Karakteristik Genetik Serta Klasifikasi Isozyme Esterase Pada Tiga Spesies Ikan Batak di Kawasan SU
Bebas
21
0
0
Teks penuh
Gambar
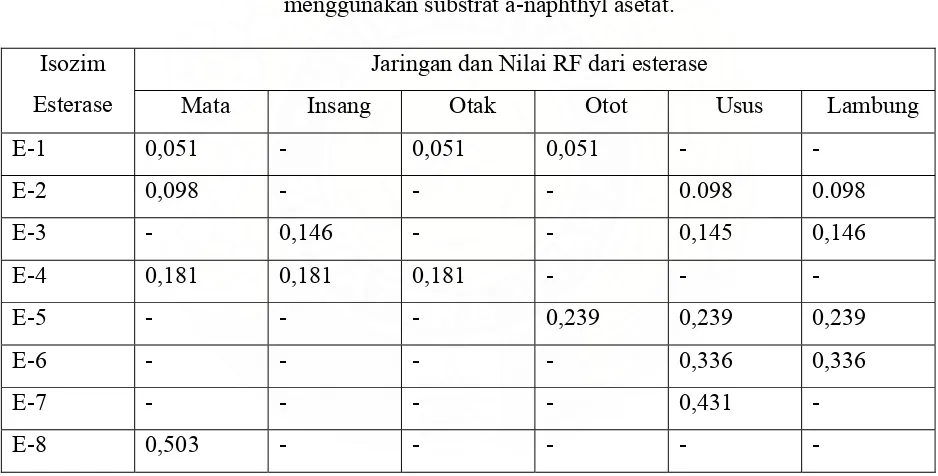
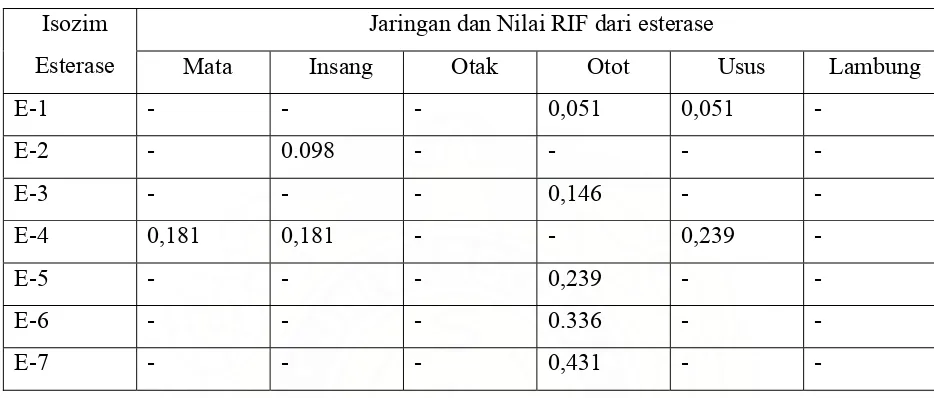
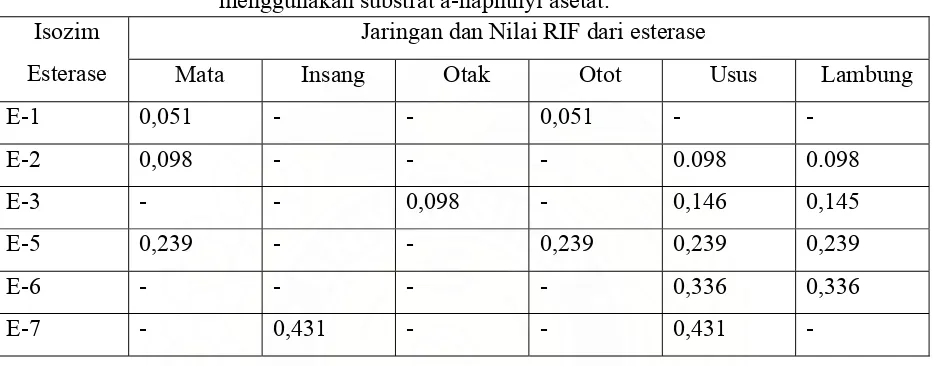
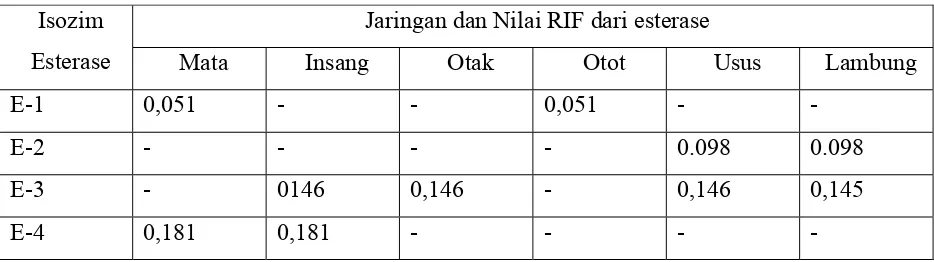
Dokumen terkait