hasil penelitian menunjukkan bahwa inkubasi selama 30 hari dalam pra-perlakuan biologis terpilih sebagai kondisi terbaik dengan indikator selektifitas delignifikasinya yang tinggi. Kondisi pra-perlakuan gelombang mikro terpilih adalah iradiasi selama 5, 10 dan 12 menit (330 W) serta 5 menit (770 W) dengan mempertimbangkan tingkat kehilangan berat dan kehilangan hemiselulosa. Pra-perlakuan menyebabkan struktur matriks lignoselulosa menjadi lebih terbuka, lunak serta terjadi pemisahan antar serat dan menyebabkan lebih tereksposnya selulosa. Terjadi penurunan intensitas absorbansi gugus fungsional setelah pra-perlakuan meskipun pada umumnya tidak terdapat perubahan gugus fungsi yang teridentifikasi. Hilangnya sebagian lignin setelah pra-perlakuan didukung dengan adanya penurunan intensitas gugus aromatik lignin dengan syringil lebih rendah dibandingkan dengan guaiacyl. Terjadi penurunan intensitas pada pola difraksi sinar X dan pra-perlakuan menyebabkan indeks kristalinitas bahan meningkat terkait dengan hilangnya bagian amorf yaitu lignin dan hemiselulosa dan efek hornifikasi terkait dengan kondisi pengujian sampel. Transformasi struktur kristal selulosa alomorf dari monoklinik menjadi triklinik terjadi pada semuapra-perlakuan.
Rendemen gula pereduksi hidrolisis enzimatis pada ketiga jenis pra-perlakuan dibawah 5% per bambu awal, meskipun terjadi peningkatan dibandingkan dengan kontrol. Rendemen ini dapat diperbaiki dengan hidrolisis asam-gelombang mikro. Penggunaan konsentrasi asam 1% direkomendasikan untuk digunakan dalam hidrolisis asam-gelombang mikro pada bambu. Peningkatan rendemen tertinggi terhadap kontrol dihasilkan pada proses hidrolisis gelombang mikro selama 12.5 menit dari hasil kombinasi pra-perlakuan biologis (inokulum 5%) dan gelombang mikro (5 menit, 330 W) yaitu sebesar 8.4 kali (16.65% per bambu awal). Rendemen ini lebih rendah dari rendemen gula pereduksi tertinggi dari pra-perlakuan gelombang mikro selama 12.5 menit (330 W) yaitu sebesar 25.81% per bambu awal dengan peningkatan rendemen terhadap kontrol 6.2 kali. Penambahan karbon aktif dalam hidrolisis asam-gelombang mikro hanya berpengaruh terhadap efek penghambatan pembentukan senyawa inhibitor yaitu senyawa coklat namun rendemen gula pereduksinya menurun.
SUMMARY
WIDYA FATRIASARI. Reducing Sugar Production through Pretreatment Process Engineering of Betung Bamboo (Dendrocalamus asper (Schult. f)). Under the supervision of WASRIN SYAFII, NYOMAN J WISTARA, KHASWAR SYAMSU, BAMBANG PRASETYA.
Lignin and crystalline nature of cellulose are major recalcitrance in the conversion of lignocellulosic materials to reducing sugar. Pretreatment of raw material is required to improve enzyme accessibility into the lignocellulosic structure . Many pretreatment methods have been developed including that of environmental-friendly pretreatments such as biological and microwave pretreatments in water. In general, lignocellulosic conversion into bioethanol is carried out with single pretreatment and then proceeded with enzymatic hydrolysis. However, low yield of reducing sugar frequently resulted from the method. The pretreatment sequences of biological-microwave could be useful to improve the yield of hydrolysis. Biological-microwave pretreatment was thought to bring about lignin degradation, hemicelluloses removal, substrate softening and an increase of substrate porosity. These effects will possibly improve the hydrolysis performance of lignocellulosic based materials. Bamboos belong to the graminae family and are potential alternative for bioenergy production. The potentials are indicated by the facts that bamboo retains high biomass productivity, fast and readily grown, highly available, capable of growing well in marginal land and high in hollocellulose content. Bamboos mainly grow in Asia including Indonesia. The country is ranked the 3rd after China and India in bamboo plantation. Betung bamboo is the most important species in Indonesia. The species morphologically and chemically suitable for the bioethanol production.
This research was aimed to evaluate the effect of single (biological and microwave) and combined (biological-microwave) pretreatments on lignocellulosic substrates to the structural change of the substrate cellulose and lignin. The changes were then used to explain the performance of the enzymatic and microwave assisted acid hydrolysis. The hydrolysis performance of single pretreatment, combined pretreatment and control (hydrolysis of the untreated sample) were compared. In the present works, the effect of activated carbon addition to the performance of the microwave-assisted acid hydrolysis was also studied.
performance of the enzymatic and microwave-assisted acid hydrolysis on the resulting substrate of the combined pretreatment. The hydrolysis performance was evaluated based on the yield of reducing sugar and the formed brown compound. The effect of activated carbon addition in the microwave-assisted acid hydrolysis was also discussed.
Based on its delignification selectivity, biological pretreatment for 30 days incubation was considered the best condition. The best condition of microwave pretreatment based on weight loss and hemicelluloses removal was found with irradiation power of 330 W for 5, 10, and 12 minutes, and with irradiation power of 770 W for 5 minutes. Pretreatments brought about a more exposed and open structure of cellulose and fiber disintegration. Following pretreatment, a decreasing peak intensity of FTIR spectra were found, but no change in the functional group was identified. Decreasing peak intensity was indicative to the decreasing of the lignin aromatic ring, in which the content of syringyl moieties was lower than that of guaiacyl moieties. Pretreatment increased the crystallinity of the substrate and was thought due to the removal of amorphous lignin and hemicelluloses components, as well as the occurrence of hornification due to substrate drying during X-ray examination. Both single and combined pretreatments transformed the cellulose monoclinic structure into the cellulose triclinic structure.
The reducing sugar yield of enzymatic hydrolysis of substrate from all method of pretreatment was higher than that of control, but was less than 5% of the initial substrate weight. The yield improved when microwave-assisted acid hydrolysis was applied. A 1% acid concentration was recommended in microwave-assisted acid hydrolysis of bamboo. The highest reducing sugar yield of the microwave-assisted acid hydrolysis of the combined pretreated substrate was 16.5 % of the initial weight of the substrate or 8.4 fold of its control yield. The hydrolysis was carried out with 12.5 minutes microwave irradiation of the substrate pretreated with 5% inoculum of TV proceeded with 330 W microwave irradiation power for 5 minutes. On the other hand, the highest reducing sugar yield of the microwave-assisted acid hydrolysis of the microwave pretreated substrate was 25.81% of the initial weight of substrate or 6.2 fold of its control. The substrate was irradiated for 12.5 minutes at the power of 330 W. Activated carbon addition in the microwave-assisted acid hydrolysis merely influenced the formation of inhibitor compound such as brown compound. Activated carbon addition, in fact, reduced the reducing sugar yield.
© Hak Cipta Milik IPB, Tahun 2014
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor
pada
Program Studi Teknologi Serat dan Komposit
PRODUKSI GULA PEREDUKSI MELALUI REKAYASA
PROSES PRA-PERLAKUAN BAMBU BETUNG
(Dendrocalamus asper (Schult.f))
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
Penguji Ujian Tertutup : Dr. Ir. Euis Hermiati, M.Sc Prof. Dr. Ani Suryani, DEA
NIM : E262100051
Disetujui oleh Komisi Pembimbing
Prof. Dr. Ir. Wasrin Syafii, M.Agr Ketua
Nyoman J. Wistara, Ph.D. Anggota
Prof. Dr. Ir. Khaswar Syamsu, M.Sc.St Anggota
Prof. Dr. Ir. Bambang Prasetya, M.Sc Anggota
Diketahui oleh
Ketua Program Studi
Teknologi Serat dan Komposit
Prof. Dr. Ir. Fauzi Febrianto, MS
Dekan Sekolah Pascasarjana
Dr. Ir. Dahrul Syah, M.Sc.Agr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Judul yang dipilih dalam penelitian ini ialah Produksi gula pereduksi melalui rekayasa proses pra-perlakuan bambu betung (Dendrocalamus asper (Schult f.)).
Pada kesempatan ini penulis menyampaikan rasa terima kasih kepada yang terhormat Prof. Dr. Ir. Wasrin Syafii, M.Agr selaku Ketua Komisi Pembimbing, Nyoman J. Wistara, Ph.D., Prof. Dr. Ir. Khaswar Syamsu, M.Sc.St, dan Prof. Dr. Ir. Bambang Prasetya, M.Sc. selaku Anggota Komisi Pembimbing yang telah banyak memberi saran, arahan dan dorongan kepada penulis, sehingga penulis dapat menyelesaikan penyusunan karya ilmiah ini.
Di samping itu, penghargaan penulis sampaikan kepada ketua program studi Teknologi Serat dan Komposit (TSK), Dekan Fakultas Kehutanan IPB, Sekolah Pasca Sarjana IPB yang telah banyak membantu dalam kelancaran studi S3 di IPB. Ungkapan terima kasih juga disampaikan kepada Pengelola Program Karyasiswa KNRT atas beasiswa S3 di Sekolah Pascasarjana IPB, kepada Kepala Puslit Biomaterial LIPI yang telah memberikan kemudahan untuk melakukan penelitian, serta kepada rekan-rekan Puslit Biomaterial LIPI khususnya Dr. Ir. Euis Hermiati, M.Sc., teman-teman laboratorium Teknologi Proses Biomasa dan Bioremediasi dan teman-teman di Puslit Biomaterial LIPI atas segala dukungan selama program S3. Ucapan yang sama penulis sampaikan kepada PT MTI (Multi Teknindo Infotronika) atas bantuan pengambilan gambar SEM. Ungkapan terimakasih tak lupa penulis ucapkan kepada SEAMEO BIOTROP atas bantuan pendanaan penelitian melalui program PhD Research Grant tahun 2013. Penulis juga berterimakasih atas saran dan masukan dari Dr.Ir.Euis Hermiati, M.Sc dan Prof. Dr. Ani Suryani, DEA sebagai penguji pada sidang tertutup serta Prof. Dr. Ir. Kurnia Sofyan dan Dr. Siswa Setyahadi selaku penguji sidang terbuka serta Ketua Program Studi TSK dan pimpinan sidang ujian tertutup dan terbuka. Ucapan yang sama penulis sampaikan kepada teman-teman mahasiswa S3 TSK (Teknologi Serat dan Komposit) IPB angkatan 2009, 2010, dan 2013 atas kerjasama dan dorongan semangat yang diberikannya.
Terimakasih yang tak terhingga penulis ucapkan untuk ayah (Samsuri), ibunda (Istiyah), kakak (dr Diana), suami (Subur Prayitno), anak-anakku (Arkananta, Audrey, Aretha) atas doa, pengertian, kesabaran, pengorbanan dan dukungan kepada penulis selama menempuh program S3.
Penulis menyadari bahwa karya ini tidak sempurna. Kritik dan saran sangat diharapkan. Penulis berharap semoga karya ilmiah ini bermanfaat.
Bogor, November 2014
DAFTAR ISI
DAFTAR TABEL xv
DAFTAR GAMBAR xv
1 PENDAHULUAN
1.1 Latar Belakang 1
1.2 Perumusan Masalah 3
1.3 Tujuan Penelitian 3
1.4 Manfaat Penelitian 4
1.5 Hipotesis Penelitian 4
1.6 Novelty Penelitian 5
1.7 Kerangka Pemikiran 5
1.8 Ruang Lingkup Penelitian 7
2 PENGARUH KONSENTRASI INOKULUM DAN WAKTU
INKUBASI PADA PRA-PERLAKUAN BIOLOGIS TERHADAP PERUBAHAN KARAKTERISTIK PADA BAMBU BETUNG
2.1 Pendahuluan 9
2.2 Bahan dan Metode 10
2.3 Hasil dan Pembahasan 13
2.4 Simpulan 24
3 PERUBAHAN STRUKTUR LIGNIN DAN SELULOSA BAMBU BETUNG SETELAH IRADIASI GELOMBANG MIKRO : PENGARUH LAMA IRADIASI DAN DAYA GELOMBANG MIKRO
3.1 Pendahuluan 25
3.2 Bahan dan Metode 26
3.3 Hasil dan Pembahasan 27
3.3 Simpulan 39
4 PENGARUH PRA-PERLAKUAN BIOLOGIS-GELOMBANG MIKRO PADA PADA BAMBU BETUNG TERHADAP PERUBAHAN STRUKTUR LIGNIN DAN SELULOSA
4.2 Pendahuluan 40
4.2 Bahan dan Metode 41
4.3 Hasil dan Pembahasan 42
4.4 Simpulan 54
5 KINERJA HIDROLISIS ENZIMATIS DAN ASAM-GELOMBANG MIKRO PADA BAMBU BETUNG BIOLOGIS SETELAH PRA-PERLAKUAN BIOLOGIS
5.1 Pendahuluan 55
5.2 Bahan dan Metode 56
5.3 Hasil dan Pembahasan 57
6 KINERJA HIDROLISIS ENZIMATIS DAN ASAM-GELOMBANG MIKRO PADA BAMBU BETUNG SETELAH PRA-PERLAKUAN GELOMBANG MIKRO
6.1 Pendahuluan 65
6.2 Bahan dan Metode 66
6.3 Hasil dan Pembahasan 67
6.4 Simpulan 73
7 HIDROLISIS ENZIMATIS DAN GELOMBANG MIKRO BAMBU BETUNG SETELAH KOMBINASI PRA-PERLAKUAN BIOLOGIS-GELOMBANG MIKRO
7.1 Pendahuluan 75
7.2 Bahan dan Metode 76
7.3 Hasil dan Pembahasan 77
7.4 Simpulan 83
8. PEMBAHASAN UMUM
8.1 Pengaruh Pra-perlakuan Terhadap Perubahan Komponen Kimia 85 8.2 Pengaruh Pra-perlakuan Terhadap Perubahan Karakteristik
Selulosa dan Lignin 86
8.3 Pengaruh Pra-perlakuan Terhadap Indeks Kristalinitas Bahan dan
Ukuran Kristal Selulosa 87
8.4 Pengaruh Pra-perlakuan Terhadap Pola Degradasi Selulosa 87 8.5 Pengaruh Pra-perlakuan Terhadap Perubahan Struktur Kristal
Selulosa Alomorf 88
8.6 Pengaruh Pra-perlakuan Terhadap Morfologi Serat dan Elemen
Penyusunnya 88
8.7 Pengaruh Perbedaan Metode Hidrolisis Terhadap Rendemen Gula
Pereduksi 89
8.8 Pengaruh Penambahan Karbon Aktif Terhadap Rendemen Gula Pereduksi, Senyawa Coklat dan pH Hidrolisat 91
9. SIMPULAN UMUM 92
DAFTAR PUSTAKA 93
DAFTAR TABEL
2.1 Perubahan berat elemen bambu sebelum dan setelah pra-perlakuan
biologis 18
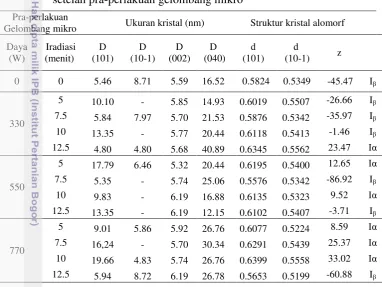
2.2 Struktur kristal selulosa alomorf pada bambu setelah pra-perlakuan
biologis 19
2.3 Nisbah intensitas antara lignin dengan karbohidrat pada bambu setelah
pra-perlakuan biologis 20
2.4 Lateral order index (LOI) dan indeks kristalinitas (CI) bambu setelah
pra-perlakuan biologis 22
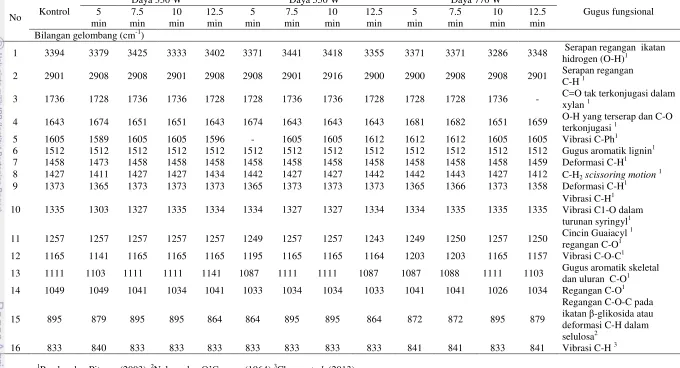
2.5 Ukuran kristal bambu setelah pra-perlakuan biologis 23 3.1 Gugus fungsional dari spektrum pita IR pada berbagai kondisi
pra-perlakuangelombang mikro pada bambu 31
3.2 Perubahan berat elemen pada bambu setelah pra-perlakuangelombang
mikro 33
3.3 Struktur kristal selulosa alomorf pada bambu setelah pra-perlakuan
gelombang mikro 34
3.4 Pola degradasi bambu setelah pra-perlakuan gelombang mikro pada
berbagai kondisi daya dan waktu iradiasi 36
3.5 Perubahan CI dan LOI pada berbagai kondisi pra-perlakuan
gelombang mikro 37
4.1 Gugus fungsional dari spektra pita IR bambu setelah pra-perlakuan
biologis-gelombang mikro 46
4.2 Perubahan berat elemen penyusun bambu setelah pra-perlakuan
biologis-gelombang mikro 48
4.3 Struktur kristal selulosa alomorf bambu setelah pra-perlakuan
biologis-gelombang mikro 49
4.4 Nisbah intensitas lignin berasosiasi dengan pita karbohidrat bambu
setelah pra-perlakuan biologis-gelombang mikro 51
4.5 CI dan LOI bambu setelah pra-perlakuanbiologis-gelombang mikro 52 4.6 Ukuran kristal bambu setelah pra-perlakuan biologis-gelombang
mikro 52
5.1 Peningkatan CI bahan dan komponen kimia setelah perlakuan jamur 59 6.1 Perbedaan rendemen gula pereduksi antara hidrolisis enzimatis dan
gelombang mikro 70
7.1 Perbandingan sifat karbon aktif sebelum dan setelah reaktivasi 81
DAFTAR GAMBAR
1.1 Kerangka pemikiran penelitian 6
1.2 Ruang lingkup penelitian 8
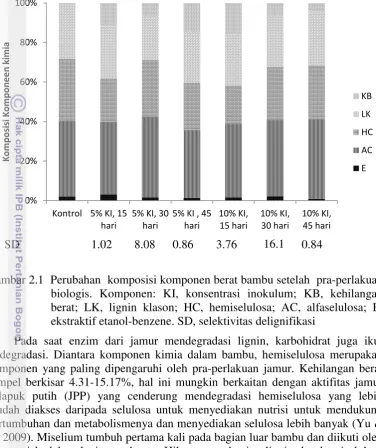
2.1 Perubahan komposisi komponen berat bambu setelah pra-perlakuan biologis
versicolor dengan inokulum 5% (A) dan (B) inokulum 10% pada
berbagai waktu inkubasi 15
2.3 Mikrograf SEM contoh setelah pra-perlakuan biologis dengan kondisi terpilih
17 2.4 Spektra FTIR pada bilangan gelombang 770-760 cm-1 (vibrasi CH2
dalam selulosa Iα) dan 710 cm-1(vibrasi CH2 dalam selulosa Iβ) 19
2.5 Spektrum FTIR bambu setelah pra-perlakuan biologis dengan TV (A) 5% dan 10% (B) pada berbagai waktu inkubasi (1) 1736 cm-1 (2) 1512 cm-1, 1373 cm-1, (4) 1165 cm-1, (5) 897 cm-1 21 2.6 Spektra XRD bambu setelah pra-perlakuan dengan TV (A) inokulum
10%) dan (B) (inokulum 5%) untuk berbagai waktu inkubasi 23 3.1 Perubahan komponen kimia bambu setelah pra-perlakuan dengan
gelombang mikro 28
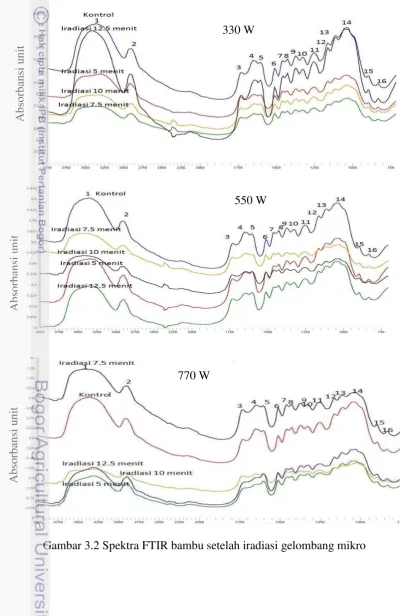
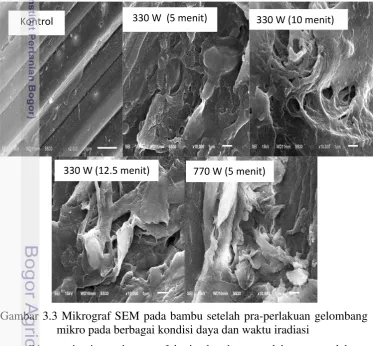
3.2 Spektra FTIR bambu setelah iradiasi gelombang mikro 30 3.3 Mikrograf SEM bambu setelah pra-perlakuan gelombang mikro pada
berbagai kondisi daya dan waktu iradiasi 32
3.4 Spektra FTIR bambu setelah pra-perlakuan gelombang mikro pada (1) 1736 cm-1 (2) 1512 cm-1, 1373 cm-1, (4) 1165 cm-1, (5) 897 cm-1 35 3.5 Spektra XRD bambu setelah pra-perlakuan gelombang mikro pada
berbagai kondisi daya dan lama iradiasi 38
4.1 Perubahan komponen kimia bambu setelah pra-perlakuan dengan
biologis-gelombang mikro 43
4.2A Spektra FTIR bambu setelah pra-perlakuan biologis (inokulum 5% inkubasi 30 hari) dilanjutkan pra-perlakuangelombang mikro 44 4.2B Spektra FTIR bambu setelah pra-perlakuan biologis (inokulum 10%
inkubasi 30 hari) dilanjutkan pra-perlakuangelombang mikro 45 4.3A Mikrograf SEM bambu setelah pra-perlakuan biologis (inokulum 10%
inkubasi 30 hari) dilanjutkan dengan pra-perlakuan gelombang mikro
pada perbesaran 10.000x 47
4.3B Gambar SEM bambu setelah pra-perlakuan biologis (inokulum 5% inkubasi 30 hari) dilanjutkan dengan pra-perlakuan gelombang mikro
pada perbesaran 10.000x 48
4.4A Spektra FTIR bambu setelah pra-perlakuan biologis (inokulum 10% inkubasi 30 hari) dilanjutkan pra-perlakuangelombang mikro 50 4.4B Spektra FTIR bambu setelah pra-perlakuan biologis inokulum (5%
inkubasi 30 hari) dilanjutkan pra-perlakuangelombang mikro 50 4.5A Spektra XRD bambu setelah pra-perlakuan biologis (inokulum 5%
inkubasi 30 hari) dilanjutkan dengan pra-perlakuangelombang mikro 53 4.5B Spektra XRD bambu setelah pra-perlakuan biologis (inokulum 10%
inkubasi 30 hari) dilanjutkan dengan pra-perlakuangelombang mikro 53 5.1 Rendemen gula pereduksi dan nisbah hidrolisis bambu setelah
pra-perlakuan pada hidrolisis enzimatis 58
5.2 Rendemen gula pereduksi dari hidrolisis asam-gelombang mikro bambu dengan pra-perlakuan biologis inokulum 5% (A) dan 10% (B) terhadap
bambu awal 60
pada inokulum 5% (A) dan 10% (B) selama hidrolisis asam-gelombang mikro
62 5.5 pH pada hidrolisat bambu setelah pra-perlakuan biologis pada inokulum
5% (A) dan 10% (B) selama hidrolisis asam-gelombang mikro 63 6.1 Rendemen gula pereduksi setelah pra-perlakuan gelombang mikro
dengan hidrolisis enzimatis, dan nisbah hidrolisis 68 6.2 Rendemen gula pereduksi berbasis bambu awal tanpa karbon aktif (A),
dan dengan karbon aktif (B) 69
6.3 Senyawa coklat yang terbentuk selama hidrolisis asam-gelombang mikro tanpa karbon aktif (A) dan dengan karbon aktif (B) 72 6.4 pH yang terbentuk selama hidrolisis asam-gelombang mikro tanpa
karbon aktif (A) dan dengan karbon aktif (B) 73 7.1 Rendemen gula pereduksi dan nisbah hidrolisis pada hidrolisis enzimatis 77 7.2 Rendemen gula pereduksi per bambu awal dari hidrolisis
asam-gelombang mikrotanpa karbon (A) dan dengan karbon aktif (B) 79 7.3 Nisbah hidrolisis per bambu awal dari hidrolisis asam-gelombang mikro
tanpa karbon aktif (A) dan dengan karbon aktif (B) 80 7.4 Senyawa coklat yang terbentuk pada hidrolisis asam-gelombang mikro
tanpa karbon aktif (A) dan dengan karbon aktif (B) 82 7.5 Perubahan nilai pH pada hidrolisis asam-gelombang mikro tanpa karbon
1
PENDAHULUAN
1.1 Latar Belakang
Ketersediaan bahan bakar fosil semakin berkurang sehingga perlu dicari alternatifnya. Bahan nabati yang telah dikonversi menjadi bahan bakar nabati (BBN) dapat menjadi substitusi bahan bakar fosil. Bioetanol merupakan salah satu jenis BBN yang berpotensi sebagai substitusi bensin. Kecenderungan penurunan cadangan minyak bumi pada periode tahun 2004-2013 merupakan fakta. Indonesia telah menjadi net importer BBM (bahan bakar minyak) sejak tahun 2004 (ESDM 2006). Produksi BBM pada tahun 2013 sebesar 825 ribu barel per hari (bpd) dengan konsumsi sebesar 825 ribu bph. Hal ini mengindikasikan terdapat defisit kebutuhan BBM sebesar 798 ribu bph (BP 2014). Berdasarkan peraturan presiden No 5 tahun 2006 tentang blue print pengelolaan energi nasional 2006-2025, penggunaan energi baru dan terbarukan (EBT) diproyeksikan sebesar 17% pada tahun 2025, dengan bahan bakar nabati menyumbang sebesar 5%. Hal ini berarti upaya pengembangan bioetanol dari bahan bakar nabati memiliki pijakan yang kuat. Bahan berlignoselulosa dari limbah pertanian, perkebunan, dan kehutanan merupakan sumber potensial untuk dikonversi menjadi bioetanol generasi kedua (Sun dan Cheng 2002). Peningkatan nilai tambah biomassa dalam konteks biorefinery didekati dengan pembentukan produk antara (intermediate) sebelum konversi berbagai polimer turunannya sebagai produk utama maupun produk sampingnya. Karbohidrat (selulosa dan hemiselulosa) merupakan sumber gula yang dapat dikonversi menjadi produk kimia yang bernilai tinggi seperti bioetanol. Konversi bahan-bahan ini menjadi bioetanol tidak mengganggu ketersediaan bahan baku pangan berkarbohidrat.
Bambu merupakan salah satu jenis lignoselulosa dari kelompok rumput-rumputan (graminae) sebagai sumber serat potensial yang memiliki pemanfaatan yang luas. Produksi pulp dan kertas, asam laktat, penguat komposit, metana, konstruksi, tekstil, makanan dan bioenergi merupakan contoh aplikasi dari pemanfaatan bambu. Mayoritas populasi bambu berada di Asia dan produksi bambu di Indonesia menduduki urutan ketiga setelah India dan Tiongkok (Lobovikov et al.2007). Produktivitas biomasa bambu adalah yang tertinggi (Kant 2010) dibandingkan dengan tanaman bioenergi lain (poplar, switch grass,
mischantus, common reed, dan bagas (Sathitsuksanoh et al. 2010; Zhang 2008). Selain itu bambu mudah dan cepat diproduksi (Scurlock et al. 2000; Gratani et al. 2008).
Berbagai pilihan jenis pra-perlakuan dapat dilakukan dengan kelebihan dan kekurangannya. Metode dan kondisi terbaik dari pra-perlakuan sangat bergantung kepada jenis lignoselulosa (Taherzadeh dan Karimi 2008). Pra-perlakuan yang efektif jika tujuannya tercapai, tidak terjadi degradasi karbohidrat dan pembentukan produk hasil samping sebagai penghambat proses hidrolisis dan fermentasi, serta efektif biayanya (Sun dan Cheng 2002; Kheswani 2009).
Pra-perlakuangelombang mikro dan biologis merupakan jenis pra-perlakuan yang relatif ramah lingkungan. Jamur pelapuk putih merupakan agen penurun kadar lignin dalam pra-perlakuan biologis (Chen et al. 2009; Zang et al. 2007) dan diharapkan mengubah kelimpahan komposisi monomer penyusun lignin sehingga laju delignifikasinya meningkat. Peningkatan porositas substrat pada pra-perlakuan gelombang mikro merupakan efek utama terhadap peningkatan aksesibilitas enzim. Untuk meningkatkan kinerja pra-perlakuan ini dapat ditambahkan asam, ataupun basa dengan efek yang berbeda terutama pada pelarutan hemiselulosa dan lignin (Kheswani 2009).
Proses pra-perlakuan biologis ini telah diaplikasikan pada berbagai bahan berlignoselulosa. Namun hasil hidrolisis enzimatis dari pra-perlakuan ini belum optimal (selektifitas delignifikasi lebih kecil dari dua) (Giles et al. 2011), sehingga perlu diperbaiki misalnya dengan pemilihan strain jamur yang tepat. Aplikasi jamur Trametes versicolor (TV) selama 45 hari pada bambu betung dalam proses biopulping memberikan delignifikasi yang lebih baik dari aplikasi
Pleorotus ostreatus (PO) dan Phenerochaete chrysosporium (PC)(Fatriasari et al. 2011; Falah et al. 2011).
Kehadiran molekul polar sangat penting dalam iradiasi gelombang mikro. Iradiasi gelombang mikromenyebabkan molekul polar bervibrasi pada kecepatan tinggi dan terjadi friksi antara molekul polar dan melingkupi medium ketika dipanaskan (Kheswani 2009). Gelombang mikro dengan selektif memanaskan
bagian yang lebih polar dan menciptakan “hot spot” dengan bahan yang tidak homogen. Pra-perlakuan gelombang mikro ini telah diaplikasikan pada
Pada konversi bahan berlignoselulosa menjadi bioetanol umumnya menggunakan pra-perlakuan tunggal dilanjutkan dengan hidrolisis enzimatik. Namun, rendemen gula yang diperoleh kurang maksimal. Penggunaan pra-perlakuantunggal (biologis atau gelombang mikro) pada bambu dengan hidrolisis berbantu gelombang mikro untuk memperbaiki rendemen gula hidrolisis enzimatis belum pernah dilaporkan sebelumnya. Selain itu, kombinasi pra-perlakuan biologis-gelombang mikro merupakan alternatif yang dapat diterapkan untuk memperbaiki rendemen gula, sehingga proses hidrolisisnya lebih efisien. Hal ini mengingat lebih optimalnya aktivitas degradasi lignin, pelarutan hemiselulosa, pelunakan substrat dan peningkatan porositasnya. Teknologi proses hidrolisis asam-gelombang mikro dengan katalis karbon aktif dapat menjadi pilihan metode untuk meningkatkan rendemen gula dan menurunkan kadar HMF, dan senyawa coklat. Sejauh ini belum ada penelitian yang melaporkan penggunaan kombinasi pra-perlakuanbiologis-gelombang mikro dengan hidrolisis asam berbantu gelombang mikro (asam-gelombang mikro) dengan atau tanpa karbon aktif ataupun hidrolisis enzimatis untuk mengkonversi bambu betung menjadi gula pereduksi.
1.2Perumusan Masalah
Permasalahan dalam penelitian ini dapat dirumuskan sebagai berikut : 1. Bagaimana perubahan karakteristik selulosa dan lignin pada bambu setelah
pra-perlakuan biologis, gelombang mikro dan kombinasi biologis-gelombang mikro?
2. Bagaimana kondisi pra-perlakuan biologis dan gelombang mikro yang tepat untuk memperbaiki kinerja hidrolisis?
3. Apakah kombinasi pra-perlakuan biologis dan gelombang mikro dapat memperbaiki kinerja hidrolisis enzimatis dan asam-gelombang mikro?
4. Bagaimana kondisi terbaik dari hidrolisis enzimatis dan asam-gelombang mikro terhadap bambu setelah pra-perlakuan biologis, gelombang mikro, dan biologis-gelombang mikro?
5. Bagaimana pengaruh penambahan karbon aktif dalam hidrolisis asam-gelombang mikro dalam menurunkan kadar penghalang yang terbentuk selama proses hidrolisis asam-gelombang mikro?
1.3 Tujuan Penelitian
Tujuan Umum
Tujuan Khusus
1. Menganalisis perubahan karakteristik bambu setelah pra-perlakuan melalui analisis komponen kimia, morfologi, elemen penyusun, dan struktur selulosa alomorf.
2. Menganalisis perubahan jumlah lignin, kemudahan pengaruh pra-perlakuan terhadap syringyl dan guiacyl serta nisbah lignin/karbohidrat setelah pra-perlakuan melalui analisis komponen kimia dan FTIR.
3. Meningkatkan rendemen gula pereduksi melalui hidrolisis asam-gelombang mikro dan menurunkan kadar senyawa coklat dalam hidrolisat dengan penambahan karbon aktif.
4. Memperoleh kondisi pra-perlakuan dengan indikator karakteristik selulosa dan lignin dan kondisi hidrolisis terbaik berdasarkan perolehan rendemen gula pereduksi dan kadar senyawa inhibitor.
1.4 Manfaat Penelitian
1. Informasi ilmiah terkait kondisi terbaik pra-perlakuan biologis, gelombang mikro dan kombinasi pra-perlakuan biologis-gelombang mikro pada bambu betung.
2. Informasi ilmiah pola perubahan karakteristik selulosa dan lignin yang terjadi pada bambu betung akibat pra-perlakuantersebut.
3. Informasi ilmiah perbandingan kinerja hidrolisis gelombang mikrodengan atau tanpa karbon aktif dan hidrolisis enzimatis.
1.5 Hipotesis Penelitian
1. Pra-perlakuan akan meningkatkan kadar relatif selulosa dan penurunan derajat kristalinitas selulosa, dan kemungkinan terjadi perubahan struktur selulosa alomorf Iα (triklinik) dari struktur selulosa Iβ (monoklinik). Terjadi penurunan
kadar kadar lignin sehingga kinerja hidrolisis lebih baik setelah pra-perlakuan.
2. Jamur pelapuk putih menyebabkan lignin terdegradasi sedangkan pra-perlakuan gelombang mikro peningkatan ketercernaan substrat akibat peningkatan porositas dan pelarutan hemiselulosa sehingga kinerja hidrolisisnya meningkat.
3. Kombinasi pra-perlakuan biologis-gelombang mikro memberikan kinerja hidrolisis yang lebih baik dibandingkan pra-perlakuan tunggal.
4. Rendemen gula pereduksi dari hidrolisis asam-gelombang mikro lebih tinggi dibandingkan dengan hidrolisis enzimatis.
1.6 Novelty Penelitian
1. Informasi terkait metode pra-perlakuan alternatif (kombinasi pra-perlakuan biologis-gelombang mikro) terhadap bambu betung untuk memperbaiki ketercernaan bahan dalam proses hidrolisis enzimatik dan asam-gelombang mikro.
2. Informasi peranan pra-perlakuan biologis, gelombang mikro, dan kombinasi pra-perlakuan biologis-gelombang mikro terhadap perubahan karakteristik bambu betung berupa penurunan kadar lignin dan hemiselulosa, peningkatan porositas serat, kerusakan serat, perubahan indeks kristalinitas bahan dan struktur selulosa alomorf.
3. Informasi peranan penambahan karbon aktif dalam hidrolisis asam-gelombang mikro untuk menurunkan kadar senyawa coklat.
1.7 Kerangka Pemikiran
Latar Belakang
Bambu potensial dimanfaatkan -Pemanfaatannya luas
-1% luas hutan dunia (Kant 2010)
-Fotosintesa efisien (C4)
-Produktivitas tertinggi (20-40 ton/ha/tahun) (Khan 2010) -Produksi selulosa tinggi
-Cepat dan mudah diproduksi (Scurlock et al. 2000; Gratani
et al. 2008)
-Delignifikasi TV pada biopulping bambu lebih baik dibandingkan PO dan PC (Fatriasari et al. 2011; Falah
et al.2011)
Metode umum pra-perlakuan:
-kimia (asam, alkali,
green solvent) -biologis
-fisik-kimia (steam explotion, afex, gelombang mikro-kimia)
-fisik
Metode umum hidrolisis:
-asam -enzimatis
Permasalahan
Struktur lignoselulosa sebagai penghalang aksesibilitas enzim
- Lignin
-Struktur kristalin selulosa
Pra-perlakuan kimia: tidak ramah lingkungan
Pra-perlakuan fisik-kimia: konsumsi energi tinggi, tidak ramah lingkungan Pra-perlakuan biologis: lama waktunya
Proses asam :
-Tidak ramah
lingkungan
-Senyawa penghalang Proses enzimatis -mahal
-lama
-rendemen gula rendah
Pemecahan Masalah
-Pemanfaatan biomassa bambu sebagai bahan baku gula
-Pemilihan metode pra-perlakuan dan hidolisis yang tepat
Rekayasa proses pra-perlakuan : kombinasi pra-perlakuan biologis dan gelombang mikro (relatif ramah lingkungan) untuk mendegradasi lignin dan hemiselulosa
Rekayasa proses hidrolisis
-iradiasi gelombang mikro medium asam -karbon aktif untuk adsorbsi penghalang dan meningkatkan rendemen gula Tujuan dan
hasil yang diharapkan
-Informasi karakteristik bahan awal dan setelah pra-perlakuan -Konsentrasi dan
rendemen gula hasil hidrolisis, senyawa coklat
-Disain kombinasi proses pra-perlakuan
-Kondisi terpilih pra-perlakuan
-Informasi perubahan karakteristik spesifik hasil pra-perlakuan lignin dan selulosa
-Informasi pengaruh karbon aktif dalam memperbaiki proses hidrolisis
-Disain proses hidrolisis terbaik
Bahan Pretreatment Hidrolisis
1.8 Ruang Lingkup Penelitian
Penelitian ini terdiri atas enam tahap yang meliputi perubahan karakteristik lignin dan selulosa setelah pra-perlakuan biologis, pra-perlakuan gelombang mikro dalam medium air, setelah kombinasi pra-perlakuan biologis-gelombang mikro, evaluasi kinerja hidrolisis enzimatis dan gelombang mikro pada pra-perlakuan biologis, setelah pra-perlakuan gelombang mikro, dan setelah kombinasi pra-perlakuan biologis-gelombang mikro.
Persiapan sampel Drum chipper
Ring Flaker Hammer mill Disk mill
Bambu 2 tahun tanpa kulit
serbuk berukuran 40-60 mesh
Karakteristik awal Pra-perlakuanbiologis
Persiapan inokulum stok
Hasil pra-perlakuan
biologis Karakteristik selulosa dan lignin
SEM-EDS, FTIR, XRD
Pra-perlakuan gelombang mikro
Persiapan sampel b
Inokulasi
Penyaringan
Residu
Pulp
Karakteristik selulosa dan lignin
SEM-EDS, FTIR, XRD Kondisi terpilih (A)
Kondisi terpilih (B)
Persiapan sampel
Iradiasi gelombang mikro
Hasil pra-perlakuanbiologis-gelombangmikro (C)
Kondisi terpilih A/B/C
Hidrolisis asam-gelombangmikro Hidrolisis enzimatis
Hidrolisis tanpa karbon aktif
Hidrolisis dengan karbon aktif
Penyaringan Hidrolisat Residu
Hidrolisat Residu Karakteristik
hasil hidrolisis
Rendemen gula, konsentrasi gula, nisbah hidrolisis, senyawa coklat
Penelitian Tahap-1, 2 dan 3
Penelitian tahap-4,5,6
2
PENGARUH KONSENTRASI INOKULUM DAN WAKTU
INKUBASI PADA PRA-PERLAKUAN BIOLOGIS
TERHADAP PERUBAHAN KARAKTERISTIK PADA
BAMBU BETUNG
2.1 Pendahuluan
Peningkatan perhatian terhadap dampak lingkungan mendorong pengembangan pendekatan pra-perlakuan yang memiliki input energi rendah, kondisi proses yang lunak, tanpa konsumsi bahan kimia dan kebutuhan prosedur dan peralatan yang sederhana (Sun dan Cheng 2002). Pra-perlakuan biologis merupakan teknik yang menjanjikan yang menggunakan mikroorganisme yang menghilangkan lignin secara efisien dari dinding sel tanaman (Hakala et al. 2005; Eriksson et al. 1990). Jamur pelapuk putih merupakan famili “basidiomycetes” yang dapat digunakan untuk pra-perlakuan biologis bahan berlignoselulosa (Sun dan Cheng 2002; Hakala et al. 2005; Eriksson et al. 1990). Jamur ini mengeluarkan enzim pendegradasi ligninolitik seperti lignin peroxidase (LiP),
laccase, mangan peroxidase (MnP) dan versatile peroxidase untuk berpenetrasi dalam dinding sel tanaman yang kemudian melakukan aktivitas degradasi bahan berlignoselulosa (Messner dan Srebotnik 1994). Proses degradasi biologis lignoselulosa merupakan proses yang kompleks dimana jenis jamur, kondisi kultur, mekanisme oksidatif dan sekresi enzimatis jamur mempengaruhi hasil yang diperoleh (Guillen et al. 2000; Wan dan Li 2010). Faktor ini dapat mempengaruhi selektifitas delignifikasi yang akan mendorong peningkatan rendemen gula dari hidrolisis enzimatis jika nilai selektifitasnya lebih besar dari dua (Giles et al. 2011). Penelitian sebelumnya melaporkan bahwa aplikasi jamur TV pada bambu betung menunjukkan kadar lignin sisa yang lebih rendah daripada aplikasi jamur Pleurotus ostreatus (PO) dan Phanerochaete chrysosporium (PC) (Fatriasari et al. 2011; Falah et al. 2011).
Bambu merupakan tanaman yang termasuk dalam kelompok rumput-rumputan yang memiliki kemampuan fotosintesa yang efisien yang ditunjukkan oleh produktivitas biomasa yang tinggi (20-40 ton/ha/tahun). Lebih lanjut, dibandingkan dengan bahan sumber bioenergi lain (poplar, switchgrass, miscanthus, common reed, dan bagas tebu) (Shathitsuksanoh et al. 2010; Zhang 2008), produksi biomasa bambu tergolong unggul. Tanaman bambu ini meliputi 1% luas hutan dunia (Kant 2010) dan 65% dari populasinya terdapat di Asia. Produksi bambu Indonesia menduduki rangking ketiga (5%) setelah India (30%) dan China (14%) (Lobovikov et al. 2007). Bambu mudah dan cepat diproduksi (Scurlock et al. 2000; Gratani et al. 2008). Bambu betung merupakan jenis bambu yang sangat penting di Indonesia (Dransfield dan Widjaja 1995) dan merupakan kultivar endemik Indonesia.
substrat diberikan perlakuan waktu inkubasi dan konsentrasi inokulum TV karena perlakuan ini kemungkinan dapat mempengaruhi kinerja hidrolisis.
2.2 Bahan dan Metode
2.2.1 Bahan
Bambu betung (Dendrocalamus asper) segar tanpa kulit yang berumur kurang lebih 2 tahun dari kebun bambu Puslit Biomaterial LIPI, Cibinong, Indonesia digunakan dalam penelitian ini. Bambu tersebut dikuliti, dicacah dan dibuat serpih dengan drum chipper, ring flaker, hammermill dan selanjutnya dibuat serbuk dengan disk mill dan disaring untuk memperoleh ukuran 40-60 mesh. Serbuk ini selanjutnya akan digunakan dalam proses pra-perlakuan.
2.2.2 Metode
2.2.2.1 Pra-perlakuan Biologis
Serbuk bambu (40-60 mesh) diberi air dengan perbandingan 1:4 sambil diaduk hingga rata. Serbuk bambu yang telah basah kemudian dimasukkan ke dalam botol selai dan dikukus selama 30 menit pada suhu ± 100 °C. Setelah dingin, botol selai yang berisi serbuk yang telah dikukus tersebut disterilisasi dengan autoclaf selama 20 menit pada suhu 1210C dengan tekanan 1 atm.
Biakan jamur pelapuk putih, TV yang diperoleh dari Puslit Kimia LIPI Serpong dikultur pada medium slant Malt Extract Agar (MEA) (8.875 g MEA dilarutkan ke dalam 250 ml aquades selama 7-14 hari. Sebanyak 5 ml medium JIS
(Japan Industrial Standard) Broth dimasukkan ke dalam setiap slant, jamur kemudian dirontokkan dengan ose. Suspensi tersebut kemudian dituangkan ke dalam 95 ml medium JIS Broth (dalam 1 L aquades ditambahkan 3 g KH2PO4, 2 g
MgSO4.7H2O, 25 g glukosa, 5 g pepton, dan 10 g malt extract) dan diinkubasi
selama 7-8 hari dalam kondisi stasioner. Sebanyak 10 g corn steep liquor (CSL) dituangkan ke dalam 100 ml inokulum, kemudian dihomogenkan dengan waring blender model HGB TWT (tegangan 240 VAC, frekuensi 50-60 Hz, arus 3.6 A) pada kecepatan tinggi (22.000 RPM) selama dua kali 20 detik. Campuran homogen ini digunakan sebagai stok inokulum.
Serbuk bambu sebanyak 15 g yang telah dikukus dan disterilisasi, kemudian didinginkan dan media siap diinokulasi dengan variasi inokulum sebesar 5 dan 10% (w/v). Media bambu kemudian diinkubasi pada suhu 270C dalam inkubator selama 15, 30 dan 45 hari.
2.2.2.2 Perubahan Karakteristik Selulosa dan Lignin
2.2.2.2.1 Penentuan Komponen Kimia
Sampel hasil holoselulosa selanjutnya dianalisis kadar alfa selulosa dengan Mokushitsu Kagaku Jiken Manual (2000). Hemiselulosa diperoleh sebagai hasil pengurangan holoselulosa dengan alfa selulosa. Penentuan kehilangan berat mengacu pada Pandey dan Pitman (2003), sedangkan selektifitas delignifikasi dihitung sebagai nisbah kehilangan lignin terhadap kehilangan selulosa (Yu et al. 2009). Komposisi komponen kimia dihitung berdasarkan persamaan 2.1, sedangkan kehilangan berat komponen kimia dihitung dengan memperhatikan kehilangan berat setelah pra-perlakuan sebagai faktor pengurang (persamaan 2.2). Komponen kimia akhir (%) =
(2.1)
Kehilangan komponen kimia (%) =
(2.2)
2.2.2.2.2 Perubahan Struktur Selulosa dan Lignin
Perubahan struktur selulosa dan lignin bambu diobservasi melalui analisis gugus fungsional dari spektra infra merah yang diambil dengan Fourier Transform Infrared Spectrometry (FTIR) ABB MB 300. Serbuk bambu kering sebanyak 4 mg dicampur dengan 200 mg KBr (Kalium Bromida) grade spektroskopi, dan dibentuk pellet dengan tekanan 6 ton sehingga menjadi pelet berdiameter sekitar 1.3 cm dan tebal sekitar 0.5 cm. Pola spektrum inframerahnya dianalisis pada 5 pemindaian dengan resolusi 16 cm-1. Spektrum dicatat dalam modus absorpsi dalam kisaran 3500-700 cm-1.
2.2.2.2.3 Penentuan Indeks Kristalinitas Bahan
Sampel berukuran 40-60 mesh diletakkan di atas pegangan gelas sampel dan dianalisis di bawah kondisi ruang X-ray diffraction (XRD) direkam dengan seri Shimadzu XRD-700 MaximaX. Radiasi NI disaring dengan CuKα pada bilangan gelombang 0.1542 nm. Sinar X dioperasikan pada voltase 40 kV dan 30 mA. Scan
sudut 2Ɵ sebesar10-40o setiap 2o per menit.
Penghitungan indeks kristalinitas (CI) bahan berdasarkan formula Zao et al.(2005) dan Foscher et al.(2001) yang dihitung dari data intensitas difraksi dengan rumus berikut:
CI=
(2.3)
Dimana Fc merupakan daerah kristalin, dan Fa merupakan daerah non kristalin/amorf. Intensitas total dihitung menggunakan software yang terdapat dalam Shimadzu MaximaX diffractometer.
2.2.2.2.4 Penentuan Struktur Kristal Selulosa Alomorf
Penentuan fungsi Z-diskriminan mengikuti formula yang dikembangkan oleh Wada et al.(2001) untuk membedakan struktur kristalin selulosa (monoklinik dan triklinik). Fungsi Z-diskriminan digunakan dengan memisahkan selulosa Iα
equator jarak d: 0.59-0.62 nm (d1) dan 0.52-0.55 nm (d2). Formula untuk
menentukan fungsi Z-diskriminan tersebut adalah:
Z =1693 d1 (nm)-902 d2 (nm)-549 (2.4)
Dimana Z>0; mengindikasikan tipe bakteri alga (Iα, kaya struktur triklinik) dan Z <0 mengindikasikan tipe kapas-rami (didominasi struktur Iβ/monoklinik) (Wada et al. 2001).
2.2.2.2.5 Ukuran Kristal Selulosa
Penentuan ukuran kristal ditentukan menggunakan pola difraksi yang diperoleh dari ketiga bidang kisi (101, 10-1 dan 002) dan (040) dengan formula sebagai berikut.
D(hkl) (2.5)
Dimana D(hkl) merupakan ukuran kristalin rata-rata, k merupakan konstanta Scherrer (0.84), λ merupakan panjang gelombang X-ray (1.5420A), B(hkl)
merupakan FWHM (full width half maximum) dari refleksi hkl yang diukur dari
2θ (theta) sesuai dengan sudut Bragg (17.250) (Oh et al.2005; Ahtee et al.1988). 2.2.2.2.6 Analisis Morfologi (SEM-EDS)
Morfologi sampel bambu sebelum dan setelah pra-perlakuan biologis dengan pra-perlakuan terpilih dikarakterisasi menggunakan SEM (Scanning Electron Microscope) Bruker 133 eV dengan lensa Carl Zeiss. Sampel bambu dipasang pada tempat sampel (stub) menggunakan spuntter canter dan kemudian dipindai pada tegangan 5 kV. Untuk mengetahui jenis dan persentase relatif kandungan unsur pada sampel digunakan Electron Dispersive Spectroscopy (EDS) dengan kondisi pengujian sama dengan SEM. Hamburan elektron contoh uji berupa sinar-X ditangkap oleh detektor EDS dan dipetakan dalam bentuk unsur berdasarkan perbedaan energi yang diterima.
2.2.2.2.7 Pola Biodegradasi dengan Analisis FTIR
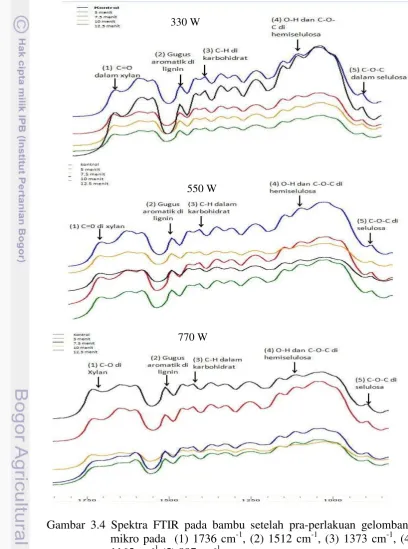
Analisis pola degradasi bambu setelah pra-perlakuan diamati dengan menggunakan spektrum dari FTIR ABB MB 300 pada bilangan gelombang 800-400 cm-1, resolusi 16 cm-1 dan 5 pemindaian. Tinggi puncak dan luas area ditentukan dengan software Horizon MB dan perubahan relatif dalam intensitas pita karakteristik karbohidrat dan lignin dapat dianalisis dengan metode Pandey (Pandey dan Pitman 2003). Intensitas puncak lignin muncul pada kisaran bilangan gelombang 1512 cm-1 berasal dari kerangka aromatis (aromatic skeletal) dalam lignin. Perubahan relatif dalam intensitas puncak lignin pada 1512 cm-1 terhadap puncak karbohidrat pada 1736 (C=0 dalam xylan, 1373 (C-H dalam selulosa dan lignin), 1165 (C-O-C) dalam selulosa dan hemiselulosa dan 897 (C-H dalam selulosa) cm-1 menggunakan analisis dengan software Horizon MB.
2.2.3 Analisis Data
2.3 Hasil dan Pembahasan
2.3.1 Perubahan Komponen Kimia
Pra-perlakuan jamur yang diberikan dalam penelitian ini mengubah komposisi komponen kimia bambu betung (Gambar 2.1). Bahan ini memiliki holoselulosa (selulosa dan hemiselulosa) yang tinggi. Fraksi ini merupakan polimer gula dan merupakan sumber gula potensial untuk memproduksi etanol. Secara teori, potensi rendemen gula yang bisa dihasilkan dari bambu sebesar 77.28-73.34%, dengan faktor konversi 1.111. Secara alami, lignoselulosa tahan terhadap degradasi dan stabil terhadap hidrolisis, terutama dikarenakan ikatan silang lignin melalui ikatan ester dan eter (Verma et al. 2011). Lignin merupakan komponen utama dinding sel tanaman yang dapat menghambat proses hidrolisis. Pra-perlakuan biologis dengan jamur pelapuk putih dapat digunakan untuk memfasilitasi enzim hidrolitik untuk mengakses struktur kristalin selulosa dengan pemecahan ikatan kompleks selulosa-lignin melalui ekstraksi atau dekomposisi dari lignin (Zadrazil et al.1999).
Pra-perlakuan biologis juga menyebabkan kehilangan berat sampel (Gambar 2.1), sedangkan penambahan waktu inkubasi pada inokulum 10% cenderung menurunkan kehilangan berat. Hal ini kemungkinan terkait dengan kehilangan lignin yang lebih besar dari kehilangan karbohidrat atau jamur lebih selektif mendegradasi lignin dibandingkan dengan karbohidrat. Selektifitas delignifikasi pra-perlakuan TV pada tingkat inokulum 5 dan 10% selama 30 hari lebih baik dari selektifitas delignifikasi pra-perlakuan lain. Jamur memiliki kemampuan mendegradasi lignin secara lebih selektif yang ditunjukkan oleh peningkatan signifikan dalam nilai selektifitas delignifikasi dengan meningkatnya waktu inkubasi sampai 30 hari. Selektivitas delignifikasi bambu akibat pra-perlakuan menurun setelah inkubasi 30 hari. Hasil ini mengindikasikan bahwa TV mendegradasi lignin hanya pada tahap awal pra-perlakuan dan menjadi tidak selektif dengan bertambahnya waktu perlakuan. Hasil ini sejalan dengan pra-perlakuan TV pada kayu karet dan pra-pra-perlakuan kultur campur TV dan C. subvermisphora (Nazarpour et al. 2013) serta pra-perlakuan Ganoderma austrael
Gambar 2.1 Perubahan komposisi komponen berat bambu setelah pra-perlakuan biologis. Komponen: KI, konsentrasi inokulum; KB, kehilangan berat; LK, lignin klason; HC, hemiselulosa; AC, alfaselulosa; E, ekstraktif etanol-benzene. SD, selektivitas delignifikasi
Pada saat enzim dari jamur mendegradasi lignin, karbohidrat juga ikut terdegradasi. Diantara komponen kimia dalam bambu, hemiselulosa merupakan komponen yang paling dipengaruhi oleh pra-perlakuan jamur. Kehilangan berat sampel berkisar 4.31-15.17%, hal ini mungkin berkaitan dengan aktifitas jamur pelapuk putih (JPP) yang cenderung mendegradasi hemiselulosa yang lebih mudah diakses daripada selulosa untuk menyediakan nutrisi untuk mendukung pertumbuhan dan metabolismenya dan menyediakan selulosa lebih banyak (Yu et al. 2009). Miselium tumbuh pertama kali pada bagian luar bambu dan diikuti oleh penetrasi kedalam lapisan substrat. Hilangnya sebagian lignin dan hemiselulosa merusak komplek karbohidrat-lignin yang menyebabkan rusaknya ikatan hidrogen dalam selulosa (Li et al. 2010), aktivitas ini menyebabkan kehilangan berat setelah pra-perlakuan.
2.3.2 Perubahan Struktur Selulosa pada Bambu
Spektroskopi FTIR digunakan sebagai alat analitik kualitatif untuk menentukan perubahan kimia di permukaan bambu untuk melengkapi dan memahami hasil investigasi dengan XRD. Gambar 2.2 menunjukkan spektrum inframerah (IR) bambu setelah pra-perlakuan pada daerah bilangan gelombang 800-4000 cm-1.
0% 20% 40% 60% 80% 100%
Kontrol 5% KI, 15 hari
5% KI, 30 hari
5% KI , 45 hari
10% KI, 15 hari
10% KI, 30 hari
10% KI, 45 hari
K
o
m
p
o
si
si
Kom
p
o
n
e
e
n
ki
m
ia
KB
LK
HC
AC
E
SD 1.02 8.08 0.86 3.76 16.1
9
[image:33.595.65.513.76.506.2]
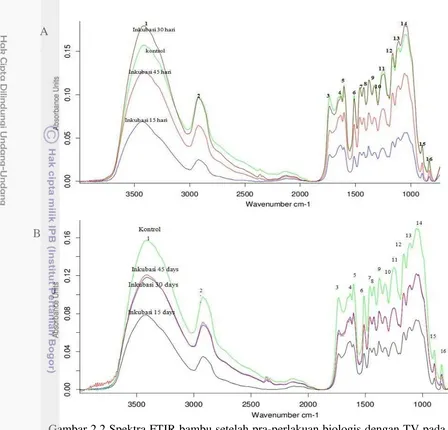
Gambar 2.2 Spektra FTIR bambu setelah pra-perlakuan biologis dengan TV pada tingkat inokulum 5% (A) dan inokulum 10% (B) yang diinkubasi bervariasi selama 15, 30, dan 45 hari
Perlakuan jamur pada bambu tidak menyebabkan perubahan gugus fungsional, tetapi hanya menyebabkan perubahan intensitas serapan. Ikatan hidrogen (O-H) uluran yang kuat dan lebar terlihat pada 3420 cm-1 (1) dan serapan uluran C-H tampak pada bilangan gelombang sekitar 2920 cm-1 (2). Pada daerah finger print antara 1750-800 cm-1, beberapa puncak ditemukan. Puncak-puncak di daerah ini adalah 1736 cm-1 (3) untuk C=O yang tidak terkonjugasi dalam xylan (hemiselulosa), 1635 cm-1 (4) untuk O-H yang diabsorbsi dan C-O terkonjugasi, kedua bilangan gelombang 1605 cm-1 (5) dan 1512 cm-1 (6) untuk gugus aromatik skeletal di lignin, 1462 cm-1 (7) untuk deformasi C-H di lignin dan selulosa, 1426 cm-1 (8) untuk C-H2 scissoring motion di selulosa, 1378 cm-1
(9) untuk deformasi C-H di selulosa dan hemiselulosa, 1328 cm-1 (10) untuk vibrasi C-H di selulosa dan vibrasi C1-O di turunan syringyl, 1246 cm-1 (11) untuk
cincin guaiacyl dan uluran C-O di lignin dan xylan, 1164 cm-1 (12) untuk vibrasi C-O-C di selulosa dan hemiselulosa, 1110 cm-1 (13) untuk gugus aromatik A
skeletal dan uluran C-O, 1053 cm-1 (14) untuk regangan C-O di selulosa dan hemiselulosa, pita tajam pada 897 cm-1 (15) untuk uluran C-O-C pada
karakteristik ikatan β glikosida pada selulosa (Li et al. 2010) atau deformasi C-H di selulosa (Pandey dan Pitman 2003), dan 833 cm-1 (16) untuk vibrasi C-H di lignin (Cheng et al. 2013).
Gugus O-H pada daerah bilangan gelombang 3000-3600 cm-1 pada spektrum bambu identik daerah regangan O-H region dari selulosa I (Gambar 2.2). Vibrasi ikatan hidrogen dari gugus OH pada selulosa I merupakan gabungan tiga ikatan hidrogen yang berbeda yaitu ikatan hidrogen intramolekul 2-OH...O-6, ikatan hidrogen intramolekul 3-OH....O-5 dan ikatan hidrogen intramolekul 6-OH....O-3 (Oh et al. 2005). Absorbansi pada bilangan gelombang ini menurun sepanjang waktu inkubasi, mengindikasikan penurunan ikatan hidrogen. Pita pada 1635-1640 cm-1, yang dihubungkan dengan vibrasi tekukan molekul air yang diserap, juga menurun setelah perlakuan jamur. Penemuan ini mirip dengan pra-perlakuandengan larutan ionik pada Asclepias syriaca dan kayu poplar (Spiridon
et al. 2010). Intensitas relatif semua pita pada seluruh pra-perlakuan bambu pada inokulum 10% lebih rendah daripada inokulum 5% karena aktivitas degradasi jamur pelapuk putih. Pita spektrum IR pada 1378 cm-1, 1735 cm-1, 1164 cm-1 dan 897 cm-1 sebagai gugus fungsional selulosa dan hemiselulosa menunjukkan penurunan intensitas dibandingkan dengan kontrol. Hal ini mengindikasikan bahwa pra-perlakuan juga menghilangkan hemiselulosa dan selulosa pada aktivitas degradasi lignin oleh jamur.
Puncak pita lignin dari sampel setelah pra-perlakuanpada 1512 cm-1 (cincin aromatik) (Pandey dan Pitman 2003) cenderung menurun. Hal ini terkait dengan pelepasan lignin dari matriks lignoselulosa. Fakta ini sejalan dengan penelitian sebelumnya oleh Nazarpour et al. (2013) yang mengindikasikan intensitas puncak lignin menurun signifikan dibandingkan dengan karbohidrat. Spektrum IR dari tandan kosong kelapa sawit (TKKS) dengan pra-perlakuan jamur, P. florindanus
mengindikasikan penyerangan ikatan lignin dan hemiselulosa dari jamur ini (Faix
1991). Perubahan ini terkait dengan terpisahnya antara atom karbon benzylic α-
aktifitas yang lebih selektif ketika sampel dipaparkan selama 30 dan 45 hari. Jumlah relatif selulosa kristalin dan amorf telah didiskripsikan sebelumnya oleh Wistara et al. (1999) menggunakan nisbah puncak inframerah.
Lateral order index (LOI) yang merupakan nisbah antara absorbansi pada bilangan gelombang 1420 dan 893 cm-1 juga dapat digunakan untuk menghitung CI dari selulosa I karena sensitifitas jumlah struktur kristalin terhadap amorf dalam selulosa (Oh et al. 2005). LOI menunjukkan korelasi linier dengan kadar hemiselulosa karena pita pada 893 cm-1 yang diidentikkan dengan frekuensi gugus karbon pada selulosa dan hemiselulosa (Ramos 2003). Dalam penelitian ini, pasangan pita puncak pada bilangan gelombang 1426 cm-1 (kristalin) dan 833 cm
-1
(amorf) tampak jelas pada sampel (Gambar 2.2).
Pola spektrum FTIR bambu dengan inokulum 5 dan 10% menunjukkan kecenderungan yang sama kecuali inokulum 5% selama 30 hari. Penurunan kadar xylan (1736 cm-1) dari pra-perlakuan selama 30 hari lebih rendah dibandingkan dengan sampel yang diinkubasi selama 15 dan 45 hari (Gambar 2.2). Intensitas puncak pada bilangan gelombang 1512 cm-1 (gugus aromatik pada lignin) untuk inkubasi 30 hari tampak yang tertinggi. Hal ini berarti jamur ini lebih bersifat menyerang lignin daripada hemiselulosa.
2.3.3 Karakteristik Morfologi Bambu
SEM digunakan untuk mengobservasi secara visual perubahan morfologi struktur pada permukaan bambu setelah pra-perlakuan biologis. Sampel yang diinkubasi selama 30 hari dipilih untuk observasi karena selektifitas delignifikasinya lebih baik daripada pra-perlakuan lain. Seperti tampak pada Gambar 2.3, perlakuan jamur TV menyebabkan kerusakan serat bambu. Bambu tanpa perlakuan menunjukkan struktur serat yang kaku dan kompak sedangkan bambu setelah pra-perlakuan mengalami kerusakan serat dan memiliki stuktur yang terpisah. Kerusakan pada matriks dinding sel ini meliputi pemutusan ikatan antara karbohidrat dan lignin, depolimerisasi lignin dan terlarutnya polimer hemiselulosa (Ramos 2003).
Gambar 2.3 Mikrograf SEM bahan dengan pra-perlakuan biologis pada kondisi terpilih.
Kontrol 5% inokulum
Degradasi lignin disebabkan oleh pra-perlakuan jamur dapat meningkatkan ukuran pori dalam substrat dan menyediakan daerah permukaan yang lebih mudah diakses oleh selulase. Perlakuan dengan jamur pada TKKS telah terbukti merusak permukaan serat akibat menghilangnya bagian silika yang meninggalkan ruang kosong sehingga memudahkan penetrasi miselium ke dalam lapisan sampel (Isroi
et al. 2010). Pada serat jerami, luas permukaan serat meningkat akibat degradasi oleh Pleurotus ostreatus (Taniguchi et al. 2010). Degradasi lignin berpotensi meningkatkan ketercernaan selulosa (Nazarphour et al.2013). Sebagian serat terpisah oleh pra-perlakuan biologis, diasumsikan karena terputusnya beberapa ikatan eter dalam lignin dan ikatan lignin-karbohidrat. Tidak ada perbedaan signifikan pada tingkat kerusakan antara konsentrasi inokulum 5 dan 10% selama 30 hari. Observasi lanjut pada tingkat ultrastruktur diperlukan untuk mempelajari perubahan dinding sel akibat degradasi lignin oleh aktifitas jamur. Degradasi dinding sel mungkin terjadi dan mekanisme dari proses ini dinyatakan oleh Akhtar et al. (1998). Jamur mungkin berkolonisasi dalam lapisan substrat dan mengambil nutrisi untuk mengembangkan jaringan dan mendistribusikan miseliumnya.
Jumlah elemen yang ada dan jenisnya direkapitulasi pada Tabel 2.1 berdasarkan analisis EDS. Fluorin dan silikon tidak ditemukan setelah sampel diberikan perlakuan jamur, sedangkan elemen lain masih ada dengan tingkat yang berbeda-beda. Karbon dan oksigen adalah elemen dominan kemungkinankarena bambu adalah polimer lignoselulosa dengan komponen utama karbon, hidrogen dan oksigen. Dalam jumlah sedikit, magnesium dan aluminium masih ditemukan dalam sampel setelah pra-perlakuan. Pra-perlakuan jamur menurunkan kadar karbon, karena karbon digunakan oleh jamur sebagai nutrisi untuk mendukung pertumbuhannya.
Tabel 2.1 Perubahan berat elemen bambu sebelum dan setelah pra-perlakuan biologis
Elemen Kontrol % b/v Inokulum 5 % b/v
Inokulum 10% b/v
Karbon (C) 44.65±13.7 42.30±13.0 43.05±13.2
Oksigen (O) 44.71±13.8 55.72±17.2 55.30±17.7
Fluorin (F) 9.05±3.2 - -
Magnesium (Mg) 0.84±0.1 1.11±0.1 0.96±0.1
Aluminium (Al) 1.11±0.1 0.87±0.1 0.69±0.1
Silikon (Si) 0.87±0.1 - -
Total 100 100 100
2.3.4 Struktur Kristal Selulosa Alomorf
Tabel 2.2 Struktur kristal selulosa alomorf dari bambu setelah pra-perlakuan biologis
Inokulum (%)
Inkubasi (hari)
Struktur kristal alomorf Jenis
kristal alomorf
d (101) nm d (10-1) nm Z
0 45 0.566041 0.534862 -73.13 Iβ
5
15 0.588426 0.522358 -23.96 Iβ
30 0.615201 0.521747 21.92 Iα
45 0.615201 0.546641 -0.53 Iβ
10
15 0.588426 0.566041 -63.36 Iβ
30 0.616903 0.520533 25.89 Iα
45 0.602733 0.535504 -11.59 Iβ
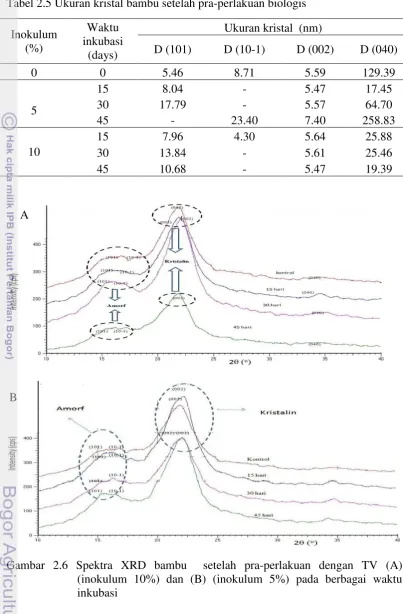
Selulosa monoklinik (Iβ) secara termodinamika lebih stabil dibandingkan dengan triklinik (Iα), karena tidak hanya padat, tetapi cenderung menjadi produk akhir dalam pemanasan dari semua selulosa (Sassi et al. 2000). Struktur monoklinik dari selulosa berubah menjadi struktur triklinik pada pra-perlakuan jamur selama 30 hari. Penentuan Z-diskriminan dilakukan berbasis pada bidang (101) and (10-1) dari sampel bambu. Struktur triklinik diharapkan beprengaruh lebih baik terhadap kinerja hidrolisis karena fase Iα lebih mudah terdegradasi daripada Iβ
(Wada dan Okano 2001). Selulosa Iα bersifat meta-stabil dan lebih reaktif daripada Iβ (O’Sullivan 1997). Selain itu, selulase Tricoderma viride lebih memilih mendegradasi selulosa Iα dibandingkan selulosa Iβ dan dengan bertambahnya waktu, mikro kristalin yang tertinggal menjadi kaya Iβ (Hayashi et al.1998a,b). Selulosa fase Iα memiliki reaktivitas yang lebih tinggi daripada fase
Iβ (Sassi et al. 2000) dan fase ini lebih banyak pada permukaan mikro kristal (Hayashi et al. 1998a,b).
Gambar 2.4 Spektra FTIR pada bilangan gelombang 750-760 cm-1 (vibrasi CH2
dalam selulosa Iα) dan 710 cm-1(vibrasi CH2 dalam selulosa Iβ)
Puncak pada bilangan gelombang sekitar 750-760 cm-1 dan 710 cm-1 diidentifikasi sebagai vibrasi CH2 dalam selulosa Iα dan selulosa Iβ. Seperti
cm-1 dan 710 cm-1. Perubahan sruktur kristal selulosa alomorf hanya tampak berdasarkan analisis XRD, tetapi tidak tampak pada analisis FTIR.
2.3.5 Pola Biodegradasi
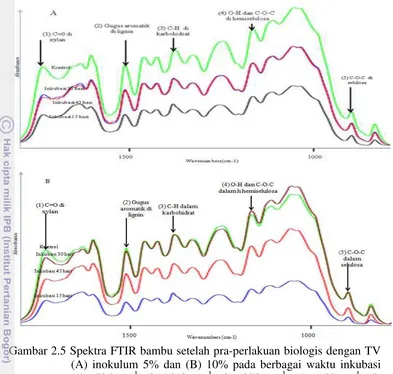
Pola biodegradasi bambu selama pra-perlakuan biologis dengan TV dievaluasi dengan analisis FTIR berdasarkan analisis metoda Pandey (Pandey dan Pitman 2003). Tabel 2.3 menunjukkan perubahan relatif dalam intensitas puncak lignin pada bilangan gelombang 1512 cm-1 terhadap puncak karbohidrat pada bilangan gelombang 1736, 1373, 1165, 897 cm-1 yang dihitung dari tinggi puncak dan luasnya. Empat puncak karbohidrat yang teridentifikasi tidak terkonjugasi dan gugus aromatik skeletal dalam lignin ditunjukkan pada Gambar 2.5.
Empat puncak karbohidrat untuk ikatan tidak terkonjugasi yaitu C=O dalam xylan, deformasi C-H dalam hemiselulosa, vibrasi C-O-C dalam selulosa dan hemiselulosa dan deformasi C-H atau uluran C-O-C pada ikatan β glikosida yang merupakan karakteristik dalam selulosa (Pandey dan Pitman 2003).
Tabel 2.3 Nisbah intensitas antara lignin dan karbohidrat pada bambu setelah pra-perlakuanbiologis.
Inokulum (%)
Inkubasi
(hari) Intensitas relatif
a
dari vibrasi gugus aromatik (I1512) terhadap tipe pita dari karbohidrat
I1512/I1736 I1512/I1373 I1512/I1165 I1512/I897
0 45 1.02(0.91) 1.05(1.15) 1.02(1.20) 1.33(1.25) 5
15 1.14(1.02) 0.89(0.98) 0.80(0.94) 8.00(1.25) 30 1.25(1.03) 1.00(1.05) 0.83(1.00) 5.00(1.29) 45 1.42(1.06) 1.11(1.00) 1.00(1.03) 1.43(1.24) 10
15 1.04(1.08) 1.02(1.08) 0.98(0.96) 1.28(1.3) 30 1.67(1.08) 1.67(1.00) 0.83(0.96) 5.00(1.29) 45 0.97(0.26) 0.78(1.00) 0.70(0.97) 7.00(1.25)
a
Intensitas relatif dihitung menggunakan tinggi puncak (diluar tanda kurung) dan luas (dalam kurung)
Umumnya terdapat peningkatan nisbah lignin/karbohidrat dari bambu dengan pra-perlakuan TV dengan bertambahnya waktu inkubasi, yang mengindikasikan jamur kurang selektif mendegradasi lignin. Studi sebelumnya oleh Zhang et al. (2007) pada bambu dan Ferraz et al. (2000) pada Pinus radiata melaporkan bahwa TV G20 dan Gonoderma australe
lebih bersifat mendegradasi lignin hanya pada tahap awal dan sektifitasnya menurun dengan bertambahnya waktu inkubasi. Namun, pra-perlakuan bambu dengan E.taxodii 2538 cenderung menurun nisbah ini secara signifikan dengan bertambahnya waktu. Hal ini mengindikasikan bahwa
E.taxodii 2538 lebih selektif mendegradasi lignin. Dalam penelitian ini hanya nisbah I1512/I897 pada inokulum 5% yang mengindikasikan
Gambar 2.5 Spektra FTIR bambu setelah pra-perlakuan biologis dengan TV (A) inokulum 5% dan (B) 10% pada berbagai waktu inkubasi (1) 1736 cm-1, (2) 1512 cm-1, (3) 1373 cm-1, (4) 1165 cm-1, (5) 897 cm-1
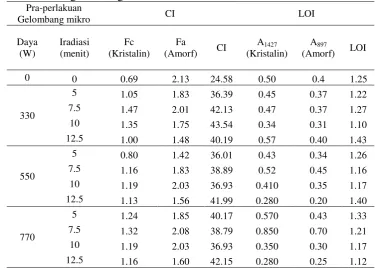
2.3.6 Indeks Kristalinitas Bahan
Lateral order index (LOI) dan Indeks kristalinitas (CI) bambu sebelum dan setelah pra-perlakuan ditabulasikan pada Tabel 2.4. Secara alami rantai selulosa terdiri dari daerah kristalin dan amorf. Struktur kristalin selulosa dapat menghambat produksi gula dalam konversi lignoselulosa. Indeks kristalinitas menggambarkan jumlah relatif bagian kristalin dalam selulosa, yang merupakan parameter sangat penting, yang dapat diukur dengan XRD, solid-state 13C NMR, spektroskopi IR dan raman spectroscopy. Intensitas minimum spektrum XRD dari puncak bidang (101) pada 2θ sekitar 18o yang mewakili bagian selulosa amorf dan
intensitas maksimum puncak bidang (002) pada 2θ sekitar 220
Highley et al. 1989; Kleman-Leyer et al.1992).Kehilangan komponen lignin dan hemiselulosa dapat menyebabkan hornifikasi pada selulosa ketika dikeringkan dan pengujian kristalinitas dilakukan pada sampel yang kering. Hornifikasi meningkatkan indeks kristinitas lignoselulosa. Setelah inkubasi 30 hari, indeks kristalinitas mulai menurun pada inokulum 5%, mungkin karena berlanjutnya serangan jamur pada bagian selulosa kristalin setelah mendegradasi bagian amorf yang lebih mudah terakses untuk digunakan sebagai nutrisi. Namun, fenomena ini tidak terjadi pada inokulum 10% dimana indeks kristalinitas cenderung meningkat setelah inkubasi 30 hari. Penyebab pasti perbedaan fenomena ini belum diketahui pasti. Tetapi indeks kristalinitas sampel setelah pra-perlakuan masih lebih tinggi daripada indeks kristalinitas kontrol.
Tabel 2.4 Lateral order index (LOI) dan indeks kristalinitas (CI) bambu setelah pra-perlakuanbiologis
Inoku lum (%)
Inku basi (hari)
CI LOI
Fc (Kristalin)
Fa (Amorf)
CI A1427
(Kristalin) A897
(Amorf) LOI
0 45 0.662 1.513 30.43 0.42 0.33 1.27
5
15 0.759 1.437 34.57 0.60 0.48 1.25
30 0.982 1.575 38.39 0.38 0.31 1.23
45 0.471 1.002 32.00 0.31 0.25 1.24
10
15 0.991 1.715 36.62 0.50 0.40 1.25
30 0.698 1.567 30.83 0.27 0.21 1.29
45 0.926 1.545 37.48 0.59 0.48 1.23
Ketercernaan bambu setelah pra-perlakuanberkorelasi negatif dengan nilai LOI. Nilai LOI dari sampel setelah pra-perlakuan lebih rendah daripada kontrol kecuali pada inokulum 10% selama 30 hari (Tabel 2.4). Pada kondisi inkubasi 30 hari terjadi perubahan fase kristal selulosa menjadi triklinik dari fase monoklinik pada kontrol dan perlakuan dengan inkubasi selama 15 hari. Hal ini mengindikasikan bahwa pra-perlakuan jamur dapat memperbaiki aksesibilitas permukaan selulosa dan secara teori akan mendukung proses hidrolisis selulosa menjadi lebih efisien pada sampel setelah pra-perlakuan. Jamur lebih mudah mendegradasi hemiselulosa dan menyisakan lebih banyak selulosa (Yu et al. 2009). Transformasi struktur kristalin selulosa menjadi bentuk amorf dapat meningkatkan nilai LOI. Kecenderungan yang sama juga terjadi dengan pra-perlakuan larutan ionik terhadap selulosa dimana pra-perlakuan menyebabkan konversi struktur kristalin menjadi amorf (Li et al. 2010). Selain melalui nilai LOI dan CI, perubahan kristalinitas dapat diduga melalui pendekatan nisbah antara A3308/A1330, yang menunjukkan intensitas ikatan hidrogen (HBI). Nilai ini
dapat dihitung dari absorbansi IR (Oh et al. 2005; Siroky et al. 2010).
Tabel 2.5 Ukuran kristal bambu setelah pra-perlakuan biologis
Gambar 2.6 Spektra XRD bambu setelah pra-perlakuan dengan TV (A) (inokulum 10%) dan (B) (inokulum 5%)