ISOLASI DAN IDENTIFIKASI BAKTERI KITINOLITIK
PENGHAMBAT PERTUMBUHAN CENDAWAN PATOGEN
ASAL KOKON Cricula trifenestrata
PRIMA NOVITASARI
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ISOLASI DAN IDENTIFIKASI BAKTERI KITINOLITIK
PENGHAMBAT PERTUMBUHAN CENDAWAN PATOGEN
ASAL KOKON Cricula trifenestrata
PRIMA NOVITASARI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Biologi
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul : Isolasi dan Identifikasi Bakteri Kitinolitik Penghambat Pertumbuhan
Cendawan Patogen Asal Kokon
Cricula trifenestrata
Nama : Prima Novitasari
NIM : G34080006
Menyetujui
Pembimbing I,
Pembimbing II,
(Dr. Nisa Rachmania Mubarik, M.Si.)
(Dr. Sri Listiyowati, M.Si.)
NIP: 196711271993022001
NIP: 196407141990022001
Mengetahui
Ketua Departemen,
(Dr. Ir. Ence Darmo Jaya Supena, M.Si.)
NIP: 196410021986031002
ABSTRAK
PRIMA NOVITASARI. Isolasi dan Identifikasi Bakteri Kitinolitik Penghambat Pertumbuhan Cendawan Patogen Asal Kokon Cricula trifenestrata. Dibimbing oleh NISA RACHMANIA MUBARIK dan SRI LISTIYOWATI.
Kokon sutera emas adalah pelindung pupa yang merupakan salah satu fase dari siklus hidup ngengat Cricula trifenestrata. Hasil olahan kokon ini memiliki nilai jual yang tinggi karena memiliki warna keemasan yang khas tidak dimiliki oleh benang sutera lain. Pengolahan benang sutera sering mengalami kendala dalam penurunan kualitas dan kuantitas kokon akibat serangan penyakit, di antaranya ialah cendawan patogen. Pengendalian secara kimia telah dilakukan akan tetapi penggunaan yang berlebihan akan menimbulkan dampak negatif. Oleh karena itu perlu dilakukan pengendalian hayati, salah satunya dengan memanfaatkan bakteri penghasil kitinase yang merujuk salah satu komponen terbesar dari dinding sel cendawan ialah kitin. Penelitian ini bertujuan untuk mengisolasi dan mengidentifikasi bakteri kitinolitik penghambat cendawan patogen yang berasal dari kokon Cricula trifenestrata. Isolat bakteri yang didapatkan sebanyak 17 isolat dari dua jenis sampel kokon asal pupa sehat dan sakit. Sebanyak empat isolat hasil seleksi, yaitu CH2, CH10, CS1, dan CS4 berdasarkan Indeks Kitinolitik (IK) paling tinggi. Isolat dengan aktivitas kitinase paling tinggi (CH10 dan CS1) diidentifikasi menggunakan MicrogenTM GnA+B ID System, keduanya termasuk spesies Shewanella putrefaciens. Cendawan patogen Scopulariopsis sp. dan FC (belum diidentifikasi) diisolasi dari kokon asal pupa sakit. Isolat CH10 berhasil menghambat pertumbuhan FC pada kultur berpasangan, tetapi tidak terjadi penghambatan oleh kitinase ekstrak kasarnya. Scopulariopsis sp. dapat dihambat oleh kultur maupun kitinase keempat isolat bakteri.
Kata kunci: Kokon Cricula trifenestrata, kitinase, indeks kitinolitik, cendawan patogen.
ABSTRACT
PRIMA NOVITASARI. Isolation and Identification of Chitinolytic Bacteria as an Inhibitor the Growth of Pathogenic Fungi Isolated from Cricula trifenestrata Cocoon. Supervised by NISA RACHMANIA MUBARIK and SRI LISTIYOWATI.
Gold Silkworm’s cocoon is a coat of Cricula trifenestrata’s pupa that include part of life cycle. The fiber which is resulted from processing cocoon has high economic value because the specific of gold colour that doesn’t have any else cocoon. However, on the process often occurred many problems that could decrease quantity and quality of cocoon, such as attack from pathogenic fungi. For controlling of the pathogenic fungi has been used chemical agent, but using regularly caused some negative effects. As alternative using chitinolytic bacteria as biocontrol, because of chitin is the highest component of the fungi cell wall. Therefore the objectives of this research were to isolate and identify of chitinolytic bacteria that inhibited fungi pathogen which isolated from cocoon of C. trifenestrata. 17 strains chitinolytic bacteria were isolated from two samples cocoon of health pupa and sick pupa. There were four isolates CH2, CH10, CS1, and CS4 had highest value of chitinolytic index. CH10 and CS1 strains showed highest chitinase activity, both isolates were identified using MicrogenTM GenA+B ID System and categorized as Shewanella putrefaciens. Scopulariopsis sp. and FC (unidentified) were isolated from cocoon of sick pupa. Strain CH10 inhibited FC on culture bacteria test but not from its in chitinase. Scopulariosis sp. had been inhibited by culture and its chitinase.
i
PRAKATA
Alhamdulillah segala puji dan syukur penulis panjatkan kehadirat Allah SWT atas segala rahmat dan karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan.Tema yang dipilih pada penelitian ini ialah Isolasi dan Identifikasi Bakteri Kitinolitik Penghambat Cendawan Patogen Asal Kokon Cricula trifenestrata.
Terima kasih penulis ucapkan kepada Dr. Nisa Rachmania Mubarik, M.Si. dan Dr. Sri Listiyowati, M.Si. yang telah membimbing pada penelitian ini. Terima kasih juga
disampaikan kepada dosen penguji Dr. Ir. Sulistijorini, M.Si. atas saran dan diskusi yang diberikan. Ucapan terima kasih disampaikan kepada Ibu, Bapak, Kakek, Nenek, om Taufik, mas Ayik, serta seluruh keluarga atas do’a dan kasih sayang yang telah diberikan. Terima kasih juga penulis ucapkan kepada teknisi dan teman-teman, antara lain Mbak Heni, Pak Jaka, Kak Maisya, Kak Yessi, Raya, Abdi, Aida, Nurul, Zuhay, Andri, Nia, Yani, Elsa, Inggit, Fika, Fenni, Via, Ida, Nurul, Neri, Yusti, dan teman-teman Biologi 45 atas do’a dan dukungannya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Pebruari 2013
RIWAYAT HIDUP
Penulis dilahirkan di Batang pada tanggal 31 Januari 1990 sebagai anak tunggal dari kedua orang tua yang bernama Mahfud Fauzi dan Nanik Dwi Hayanti.
Penulis menyelesaikan pendidikan dasar pada tahun 2002 di SDN 1 Adinuso Batang. Pendidikan menengah pertama diselesaikan tahun 2005 di SLTPN 1 Reban, Batang. Pendidikan menengah atas diselesaikan tahun 2008 di SMAN 1 Batang. Pada tahun 2008, penulis diterima di Institut Pertanian Bogor (IPB) melalui jalur undangan seleksi masuk IPB (USMI). Penulis memilih Mayor Biologi, Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam.
i
DAFTAR ISI
Halaman
DAFTAR TABEL ... i
DAFTAR GAMBAR ... i
DAFTAR LAMPIRAN ... i
PENDAHULUAN Latar Belakang ... 1
Tujuan ... 1
BAHAN DAN METODE Waktu dan Tempat... 2
Bahan ... 2
Isolasi dan Seleksi Bakteri Kitinolitik... 2
Isolasi dan Identifikasi Cendawan Patogen Asal Kokon ... 2
Uji Daya Hambat Bakteri Hasil seleksi dan Uji Enzim Kitinase Ekstrak Kasarnya terhadap Cendawan Patogen secara In vitro ... 2
Produksi dan Uji Aktivitas Kitinase serta Pengukuran Kadar Protein ... 3
Identifikasi Isolat Bakteri dengan Aktivitas Kitinase Paling tinggi. ... 3
HASIL Isolasi dan Seleksi Bakteri Kitinolitik... 3
Isolasi dan Identifikasi Cendawan Patogen Asal Kokon ... 4
Uji Daya Hambat Bakteri Hasil seleksi dan Uji Enzim Kitinase Ekstrak Kasarnya terhadap Cendawan Patogen secara In vitro ... 4
Produksi dan Uji Aktivitas Kitinase serta Pengukuran Kadar Protein ... 5
Identifikasi Isolat Bakteri dengan Aktivitas Kitinase Paling tinggi ... 5
PEMBAHASAN... 6
SIMPULAN ... 7
DAFTAR PUSTAKA ... 7
DAFTAR TABEL
Halaman
1 Indeks kitinolitik 17 isolat bakteri asal kokon Cricula trifenestrata pada umur 4 HSI ... 4 2 Zona hambat isolat bakteri CH10, CS1, dan CS4 terhadap FC dan Scopulariopsis sp. umur
14 HSI ... 4
3 Zona hambat pengujian kitinase ekstrak kasar dari empat isolat bakteri terhadap Scopulariopsis sp. umur 14 HSI ... 4
4 Hasil pengukuran aktivitas kitinase, kadar protein, dan aktivitas spesifik empat isolat bakteri setelah umur 24 jam ... 5
5 Hasil uji biokimia isolat CH10 dan CS1 dengan kit MicrogenTM GnA+B–ID ... 5
DAFTAR GAMBAR
Halaman
1 Kokon C. trifenestrata (a) kokon asal pupa sehat, (b) kokon asal pupa sakit ... 2 2 Morfologi cendawan patogen dari kokon asal pupa sakit (a) Scopulariopsis sp.,
(b) FC (Perbesaran 400x). ... 4
3 Penghambatan cendawan oleh isolat bakteri CH10, (a) FC umur 14 HIS, (b) kontrol. ... 4
4 Hasil pewarnaan Gram isolat bakteri, (a) CH10 dan (b) CS1 (Perbesaran 1000x). ... 5
DAFTAR LAMPIRAN
Halaman
1 Metode pengujian aktivitas kitinase (Spindler 1997) ... 10
2 Metode pengukuran kadar protein (Bradford 1976) ... 11 3 Aktivitas kitinase dan kadar protein empat isolat bakteri hasil seleksi: CH2, CH10, CS1,
dan CS4 ... 11
4 Isolat bakteri kitinolitik hasil seleksi dengan IK paling tinggi setelah inkubasi 4 hari. (a) CS1, (b) CS4,(c) CH2, (d) CH10 ... 12 5 Hasil uji daya hambat isolat bakteri terhadap cendawan patogen ... 12 6 Hasil uji daya hambat kitinase ekstrak kasar bakteri terhadap cendawan patogen. ... 13 7 Kit MicrogenTM GnA+B–ID bertipe GNB 24E yang terdiri atas GNB 12A/12E dan GNB
PENDAHULUAN
Latar Belakang
Cricula trifenestrata (Lepidoptera: Saturniidae) merupakan salah satu jenis ulat sutera liar yang sering dikenal dengan nama ulat kipat atau ulat alpukat. Ulat ini bersifat polifag, menyukai berbagai jenis inang, seperti alpukat, jambu mete, kayumanis, kedondong, mangga, kenari, cokelat, dan kina (Kalshoven 1981; Holloway 1987).
Di beberapa daerah seperti Yogyakarta dan Bali, kepompongnya mempunyai nilai ekonomi yang tinggi. Pupa diolah menjadi sumber nutrisi, sedangkan kokon dijadikan bahan kerajinan seperti ornamen, bunga, dan benang sutera (Wikardi & Djuwarso 2000; Purwanti 2005) tetapi ulat dan kokon sutera ini dapat terserang penyakit.
Penyakit yang banyak menyerang ulat sutera antara lain: Nuclear Polyhedrosis Virus
(NPV), Cytoplasmic Polyhedrosis Virus
(CPV), protozoa Nozema bombycis, Beauveria bassiana, dan Aspergillus spp. (Guntoro 1994). Serangan penyakit-penyakit tersebut mengakibatkan kualitas dan kuantitas benang sutera yang dihasilkan mengalami penurunan. Di Sulawesi Selatan lebih dari 60% ulat sutra Bombyx mori terserang oleh cendawan Aspergillus, sedangkan di India kegagalan panen kokon akibat penyakit mencapai 30 - 40% (Hee1998).
Aspergillus spp. menyerang pada fase ulat sutera. Spora cendawan tersebut menempel pada kulit larva kemudian berkembang masuk ke badan sampai masa inkubasi. Larva yang terserang menjadi cokelat kehitaman, lembek, dan mengeluarkan cairan pencernaan.
Penularan terjadi melalui kulit dengan perantara angin atau kontak badan antara ulat yang sakit dengan ulat sehat (Guntoro 1994).
Dinding sel cendawan tersusun atas kompleks kitin (polimer dari N-asetilglukosamin) dan variasi mannoprotein bersama dengan ikatan α- dan β-1,3-D- glukan. Dinding sel ini merupakan target penting untuk agens anti cendawan (Hanson 2008). Pengendalian penyakit yang diakibatkan cendawan dapat dilakukan menggunakan fungisida tetapi pemberian yang berlebihan dalam jangka waktu yang panjang dapat memberikan dampak negatif.
Penggunaan fungisida dapat menimbulkan masalah lingkungan karena residunya tidak dapat terdegradasi oleh organisme kecuali mikroorganisme tertentu. Residu tersebut terakumulasi dalam sel atau jaringan
organisme sehingga dapat meracuni organisme yang bersangkutan. Pengendali hayati dapat digunakan karena lebih aman dari pada penggunaan bahan kimia, di antaranya dengan memanfaatkan bakteri penghasil enzim pendegradasi kitin yang menghambat pertumbuhan cendawan (Neuhaus 1999).
Kitinase adalah mengkatalisis reaksi pemecahan memoton terhadap Kitinase terdiri atas dua jenis, yaitu endokitinase yang dapat memotong bagian dalam molekul kitin dan eksokitinase yang hanya memotong ujung terminal-N (ujung diperoleh dari berbagai makhluk hidup
seperti, hewan
Aronson 2007). Bakteri penghasil kitinase banyak berada pada habitat yang memiliki kandungan kitin tinggi, seperti kompos yang mengandung kitin, eksoskeleton crustaceae, air laut, sedimen laut (Donderski & Brzezinska 2001), dan tanah (Chernin 1995).
Strategi seleksi bakteri kitinolitik untuk pengendali hayati didasarkan pada kemampuan kolonisasi, kompetisi, dan kemampuan menyesuaikan diri dengan lingkungan (McQuilken et al. 1998). Bakteri kitinolitik dalam penelitian ini diisolasi dari substrat kokon C. trifenestrata. Hal ini diasumsikan bahwa bakteri tersebut telah beradaptasi dengan kondisi kokon. Gunawan (2012) melaporkan isolasi bakteri dari kokon C. trifenestrata diperoleh bakteri proteolitik sebanyak 26 isolat. Sebanyak 18 isolat dari sampel kokon asal pupa sehat dan 8 isolat dari sampel kokon asal pupa sakit. Sebanyak dua isolat dengan indeks proteolitik paling tinggi diidentifikasi sebagai Aeromonas salmonicida ssp. salmonicida dan Vibrio fluvialis (Gunawan 2012). Sebanyak 15 isolat bakteri amilolitik juga diisolasi dari pupa sutera Attacus atlas. Sebanyak dua isolat dengan indeks amilolitik paling tinggi diidentifikasi sebagai Burkholderia pseudomalei (Soraya 2012).
Tujuan
2
penghasil enzim kitinase penghambat cendawan patogen yang berasal dari kokon ulat sutera Cricula trifenestrata.
BAHAN DAN METODE Waktu dan Tempat
Penelitian dilaksanakan pada bulan Januari - Oktober 2012 bertempat di Laboratorium Mikrobiologi dan Mikologi Departemen Biologi, FMIPA, IPB.
Bahan
Kokon ulat sutera C. trifenestrata berasal dari pohon alpukat yang tumbuh di Desa Cikoneng, Bogor, Jawa Barat.
Isolasi dan Seleksi Bakteri Kitinolitik Sampel yang digunakan dalam isolasi dan seleksi bakteri kitinolitik ialah kokon C. trifenestrata asal pupa sakit dan kokon asal pupa sehat sebagai pembanding. Kokon C. trifenestrata memiliki bentuk seperti jaring, memiliki pori yang besar, berwarna kuning, dan tidak berbau (Kalshoven 1981). Sampel kokon asal pupa sakit memiliki ciri berbeda, yaitu tampak berwarna cokelat kehitaman dibandingkan dengan kokon asal pupa sehat yang bersih dan berwarna kuning keemasan (Gambar 1).
Gambar 1 Kokon C. trifenestrata. (a) Kokon asal pupa sehat, (b) kokon asal pupa sakit.
Sebanyak masing-masing 3 g kokon dimasukkan ke dalam 100 ml Nutrient Broth (NB) yang mengandung 0,5% koloidal kitin
dalam erlenmeyer 500 ml, kemudian
diinkubasi pada mesin penggoyang pada suhu ruang selama 24 jam. Isolasi bakteri kitinolitik diawali dengan melakukan pengenceran serial dari 10-1 sampai 10-8 dalam garam fisiologis NaCl 0,85%. Suspensi dituang pada medium agar-agar kitindan diinkubasi selama 24 jam pada suhu 37oC.
Penggoresan kuadran dilakukan untuk mendapatkan isolat murni. Masing-masing isolat diinokulasi kembali pada agar-agar kitin dan diinkubasi selama empat hari pada suhu 37oC. Isolat diamati berdasarkan indeks kitinolitik (IK) dengan membandingkan antara
zona bening di sekitar koloni bakteri dengan diameter koloni.
Identifikasi hasil isolat bakteri meliputi ciri-ciri morfologi dan pewarnaan Gram. Isolat dengan IK paling tinggi dari masing-masing sampel digunakan dalam uji daya hambat terhadap cendawan patogen.
Isolasi dan Identifikasi Cendawan Patogen Asal Kokon
Isolasi cendawan patogen diawali dengan memotong kokon asal pupa sakit secara membujur menjadi dua bagian. Potongan
direndam di dalam campuran 2 ml
kloroxilenol 0,8% dan 100 ml akuades steril. Potongan kokon diletakkan di atas medium Potato Dextrose Agar (PDA) yang telah ditambahkan dengan kloramfenikol 500 mg/L kemudian diinkubasi selama lima hari pada
suhu 25oC. Cendawan yang tumbuh
dipindahkan pada medium PDA yang baru sehingga didapatkan kultur cendawan murni.
Langkah awal pembuatan preparat cendawan patogen dilakukan dengan metode Riddle (Riddle 1950). Sepotong agar-agar (1cm x 1 cm x 0,5 cm) diletakkan pada bagian tengah kaca objek steril dan keempat sisinya digores inokulasi cendawan dengan jarum inokulasi. Kaca penutup steril diletakkan pada bagian atas potongan agar-agar yang telah diinokulasikan cendawan. Kaca objek ini diletakkan di dalam cawan Petri steril yang berisi kertas saring lembap dan diinkubasi selama tiga hari pada suhu ruang.
Cendawan yang telah tumbuh pada kaca penutup dan kaca objek dipindahkan untuk dibuat preparat semipermanen menggunakan laktofenol kemudian diamati menggunakan mikroskop. Identifikasi menggunakan buku identifikasi Barnett (Barnett & Hunter 1987).
Uji Daya Hambat Bakteri Hasil Seleksi dan Uji Enzim Kitinase Ekstrak Kasarnya terhadap Cendawan Patogen secara In vitro
Isolat bakteri dengan IK paling tinggi dari hasil yang sudah ada digunakan dalam metode ini. Pengujian dilakukan dengan metode kultur berpasangan, yaitu dengan cara bakteri ditumbuhkan pada medium NB dan diinkubasi selama 24 jam. Sebanyak 50 µl kultur bakteri 105 sel diteteskan di atas permukaan kertas cakram yang diletakkan pada permukaan atas medium PDA. Potongan biakan murni cendawan (Φ 0,5 cm) diinokulasikan berhadapan dengan cakram yang berisi kultur dari bakteri dengan jarak 3 cm.
Uji kitinase ekstrak kasar dilakukan sama seperti uji daya hambat yang menggunakan
3
kultur bakteri, akan tetapi kultur digantikan dengan enzim kitinase ekstrak kasar dari bakterinya. Kitinase ekstrak kasar dari bakteri diperoleh dengan menumbuhkan 1 ml suspensi kultur bakteri pada 20 ml NB yang mengandung 0,5% koloidal kitin pada mesin penggoyang dengan suhu 37oC selama 24 jam. Kultur disentrifugasi dengan kecepatan 10.000 rpm (rotor FE13) selama 10 menit. Sebanyak 50 µl supernatannya diteteskan di atas permukaan kertas cakram dan diletakkan pada permukaan atas medium PDA. Potongan biakan murni cendawan (Φ 0,5 cm) diinokulasikan berhadapan dengan cakram yang berisi kitinase dari bakteri dengan jarak 3 cm. Sebagai kontrol, cakram diisi dengan akuades steril. Masing-masing perlakuan dilakukan sebanyak dua kali ulangan. Biakan diinkubasi pada suhu ruang selama tujuh hari. Pertumbuhan cendawan diketahui dengan mengukur diameter zona hambat di sekitar koloni. Perhitungan zona hambat dapat ditentukan dengan cara: Zona pertumbuhan cendawan =
2 D2
D1+ , D1 adalah diameter
zona pertumbuhan miselium terpanjang, D2 adalah diameter pertumbuhan miselium
terpendek. Persentase penghambatan (% Z) = [(C-T)/C] x 100% (Udomsilp et al. 2009); % Z: persen zona hambat, C: rata-rata diameter miselium pada kontrol, T: rata-rata diameter miselium pada perlakuan.
Produksi dan Uji Aktivitas Kitinase serta Pengukuran Kadar Protein
Sebanyak dua lup isolat bakteri
diinokulasikan ke dalam 20 ml NB yang mengandung 0,5% koloidal kitin. Kultur diinkubasi pada mesin penggoyang dengan suhu 37oC dan 100 rpm selama 24 jam. Kultur kemudian disentrifugasi pada 10.000 rpm (rotor FE13) selama 10 menit untuk mendapatkan enzim kitinase ekstrak kasar di dalam supernatan.
Aktivitas kitinase diukur sebagaimana dijelaskan oleh Spindler (1997), yaitu sebanyak 225 µl enzim ditambahkan pada 450 µl 0,3% koloid kitin dan 225 µl 0,1 M bufer fosfat pH 7. Campuran enzim diinkubasikan
pada suhu 55oC selama 30 menit. Reaksi enzim dihentikan pada suhu 100oC selama 10 menit kemudian disentrifugasi pada 10.000 rpm (rotor FE13) selama 10 menit. Sebanyak 300 µl filtratnya ditambahkan pada 750 µl aquades steril dan 1500 µl reagen Schales dan campuran didihkan pada suhu 100oC selama 10 menit (Lampiran 1). Aktivitas enzim ditentukan dengan pengukuran absorbansi
pada λ 420 nm. Satu unit aktivitas enzim didefinisikan sebagai rata-rata jumlah enzim yang dihasilkan dari 1 µmol gula (N-asetil glukosamin) per menit. Konsentrasi N-asetilglukosamin dihitung berdasarkan kurva standar N-asetilglukosamin (Lampiran 3).
Kadar protein diukur dengan metode
Bradford (Bradford 1976), yaitu dengan cara mencampurkan 100 µl enzim kasar yang dihasilkan oleh masing-masing isolat dengan 5 ml pereaksi Bradford. Campuran kemudian dikocok dengan vorteks dan diinkubasi pada suhu kamar selama 15 menit. Absorbansi larutan dibaca pada λ 595 nm (Lampiran 2). Kadar protein dihitung berdasarkan kurva standar Bovin Serum Albumin (BSA) (Lampiran 3). Kadar protein yang diperoleh digunakan untuk menentukan aktivitas spesifik enzim kitinolitik.
Identifikasi Isolat Bakteri dengan Aktivitas Kitinase Paling tinggi
Sebanyak dua isolat bakteri dengan aktivitas kitinase paling tinggi diidentifikasi menggunakan kit MicrogenTM GnA+B–ID System. Sebanyak satu lup bakteri digoreskan pada medium NA di agar-agar miring dan diinkubasi selama 24 jam. Sebanyak dua lup kultur bakteri diencerkan ke dalam 3 ml larutan garam fisiologis 0,85 % steril kemudian dikocok dengan vorteks. Sebanyak 200 µl kultur hasil pengenceran dimasukkan ke dalam setiap lubang pada kit dan diinkubasi selama 24 jam.
Pengamatan dilakukan berdasarkan perubahan warna yang disesuaikan dengan prosedur kit. Hasil yang diperoleh kemudian diproses menggunakan software MicrogenTM GnA+B–ID System 2009.
HASIL
4
Tabel 1 Indeks kitinolitik 17 isolat bakteri asal kokon Cricula trifenestrata pada umur 4 HSI
Isolat Bakteri Φ Koloni (cm) Φ Zona Bening (cm) Indeks Kitinolitik
CS1 2,43 0,46 0,19
CS2 2,46 0,32 0,16 CS3 1,93 0,17 0,09
CS4 1,61 0,25 0,17
CH1 1,67 0,40 0,24
CH2 0,83 0,47 0,60
CH3 1,13 0,53 0,45 CH4 1,13 0,53 0,40 CH5 1,13 0,53 0,45 CH6 1,43 0,53 0,37 CH7 0,70 0,20 0,30 CH8 0,70 0,20 0,30 CH9 1,50 0,40 0,50
CH10 1,16 0,80 0,74
CH11 1,80 0,40 0,10 CH12 2,20 0,60 0,30 CH13 1,27 0,20 0,20
Isolasi dan Identifikasi Cendawan Patogen Asal Kokon
Isolasi cendawan patogen dari kokon asal pupa sakit diperoleh dua isolat dengan kenampakan koloni yang berbeda (Gambar 2). Identifikasi secara morfologi menunjukkan isolat tersebut adalah Scopulariopsis sp. dan satu isolat belum dapat diidentifikasi (FC, fungi diisolasi dari Cricula trifenestrata).
Gambar 2 Morfologi cendawan patogen dari kokon asal pupa sakit (a) Scopulariopsis sp., (b) FC (Perbesaran 400x).
Uji Daya Hambat Bakteri Hasil seleksi dan Uji Enzim Kitinase Ekstrak Kasarnya terhadap Cendawan Patogen secara In vitro
Sebanyak empat isolat bakteri hasil seleksi (CH2, CH10, CS1, dan CS4) mampu menghambat Scopulariopsis sp. (Lampiran 5), akan tetapi hanya CH10 yang mampu menghambat FC (Gambar 3).
Gambar 3 Penghambatan cendawan oleh isolat bakteri CH10 (a) FC umur 14 HSI, (b) kontrol.
Zona hambat isolat bakteri CS1 terhadap Scopulariopsis sp. paling besar (19,33%) daripada isolat CH2 (3,40%), CH10 (11,31%), dan CS4 (17,60%) (Tabel 2).
Tabel 2 Zona hambat isolat bakteri CH10, CS1, dan CS4 terhadap FC dan Scopulariopsis sp. umur 14 HSI
Isolat
Bakteri Isolat Cendawan
Zona Hambat
(%)
CH10 FC 11,31
CH10 Scopulariopsis sp. 16,48 CH2 Scopulariopsis sp. 3,40 CS1 Scopulariopsis sp. 19,33 CS4 Scopulariopsis sp. 17,60
Kitinase ekstrak kasar asal empat isolat bakteri hasil seleksi mampu menghambat Scopulariopsis sp. Isolat CH2 memiliki persen penghambatan yang lebih besar (19,88%) terhadap Scopulariopsis sp. dibandingkan dengan tiga isolat lain (Tabel 3). Namun, kitinase ekstrak kasar CH10 tidak mampu menghambat pertumbuhan FC (Lampiran 6)
Tabel 3 Zona hambat pengujian kitinase ekstrak kasar dari empat isolat bakteri terhadap Scopulariopsis sp. umur 14 HSI
Sumber enzim Zona hambat (%)
CH10 14,21
CH2 19,88
CS1 6,26
CS4 5,11
a
b
a b
FC
CH10
5
Produksi dan Uji Aktivitas Kitinase serta Pengukuran kadar Protein
Pengukuran aktivitas kitinase secara kuantitatif bertujuan untuk mengetahui hubungan antara IK dan zona penghambatan oleh isolat maupun kitinase ekstrak kasarnya terhadap pertumbuhan cendawan patogen. Isolat bakteri CS1 memiliki aktivitas kitinase (0,09 U/ml), kadar protein (0,08 mg/ml), dan aktivitas spesifik (2,84 U/ml) yang lebih tinggi dibandingkan dengan tiga isolat lain (Tabel 4).
Tabel 4 Hasil pengukuran aktivitas kitinase, kadar protein, dan aktivitas spesifik empat isolat bakteri setelah umur 24 jam Isolat Bakteri Aktivitas Kitinase (U/ml) Kadar Protein (mg/ml) Aktivitas Spesifik (U/ml)
CH2 0,06 0,05 1,19
CH10 0,08 0,07 1,20
CS1 0,09 0,08 2,84
CS4 0,03 0,08 0,41
Identifikasi Isolat Bakteri dengan Aktivitas Kitinase Paling tinggi
Kedua isolat dengan aktivitas kitinase paling tinggi (CS1 dan CH10) memiliki koloni dengan bentuk bundar dan warna putih. Tepian koloni CS1 berombak, sedangkan CH10 tak beraturan. Berdasarkan pewarnaan Gram, kedua isolat berbentuk batang dan Gram negatif (Gambar 5).
Gambar 5 Hasil pewarnaan Gram isolat bakteri CH10 dan (b) Isolat bakteri CS1 (Perbesaran 1000x).
Identifikasi menggunakan kit MicrogenTM GnA+B–ID System dilakukan berdasarkan uji biokimiawi yang terkandung di dalam bakteri (Tabel 5)
Tabel 5 Hasil uji biokimia isolat CH10 dan CS1 dengan kit MicrogenTM GnA+B– ID
Uji Biokimia
Isolat
CH10 CS1
Nitrat + +
Lisin - -
Ornitin - -
H2S + +
Glukosa - -
Mannitol - -
Xilosa - -
ONPG - -
Indol - -
Urease - -
VP - -
Sitrat - -
TDA - -
Gelatin + +
Malonat - -
Inositol - -
Sorbitol - -
Ramnosa - -
Sukrosa - -
Laktosa - -
Arabinosa - -
Adonitol - -
Rafinosa - -
Salisin - -
Arginin - +
Berdasarkan identifikasi menggunakan kit MicrogenTM GnA+B–ID System, CH10 dan CS1 memiliki kemiripan dengan Shewanella putrefaciens masing-masing sebesar 99% dan 94%.
PEMBAHASAN
Sebanyak 17 isolat bakteri kitinolitik berhasil diisolasi menggunakan agar-agar nutrien dengan penambahan koloidal kitin sebagai penginduksi. Koloidal kitin merupakan salah satu substrat yang umum digunakan untuk menginduksi enzim hidrolitik seperti kitinase (Haran & Chet 1995). Suspensi koloidal kitin ini merupakan kitin yang dilarutkan dalam asam klorida pekat seperti yang telah dipelajari oleh Hsu dan Lockwood (1975) sebagai media selektif untuk mendapatkan Actinomycetes dari air dan tanah. Mikroorganisme kitinolitik dapat dideteksi dengan adanya zona bening di sekitar koloni. Sebanyak empat isolat bakteri (CH2, CH10, CS1, dan CS4) merupakan hasil seleksi dari 17 isolat berdasarkan indeks kitinolitik (IK) tertinggi. Isolat bakteri CH10 memiliki IK paling tinggi dengan nilai 0,74. Indeks kitinolitik isolat bakteri CH10 (0,74)
a 6 µm b
6
memiliki nilai yang lebih rendah dibandingkan dengan nilai rata-rata IK bakteri kitinolitik hasil isolasi dari perakaran tanaman cabai yaitu sebesar 1,00 (Nurdebyandaru et al. 2008). Namun demikian, isolat bakteri CH10
memiliki nilai IK yang lebih tinggi
dibandingkan dengan isolat bakteri kitinolitik dari rizosfer tanaman karet. Hasil penapisan bakteri kitinolitik dari rizosfer tanaman karet Sembawa Kabupaten Banyuasin memiliki nilai IK paling besar 0,52 (Muharni & Widjajanti 2011). Sebanyak 17 isolat bakteri asal rumen memiliki nilai rata-rata sebesar 0,7 dengan kisaran 0,2 sampai dengan 2,5 (Rahayu et al. 2003). Isolat bateri CH10 termasuk dalam kisaran tersebut. Keempat isolat bakteri hasil seleksi (CH2, CH10, CS1, dan CS4) digunakan sebagai isolat uji dalam proses penghambatan pertumbuhan cendawan patogen dari kokon asal pupa sakit.
Sebanyak dua cendawan patogen hasil isolasi memiliki kenampakan morfologi yang berbeda. Scopulariopsis sp. termasuk dalam filum Ascomycota yang bersifat kosmopolitan di tanah, tanaman, dan serangga. Cendawan ini juga berhasil diisolasi dari permukaan tubuh tungau Psoroptes cuniculi yang merupakan ektoparasit pada kelinci, kambing, dan kuda (Perucci et al. 2008). Scopulariopsis sp. tumbuh dalam waktu cukup cepat dan matang dalam waktu lima hari. Koloni berbentuk seperti bubuk dengan permukaan berwarna putih yang lama kelamaan berubah menjadi cokelat keabu-abuan. Beberapa spesies dalam genus Scopulariopsis memiliki warna koloni gelap (McGinnis & Pasarell 1998). Berdasarkan kenampakan secara mikroskopik, hifa Scopulariopsis sp. berseptat dan hialin. Konidiofor gelap, tersusun secara sederhana atau bercabang. Konidium hialin atau gelap, satu sel, dan tersusun secara basipetal (Tosti et al. 1996). Genus ini menyerang larva lebah, ulat sutera, dan menyebabkan penyakit onychomycosis pada manusia (Tosti et al. 1996).
Koloni cendawan FC (belum teridentifikasi) bertekstur seperti bubuk dan awalnya berwarna putih kemudian berubah menjadi cokelat dan hitam. Kenampakan cendawan FC secara mikroskopis seperti berpiknidium dengan spora berbentuk batang berwarna hialin. Cendawan Scopulariopsis sp. dan FC digunakan sebagai isolat uji dalam penghambatan oleh empat isolat bakteri hasil seleksi.
Sebanyak empat isolat bakteri hasil seleksi (CH2, CH10, CS1, dan CS4) dan kitinase ekstrak kasarnya mampu
menghambat pertumbuhan Scopulariopsis sp. Hal ini membuktikan bahwa enzim kitinase berperan dalam mekanisme penghambatan Scopulariopsis sp. CS1 memiliki zona penghambatan paling besar pada pengujian kultur, yaitu sebesar 19,33% tetapi hanya 6,25% pada pengujian enzim kitinase ekstrak kasarnya. Kultur bakteri CH10 berhasil menghambat pertumbuhan FC, akan tetapi pada pengujian enzim kitinase ekstrak kasarnya tidak mampu menghambat cendawan FC. Perbedaan hasil tersebut dapat diasumsikan bahwa enzim kitinase ekstrak kasar tidak dapat menghambat FC.
Variasi perbedaan penghambatan ini, di antaranya disebabkan oleh kespesifikan spesies, perbedaan aktivitas kitinase bakteri, komposisi kitin dari dinding sel cendawan, dan keberadaan metabolit anticendawan. Dinding sel cendawan pada umumnya tidak hanya tersusun atas kitin tetapi juga tersusun oleh gula lain, misalnya adalah β-1,3 glukan yang berikatan dengan kitin. Dinding sel cendawan tersusun atas kitin dan glukan sehingga β-1,3 glukanase dan kitinase adalah enzim yang bertanggung jawab dalam degradasi dinding sel cendawan (Anand & Reddy 2009; Gohel et al. 2006).
Aktivitas enzim kitinase isolat CS1 memiliki nilai paling tinggi dibandingkan dengan tiga isolat lain, yaitu sebesar 0,091 U/ml. Ketidaksesuaian besar aktivitas enzim dengan zona hambat CS1 pada pengujian enzim kitinase (6,25%) membuktikan keduanya tidak saling berhubungan. Aktivitas kitinase yang tinggi tidak selalu berkorelasi dengan kemampuan menghambat pertumbuhan cendawan patogen. Hal ini karena perbedaan struktur pada substrat yang digunakan. Struktur kitin pada dinding sel cendawan lebih kompleks dibandingkan dengan koloidal kitin yang digunakan sebagai sumber karbon pada uji aktivitas kitinase. Hal yang sama juga terjadi pada Trichoderma harzianum T-35 dan T-203. Kedua galur mensekresikan enzim kitinase ketika ditumbuhkan pada medium yang mengandung kitin. T-203 mensekresikan kitinase 90% lebih banyak dibandingkan dengan T-35. Namun demikian, pada uji in vivo hanya T-35 yang efektif sebagai mikoparasit atau agen biokontrol. Hal ini membuktikan bahwa persentase produksi kitinase tidak selalu berkorelasi dengan kemampuannya untuk melakukan penghambatan atau sebagai agen biokontrol (Shivan & Chet 1989).
7
System, kedua isolat (CH10 dan CS1) dengan
aktivitas kitinase paling tinggi hanya berbeda pada pengujian arginin. Uji ini untuk mengetahui adanya perubahan arginin menjadi ornitin, ammonia, dan CO2 oleh arginin dihidrolase (Lapageet al. 1973). Hasil identifikasi menunjukkan kedua isolat memiliki kemiripan dengan spesies Shewanella putrefaciens. Isolat CH10 dan CS1 masing-masing memiliki kemiripan sebesar 99% dan 94%. Menurut Gooday (1990), prokariot pendegradasi kitin pada umumnya adalah Pseudomonas, Vibrio, Photobacterium, Actinomycetes, Bacillus, Clostridium, dan Enterobacter. Shewanella putrefaciens termasuk dalam kelompok Gram negatif dan berbentuk batang. Panjang sel sekitar 2-3 µm dengan diameter 0,4-0,7 µm dan bersifat motil (Venkateswaran et al. 1999).
Biakan S. putrefaciens merupakan bakteri penghasil enzim kitinase dengan aktivitas paling tinggi pada hari ke- 4-7 inkubasi. Shewanella putrefaciens memiliki tingkat virulensi yang lebih rendah dibandingkan dengan Shewanella algae (Khashe & Janda 1998). Selain penghasil enzim kitinase, S. putrefaciens juga menghasilkan enzim hyaluronidase, lipase, protease, α-glucosidase, aminopeptidase, dan galaktinase (Khashe & Janda 1998). Khashe dan Janda (1998) membuktikan secara biokimia bahwa S. putrefaciens menghasilkan siderofor yang merupakan salah satu agens biokontrol. Produksi siderofor terjadi pada hari ke-3-7 inkubasi. Hal inilah yang diduga menjadi penyebab S. putrefaciens hanya mampu menghambat FC pada pengujian kultur tetapi pada kitinase ekstrak kasarnya tidak terjadi penghambatan.
Salah satu senyawa yang dihasilkan bakteri antagonis adalah siderofor. Menurut Maurhofer et al. (1998) siderofor merupakan molekul berbobot molekul relatif rendah yang dihasilkan oleh plant growth promoting rhizobacteria sebagai agens pengelat ion Fe. Siderofor dihasilkan oleh bakteri maupun cendawan yang tumbuh dalam keadaan cekaman Fe rendah. Molekul ini merupakan senyawa organik yang dapat digunakan dalam pengendalian penyakit tanaman. Mekanisme pengendalian bakteri terhadap cendawan penyebab penyakit tanaman diduga secara kompetisi. Bakteri dan cendawan dapat menghasilkan siderofor untuk mengelat ion Fe dari lingkungan, tetapi daya kelat bakteri terhadap Fe lebih tinggi dibandingkan dengan cendawan sehingga pertumbuhan cendawan
terhambat akibat kekurangan ion Fe (Maurhofer et al. 1998).
Sifat antagonis bakteri penghasil siderofor inilah yang diduga mampu menghambat pertumbuhan cendawan. Pseudomonas memiliki kemampuan untuk menghasilkan siderofor dalam jumlah yang tinggi. Siderofor ini diketahui efektif menekan pertumbuhan penyakit Fusarium oxysporum. Hal ini karena ion Fe yang dibutuhkan F. oxysporum untuk berkecambah tidak tersedia akibat dikelat oleh siderofor dari Pseudomonas (Maurhofer et al. 1998).
SIMPULAN
Sebanyak 17 isolat bakteri kitinolitik berhasil diisolasi dari sampel kokon Cricula trifenestrata. Ada empat isolat yang memiliki indeks kitinolitik tertinggi, yaitu: CH2, CH10, CS1, dan CS4. Isolat CH10 dan CS4 menunjukkan aktivitas kitinase paling tinggi dan diidentifikasi sebagai Shewanella putrefaciens. Cendawan Scopulariopsis sp. dan FC (belum teridentifikasi) berhasil diisolasi dari sampel kokon asal pupa sakit. Keempat kitinase ekstrak kasar yang dihasilkannya mampu menghambat pertumbuhan Scopulariopsis sp. Hanya isolat bakteri CH10 yang berhasil pertumbuhan FC dalam kultur berpasangan, tetapi tidak terjadi penghambatan oleh kitinase ekstrak kasarnya.
DAFTAR PUSTAKA
Anand S, Reddy J. 2009. Biocontrol potential of Sp. against plant pathogens. Int J Agric Sci 1: 30-39.
Barnett HL, Hunter BB. 1987. Illustratred Genera of Imperfecti Fungi, FourthEdition. New York: Macmilan Publishing Company.
Bradford MM. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72: 248-254.
Funkhouser JD, Aronson NN. 2007. Chitinase family GH18: evolutionary insights from the genomic history of a diverse protein family. BMC Evol Biol 7: 96-111. Gohel V, Singh A, Vimal M, Ashwini P,
8
Gooday GW. 1990. The ecology of chitin degradation advance. Microbiol Ecol 11: 387-430.
Gunawan A. 2012. Isolasi dan karakterisasi bakteri proteolitik asal kokon Cricula trifenestrata yang mempengaruhi kualitas sutera emas [skripsi]. Bogor: Institut Pertanian Bogor.
Guntoro S. 1994. Budidaya Ulat Sutera. Kanisius: Yogyakarta.
Hanson JR. 2008. The Chemistry of Fungi. Cambridge: RSC Publishing.
Haran S, Chet I. 1995. New components of the chitinolytic system of Trichoderma harzianum. Mycol Rev 94: 441-446. Hee RC. 1998. Panduan Teknis Persuteraan
Alam Indonesia. Surabaya: PT Indojado Sutera Pratama, Silk Industri.
Holloway JD. 1987. The moth of Borneo Part 3: Supefamily Bomnicoidea: families Lasiocampidae, Eupterotidae, Bombycidae, Brahmaeidae, Saturniidae, Sphingidae. Kuala Lumpur: Southene Shn Bhd.
Hsu SC, Lockwood JL. 1975. Powdered chitin agar as a selective medium for enumeration of actinomycetes from water and soil. J Applied Microbiol 29: 422-426.
Kalshoven LGE. 1981. Pest of Crop in Indonesia. Laan PA van der, penerjemah. Jakarta:Ichtiar baru/Van Hoeve.
Khashe S, Janda M. 1998. Biochemical and pathogenic properties of Shewanella alga and Shewanella putrefaciens. J Clin Microbiol 36: 783-787.
Lapage SP, Bascombe S, Willcox WR, Curtis MA. 1973. Identification bacteria by computer: general aspects and perspectives. J Gen Microbiol 77: 273-290.
Maurhofer M, Reimman C, Schmidli-Sacherer P, Heep S, Defago G. 1998. Salysilic acid biosynthetic genes expressed in
Pseudomonas fluorescens strain P3
improve the induction of systemic resistance in tobacco against Tobacco
necrosis virus. Phytopathology
88:678-684.
McGinnis MR, Pasarell L. 1998. In vitro testing of susceptibilities of filamentous Ascomycetes to voriconazole, itraconazole, and amphotericin B, with consideration of phylogenetic implications. J Antimicrob Agents Chemother 36:2353-2355.
McQuilken MP, Halmer P, Rhodes DJ. 1998. Application of microorganisms to seeds. In Formulation of microbial biopesticides: Beneficial microorganisms, nematodes and seed treatments. Dordrecht: Kluwer Academic Press. Muharni, Widjajanti H. 2011. Skrining bakteri
kitinolitik antagonis terhadap pertumbuhan jamur akar putih (Rigidopus lignosus) dari rizosfer tanaman karet. J Penel Sains 14: 51-56.
Neuhaus JM. 1999. Plant chitinase (3, PR-4, PR-8, PR-11). Di dalam: Datta SK, Muthukrishnan S, editor. Pathogenesis-Related Proteins in Plant. London: CRC Pr. hlm 77-105.
Nurdebyandaru N, Mubarik NR, Prawasti TS. 2008. Chitinolytic bacteria isolated from chili rhizospher: chitinase characterization and application as biocontrol for Aphis gossypii. J Microbiol Indonesi 4: 103-107.
Perucci S, Zini S, Donadio E, Mancianti F, Fichi G. 2008. isolation of Scopulariopsis spp. fungi from Psoroptes cuniculi body surface and evaluation of their entomopathogenic role. J Parasitol Res 102: 957-962.
Purwanti. 2005. Analisis kadar protein dan lemak kepompong ulat sutera emas (Cricula trifenestrata) [skripsi]. Malang: Departemen Biologi Universitas Muhammadiyah Malang.
Rahayu S, Suhartati FM, Rimbawanto EA, Iriyanti N. 2003. Isolasi dan identifikasi bakteri kitinolitik asal rumen. J Anim Prod 5: 73-78.
Riddle RW. 1950. Permanent stained mycological preparation obtained by slide culture. Mycologia 42: 265-70.
Shivan A, Chet I. 1989. Degradation of fungal cell walls by lytic enzymes of Trichoderma harzianum. J Gen Microbiol 135: 675-682.
Soraya F. 2012. Isolasi dan karakterisasi bakteri amilolitik asal pupa ulat sutera Attacus atlas [skripsi]. Bogor: Institut Pertanian Bogor.
Spindler KD. 1997. Chitinase and chitonase assays. Di dalam: Muzarelli RAA, Peter MG, editor. Chitin Handbook. Grottamare: Alda Tecnografica. hlm 229-35.
9
Udomsilp J, Piyo A, Khang-Khun P, Thobunluepop P. 2009. Antifungal properties of essential oils from Thai medical plants againts rice pathogenic fungi. J Food Ag-Ind 24-30.
Venkateswaran K et al. 1999. Polyphasic taxonomy of the Shewanella and description of Shewanella oneidensis sp. nov. Int J Syst Bacteriol 49: 705-724.
11
Lampiran 1 Metode pengujian aktivitas kitinase (Spindler 1997)
Kultur sel disentrifugasi (10.000 rpm, 10’)
Supernatan (enzim ekstrak kasar)
Bahan Sampel (µl) Kontrol-A (µl) Kontrol-B (µl)
Koloidal kitin 450 450 -
Buffer fosfat 225 225 -
Enzim ekstrak kasar 225 - 225
Inkubasi (55oC, 30’)
Inkubasi dihentikan (100oC, 10’)
Didinginkan 10’
Kontrol-A dicampurkan dengan kontrol-B menjadi kontrol
Sampel dan kontrol masing-masing dimasukkan ke dalam tabung mikro
kemudian disentrifugasi (10.000 rpm, 5’)
Supernatan yang dihasilkan merupakan enzim campuran
Bahan Sampel (µl) Kontrol (µl) Blanko (µl)
Enzim campuran 300 300 -
Akuades steril 750 750 1050
Reagen Schales 1500 1500 1500
Reaksi dihentikan (100oC, 10’)
12
Lampiran 2 Metode pengukuran kadar protein (Bradford 1976)
Pereaksi Blanko (ml) Standar (ml) Sampel (ml)
Standar protein - 0,1 -
Akuades 0,1 - -
Enzim - - 0,1
Bradford 5 5 5
Campuran dikocok kuat dengan vortex, didiamkan 20’ Absorbansi diukur pada λ 595 nm
Lampiran 3 Aktivitas kitinase dan kadar protein empat isolat bakteri terpilih (CH2, CH10, CS1, dan CS4)
Kurva standar N-asetilglukosamin
13
Lampiran 4 Isolat bakteri kitinolitik terpilih dengan IK tertinggi setelah inkubasi 4 hari. (a) CS1, (b) CS4,(c) CH2, (d) CH10.
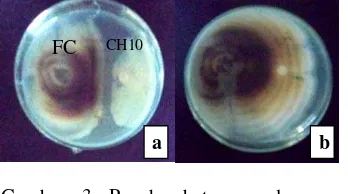
Lampiran 5 Hasil uji daya hambat isolat bakteri (kanan) terhadap cendawan patogen (kiri). (a-d) isolat bakteri CH2, CH10, CS1, dan CS4 (berurutan dari kiri ke kanan) terhadap cendawan FC, (e) kontrol akuades steril terhadap FC, (f-i) isolat bakteri CH2, CH10, CS1, dan CS4 (berurutan dari kiri ke kanan) terhadap Scopulariopsis sp., (j) kontrol akuades steril terhadap Scopulariopsis sp. (14 HSI).
Zona hambat isolat bakteri CH10, CS1, dan CS4 terhadapFC dan Scopularipsis sp.
IsolatBakteri Isolat Cendawan Diameter Cendawan pada kontrol (cm2)
Diameter Cendawan pada Perlakuan (cm2)
Zona Hambat (%)
CH10 FC 3,98 3,53 11,31
Scopulariopsis sp. 2,93 2,45 16,48
CH2 Scopulariopsis sp. 2,93 2,83 3,40
CS1 Scopulariopsis sp. 2,93 2,37 19,33
CS4 Scopulariopsis sp. 2,93 2,42 17,60
Rumus:
Diameter Cendawan =
2
g terpanjan hifa
terpendek
hifa φ
φ +
% Zona Hambat =
kontrol pada cendawan
perlakuan
pada cendawan
-kontrol pada cendawan
φ
φ φ
d
a b c
b a
g f
e d
c
j i
h
1 cm
1 cm
1 cm
14
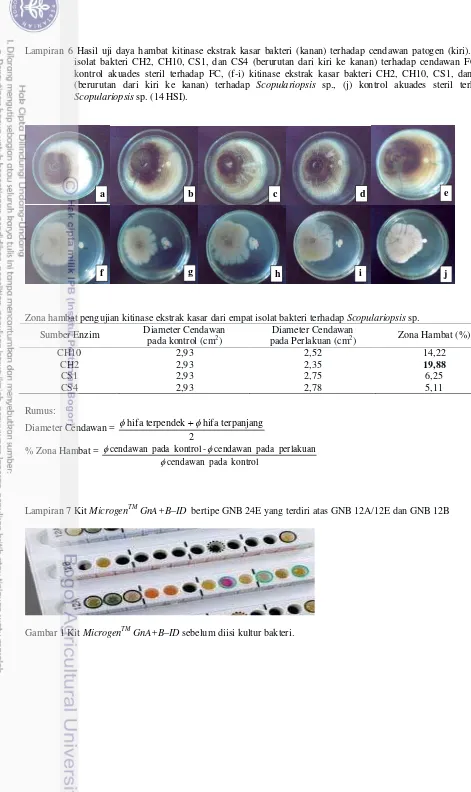
Lampiran 6 Hasil uji daya hambat kitinase ekstrak kasar bakteri (kanan) terhadap cendawan patogen (kiri). (a-d) isolat bakteri CH2, CH10, CS1, dan CS4 (berurutan dari kiri ke kanan) terhadap cendawan FC, (e) kontrol akuades steril terhadap FC, (f-i) kitinase ekstrak kasar bakteri CH2, CH10, CS1, dan CS4 (berurutan dari kiri ke kanan) terhadap Scopulariopsis sp., (j) kontrol akuades steril terhadap Scopulariopsis sp. (14 HSI).
Zona hambat pengujian kitinase ekstrak kasar dari empat isolat bakteri terhadap Scopulariopsis sp.
Sumber Enzim Diameter Cendawan
pada kontrol (cm2)
Diameter Cendawan
pada Perlakuan (cm2) Zona Hambat (%)
CH10 2,93 2,52 14,22
CH2 2,93 2,35 19,88
CS1 2,93 2,75 6,25
CS4 2,93 2,78 5,11
Rumus:
Diameter Cendawan =
2
g terpanjan hifa
terpendek
hifa φ
φ +
% Zona Hambat =
kontrol pada cendawan
perlakuan
pada cendawan
-kontrol pada cendawan
φ
φ φ
Lampiran 7 Kit MicrogenTM GnA+B–ID bertipe GNB 24E yang terdiri atas GNB 12A/12E dan GNB 12B
Gambar 1 Kit MicrogenTM GnA+B–ID sebelum diisi kultur bakteri.
a b c d e
15
Gambar 2 Kit MicrogenTM GnA+B–ID setelah diisi kultur bakteri CH10 dan CS1.
Keterangan:
A: Lisin M: Gelatin
B: Orinitin N: Malonat
C: H2S O: Inositol
D: Glukosa P : Sorbitol
E: Mannitol Q: Rhamnosa
F: Xilosa R: Sukrosa
G: ONPG S: Laktosa
H: Indol T: Arabinosa
I: Urease U: Adonitol
J: VP V: Raffinosa
K: Sitrat W:Salisin
L : TDA X: Arginin