DEPRESI PENANGKARANDALAM EMPAT GENERASI
PENYERBUKAN TERTUTUP TANAMAN KELAPA DALAM
MAPANGET No.32 BERDASARKAN SIFAT MORFOLOGI
DAN PENANDA MIKROSATELIT
Donata Suriaty Pandin
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi yang berjudul Depresi Penangkarandalam Empat Generasi Penyerbukan Tertutup Tanaman Kelapa Dalam Mapanget No.32 Berdasarkan Sifat Morfologi dan Penanda Mikrosatelit adalah karya saya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi manapun. Sumber informasi yang berasal dan dikutip dari karya yang diterbitkan mau pun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Februari 2009
ABSTRACT
DONATA SURIATY PANDIN. Inbreeding Depression of Four Generations Close-Pollinated of Mapanget Tall Coconut Trees No.32 based on morphological characters and microsatellite markers. Supervised by ALEX HARTANA, HAJRIAL ASWIDINNOOR, and ASEP SETIAWAN.
Coconuts (Cocos nucifera L) are one of the major plantation crops in the tropics, where their products play an important economical role in many developping country. The objectives of this study were to determine inbreeding depression, parentage analysis, and to identify DNA specific fragment related to coconut fruits production based on morphological and microsatellite (SSR) markers of second, third, and fourth generation close pollinated of Mapanget Tall coconut trees No.32 (DMT-32). The study was conducted using 30, 9, 40, and 38 trees of open-pollinated Mapanget Tall coconut (DMT OP), second, third, and fourth generations of close pollinated DMT-32 (DMT-32 S2, DMT-32 S3, and DMT-32 S4 respectively. Inbreeding depression expressed in both of two markers, morphological and microsatellite (SSR). Inbreeding depression occured in almost of all characters studied and have tendency to increase from generation to the next generation of DMT-32 S2, DMT-32 S3 and DMT-32 S4. Inbreeding depression was also assessed by SSR markers and show the same pattern with morphological marker, increase from generation to the next generation of DMT-32. Fifteen SSR primers used were able to analyze the tree parents of DMT-32 S4 from DMT-32 S3, and DMT-32 S3 from DMT-32 S2. All of 9 coconut trees of DMT-32 S2 analyzed are the parents of DMT-32 S3, but some of those coconut palms of DMT-32 S3 are not the parents of DMT-32 S4. Each SSR locus spreads in 15 chromosomes. Two of 15 primers used, CNZ21 and CNZ51, were able to show the specific DNA fragment related to fruitless of coconut. There were indication that CNZ21-270bp and CNZ51-110bp fragments associate with coconut fruit production.
RINGKASAN
DONATA SURIATY PANDIN. Depresi Penangkarandalam Empat Generasi Penyerbukan Tertutup Tanaman Kelapa Dalam Mapanget No.32 Berdasarkan Sifat Morfologi dan Penanda Mikrosatelit. Di bawah bimbingan ALEX HARTANA, HAJRIAL ASWIDINNOOR, dan ASEP SETIAWAN.
Indonesia memiliki keanekaragaman kelapa yang sangat tinggi, dengan luas pertanaman tertinggi di dunia, tetapi rata-rata produksi kelapa per hektar per tahun masih sangat rendah. Pengetahuan tentang karakter-karakter yang bernilai ekonomi tinggi serta identifikasi dini pohon-pohon yang berpotensi produksi tinggi sangat penting, mengingat umur mulai berbuah kelapa yang lama. Penelitian untuk mendapatkan karakter-karakter yang homozigot telah dilakukan menggunakan serbuksari campuran dari pohon-pohon terpilih dari setiap ge nerasi kelapa DMT-32 sampai pada generasi keempat. Penyerbukan antar pohon-pohon tertentu dalam generasi yang sama ini mengakibatkan terjadinya depresi penangkarandalam, dan karena serbuksari yang digunakan adalah campuran dari pohon-pohon terpilih sehingga identitas dari masing- masing pohon pada setiap generasi tidak jelas.
jumlah daun. Depresi penangkarandalam pada karakter generatif memiliki kecenderungan yang sama dengan karakter vegetatif yaitu semakin banyak karakter yang mengalami depresi penangkarandalam dengan persentase semakin meningkat dari generasi ke generasi hasil penyerbukan tertutup pohon kelapa menggunakan campuran polen dari pohon kelapa generasi yang sama. Pada generasi DMT-32 S2 ada 3 karakter yang tidak mengalami depresi penangkarandalam, sedangkan pada DMT-32 S3 tinggal 2 yang tidak mengalami depresi penangkarandalam. Pada DMT-32 S4 semua karakter generatif telah mengalami depresi penangkarandalam yang persentasenya semakin meningkat. Pada sifat komponen buah, penyerbukan tertutup tidak selalu menyebabkan depresi penangkarandalam pada generasi kedua, ketiga, dan keempat hasil penyerbukan tertutup kelapa DMT-32. Pada generasi kedua hanya satu komponen buah yang tidak mengalami depresi yaitu sabut buah. Sedangkan pada generasi ketiga air kelapa dan tempurung kelapa bertambah berat. Pada generasi keempat semua komponen buah mengalami depresi penangkarandalam. Hasil penelitian ini juga memperlihatkan bahwa persentase depresi penangkarandalam untuk komponen buah pada DMT-32 S3 lebih rendah kecuali pada berat sabut dibandingkan dengan DMT-32 S2 dan meningkat lagi pada DMT-32 S4. Secara umum depresi penangkarandalam meningkat persentasenya dari DMT-32 S2 ke DMT-32 S4.
kelapa zuriat dari DMT-32 S2 No.3 menjadi tetua jantan dan betina dari 18 individu DMT-32 S4. DMT-32 S2 No.4 memiliki 5 zuriat yaitu sebagai tetua jantan dari 4 individu dan tetua betina dari satu individu. Dua nomor pohon diantaranya menjadi tetua jantan dan betina dari 6 individu DMT-32 S4. DMT-32 S2 No.5 merupakan tetua jantan dari 6 individu DMT-32 S3 dan tetua betina dari 4 individu DMT-32 S3. Enam nomor pohon diantaranya menjadi tetua jantan dan betina dari 24 individu DMT-32 S4. Pohon kelapa DMT-32 S4 No.8 dan No.22 merupakan hasil zigot dengan polen dari pohon yang sama yaitu DMT-32 S3 No.28. DMT-32 S2 No.6 adalah tetua jantan dari 3 individu DMT-32 S3 dan tetua betina dari satu individu DMT-32 S3. Hanya pohon DMT-32 S3 No.35 yang menjadi tetua dari 4 individu DMT-32 S4, yang satu individu merupakan hasil zigot dengan polen dari pohon tersebut yaitu DMT-32 S4 No.30. Pohon kelapa DMT-32 S2 No.7 merupakan tetua jantan dari merupakan tetua betina dan jantan dari masing- masing 5 individu DMT-32 S3. Lima diantaranya merupakan tetua jantan dan betina dari 20 individu DMT-32 S4. Pohon kelapa DMT-32 S4 No.11 merupakan hasil zigot dengan polen dari pohon DMT-32 S3 No.32. Pohon kelapa DMT-32 S2 No.8 adalah tetua jantan dari 5 individu DMT-32 S3, tetua betina dari 6 individu DMT-32 S3, dan tetua betina sekaligus tetua jantan dari satu individu yaitu DMT-32 S3 No.25. Empat nomor pohon DMT-32 S3 diantaranya adalah tetua jantan dan betina dari 15 individu 32 S4. Pohon-pohon kelapa DMT-32 S4 No.8, No.10, dan No.22 merupakan zigot dengan polen dari pohon yang sama yaitu DMT-32 S3 No.28. Pohon kelapa DMT-32 S2 No.9 merupakan tetua jantan dari 6 individu DMT-32 S3 dan tetua betina dari satu individu DMT-32 S3. Empat nomor dari zuriat DMT-32 S2 No.9 adalah tetua jantan dan betina dari 17 individu DMT-32 S4.
Penelitian untuk mendeteksi adanya pita atau fragmen DNA spesifik yang berkaitan dengan kemampuan berbuah pada tanaman kelapa menggunakan populasi kelapa DMT No.32 hasil penyerbukan tertutup generasi kedua (DMT-32 S2), generasi ketiga (DMT-32 S3), dan generasi keempat (DMT-32 S4). Sebanyak 15 lokus mikrosatelit (SSR) kelapa digunakan untuk mendeteksi adanya fragmen spesifik. Untuk mengetahui hubungan antar lokus yang dipakai dalam penelitian ini dan korelasi antara sifat morfologi dengan penanda molekular dilakukan analisis korelasi Pearson dengan Program komputer Minitab 14. Peta keterpautan (linkage map) menggunakan MAPMAKER dengan nilai LOD 3. Hasil penelitian menunjukkan bahwa tidak ada hubungan antar lokus dari 15 lokus SSR yang digunakan, dan 15 lokus SSR tersebut tersebar dalam 15 kelompok sehingga diduga 15 lokus tersebut berada pada 15 kromosom berbeda. Kemampuan produksi buah pada tanaman kelapa DMT-32 kemungkinan berhubungan dengan ada tidaknya pita DNA yang hilang pada CNZ21-270bp dan CNZ51-110b.
© Hak cipta milik IPB, tahun 2009 Hak cipta dilindungi
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB 2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya
DEPRESI PENANGKARANDALAM EMPAT GENERASI
PENYERBUKAN TERTUTUP TANAMAN KELAPA DALAM
MAPANGET No.32 BERDASARKAN SIFAT MORFOLOGI
DAN PENANDA MIKROSATELIT
Donata Suriaty Pandin
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Departemen Agronomi dan Hortikultura
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Judul Disertasi : Depresi Penangkarandalam Empat Generasi Penyerbukan Tertutup Tanaman Kelapa Dalam Mapanget No.32 Berdasarkan Sifat Morfologi dan Penanda Mikrosatelit.
Nama : Donata Suriaty Pandin
NIM : P036.00008
Disetujui Komisi Pembimbing
Prof. Dr. Ir. Alex Hartana, MSc Ketua
Dr. Ir. Hajrial Aswidinnoor, MSc Dr. Ir. Asep Setiawan, MS Anggota Anggota
Diketahui
Ketua Program Studi Agronomi Dekan Sekolah Pascasarjana
Dr.Ir. Munif Ghulamahdi, MS Prof. Dr.Ir. Khairil Anwar Notodiputro, MS
Penguji Luar Komisi pada Ujian Tertutup
Dr. Ir. Ade Wachyar, MS
Pengajar pada Departemen Agronomi dan Hortikultura Fakultas Pertanian, IPB
Penguji Luar Komisi pada Ujian Terbuka
Prof. Dr. Ir. Sudirman Yahya, MSc
Guru besar Departemen Agronomi dan Hortikultura Fakultas Pertanian, IPB
Dr. Ir. Hengki Novarianto, MS
Ahli Peneliti pada Balai Penelitian Tanaman Kelapa dan Palma Lain Pusat Penelitian dan Pengembangan Tanaman Industri
PRAKATA
Depresi Penangkarandalam Empat Generasi Penyerbukan Tertutup
Tanaman Kelapa Dalam Mapanget No.32 Berdasarkan Sifat Morfologi dan
Penanda Mikrosatelit merupakan judul disertasi yang dibuat penulis sebagai salah
satu syarat untuk memperoleh gelar doktor pada Departemen Agronomi dan
Hortikultura, Program Pascasarjana Institut Pertanian Bogor.
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Pengasih dan
Pemurah atas segala anugerahNya sehingga karya ilmiah ini dapat diselesaikan.
Penghargaan, terima kasih, dan rasa hormat yang dalam saya sampaikan kepada
komisi pembimbing, Prof. Dr. Ir. Alex Hartana, MSc, Dr Ir. Hajrial Aswidinnoor,
MSc, dan Dr. Ir. Asep Setiawan MS, atas waktu, perhatian dan arahan-arahan
yang diberikan sehingga karya ilmiah ini berhasil diselesaikan.
Terima kasih dan penghargaan juga disampaikan kepada Dr.Ir. Ade
Wachyar MS, Prof. Dr. Ir. Sudirman Yahya MSc, dan Dr.Ir. Hengki Novarianto
MS, sebagai penguji luar komisi pada ujian tertutup dan terbuka penulis. Masukan
yang diberikan telah memperkaya dan menajamkan pembahasan dalam tulisan ini.
Terima kasih dan penghargaan disampaikan kepada Kepala Badan Litbang
Pertanian, Kepala Pusat Penelitian dan Pengembangan Tanaman Perkebunan dan
kepala Balai Peneltian Tanaman Kelapa dan Palma Lain, yang telah memberikan
kesempatan pada penulis untuk melanjutkan pendidikan.
Terima kasih dan penghargaan juga disampaikan kepada Pimpinan Institut
Pertanian Bogor dan Sekolah Pascasarjana, yang telah menerima penulis untuk
belajar dan menyelesaikan studi di Lembaga ini.
Kepada Komisi Pembinaan Tenaga Badan Penelitian dan Pengembangan
Pertanian Departemen Pertanian melalui Proyek PAATP Badan Litbang Pertanian
atas beasiswa dan bantuan dana penelitian yang telah diberikan, disampaikan
terima kasih.
Ucapan terima kasih penulis sampaikan kepada Kepala Laboratorium
Biologi Tumbuhan dan Kepala Pusat Penelitian Sumberdaya Hayati dan
Ucapan terima kasih disampaikan pula kepada Dr.Ir. David Allorerung,
Dr.Ir. H Novarianto, Ir. HT Luntungan MSc, Ir. E Tenda MS, Ir. H Tampake MS,
Ir. Ismail Maskromo MSi, Ir Miftahorrachman, Ir. J Kumaunang MSc, serta
rekan-rekan peneliti dan staf lainnya di Balitka Manado atas bantuan dan
dorongan moril yang diberikan.
Kepada Dr. Ir. Ni Made Armini Wiendi MS, Dr.Ir. Endah R Palupi MSc,
Dr.Ir. Trikoesoemaningtyas MSc, Prof. Dr. Ir. Sudarsono MSc, Dr.Ir Dirvamena
Boer, Dr. Fitmawati MSi, Dr. Soaloon Sinaga MSi, Dr. Amin Retnoningsih MSi,
Pak Sutiyo serta rekan-rekan lain di Lab Biologi Tumbuhan dan Bioteknologi
PPSHB yang tidak sempat penulis tuliskan satu per satu diucapkan terima kasih
atas bantuan dan kerjasama yang baik selama ini.
Kepada rekan-rekan mahasiswa Sulut khususnya Hasnawaty P, Alfred L
Susana R, Anderson K, Hengky S, Jack M, Erly K, Ben J T, Emma M, Agung W
dan Meiliana K tak lupa diucapkan terima kasih.
Terima kasih kepada kedua orang tua penulis S Pandin (alm) dan LR
Tandirerung (almh) untuk perhatian berlimpah dan kasih yang tulus yang
diberikan kepada penulis selama hidup mereka. Kepada kakak-kakak dan
adik-adik serta segenap keluarga yang senantiasa mendukung dalam doa dan dana,
dengan hati yang tulus penulis menyampaikan banyak terima kasih. Tanpa
dukungan keluarga penulis tidak akan mungkin dapat menyelesaikan disertasi ini.
Akhirnya dengan segala kekurangan yang ditemukan, penulis berharap
semoga tulisan ini dapat memberikan informasi dan bermanfaat bagi
pembangunan dan pengembangan pertanian di Indonesia.
Bogor, Februari 2009.
RIWAYAT HIDUP
Penulis dilahirkan di Palopo pada tangal 22 Agustus 1955 sebagai anak
ketujuh dari pasangan Sampe Pandin (Alm) dan Ibu L.R. Tandirerung (Almh).
Pendidikan sarjana ditempuh pada Jurusan Teknik Pertanian, Fakultas Pertanian
Universitas Hasanuddin Makassar dan lulus tahun 1986. Pendidikan Master
ditempuh pada Program Studi Agronomi Sekolah Pascasarjana IPB, Institut
Pertanaian Bogor dan tamat tahun 2000. Sejak tahun 1986 penulis telah bekerja
sebagai staf peneliti pada Balai Penelitian Tanaman Kelapa dan Palma Lain,
Manado.
Pada tahun 2000 penulis diberi kesempatan mengikuti program doktor pada
DAFTAR ISI
Halaman
DAFTAR TABEL ……….... viii
DAFTAR GAMBAR ……… ix
I. PENDAHULUAN ………. 1
Latar Belakang ... 1
Tujuan Penelitian ... 6
II. TINJAUAN PUSTAKA ………. 7
Asal dan Keanekaragaman Kelapa ………... 7
Pemuliaan Tanaman Kelapa di Indonesia ... 10
Depresi Penangkarandalam dan Heterosis pada Tanaman Kelapa.. 14
Penanda Mikrosatelit (SSR Simple Sequence Repeats)………. 16
Variabilitas dalam mikrosatelit dan mekanismenya ... 19
Kegunaan potensial mikrosatelit untuk pemetaan gen ... 20
Sistem Aliran Informasi Genetika (Gene Flow) ... 21
III. BAHAN DAN METODE ………... 25
Tempat Penelitian ... 25
Bahan Penelitian ………... 25
Metode Penelitian ... 26
Pengamatan Karakter Morfologi Tanaman ... 26
Karakter Vegetatif ... 26
Karakter Generatif ... 26
Komponen Buah ... 26
Analisis Molekular ……… 26
Isolasi DNA Total ……… 26
Uji Konsentrasi dan Kemurnian DNA ……… 27
Analisis Mikrosatelit (Simple Sequence Repeat /SSR) ... 27
Elektroforesis Gel PAGE ………. 27
Pewarnaan Perak (Silver staining) ... 28
Identifikasi Fragmen DNA spesifik ... 29
Analisis Data ... 30
Depresi Penangkarandalam (Inbreeding Depression) ... 30
Penanda Morfologi ... 30
Rata-rata Heterozigot Observasi ……….. 30
Halaman
IV. HASIL DAN PEMBAHASAN ………. 32
1. Depresi Penangkarandalam (Inbreeding Depression) Kelapa Dalam Mapanget No.32 (DMT-32) Hasil Penyerbukan Selama Empat Generasi ……… 32 Depresi Penangkarandalam Berdasarkan Penanda Morfologi ……… 32
Karakter Vegetatif ……….. 32
Karakter Generatif ……….. 35
Komponen Buah ………. 39
Depresi Penangkarandalam Berdasarkan Penanda Molekular ………. 41
2. Pelacakan Tetua Kelapa Dalam Mapanget No.32 Melalui Aliran Gen (Gene Flow) Berdasarkan Penanda Mikrosatelit (SSR) ………. 45
3. Identifikasi Pita Spesifik Terpaut Dengan Karakter Produksi Buah Pada Tanaman Kelapa Berdasarkan Penanda Mikrosatelit (SSR) ……… 57
V. SIMPULAN ……….. 65
DAFTAR TABEL
Halaman
1 Nama 5 primer SSR (Rivera et al. 1999) dan urutan
basa nukleotidanya ……… 29
2 Nama 14 primer SSR CIRAD (2002) dan urutan
basa nukleotidanya ... 29
3 Depresi penangkarandalam pada karakter vegetatif generasi kedua, ketiga, dan keempat kelapa Dalam Mapanget
No.32 (DMT-32 ………... 33
4 Depresi penangkarandala m pada karakter generative generasi Kedua, ketiga, dan keempat kelapa Dalam Mapanget
No 32 (DMT-32) ………... 36
5 Depresi penangkarandalam pada karakter generatif generasi kedua, ketiga, dan keempat kelapa Dalam Mapanget
No.32 (DMT-32) ………... 40
6 Jumlah alel, Frekuensi heterozigot, Indeks fiksasi dan depresi penangkarandalam DMT-32 S2, DMT-32 S3 dan DMT-32 S4
berdasarkan penanda SSR ………. 42
7 Jumlah alel, individu heterozigot, individu homozigot, tingkat polimorfisme dan kesetimbangan Hardy-Weinberg ………. 45
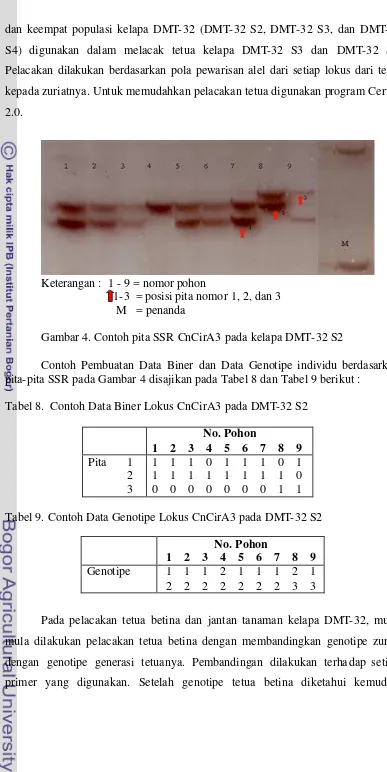
8 Contoh Data Biner Lokus CnCirA3 pada DMT-32 S2 ... 46
9 Contoh Data Genotipe Lokus CnCirA3 pada DMT-32 S2 ... 47
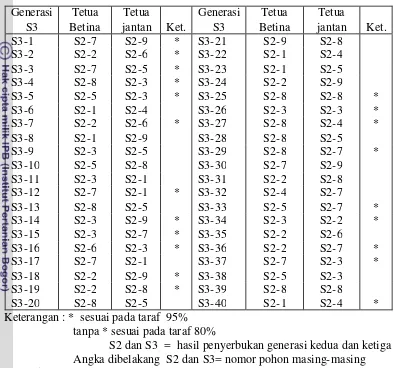
10 Pelacakan Tetua betina dan jantan Kelapa DMT-32 generasi S3 berdasarkan 15 lokus mikrosatelit (SSR) ……….. 48
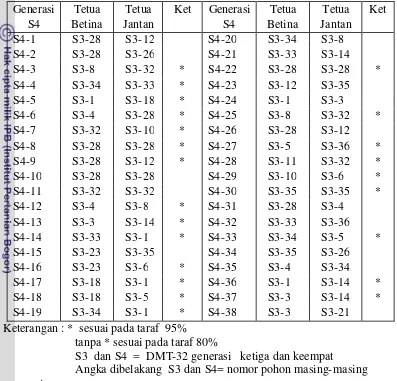
11 Pelacakan Tetua betina dan jantan Kelapa DMT-32 generasi S4
berdasarkan 15 lokus mikrosatelit (SSR) ……….. 49
12 Produksi buah dan tandan per pohon per tahun pada setiap generasi menyerbuk sendiri kelapa DMT-32 ……… 58
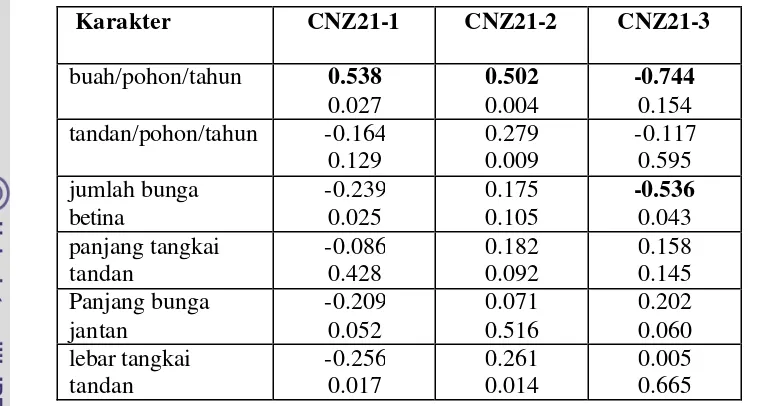
13 Korelasi antar alel (pita) dari CNZ21 dan CNZ51 ……… 62
14 Korelasi antara sifat generatif dengan Lokus CNZ21 ………… 63
DAFTAR GAMBAR
Halaman
1 Penampilan pohon dengan jumlah buah terbanyak dan tersedikit
pada generasi DMT -32 S2 ……… 38
2 Penampilan pohon dengan jumlah buah terbanyak dan tersedikit
pada generasi DMT -32 S3 ……… 38
3 Penampilan pohon dengan jumlah buah terbanyak, buah sedang
dan tidak berbuah pada generasi DMT -32 S4 ……… 39
4 Contoh pita SSR CnCirA3 pada kelapa DMT-32 S2 ………….. 46
5 Zuriat dari individu pohon nomor satu generasi kedua DMT-32
(DMT-32 S2 No.1) ……….. 50
6 Zuriat dari individu pohon nomor dua generasi kedua DMT-32
(DMT-32 S2 No.2) ………. 51
7 Zuriat dari individu pohon nomor tiga generasi kedua DMT-32
(DMT-32 S2 No.3) ……….. 52
8 Zuriat dari individu pohon nomor empat generasi kedua
DMT-32 (DMT-DMT-32 S2 No.4) ………. 53
9 Zuriat dari individu pohon nomor lima generasi kedua DMT-32
(DMT-32 S2 No.5) ……….. 53
10 Zuriat dari individu pohon nomor enam generasi kedua
DMT-32 (DMT-DMT-32 S2 No.5) ……… 54
11 Zuriat dari individu pohon nomor tujuh generasi kedua
DMT-32 (DMT-DMT-32 S2 No.7) ……… 55
12 Zuriat dari individu pohon nomor delapan generasi kedua
DMT-32 (DMT-32 S2 No.8) ……….. 55
13 Zuriat dari individu pohon nomor sembilan generasi kedua
DMT-32 (DMT-32 S2 No.9) ………... 56
14 Pita spesifik yang berkaitan dengan produksi buah pada kelapa
Halaman
15 Profil Pola Pita Lokus CNZ21 pada populasi DMT-32 S4…….. 60
16 Pita spesifik yang berkaitan dengan produksi buah pada kelapa
DMT-32 menggunakan Lokus CNZ51 ……… 60
PENDAHULUAN
Latar Belakang
Tanaman kelapa adalah salah satu tanaman strategis yang berperan bukan
saja dalam bidang ekonomi tetapi juga dalam kehidupan sosial dan budaya
masyarakat Indonesia. Seluruh bagian tanaman ini dapat dimanfaatkan baik
sebagai bahan makanan maupun bukan makanan, sehingga mendapat julukan
sebagai the tree of life.
Peluang pengembangan agribisnis kelapa sangat tinggi karena berbagai
produk bernilai ekonomi tinggi dapat dikembangkan dari tanaman kelapa.
Berbagai alternatif produk yang dapat dikembangkan adalah virgin coconut oil
(VCO), oleochemical (OC), desicated coconut (DC), coconut milk/cream
(CM/CC), coconut charcoal (CCL), activated carbon (AC), brown sugar (BS),
coconut fiber (CF), dan coconut wood (CW). Pelaku agribisnis kelapa dengan diversifikasi produk tersebut dapat meningkatkan pendapatannya 5-10 kali
dibandingkan bila hanya menjual kopra (Ditjenbun 2007).
Indonesia adalah Negara dengan pertanaman kelapa terluas di dunia.
Pentingnya tanaman kelapa bagi masyarakat Indonesia juga tercermin dari
luasnya pertanaman kelapa rakyat yang mencapai 97% (Ditjenbun 2007).
Pertanaman kelapa tersebar di seluruh kepulauan Indonesia. Sentra kelapa tersebar
di Sumatera (34.5%), Jawa (23.2%), Sulawesi (19.6%), Kalimantan (7.5%),
Maluku dan Papua (7.5%), serta Bali, NTB, dan NTT (8.0%). Namun sayang
produktivitas tanaman kelapa masih sangat rendah yaitu 0.8-1.2 ton kopra per
hektar. Penggunaan kelapa Dalam unggul akan dapat meningkatkan produksi
menjadi minimal 2.5-3.0 ton kopra per hektar per tahun dengan pemeliharaan
semi intensif (Ditjenbun 2007).
Migrasi suatu individu atau populasi tanaman dari satu tempat ke tempat
lain atau dari satu benua ke benua lain yang diikuti oleh isolasi geografi dan
hibridisasi dapat menyebabkan terjadinya aliran gen (gene flow). Konsekuensi dari adanya aliran gen antar populasi tanaman akan menyebabkan meningkatnya
keanekaragaman karakter genetik, menimbulkan kombinasi gen baru, dan
populasi lainnya (Nagy 1997). Keanekaragaman karakter genetik suatu populasi
semakin meningkat dengan adanya mutasi.
Keanekaragaman karakter genetik akibat migrasi, isolasi geografi, seleksi,
hibridisasi, dan mutasi telah terjadi pada tana man kelapa. Dari satu spesies Cocos nucifera L telah terbentuk berbagai macam populasi yang umumnya diberi nama berdasarkan tempat tumbuhnya, warna buah dan tipe tanaman kelapa. Secara
umum tanaman kelapa yang tumbuh secara lokal (indigenous) di suatu daerah diklasifikasi dalam 3 tipe. Ketiga tipe kelapa tersebut adalah Typica (Kelapa
Dalam), Nana (Kelapa Genjah) dan Aurantica (intermediate antara Typica dan
Nana), namun yang umum dikembangkan adalah Typica dan Nana (Liyanage & Corputy 1976).
Inventarisasi populasi kelapa yang dilakukan oleh COGENT, CGR (The International Coconut Genetic Resources Network, Coconut Genetic Resources) dari 17 negara dilaporkan sebanyak 936 populasi dan 309 diantaranya berasal dari
Asia Tenggara. Dari 309 aksesi yang berasal dari Asia Tenggara, 105 populasi
berasal dari Indonesia atau setara dengan 11.22% dari seluruh populasi kelapa
yang telah dilaporkan (Batugal 1998).
Keanekaragaman karakter genetik yang tinggi dari suatu populasi tanaman
sangat bermanfaat sebagai sumber keanekaragaman gen untuk program pemuliaan
tanaman dalam usaha perbaikan produksi tanaman pertanian dan pemeliharaan
kesinambungan sumber gen yang ada.
Kegiatan pemuliaan tanaman kelapa di Indonesia telah dilakukan melalui
eksplorasi, koleksi, dan hibridisasi. Pemuliaan tanaman kelapa di Indonesia telah
dilakukan sejak tahun 1911 saat pemerintah Belanda mengoleksi beberapa
populasi kelapa Dalam di pulau Jawa. Pada tahun 1927 pemerintah Belanda
mendirikan Klapper Proef Station di Manado dengan kegiatan eksplorasi, seleksi, dan koleksi plasma nutfah kelapa yang berada disekitar Desa Mapanget oleh
seorang ahli agronomi yaitu Dr. P.L.M. Tammes, dan ditanam di kebun percobaan
Mapanget. Dari kegiatan tersebut ditemukan 100 nomor yang masing- masing
nomor terdiri atas 10 pohon sehingga seluruhnya berjumlah 1000 pohon. Koleksi
ini kemudian dikenal sebagai Koleksi Tammes. Pada tahun 1955, seorang pemulia
negatif berdasarkan produksi tiap pohon dari koleksi Tammes dan didapatkan 42
nomor pohon terpilih. Hasil seleksi negatif tersebut selanjutnya diseleksi lagi dan
didapatkan 29 nomor pohon yang digunakan sebagai tetua dalam persilangan
terbuka dan buatan untuk uji keturunan (Novarianto et al. 1989)
Balai Penelitian Tanaman Kelapa dan Palma Lain (BALITKA) di Manado,
telah mengoleksi buah kelapa dari berbagai daerah di Indonesia. Populasi kelapa
yang dikoleksi umumnya diberi nama sesuai dengan daerah asal misalnya kelapa
Dalam Mapanget berasal dari desa Mapanget, Kabupaten Minahasa, Sulawesi
Utara, dan kelapa Genjah Kuning Nias berasal dari Pulau Nias, Sumatera Utara.
Beberapa nama aksesi lainnya tidak menggunakan nama daerah asalnya jika pada
saat pengoleksian dilakukan, aksesi tersebut sudah diberi na ma oleh masyarakat
setempat seperti kelapa Dalam Takome dan Genjah Raja yang berasal dari
Maluku, dan kelapa Genjah Salak yang berasal dari Kalimantan Selatan.
Untuk dapat digunakan dalam program pemuliaan, populasi kelapa yang
telah dikoleksi tersebut dik arakterisasi. Pengkarakterisasian tanaman kelapa yang
dikoleksi di BALITKA dilakukan terhadap sifat seperti: kecepatan berbunga
pertama, jumlah daun, jumlah tandan, jumlah buah, kadar kopra, dan kadar
minyak. Kelapa yang cepat berbunga adalah kelapa Genjah Salak (GSK), Genjah
Kuning Nias (GKN), dan Dalam Sawarna (DSA) berturut-turut 17 bulan, 28
bulan, dan 36 bulan. Kelapa yang berbuah banyak adalah kelapa Dalam Takome
(DTE) berkisar 200-230 btr/phn/thn dan kelapa GKN sekitar 80-120 btr/phn/thn.
Kelapa yang memiliki kadar kopra tinggi adalah DMT, DTA, DBI, DPU, DSA,
dan Dalam Riau (DRU) berkisar 270-300 gram/butir. Kelapa yang mempunyai
kadar minyak tinggi adalah DMT berkisar 67-71 % (Rompas 1993).
Hibridisasi yang telah dilakukan adalah antara kelapa tipe Genjah dengan
tipe Dalam dan kelapa tipe Dalam dengan Dalam. Hibridisasi antara kelapa tipe
Genjah dengan tipe Dalam bertujuan menggabungkan sifat baik yang dimiliki
oleh kedua tipe kelapa tersebut, sedangkan persilangan kelapa tipe Dalam dengan
tipe Dalam bertujuan untuk mendapatkan pohon atau populasi kelapa yang
berproduksi tinggi dengan pemeliharaan sederhana. Persilangan antara kelapa
Genjah dengan Dalam menghasilkan kelapa unggul yang diberi nama Khina-1,
jumlah buah banyak, kadar kopra dan kadar minyak tinggi diseleksi dan beberapa
nomor pohon terpilih disilangkan. Persilangan antara nomor-nomor terpilih kelapa
Dalam Mapanget menghasilkan Kelapa Baru (KB) yaitu KB-1, KB-2, KB-3, dan
KB-4. Kedelapan hasil persilangan tersebut dilepas oleh Menteri Pertanian pada
tahun 1984 (Novarianto 1996).
Selain menghasilkan kelapa hibrida hasil silangan kelapa tipe Genjah
dengan Dalam dan tipe Dalam dengan Dalam, dilakukan pula penyerbukan di
antara individu-individu terseleksi populasi kelapa Dalam Mapanget (DMT).
Koleksi Kelapa Dalam Mapanget (DMT) adalah salah satu kelapa dalam unggul
yang berproduksi tinggi dengan kadar minyak dan protein yang baik. Selain
digunakan sebagai tetua jantan dalam pembuatan kelapa hibrida, pohon-pohon
terseleksi kelapa Dalam Mapanget (DMT No 10, No 32, No 55) diserbuki oleh
campuran polen dari pohon-pohon tersebut. Buah kelapa dari hasil penyerbukan
ini ditanam di Kebun Percobaan Mapanget membentuk populasi DMT ge nerasi
pertama (DMT S1) pada 1957. Populasi DMT S1 ini diseleksi lagi dan individu
pohon terseleksi diserbuki oleh campuran polen dari pohon kelapa terseleksi
tersebut, dan buah kelapa yang diperoleh ditanam di Kebun Percobaan Kima Atas
pada tahun 1969, me mbentuk populasi generasi kedua DMT (DMT S2). Dengan
cara yang sama diperoleh populasi DMT generasi ketiga (DMT S3) pada tahun
1979, dan untuk generasi keempat (DMT S4) dilakukan penyerbukan individual
yaitu polen dari pohon terseleksi pada generasi ketiga diserbuki dengan polen dari
pohon yang sama dibuat pada tahun 1993. Karena polen yang digunakan
merupakan campuran polen dari pohon kelapa terseleksi sampai pada generasi
ketiga, dan benih yang diperoleh pada setiap generasi tidak dilabel dengan baik
sehingga identitas individu tetua pohon sebagai jantan dari setiap zuriat generasi
tidak diketahui.
Untuk mempelajari kontrol pewarisan suatu karakter pada tanaman, maka
diperlukan silsilah yang lengkap dan jelas asal usul persilangannya pada setiap
generasi. Suatu karakter yang dikontrol oleh gen, pola pewarisannya akan dapat
ditelusuri melalui silsilah hasil persilangan dari beberapa generasi yang jelas
polanya, akan tetapi bila pola pesilangan tidak jelas maka perlu dicari suatu
pencerminan genotipenya yang sedikit sekali dipengaruhi oleh lingkungan.
Pewarisan genotipe ini yang dicerminkan oleh fenotipenya dari satu generasi ke
generasi berikutnya, merupakan pelacakan aliran gen terutama untuk silsilah yang
tetuanya tidak jelas.
Penyerbukan sendiri atau penyerbukan oleh polen tanaman itu sendiri,
walaupun berupa campuran polen dari pohon terpilih, pada tanaman yang bersifat
menyerbuk terbuka dapat mengakibatkan terjadinya penghanyutan gen.
Penghanyutan genetik dan migrasi adalah proses netral dan mempengaruhi
frekuensi gen pada populasi alami dan terjadi pada seluruh genom. Sedangkan
rekombinasi dan seleksi menginduksi terjadinya perubahan genetik pada lokus
target (Cavalli Sforza & Bodmer 1971).
Penanda DNA adalah bagian kecil dari DNA yang memperlihatkan
polymorfisme sekuen pada individu- individu berbeda dalam suatu spesies.
Dengan berkembangnya teknologi penanda DNA, pengkarakterisasian
keanekaragaman genetik dan aliran gen dalam plasma nutfah kelapa pada taraf
DNA dapat dilakukan. Penanda DNA menawarkan alternatif analisis yang lebih
baik untuk melengkapi kelemahan penanda morfologi. Penanda DNA mampu
memberikan polimorfisme pita DNA dalam jumlah yang banyak, konsisten, dan
tidak dipengaruhi oleh lingkungan.
Penanda DNA telah banyak digunakan untuk identifikasi suatu individu
atau genotipe, derajat kekerabatan antar genotype, dan adanya variasi genetik
suatu populasi tanaman (Brown et al. 1996); determinasi adanya suatu gen atau kompleks gen yang diinginkan dalam suatu genotipe spesifik, dan pengembangan
varietas tanaman baru melalui transformasi (Henry 1887; Lande & Thompson
1990; Preston et al. 1999). Penanda DNA dapat digunakan untuk mengetahui adanya introgresi gen, melakukan pemetaan gen, gen tagging, dan konservasi
plasma nutfah (Gupta et al. 1999). Penanda DNA dapat pula digunakan untuk DNA fingerprinting tetua untuk memperkirakan penampilan zuriatnya (hybrid),
transgen backcross, homosigositas, dan peta genetik QTL (Quantitative Trait Loci) (Lee 1995).
Karena lamanya waktu yang dibutuhkan dalam pemuliaan tanaman kelapa,
digunakan sebagai alat untuk menyeleksi dan mengidentifikasi sifat-sifat genetik
dalam program pemuliaan kelapa.
Berbagai teknik penanda DNA telah digunakan dalam analisis keragaman
genetik kelapa seperti RAPD, RFLP (Lebrun et al 1998) dan AFLP (Perera et al.
1998), SSR (Teulat et al. 2000), dan beberapa sifat QTL (Herran et al. 2000; Lebrun et al. 2001).
Penanda DNA ruas berulang utamanya DNA ruas berulang sederhana
(Simple Sequence Repeats, SSR) adalah suatu penanda DNA yang dapat
memberikan polimorfik tinggi, merupakan lokus tunggal, kodominan dan relatif
mudah dianalisis (Innan et al. 1997; Crouch et al. 1998), sehingga berpotensi tinggi untuk digunakan dalam menganalisis keragaman genetik suatu populasi
(Szewe-McFaden et al. 996; Powell et al. 1996), dan menganalisis aliran gen dari tetua ke turunannya.(Perera et al. 2000; Lebrun et al. 2001), serta pemetaan genom suatu populasi (Herran et al. 2000).
Tujuan Penelitian
Penelitian ini bertujuan untuk (1) Mempelajari besarnya depresi
penangkarandalam (inbreeding depression) yang terjadi pada karakter vegetatif, generatif dan komponen buah kelapa Dalam Mapanget No.32 (DMT-32) hasil
penyerbukan tertutup buatan selama empat generasi, (2) Melacak tetua dari
DMT-32 hasil penyerbukan tertutup buatan generasi ketiga (DMT-DMT-32 S3) dan keempat
(DMT-32 S4) melalui analisis aliran gen menggunakan penanda mikrosatelit
(SSR), (3) Mengidentifikasi pita-pita yang berkaitan dengan karakter kuantitatif
I. TINJAUAN PUSTAKA
Asal dan Keanekaragaman Kelapa
Tanaman kelapa termasuk genus Cocos yang hanya memiliki satu spesies
yaitu Cocos nucifera L, tetapi memiliki fenotipik yang sangat beragam.
Keanekaragaman tanaman ini terutama pada sifat kecepatan berbunga pertama,
warna buah, bentuk dan ukuran buah, jumlah buah per tandan, tinggi batang hasil
dan kualitas kopra.
Daerah asal tanaman kelapa belum diketahui secara pasti. Namun dari
informasi etnobotani dan fakta, tingginya keanekagaman genetik kelapa yang ada
di wilayah Asia- Tenggara termasuk wilayah Indo-Malaya yang meliputi Malaysia
dan Indonesia diduga sebagai pusat asal dan sumber keanekaragaman genetik
kelapa di dunia. Beberapa alasan bahwa daerah Asia-Melanesia sebagai pusat asal
kelapa adalah 1) adanya penemuan fosil buah spesies dari Cocos di Plelocence North Auckland, New Zealand, 2) ditemukan fosil buah kelapa di dalam aspal di
Buton, Sulawesi Tenggara, 3) nama-nama lokal dan prosesing untuk kelapa lebih
banyak jenisnya di Asia, 4) adanya hewan-hewan pemakan kelapa seperti Birgus letro di Melanesia dan 5) variasi genetik kelapa lebih besar di Asia. Teori ini sejalan dengan adanya pusat asal (origin centre) tumbuhan di bumi yang dikemukakan oleh botanis Rusia Vavilov (1949), yaitu terdapat 8 sentra asal-usul
tanaman, dan Indo-Malaya merupakan salah satu sentra asal tanaman yang
mencakup pisang (Musa spp.), kelapa (Cocos nucifera), tebu (Saccharum offinarum), yam (Dioscorea spp.) dan jeruk (Citrus maxima).
Penyebaran kelapa mencakup daerah yang sangat luas. Luasnya
penyebaran kelapa kemungkinan disebabkan oleh penggunaan kelapa yang telah
lama memasyarakat, struktur buah yang memungkinkan dapat tahan selama
transportasi dalam jangka waktu yang agak lama, ketahanannya pada berbagai
kondisi, serta daya adaptasinya yang besar pada berbagai kondisi lingkungan.
Penyebaran yang paling cepat dan luas adalah melalui campur tangan
manusia, melalui perpindahan penduduk dari satu tempat ke tempat lain atau
migrasi dapat berbentuk (1) migrasi satu arah (one-way migration) yaitu migrasi yang terjadi satu arah dari populasi daratan yang besar dengan genotipe tertentu
ke sebuah populasi pulau yang frekuensi genotipenya bervariasi, (2) migrasi
simetrik (simetric migration) yaitu dua populasi berubah dengan frekwensi persentase yang sama dari migran setiap ge nerasi (Servedio & Kirkpatrick 1997).
Migrasi suatu individu atau populasi tanaman dari satu benua ke benua lain atau
dari satu tempat ke tempat lain, yang diikuti oleh terjadinya isolasi geografi dan
hibridisasi menyebabkan terjadinya aliran gen (gene flow). Aliran gen antar populasi tanaman akan menimbulkan konsekuensi evolusi yang nyata dan dapat
menyebabkan meningkatnya keanekaragaman karakter genetik, dapat
menimbulkan kombinasi gen baru, dan memindahkan kemampuan beradaptasi di
suatu tempat dari suatu populasi ke populasi lainnya (Nagy 1997).
Penyebaran kelapa ini berawal dari Asia ke arah Timur menuju Pasifik
dan Amerika, serta ke Barat menuju Afrika (Perera et al. 2000; Teulat et al.
2000). Hasil penelitian menggunakan RFLP membuktikan bahwa kelapa bergerak
dari Asia Tenggara menuju Pasifik dan pantai barat Amerika (Lebrun et al. 1998). Keanekaragaman fenotipe kelapa yang besar disebabkan oleh bunga
tanaman yang bertipe monocious yaitu disetiap pohon menghasilkan bunga betina dan jantan yang terpisah dalam satu tandan bunga yang sama. Bunga jantan
terdapat pada bagian ujung dan bunga betina pada bagian pangkal tandan bunga.
Masa antesis bunga jantan dan masa reseptif bunga betina pada kelapa Dalam
tidak bersamaan sehingga lebih cenderung bersifat kawin silang. Pada tipe Genjah
terdapat masa tumpang tindih antara masa reseptif bunga betina dan antesis bunga
jantan sehingga tipe Genjah lebih cenderung kawin sendiri. Selain itu antar
populasi kelapa dan antar pohon dalam satu populasi kelapa terdapat perbedaan
waktu awal dan lamanya periode antesis bunga jantan dan reseptif bunga
betinanya.
Perbedaan letak dan periode reseptif bunga betina dan antesis bunga jantan
menyebabkan tanaman kelapa memiliki empat kemungkinan tipe penyerbukan
yaitu (1) tipe menyerbuk silang (Strict allogami) dengan ciri tidak ada tumpang tindih antara periode reseptif bunga betina dan antesis bunga jantan, baik dalam
menyerbuk sendiri tidak langsung (Indirect autogami) dengan ciri periode reseptif bunga betina pendek sehingga tidak ada tumpang tindih periode reseptif bunga
betina dan antesis bunga jantan dalam tandan yang sama, tetapi terjadi tumpang
tindih periode reseptif bunga betina dan antesis bunga jantan pada tandan
berikutnya, (3) Tipe menyerbuk sendiri (Direct autogami) dengan ciri periode reseptif bunga betina dan antesis bunga jantan tumpang tindih dalam tandan yang
sama, dan (4) Tipe menyerbuk sendiri semi tidaklangsung (Semi indirect autogami) denga n ciri periode masak bunga betina pendek, terjadi tumpang tindih periode masak bunga betina dan jantan dalam tandan yang sama, maupun dalam
tandan bunga berikutnya (Sangare et al. 1978).
Berdasarkan kecepatan berbunga pertama, tanaman kelapa digolongkan
atas 2 tipe yaitu kelapa tipe Dalam dan tipe Genjah. Kelapa tipe Dalam umumnya
memiliki batang yang tinggi, sekitar >15 meter dan bagian pangkal membengkak
(disebut bole), mahkota daun terbuka penuh berkisar 30 - 40 daun, panjang daun berkisar 5 - 7 meter, berbunga pertama lambat berkisar 7 - 10 tahun setelah tanam,
buah masak sekitar 12 bulan setelah penterbukan, umur tanaman dapat
mencapai 80 - 90 tahun, lebih toleran terhadap macam- macam jenis tanah dan
kondisi iklim, kualitas kopra dan minyak, serta sabut umumnya baik, pada
umumnya menyerbuk silang (Menon & Pandalai 1958; Child 1974; Fremond et al. 1966; Foale 1992; Rompas et al. 1988; Santos et al. 1996).
Kelapa tipe Genjah pada umumnya memiliki batang pendek berkisar 12
meter dan agak kecil, tidak memiliki bole, panjang daun berkisar 3 - 4 meter, berbunga pertama cepat berkisar 3 - 4 tahun setelah tanam, buah masak berkisar
11-12 bulan sesudah penyerbukan, umur tanaman dapat mencapai 35 - 40 tahun,
kualitas kopra dan minyak serta sabut kurang baik (Rompas et al. 1988), umumnya menyerbuk sendiri (Foale 1992).
Kelapa Dalam dapat dibagi lagi dalam tiga kelompok yaitu (1) kelompok
typical yang memiliki ciri menyerbuk silang, pembungaan lambat, bunga normal, dan ukuran buah beragam dari sedang sampai besar, (2) kelompok spicata
memiliki ciri menyerbuk silang, pembungaan lambat dan ukuran buah sedang, (3)
Kelapa Genjah dapat dibedakan atas 2 kelompok yaitu (1) kelompok Nana
dengan ciri menyerbuk sendiri, pembungaan cepat, dan ukuran buah kecil, dan (2)
kelompok Jamaica dengan ciri menyerbuk sendiri, pembungaan cepat, dan ukuran buah sedang (Santos 1983).
Selain tipe kelapa Dalam dan Genjah, beberapa jenis kelapa yang
dianggap unik adalah (1) kelapa Hibrida, adalah jenis kelapa hasil persilangan
antara tipe kelapa Genjah dan kelapa Dalam dengan sifat lebih unggul dari kedua
tetuanya, (2) kelapa Kopyor (Genjah maupun Dalam), adalah jenis kelapa yang
mengalami kelainan pada endospermanya (daging buahnya sangat lunak dan
mudah lepas dari tempurungnya) sehingga perbanyakannya harus melalui kultur
embrio, (3) kelapa Kenari, mempunyai struktur daging buah berbeda dari kelapa
normal, rasanya sangat renyah sehingga sangat cocok untuk dikonsumsi sebagai
kelapa muda atau campuran fruit cocktail, (4) kelapa Sabut Merah, terdapat warna
merah muda (pink) pada daerah basal (pangkal buah), (5) kelapa Bercabang,
anakan muncul dari pangkal batang pohon pertama, (6) kelapa yang memiliki
lembaga lebih dari satu (Novarianto & Miftahorrachman 2000).
Tingginya keanekaragaman tanaman kelapa ini merupakan materi dasar
yang dapat digunakan dalam program pemuliaan kelapa, oleh sebab itu pelestarian
plasmanutfah kelapa adalah faktor yang sangat penting agar tidak terjadi erosi
genetik atau hilangnya sumber gen potensial. Penyebab terjadinya erosi genetik
kelapa di Indonesia diakibatkan oleh berbagai faktor antara lain pengalihan fungsi
lahan kebun kelapa menjadi perumahan, pemakaian kelapa hibrida, penggantian
tanaman kelapa dengan komoditas lain yang dianggap lebih bernilai ekonomi
tinggi, organisasi dibidang konservasi genetik kelapa belum baik, dan pemulia
kelapa masih sangat sedikit.
Pemuliaan Tanaman Kelapa di Indonesia
Untuk berbagai tanaman seperti kelapa kegiatan pemuliaan merupakan
proses yang sangat lama dan mahal. Pengujian satu keturunan tunggal
membutuhkan minimal 70 pohon atau membutuhkan lahan setara 0.5 Ha untuk
masa penelitian minimal selama 10 tahun setelah tanam. Selain itu jumlah zuriat
al. 1996). Pada kondisi ini diperlukan metode yang dapat meningkatkan efisiensi untuk pemuliaan tanaman kelapa.
Karakterisasi dan evaluasi standar suatu aksesi secara rutin dapat
dilakukan dengan berbagai metode termasuk cara-cara tradisional seperti list
karakter morfologi, dan evaluasi terhadap karakter agronomis pada kondisi
lingkungan bervariasi hingga karakterisasi yang melibatkan profil isozim, protein
dan sekuen DNA spesifik melalui pendekatan genom berbeda (Vicente de et al.
2005).
Karakterisasi adalah deskripsi suatu karakter atau kualitas suatu individu
(Merriam-Webster 1991). Karakterisasi sumber genetik merujuk pada proses
identifikasi atau diferensiasi aksesi. Terminologi yang disepakati oleh Genebanks
dan Germplasm management diartikan sebagai deskripsi karakter yang pewarisannya tinggi, mudah dilihat secara kasat mata, dan terekspresi stabil pada
berbagai lingkungan (IPGRI 2003).
Pemuliaan tanaman kelapa di Indonesia telah dilakukan sejak 1911/1912
saat pemerintah Belanda mengoleksi 240 buah kelapa yang berasal dari
pohon-pohon kelapa Dalam terpilih di pulau Jawa (Reyne 1948). Pada 1927 pemerintah
Belanda mendirikan Klapper Proef Station di Manado dengan kegiatan eksplorasi, seleksi dan koleksi plasma nutfah kelapa yang berada disekitar Desa Mapanget
oleh seorang ahli agronomi yaitu Dr. P.L.M. Tammes. Dari kegiatan tersebut
ditemukan 100 nomor. Seleksi dilakukan berdasarkan produksi buah banyak per
pohon dan terseleksi 100 pohon, dari setiap pohon diambil buahnya untuk ditanam
di Kebun Percobaan Mapanget yang sekarang disebut Kebun Instalasi Penelitian
Mapanget (KIP Mapanget). Setiap nomor tetua ditanam dalam bentuk barisan
sebanyak 10 poho n sehingga seluruhnya berjumlah 1000 pohon (Reyne 1948).
Populasi ini kemudian dikenal sebagai Koleksi Tammes. Pada tahun 1955,
seorang pemulia tanaman berkebangsaan Jerman yaitu Dipl. Ing. A.F. Ihne
melakukan seleksi negatif berdasarkan produksi pada setiap pohon dari koleksi
Tammes dan didapatkan 42 nomor pohon terpilih. Hasil seleksi negatif tersebut
selanjutnya diseleksi lagi dan didapatkan 29 nomor pohon yang digunakan
sebagai tetua dalam persilangan terbuka dan buatan untuk uji keturunan
Balai Penelitian Tanaman Kelapa dan Palma Lain (BALITKA) di Manado,
telah mengoleksi tanaman kelapa dari berbagai daerah di Indonesia. Nama
populasi kelapa yang dikoleksi umumnya diberikan sesuai dengan daerah asal
misalnya kelapa Dalam Mapanget berasal dari desa Mapanget, Kabupaten
Minahasa, Sulawesi Utara, dan kelapa Genjah Kuning Nias berasal dari Pulau
Nias, Sumatera Utara. Beberapa nama aksesi lainnya tidak menggunakan nama
daerah asalnya jika pada waktu pengoleksian dilakukan, aksesi tersebut sudah
diberi nama oleh masyarakat setempat. Nama itu selanjutnya digunakan sebagai
nama populasi/aksesi dari koleksi tersebut contohnya kelapa Dalam Takome dan
Genjah Raja yang berasal dari Maluku, dan kelapa Genjah Salak yang berasal dari
Kalimantan Selatan.
Program pemuliaan kelapa di Indonesia bertujuan untuk menghasilkan
bahan tanaman yang memiliki karakteristik antara lain: cepat berbunga, habitat
pohon pendek, resisten terhadap hama dan penyakit, hasil kopra per satuan luas
tinggi dengan pemupukan rendah, ukuran buah besar, daging buah tebal,
kandungan minyak tinggi, dan kandungan air rendah (Rompas et al. 1998)
Untuk dapat digunakan dalam program pemuliaan, populasi kelapa yang
telah dikoleksi tersebut dikarakterisasi. Untuk mendukung studi tentang
keanekaragaman kelapa dan pengkarakterisasian kelapa berdasarkan penanda
morfologi, COGENT telah menerbitkan Manual on Standardized Research Technique in Coconut Breeding (Stantech Manual) untuk menjadi pedoman dalam melakukan eksplorasi dan karakterisasi plasma nutfah kelapa (Santos et al. 1996).
Pengkarakterisasian tanaman kelapa yang dikoleksi di BALITKA
dilakukan terutama terhadap sifat morfologi seperti: kecepatan berbunga pertama,
jumlah daun, jumlah tandan, jumlah buah, kadar kopra, dan kadar minyak.
Kelapa yang cepat berbunga adalah kelapa Genjah Salak (GSK), Genjah Kuning
Nias (GKN), dan Dalam Sawarna (DSA) berturut-turut 17 bulan, 28 bulan, dan 36
bulan. Kelapa yang berbuah banyak adalah kelapa Dalam Takome (DTE) berkisar
200 - 230 butir per pohon per tahun dan kelapa GKN sekitar 80-120 butir per
pohon per tahun. Kelapa yang memiliki kadar kopra tinggi adalah Dalam
Mapanget (DMT), Dalam Tenga (DTA), Dalam Palu (DPU), Dalam Sawarna
yang mempunyai kadar minyak tinggi adalah DMT berkisar 67-71 % (Rompas
et al. 1989; Rompas 1993).
Beberapa koleksi yang tergolong unik adalah Dalam Mamuaya (Sulut),
Dalam Palapi (Sulteng), dan Dalam Dobo yaitu berbuah besar, sabut tip is dengan
potensi hasil 4-5 ton kopra/ha. Kelapa Dalam Santongbolang (Sulut) berbuah
banyak yaitu 60-100 buah /tandan. Kelapa Kenari ditemukan secara individu
dalam suatu populasi memiliki keunikan pada struktur daging buah yang berbeda
dari buah normal, lebih renyah sehingga sangat cocok untuk campuran es buah.
Kelapa bersabut merah ditemukan di Gorontalo dan NTT, pada bagian basal
kelapa (pangkal buah kelapa) berwarna merah muda (pink). Kelapa bertunas
ditemukan di Sulut dan Sulteng, keunikannya terletak pada munculnya anakan
pada umur setahun setelah tanam, dan setiap 1-2 tahun muncul anakan baru yang
semuanya dapat berbuah normal. Kelapa kopyor ditemukan secara alami di Jawa
dan Sumatera (Novarianto & Miftahorrachman 2000).
Berdasarkan hasil karakterisasi maka populasi kelapa GKN dipilih sebagai
tetua betina dan tiga populasi kelapa tipe Dalam yaitu DTA, DBI, DPU digunakan
sebagai tetua jantan untuk menghasilkan empat kelapa hibrida. Sifat unggul yang
diharapkan dari persilangan antara kedua tipe kelapa ini adalah kelapa hibrida
yang cepat berbuah dan berproduksi tinggi. Kelapa Hibrida hasil persilangan
antara GKN dengan populasi DTA, DBI, dan DPU masing- masing diberi nama
KHINA-1, KHINA-2, dan KHINA-3. Ketiga kelapa hibrida tersebut rata-rata
mulai berbunga pada umur 3 tahun setelah tanam, dengan produksi kopra 3.66
- 4.09 ton kopra per hektar per tahun (Mahmud 1993). Selain keempat kelapa
hibrida unggul tersebut berbagai silangan antara beberapa populasi kelapa tipe
Genjah dengan tipe Dalam sedang dalam pengujian.
Kelapa Dalam Mapanget (DMT) yang memiliki jumlah buah banyak,
kadar kopra dan kadar minyak tinggi diseleksi dan beberapa nomor pohon terpilih
disilangkan. Persilangan nomor-nomor terpilih tersebut dihasilkan kelapa Dalam
unggul yang diberi nama Kelapa Baru (KB) yaitu KB-1 (32 x 32), KB-2 (32 x 2),
KB-3 (32 x 83) dan KB-4 (32 x 99) dengan potensi produksi 3 - 3.5 ton kopra per
Balitka selain melakukan seleksi, koleksi, dan hibridisasi pada tanaman
kelapa dilakukan pula penyerbukan tertutup pada kelapa Dalam Mapanget
(DMT). Penyerbukan tertutup kelapa Dalam Mapanget telah dilakukan hingga
mencapai generasi keempat (S4). Kelapa DMT yang tersisa 29 nomor hasil seleksi
massa negatif diseleksi lagi. Kriteria seleksi yang digunakan adalah jumlah buah
per tandan dan jumlah tandan per pohon per tahun banyak. Beberapa nomor
terpilih yang berproduksi tinggi di antaranya adalah No.2, No.10, No.32, No.55,
No.83, dan No.99. Setiap nomor terdiri atas 10 pohon. Selanjutnya kelapa DMT
No 10, No 32, dan No 55, dibuat menyerbuk sendiri menggunakan campuran
serbuksari (polen bulk) pohon-pohon terseleksi dari setiap nomor tersebut. Zuriat
hasil penyerbukan sendiri itu ditanam di Kebun Percobaan Mapanget (KIP
Mapanget) membentuk populasi DMT S1. Populasi DMT S1 ini diseleksi lagi dan
individu terseleksi dibuat menyerbuk sendiri menggunakan polen campuran dari
pohon-pohon terseleksi tersebut. Zuriat yang diperoleh ditanam di Kebun
Percobaan Kima Atas (KIP Kima Atas) pada tahun 1969, yang membentuk
populasi DMT generasi kedua (DMT S2). Dengan cara yang sama didapatkan
populasi DMT generasi ketiga (DMT S3). Untuk generasi keempat dilakukan
penyerbukan individual yaitu polen dari pohon terseleksi dari generasi ketiga
diserbukan kembali pada pohon yang sama dan diperoleh populasi DMT generasi
keempat (DMT S4).
Depresi Penangkarandalam dan Heterosis pada Tanaman Kelapa
Kelapa Dalam pada umumnya bersifat menyerbuk terbuka sedangkan
kelapa Genjah pada umumnya menyerbuk sendiri (Child 1974; Thampan 1981).
Penyerbukan terbuka pada tanaman kelapa Dalam disebabkan oleh adanya
perbedaan masa reseptif bunga betina dan masa antesis bunga jantan. Masa
reseptif bunga betina mulai kurang lebih 3 (tiga) minggu setelah seludang terbuka
atau 3-6 hari setelah masa antesis bunga jantan. Pada tanaman kelapa Genjah
dilaporkan adanya masa tumpang tindih antara fase reseptif bunga betina dan fase
Dalam upaya merakit kelapa hibrida yang unggul karena heterosis
diperlukan tetua-tetua homosigot. Persilangan antar dua tetua homosigot yang
berbeda akan menghasilkan turunan yang unggul akibat pengaruh heterosis
(Allard 1960). Kelapa tipe Dalam pada umumnya menyerbuk silang sehingga
keturunannya memiliki penampilan beragam karena genotipe yang heterosigot
(Menon & Pandalai 1960; Child 1974; Fremond et al. 1966; Foale 1992). Untuk mendapatkan tetua homosigot pada kelapa Dalam dapat diperoleh melalui teknik
penyerbukan tertutup sampai beberapa generasi. Zuriat yang dihasilkan dari
proses penyerbukan tertutup akan memiliki derajat homozigot yang meningkat
dengan penurunan vigor sebagai akibat adanya peristiwa depresi
penangkarandalam.
Depresi penangkarandalam dan heterosis adalah dua fenomena bertolak
belakang yang banyak dipelajari pada tanaman dan binatang. Depresi
penangkarandalam berkaitan dengan menurunnya kekekaran turunan hasil
penyerbukan sendiri (inbreeding) pada beberapa generasi tanaman menyerbuk terbuka (Stebbins 1958). Sebaliknya heterosis berkaitan dengan keunggulan
hibrida (F1) melebihi nilai atau rata-rata kedua tetuanya bila dua galur murni yang
berbeda disilangkan.
Pada populasi kelapa Dalam Mapanget terutama No.10, No.32, dan No.55
penyerbukan tertutup di antara pohon-pohon terpilih dari masing- masing nomor
pohon telah mencapai generasi keempat (S4), sehingga diduga tingkat
homozigositas telah mencapai 93.75%. Pada tingkat ini bila dilakukan seleksi
individu dan dilanjutkan dengan persilangan dengan kelapa Genjah terpilih,
diharapkan akan diperoleh kelapa dengan efek heterosis tinggi (Novarianto et al.
1989). Bila dua galur murni yang berbeda disilangkan, ketegaran hibrida akan
kembali (Allard 1960).
Perkawinan antar individu- individu yang berkerabat dekat dalam populasi
kelapa Dalam akan mengakibatkan 2 hal yaitu depresi penangkarandalam akibat
peningkatan homozigositas dan penurunan heterozigositas. Generasi S3 dari
kelapa DMT memiliki batang yang lebih pendek dibandingkan populasi tetuanya
Bago-Oshiro Tall (keduanya populasi kelapa lokal Filipina) berbunga lebih awal
dibandingkan tetuanya yang menyerbuk terbuka (Santos & Sangare 1992).
Untuk mempelajari depresi penangkarandalam dapat dilakukan melalui 2
cara yaitu (1) studi komparatif sifat-sifat morfologi menggunakan populasi
menyerbuk terbuka (open pollination, OP) dan generasi menyerbuk tertutup dari kelapa Dalam, dan (2) penanda molekular (Ritland 1990; 1996). Untuk tujuan ini
Ritland (1996) menganjurkan SSR (simple sequence repeats) sebagai penanda DNA yang tepat, karena sifatnya yang kodominan sehingga memungkinkan
membedakan individu-individu homozigot dan heterozigot suatu populasi.
Penanda Mikrosatelit (SSR Simple Sequence Repeats)
DNA genom terdiri atas DNA ruas khas yang merupakan gen struktural
dan DNA ruas berulang. DNA ruas berulang adalah suatu komponen yang
terintegrasi di antara fragmen- fragmen DNA ruas khas dalam genom eukariot.
Fragmen ruas berulang sederhana (SSR) ini dapat mencapai > 90% DNA total
yang ada pada genom tanaman (Weising et al. 1995). Makin besar ukuran genom tanaman, makin besar pula proporsi DNA ruas berulangnya. Proporsi DNA ruas
berulang dalam genom tanaman jagung mencapai 60% (Gupta et al. 1984), kedelai mencapai 60% (Walbot dan Goldberg, 1979), gandum mencapai 70%
(Flavel 1980), dan padi mencapai 50% (McCouch et al. 1988).
Pengulangan ruas berulang DNA dapat berupa pengulangan ruas secara
moderat atau selang seling (interspace) dan pengulangan ruas secara berurutan
(tandem).
Pengulangan ruas secara tandem diklasifikasikan berdasarkan panjang dan
jumlah ruas berulang di dalam genom yang dapat berupa (1) DNA Satelit, adalah
DNA yang memiliki pengulangan sangat tinggi biasanya antara 1000 – 100 000
kopi, sering berada pada bagian heterokromatin, (2) Minisatelit, memiliki
pengulangan yang lebih sedikit yaitu 10 – 60 pasang basa, (3) Mikrosatelit (SSR),
disebut juga fragmen berulang sederhana atau fragmen berulang tandem
sederhana, memiliki pengulangan lebih pendek dengan derajat pengulangan lebih
(4) Midisatelit, memiliki ruas berulang yang merupakan kombinasi dari satelit dan
minisatelit (Weising et al. 1995; Karp et al. 1995; )
DNA ruas berulang yang memiliki variasi paling tinggi dalam genom
tanaman adalah sekuen berulang dengan fragmen berulang sederhana atau pendek.
Fragmen ini dikenal dengan nama minisatelit dan mikrosatelit. Mikrosatelit (SSR)
terdapat dalam jumlah banyak dan menyebar di dalam genom. Mikrosatelit
memiliki unit berulang berkisar antara 1 – 6 pasang basa, dan minisatelit memiliki
unit berulang yang lebih panjang.
Variasi fragmen-fragmen ini biasanya merupakan hasil perubahan dalam
jumlah kopi dari perulangan asal dan sering dikategorikan sebagai Variable number of tandem repeats (VNTR). Karena level polimorfisme yang sangat tinggi dapat dideteksi dengan fragmen ini, VNTR diakui sebagai alat yang manjur untuk
sidik jari dan identitas kultivar tanaman (Karp et al. 1995). Fragmen ini juga dapat digunakan untuk mempelajari keragaman antar dan intra populasi, studi ekologi,
menghitung jarak genetik, dan mempelajari evolusi tanaman (Perera et al. 2000). Mikrosatelit dikenal dengan beberapa nama seperti simple sequence
repeat (SSR), simple tandem repeat (STR), sequence tagged microsatellite site
(STMS), dan simple sequence length polimorphism (SSLP).
Simple sequence repeat (SSR) memberikan kandungan informasi yang tinggi, pada umumnya single lokus, bersifat kodominan, membutuhkan jumlah
DNA yang sangat sedikit, relatif sederhana, dan deteksi yang didasarkan pada
PCR menandakan bahwa SSR merupakan alat ideal untuk banyak aplikasi genetik
(Karp et al. 1995; Rivera et al. 1999; Saghai-Maroof et al. 1994; Morgante & Olivieri 1993).
Sekuen mikrosatelit (SSR) yang pendek dan dengan sekuen DNA pengapit
bersifat conserved, sehingga sekuen ini telah digunakan mendesain primer untuk mengamplifikasi situs spesifik menggunakan PCR. Jika primer-primer tersebut
digunakan mengamplifikasi lokus- lokus SSR tertentu, maka setiap primer akan
menghasilkan polimorfisme dalam bentuk perbedaan panjang hasil amplifikasi
yang dikenal dengan SSLP (Simple Sequence Length Polymorphism). Setiap panjang mewakili satu alel dari suatu lokus. Perbedaan panjang terjadi karena
Olivieri 1993; Gupta et al. 1996; Karp et al. 1997; Liu 1998). Keanekaragaman jumlah ulangan pada mikrosatelit dapat dideteksi dengan mengelektroforesis
produk DNA yang sudah diamplifikasi di dalam sekuen gel standar, yang dapat
memisahkan fragmen- fragmen yang membedakan tiap-tiap nukleotida.
Mikrosatelit (SSR) terdapat dalam jumlah banyak dan menyebar di dalam
genom. Bentuk umum pengulangan SSR adalah pengulangan dua basa secara
sederhana seperti (CA)n; (AC)n; (GT)n; (GA)n; (CT)n; (CG)n; (GC)n; (AT)n;
dan (TA)n, n adalah jumlah pengulangan. Mikrosatelit dengan pengulangan
3-basa dan 4-3-basa ditemukan juga tetapi frekuensinya lebih rendah dibandingkan
pengulangan 2-basa (Liu 1998; Preston et al. 1999). Pengulangan SSR paling banyak adalah (AT)n diikuti oleh (A)n, (AG)n, (AAT)n, (AAC)n, (AGC)n,
(AAG)n, (AATC)n, (AC)n (Brown et al. 1996). Pada kelapa, sekuensing yang dilakukan pada 197 klon pustaka genom dari kelapa Tagnanan menunjukkan
bahwa 75% mengandung sekuen mikrosatelit dan 64% dari SSR tersebut adalah
pengulangan 2-basa GA/CT, CA/GT dan GC/CG; 6 % merupakan pengulangan
3-basa dan 30 % merupakan pengulangan campuran (Rivera et al. 1999).
Berdasarkan teknik PCR, primer-primer SSR yang sangat informative
telah dapat dikembangkan pada berbagai tanaman. Primer-primer SSR yang telah
berhasil dibuat pada tanaman kelapa merupakan sesuatu yang sangat berharga
bagi pemulia kelapa. Dengan penggunaan PCR multipleks (aplikasi secara
simultan dari beberapa penanda genetik dalam suatu reaksi tunggal) yang
digabungkan dengan deteksi DNA berdasarkan teknologi fluorescence dan ukuran
alel semi otomatis, memungkinkan menghasilkan informasi genetik dengan
kuantitas yang lebih besar dan akurat berdasarkan penanda-penanda DNA SSR
(Mitchell et al. 1997).
Sejak mikrosatelit ditemukan dalam jumlah yang sangat banyak di dalam
genom, dengan tingkat polimorfisme yang tinggi dan mudah dianalisis,
mikrosatelit menjadi penanda pilihan untuk pemetaan genetik dan analisis
keterpautan pada hampir semua spesies.
Penggenotipan SSR akan memberikan seleksi yang efisien dari tetua
untuk program pemuliaan kelapa, dimana pemilihan tetua-tetua yang divergen
hybrid (Perera et al. 2000). Perera et al. (2000) menganalisis populasi kelapa dari berbagai daerah di dunia dan hasilnya menguatkan evolusi kelapa dan lintasan
penyebarannya ke seluruh dunia. Herran et al. (2000) menggunakan SSR untuk membuat peta keterpautan dan analisis lokus sifat kuantitatif (quantitative trait loci) kecepatan berkecambah pada kelapa. Ritter et al (2000) menggunakan SSR membuat peta keterpautan produksi daun dan lingkar batang. Lebrun et al (2001) menggunakan penanda yang sama untuk membuat peta keterpautan dan analisis
QTL untuk karakter-karakter produksi buah. Semua hasil ini memberikan
gambaran bahwa SSR memberikan implikasi untuk tinjauan konservasi, karena
primer-primer SSR dapat digunakan dalam mengidentifikasi genepool core untuk
konservasi ex situ yang sangat berguna bagi program pemuliaan kelapa di masa
mendatang.
Variabilitas dalam mikrosatelit dan mekanismenya
Keragaman mikrosatelit tergantung pada variasi dalam jumlah ulangan
pada pusat sekuen. Derajat keragaman secara positif sangat berkorelasi dengan
panjang ulangan sekuen (Weber & May, 1989), dengan panjang mikrosatelit
kurang dari 20 bp maka akan kurang tingkat polimorfismenya. Variasi yang tinggi
ditemukan pada genom inti bila diband ingkan dengan cpDNA menggunakan
penanda RAPD, AFLP dan ISSR (Viard et al. 2001; Hultquist et al. 1996; Panda
et al. 2003, Cronn et al. 2002). Beberapa penelitian menunjukkan bahwa mikrosatelit (SSR) memiliki tingkat diversitas yang tinggi pada berbagai tanaman
yang diuji (Perera et al. 2000; Narvel et al. 2000; Burstin et al. 2001, Cipriani et al. 2002).
Walaupun fungsinya masih belum jelas, namun banyak dari sekuen ini
berlokasi di daerah sentromer, telomer dan di bagian dasar loop kromatin yang
mempunya i peranan dalam perpasangan dan synopsis kromosom (Weising et al.
1995). Naylor dan Clark (1990) melaporkan bahwa ulangan sekuen (GT)n
memiliki aktivitas peningkatan transkripsi.
Mekanisme mutasi menghasilkan perubahan pada jumlah ulangan cetakan
penyebab mutasi pada fragmen sekuen berulang sederhana ini adalahterjadinya
Strand slippage selama replikasi atau perbaikan DNA (Levinson & Gutman 1987). Kebanyakan mutasi yang terjadi pada sekuen mikrosatelit merupakan hasil
dari kejadian-kejadian intrakromosom tanpa adanya sekuen homolog pada
kromosom-kromosom lain (Webber 1989).
Kegunaan potensial mikrosatelit untuk pemetaan gen
Sejak mikrosatelit ditemukan dalam jumlah yang sangat banyak di dalam
genom, dengan tingkat polimorfisme yang tinggi dan mudah dianalisis,
mikrosatelit menjadi penanda pilihan untuk pemetaan genetik dan analisis
keterpautan pada hampir semua spesies.
Moran (1993) mengidentifikasi 23 model gen yang mengandung sekuen
ulangan mono-, di-, tri-, dan tetra nukleotida. Ulangan- ulangan tersebut berada
pada 5’-tidak terkode, 3’-tidak terkode, 3’-tidak terartikan, 5’-tidak terartikan, dan
daerah-daerah pengkodean. Sebuah daerah yang membawa sekuen mikrosatelit
dapat diketahui sehingga sekuen-sekuen pengapit (flanking) mikrosatelit digunakan untuk mendesain primer-primer untuk amplifikasi PCR.
Polimorfisme dapat dideteksi dengan mengamplifikasi DNA genom dari
individu- individu yang akan dipelajari menggunakan primer pada reaksi PCR.
Penggunaan pendekatan semacam ini untuk memproduksi penanda-penanda
mikrosatelit sudah dilakukan pada berbagai spesies baik tumbuhan seperti kelapa
(Rivera et al. 1999: Teulat et al. 2000; Lebrun et al. 2001; Akuba 2002), Akasia (Prihartini et al. 2006), Jati (Boer 2007) maupun hewan seperti sapi dan domba (Moore et al. 1992).
Lokasi dari sekuen berulang sederhana seperti mikrosatelit di antara gen
atau yang berdekatan dengan suatu gen sangat penting, agar dapat digunakan
sebagai penanda dari sifat yang disandi oleh gen tersebut. Beberapa fragmen
sekuen berulang (SSR) yang terletak di dalam atau berdekatan dengan gen
fungsional sudah ditemukan. Beberapa fragmen sekuen berulang (SSR) yang
terletak di dalam atau berdekatan dengan gen fungsional sudah ditemukan
dihasilkan dengan mengunakan informasi sekuen dari spesies lain (Rotwein et al.
1986; Shimatsu & Rotwein 1987).
Kemudahan dan kecepatan untuk mengetahui mikrosatelit, menggunakan
teknologi PCR, mudah diinterpretasikan, membuat mikrosatelit menjadi penanda
paling baik dalam pemetaan gen. Mikrosatelit memiliki polimorfisme yang tinggi
yang berasal dari berbagai populasi dan aksesi untuk meningkatkan kemungkinan
heterozigositas pada populasi F1. Kebanyakan mikrosatelit hipervariabel,
sehingga jumlah genotipe individu F2 untuk analisis segregasi dapat dikurangi.
Karakteristik mikrosatelit yang menarik ini telah mengurangi secara besar-besaran
penggunaan dari penand a RFLP dalam penelitian pemetaan genetik (Muladno,
2006).
Keanekaragaman mikrosatelit dapat dilihat dengan menggunakan PCR,
dimana sekuen unik digunakan sebagai primer-primer untuk mengamplifikasi
daerah mikrosatelit. Perbedaan pada ukuran fragmen DNA yang diamplifikasi
dapat divisualisasi dengan pewarnaan etidium bromida setelah elektroforesis
dengan gel agaros (Sambrook et al. 1989), atau pewarnaan perak dengan gel PAGE (Creste et al. 2001).
Sistem Aliran Informasi Genetik (Gene Flow)
Aliran gen melalui polinasi adalah proses alami yang karenanya gen-gen
berpindah di antara tanaman. Proses aliran gen dapat terjadi jika tanaman
domestik memiliki keserasian secara seksual dengan baik antara tanaman
domestik itu sendiri maupun kerabat liarnya. Pada tanaman proses ini dapat terjadi
melalui pemuliaan secara konvensional dan bioteknologi modern. Dengan
berkembangnya bioteknologi maka potensi terjadinya aliran gen dari suatu
tanaman ke tanaman lainnya ataupun kerabat liarnya menjadi sangat besar.
Aliran gen antar tanaman budidaya dengan kerabat liarnya adalah suatu
proses penting yang memiliki implikasi kuat terhadap konservasi keanekaragaman
genetik maupun pemuliaan tanaman. Introgresi antara populasi domestik dan
populasi liar merupakan suatu fenomena yang menyebar luas pada kebanyakan
seleksi pada struktur keanekaragaman genetik populasi liar dan domestik perlu
dipelajari.
Sepanjang evolusi yang termasuk di dalamnya penghanyutan genetik
(genetic drift), mutasi, seleksi maka proses migrasi juga menyumbang terjadinya
perubahan frekuensi gen dalam populasi alami. Penyimpangan antar populasi
dapat diakibatkan oleh penghanyutan genetik karena adanya isolasi parsial
ataupun penuh, atau oleh seleksi heterogenous. Migrasi individu (misalnya biji)
atau gamet (misalnya polen) antara subpopulasi menghalangi penyimpangan antar
populasi, menyebabkan reduksi dalam keragaman genetik antar subpopulasi, dan
meningkatkan tingkat keragaman genetik dalam subpopulasi.
Penghanyutan genetik dan migrasi adalah proses netral dan mempengaruhi
frekuensi gen pada populasi alami dan terjadi pada seluruh genom. Sedangkan
rekombinasi dan seleksi menginduksi terjadinya perubahan genetik hanya pada
lokus target (Cavalli-Sforza 1971; Lewontin 1988).
Aliran gen (Gene flow) adalah perpindahan informasi genetik melalui transportasi serbuk sari (penyebaran gamet jantan) dan transportasi melalui biji
atau benih (migrasi). Aliran gen berperan penting dalam distribusi informasi
genetik dalam populasi (intrapopulation gene flow) dan antar populasi (interpopulation gene flow). Efisiensi aliran gen melalui serbuk sari dan biji merupakan hal yang sangat penting berkaitan dengan ukuran populasi efektif
dalam bereproduksi. Ukuran populasi sangat penting untuk menggambarkan pola
penyebaran keragaman genetik dan perbedaan genetik di antara populasi
(Hamrick & Nason 2000, dan Gailing et al. 2003).
Banyaknya tanaman, jarak, serta arah aliran gen melalui serbuk sari dalam
setiap individu menentukan apakah perkawinan yang terjadi antar tanaman yang
berbeda (outcrossing) atau dalam tanaman itu sendiri (selfing).
Transfer serbuk sari pada tanaman kelapa masih dalam perdebatan.
Menurut Menon dan Pandalai (1960) dan Patel (1938) transfer serbuk sari
terutama melalui angin (anemophilous), sedangkan menurut Davis et al (1985) dan Ashburner et al (2001) terutama melalui serangga (entomophilous).
organisme tersebut sangat terbatas. Namun demikian setiap organisme mempunyai
potensi untuk menurunkan informasi genetik yang dimilikinya ke keturunannya melalui pertukaran garnet dan hal ini akan menghasilkan rekombinasi baru. Dengan demikian dinamika dari struktur genetik tidak dapat diamati ditingkat organisme
tunggal, tetapi diamati ditingkat populasi dimana setiap anggota dari populasi tersebut saling bertukar garnet.
Dinamika struktur genetik ditentukan dari komposisi gen berupa frekuensi alel dan frekuensi genotipe yang menyusun populasi tersebut. Penyebaran frekuensi dari
genotipe-genotipe dalam populasi disebut sebagai struktur genotipe dan penyebaran frekuensi dari alel-alel dalam satu populasi disebut struktur alel. Struktur genetik ini bersifat dinamik yaitu dalam kondisi kesetimbangan atau mengalami perubahan atau berevolusi bila terdapat kekuatan yang dapat merubah kesetimbangan seperti adanya mutasi, aliran gen (migrasi), penghanyutan genetik (genetic drift), seleksi dan model dari sistem perkawinan.
Untuk mempelajari terjadinya aliran gen pada populasi perlu
memperhatikan (1) antar tanaman yang dipelajari harus serasi secara seksual (sex compatible), ketidakserasian akan menyebabkan aliran gen tidak akan terjadi, (2) harus ada karakter yang berasosiasi dengan gen target. Modifikasi harus dapat
meningkatkan kemampun tanaman untuk bertahan (survive) dan berkembang biak