(HYMENOPTERA: EUPELMIDAE) SEBAGAI DASAR
PENGENDALIAN HAYATI KEPIK PENGISAP BUAH LADA
Dasynus piperis CHINA (HEMIPTERA: COREIDAE)
IWA MARA TRISAWA
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi Kajian Ekologi Parasitoid
Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae) Sebagai Dasar Pengendalian Hayati Kepik Pengisap Buah Lada Dasynus piperis China (Hemiptera: Coreidae) adalah karya saya dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi manapun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Januari 2011
Iwa Mara Trisawa
© Hak cipta milik IPB, tahun 2011
Hak Cipta dilindungi undang-undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
(HYMENOPTERA: EUPELMIDAE) SEBAGAI DASAR
PENGENDALIAN HAYATI KEPIK PENGISAP BUAH LADA
Dasynus piperis CHINA (HEMIPTERA: COREIDAE)
IWA MARA TRISAWA
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Entomologi - Fitopatologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup: Dr. Ir. Teguh Santoso, D.E.A. Dr. Ir. Ade Wachjar, M.S.
Penulis panjatkan puji dan syukur kepada Allah Subhanahu Wataalla atas segala rahmat dan karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Penelitian yang berjudul Kajian Ekologi Parasitoid Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae) Sebagai Dasar Pengendalian Hayati Kepik Pengisap Buah Lada Dasynus piperis China (Hemiptera: Coreidae), dilaksanakan sejak Mei 2008 sampai Desember 2009 di Bogor dan Bangka.
Bagian dari disertasi ini telah diterbitkan di Jurnal Penelitian Tanaman Industri Vol 16 No. 3 tahun 2010, dengan judul Kesesuaian Telur Kepik Kedelai untuk Pembiakan Massal Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae), Parasitoid Telur Kepik Lada.
Penulis mengucapkan terimakasih kepada Bapak Prof. Dr. Ir. Aunu Rauf, M.Sc., Bapak Prof. Dr. Ir. Utomo Kartosuwondo, M.S., Ibu Dr. Ir. Nina Maryana, M.Si. dan Bapak Dr. Ir. Ali Nurmansyah, M.Si. atas bimbingan dan pengarahan kepada penulis mulai dari perencanaan penelitian hingga penyelesaian disertasi. Terimakasih penulis ucapan juga kepada Kepala Badan Penelitian dan Pengembangan Pertanian, Kepala Pusat Penelitian dan Pengembangan Perkebunan, Kepala Balai Penelitian Tanaman Rempah dan Aneka Tanaman Industri, Dekan Sekolah Pasca Sarjana IPB yang telah memberikan kesempatan kepada penulis untuk melanjutkan pendidikan program doktor. Kepada Ketua Program Studi Entomologi/Fitopatologi dan seluruh dosen di Departemen Proteksi Tanaman IPB, penulis mengucapkan terimakasih atas segala arahan dan didikan.
Kepada Kepala Balai Penelitian Tanaman Obat dan Aromatik (Balittro) Bogor, dan Kepala Balai Pengkajian dan Teknologi Pertanian Kepulauan Bangka Belitung yang telah memberikan kesempatan dan fasilitas penelitian, penulis ucapkan terimakasih. Di samping itu, terimakasih penulis sampaikan kepada The Indonesian International Education Foundation (IIEF) melalui program
Indonesian Scholarship Dissertation Award (ISDA) yang telah membantu memberikan beasiswa penelitian.
Terimakasih penulis ucapkan kepada Bapak Prof. (R) Dr. Ir. I Wayan Laba, M.Sc, Dr. Ir. Wiratno, M.Env.Mgt., dan Ir. Fardedi, M.Si. atas segala dukungan dan kerjasama yang baik, serta kepada Sdr. Ahyar dan Bapak Muchyadi yang telah membantu pelaksanaan penelitian.
Kepada orangtua, ayahanda E. Sukarsawinata (Alm.) dan Ibunda R.E. Fatimah (Alm.), serta mertua Muchtar (Alm) dan R. Martha S. disampaikan terimakasih atas pendidikan, dukungan, nasihat, dan doa sehingga penulis dapat menyelesaikan pendidikan ini. Kepada istri Tini Mardiyana Ruliyanti dan ketiga putri tercinta Edvami Maradiyana Praeswitari, Vidina Diniarti Hanifa, dan Clarinta Tidara Estriva serta seluruh keluarga yang selalu mendoakan dan memberikan dukungan dalam menyelesaikan pendidikan ini, penulis mengucapkan terimakasih.
Semoga karya ilmiah ini bermanfaat.
Bogor, Januari 2011
Penulis dilahirkan di Bogor, pada tanggal 29 Juli 1963 sebagai anak ketiga dari pasangan E. Sukarsawinata (Alm) dan RE. Fatimah (Alm.). Pendidikan sarjana ditempuh di Jurusan Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam Universitas Pakuan Bogor, lulus pada tahun 1988. Pada tahun 2002, penulis diterima di Program Studi Entomologi/Fitopatologi pada Sekolah Pascasarjana IPB dan menamatkannya pada tahun 2005. Kesempatan untuk melanjutkan ke program doktor pada program studi dan perguruan tinggi yang sama diperoleh pada tahun 2006. Beasiswa pendidikan pascasarjana diperoleh dari Badan Penelitian dan Pengembangan Pertanian, Departemen Pertanian Republik Indonesia. Penulis juga merupakan salah satu dari enam mahasiswa program doktor IPB sebagai penerima Indonesian Scholarship Dissertation Award dari program kerjasama Ford Foundation dan Indonesian International Education Foundation.
Hasil dan Pembahasan ... Kesimpulan ... Daftar Pustaka ... 6 PENGARUH VEGETASI LIAR BERBUNGA TERHADAP PARA- SITOID Anastatus dasyni FERR. (HYMENOPTERA: EUPELMI- DAE ... Abstrak ... Abstract ... Pendahuluan ... Bahan dan Metode ... Hasil dan Pembahasan ... Kesimpulan ... Daftar Pustaka ... 7 SINTESIS ... 8 KESIMPULAN DAN SARAN ... DAFTAR PUSTAKA ...
51 55 55
Halaman 3.1 Karakteristik perkembangan pradewasa dan kehidupan imago beti-
na A. dasyni pada dua jenis inang alternatif ... 3.2 Dimensi tubuh A. dasyni betina yang dipelihara pada telur kepik
kedelai ... 3.3 Dimensi tubuh A. dasyni jantan yang dipelihara pada telur kepik
kedelai ... 3.4 Berbagai statistik neraca hayati A. dasyni ... 3.5 Proporsi fase perkembangan A. dasyni pada sebaran stabil ... 4.1 Nilai indeks preferensi masing-masing jenis inang pada pengujian preferensi di laboratorium dan lapangan ... 4.2 Tingkat parasitisasi A. dasyni pada umur inang alami ... 4.3 Tingkat parasitisasi A. dasyni pada berbagai lama penyimpanan inang ... 5.1 Rataan inang yang diparasit A. dasyni pada berbagai kerapatan
inang ... 5.2 Hasil analisis regresi logistik proporsi telur kepik D. piperis yang terparasit oleh A. dasyni ... 5.3 Nilai penduga parameter laju pencarian seketika (a) dan masa pe- nanganan inang (Th) berdasarkan model tanggap fungsional tipe II 6.1 Lama hidup imago A. dasyni yang dipelihara pada bunga vegetasi
liar ... 6.2 Jumlah keturunan dan nisbah kelamin A. dasyni yang dipelihara pada bunga vegetasi liar ... 6.3 Kadar gula tereduksi pada berbagai bunga vegetasi liar ...
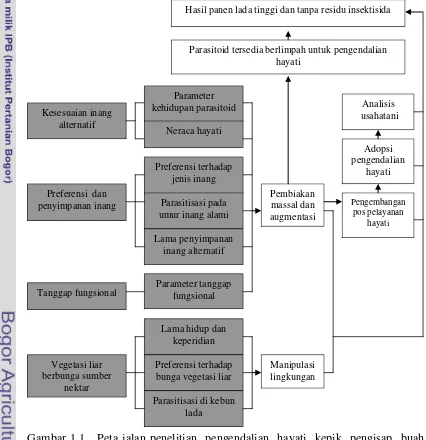
Halaman 1.1 Peta jalan penelitian pengendalian hayati kepik pengisap buah lada
D. piperis dengan parasitoid A. dasyni ... 5.1 Rataan nilai pengamatan proporsi inang yang diparasit dan pendu-
ga berdasarkan hasil analisis regresi logistik ... 5.2 Kurva tanggap fungsional parasitoid A. dasyni terhadap peningkat- an kerapatan inang ... 6.1 Pilihan parasitoid A. dasyni betina yang lapar terhadap bunga vege- tasi liar dan inang ... 6.2 Pilihan parasitoid A. dasyni jantan yang lapar terhadap bunga vege-
tasi liar ... 6.3 Lama kunjungan parasitoid A. dasyni betina yang lapar pada bunga vegetasi liar ... 6.4 Lama kunjungan parasitoid A. dasyni jantan yang lapar pada bunga
vegetasi liar ... 6.5 Pilihan parasitoid A. dasyni betina yang kenyang terhadap bunga
dan inang ... 6.6 Lama kunjungan parasitoid A. dasyni betina yang kenyang pada
bunga dan inang ... 6.7 Pilihan parasitoid A. dasyni betina yang lapar antara bunga vegeta- si liar dan inang pada pengujian lorong “Y” ... 6.8 Pilihan parasitoid A. dasyni betina yang kenyang antara bunga ve-
getasi liar dan inang pada pengujian lorong “Y” ... 6.9 Rataan tingkat parasitisasi oleh kompleks parasitoid di kebun lada
yang disiangi dan tidak ... 6.10Rataan tingkat parasitisasi A. dasyni di kebun lada yang disiangi
Halaman 1 Bunga vegetasi liar dan tanaman penutup tanah yang umum dijumpai
di kebun lada ... 2 Berbagai kondisi kebun lada ...
PENDAHULUAN
Latar Belakang
Lada (Piper nigrum L.) merupakan salah satu tanaman rempah yang
sebagian besar (99.89%) diusahakan dalam bentuk perkebunan rakyat, sedangkan
sisanya (0.11%) dalam bentuk perkebunan besar swasta (Ditjenbun 2008).
Meskipun bukan tanaman asli Indonesia, tanaman yang diintroduksi dari India ini
tumbuh dan berkembang serta memiliki peranan penting dalam perekonomian
nasional. Lada merupakan produk rempah-rempah pertama Indonesia yang
diperdagangkan ke Eropa melalui Arab dan Persia. Sebelum perang dunia ke-dua,
Indonesia bahkan tercatat sebagai penghasil lada terbesar dunia dan memasok
80% kebutuhan lada dunia (Wahid 1996). Pada tahun 2007 volume dan nilai
ekspor lada Indonesia tercatat 38.447 ton dengan nilai US $ 132.495.000,- yang
terdiri atas 15.544 ton lada putih, 20.881 ton lada hitam, dan 2.022 ton dalam
bentuk lada lainnya (Ditjenbun 2008). Lada digunakan sebagai bumbu makanan
baik pada sektor rumah tangga maupun industri makanan.
Sentra pertanaman lada di Indonesia terdapat di Lampung, Bangka,
Kalimantan Barat, Kalimantan Timur, dan Sulawesi Selatan. Total luas areal
pada tahun 2009 diperkirakan 191.608 ha dengan produksi 81.660 ton.
Dibandingkan tahun 2003 yang mencapai 204.128 ha, luas areal pada beberapa
tahun belakangan ini mengalami penyusutan (Ditjenbun 2008). Di antara
faktor penyebabnya adalah kondisi perekonomian nasional dan dunia, situasi
politik dan keamanan serta harga lada. Kondisi tersebut ditambah dengan teknik
budidaya yang minimal menyebabkan produksi tanaman lada menurun.
Penurunan produksi dan munculnya negara pesaing baru seperti Vietnam
menyebabkan Indonesia tidak lagi menjadi pemasok utama lada dunia. Sebagai
gambaran, saat ini Indonesia menempati peringkat ke-tiga sebagai negara
penghasil lada hitam di dunia setelah Vietnam dan India, sedangkan untuk lada
putih Indonesia tetap produsen utama (Manohara et al. 2007).
Salah satu kendala penurunan produksi lada adalah gangguan hama dan
dari 10 milyar rupiah (Ditlintanbun 2004). Di antara hama pada tanaman lada
yang sering menimbulkan kerusakan adalah kepik pengisap buah lada, Dasynus
piperis China (Hemiptera: Coreidae) (Deciyanto et al. 1993; Trisawa et al. 2007). Nimfa dan imago D. piperis mengisap cairan buah lada. Buah yang
diisap menunjukkan gejala bercak hitam, hampa, kering, dan kemudian gugur.
Kondisi buah terserang dapat juga diperburuk oleh kehadiran mikroorganisme
seperti cendawan dan bakteri yang menyebabkan buah menjadi busuk (Deciyanto
& Wikardi 1989; Wikardi & Asnawi 1996). Buah lada mulai diserang saat
berumur 4 bulan sampai 5 bulan. Namun demikian, buah lada umur 6 bulan
sampai 9 bulan paling sesuai untuk perkembangan D. piperis karena imago
hidup lebih lama, bertelur lebih banyak, persentase tetas telur dan jumlah nimfa
yang menjadi imago lebih tinggi (Suprapto & Thomas 1989).
Hasil survei Laba et al. (2004) di Bangka menunjukkan rataan tingkat
serangan 36.80%. Di Lampung dilaporkan serangan D. piperis mengakibatkan kerugian produksi sebesar 15% (Suprapto & Thomas 1989). Survei yang sama
oleh Trisawa et al. (1992) pada beberapa kecamatan di Kabupaten Sambas,
Kalimantan Barat menunjukkan tingkat serangan berkisar antara 13.52% sampai
18.68%.
Petani lada umumnya mengandalkan penggunaan insektisida untuk
mengendalikan D. piperis, dengan frekuensi 1 kali sebulan sejak berbunga hingga
panen atau sekitar 10 kali penyemprotan dalam setahun. Selain menimbulkan
dampak buruk terhadap lingkungan seperti munculnya hama sekunder, resistensi
hama, dan terbunuhnya musuh alami, aplikasi insektisida meninggalkan residu
pada hasil panen. Hal yang disebut terakhir ini dapat berdampak pada daya saing
yang rendah di pasar global. Salah satu pendekatan yang dapat dilakukan untuk
mengurangi penggunaan insektisida adalah pengendalian hayati dengan
memanfaatkan musuh alami hama. Tercatat ada tiga jenis parasitoid telur
D. piperis yang potensial yaitu Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae), Gryon dasyni Nix. (Hymenoptera: Scelionidae) dan Ooencyrtus
malayensis Ferr. (Hymenoptera: Encyrtidae). A. dasyni merupakan jenis yang paling dominan dibandingkan dua parasitoid lainnya, dengan tingkat parasitisasi
Peta Jalan Penelitian
Pemanfaatan parasitoid A. dasyni dalam pengendalian hayati kepik
pengisap buah lada memerlukan pemahaman yang mendasar tentang berbagai
aspek biologi dan ekologi parasitoid. Gambar 1.1 menyajikan tahapan penelitian
atau kajian yang perlu ditempuh dalam rangka pengembangan pengendalian
hayati D. piperis dengan memanfaatkan parasitoid A. dasyni. Uraian di bawah ini
dibatasi pada tahapan penelitian yang secara langsung menyediakan landasan bagi
upaya augmentasi dan manipulasi lingkungan.
Gambar 1.1 Peta jalan penelitian pengendalian hayati kepik pengisap buah lada D. piperis dengan parasitoid A. dasyni. Kotak berwarna gelap adalah tahapan penelitian yang merupakan bagian dari disertasi.
Hasil panen lada tinggi dan tanpa residu insektisida
Penyediaan parasitoid dalam jumlah yang banyak memerlukan penyediaan
inang yang banyak pula. Pengalaman selama ini menunjukkan bahwa cukup
banyak kendala dalam membiakkan parasitoid A. dasyni dengan menggunakan
telur D. piperis. Kendala tersebut di antaranya adalah sulitnya mendapatkan telur
D. piperis dari hasil pembiakan massal. Oleh karena itu, upaya mendapatkan inang alternatif yang sesuai untuk membiakkan parasitoid A. dasyni merupakan hal yang sangat penting. Penelitian pendahuluan di laboratorium mendapatkan
bahwa parasitoid A. dasyni dapat dibiakkan dengan mudah pada telur kepik
polong kedelai, Riptortus linearis (Hemiptera: Alydidae) dan Nezara viridula
(Hemiptera: Pentatomidae). Sebelum telur R. linearis dan N. viridula dijadikan
inang alternatif untuk pembiakan massal parasitoid A. dasyni, perlu diteliti sejauh
mana pengaruh inang alternatif ini terhadap kehidupan parasitoid. Pendekatan
yang dapat dilakukan adalah melalui pengkajian neraca hayati. Dari neraca hayati
dapat dihitung beberapa statistik demografi. Salah satu statistik demografi yang
paling handal untuk mengukur potensi musuh alami adalah laju pertambahan
intrinsik (r), karena di dalamnya telah mempertimbangkan karakteristik kehidupan
serangga seperti masa hidup, keperidian, sintasan, dan nisbah kelamin (Carey
1993). Nilai r merupakan salah satu kriteria yang penting untuk mengevaluasi
keefektifan atau potensi dari agens pengendalian hayati (Lee & Ahn 2000), serta
dapat digunakan untuk menduga potensi pertumbuhan populasi parasitoid (Lysyk
2000).
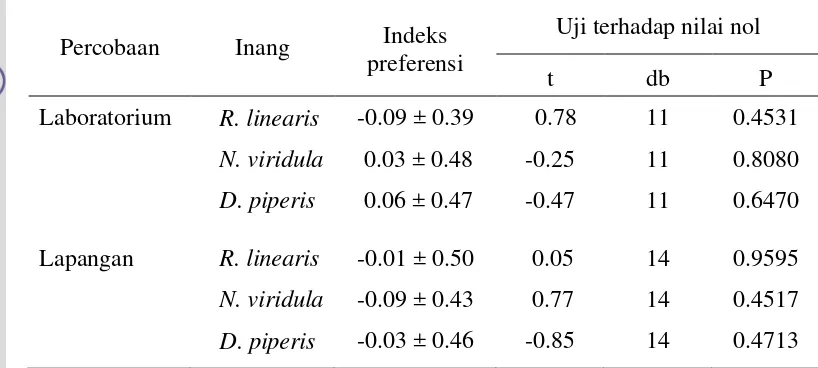
Parasitoid A. dasyni hasil pembiakan pada inang alternatif, sebelum dilepas ke lapangan perlu dikaji tingkat preferensinya baik terhadap inang alami
maupun inang alternatif. Di samping itu, perlu dikaji tingkat parasitisasinya pada
beberapa umur inang. Hal ini penting untuk memastikan bahwa parasitoid
tersebut efektif dan mampu berkembang sesuai dengan fenologi inang di
lapangan. Informasi yang diperoleh menjadi acuan dalam penerapan salah satu
pendekatan pengendalian hayati yaitu augmentasi (pelepasan parasitoid).
Selanjutnya, untuk efisiensi pembiakan massal A. dasyni diperlukan kajian
tentang pengaruh lama penyimpanan inang dalam suhu dingin terhadap
kemampuan parasitisasi. Hal ini penting mengingat inang yang dibiakkan
mendatang. Penyimpanan inang ini ditujukan sebagai stok yang dapat langsung
digunakan ketika parasitoid perlu segera dibiakkan. Cara ini dapat memangkas
waktu penyediaan inang dibandingkan jika inang diperoleh melalui proses
pemeliharaan dan pembiakan serangga dewasa.
Keefektifan parasitoid juga perlu dikaji berdasarkan tanggapnya terhadap
peningkatan kerapatan inang. Secara umum individu parasitoid biasanya akan
memberikan tanggap terhadap peningkatan kelimpahan hama, yang disebut
tanggap fungsional. Dari kegiatan penelitian ini dapat ditentukan apakah
A. dasyni memperlihatkan tanggap fungsional tipe I, II, atau III. Parameter penting dari tanggap fungsional adalah laju pencarian seketika (a) dan masa
penanganan inang (Th). Parasitoid yang potensial adalah yang memiliki nilai a
yang tinggi dan nilai Th yang rendah (Hassell 2000).
Keberhasilan pemanfaatan A. dasyni dalam pengendalian hayati D. piperis
di lapangan perlu didukung oleh ketersediaan pakan bagi imago betina parasitoid.
Salah satu sumber pakan bagi parasitoid di lapangan adalah nektar yang terdapat
pada bunga vegetasi liar. Pada pertanaman lada tumbuh beberapa vegetasi liar
berbunga dan belum diketahui bagaimana peranan bunga vegetasi liar tersebut
sebagai sumber nektar parasitoid A. dasyni. Hal ini perlu diteliti, termasuk juga
terhadap bunga A. pintoi yang selama ini dianjurkan ditanam pada kebun lada
karena dianggap dapat meningkatkan tingkat parasitisasi parasitoid. Dari
penelitian ini dapat ditentukan jenis vegetasi liar yang dapat mendukung
kehidupan parasitoid. Pengetahuan ini diperlukan untuk mengelola ekosistem
lada yang menunjang pengendalian hayati kepik pegisap buah lada.
Tujuan Penelitian
Secara umum penelitian bertujuan memanfaatkan parasitoid A. dasyni
sebagai agens pengendalian hayati kepik pengisap buah lada D. piperis. Secara
lebih khusus penelitian bertujuan (1) mengkaji kesesuaian telur kepik kedelai
R. linearis dan N. viridula sebagai inang alternatif untuk pembiakan massal parasitoid A. dasyni berdasarkan berbagai parameter hayati; (2) mengkaji tingkat
preferensi parasitoid A. dasyni hasil pembiakan pada inang alternatif, serta pengaruh lama penyimpanan inang pada suhu dingin terhadap parasitisasi; (3)
telur kepik buah lada; dan (4) mengkaji pengaruh berbagai bunga vegetasi liar
sebagai sumber nektar terhadap lama hidup, keperidian, dan perilaku parasitoid di
laboratorium, serta pengaruh keberadaan vegetasi liar terhadap tingkat parasitisasi
TINJAUAN PUSTAKA
Kepik Pengisap Buah Lada
Telur D. piperis berwarna coklat muda sampai tua, berbentuk lonjong, berukuran panjang 1.5 mm, lebar 1.0 mm, dan tinggi 0.9 mm. Telur diletakkan
oleh imago betina secara berkelompok antara 3 butir sampai 10 butir (Kalshoven
1981). Hasil penelitian Karmawati (1988) menunjukkan bahwa 50% telur
diletakkan pada bagian tengah tajuk tanaman, menyebar secara mengelompok
pada daun atau bulir buah. Sebanyak 81% telur diletakkan pada permukaan atas
dan bawah daun. Stadium telur berlangsung antara 7 hari sampai 8 hari
(Kalshoven 1981).
Nimfa instar-1 yang baru keluar dari telur berukuran kurang lebih 2.0 mm,
berwarna kuning kecoklatan, tidak bersayap, dan memiliki antena yang lebih
panjang dibandingkan panjang tubuhnya. Pada antena terdapat dua ruas yang
menebal. Nimfa selanjutnya segera mencari makanan dan 98% nimfa instar muda
terdapat pada buah (Kalshoven 1981). Nimfa umumnya (65%) menyukai buah
yang terdapat pada bagian tengah tajuk. Pola sebaran nimfa tersebut tidak
berbeda jauh dengan pola sebaran telur, karena mobilitas nimfa yang tidak terlalu
aktif (Karmawati 1988). Stadium nimfa berlangsung antara 3 minggu sampai 4
minggu dan mengalami empat kali pergantian kulit sampai terbentuk imago.
Lama stadium nimfa dapat dipengaruhi oleh umur buah lada yang menjadi
makanannya. Stadium nimfa berlangsung antara 26 hari sampai 33 hari pada buah
lada umur 4.5 bulan sampai 6 bulan, sedangkan pada buah umur 6 bulan sampai 9
bulan antara 19 hari sampai 25 hari (Kalshoven 1981; Suprapto & Thomas 1989).
Imago berwarna hijau kecoklatan. Panjang tubuh antara 12.0 mm sampai
13.0 mm dan lebar 4.0 mm sampai 5.0 mm. Imago jantan dan betina dapat
dibedakan berdasarkan ukuran tubuh. Imago jantan lebih kecil dan ramping,
sedangkan imago betina lebih besar. Lama hidup imago kurang lebih 3 bulan.
Imago betina meletakkan telur pertama pada umur 14 hari dan selama hidupnya
sampai 18:00 (Kalshoven 1981). Tempat yang rimbun dan agak gelap lebih
disukai sebagai tempat peletakan telur (Karmawati 1988).
Kepik pengisap buah lada D. piperis selalu dijumpai sepanjang tahun pada
tanaman lada. Namun demikian, populasinya bergantung pada musim buah.
Populasi D. piperis paling tinggi pada bulan Juni dan Nopember, sedangkan populasi rendah terjadi pada bulan Juli sampai September karena buah telah
dipanen (Karmawati 1988; Deciyanto 1991). Setiap fase perkembangan D. piperis
dapat dijumpai secara bersamaan di lapangan dan menyebar pada tajuk tanaman.
Parasitoid Anastatus dasyni Ferr.
Fase perkembangan A. dasyni pada telur D. piperis diuraikan oleh Trisawa
(2005) sebagai berikut: telur berbentuk lonjong, berwarna putih kotor, berukuran
panjang 0.38 mm dan lebar 0.16 mm. Larva berwarna putih kekuningan. Larva
instar awal berukuran panjang 0.57 mm dan lebar 0.21 mm, sedangkan larva instar
lanjut berukuran panjang 1.06 mm dan lebar 0.47 mm. Prapupa berwarna putih
kekuningan dan lebih gelap dibandingkan warna larva. Panjang prapupa awal
1.06 mm dan lebar 0.46 mm, sedangkan satu hari menjelang pupa panjangnya
1.12 mm dan lebar 0.48 mm. Pupa yang baru terbentuk berwarna putih
kecoklatan. Bagian tubuh seperti mata, tungkai, antena, sayap, ruas abdomen
sudah terbentuk dan terlihat jelas. Pupa kemudian berubah warna menjadi
kehitaman dan organ tubuh sudah terbentuk lengkap. Panjang pupa 1.49 mm dan
lebar 0.52 mm. Di sekitar kepala terdapat selubung warna coklat muda.
Perkembangan pradewasa A. dasyni mulai dari telur sampai pupa berlangsung di
dalam telur inang. Waktu yang dibutuhkan mulai telur diletakkan sampai imago
keluar adalah 15.57 hari.
Imago A. dasyni keluar dengan cara merobek korion telur D. piperis. Imago keluar mulai hari ke 13 sampai ke 18 setelah telur diletakkan. Imago
jantan dan betina berwarna hitam dan mudah dibedakan, terutama dari ukuran
tubuh. Imago betina lebih besar dibandingkan jantan. Di samping itu venasi
sayap imago betina berwarna coklat muda sedangkan imago jantan transparan.
Satu ekor imago betina A. dasyni mampu menghasilkan keturunan antara 60 ekor
sampai 136 ekor. Berdasarkan pengamatan perilaku kopulasi, imago betina
sudah kopulasi akan menolak kehadiran jantan. Kopulasi berlangsung sangat
singkat, sekitar 2 detik sampai 3 detik.
Imago betina dapat hidup selama 1 bulan (Deciyanto et al. 2000; Trisawa
et al. 2007). Pemberian madu dapat memperpanjang lama hidup parasitoid A. dasyni di laboratorium (Deciyanto & Wikardi 1989; Trisawa et al. 2007). Menurut Mendel et al. (1987), pemberian madu dapat meningkatkan lama hidup
parasitoid Anastatus seperti juga yang terjadi pada A. semiflavidus (Hymenoptera:
Eupelmidae). Pada kasus parasitoid yang lain, Schmale et al. (2001) melaporkan
bahwa parasitoid Dinarmus basalis (Hymenoptera: Pteromalidae) yang diberi
pakan madu, meningkat lama hidupnya tiga kali lipat.
Preferensi, Umur Inang dan Nisbah Kelamin Parasitoid
Preferensi parasitoid terhadap inang menunjukkan suatu kemampuan
parasitoid dalam membedakan antara inang yang sesuai untuk oviposisi dan yang
tidak. Hal ini menunjukkan terjadinya diskriminasi terhadap inang. Perilaku
seperti ini merupakan ciri keefektifan parasitoid dalam menekan kepadatan inang
(van Lenteren et al. 1978; van Alphen & Jervis 1997).
Preferensi parasitoid terhadap inang dipengaruhi oleh beberapa faktor, di
antaranya adalah umur inang. Pada parasitoid telur, umur inang sangat penting
bagi perkembangan pradewasa parasitoid karena embriogenesis pada inang
berpengaruh terhadap nutrisi. Kualitas nutrisi inang akan semakin turun seiring
dengan meningkatnya umur inang (Vinson 1994). Di samping berpengaruh
terhadap perkembangan embrio, umur inang yang meningkat juga akan
menyebabkan kulit inang semakin keras sehingga mempersulit parasitoid untuk
melakukan oviposisi (Gross 1993).
Contoh dari pengaruh umur inang terhadap pilihan parasitoid dalam
meletakkan telurnya ditunjukkan oleh parasitoid A. dasyni. Parasitoid tersebut
lebih memilih telur D. piperis umur ≤ 3 hari dibandingkan umur telur yang lebih
tua untuk peletakan telurnya. Hal ini karena pada umur yang lebih tua, bakal
nimfa D. piperis sudah terbentuk (Trisawa et al. 2007). Selama masa pradewasa,
parasitoid menggunakan nutrisi inang untuk perkembangannya. Nutrisi inang
yang sesuai akan menghasilkan imago parasitoid yang lebih bugar (Mackauer &
Pada inang yang sudah dipilih, parasitoid akan meletakkan telur pada
inang tersebut untuk jenis kelamin tertentu dari keturunanannya. Dalam alokasi
jenis kelamin keturunan, faktor yang mempengaruhi adalah umur dan ukuran
inang, jumlah perkawinan, faktor lingkungan yaitu temperatur dan kelembapan
(Colazza & Wajnberg 1998; Gordh et al. 1999).
Betina parasitoid akan mengalokasikan jenis kelamin betina pada ukuran
inang yang relatif lebih besar dan jenis kelamin jantan pada inang yang relatif
lebih kecil (Charnov et al. 1981; Lee 2009). Sebagai contoh, parasitoid Syngaster
lepidus (Hymenoptera: Braconidae) mengalokasikan 80% keturunan betina pada inangnya yaitu larva Phoracantha spp. yang berukuran besar, sedangkan pada
inang yang berukuran kecil mengalokasikan keturunan jantan. Ukuran parasitoid
yang dihasilkan juga meningkat dengan meningkatnya ukuran inang (Joyce et al.
2002).
Neraca Hayati
Perkembangan populasi serangga dapat dipelajari dengan menyusun
neraca hayati. Neraca hayati menunjukkan perkembangan kehidupan serangga
yang memberikan informasi mengenai kelahiran dan kematian, serta peluang
untuk berkembangbiak (Tarumingkeng 1994; Price 1997). Potensi
perkembangbiakan serangga penting untuk diketahui, misalnya untuk meramalkan
kenaikan populasinya pada waktu tertentu dan dampak yang mungkin
ditimbulkannya pada tanaman (Waters 1969; Royama 1981).
Dari neraca hayati dapat dibuat kurva kesintasan (survivorship curve)
populasi serangga yang diamati. Kurva tersebut akan menggambarkan pola
bertahan hidup serangga. Menurut Price (1997) terdapat tiga pola bertahan hidup,
yaitu tipe I yang menunjukkan kematian serangga pada umur muda dalam jumlah
sedikit dan kematian serangga pada umur yang lebih tua dalam jumlah banyak;
tipe II yang menunjukkan laju kematian konstan; dan tipe III yang menunjukkan
kematian serangga pada umur muda dalam jumlah banyak.
Dari neraca hayati juga dapat dihitung beberapa statistik demografi yaitu
laju reproduksi bersih (Ro), waktu satu generasi (T), laju pertambahan intrinsik
Laju reproduksi bersih (Ro) adalah jumlah keturunan betina yang berhasil
menjadi imago, atau kelipatan populasi per generasi. Suatu populasi dikatakan
stabil bila Ro = 1, populasi bertambah bila Ro > 1, dan populasi berkurang bila Ro
< 1. Bila Ro suatu spesies diketahui maka waktu satu generasi (T) dan laju
pertambahan intrinsik (r) dapat dihitung (Price 1997). Waktu satu generasi (T)
adalah waktu yang dibutuhkan sejak telur tersebut diletakkan sampai imago yang
berasal dari telur tersebut menghasilkan separuh keturunannya, sedangkan laju
pertambahan intrinsik (r) adalah laju pertambahan populasi pada keadaan
sumberdaya tak terbatas. Bila nilai r diketahui maka laju pertambahan terbatas (λ)
yang merupakan kelipatan populasi per satuan waktu juga dapat diketahui (Price
1997; Carey 1993).
Laju pertambahan intrinsik (r) merupakan statistik demografi yang paling
handal untuk mengukur potensi musuh alami karena di dalamnya telah
mempertimbangkan masa hidup, keperidian, sintasan, dan nisbah kelamin (Carey
1993). Nilai r merupakan salah satu kriteria yang penting untuk mengevaluasi
keefektifan atau potensi agens pengendalian hayati (Lee & Ahn 2000), serta dapat
digunakan untuk menduga potensi pertumbuhan populasi parasitoid tersebut
(Lysyk 2000). Makin tinggi persentase telur yang diletakkan pada kelompok
umur muda maka makin besar nilai laju pertambahan intrinsik (Birch 1948).
Dalam pertumbuhan populasi serangga maka nilai reproduksi (RVx) dan
persebaran umur stabil (px) juga merupakan ciri yang penting. Nilai reproduksi
adalah sumbangan relatif suatu individu serangga pada umur tertentu terhadap
populasi. Nilai reproduksi pada individu yang berumur muda lebih rendah
dibandingkan kelompok umur yang lebih tua. Namun demikian, nilai reproduksi
tertinggi akan terjadi pada umur individu tertentu kemudian nilai reproduksi
menurun sampai nol (pasca reproduksi) (Carey 1993).
Sebaran umur stabil dapat menunjukkan proporsi individu-individu yang
berumur tertentu dari populasi. Pertumbuhan populasi yang memiliki sebaran
umur stabil sepanjang waktu memiliki proporsi jumlah individu yang tidak
berubah, sehingga proporsi masing-masing kelompok umur akan relatif sama
Tanggap Fungsional
Tanggap fungsional menyatakan perubahan jumlah inang atau mangsa
yang diserang oleh individu parasitoid atau predator akibat perubahan kerapatan
populasi inang atau mangsa per satuan waktu. Tanggap ini penting dalam
interaksi antara inang atau mangsa dengan parasitoid atau predator (Hassel 2000).
Tanggap fungsional kemudian menjadi salah satu ukuran untuk
menentukan keefektifan suatu parasitoid atau predator dalam mengendalikan
populasi hama atau kemampuannya mengatur keseimbangan populasi hama.
Keefektifan tersebut dipengaruhi oleh beberapa faktor seperti karakteristik inang
atau mangsa dan persebarannya pada tanaman (Kumar et al. 1999; Yasuda &
Ishikawa 1999).
Jumlah inang atau mangsa yang diparasit atau dimangsa pada kerapatan
inang atau mangsa tertentu merupakan aspek penting untuk dipelajari, sehingga
diperoleh gambaran tentang kemampuan parasitoid atau predator dalam
menangani inang atau mangsanya (Pervez & Omkar 2005; Rahman et al. 2009).
Holling (1959) mengidentifikasi tiga macam tipe fungsional: (1) tipe I
(linear) yaitu proporsi inang terparasit bersifat konstan, sehingga hubungan antara
banyaknya inang yang terparasit dan kerapatan inang bersifat linear, (2) tipe II
(hiperbolik) yaitu proporsi inang yang terparasit menurun tajam dengan
bertambahnya kerapatan inang, dan (3) tipe III (sigmoid) yaitu proporsi inang
yang terparasit awalnya meningkat, tetapi kemudian secara berangsur menurun
dengan meningkatnya kerapatan inang.
Secara umum individu parasitoid biasanya akan memberikan tanggap
terhadap peningkatan kerapatan inang. Pengetahuan tentang tanggap fungsional
dapat digunakan untuk menapis musuh alami yang potensial dan memperkirakan
potensi pengendalian hayati (Parella & Horsburgh 1983; Houck & Strauss 1985).
Parameter penting dari tanggap fungsional adalah laju pencarian seketika (a) dan
masa penanganan inang (Th). Parasitoid yang potensial adalah yang memiliki
nilai a yang tinggi dan nilai Th yang rendah (Hassell 2000).
Tanggap fungsional dapat dipengaruhi oleh beberapa faktor seperti stadia
mangsa, fase pertumbuhan tanaman, cuaca, kehadiran mangsa alternatif, dan
mempengaruhi laju pencarian mangsa dan lama predator menetap (Messina &
Hanks 1998; O’Neil 1998).
Pembiakan Massal dan Pelepasan Parasitoid
Parasitoid yang sudah ada di lapangan suatu saat populasinya dapat
rendah, tidak ada, atau terlambat kehadirannya sehingga perlu dilepas parasitoid
hasil pembiakan di laboratorium. Proses ini disebut augmentasi. Augmentasi
dilakukan dalam dua tipe, yaitu (1) inokulasi, yaitu pelepasan musuh alami dalam
jumlah yang sedikit sebagai inokulan dan pengendalian hama terjadi oleh generasi
musuh alami berikutnya, dan (2) inundasi, yaitu musuh alami dilepas dalam
jumlah yang lebih banyak dan terjadi pengendalian langsung oleh musuh alami
tersebut (van Driesche & Bellows 1996).
Augmentasi musuh alami banyak dilakukan untuk mengendalikan hama
tanaman dengan hasil yang cukup memuaskan (Wright et al. 2001; Head &
Walters 2002; Mills 2002). Di Indonesia, contoh augmentasi inundatif adalah
pelepasan parasitoid Trichogramma spp. untuk mengendalikan penggerek batang
tebu, sedangkan untuk augmentasi inokulatif yang dilepas adalah parasitoid
Diatraeophaga striatalis (Sosromarsono 1999).
Dalam pembiakan massal parasitoid digunakan inang alternatif. Dalam
pemilihan inang alternatif perlu dipertimbangkan kemudahan dalam pembiakan,
biaya murah, dan kesesuaian inang sehingga teknik pengendalian hayati yang
dikembangkan memang layak dilakukan. Inang alternatif yang demikian dikenal
dengan istilah factitious host (van Lenteren 1989; van Driesche & Bellows 1996).
Kesesuaian inang dapat diukur dari reproduksi parasitoid dan nisbah kelamin
betina yang tinggi (Godfray 1994).
Dalam pembiakan parasitoid sering diperoleh jumlah inang alternatif yang
melebihi kebutuhan. Kelebihan inang tersebut dapat diawetkan dengan
menyimpannya pada suhu dingin dalam jangka waktu tertentu. Sebagai contoh,
Tengkano (2000) menyatakan bahwa telur Riptortus linearis dapat diawetkan di
lemari es pada temperatur -2oC selama 34 hari. Telur yang diawetkan tersebut dapat digunakan untuk pembiakan massal parasitoid Ooencyrtus sp. untuk
Pemanfaatan Bunga Vegetasi Liar untuk Parasitoid
Berbagai cara dapat dilakukan untuk meningkatkan peran parasitoid di
lapangan. Manipulasi lingkungan yang bertujuan mengonservasi musuh alami
merupakan salah satu upaya mempertahankan dan melestarikan musuh alami yang
sudah ada di suatu tempat atau ekosistem dan membuatnya lebih efektif dalam
fungsinya (van Driesche & Bellows 1996). Manipulasi lingkungan dapat
dilakukan dengan menanam atau mengelola vegetasi penghasil nektar dan polen
di sekitar tanaman utama (van Driesche & Bellows 1996; Landis et al. 2000).
Nektar merupakan sumber pakan bagi parasitoid betina yang dapat meningkatkan
reproduksi parasitoid (Jervis et al. 1996; Lewis et al. 1998; Corteserro et al.
2000).
Manfaat vegetasi berbunga bagi parasitoid dikemukakan oleh Baggen dan
Gurr (1998). Bunga saba (Fagopyrum esculentum) mampu meningkatkan
keperidian Copidosoma koehleri (Hymenoptera: Encyrtidae), yang merupakan parasitoid telur Phthorimaea operculella (Lepidoptera: Gelechiidae). Di samping
itu, Idris dan Grafius (1995) menyatakan bahwa vegetasi berbunga seperti
beberapa anggota famili Brasicaceae mampu meningkatkan lama hidup dan
keperidian parasitoid Diadegma insulare (Hymenotera: Ichneumonidae).
Pemanfaatan vegetasi berbunga yang mampu meningkatkan keperidian
parasitoid betina dapat dipahami dari segi keseimbangan antara inang dan
kebutuhan pakan. Hal tersebut sangat penting dalam pengendalian hayati (Lewis
et al. 1998). Parasitoid yang mendapatkan pakan cukup akan meletakkan telur lebih banyak dibandingkan parasitoid yang lapar (Takasu & Lewis 1993). Hasil
penelitian Stapel et al. (1997) menunjukkan bahwa persentase parasitisasi
Microplitis croceipes (Hymenoptera: Braconidae) pada larva Helicoverpa zea (Lepidoptera: Noctuidae) lebih tinggi pada parasitoid betina yang kenyang
dibandingkan yang lapar. Pakan tambahan berupa nektar atau embun madu
merupakan faktor penting dalam meningkatkan daya bertahan hidup parasitoid, di
samping untuk produksi telur (Sosromarsono 2002).
Vegetasi berbunga perlu mudah didapat oleh parasitoid, karena parasitoid
dapat pergi ke tempat lain jika tidak ada pakan di sekitar tempat hidupnya. Jika
menjadi berkurang (Baggen & Gurr 1998). Oleh karena itu, sangat penting jika
keberadaan sumber pakan dekat dengan lokasi inang (Lewis et al. 1998). Di
samping itu, perlu diperhatikan bentuk bunga yang mudah dimanfaatkan oleh
parasitoid. Sebagai contoh, diameter mahkota bunga yang terbuka berpengaruh
positif terhadap lama hidup dan keperidian parasitoid D. insulare (Idris & Grafius
1995). Warna bunga juga menarik perhatian parasitoid. Menurut Kartosuwondo
(1993), bunga Nasturtium yang berwarna kuning cerah menarik perhatian imago
D. semiclausum. Parasitoid tersebut menurut Keller dan Baker (2002) juga tertarik pada bunga alyssum (Lobularia maritima) dan pakchoi (Brassica
campestris).
Pada pertanaman lada dianjurkan untuk menanam tanaman penutup
tanah Arachis pintoi yang berbunga terus menerus. Tanaman A. pintoi dapat meningkatkan parasitisasi parasitoid (Suprapto 2000, Trisawa et al. 2006).
Tingkat parasitisasi Spathius piperis (Hymenoptera: Braconidae) pada larva
penggerek batang lada Lophobaris piperis (Coleoptera: Curculionidae) berkisar
antara 25.0% sampai 50% pada tanaman lada dengan A. pintoi, sedangkan tanpa
A. pintoi hanya 5.2% sampai 10,8% (Suprapto 2000). Penanaman tanaman A. pintoi juga dapat meningkatkan parasitisasi total parasitoid telur D. piperis. Hasil survei Trisawa (2005) pada pertanaman lada di Bangka
menunjukkan bahwa tingkat parasitisasi tersebut mencapai 70.0%.
Beberapa jenis tumbuhan lain penghasil nektar untuk meningkatkan
populasi dan tingkat parasitisasi parasitoid pada tanaman lada adalah kopi (Coffea
arabica), sentrosema (Centrocema sp.), kalopogonium (Callopogonium sp.), kumis kucing (Orthosiphon aristatus), dan babadotan (Ageratum conyzoides)
(Suprapto & Yufdy 1988). Tingkat parasitisasi oleh kompleks parasitoid telur
KESESUAIAN TELUR KEPIK KEDELAI UNTUK
PEMBIAKAN MASSAL
Anastatus dasyni
FERR.
(HYMENOPTERA: EUPELMIDAE), PARASITOID TELUR
KEPIK LADA
[Suitability of soybean bug eggs for mass rearing of Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae), an egg parasitoid of pepper bug]
Abstrak
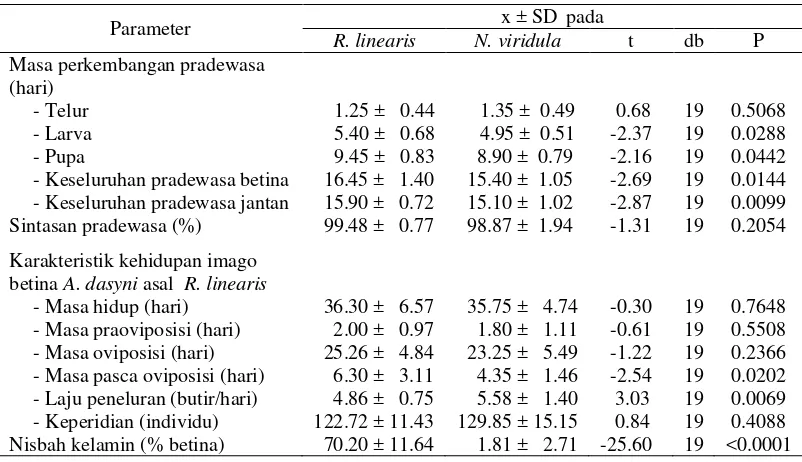
Penelitian bertujuan mengkaji kesesuaian telur kepik kedelai Riptortus linearis dan Nezara viridula sebagai inang untuk pembiakan massal parasitoid A. dasyni. Imago parasitoid A. dasyni yang berasal dari lapangan dipelihara secara terpisah pada telur dari kedua jenis kepik kedelai. Pengamatan dilakukan terhadap biologi A. dasyni yang meliputi masa perkembangan pradewasa dan berbagai karakteristik kehidupan imago betina. Selain itu, dilakukan analisis neraca hayati dengan menggabungkan data perkembangan dan sintasan pradewasa, masa hidup imago dan reproduksi, serta nisbah kelamin. Hasil penelitian menunjukkan bahwa masa perkembangan larva dan pupa A. dasyni pada telur N. viridula lebih singkat serta laju peneluran lebih tinggi dibandingkan pada telur R. linearis. Namun imago betina A. dasyni yang keluar dari telur N. viridula hanya 1.81%, sedangkan dari telur R. linearis sebanyak 70.20%. Oleh karena itu, parameter neraca hayati hanya dapat dihitung dari parasitoid yang dipelihara pada telur R. linearis. Laju pertambahan intrinsik parasitoid (r) 0.1870 betina/induk/hari, masa generasi (T) 27.51 hari, reproduksi bersih (Ro) 84.29 betina/induk/generasi; laju pertambahan terbatas (λ) 1.21 betina/induk/hari dan nilai reproduksi (RVx) 402.51 individu. Proporsi persebaran usia stabil (px) adalah 17.06% telur, 50.41% larva, 26.53% pupa, dan 6.02% imago. Telur kepik kedelai R. linearis dapat digunakan untuk pembiakan massal A. dasyni.
Kata kunci: Anastatus dasyni, Dasynus piperis, Riptortus linearis, Nezara viridula, parasitoid, pembiakan massal
Abstract
demographic statistics were only estimated for the parasitoid reared in R. linearis egg. The intrinsic rate of increase (r) was 0.1870 female/mother/days, the mean generation time (T) 27.51 days, the net reproductive rate (Ro) 84.29 female/ mother/generation, the finite rate of increase (λ) 1.21 female/mother/days, and the reproductive value (RVx) 402.51 individual. The stable stage distribution (px) was 17.06% egg, 50.41% larvae, 26.53% pupae, and 6.02% adult. R. linearis egg was the best alternative host to be used for mass rearing of A. dasyni.
Key words: Anastatus dasyni, Dasynus piperis, Riptortus linearis, Nezara viridula, parasitoid, mass rearing
Pendahuluan
Parasitoid Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae) memiliki potensi untuk dikembangkan sebagai agens pengendalian hayati hama kepik
pengisap buah lada Dasynus piperis China (Hemiptera: Coreidae), karena tingkat
parasitisasinya di lapangan dapat mencapai 84% (Deciyanto et al. 1993; Trisawa
et al. 2007). Pemanfaatan A. dasyni sebagai agens hayati yang efektif perlu diteliti dan dikembangkan lebih lanjut. Salah satunya adalah upaya
pengembangbiakan massal parasitoid ini pada inang alternatif. Hal ini penting
karena populasi parasitoid secara umum di lapangan sering terlambat
kehadirannya (van Driesche & Bellows 1996), sehingga perlu ditambah atau
dilepas dengan parasitoid hasil pembiakan di laboratorium. Pelepasan parasitoid
telah banyak dilakukan untuk mengendalikan hama tanaman dengan hasil yang
cukup memuaskan (Wright et al. 2001; Head & Walters 2002; Mills 2002).
Parasitoid A. dasyni diketahui memiliki beberapa inang selain telur D. piperis yaitu telur Riptortus linearis Fabr. (Hemiptera: Alydidae), Nezara
viridula Linn. (Hemiptera: Pentatomidae), dan Physomerus grossipes Fabr. (Hemiptera: Coreidae) (Kalshoven, 1981). Telur-telur tersebut dapat digunakan
sebagai inang alternatif untuk pembiakan A. dasyni. Namun demikian, dalam
pemilihan inang alternatif perlu dipertimbangkan kemudahan dalam pembiakan,
biaya murah, dan kesesuaian inang sehingga teknik pengendalian hayati yang
dikembangkan memang layak dilakukan (van Lenteren 1989; van Driesche &
Bellows 1996).
Kesesuaian inang alternatif dapat diukur berdasarkan beberapa ciri biologi,
di antaranya adalah reproduksi parasitoid. Reproduksi parasitoid yang tinggi
Reproduksi tersebut tidak hanya terbatas pada besarnya populasi, melainkan juga
terjadi dalam komposisi jenis kelamin dan kelompok umur.
Perubahan populasi parasitoid dapat disusun dalam neraca hayati. Dari
neraca hayati dapat dihitung beberapa statistik demografi (Birch 1948; Zeng et al.
1993; Carey 1993). Salah satu statistik demografi yang paling handal untuk
mengukur potensi musuh alami adalah laju pertambahan intrinsik (r), karena di
dalamnya telah mempertimbangkan karakteristik kehidupan serangga seperti masa
hidup, keperidian, sintasan, dan nisbah kelamin (Carey 1993). Nilai r merupakan
salah satu kriteria yang penting untuk mengevaluasi keefektifan atau potensi dari
agens pengendalian hayati (Lee & Ahn 2000), serta dapat digunakan untuk
menduga potensi pertumbuhan populasi parasitoid tersebut (Lysyk 2000).
Penelitian ini bertujuan mengkaji kesesuaian telur kepik kedelai R. linearis
dan N. viridula sebagai inang untuk pembiakan massal parasitoid A. dasyni. Kesesuaian diukur berdasarkan masa perkembangan pradewasa dan berbagai
karakteristik kehidupan imago betina A. dasyni serta neraca hayati yang meliputi reproduksi bersih (Ro), masa generasi (T), laju pertambahan intrinsik (r),
laju pertambahan terbatas (λ), nilai reproduksi (RVx) dan sebaran umur stabil
(px).
Bahan dan Metode
Penelitian dilakukan dari bulan Mei 2008 sampai dengan Februari 2009 di
laboratorium hama Balai Penelitian Tanaman Obat dan Aromatik Bogor, serta
rumah kaca Balai Pengkajian Teknologi Pertanian Kepulauan Bangka Belitung.
Pembiakan R. linearis dan N. viridula
Imago R. linearis dan N. viridula asal pertanaman kedelai masing-masing dipelihara dalam kurungan kain kasa yang berukuran panjang 35 cm, lebar 35
cm, dan tinggi 75 cm di laboratorium. Serangga diberi pakan kacang panjang
yang digantungkan pada kawat di bagian atas kurungan. Pakan diganti setiap 2
hari sekali. Untaian kain wol digantungkan di dalam kurungan sebagai tempat
Pemeliharaan D. piperis dan Pembiakan A. dasyni
Imago D. piperis asal pertanaman lada dipelihara pada bibit lada dalam pot
dan dikurung dengan plastik milar bergaris tengah 18 cm dan tinggi 40 cm serta
kurungan kayu berdinding kain kasa berukuran panjang 75 cm, lebar dan tinggi
masing-masing 50 cm. Kurungan ditempatkan di rumah kaca. Serangga diberi
pakan buah lada yang digantungkan pada kawat di bagian atas kurungan atau
dilekatkan pada bibit lada. Buah lada diganti setiap 2 hari sekali. Telur yang
diperoleh digunakan untuk pembiakan dan penelitian.
Telur D. p.iperis asal pertanaman lada dipelihara dalam tabung reaksi bergaris tengah 1.5 cm, panjang 18.0 cm di laboratorium. Tabung reaksi ditutup
dengan kapas yang dibungkus kain kasa. Parasitoid A. dasyni yang diperoleh
digunakan untuk pembiakan.
Sepasang A. dasyni yang baru keluar dari telur D. piperis dipelihara dalam
tabung reaksi bergaris tengah 1.5 cm, panjang 18.0 cm dan diberi pakan madu
10%. Setelah imago berumur 2 hari, ke dalam tabung reaksi dimasukkan 10 telur
D. piperis umur 2 hari yang diperoleh dari hasil pemeliharaan imago D. piperis. Telur D. piperis dilekatkan dengan lem kertas cair pada kertas karton (pias) ukuran 1.0 cm x 5.0 cm. Pias telur diambil setelah 24 jam dan diganti dengan pias
telur yang baru. Pias telur yang diambil dimasukkan ke dalam tabung reaksi
bergaris tengah 1.5 cm dan panjang 18.0 cm, kemudian diamati sampai parasitoid
keluar. Parasitoid A. dasyni yang diperoleh digunakan untuk penelitian.
Perkembangan Pradewasa Parasitoid pada Telur Kepik Kedelai
Setiap 10 butir telur R. linearis dan N. viridula umur dua hari, masing-masing secara terpisah dilekatkan dengan lem kertas cair pada pias. Pias telur
dimasukkan ke dalam tabung reaksi bergaris tengah 1.5 cm dan panjang 18.0 cm
yang berisi sepasang A. dasyni umur 2 hari asal telur D. piperis. Parasitoid diberi
pakan madu 10%. Tabung reaksi ditutup dengan kapas yang dibungkus kain kasa.
Pias telur dikeluarkan setelah 24 jam dan diganti dengan pias telur yang baru.
Pias telur yang diambil dimasukkan ke dalam tabung reaksi yang lain. Kegiatan
tersebut diulang pada 10 pasang parasitoid A. dasyni. Setiap hari sebanyak 20
telur yang diparasit dibedah untuk diperiksa perkembangan pradewasa A. dasyni.
perkembangan pradewasa dihitung dengan menjumlahkan umur fase telur, larva,
dan pupa.
Reproduksi Parasitoid pada Telur Kepik Kedelai
Telur R. linearis dan N. viridula umur 2 hari sebanyak 10 butir pada pias, secara terpisah dimasukkan ke dalam tabung reaksi bergaris tengah 1.5 cm dan
panjang 18.0 cm yang berisi sepasang A. dasyni umur 2 hari asal telur D. piperis.
Parasitoid diberi pakan madu 10%. Tabung reaksi ditutup dengan kapas yang
dibungkus kain kasa. Pias telur dikeluarkan setelah 24 jam dan diganti dengan
pias telur yang baru. Pias telur yang diambil dimasukkan ke dalam tabung reaksi
yang lain dan diamati sampai keluar imago A. dasyni.
Pada kegiatan ini, dari telur N. viridula tidak diperoleh imago A. dasyni
betina sehingga tidak dapat dilanjutkan terhadap sintasan imago betina A. dasyni.
Namun demikian untuk melihat perilaku peletakan telur, 20 imago betina yang
muncul dari telur R. linearis diberi perlakuan telur R. linearis dan 20 imago betina
yang lain diberi telur N. viridula.
Setiap satu pasang imago A. dasyni yang baru keluar dari telur
R. linearis dimasukkan ke dalam tabung reaksi bergaris tengah 1.5 cm dan panjang 18.0 cm serta diberi pakan madu 10%. Ke dalam tabung reaksi
kemudian dimasukkan pias berisi 10 telur masing-masing inang alternatif umur 2
hari. Tabung reaksi ditutup dengan kapas yang dibungkus kain kasa. Pias telur
dikeluarkan setelah 24 jam dan diganti dengan pias telur yang baru. Pias telur
yang diambil diamati setiap hari sampai imago A. dasyni keluar. Kegiatan tersebut dilakukan setiap hari sampai parasitoid betina mati (untuk penghitungan
masa hidup). Ditentukan pula masa praoviposisi, oviposisi, dan pasca oviposisi
berdasarkan imago A. dasyni yang muncul dari inang yang diparasit. Keperidian
dihitung dengan menjumlahkan semua telur yang diletakkan oleh seekor betina
selama hidupnya. Mengingat bahwa A. dasyni adalah parasitoid soliter, maka
jumlah imago yang keluar dari inang menunjukkan jumlah telur yang diletakkan.
Banyaknya imago yang muncul dan jenis kelaminnya dicatat untuk menentukan
nisbah kelamin. Nisbah kelamin dinyatakan dalam persentase betina. Telur inang
Analisis Data
Data perkembangan pradewasa dan karakteristik kehidupan imago
A. dasyni dianalisis menggunakan uji-t pada α = 0.05. Statistik demografi A. dasyni dihitung menurut Carey (1993) dan Lysyk (2000) sebagai berikut: Laju reproduksi bersih:
Ro = ∑ lxmx
Rataan masa generasi:
T = ∑xlxmx/∑lxmx
Laju pertambahan intrinsik:
r = Σ lxmxe-rx = 1
Laju pertambahan terbatas:
λ = er Nilai reproduksi:
RVx = (erx/lx) . (Σ e–ry lymy ), y = x
Sebaran umur stabil:
px =100β lx e-r(x+1) dan 1/β = Σ lx e-r(x+1), β = laju kelahiran terbatas
dengan x = kelas umur parasitoid betina, lx = proporsi parasitoid betina
yang hidup pada kisaran umur x, dan mx = jumlah keturunan betina pada
umur x.
Hasil dan Pembahasan
Perbandingan Parameter Hayati Parasitoid pada Dua Jenis Inang
Perbedaan jenis inang alternatif berpengaruh nyata terhadap masa
perkembangan larva dan pupa parasitoid. Dari perkembangan pradewasa pada
telur R. linearis diperoleh imago A. dasyni jantan dan betina, sedangkan dari telur N. viridula hanya jantan. Keturunan betina pada telur N. viridula diperoleh saat imago A. dasyni asal telur R. linearis dibiakkan pada telur N. viridula,
meskipun jumlahnya sangat sedikit yaitu hanya 1.81% dan berbeda nyata dengan
Tabel 3.1 Karakteristik perkembangan pradewasa dan kehidupan imago betina - Masa pasca oviposisi (hari) - Laju peneluran (butir/hari)
Perkembangan pradewasa A. dasyni berlangsung di dalam inang dan
waktu yang dibutuhkan untuk setiap fase menunjukkan kemampuan berkembang
dalam kondisi nutrisi inang yang tersedia. Waktu perkembangan larva dan pupa
yang berbeda dapat disebabkan oleh kandungan nutrisi dari ukuran inang yang
berbeda. Ukuran telur R. linearis lebih besar dibandingkan telur N. viridula, sehingga nutrisi yang tersedia pada telur R. linearis dapat dimaksimalkan oleh larva dan pupa untuk perkembangannya. Nutrisi inang memang menjadi salah
satu faktor yang mempengaruhi perkembangan pradewasa parasitoid (Vinson
1984; Beckage 1985; Godfray 1994). Kondisi ini juga yang menyebabkan waktu
perkembangan pradewasa yang menjadi imago A. dasyni pada kedua inang
alternatif tersebut berbeda.
Dari kedua jenis inang alternatif, secara umum imago jantan muncul satu
hari lebih cepat dibandingkan betina. Kemunculan jantan yang lebih awal
tersebut diduga berkaitan dengan strategi perkawinan. Jantan yang sudah siap
kawin akan menunggu betina muncul kemudian terjadi kopulasi. Betina hanya
melakukan 1 kali kopulasi selama hidupnya. Betina yang sudah berkopulasi akan
menolak kehadiran jantan. Jika betina A. dasyni yang muncul lebih dahulu dan
tidak menemukan jantan, maka betina akan cenderung menolak kopulasi terutama
A. dasyni karena telur yang tidak dibuahi (tidak terjadi kopulasi) berkembang menjadi jantan, sedangkan yang dibuahi menjadi imago betina. Reproduksi
parasitoid seperti ini (van Driesche & Bellows 1996) termasuk ke dalam tipe
arenotoki. Oleh karena itu, kemunculan jantan lebih awal merupakan salah satu
strategi yang penting untuk berlangsungnya kopulasi (Godfray 1994; Colazza &
Wajnberg 1998).
Betina A. dasyni yang muncul dari telur R. linearis memiliki kemampuan
reproduksi yang tidak berbeda pada kedua jenis inang alternatif. Perbedaan hanya
terjadi pada alokasi jenis kelamin keturunan. Pada telur R. linearis, betina
A. dasyni akan meletakkan 70.20% keturunannya berkelamin betina, sedangkan
pada telur N. viridula hampir 100% jantan. Di samping berpengaruh terhadap
perkembangan pradewasa, ukuran inang juga menjadi faktor yang mempengaruhi
perilaku peletakan telur parasitoid. Parasitoid A. dasyni akan meletakkan keturunan jantan sebanyak-banyaknya pada inang yang berukuran kecil karena
inang dinilai kurang cocok untuk perkembangan betina. Proporsi keturunan
berkelamin jantan akan lebih banyak dibandingkan betina pada inang yang lebih
kecil (Gauld & Fitton 1987; Heinz & Parella 1990; Joyce et al. 2002).
Nisbah kelamin A. dasyni yang diperoleh pada dua jenis inang alternatif,
juga sesuai dengan teori Charnov et al. (1981) yang memprediksi bahwa
parasitoid betina akan meletakkan jantan pada inang yang kecil dan betina pada
inang yang lebih besar. Namun demikian, teori ini tidak mutlak karena parasitoid
dapat menghasilkan keturunan berkelamin jantan dan betina, baik pada inang yang
berukuran besar maupun kecil. Hanya saja terjadi perbedaan di dalam nisbah
kelamin. Hal ini terbukti dari kemunculan betina A. dasyni pada telur
N. viridula. Kemunculan betina tersebut kemungkinan terjadi pada telur N. viridula yang memiliki ukuran lebih besar di antara telur yang disediakan. Telur N. viridula memiliki ukuran panjang berkisar antara 0.98 mm sampai 0.99
mm dan lebar antara 0.81 mm sampai 0.83 mm.
Keturunan jantan yang lebih banyak dalam pembiakan massal parasitoid
tidak dikehendaki karena kegiatan pemarasitan inang hanya dilakukan oleh
parasitoid betina (Heinz 1998). Hasil penelitian memperlihatkan bahwa parasitoid
adalah betina. Dengan demikian inang alternatif tersebut sesuai untuk pembiakan
massal A. dasyni.
Imago betina A. dasyni asal telur R. linearis yang diberi inang berbeda
tidak menunjukkan perbedaan yang nyata terhadap lama hidup. Lama hidup,
sebagaimana yang dikemukakan oleh Trisawa et al. (2007) dipengaruhi oleh
ketersediaan pakan. Madu merupakan faktor penting bagi kelangsungan hidup
dan reproduksi A. dasyni. Keberadaan inang hanya dapat memberikan kesempatan
kepada imago betina A. dasyni untuk menunjukkan potensi bertelurnya.
Masa praoviposisi, oviposisi, dan pasca oviposisi relatif lebih cepat pada
betina A. dasyni yang diberi telur N. viridula dibandingkan yang diberi telur
R. linearis. Hal ini dapat disebabkan oleh pengaruh spesies inang yang secara tidak langsung mempengaruhi fisiologi parasitoid (van Alphen & Jervis 1997;
Corteserro et al. 2000).
Satu ekor betina A. dasyni mampu menghasilkan keturunan rata-rata
sebesar 122.72 ± 11.43 ekor pada telur R. linearis, dan 129.85 ± 15.15 ekor pada
telur N. viridula. Data tersebut berdasarkan jumlah imago parasitoid baik yang keluar maupun yang gagal keluar dari setiap telur inang yang diparasit. Parasitoid
yang gagal keluar sangat rendah, hanya 0.52% pada telur R. linearis dan 1.13%
pada telur N. viridula. Mengingat bahwa A. dasyni merupakan parasitoid soliter,
maka jumlah keturunan tersebut menunjukkan jumlah telur yang diletakkan
(keperidian). Keperidian ini menurut Trisawa et al. (2007) berada pada kisaran keperidian A. dasyni pada inang alami yaitu antara 60 sampai 136 butir.
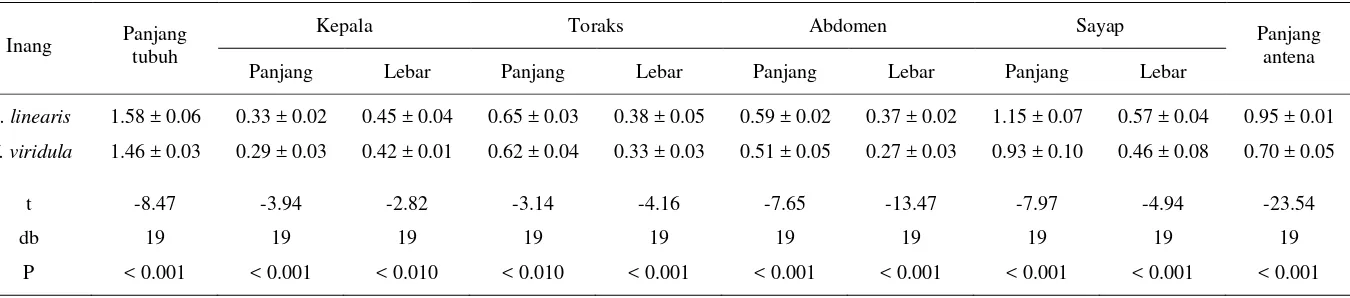
Perbandingan Berbagai Dimensi Tubuh Parasitoid
Ukuran tubuh parasitoid betina dan jantan meningkat seiring
meningkatnya ukuran inang. Di samping itu, ukuran tubuh parasitoid betina
selalu lebih besar dibandingkan jantan pada kedua jenis inang alternatif.
Pengaruh jenis inang alternatif terhadap ukuran parasitoid menunjukkan bahwa
semua parameter ukuran parastoid betina asal telur R. linearis lebih panjang dan
lebih lebar dibandingkan betina asal telur N. viridula. Demikian pula dengan
parasitoid jantan yang berkembang pada telur R. linearis juga memiliki tubuh
yang lebih panjang dan lebar dibandingkan jantan yang berkembang pada inang
Tabel 3.2 Dimensi tubuh A. dasyni betina (x ± SD mm) yang dipelihara pada telur kepik kedelai
Inang Panjang
tubuh
Kepala Toraks Abdomen Sayap Panjang
antena
Panjang Lebar Panjang Lebar Panjang Lebar Panjang Lebar
R. linearis 2.23 ± 0.06 0.46 ± 0.04 0.62 ± 0.02 0.89 ± 0.07 0.53 ± 0.03 0.83 ± 0.02 0.50 ± 0.02 1.36 ± 0.04 0.49 ± 0.04 1.14 ± 0.02 N. viridula 1.73 ± 0.10 0.33 ± 0.02 0.50 ± 0.04 0.68 ± 0.07 0.40 ± 0.04 0.58 ± 0.08 0.35± 0.07 1.24 ± 0.05 0.40 ± 0.04 0.84 ± 0.06
t -19.58 -7.02 -11.02 -9.53 -12.03 -14.46 -8.59 -8.45 -7.70 -20.78
db 19 19 19 19 19 19 19 19 19 19
P < 0.001 < 0.001 < 0.001 < 0.001 < 0.001 < 0.001 < 0.001 < 0.001 < 0.001 < 0.001
Tabel 3.3 Dimensi tubuh A. dasyni jantan (x ± SD mm) yang dipelihara pada telur kepik kedelai
Inang Panjang tubuh
Kepala Toraks Abdomen Sayap Panjang
antena
Panjang Lebar Panjang Lebar Panjang Lebar Panjang Lebar
R. linearis 1.58 ± 0.06 0.33 ± 0.02 0.45 ± 0.04 0.65 ± 0.03 0.38 ± 0.05 0.59 ± 0.02 0.37 ± 0.02 1.15 ± 0.07 0.57 ± 0.04 0.95 ± 0.01 N. viridula 1.46 ± 0.03 0.29 ± 0.03 0.42 ± 0.01 0.62 ± 0.04 0.33 ± 0.03 0.51 ± 0.05 0.27 ± 0.03 0.93 ± 0.10 0.46 ± 0.08 0.70 ± 0.05
t -8.47 -3.94 -2.82 -3.14 -4.16 -7.65 -13.47 -7.97 -4.94 -23.54
db 19 19 19 19 19 19 19 19 19 19
Gambar 3.1 Perbandingan ukuran parasitoid A. dasyni betina dan jantan asal inang alternatif. Kiri: parasitoid asal telur N. viridula, kanan:
parasitoid asal telur R. linearis
Ukuran tubuh A. dasyni yang meningkat dapat dipengaruhi oleh sumber
energi yang dibawa saat fase larva sehingga dewasa yang besar akan memiliki
cadangan energi. Kondisi ini dapat menyebabkan imago betina memiliki peluang
hidup lebih lama dengan keperidian yang lebih tinggi. Betina yang berukuran
lebih kecil, kurang baik jika digunakan sebagai induk dalam pembiakan massal.
Ukuran tubuh betina yang kecil akan memiliki kebugaran yang rendah yang
berdampak pada rendahnya produksi telur (Rosenheim & Rosen 1991; Godfray
1994; van Alphen & Jervis 1997).
Sintasan dan Neraca Hayati A. dasyni pada Telur R. linearis
Sintasan dan neraca hayati parasitoid yang ditampilkan adalah A. dasyni
asal inang alami yang dibiakkan pada telur R. linearis karena dari sintasan
pradewasa pada telur N. viridula sebelumnya tidak menghasilkan imago betina.
Keberhasilan hidup setiap fase pradewasa mulai dari telur sampai pupa dapat
diukur dari imago yang berhasil keluar dari telur inang. Dengan demikian
pengamatan menunjukkan bahwa 100% telur R. linearis yang diparasit berhasil
mengeluarkan imago parasitoid.
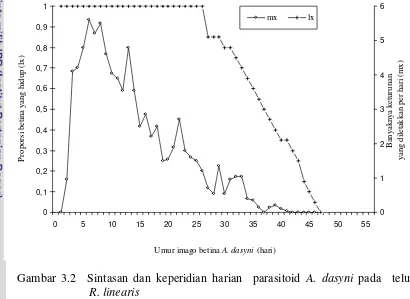
Kesintasan imago betina A. dasyni pada inang R. linearis menunjukkan
bahwa proporsi imago yang masih hidup (lx) menurun mulai hari ke 27. Jumlah
keturunan betina yang diletakkan setiap hari (mx
0
Umur imago betina A. dasyni (hari)
Ba
) berfluktuasi. Puncak peneluran
A. dasyni terjadi pada individu yang berumur 21 hari atau imago betina yang berumur 6 hari (Gambar 3.2).
Gambar 3.2 Sintasan dan keperidian harian parasitoid A. dasyni pada telur R. linearis
Peneluran menurun seiring dengan meningkatnya umur betina parasitoid.
Hal tersebut berhubungan dengan umur fisiologis parasitoid. Semakin tua umur
parasitoid, semakin menurun keperidiannya. Parasitoid umumnya tidak
meletakkan telur antara 3 - 14 hari sebelum mati. Kematian yang terjadi pada
parasitoid umur tua menyebabkan kurva peluang hidup A. dasyni yang terbentuk
memperlihatkan pola tipe I sebagaimana yang dikemukakan oleh Price (1997).
Pola seperti ini terjadi pada serangga yang dipelihara di laboratorium karena
Neraca hayati A. dasyni yang dibiakkan pada telur R. linearis disajikan
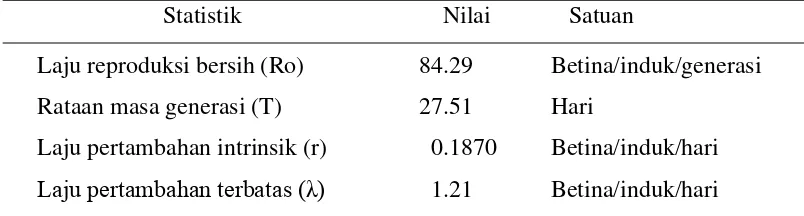
pada Tabel 3.4. Nilai laju reproduksi bersih (Ro) menggambarkan rata-rata
jumlah keturunan betina yang dihasilkan oleh seekor induk parasitoid. Dengan
demikian, maka seekor imago betina A. dasyni dapat menghasilkan 84.29
betina/induk/generasi atau terjadi kelipatan populasi A. dasyni 84.29 kali dalam
setiap generasi. Parasitoid A. dasyni yang dibiakkan pada telur R. linearis memiliki rata-rata waktu satu generasi (T) 27.51 hari. Waktu tersebut
menunjukkan waktu yang dibutuhkan sejak telur diletakkan sampai imago yang
berasal dari telur tersebut menghasilkan separuh keturunannya. Nilai laju
pertambahan intrinsik (r) adalah 0.1870 betina/induk/hari dengan nilai laju
pertambahan terbatas (λ) adalah 1.21 betina/induk/hari. Nilai ini menunjukkan
besarnya kelipatan populasi A. dasyni per hari.
Tabel 3.4 Berbagai statistik neraca hayati A. dasyni
Statistik Nilai Satuan
Laju reproduksi bersih (Ro) 84.29 Betina/induk/generasi
Rataan masa generasi (T) 27.51 Hari
Laju pertambahan intrinsik (r) 0.1870 Betina/induk/hari
Laju pertambahan terbatas (λ) 1.21 Betina/induk/hari
Hasil perhitungan nilai reproduksi (RVx) parasitoid A. dasyni pada telur
R. linearis 402.51 individu. Nilai reproduksi meningkat pada awal reproduksi sampai puncaknya pada individu parasitoid yang berumur 19 hari atau imago
umur 4 hari, kemudian menurun sesuai dengan bertambahnya umur parasitoid
(Gambar 3.3).
Nilai reproduksi memberikan gambaran tentang sumbangan relatif suatu
individu pada umur tertentu terhadap populasi. Nilai reproduksi tersebut biasanya
meningkat pada awal reproduksi kemudian menurun seiring bertambahnya umur
serangga (Nakamura 1984). Nilai reproduksi juga dapat digunakan sebagai acuan
dalam pelepasan imago parasitoid hasil pembiakan ke lapangan. Acuan tersebut
berdasar pada umur imago parasitoid yang memiliki nilai reproduksi tertinggi.
Gambar 3.3 Nilai reproduksi A. dasyni menurut umur pada telur R. linearis
Hasil perhitungan untuk sebaran umur stabil (px) menunjukkan bahwa
fase larva A. dasyni memiliki proporsi yang lebih tinggi (50.41%) dibandingkan
fase lainnya (Tabel 3.5). Hasil ini menunjukkan bahwa sebagian besar anggota
populasi A. dasyni merupakan serangga pradewasa. Menurut Price (1997) populasi yang sebagian besar tersusun dari individu-individu yang berumur muda
akan tumbuh dengan cepat.
Tabel 3.5 Proporsi fase perkembangan A. dasyni pada sebaran stabil
Fase Proporsi (%)
Telur 17.06
Larva 50.41
Pupa 26.53
Imago 6.02
Kesimpulan
Parasitoid A. dasyni yang dibiakkan pada telur N. viridula memperlihatkan
masa perkembangan larva dan pupa yang lebih singkat serta laju peneluran yang
lebih tinggi dibandingkan pada telur R. linearis. Namun demikian, hampir
seluruh keturunannya adalah jantan. Oleh karena itu, untuk keperluan pembiakan
parasitoid yang dihasilkan dari telur R. linearis sebagian besar adalah betina,
dengan laju pertambahan intrinsik 0.1870, rataan masa generasi (T) 27.51 hari,
dan populasinya meningkat 84.29 kali lipat per generasi.
Daftar Pustaka
Beckage NE. 1985. Endocrine interactions between endoparasitic insect and their host. Ann Rev Entomol 30:371-413.
Birch LC. 1948. The intrinsic rate of natural increase of an insect population. J Anim Ecol 17:15-26.
Carey JR. 1993. Applied Demography for Biologist with Special Emphasis on Insects. New York. Oxford Univ Press.
Charnov EL, Los-den Hartogh RL, Jones WT, van den Assem J. 1981. Sex ratio evolution in a variable environment. Nature 289:27-33.
Colazza S, Wajnberg E. 1998. Effect of host egg mass size on sex ratio and oviposition sequence of Trissolcus basalis (Hymenoptera: Scelionidae). Pop Ecol 27:329-336.
Corteserro AM, Stapel JO, Lewis WJ. 2000. Understanding and manipulating plant attributes to enhance biological control. Biol Control 17:35-49.
Deciyanto S, Trisawa IM, Muchyadi. 1993. Parasitism fluctuation of egg-parasitoids of pepper bug (Dasynus piperis China) in Bangka. J Spice Medic Crops 1(2):33-36.
Gauld ID, Fitton MG. 1987. Sexual dimorphism in the Ichneumonidae: a response to hurlbutt. Biol J Linn Soc 31:291-300.
Godfray HCJ. 1994. Parasitoids: Behavioral and Evolutionary Ecology. New Jersey: Princenton Univ Press.
Head J, Walters KFA. 2002. Augmentation biological control using the entomopathogenic nematode Steinernema feltiae against the south american leafminer Liriomyza huidobrensis. International Symposium on Biological Control of Arthropods; Honolulu, January 14-18, 2002. West Virginia: Forest Health Technology Enterprise Team. hlm 136-140.
Heinz KM. 1998. Host size-dependent sex allocation behavior in a parasitoid: implications for Catolaccus grandis (Hymenoptera: Pteromalidae) mass rearing program. Bull Entomol Res 88:37-45.
Heinz KM, Parella MP. 1990. The influence of host size on sex ratios in the parasitoid Diglyphus begini (Hymenoptera: Eulophidae). Ecol Entomol 15: 391-399.
Kalshoven LGE. 1981. Pests of Crops in Indonesia. van der Laan PA, penerjemah. Jakarta: Ichtiar Baru-Van Hoeve. Terjemahan dari: De Plagen van de Cultuurgewassen in Indonesie.
Lee, JH, Ahn JJ. 2000. Temperature effects on development, fecundity, and life table parameters of Amblyseius womersleyi (Acari: Phytoseiidae). Environ Entomol 29(2):265-271.
Lysyk TJ. 2000. Relationship between temperature and life history parameters of Muscidifurax raptor (Hymenoptera: Pteromalidae). Environ Entomol 29(3):596-605.
Mills NJ. 2002. Augmentation in orchards: improving the efficacy of Trichogramma inundation. International Symposium on Biological Control of Arthropods; Honolulu, January 14-18, 2002. West Virginia: Forest Health Technology Enterprise Team. hlm 130-135.
Nakamura, N. 1984. Survivorship and fertility schedules of two Epilachnine ”spesies” feeding on cucurbitaceous plant under laboratory conditions (Coleoptera: Coccinellidae). Appl Ent Zool 19(1):59-66.
Price PW. 1997. Insect Ecology.3rd. New York: John Wiley & Sons.
Rosenheim JA, Rosen D. 1991. Foraging and oviposition decisions in the parasitoid Aphytis lingnanensis: Distinguishing the influenches of egg load and experience. J Animal Ecol 60:873-894.
Trisawa IM, Rauf A, Kartosuwondo U. 2007. Biologi parasitoid Anastatus dasyni Ferr. (Hymenoptera: Eupelmidae) pada telur Dasynus piperis China (Hemiptera: Coreidae). Hayati 14(3):81-86.
van Alphen JJM, Jervis MA. 1997. Foraging behaviour. Di dalam: Jervis M, Kidd N, editor. Insect Natural Enemies. Practical Approaches to Their Study and Evaluation. London: Chapman & Hall. hlm 1-62.
van Driesche RG, Bellows JTS. 1996. Biological Control. New York: Chapman & Hall.
van Lenteren JC. 1989. Parasitoids in the greenhouse: successes with seasonal inoculative release systems. Di dalam: Waage J, Greathead D, editor. Insect Parasitoids. London : Academic Press. hlm 341-374.
Vinson SB. 1984. Parasitoid-host relationship. Di dalam: William JB, Carde RT, editor. Chemical Ecology of Insect. London: Chapman & Hall Ltd. hlm 205-225.
Wright MG, Hoffmann MP, Chenus SA, Gardner J. 2001. Dispersal behavior of Trichogramma ostriniae (Hymenoptera: Trichogrammatidae) in sweet corn fields: implications for augmentative releases against Ostrinia nubilalis (Lepidoptera: Crambidae). Biol Control 22:29-37.