[Glycine max (L.) Merr.] MELALUI IRADIASI SINAR GAMMA DOSIS RENDAH
DIANA SOFIA HANAFIAH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
iii
SURAT PERNYATAAN
Saya menyatakan dengan sebenar-benarnya, bahwa segala pernyataan dalam disertasi yang berjudul :
“PERBAIKAN KARAKTER AGRONOMI DAN ADAPTASI TERHADAP
CEKAMAN KEKERINGAN PADA KEDELAI [Glycine max (L.) Merr.]
MELALUI IRADIASI SINAR GAMMA DOSIS RENDAH”
adalah benar merupakan hasil karya saya sendiri dengan arahan komisi pembimbing. Semua data dan informasi yang dikutip dari karya yang diterbitkan oleh penulis lain telah disebutkan dalam teks dan dicantumkan dalam daftar pustaka.
Bogor, Agustus 2012
v ABSTRACT
DIANA SOFIA HANAFIAH. Agronomic Improvement Characters and Adaptation to Drought Stress on Soybean [Glycine max (L) Merr] through Low Doses of Gamma Rays Irradiation. Supervised by Trikoesoemaningtyas, Sudirman Yahya and Desta Wirnas.
The increase in genetic variability of soybean plants improve the probability to select for desired characters, for example, adaptation to drought stress. Mutation breeding is a breeding methods that can be applied to improve genetic variability of soybean in a relatively short time. Similarly, mutation induction can also increase genetic variability of soybean by using low doses of gamma rays irradiation. The objectives of the research were : (1) to obtain information on the changes in morphological and agronomic characters due to the treatment of low doses of gamma rays irradiation on plant population; (2) to obtain information on genetic variability among irradiation doses in plant population; (3) to obtain mutant putative genotype with good morphological and agronomic characters on optimum as well as stress condition; (4) to obtain information on the character of physiological adaptation to drought and plants; and (5) to obtain to a mutant putative line which can tolerate drought and high yielding. The study result showed that in Lethal Dose 50 (LD50) value there were 457,13 Gy. Gamma ray irradiation produced four populations that showed different growth and development from the non-irradiated control (Argomulyo). Genetic variability of each trait was different in each population at M2 generation. The selection based on number of productive pods and weight of seed per plant resulted in a mean improvement of selected genotype compared to the mean of base populations in M3 generation, respectively. Putative mutant lines on M4 that was from M3 selection had a mean improvement of plant height, number of productive nodes and number of productive pods better than that of Argomulyo variety. There were different responses of drought tolerant varieties (Ceneng and Tanggamus) and drought sensitive varieties (Anjasmoro and Argomulyo) to soil water content. The escape mechanism was found as earlier maturity in Argomulyo variety. Avoidance mechanism due to the increase in root volume was found in Anjasmoro, Ceneng and Tanggamus varieties at 40 % field capacity. Tolerance mechanism was marked by the reduction of evapotranspiration in Tanggamus variety at 40 % field capacity. The research results showed that the performance of agronomic characters on drought condition were lower than that of in optimum condition in M5 population. The selection result on M5 generation indicated that the selected mutant strains had good agronomic characters and high yielding. They had good adaptability to drought and were high yielding in optimum conditions and drought conditions.
vii RINGKASAN
DIANA SOFIA HANAFIAH. Perbaikan Karakter Agronomi dan Adaptasi terhadap Cekaman Kekeringan pada Kedelai [Glycine max (L.) Merr.] Melalui Iradiasi Sinar Gamma Dosis Rendah. Dibimbing oleh Trikoesoemaningtyas, Sudirman Yahya dan Desta Wirnas.
Peningkatan keragaman genetik tanaman kedelai akan mempermudah usaha dalam menyeleksi tanaman untuk mendapatkan suatu tanaman dengan sifat yang diinginkan, misalnya karakter tanaman untuk ketahanan terhadap cekaman kekeringan. Pada penelitian ini diharapkan diperoleh galur tanaman yang toleran kekeringan dan berdaya hasil tinggi melalui pemuliaan mutasi dengan menggunakan iradiasi sinar gamma dosis rendah. Tujuan penelitian ini adalah : (1) memperoleh informasi tentang perubahan karakter morfologi dan agronomi yang disebabkan perlakuan iradiasi sinar gamma dosis rendah dalam populasi tanaman; (2) memperoleh informasi tentang keragaman genetik antar perlakuan dosis iradiasi dalam populasi tanaman; (3) memperoleh genotip putatif mutan dengan karakter morfologi dan agronomi baik pada kondisi cekaman kekeringan dan optimum; (4) memperoleh informasi tentang karakter fisiologi adaptasi kekeringan pada tanaman; dan (5) mendapatkan galur putatif mutan yang toleran kekeringan dan berdaya hasil tinggi.
Hasil penelitian ini menunjukkan bahwa nilai Lethal Doses 50 (LD50) terdapat pada dosis 457,13 Gy. Iradiasi sinar gamma menghasilkan empat populasi iradiasi yang menunjukkan pertumbuhan dan perkembangan yang berbeda pada setiap karakter agronomi dengan populasi kontrol (Argomulyo).
Hasil penelitian ini menunjukkan bahwa keragaman genetik yang dihasilkan pada setiap karakter berbeda-beda dari masing-masing populasi iradiasi pada generasi M2. Seleksi individu terpilih berdasarkan jumlah polong bernas dan bobot biji per tanaman menghasilkan perbaikan nilai tengah genotipe terpilih dibandingkan nilai tengah populasi dasarnya dari generasi M3. Galur putatif mutan M4 yang berasal dari seleksi M3 memiliki nilai tengah karakter tinggi tanaman, jumlah buku produktif dan jumlah polong bernas yang lebih baik dibanding varietas asal Argomulyo.
Hasil penelitian menunjukkan terdapat respon yang berbeda dari masing-masing varietas peka kekeringan (Anjasmoro dan Argomulyo) dan varietas toleran kekeringan (Ceneng dan Tanggamus) terhadap ketersediaan air. Mekanisme
escape terjadi dengan adanya karakter umur panen yang genjah pada varietas
Argomulyo. Mekanisme avoidance terjadi dengan adanya kemampuan
mempertahankan volume akar pada varietas Anjasmoro, Ceneng dan Tanggamus pada ketersediaan air 40% KL. Mekanisme toleransi ditandai dengan nilai evapotranspirasi rendah pada varietas Tanggamus pada ketersediaan air 40% KL.
Hasil penelitian ini juga menunjukkan bahwa penampilan karakter agronomi mutan pada kondisi kekeringan tidak sebaik pada kondisi optimum pada populasi M5. Hasil seleksi generasi M5 diperoleh galur-galur mutan terpilih yang mempunyai karakter agronomi baik serta berdaya hasil tinggi dan galur-galur mutan terpilih yang mempunyai karakter adaptasi terhadap kekeringan dan berdaya hasil tinggi pada pada kondisi optimum dan kondisi kekeringan.
ix
© Hak cipta milik Institut Pertanian Bogor, tahun 2012 Hak cipta dilindungi Undang-undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumber
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau tinjauan suatu masalah
b. Pengutipan tidak merugikan kepentingan yang wajar IPB
xi
PERBAIKAN KARAKTER AGRONOMI DAN ADAPTASI TERHADAP CEKAMAN KEKERINGAN PADA KEDELAI
[Glycine max (L.) Merr.] MELALUI IRADIASI SINAR GAMMA DOSIS RENDAH
DIANA SOFIA HANAFIAH
Disertasi
Sebagai salah satu syarat memperoleh Gelar Doktor
pada Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
xii
Penguji Luar Komisi
Pada Ujian Tertutup : Jumat 6 juli 2012
1. Prof. Dr. Ir. Munif Ghumaladi
2. Dr. Ir. Yudiwanti Wahyu EK, MSi
Penguji Luar Komisi
Pada Ujian Terbuka : Selasa 24 juli 2012
1. Dr. Ir. Syarifah Iis Aisyah, MSc. Agr.
xiii
Judul Disertasi : Perbaikan Karakter Agronomi dan Adaptasi terhadap Cekaman Kekeringan pada Kedelai [Glycine max (L.) Merr.] Melalui Iradiasi Sinar Gamma Dosis Rendah
Nama Mahasiswa : Diana Sofia Hanafiah
Nomor Pokok : A263070041
Disetujui, Komisi Pembimbing
Dr. Ir. Trikoesoemaningtyas, M.Sc. Ketua
Prof. Dr. Ir. Sudirman Yahya, M.Sc. Dr. Desta Wirnas, SP, M.Si
Anggota Anggota
Diketahui,
Koordinator Program Studi Dekan Sekolah Pascasarjana Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, M.Sc. Dr. Ir. Dahrul Syah, MSc. Agr.
xv PRAKATA
Syukur alhamdulillah, penulis panjatkan kehadlirat Allah SWT yang telah nemberikan rahmat dan karunia-Nya, sehingga penulis dapat menyelesaikan penelitian
dan penulisan disertasi yang berjudul : “Perbaikan Karakter Agronomi dan Adaptasi terhadap Cekaman Kekeringan pada Kedelai [Glycine max (L.) Merr.] Melalui Iradiasi Sinar Gamma Dosis Rendah.
Penelitian dan penulisan disertasi ini berlangsung dibawah bimbingan Dr. Ir. Trikoesoemaningtyas, M.Sc. selaku ketua komisi pembimbing dan Prof. Dr. Ir. Sudirman Yahya, MSc, serta Dr. Desta Wirnas, SP, M.Si selaku anggota komisi pembimbing. Pada kesempatan ini penulis menyampaikan terima kasih dan penghargaan yang tulus atas waktu dan kesempatan yang telah diluangkan dalam mengarahkan dan membimbing penulis.
Penelitian dan penulisan disertasi ini didanai oleh Direktorat Jendral Pendidikan Tinggi, Kementrian Pendidikan Nasional melalui beasiswa BPPS tahun 2007-2010, Hibah SINTA tahun 2009 dan Penelitian IMHERE tahun 2010 serta dari Rektor Universitas Sumatera Utara (USU). Oleh karena itu penulis menyampaikan terima kasih kepada Dirjen Dikti dan Rektor USU selaku pemberi dana, sehingga penelitian dan penulisan disertasi ini dapat diselesaikan.
Penghargaan yang setingi-tingginya serta terima kasih yang tulus, penulis sampaikan juga kepada :
1. Dirjen Direktorat Pendidikan Tinggi (DIKTI) yang telah memberikan beasiswa BPPS.
2. Rektor Universitas Sumatera Utara (USU) dan Dekan Fakultas Pertanian USU yang telah memberikan izin tugas belajar.
3. Rektor Institut Pertanian Bogor (IPB), Dekan Sekolah Pascasarjana IPB dan Ketua Program Studi Pemuliaan dan Bioteknologi Tanaman yang telah menerima penulis untuk melanjutkan studi di Institut Pertanian Bogor.
4. Kepala dan Staf Laboratorium Iradiasi Pusat Antar Universitas (PAU) IPB atas bantuan dan kerjasamanya.
xvi
6. Kepala dan Staf Kebun Percobaan Balai Penelitian Kacang-kacangan dan umbi-umbian (BALITKABI) Malang atas bantuan dan kerjasamanya.
7. Kepala dan Staf Laboratorium Research Group on Crop Improvement (RGCI) Faperta IPB atas bantuan dan kerjasamanya.
8. Kepala dan Staf Laboratorium Pemuliaan Tanaman Faperta IPB atas bantuan dan kerjasamanya.
9. Teman-teman seperjuangan pada Mayor PBT dan Mayor AGH IPB atas persahabatan dan kebersamaannya.
10. Ayahanda Alm. Prof. Dr. Ir. H. T.M. Hanafiah Oeliem, DAA dan ibunda Prof. Dr. Ir. Hj. Asmarlaili S. Hanafiah, MS, DAA yang telah membesarkan, mendidik, mendoakan dan menyertai penulis dengan penuh kesabaran serta kasih sayangnya. 11. Nenek Hj. Rosiah Sahar; Bibi drg. Rosmawati dan Om dr.Sona, Sp. THT,
Kakanda drg. Olivia Avriyanti Hanafiah, Sp.BM dan keluarga; adinda Ully Irma Maulina Hanafiah, ST, MT dan keluarga; dan adinda dr. Ghamal Arief Hanafiah dan keluarga atas doa, bantuan, perhatian dan motivasinya
12. Suami tercinta dan anak-anak tersayang atas doa, dorongan, pengertian dan pengorbanannya.
13. Semua pihak yang telah memberikan dukungan dan bantuannya.
Semoga karya ini bermanfaat bagi pengembangan ilmu pengetahuan, khususnya di bidang pertanian. Amin.
Bogor, Agustus 2012
xvii
RIWAYAT HIDUP
Penulis dilahirkan di Medan pada tanggal 30 Agustus 1974 sebagai anak kedua dari empat bersaudara dari ayah Alm. Prof. Dr. Ir. T.M. Hanafiah Oeliem, DAA dan ibu Prof. Dr. Ir. Asmarlaili S. Hanafiah, MS, DAA. Penulis menikah dengan Dr. Muhdi, S.Hut, M.Si dan telah dikarunia tiga orang putra dan putri yaitu Muhammad Hafizh Abdurrohman, Muhammad Hafazh Abdurrohim dan Shadrina Alyani.
Pendidikan sarjana pertanian ditempuh di Program Studi Pemuliaan Tanaman, Jurusan Budidaya Pertanian, Fakultas Pertanian Universitas Sumatera Utara, lulus pada tahun 1997. Jenjang Magister diikuti pada Program Pascasarjana Universitas Sumatera Utara Program Studi Agronomi, lulus pada tahun 2001. Selanjutnya, pada tahun 2007 mengikuti pendidikan program doktor pada Program Studi Pemuliaan dan Bioteknologi Tanaman, Sekolah Pascasarjana, Institut Pertanian Bogor melalui beasiswa BPPS dari Kementrian Pendidikan Nasional RI. Penulis adalah staf pengajar Fakultas Pertanian Universitas Sumatera Utara sejak tahun 1999 sampai sekarang.
Sebagian dari disertasi ini telah dipresentasikan dalam Seminar Nasional Perhimpunan Agronomi Indonesia (PERAGI) 2009 di Fakultas Pertanian Universitas Sumatera Utara dengan judul Agronomic Improvement of Argomulyo Soybean Variety [Glycine max (L) Merr] Throught Induced Mutation by Gamma Irradiation in M1 AND M2 Generation dan
xix DAFTAR ISI
Hal.
KATA PENGANTAR ... xi
DAFTAR ISI ... xiii
DAFTAR TABEL ... xvi
DAFTAR GAMBAR ... xix
DAFTAR LAMPIRAN ... xxi
PENDAHULUAN ... 1
Latar Belakang ... 1
Perumusan Masalah ... 3
Tujuan Penelitian ... 4
Alur Penelitian ... 4
TINJAUAN PUSTAKA ... 7
Asal Usul dan Taksonomi Tanaman Kedelai ... 7
Prospek dan Arah Pengembangan Kedelai... ... 7
Usaha Peningkatan Keragaman Genetik Tanaman Kedelai ... 9
Cekaman Kekeringan pada Tanaman Kedelai ... 11
Mekanisme Adaptasi terhadap Kekeringan ... 13
Pemuliaan Bagi Lingkungan Bercekaman ... 15
RADIOSENSITIVITAS DAN PENGARUH IRADIASI DOSIS RENDAH SINAR GAMMA PADA VARIETAS ARGOMULYO ... 19
ABSTRAK... 19
ABSTRACT ... 19
PENDAHULUAN ... 19
METODE PENELITIAN ... 21
HASIL PENELITIAN ... 21
PEMBAHASAN ... 23
KESIMPULAN ... 25
KERAGAMAN KARAKTER AGRONOMI DAN SELEKSI GENOTIPE PUTATIF MUTAN HASIL IRADIASI SINAR GAMMA DOSIS RENDAH ... 27
ABSTRAK... 27
ABSTRACT ... 27
PENDAHULUAN ... 28
METODE PENELITIAN ... 29
HASIL PENELITIAN ... 33
PEMBAHASAN ... 46
xx
KERAGAAN GENERASI M3 PADA KONDISI OPTIMUM DAN KONDISI
CEKAMAN KEKERINGAN ... 53
ABSTRAK... 53
ABSTRACT ... 53
PENDAHULUAN ... 54
METODE PENELITIAN ... 55
HASIL PENELITIAN ... 57
PEMBAHASAN ... 62
KESIMPULAN ... 64
MEKANISME ADAPTASI KEDELAI TERHADAP KEKERINGAN ... 65
ABSTRAK... 65
ABSTRACT ... 65
PENDAHULUAN ... 65
METODE PENELITIAN ... 67
HASIL PENELITIAN ... 71
PEMBAHASAN ... 86
KESIMPULAN ... 92
SELEKSI PADA POPULASI M5 UNTUK KARAKTER ADAPTASI KEKERINGAN DAN DAYA HASIL TINGGI ... 93
ABSTRAK... 93
ABSTRACT ... 93
PENDAHULUAN ... 94
METODE PENELITIAN ... 96
HASIL PENELITIAN ... 97
PEMBAHASAN ... 104
KESIMPULAN ... 106
PEMBAHASAN UMUM ... 107
KESIMPULAN DAN SARAN ... 115
DAFTAR PUSTAKA ... 117
xxi
DAFTAR TABEL
. Hal.
1. Persentase perkecambahan dan tinggi kecambah dua minggu setelah tanam. .. 22 2. Pengaruh iradiasi sinar gamma terhadap karakter agronomi varietas
Argomulyo ... 33 3. Nilai tengah karakter agronomi populasi M2 hasil iradiasi sinar gamma
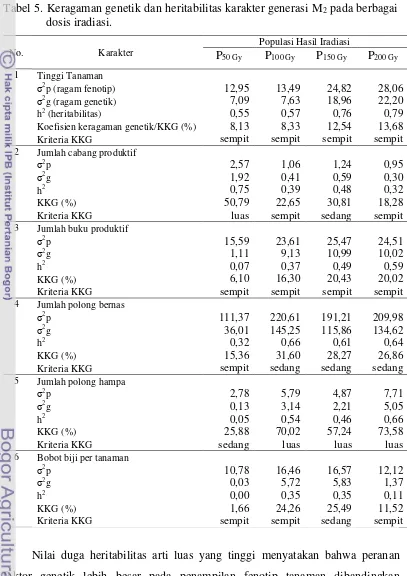
dosis rendah.. ... 37 4. Kisaran nilai karakter agronomi populasi M2 hasil iradiasi sinar gamma. ... 38 5. Keragaman genetik dan heritabilitas karakter generasi M2 pada berbagai dosis iradiasi. ... 39 6. Nilai tengah karakter agronomi populasi M3 hasil iradiasi sinar gamma
dari varietas Argomulyo ... 40 7. Kisaran nilai karakter agronomi populasi M3 hasil iradiasi sinar gamma ... 41 8. Komponen ragam dan keragaman karakter tinggi tanaman, jumlah cabang produktif, dan jumlah buku produktif kedelai generasi M3 hasil iradiasi sinar gamma dari varietas Argomulyo pada kondisi optimum ... 42 9. Kemajuan genetik akibat seleksi genotipe kedelai generasi M3 pada kondisi optimum ... 43 10. Nilai tengah karakter agronomi 291 galur putatif mutan, varietas Anjasmoro,
Argomulyo, Ceneng dan Tanggamus pada kondisi optimum ... 44 11. Pendugaan ragam, koefisien keragaman genetik dan heritabilitas arti
luas karakter agronomi populasi M4 hasil iradiasi sinar gamma dari varietas Argomulyo ... 45 12. Nilai tengah karakter agronomi 150 galur putatif mutan terpilih, varietas
Anjasmoro, Argomulyo, Ceneng dan Tanggamus pada kondisi optimum ... 45 13. Nilai tengah karakter agronomi populasi M3 hasil iradiasi sinar gamma
pada kondisi optimum dan pada kondisi kekeringan ... 57 14. Komponen ragam dan keragaman karakter tinggi tanaman, jumlah cabang produktif, dan jumlah buku produktif kedelai generasi M3 hasil iradiasi sinar gamma pada kondisi optimum dan kekeringan ... 59 15. Kemajuan genetik genotipe kedelai generasi M3 hasil seleksi ketahanan
terhadap kondisi optimum dan kekeringan ... 61 16. Karakter luas daun trifoliat varietas kedelai pada berbagai ketersediaan
air. ... 71 17. Karakter kerapatan stomata varietas kedelai pada berbagai ketersediaan
xxii
19. Karakter panjang akar varietas kedelai pada berbagai ketersediaan air ... 74 20. Karakter bobot kering akar varietas kedelai pada berbagai ketersediaan air .... 74 21. Karakter bobot kering total varietas kedelai pada berbagai ketersediaan air .... 75 22. Perubahan terhadap kontrol (%) pada karakter bobot kering total ... 75 23. Pengaruh cekaman air terhadap karakter volume akar ... 77 24. Karakter nisbah tajuk akar varietas kedelai pada berbagai ketersediaan air. .... 78 25. Karakter kehijauan daun saat awal cekaman dan pada tahap R6 pada
berbagai ketersediaan air ... 79 26. Karakter kandungan klorofil a dan klorofil b varietas kedelai pada
berbagai ketersediaan air ... 80 27. Pengaruh cekaman air terhadap karakter evapotranspirasi ... 81 28. Karakter efisiensi penggunaan air varietas kedelai pada berbagai
ketersediaan air. ... 82 29. Karakter kandungan gula total varietas kedelai pada berbagai ketersediaan air. ... 83 30. Karakter umur panen varietas kedelai pada berbagai ketersediaan air ... 84 31. Karakter bobot biji per tanaman varietas kedelai pada berbagai
ketersediaan air ... 85 32. Keragaan karakter agronomi 150 galur putatif mutan, varietas Anjasmoro,
Argomulyo, Ceneng dan Tanggamus pada kondisi optimum dan kekeringan. ... 97 33. Karakter kehijauan daun R1/awal cekaman dan kehijauan daun R6 150 galur
putatif mutan, varietas Anjasmoro, Argomulyo, Ceneng dan Tanggamus
pada kondisi optimum dan kekeringan... 98 34. Pendugaan ragam, koefisien keragaman genetik dan heritabilitas arti luas
karakter agronomi populasi M5 pada lingkungan optimum dan kekeringan ... 100 35. Koefisien korelasi parsial antar karakter agronomi populasi M5 pada kondisi kekeringan ... 101 36. Nilai tengah karakter agronomi populasi dasar M5, galur mutan hasil seleksi
dan varietas Argomulyo pada kondisi optimum. ... 101 37. Keragaan karakter agronomi 30 galur putatif mutan hasil seleksi pada kondisi
optimum ... 102 38. Nilai tengah karakter agronomi populasi dasar M5, galur mutan hasil seleksi
dan varietas Argomulyo pada kondisi kekeringan ... 103 39. Keragaan karakter agronomi 31 galur putatif mutan hasil seleksi berdasarkan
indeks sensitifitas kekeringan pada kondisi kekeringan ... 103 40. Differensial seleksi dari indeks terboboti, karakter bobot biji per tanaman
xxiii
DAFTAR GAMBAR
Hal. 1. Bagan alir penelitian ... 5 2. Perkecambahan tanaman kedelai varietas Argomulyo yang diiradiasi sinar gamma pada dosis 0, 200, 400, 600, 800 dan 1000 Gy... 22 3. Grafik pengaruh iradiasi sinar gamma terhadap persentase perkecambahan.... 23 4. Hubungan antara dosis iradiasi (Gy) dengan tinggi tanaman (cm) pada
generasi M1... 34 5. Rasim bunga dan tunas tidak berkembang. ... 34 6. Hubungan antara dosis iradiasi (Gy) dengan jumlah polong bernas pada
generasi M1... 35 7. Hubungan antara dosis iradiasi (Gy) dengan jumlah biji per tanaman
pada generasi M1 ... 35 8. (a). Daun trifoliate dan oval varietas Argomulyo (b). Daun bifoliat dan
memanjang tanaman hasil iradiasi (c). Daun unifoliat dan memanjang tanaman hasil iradiasi (d).Rasim bunga dan tunas (tidak berkembang) tanaman hasil iradiasi (e). Bunga ungu (varietas Argomulyo) (f).Bunga
putih (tanaman hasil iradiasi) ... 36 9. Hubungan antara volume akar (ml) varietas kedelai dengan persentase
ketersediaan air... 76 10. Hubungan antara evapotranspirasi varietas kedelai dengan ketersediaan
xxv
DAFTAR LAMPIRAN
Hal. 1. Hasil analisis ragam pengaruh varietas terhadap luas daun trifoliat, stomata,
bobot kering tajuk, panjang akar, bobot kering akar, dan rasio tajuk-akar pada berbagai ketersediaan air ... 129 2. Hasil analisis ragam pengaruh varietas terhadap kehijauan daun saat awal
cekaman dan tahap R6 padaberbagai ketersediaan air ... 129 3. Hasil analisis ragam pengaruh varietas terhadap klorofil a, klorofil b,
evapotranspirasi, efisiensi penggunaan air dan gula total pada berbagai
ketersediaan air ... 129 4. Hasil analisis polinomial ortogonal pengaruh varietas terhadap karakter
volume akar dan evapotranspirasi pada berbagai ketersediaan air ... 130 5. Hasil analisis ragam pengaruh varietas terhadap karakter umur panen dan
bobot biji per tanaman pada berbagai ketersediaan air ... 130 6. Koefisien korelasi parsial antar karakter pada semua varietas... 131 7. Koefisien korelasi parsial antar karakter pada Anjasmoro ... 132 8. Koefisien korelasi parsial antar karakter pada Argomulyo ... 133 9. Koefisien korelasi parsial antar karakter pada Ceneng ... 134 10. Koefisien korelasi parsial antar karakter pada Tanggamus. ... 135 11. Nilai persentase perubahan terhadap kontrol pada varietas Anjasmoro ... 136 12. Nilai persentase perubahan terhadap kontrol pada varietas Argomulyo ... 136 13. Nilai persentase perubahan terhadap kontrol pada varietas Ceneng ... 137 14. Nilai persentase perubahan terhadap kontrol pada varietas Tanggamus ... 137 15. Penetapan kadar air tanah melalui metode pengeringan (oven) ... 138 16. Penetapan kadar air pada kapasitas lapang dengan metode Bouyoucos
dan penetapan tanah kering udara dalam pot berdasarkan kadar air
PENDAHULUAN
Latar belakang
Kedelai merupakan tanaman pangan terpenting ke tiga setelah padi dan jagung. Perkembangan industri pangan berbahan baku kedelai telah menyebabkan permintaan kedelai terus meningkat melampaui produksi dalam negeri (Sudaryanto dan Swastika 2007). Sampai saat ini hanya 20 – 30 % kebutuhan nasional yang dapat dipenuhi oleh produksi kedelai nasional, sedangkan sisanya dipenuhi dengan melakukan impor kedelai (Purna et al. 2009). Produksi kedelai nasional tahun 2011 (ARAM II) diperkirakan mencapai 819,45 ribu ton biji kering yang menurun sebanyak 87,59 ribu ton dibandingkan dengan produksi kedelai tahun 2010. Penurunan produksi kedelai tahun 2011 diperkirakan terjadi di Jawa sebesar 85,25 ribu ton dan di luar Jawa sebesar 2,34 ribu ton. Salah satu penyebab penurunan produksi kedelai adalah penurunan luas panen sebesar 68,79 ribu hektar (BPS 2011).
Penurunan luas areal tanam kedelai yang cukup tajam terjadi di pulau Sumatera sebesar 21,1% dari tahun 1992 sampai 2003. Pulau Jawa masih tetap merupakan sentra produksi dengan peningkatan luas lahan sebesar 18,25 % dari tahun 1992 sampai 2003. Posisi Sumatera pada tahun 2003 diambil alih oleh Bali dan NTB dengan luas areal tanam pada tahun 1992 sebesar 9,15 % meningkat menjadi 14,04 % pada tahun 2003 (Sudaryanto dan Swastika 2007).
Usaha meningkatkan produksi kedelai dapat dilakukan melalui peningkatan produktivitas dan perluasan areal tanam. Upaya peningkatan produktivitas kedelai di lahan kering dapat ditempuh melalui penggunaan teknik budidaya yang sesuai dan penanaman varietas unggul. Teknik budidaya digunakan berdasarkan agroekologi daerah. Peningkatan produktivitas dapat diupayakan melalui penggunaan varietas unggul yang adaptif dan berpotensi hasil baik terhadap agroekologi daerah (Arsyad et al. 2007).
Cekaman kekeringan pada tanaman dapat terjadi karena kekurangan suplai air di daerah perakaran dan laju evapotranspirasi melebihi laju absorbsi air di perakaran (Levitt 1980).
Varietas unggul diperoleh melalui pemuliaan tanaman dengan melakukan perbaikan daya hasil dan adaptasi tanaman. Perakitan varietas baru memerlukan populasi dasar yang memiliki keragaman genetik yang tinggi. Keragaman genetik kedelai di Indonesia rendah, sehingga perlu upaya peningkatan keragaman genetik tanaman. Upaya peningkatan keragaman genetik kedelai dapat dilakukan melalui introduksi, persilangan, transformasi genetik dan mutasi (Arsyad et al.
2007).
Pemuliaan mutasi berguna untuk memperbaiki karakter tanaman jika karakter yang diinginkan tidak terdapat pada suatu plasma nutfah suatu spesies tanaman. Induksi mutasi dilakukan dengan menggunakan mutagen. Mutagen terdiri dari mutagen kimia dan fisik. Salah satu mutagen fisik adalah iradiasi sinar gamma. Mutasi iradiasi dapat dilakukan secara mutasi makro (macro mutation) dan mutasi secara mikro (micro mutation). Mutasi makro menggunakan dosis iradiasi yang tinggi yaitu dosis yang berada diatas
Lethal Doses50, biasanya menyebabkan ketidakstabilan genetik. Adapun mutasi mikro yaitu mutasi yang menggunakan dosis rendah yaitu dosis di bawah LD50 (Van Harten 1998).
Iradiasi sinar gamma dosis rendah (mutasi mikro) telah digunakan untuk meningkatkan keragaman genetik, karakter kuantitatif dan daya adaptasi serta meningkatkan kualitas dan nutrisi dari beberapa tanaman kacang-kacangan (Manjaya dan Nandanwar 2007; Kaveri dan Nadaf 2009; Diouf et al. 2010; Moussa 2011). Mutasi mikro mengubah karakter kuantitatif yang diturunkan dan lebih bermanfaat bagi pemulia, karena mutasi mikro sedikit merusak walaupun mutasi ini sulit dideteksi (Sakin 2002).
Perumusan Masalah
Masalah yang dihadapi dalam pengembangan kedelai untuk toleransi terhadap kekeringan adalah masih terbatasnya karakter-karakter tertentu seperti karakter adaptasi terhadap kekeringan pada varietas kedelai dengan ukuran biji besar seperti varietas Argomulyo pada plasma nutfah varietas kedelai lokal Indonesia. Untuk meningkatkan keragaman genetik tanaman kedelai agar dapat beradaptasi dengan agroekologi di Indonesia dilakukan cara antara lain dengan mutasi. Pemuliaan mutasi berguna untuk memperbaiki karakter tanaman jika karakter yang diinginkan tidak terdapat pada suatu plasma nutfah suatu spesies tanaman.
Mutasi dapat dilakukan dengan menggunakan sinar gamma. Pengaruh dari iradiasi sinar gamma pada bagian tanaman seperti tunas, biji dan serbuk sari dapat diukur dari nilai radiosensitivitas yang diperoleh. Radiosensitivitas dari setiap bagian
tanaman yang diiradiasi berbeda-beda pada masing-masing tanaman.
Radiosensitivitas menentukan nilai Lethal Dose 50 (LD50) dari pengaruh iradiasi sinar gamma. Nilai LD50 digunakan untuk menentukan dosis iradiasi sinar gamma yang digunakan pada penelitian ini.
Pada penelitian ini digunakan iradiasi sinar gamma dosis rendah (mutasi mikro) yang diharapkan dapat menginduksi keragaman genetik pada kedelai varietas Argomulyo. Iradiasi sinar gamma dosis rendah menggunakan dosis di bawah LD50, mempengaruhi perubahan karakter kuantitatif tanaman dan sedikit mempengaruhi perubahan kromosom dibandingkan dengan mutasi makro.
Keragaman genetik yang terdapat pada populasi tanaman hasil iradiasi merupakan dasar untuk dilakukan seleksi tanaman terpilih yang berdaya hasil tinggi. Seleksi pada populasi hasil iradiasi lebih efektif dilakukan pada generasi setelah M2, dimana gen-gen minor yang bersifat kuantitatif telah terfiksasi pada generasi lanjut (Van Harten 1998).
tidak langsung menggunakan beberapa karakter serta mempunyai nilai heritabilitas rendah.
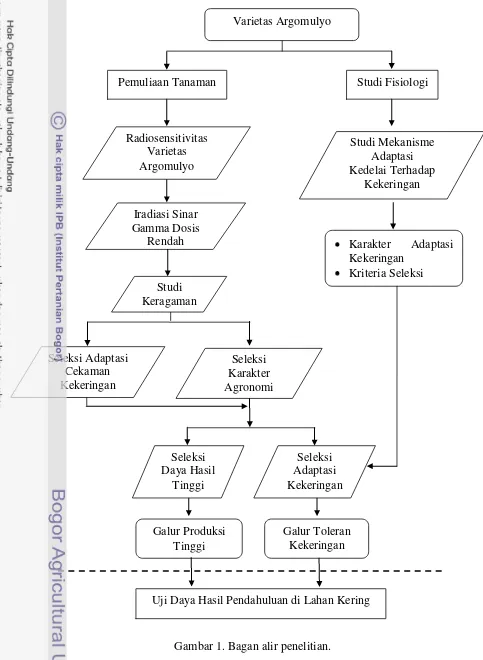
Pada penelitian ini diharapkan memperoleh galur kedelai yang toleran kekeringan dan berdaya hasil tinggi melalui pemuliaan mutasi dengan iradiasi sinar gamma dosis rendah. Kegiatan yang dilakukan dalam penelitian ini terdiri atas studi radiosensitivitas varietas Argomulyo, pengaruh iradiasi sinar gamma dosis rendah, studi keragaman pada populasi hasil iradiasi, studi mekanisme adaptasi kedelai terhadap kekeringan dan seleksi galur-galur toleran kekeringan dan berdaya hasil tinggi (Gambar 1).
Tujuan Penelitian
Tujuan utama penelitian ini untuk memperoleh galur-galur mutan toleran kekeringan serta berdaya hasil tinggi melalui iradiasi sinar gamma dosis rendah.
Tujuan khusus penelitian ini adalah :
1. Memperoleh informasi tentang perubahan karakter morfologi dan agronomi yang disebabkan perlakuan iradiasi sinar gamma dosis rendah dalam populasi tanaman. 2. Memperoleh informasi tentang keragaman genetik antar perlakuan dosis iradiasi
dalam populasi tanaman
3. Memperoleh genotip putatif mutan dengan karakter morfologi dan agronomi baik pada kondisi cekaman kekeringan dan optimum.
4. Memperoleh informasi tentang karakter fisiologi adaptasi kekeringan pada tanaman
5. Mendapatkan galur putatif mutan yang toleran kekeringan dan berdaya hasil tinggi.
Alur Penelitian
[image:30.595.54.484.2.818.2]Gambar 1. Bagan alir penelitian. Varietas Argomulyo
Studi Fisiologi Pemuliaan Tanaman
Studi Mekanisme Adaptasi Kedelai Terhadap
Kekeringan
Karakter Adaptasi Kekeringan
Kriteria Seleksi Radiosensitivitas
Varietas Argomulyo
Iradiasi Sinar Gamma Dosis
Rendah
Seleksi Adaptasi Cekaman Kekeringan
Seleksi Karakter Agronomi
Seleksi Daya Hasil
Tinggi
Seleksi Adaptasi Kekeringan Studi
Keragaman
Galur Toleran Kekeringan
Toleran Galur Produksi
Tinggi
TINJAUAN PUSTAKA
Asal Usul dan Taksonomi Tanaman Kedelai
Kedelai (Glycine max) bukan tanaman asli Indonesia dan diduga berasal dari daratan pusat dan utara Cina. Hal ini didasarkan pada adanya penyebaran Glycine ussuriensis, spesies yang diduga sebagai tetua Glycine max. Bukti sitogenetik menunjukkan bahwa Glycine max dan Glycine ussuriensis tergolong spesies yang sama. Penyebaran kedelai di kawasan Asia, khususnya Jepang, Indonesia, Filipina, Vietnam, Thailand, Malaysia, Birma, Nepal dan India dimulai sejak pada abad pertama setelah masehi bersamaan dengan semakin berkembangnya jalur perdagangan lewat darat dan laut (Adie dan Krisnawati 2007).
Klasifikasi kedelai yang disempurnakan oleh Verdcourt (1966) yang mengklasifikasikan kedelai menjadi tiga subgenus yaitu Glycine, Bracteata dan Soja. Sub genus Soja umumnya merupakan tanaman semusim. Spesies Glycine ussuriensis
dan Glycine max memiliki jumlah kromosom somatik 2 n = 40. Klasifikasi dari Glycine max (L.) Merrill adalah :
Ordo : Polypetales
Famili : Leguminosae
Sub-famili : Papilionoideae
Genus : Glycine
Sub-genus : Soja
Species : max
G.ussuriensis merupakan tanaman semusim, batangnya menjalar, daun berukuran kecil dan berbentuk lancip, bunga berwarna ungu, biji keras berwarna hitam hingga coklat tua. G.ussuriensis lebih dikenal sebagai kedelai liar (wild soybean). G.max adalah kedelai yang telah dibudidayakan dan merupakan tanaman semusim, warna bunga putih atau ungu, dan memiliki ragam bentuk dan ukuran untuk karakter daun dan biji (Adie dan Krisnawati 2007).
Prospek dan Arah Pengembangan Kedelai
areal tanam dan penanaman varietas unggul sesuai dengan agroekologi yang menjadi target area produksi (Arsyad et al. 2007).
Selain dengan memanfaatkan lahan sawah, peningkatan luas area panen dapat dilakukan dengan memanfaatkan lahan kering (tegalan), lahan perkebunan, lahan bukaan baru, lahan gambut dan lahan pasang surut yang telah direklamasi. Secara umum pengembangan kedelai dapat ditujukan pada berbagai provinsi yang pernah menjadi sentra produksi kedelai, antara lain Nangroe Aceh Darussalam, Sumatera Utara, Lampung, Sulawesi Selatan dan Sulawesi Tenggara (Rachman et al. 2007; Sumarno dan Manshuri 2007). Perluasan areal tanaman kedelai melalui pemanfaatan lahan dengan keragaman agroekosistem harus didukung oleh varietas yang sesuai untuk masing agroekosistem (Arsyad et al. 2007).
Pengembangan varietas kedelai juga harus diarah untuk perbaikan kualitas sesuai kebutuhan konsumen baik rumah maupun industri. Perbaikan kualitas diarahkan untuk perbaikan ukuran biji, kandungan protein, kandungan asam amino tertentu serta penurunan kandungan anti nutrisi.
Saat ini secara pertanian mendapat tekanan sebagai dampak dari pemanasan global. Pemanasan global adalah kejadian terperangkapnya radiasi gelombang panjang matahari (infra merah atau gelombang panas) yang dipancarkan oleh bumi, sehingga tidak dapat lepas ke angkasa dan akibatnya suhu di atmospher bumi memanas (Hairiah 2008).
Penjebak gelombang panas tersebut adalah lapisan gas yang berperan seperti
dinding kaca atau ‘selimut tebal’, antara lain adalah uap air, gas asam arang atau
karbon dioksida (CO2), gas methana (CH4), dinitrogen oksida (N2O), perfluorokarbon (PFC), hidrofluorokarbon (HFC) dan sulfurheksfluorida (SF6). Gas rumah kaca yang menyelimuti atmosfer bumi akan menyerap radiasi gelombang panjang yang memanaskan bumi (Hairiah 2008)
Untuk mengantisipasi dampak perubahan iklim terhadap penurunan produksi tanaman diperlukan tanaman atau varietas unggul yang mampu beradaptasi pada perubahan lingkungan dan kondisi bercekaman. Varietas unggul tersebut memiliki ideotype (tipe tanaman ideal) pada lingkungan bercekaman tertentu seperti abiotik dan biotik sehingga penurunan produksi tanaman dapat dikendalikan (Soepandie 2006; Irianto 2009).
Usaha Peningkatan Keragaman Genetik Tanaman Kedelai
Perbaikan produktivitas kedelai melalui pemuliaan konvensional menghadapi beberapa masalah seperti terbatasnya koleksi plasma nutfah sehingga keragaman untuk karakter tertentu rendah. Tanaman kedelai termasuk tanaman menyerbuk sendiri yang membentuk galur murni jika dari generasi ke generasi mengalami penyerbukan sendiri (Jusuf 2004). Keragaman genetik tanaman dapat ditingkatkan melalui cara introduksi, hibridisasi (persilangan), poliploidi, mutasi dan transformasi genetik tanaman. Apabila pemuliaan tanaman hanya dari bahan yang telah ada pada suatu saat secara genetik akan terbatas sehingga kemajuannya pun menjadi lambat (Jusuf 2004; Arsyad 2007 ).
Induksi mutasi salah satu metode untuk meningkatkan keragaman pada berbagai karakter. Menurut Van Harten (1998) mutasi adalah suatu perubahan yang bersifat diwariskan yang terjadi pada DNA. Mutasi dapat terjadi secara alami atau diinduksi dengan perlakuan bahanmutagen tertentu terhadap organ reproduksi tanaman seperti biji, stek batang, serbuk sari, akar rhizome, kultur jaringan dan sebagainya (BATAN 2006).
Induksi mutasi dapat dilakukan dengan menggunakan mutagen fisik dan mutagen kimia. Mutagen fisik dapat menggunakan iradiasi tanpa ionisasi (sinar UV) atau iradiasi dengan ionisasi (sinar X, gamma, alpha, beta, dan neutron). Mutagen fisik menyebabkan mutasi atau terhapusnya DNA dalam jumlah yang besar dan perubahan pada struktur kromosom. Mutagen kimia hanya mempengaruhi satu pasang nukleotida (mutasi titik). Mutagen kima yang sering digunakan adalah ethylmethane sulphonate (EMS), methyl-methane sulphonate
Di antara kedua jenis mutagen, mutasi fisik paling banyak digunakan, terutama sinar gamma. Sinar gamma merupakan iradiasi terionisasi yang bersifat elektromagnetik. Sinar gamma memiliki panjang gelombang pendek, energi yang tinggi, tidak bersifat elektrik dan tidak mempunyai massa dibandingkan partikel iradiasi lainnya (EPA 2012). Agar sinar gamma efektif untuk menginduksi mutasi sehingga menghasilkan keragaman maka perlu diketahui dosis yang sesuai. Dosis yang sesuai untuk induksi keragaman dapat diketahui dengan mempelajari radiosensitivitas dan mengukur nilai LD50 (Amano et al. 2001).
Radiosensitivitas adalah tingkat kepekaan tanaman terhadap iradiasi. Secara visual tingkat kepekaan ini dapat diamat berdasarkan daya berkecambah atau pertumbuhan tanaman setelah perlakuan iradiasi (Sanada dan Amano 1998). Lethal Doses (LD50) adalah dosis yang menyebabkan kematian 50 % dari populasi yang diradiasi. Berbagai hasil penelitian menunjukkan bahwa umumnya mutasi yang diinginkan dapat diperoleh dengan perlakuan dosis sekitar LD50 atau lebih tepatnya pada dosis sedikit di bawah LD50 (Van Harten 1998; Amano et al. 2001; Tah 2006; Manjaya dan Nandanwar 2007; Borzouei et al. 2010; Goyal dan Khan 2010).
LD50 berbeda pada setiap jenis tanaman. Nilai LD50 kultivar kedelai Pusa-16 dan PK-1042 terdapat pada nilai 377 Gy dan 467,4 Gy (Khan dan Tyagi 2005). Hal ini menunjukkan bahwa antar kultivar atau intraspesies terdapat radiosensitivitas berbeda.
Perlakuan iradiasi sinar gamma dapat diberikan dengan dua cara, yaitu
fractionated irradiation technique dan acute irradiation. Weather dan Sader (1990) menyatakan bahwa fractionated irradiation technique (iradiasi terbagi) pada eksplan pucuk gerbera membuat persentase tumbuh tunas yang lebih tinggi dibandingkan teknik acute irradiation (iradiasi tunggal). Fractionated irradiation
atau iradiasi terbagi adalah teknik permberian dosis yang diberikan dua kali, masing-masing setengah dosis atau split dose.
Teknik pemberian iradiasi lainnya yaitu dengan iradiasi berulang baik secara intermittent irradiation (jarak antar iradiasi tidak terlalu lama) atau
iradiasi tunggalnya. Pada iradiasi berulang, iradiasi diberikan sekali ataubeberapa kali lagi dengan dosis yang lebih rendah (Basiran dan Ariffin 2002).
Generasi keturunan berikutnya dari biji atau tanaman yang diperlakukan dengan mutagen disimbolkan dengan M1,M2, M3,dan seterusnya untuk membedakan dengan generasi hibridisasi yang disimbolkan dengan F1, F2, F3 dan seterusnya. Hanya hasil mutasi dominan yang terekspresikan dalam generasi M1.Mutasi akan menghasilkan kimera, yaitu dimana keturunan-keturunan sel-sel mutan dan sel-sel normal akan menghasilkan jaringan genotipe yang berbeda. Tanaman M1biasanya bersifat kimerik, melalui pertumbuhan tanaman M1,kesempatan kompetisi antara mutan dan non mutan terjadi (Nasir 2002).
Pemuliaan mutasi kedelai dimulai pada tahun 1977. Materi percobaan (induknya) dipilih kedelai varietas Orba dengan mengiradiasi varietas Orba, menggunakan sinar gamma dengan dosis 0,40 kGy. Pada percobaan tersebut didapat satu galur mutan yang hasilnya tinggi dan dilepas pada tahun 1987 sebagai varietas kedelai baru, yang diberi nama Muria (BATAN 2003).
BATAN telah menghasilkan beberapa mutan tanaman pangan yang dilepas sebagai varietas unggul oleh Departemen Pertanian, dimana sampai saat ini telah diakui empat varietas mutan tanaman kedelai, yaitu Muria, Tengger, Meratus dan terakhir yang dilepas pada tahun 2004 yaitu varietas Rajabasa (BATAN 2006).
Hasil penelitian iradiasi sinar gamma pada tanaman kedelai diantaranya telah dilakukan oleh Manjaya dan Nandanwar (2007). Dosis penyinaran sinar gamma 250 Gy berhasil menginduksi terjadinya mutasi dan menyebabkan terjadinya keragaman genetik pada kedelai cv JS 80-21. Dosis iradiasi ini berbeda untuk tiap kultivar dan spesies kedelai yang ada. Persentase perkecambahan juga dipengaruhi oleh keadaan kondisi benih baik dari segi vigor dan besarnya ukuran benih. Perbaikan keragaan tanaman dan produksi benih yang tinggi tercapai dengan dirilisnya varietas baru yang berasal dari cv JS 80-21, yaitu TAMS 98-21.
Cekaman Kekeringan pada Tanaman Kedelai
produksi tanaman. Cekaman lingkungan pada tanaman terbagi atas cekaman biotik dan abiotik. Cekaman biotik disebabkan oleh organisme (patogen, serangga dan binatang. Cekaman abiotik disebabkan oleh kelebihan dan kekurangan pada lingkungan baik secara fisik maupun kima (genangan air, kekeringan, suhu, cahaya, salinitas tanah dan cekaman unsur mineral dalam tanah) (Cassells dan Doyle 2003).
Cekaman kekeringan adalah suatu kondisi yang ditunjukkan oleh defisit air pada lingkungan tumbuh tanaman. Cekaman kekeringan terjadi ketika akar tidak mampu menyerap air dalam jumlah yang cukup. Tanaman mengalami kekeringan bila laju penyerapan air tanah oleh perakaran tidak dapat mengimbangi laju evapotranspirasi (Levitt 1980).
Tingkat cekaman kekeringan dapat berbeda-beda tergantung pada jenis tanah serta kemampuan tanah menahan air (kapasitas air tanah) (Mason et al. 1980). Gejala kekeringan ditandai oleh adanya gangguan pertumbuhan dan perkembangan tanaman diantaranya hambatan dalam penyerapan hara, pengurangan laju transpirasi dan nilai relatif luas daun. Gangguan pertumbuhan dan perkembangan juga dipengaruhi oleh penurunan laju fotosintesis, fiksasi CO2, dan daya hantar stomata seiring dengan peningkatan intensitas cekaman yang mempengaruhi produksi assimilat untuk pertumbuhan dan produksi tanaman (Kirnak et al. 2001; Purwanto 2003).
Secara umum, tanaman peka terhadap kondisi defisit air pada lingkungan tumbuhnya, terutama pada tahap pembungaan dan pengisian biji pada tanaman (Mitra 2001). De Souza et al. (1997) menambahkan cekaman kekeringan selama pengisian biji mengurangi hasil kedelai karena cekaman kekeringan mempercepat pengguguran daun dan memperpendek periode pengisian biji.
Kacang kedelai (Glycine max), seperti kebanyakan spesies tanaman, sangat peka pada kondisi air yang suboptimal selama pertumbuhan vegetatif dan reproduktif. Kacang kedelai adalah suatu spesies dehydration-sensitive yang menuntut jumlah air optimum untuk perkecambahan benih, pertumbuhan bibit dan perkembangan tanaman (Chen et al. 2006).
perkecambahan. Kedelai membutuhkan air sebanyak 300 mm hingga 450 mm atau 2,5-3,3 mm/hari selama pertumbuhan (Fagi dan Tangkuman 1985).
Kondisi air tanah untuk kebutuhan kedelai yang diukur berdasarkan kapasitas lapang berbeda-beda untuk setiap periode pertumbuhan. Kadar air lapang yang tinggi, yaitu berkisar antara 80-100 % diperlukan pada saat benih ditanam hingga berkecambah dan tanaman berdaun tunggal muncul di permukaan tanah (1-12 hst). Untuk pertumbuhan selanjutnya, tanaman kedelai akan tumbuh baik pada kapasitas lapang sekitar 75-85 % dan mulai menurun pada stadia biji mencapai ukuran maksimum (R6) dan kelembaban tanah yang mininum pada saat polong mulai matang hingga polong matang penuh (R7-R8) (Chen et al. 2006; Sumarno dan Manshuri 2007).
Mekanisme Adaptasi Terhadap Cekaman Kekeringan
Adaptasi adalah tanggap tanaman terhadap keadaan lingkungan yang kurang menguntungkan bagi pertumbuhan dan perkembangan tanaman (Levitt 1980). Mekanisme adapatasi tanaman terhadap cekaman kekeringan terbagi atas tiga kategori yaitu :
a. Melepaskan diri dari cekaman kekeringan (drought escape), yaitu kemampuan tanaman untuk dapat melengkapi siklus hidupnya sebelum mengalami defisit air yang parah. Mekanisme ini melibatkan perkembangan fenologi yang cepat (umur berbunga dan umur panen yang cepat
b. Penghindaran dari cekaman kekeringan (drought avoidance), yaitu kemampuan tanaman untuk menjaga potensial air jaringan yang tinggi. Mekanisme ini dengan meningkatkan perakaran yang lebih dalam, mengurangi kehillangan air melalui pengurangan epidermal (stomata dan lenticular), mengurangi penyerapan radiasi matahari dengan penyempitan dan penggulungan daun
c. Toleransi terhadap cekaman kekeringan (drought tolerance), yaitu kemampuan tanaman untuk bertahan pada potensial air jaringan yang rendah. Mekanisme ini menjaga tekanan turgor tanaman melalui osmotic adjustment
Pada tanaman kedelai, cekaman kekeringan akan mengurangi pengambilan air oleh akar. Jika akar tanaman tidak mampu menyerap air untuk mengimbangi kehilangan air oleh transpirasi menyebabkan tanaman menjadi layu. Tanaman akan mengurangi kehilangan air dengan penutupan stomata. Penutupan stomata membantu tanaman untuk menghindari kekeringan yang cepat. Bagaimanapun, pori-pori stomata yang tertutup juga menghambat pertukaran karbondioksida dan oksigen antara jaringan internal tanaman dan udara di luarnya. Hal ini juga menghentikan aliran air melalui tanaman yang mengurangi juga penyerapan hara. Semua faktor yang di uraikan di atas adalah penyebab tanaman kedelai mengurangi metabolismenya dalam rangka untuk mempertahankan hidupnya selama masa kekeringan (Borges 2004).
Genotipe toleran terhadap cekaman kekeringan mempunyai transpirasi lebih rendah, fotosintesis lebih tinggj, menggunakan air lebih efisien dan mampu memberikan hasil polong lebih tinggi dibanding genotipe peka terhadap cekaman kekeringan. Toleransi genotipe Singa pada kacang tanah lebih baik, antara lain disebabkan mempunyai transpirasi lebih rendah, fotosintesis lebih tinggi dan lebih efisien dalam penggunaan air, yang disebabkan adanya perbedaan transpirasi antara genotipe disebabkan oleh perbedaan penyebaran akar, penutupan kanopi dan perkembangan daun (Harsono et al. 2003).
Kemampuan tanaman mempertahankan tekanan turgor pada kondisi kekeringan dengan cara menurunkan potensial osmotiknya sebagai mekanisme toleransi terhadap cekaman kekeringan. Proses fisiologi dan biokimia dalam tumbuhan sangat dipengaruhi oleh perubahan tekanan turgor. Faktor yang dapat membantu mempertahankan turgor sel ialah : (1) penurunan potensial osmotik sel dan (2) kemampuan mengakumulasi zat-zat terlarut seperti senyawa-senyawa gula, asam-asam amino dan prolina (Salisbury dan Ross 1992).
berlebihan dapat menghasilkan peningkatan toleransi terhadap cekaman kekeringan pada tanaman (Kishor, et al. 1995).
Beberapa peneliti (Sopandie, et al. 1996; Hamim, et al. 1996; Hapsoh, et al.
2006) menyatakan bahwa pada kedelai mekanisme fisiologi terpenting dari adaptasi terhadap kekeringan dengan mempertahankan turgor melalui penurunan potensial osmotik dan akumulasi senyawa terlarut yaitu prolina serta adanya peningkatan kandungan ABA dan IAA.
Hormon pertumbuhan yang terakumulasi selama tanaman berada pada kondisi cekaman kekeringan diantaranya adalah ABA. ABA yang diproduksi dalam akar tanaman yang tercekam kekeringan berperan sebagai sinyal kimia pada tajuk sehingga mendorong penutupan stomata sebelum perubahan status air dalam daun terjadi, sehingga tanaman dapat mengoptimalkan penggunaan air pada kondisi tercekam (Zeevaart dan Creelman 1988).
Lopez-Carbonell et al. (1996) menyimpulkan bahwa peningkatan ABA pada tanaman lavender dan rosemary yang telah beradaptasi pada kondisi cekaman kekeringan bukan merupakan mekanisme umum. Hal ini disebabkan karena cekaman kekeringan pada lavender meningkatkan ABA pada awal cekaman hari kedua dan ketiga tetapi setelah hari ketiga ABA dan zeatin ribosida (sitokinin) menurun tajam tetapi IAA (auksin) cenderung meningkat. Sebaliknya pada rosemary, auksin dan sitokinin cenderung berkurang. Disimpulkan bahwa cekaman kekeringan meningkatkan ABA guna mempertahankan turgor, meningkatkan aktivitas IAA oksidase, mengurangi transpor auksin dan sitokinin dari akar. Menurut Mansfield dan McAinsh (1995), IAA menstimulasi pembukaan stomata dan pergerakan air di akar dan bertindak bersama dengan ABA dalam menghadapi berbagai cekaman yakni mempengaruhi turgor seperti salinitas dan kekeringan.
Pemuliaan bagi Lingkungan Bercekaman
faktor-faktor yang menjadi permasalahan bisa dimodifikasi atau dikendalikan. Lahan yang dianggap sub optimal antara lain adalah lahan kering masam, lahan tadah hujan dan lahan rawa (Rachman et al. 2007).
Pemuliaan tanaman pada lingkungan bercekaman memerlukan tanaman atau varietas unggul yang mampu beradaptasi pada kondisi cekaman pada daerah penanamannya. Pengembangan kedelai pada lingkungan yang mengalami cekaman dapat dicapai melalui perbaikan potensi hasil, tingkat adaptasi tanaman terhadap cekaman abiotik dan biotik dan penggunaan teknik budidaya yang berbasis pengetahuan fisiologi atau ekofisiologi tanaman. Ketiga faktor tersebut saling berkaitan untuk mendapatkan varietas unggul kedelai produksi tinggi yang spesifik agroekologi melalui pendekatan ideotype breeding. Ideotype breeding
adalah perbaikan tanaman dengan mengembangkan kombinasi spesifik dari karakter yang mendukung fotosintesis, pertumbuhan, dan produksi dengan pemahaman tentang morfologi dan fisiologi tanaman (Soepandie 2006).
Varietas unggul yang memiliki ideotype breeding pada lingkungan bercekaman tertentu seperti abiotik dan biotik, memerlukan keragaman genetik pada populasi dasarnya. Keragaman dalam suatu populasi terdiri dari keragaman fenotipe dan keragaman genotipe. Keragaman fenotipe adalah keragaman yang dapat diukur langsung dari karakter yang diamati, sedangkan keragaman genotipe adalah adalah keragaman yang tidak dapat diukur langsung, pengukuran dapat diduga melalui analisis ragam (Roy 2000).
Heritabilitas dari hasil pada kondisi cekaman kekeringan tergantung pada dua faktor yaitu (a) adanya gen-gen ketahanan kekeringan pada populasi, dimana seleksi efektif dilakukan di kondisi cekaman kekeringan, dan (b) tingkat kehomogenan kondisi kekeringan dimana seleksi dilakukan. Seleksi pada lingkungan homogen pada kondisi kekeringan serta tidak disertai adanya masalah seperti gulma, penyakit, nematoda, jenis tanah yang digunakan, menghasilkan nilai duga heritabilitas yang lebih baik dibandingkan dengan nilai duga heritabilitas pada lingkungan yang tidak homogen (Blum 2005).
Poehlman dan Sleper (1996) menyatakan bahwa keragaman genetik pada suatu populasi menentukan efektivitas seleksi untuk mendapatkan genotipe unggul dan seberapa besar sifat unggul yang diinginkan tersebut dapat diturunkan pada generasi selanjutnya. Seleksi terhadap karakter yang memiliki keragaman genetik yang luas dan kriteria heritabilitas yang sedang-tinggi akan memberikan kemajuan seleksi pada generasi selanjutnya.
Seleksi pada lingkungan seleksi dapat dilakukan dengan dua cara yaitu (a) seleksi langsung (direct breeding) dan seleksi tidak langsung (indirect breeding). Seleksi langsung pada lingkungan bercekaman menghasilkan genotipe-genotipe terpilih yang mampu beradaptasi pada lingkungan bercekaman tersebut. Keragaman genetik pada lingkungan bercekaman cenderung rendah, nilai heritabilitas yang rendah dan pengaruh interaksi antara genotipe dengan lingkungan besar sehingga mempengaruhi penampilan fenotipe tanaman dan kemajuan genetik saat seleksi (Beebe et al. 2008; Witcombe et al. 2008).
Seleksi tidak langsung dilakukan pada suatu lingkungan tidak bercekaman (optimum) lebih efisien dibandingkan seleksi langsung jika hasil relatif yang diperoleh dari dua lingkungan tidak berbeda nyata dan nilai heritabilitas karakter pada lingkungan optimum lebih tinggi dibandingkan pada lingkungan bercekaman (Witcombe et al. 2008). Seleksi tidak langsung hanya efektif jika korelasi nyata diantara karakter-karakter yang diamati dengan hasil (Wricke dan Weber 1986).
RADIOSENSITIVITAS DAN PENGARUH IRADIASI DOSIS RENDAH SINAR GAMMA PADA VARIETAS ARGOMULYO
ABSTRAK
Tujuan uji radiosensitivitas adalah untuk memperoleh informasi mengenai nilai LD50 dari kedelai varietas Argomulyo. Untuk menentukan radiosensitivitas, benih Argomulyo diiradiasi sinar gamma dengan dosis 200 Gy, 400 Gy, 600 Gy, 800 Gy, dan 1000 Gy. Pengamatan dilakukan pada populasi M1 untuk daya tumbuh kecambah untuk menentukan radiosensitivitas. Hasil penelitian menunjukkan bahwa perkecambahan varietas Argomulyo memberikan respon yang berbeda terhadap masing-masing dosis iradiasi yang diberikan. Nilai Lethal Dosis 50 (LD50) terdapat pada dosis 457,13 Gy.
Kata kunci : iradiasi sinar gamma, radiosensitivitas, LD50
ABSTRACT
The objective of this research was to find out information about the value of LD50 from soybean of Argomulyo variety. To determine the radiosensitivy, the seeds of Argomulyo variety were treated with gamma ray irradiation with doses of 200 Gy, 400 Gy, 600 Gy, 800 Gy, and 1000 Gy. The M1 population were grown and observed for mortality to determine the radiosensitivity. The results of the observation showed that the germination of Argomulyo variety gave different responses to the gamma rays for each dose of irradiation. The value of LD50 for Argomulyo was found at a dose of 457,178 Gy.
Key word : gamma ray irradiation, radiosensitivity, LD50
PENDAHULUAN
Kedelai bukan merupakan tanaman asli Indonesia, tapi merupakan tanaman
introduksi yang berasal dari Cina. Kedelai merupakan tanaman menyerbuk
sendiri yang bersifat kleistogami dan keragaman genetik kedelai di Indonesia
masih rendah (Adie dan Krisnawati 2007). Untuk meningkatkan keragaman
genetik tanaman kedelai dapat dilakukan melalui pemuliaan secara mutasi untuk memperbaiki karakter-karakter tanaman yang diinginkan. Pemuliaan mutasi merupakan suatu cara untuk memperoleh keragaman genetik dari karakter-karakter kuantitatif maupun kualitatif pada tanaman (Manjaya dan Nandawar 2007; Kavithamani et al. 2010).
Varietas Argomulyo memiliki potensi hasil tinggi yaitu dua ton per hektar, ukuran biji sama bahkan lebih besar dari kedelai impor dan kadar proteinnya lebih tinggi dari kedelai impor (Hidajat
Varietas Argomulyo adalah salah satu varietas unggul nasional berbiji besar, berwarna kuning dan direkomendasikan untuk penanaman pada lahan sawah (Ginting et al. 2009). Kompetisi penanaman dengan tanaman lain seperti padi dan jagung menyebabkan penurunan luas areal penanaman kedelai. Oleh karena itu pemanfaatan lahan kering untuk perluasan tanam kedelai dapat menjadi alternatif untuk meningkatkan produksi kedelai. Pemanfaatan lahan kering harus didukung oleh varietas yang mampu beradaptasi dan berpotensi hasil baik (Arsyad et al. 2007).
Pengembangan varietas toleran kering dimulai dengan pembentukan populasi dasar yang memiliki keragaman genetik tinggi untuk karakter yang diperbaiki. Iradiasi sinar gamma hanya mengubah satu atau beberapa karakter pada tanaman. Induksi keragaman genetik melalui iradiasi sinar gamma dapat dilakukan dengan menggunakan dosis rendah yaitu dosis di bawah LD50 (Van Harten 1998; Sakin 2002).
Proses mutasi alami terjadi secara sangat lambat dan membutuhkan waktu yang lama, maka percepatan, frekuensi dan spektrum mutasi tanaman dapat diinduksi dengan perlakuan bahan mutagen tertentu. Induksi mutasi dapat dilakukan pada tanaman dan hewan dengan perlakuan bahan mutagen secara fisik dan kimiawi. Mutagen secara fisik dapat dilakukan menggunakan iradiasi ionisasi (sinar X, gamma, alpha, beta dan neutron) dan iradiasi tanpa ionisasi (sinar UV) (Parry et al. 2009). Sinar gamma merupakan iradiasi yang bersifat ionisasi dan merupakan iradiasi elektromagnetik dengan energi yang tinggi. Tingkat energi sinar gamma berkisar dari 10 kilo elektron volts (keV) sampai beberapa ratus kilo elektron volts. Sinar gamma mempunyai daya tembus kuat daripada tipe iradiasi lainnya seperti sinar alpha dan beta (Kovacs dan Keresztes 2002).
Sinar gamma berinteraksi dengan atom atau molekul air yang menghasilkan radikal bebas dalam sel. Radikal bebas ini dapat merusak atau mempengaruhi komponen-komponen penting dalam tanaman dan dilaporkan telah mempengaruhi perubahan morfologi, anatomi, biokimia dan fisiologi dari tanaman tergantung pada tingkat iradiasi yang diberikan (Thilagavathi dan Mullainathan 2011).
tanaman. Dosis iradiasi ini berbeda untuk tiap kultivar dan varietas kedelai yang ada (Srisombun et al. 2009). Berdasarkan hal itu perlu diketahui dosis iradiasi yang tepat untuk varietas Argomulyo yang dapat menginduksi keragaman tertinggi. Tujuan penelitian ini adalah untuk memperoleh taraf radiosensitivitas tanaman kedelai varietas Argomulyo melalui iradiasi sinar gamma.
METODE PENELITIAN
Penelitian ini dilaksanakan di Institut Pertanian Bogor dari bulan Februari sampai Maret 2009. Perlakuan iradiasi dilaksanakan pada Pusat Sumberdaya Biologi dan Bioteknologi, IPB, Bogor dan penelitian lapangan dilaksanakan pada University farm IPB, Bogor. Penelitian ini menggunakan benih varietas Argomulyo. Benih diiradiasi dengan sinar gamma yang bersumber dari 137 Cs menggunakan IBL 437C type H Irradiator (CIS Bio International, France) dengan laju dosis 2,23 Gy/menit. Dosis iradiasi yang digunakan adalah 200 Gy, 400 Gy, 600 Gy, 800 Gy, dan 1000 Gy
Benih ditanam dalam bak perkecambahan dan dievaluasi persentase daya tumbuh pada dua minggu setelah penanaman. Peubah yang diamati (1) persentase daya tumbuh (2) tinggi bibit, yang dihitung dari pangkal akar sampai ujung bibit. Data persentase daya tumbuh digunakan untuk menentukan nilai Lethal Dose 50 (LD50) yang dianalisis menggunakan program Curve-fit Analysis. Program ini merupakan suatu program untuk menentukan model persamaan yang terbaik untuk persentase daya tumbuh dari suatu populasi (Finney 2000).
HASIL PENELITIAN
Tabel 1. Persentase daya tumbuh dan tinggi kecambah dua minggu setelah tanam.
No. Dosis (Gy) Persen Daya Tumbuh (%) Tinggi Kecambah (cm)
1 0 (Kontrol) 100,0 14,7±0,6
2 200 81,3 10,3±0,3
3 400 87,5 7,8±0,5
4 600 81,3 4,1±0,5
5 800 75,0 2,3±0,3
6 1000 87,5 1,6±0,4
Pengaruh peningkatan dosis sinar gamma mempengaruhi pertumbuhan kecambah kedelai. Semakin tinggi dosis iradiasi yang diberikan semakin rendah pertumbuhan tinggi kecambah (Gambar 2). Pada dosis iradiasi 1000 Gy perumbuhan tetap terjadi, tetapi pertumbuhan kecambah mengalami stagnasi serta abnormalitas yang ditunjukkan oleh batang menebal dan daun kotiledon menebal serta tinggi tanaman tidak bertambah sampai akhir pengamatan (dua minggu setelah tanam) (Tabel 1). Secara visual tingkat kepekaan ini dapat diamati pada tanggap yang diberikan tanaman, baik pada morfologi tanaman maupun pada kemampuan daya tumbuh tanaman.
Gambar 2. Pertumbuhan kecambah tanaman kedelai varietas Argomulyo pada umur 2 MST yang diiradiasi sinar gamma pada dosis 0, 200, 400, 600, 800 dan 1000 Gy.
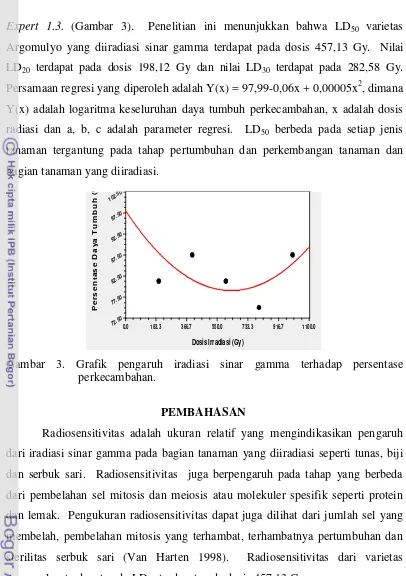
Expert 1.3. (Gambar 3). Penelitian ini menunjukkan bahwa LD50 varietas Argomulyo yang diiradiasi sinar gamma terdapat pada dosis 457,13 Gy. Nilai LD20 terdapat pada dosis 198,12 Gy dan nilai LD30 terdapat pada 282,58 Gy. Persamaan regresi yang diperoleh adalah Y(x) = 97,99-0,06x + 0,00005x2, dimana Y(x) adalah logaritma keseluruhan daya tumbuh perkecambahan, x adalah dosis radiasi dan a, b, c adalah parameter regresi. LD50 berbeda pada setiap jenis tanaman tergantung pada tahap pertumbuhan dan perkembangan tanaman dan bagian tanaman yang diiradiasi.
Gambar 3. Grafik pengaruh iradiasi sinar gamma terhadap persentase perkecambahan.
PEMBAHASAN
Radiosensitivitas adalah ukuran relatif yang mengindikasikan pengaruh dari iradiasi sinar gamma pada bagian tanaman yang diiradiasi seperti tunas, biji dan serbuk sari. Radiosensitivitas juga berpengaruh pada tahap yang berbeda dari pembelahan sel mitosis dan meiosis atau molekuler spesifik seperti protein dan lemak. Pengukuran radiosensitivitas dapat juga dilihat dari jumlah sel yang membelah, pembelahan mitosis yang terhambat, terhambatnya pertumbuhan dan sterilitas serbuk sari (Van Harten 1998). Radiosensitivitas dari varietas Argomulyo terdapat pada LD50 terdapat pada dosis 457,13 Gy.
LD50 berbeda pada setiap jenis tanaman tergantung pada tahap pertumbuhan dan perkembangan tanaman dan bagian tanaman yang diiradiasi. LD50 pada persentase perkecambahan dan daya tumbuh dari biji tomat 790 dan 640 Gy sedangkan pada biji okra 770 dan 580 Gy. Secara umum, dosis sinar gamma yang tinggi terutama pada 800 Gy mempunyai efek negatif pada
Dosis Irradiasi (Gy)
P e r s e n t a s e D a y a T u m b u h ( %
0.0 183.3 366.7 550.0 733.3 916.7 1100.0
[image:49.595.107.513.66.642.2]penampilan morfologi dan karakter dari perkecambahan tomat dan okra (Norfadzrin et al. 2007). Nilai LD50 kultivar kedelai Pusa-16 dan PK-1042 terdapat pada nilai 377 Gy dan 467,4 Gy (Khan dan Tyagi 2005).
Teknologi iradiasi digunakan oleh pemulia untuk menginduksi mutasi. Iradiasi dilakukan pada kisaran dosis kematian 50% dengan pertimbangan bahwa kerusakan fisiologis berimbang dengan perubahan genetik yang diperoleh (Sudrajat dan Zanzibar 2009). Selain untuk menentukan tingkat sensitivitas terhadap mutagen fisik maupun kimia, kisaran dosis LD50 juga berguna untuk memperkirakan dosis atau konsentrasi yang sesuai untuk menginduksi mutasi (Abdullah et al. 2009). Kisaran dosis sinar gamma yang digunakan untuk menginduksi keragaman terdapat dibawah dosis LD50, berkisar antara LD20 (198,12 Gy) dan LD30 (282,58 Gy) (Van Harten 1998).
Generasi M1 merupakan populasi yang mengalami pengaruh fisiologis akibat iradiasi. Sinar gamma menghasilkan elektron bebas yang bersifat radikal sehingga mengakibatkan kerusakan pada sel (Van Harten 1998). Hasil penelitian menunjukkan bahwa semakin tinggi dosis iradiasi yang diberikan, mempengaruhi pertumbuhan kecambah benih Argomulyo, dimana pertumbuhan kecambah terhambat dan tidak berkembang. Tah (2006) menyatakan bahwa tingkat kematian meningkat secara linier seiring dengan meningkatnya dosis pada tanaman kacang hijau [Vigna radiata (L.) Wilczek], demikian pula hasil penelitian Kumar dan Ratnam (2010) pada tanaman bunga matahari. Penelitian yang lain menyatakan bahwa pengaruh perlakuan berbagai dosis iradiasi terhadap perkecambahan benih dan kekuatan benih berhubungan secara linier antara dosis mutagen dan penurunan tingkat perkecambahan benih (Sarkar et al. 1996; Singh
et al. 1997).
elektron bebas dapat menginduksi terbentuknya radikal yang dapat bereaksi dengan makromolekul seperti protein dan lemak sehingga akan mempengaruhi proses metabolisme dan fisiologi dari sel atau tanaman (Van Harten 1998). Selain itu spesies tanaman, tingkat ploidy, tahap perkembangan tanaman dan keadaan fisiologis (adanya cekaman biotik/abiotik) akan menyebabkan respon yang berbeda terhadap iradiasi sinar gamma (Bhatia et al. 2001).
KESIMPULAN
KERAGAMAN KARAKTER AGRONOMI DAN SELEKSI GENOTIPE PUTATIF MUTAN HASIL IRADIASI SINAR GAMMA DOSIS RENDAH
ABSTRAK
Induksi mutasi dengan iradiasi sinar gamma merupakan salah satu cara dalam meningkatkan keragaman genetik tanaman. Iradiasi sinar gamma dosis rendah (micro mutation) diberikan pada benih kedelai varietas Argomulyo dengan dosis di bawah LD50 yaitu 0 Gy, 50 Gy, 100 Gy, 150 Gy dan 200 Gy. Tujuan penelitian ini untuk memperoleh informasi tentang respon pertumbuhan dan perkembangan tanaman setelah pemberian iradiasi mikro sinar gamma pada generasi M1, keragaman genetik, perubahan karakter morfologi dan agronomi akibat induksi mutasi pada generasi M2 dan seleksi genotipe putatif mutan terpilih dengan karakter agronomi yang baik dan berdaya hasil tinggi pada generasi M3 dan M4. Hasil penelitian menunjukkan bahwa iradiasi sinar gamma menghasilkan empat populasi iradiasi yang menunjukkan pertumbuhan dan perkembangan yang berbeda pada setiap karakter agronomi yang diamati dibandingkan dengan populasi kontrol (Argomulyo) pada generasi M1. Pada generasi M2, keragaman genetik setiap karakter berbeda-beda pada masing-masing populasi hasil iradiasi. Seleksi berdasarkan jumlah polong bernas dan bobot biji per tanaman, menyebabkan perbaikan nilai tengah genotipe terpilih pada masing-masing populasi hasil iradiasi dari generasi M3. Galur putatif mutan M4 yang berasal dari seleksi M3 memiliki nilai tengah karakter tinggi tanaman, jumlah buku produktif dan jumlah polong bernas yang lebih baik dibanding varietas asal Argomulyo.
Kata kunci : keragaman genetik, karakter agronomi, iradiasi sinar gamma, mutasi mikro, seleksi pedigri
ABSTRACT
Mutation induced by gamma ray irradiation is one way to increase the genetic diversity of plants. Low doses of gamma ray irradiation is applied to soybean seed varieties Argomulyo with a dose of slightly under LD50, that is, 0 Gy, 50 Gy, 100 Gy, 150 Gy and 200 Gy. The aim of this study was to obtain information about the responses of plant growth and development after being
irradiated by gamma ray in M1 generation, genetic variability, changes in
morphological and agronomic characters as a result of mutation induction in M2 generation and genotype selection of putative mutant with good agronomic characters and high yielding in M3 and M4 generations. The results of the study showed that gamma ray irradiation generated four populations with different growth and development in each agronomic character observed compared with the control population (Argomulyo) in the M1 generation. In the M2 generation, genetic variability generated were different for every character in each population. The selection based on the number of productive pods and seed weight per plant resulted in an improvement on the mean of selected genotypes compared to the
mean of basic population in M3 generation. The M4 putative mutant lines derived
Keywords: genetic variability, agronomy traits, low doses of gamma rays irradiation, pedigree selection
PENDAHULUAN
Keragaman genetik merupakan dasar dalam upaya perbaikan karakter tanaman melalui pemuliaan tanaman. Beberapa spesies tanaman budidaya mempunyai keragaman genetik yang tinggi, sementara sebagian spesies tanaman yang lain mempunyai keragaman genetik yang rendah. Tanaman menyerbuk sendiri, seperti tanaman kedelai akan membentuk galur-galur yang mantap atau tidak bersegregasi. Populasi tersusun dari galur-galur, dengan keragaman genetik intragalur sangat kecil atau hampir nol, dan keragaman antar galur sangat nyata. Keragaman genetik baru akan muncul di alam sebagai akibat mutasi atau terjadinya persilangan antar galur, walau dengan derajat yang kecil sehingga keragaman genetik kedelai rendah (Jusuf 2004).
Mutasi dengan iradiasi dosis tinggi, biasanya menyebabkan ketidakstabilan genetik (Van Harten 1998). Adapun mutasi mikro mengubah karakter kuantitatif yang diturunkan dari benih yang diiradiasi dan lebih bermanfaat bagi pemulia, karena mutasi mikro sedikit merusak walaupun mutasi ini sulit dideteksi. Mutasi mikro dapat meningkatkan keragaman pada hasil, kandungan protein, tinggi tanaman, pembungaan, produksi polong, berat biji dan hasil-hasil lain yang berhubungan dengan karakter kuantitatif yang diturunkan (Sakin 2002; Tah 2006). Dosis iradiasi ini berbeda untuk tiap kultivar dan varietas kedelai yang ada. Secara umum, dosis iradiasi sinar gamma untuk tanaman kacang-kacangan berkisar dari 100 Gy sampai 200 Gy (Bhatia et al. 2001)
Keragaman genetik pada materi pemuliaan merupakan salah satu syarat dasar untuk melakukan seleksi. Keefektifan dari seleksi tergantung pada luasnya keragaman genetik untuk karakter yang akan diseleksi dalam populasi. Parameter genetik seperti koefisien keragaman genetik dan heritabilitas merupakan cara untuk menentukan keragaman genetik dalam populasi (Hiremath et al. 2011).
Setelah diperoleh populasi dengan keragaman yang tinggi dari induksi mutasi, seleksi dapat dilakukan dengan metode seleksi pedigri. Metode pedigri dapat mengident