EFIKASI VAKSIN ULANG
Streptococcus agalactiae
PADA
INDUK IKAN NILA TERHADAP IMUNITAS MATERNAL
UNTUK PENCEGAHAN STREPTOCOCCOSIS
DENDI HIDAYATULLAH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Efikasi Vaksin Ulang Streptococcus agalactiae pada Induk Ikan Nila terhadap Imunitas Maternal untuk Pencegahan Streptococcosis adalah benar karya saya dengan arahan dari komisipembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Januari 2016
Dendi Hidayatullah
RINGKASAN
DENDI HIDAYATULLAH. Efikasi Vaksin Ulang Streptococcus agalactiae pada Induk Ikan Nila terhadap Imunitas Maternal untuk Pencegahan Streptococcosis. Dibimbing oleh SUKENDA dan SRI NURYATI.
Bakteri S. agalactiae merupakan patogen utama yang menyerang ikan nila mulai fase benih hingga dewasa. Fase benih merupakan fase rawan disebabkan banyak terjadi kematian akibat belum berkembangnya sistem imun dengan sempurna. Rekayasa transfer imunitas maternal melalui vaksinasi induk ikan merupakan teknik alternatif dalam mengantisipasi tingginya angka kematian benih. Pemberian vaksin S. agalactiae di induk ikan nila pada pemijahan pertama mampu meningkatkan imunitas induk maupun benih dan memproteksi benih dari infeksi S. agalactiae. Namun, transfer imunitas maternal pada pemijahan kedua dari induk yang telah divaksin mengalami penurunan. Hal tersebut disebabkan karena dalam jangka waktu tertentu keberadaan antibodi dalam tubuh dan proteksinya akan semakin menurun. Tujuan penelitian ini yaitu untuk menguji efikasi vaksin ulang S. agalactiae di induk ikan nila pada pemijahan kedua terhadap imunitas maternal untuk pencegahan streptococcosis.
Vaksin dibuat dengan cara menginaktivasi bakteri S. agalactiae
menggunakan formalin sebanyak 3% (v/v). Vaksin yang digunakan adalah gabungan sediaan sel utuh dan produk ekstraselular (ECP) S. agalactiae. Vaksin gabungan diinjeksi sebanyak 0,4 mL/kg induk ikan dengan perbandingan 50:50% (v/v) dari dosis penyuntikan, sedangkan kontrol diinjeksi dengan phosphate buffered saline (PBS). Vaksin diberikan ke induk ikan pada fase tingkat kematangan gonad dua (TKG 2). Penelitian terdiri dari tiga perlakuan dan tiga ulangan yaitu induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Uji tantang benih dari setiap induk perlakuan dilakukan melalui perendaman S. agalactiae 107 CFU/mL selama 30 menit pada umur benih 5, 10, 15, dan 20 hari setelah menetas. Parameter yang diamati pada induk ikan meliputi level antibodi, aktivitas lisosim, gambaran darah, dan fekunditas telur. Parameter yang diamati pada telur dan benih meliputi daya tetas telur, level antibodi, aktivitas lisosim, mortalitas benih setelah uji tantang, relative percent survival (RPS) dan perubahan patologi anatomi mikroskopis jaringan otak benih setelah infeksi S. agalaciae.
A tidak berbeda signifikan (P>0,05) dengan benih dari induk K mulai hari ke-10 hingga hari ke-20.
Fekunditas telur tidak berbeda signifikan (P>0,05) pada semua perlakuan. Namun, terdapat perbedaan yang signifikan terhadap daya tetas telur. Daya tetas telur dari perlakuan induk B (94,52%) signifikan lebih tinggi (P<0,05) dari pada perlakuan induk lainnya. Tingkat mortalitas benih perlakuan induk B signifikan lebih rendah (P<0,05) dari perlakuan lainnya hingga hari ke-20. Namun, mortalitas benih perlakuan induk B tidak berbeda signifikan (P>0,05) dengan perlakuan benih dari induk A pada hari ke-5. Nilai RPS benih dari perlakuan induk B tidak berbeda signifikan (P>0,05) dengan perlakuan benih dari induk A pada hari ke-5 tetapi, signifikan lebih tinggi (P<0,05) pada hari ke-10 hingga hari ke-20. Hasil histologi jaringan otak ikan yang terinfeksi S. agalactiae terlihat mengalami nekrosis dan degenerasi, namun tidak terlihat pada jaringan otak benih ikan normal.
Kesimpulan penelitian yaitu pemberian vaksin ulang di induk ikan nila pada pemijahan kedua dapat menstimulasi imunitas dan transfer imunitas maternal ke anaknya untuk pencegahan streptococcosis.
SUMMARY
DENDI HIDAYATULLAH. Efficacy of Streptococcus agalactiae Re-vaccine on Tilapia Broodstock to Maternal Immunity in Preventing Streptococcosis. Supervised by SUKENDA and SRI NURYATI.
The bacteria S. agalactiae is a main pathogen attacking tilapia during the fry up to adult phase. The fry phase is the most threatened phase with high mortality rate due to under developed immunity system. The manipulation maternal immunity transfer via broodstock vaccination was the best alternative technique to anticipate the high mortality rate in fry. The administration of S. agalactiae
vaccine in tilapia broodstock during the first spawning has been proven to increase the immunity of broodstock and fry against infection of S. agalactiae.
However the maternal immunity transfer in the second spawning has decreased. In addition, the presence and potection of antibodies will continue to decrease over
time. The purpose of the research was conducted to assess the efficacy of
S. agalactiae re-vaccine in tilapia broodstock before second spawning on the maternal immunity transfer to the fry in preventing streptococcosis.
The vaccine was prepared using formalin-killed whole cell and extracellular products (ECP) of S. agalactiae. Fish were vaccinated with mixed whole cell and ECP vaccine. The vaccine was administered by injection of 0.4 mL/kg fish in a 50:50% (v/v), and the control was injected by phosphate buffered saline (PBS). Vaccinated broodstock at gonad developmental stage two. The research consisted of three treatments with three replications, which were broodstock with injected PBS (K), broodstocks injected once vaccine (A), and broodstocks injected twice vaccine with interval one month (B). Challenge test for fry produced on every broodstock were performed by immersion of 107 CFU/mL S. agalactiae for 30 minutes at fry ages 5, 10, 15, and 20 day after hatching. The parameters observed in broodstock were level of antibodies, lysozyme activity, blood profile, and fecundity rate of eggs. Parameters observed in eggs and fry were hatching of eggs, level of antibodies, lysozyme activity, mortality rate of fry after challenge test, relative percent survival (RPS), and anatomic microscopic pathology changes in brain tissue of fry after infection with S. agalaciae.
The results showed total erythrocytes, hemoglobin and hematocrit in broodstock were not significantly different (P>0.05) between all treatments. However, there were significantly different (P<0.05) in total leukocytes and phagocytic activity. The total leukocytes and phagocytic activity from broodstock B treatment were significantly higher (P<0.05) than the other broodstock. Antibody level of broodstock, eggs, and fry from B treatment (OD: 0.20) were significantly higher (P<0.05) than the other treatments. Lisozyme activity in broodstock, eggs, and fry from B treatment were significantly higher (P<0.05) than the other broodstock. The lisozyme activity of fry from broodstock A and B decreased at day 10 and than rise again until day 20. The lisozyme activity of fry from broodstock A and K were not significantly different (P<0.05) from 10th until 20th day.
hatching rate. Hatching rate from broodstock B was significantly higher (P<0.05) than the other broodstock treatments. Mortality rate of fry from broodstock B was significantly lower (P<0.05) than the other treatment until 20th day. However, mortality rate of fry from broodstock B was not significantly different (P>0.05) with fry from broodstock A at 5th day. Relative percent survival of fry from broodstock B was not significanty different (P>0.05) with fry from broodstock A at 5th day but, significantly higher (P<0.05) at 10th until 20th day. There was microscopic changes in brain of fry after S. agalactiae infection. The brain tissue which S. agalactiae infection showed necrosis and degeneration, but not shown in normal fish brain.
The conclusion of this research showed re-vaccine in broodstock on second spawning can stimulate immunity and maternal immunity transfer to fry for streptococcosis prevention.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Akuakultur
EFIKASI VAKSIN ULANG
Streptococcus agalactiae
PADA
INDUK IKAN NILA TERHADAP IMUNITAS MATERNAL
UNTUK PENCEGAHAN STREPTOCOCCOSIS
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
Judul Tesis : Efikasi Vaksin Ulang Streptococcus agalactiae pada Induk Ikan Nila terhadap Imunitas Maternal untuk Pencegahan Streptococcosis Nama : Dendi Hidayatullah
NIM : C151130161
Disetujui oleh Komisi Pembimbing
Dr Ir Sukenda, MSc Ketua
Dr Sri Nuryati, SPi MSi Anggota
Diketahui oleh
Ketua Program Studi Ilmu Akuakultur
Dr Ir Widanarni, MSi
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MSc Agr
Tanggal Ujian: 14 Desember 2015
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wata’ala atas segala karunia-Nya sehingga tesis ini berhasil diselesaikan. Penelitian dilaksanakan pada bulan Januari hingga Maret 2015 dengan judul Efikasi Vaksin Ulang Streptococcus agalactiae pada Induk Ikan Nila terhadap Imunitas Maternal untuk Pencegahan Streptococcosis.
Pada kesempatan ini, penulis mengucapkan terima kasih kepada semua pihak yang telah membantu dalam penulisan tesis ini, terutama kepada Dr Ir Sukenda, MSc dan Dr Sri Nuryati, SPi MSi selaku ketua dan anggota komisi pembimbing tesis serta Dr Dinamella Wahjuningrum, SSi MSi selaku wakil dari program studi Ilmu Akuakultur dan Dr Ir Kukuh Nirmala, MSc selaku dosen penguji luar komisi. Ungkapan terima kasih juga disampaikan kepada ayah, ibu, serta seluruh keluarga atas segala doa dan kasih sayangnya. Penulis sampaikan pula terima kasih dan penghargaan kepada Pak Ranta, Pak Rahman, teman-teman BDP45 (Wahyu, Fajar, Ardina, Nurina, Tira, Tiara, Yoseph, Asep, Erriza), teman-teman Ilmu Akuakultur 2013, teman-teman-teman-teman laboratorium kesehatan organisme akuatik 2013 (Ka Ezy, Ka Dwi, Ka Vika, Windu, Abung, Asep, Ari, Lukman, Sekar, Ike, Amal, Bu Aty, Nurin), serta adik-adik tingkat (Yanti, Rudi, Ardana, Dhana, Kiki, Zani, Adel, Iqbal, Mita, Syifa, Yuri, May, Hana, Berman, Arbi) yang terus memberikan semangat, bantuan, dukungan, dan doanya kepada penulis dalam penyusunan tesis. Terimakasih juga disampaikan kepada DIKTI yang telah memberikan Beasiswa Pendidikan Pascasarjana Dalam Negeri (BPPDN) tahun 2013.
Demikian tesis ini disusun, semoga bermanfaat dan dapat berkontribusi terhadap pengembangan akuakultur di masa depan.
Bogor, Januari 2016
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vi
DAFTAR LAMPIRAN vii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 2
Manfaat Penelitian 2
2 METODE 3
Materi Uji 3
Rancangan Percobaan 3
Prosedur Penelitian 3
Parameter Penelitian 5
Analisis Data 8
3 HASIL DAN PEMBAHASAN 8
Hasil 8
Pembahasan 13
4 SIMPULAN DAN SARAN 17
Simpulan 17
Saran 17
DAFTAR PUSTAKA 17
LAMPIRAN 22
DAFTAR TABEL
1 Perlakuan pengujian efikasi vaksin ulang S. agalactiae di induk ikan nila
pada pemijahan kedua 3
2 Gambaran darah induk ikan nila setelah vaksinasi pada perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B): total eritrosit, total leukosit, hemoglobin (Hb), hematokrit (Hc), dan aktivitas fagositik (AF) 9 3 Fekunditas dan daya tetas telur ikan nila pada pemijahan kedua dari
perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). 12 4 Tingkat mortalitas setelah infeksi S. agalactiae dan relative percent survival
benih ikan nila pada pemijahan kedua dari perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua
kali dengan selang waktu satu bulan (B) 13
DAFTAR GAMBAR
1 Level antibodi induk ikan nila setelah vaksinasi pada perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Huruf yang berbeda di setiap bar menunjukkan pengaruh perlakuan yang berbeda nyata
(Tukey’s P<0,05) 9
2 Level antibodi telur ikan nila pada pemijahan kedua dari perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Huruf yang berbeda di setiap bar menunjukkan pengaruh perlakuan yang berbeda nyata
(Tukey’s P<0,05) 10
3 Level antibodi benih ikan nila umur benih 5, 10, 15, dan 20 hari setelah menetas pada pemijahan kedua dari perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Huruf yang berbeda di setiap bar pada umur benih yang sama menunjukkan pengaruh perlakuan yang berbeda
nyata (Tukey’s P<0,05) 10
4 Aktivitas lisosim induk ikan nila pada perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). setelah vaksinasi. Huruf yang berbeda di setiap bar menunjukkan pengaruh perlakuan yang berbeda nyata
(Tukey’s P<0,05) 11
5 Aktivitas lisosim telur ikan nila pada pemijahan kedua dari perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Huruf yang berbeda di setiap bar menunjukkan pengaruh perlakuan yang berbeda
6 Aktivitas lisosim benih ikan nila umur benih 5, 10, 15, dan 20 hari setelah menetas pada pemijahan kedua dari perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Huruf yang berbeda di setiap bar pada umur benih yang sama menunjukkan pengaruh perlakuan yang berbeda
nyata (Tukey’s P<0,05) 12
7 Patologi anatomi mikroskopis pada jaringan otak benih ikan nila setelah infeksi S. agalactiae: A) Jaringan otak normal; B) Jaringan otak terserang
S. agalactiae mengalami nekrosis (Ne) dan Degenerasi (De) 13
DAFTAR LAMPIRAN
1 Karakterisasi bakteri S. agalactiae menggunakan KIT API 20 Strep 22 2 Hasil pengukuran bobot molekul protein produk ekstraselular (ECP)
S. agalactiae dengan metode sodium dodecyl sulfate polyacrylamide gel
electrophoresis (SDS-PAGE) 23
3 Histologi jaringan gonad induk ikan nila pada tingkat kematangan gonad
1
PENDAHULUAN
Latar Belakang
Bakteri S. agalactiae merupakan patogen utama di ikan nila. Bakteri tersebut menyebabkan kematian massal pada budidaya ikan nila di beberapa negara produsen ikan nila seperti China, Thailand, Malaysia dan Indonesia (Sheehan et al., 2009). Bakteri ini menimbulkan penyakit streptococcosis mulai fase benih hingga fase dewasa pada ikan nila (Evans et al., 2006; Yang & Li, 2009; Jantrakajorn et al., 2014). Infeksi Streptococcus pada budidaya ikan nila intensif dapat menyebabkan tingkat kematian mencapai 90% populasi dan lebih dari 90% bakteri disebabkan oleh S. agalactiae (Ye et al., 2011; Chen et al.,
2012). Streptococcosis pada ikan yang disebabkan oleh S. agalactiae
menyebabkan septicemia dan meningoencephalitis (Mian et al., 2009). Hardi et al., (2011) melaporkan bahwa ikan yang terserang patogen S. agalactiae memiliki gejala seperti pembengkakan pada mata (exopthalmia), kekeruhan pada mata (opacity), mata memutih (purulens), perubahan pola renang (whirling), dan penjernihan operkulum (Clear operculum).
Aspek yang berpengaruh terhadap keberhasilan budidaya ikan nila salah satunya adalah kualitas benih. Hal ini erat hubungannya dengan kualitas induk ikan. Menurut Swain dan Nayak (2009), kesehatan dan status imun induk ikan sangat penting bukan hanya pada saat pemijahan tetapi juga untuk kesehatan benih yang dihasilkan. Adanya imunitas maternal yang ditransfer oleh induk menjadi esensial pada fase awal pertumbuhan benih. Hal ini penting karena pada awal pertumbuhan, kemampuan embrio dan benih ikan masih belum dapat mengembangkan respon imun dengan baik (Magnadottir et al., 2005; Zapata et al.,
2006).
Vaksinasi ikan merupakan salah satu upaya pencegahan yang dapat dilakukan untuk mengatasi masalah penyakit dengan cara menginduksi kekebalan spesifik (Evans et al., 2005; Sommerset et al., 2005; Muiswinkel, 2008; Secombes, 2011; Gudding et al., 2013; Yi et al., 2014). Beberapa penelitian menunjukkan bahwa pemberian vaksin dari sediaan antigen sel utuh dan produk ekstraselular S. agalactiae mampu merangsang terbentuknya antibodi spesifik dan memproteksi ikan nila dari infeksi S. aglactiae (Hardi et al., 2013; Amrullah et al., 2014; Dwinanti et al., 2014). Vaksin gabungan antara sel utuh dan produk ekstraselular S. agalactiae telah diketahui bersifat lebih imunogenik dibandingkan jika diberikan secara tunggal (Hardi et al., 2013). Akan tetapi vaksinasi pada stadia benih melalui perendaman belum menunjukkan proteksi yang optimal dengan nilai kelangsungan hidup relatif yang masih rendah (Taukhid et al., 2012; Sukenda et al., 2014). Padahal meningkatkan kekebalan pada ikan sedini mungkin adalah pendekatan yang lebih baik karena memberi proteksi terhadap serangan penyakit sedini mungkin (Bowden et al., 2005; Magnadottir et al., 2005).
Rekayasa transfer kekebalan maternal pada induk ikan merupakan upaya dalam mengantisipasi tingginya angka kematian anak ikan pada umur kurang dari satu bulan (Swain et al., 2006; Wang et al., 2012; Mingming et al., 2014). Salah satu alternatif yang dapat dilakukan yaitu melalui vaksinasi induk (Zang et al.,
2
dan lisosim induk ikan ditransfer dari induk ke anaknya dan memberikan proteksi yang baik (Nur et al., 2004; Hanif et al., 2004; Picchietti et al., 2004; Swain et al.,
2006, Wang et al., 2012).
Vaksinasi induk ikan nila sebelum memijah telah menunjukkan transfer imunitas maternal yang baik kepada benih di pemijahan pertama (Nur et al., 2004; Sukenda et al., 2015a). Namun transfer imunitas maternal pada pemijahan kedua dari induk yang telah divaksin ke anaknya mengalami penurunan. Hal ini disebakan karena dalam jangka waktu tertentu keberadaan antibodi dalam tubuh ikan dan tingkat proteksinya semakin menurun (Pasnik et al., 2005; Sukenda et al., 2015b). Oleh karena itu perlu dilakukan penelitian terkait pemberian vaksin ulang di induk ikan nila pada pemijahan kedua dalam upaya meningkatkan transfer imunitas maternal induk ke anak ikan.
Perumusan Masalah
Bakteri S. agalactiae diketahui sebagai patogen utama penyebab kematian pada ikan nila. Bakteri S. agalactiae menimbulkan penyakit streptococcosis mulai fase benih hingga fase dewasa. Rekayasa transfer kekebalan maternal pada induk ikan merupakan teknik alternatif dalam upaya mengantisipasi tingginya angka kematian benih ikan nila pada umur kurang dari satu bulan. Salah satu cara yang dapat dilakukan adalah melalui vaksinasi induk. Vaksinasi induk dengan gabungan antara sel utuh dan produk ekstraselular S. agalactiae dipemijahan pertama mampu meningkatkan imunitas spesifik dan non spesifik induk maupun benih yang dihasilkan terhadap infeksi S. agalactiae. Namun, dalam jangka waktu tertentu keberadaan antibodi dalam tubuh ikan dan tingkat proteksinya akan semakin menurun dan berdampak pada transfer imunitas maternal pada pemijahan selanjutnya. Oleh karena itu perlu dilakukan penelitian terkait pemberian vaksin ulang pada induk nila pada pemijahan kedua.
Tujuan Penelitian
Penelitian ini bertujuan untuk menguji efikasi vaksin ulang S. agalactiae di induk ikan nila pada pemijahan kedua terhadap imunitas maternal yang ditransfer ke anaknya untuk pencegahan streptococcosis.
Manfaat Penelitian
3
2
METODE
Materi Uji
Induk ikan nila yang digunakan adalah ikan nila strain Nirwana yang diperoleh dari Balai Pengembangan Benih Ikan Air Tawar, Wanayasa, Jawa Barat. Bobot induk ikan nila yang digunakan berukuran 200-300 gram. Induk ikan dipisahkan antara jantan dan betina. Induk ikan dipelihara dalam bak berukuran 3×2×1 m3 pada temperatur air 24-30 °C; pH 6,58-7,76; OD 4,5-6,7 ppm. Induk ikan diberi pakan dengan kadar protein 35-40% sebanyak dua kali dalam sehari secara at satiation dan dilakukan pergantian air setiap tiga hari sekali.
Bakteri S. agalactiae diperoleh dari koleksi Laboratorium Kesehatan Ikan, Instalasi Penelitian dan Pengembangan Pengendalian Penyakit Ikan BPPBAT Bogor. Bakteri S. agalactiae yang digunakan dalam penelitian dilakukan pemulihan virulensi dengan cara difasase. Bakteri S. agalactiae hasil fasase kemudian dikarakterisasi menggunakan Kit API 20 Strept (Lampiran 1). Setelah bakteri dikarakterisasi ulang kemudian siap digunakan untuk pembuatan vaksin dan uji tantang.
Rancangan Percobaan
Penelitian ini dilakukan dengan metode eksperimen menggunakan rancangan acak lengkap (RAL) dengan tiga perlakuan dan tiga ulangan. Evaluasi transfer imunitas maternal induk ke anak ikan dilakukan pada pemijahan kedua Rancangan penelitian disajikan pada Tabel 1.
Tabel 1 Perlakuan pengujian efikasi vaksin ulang S. agalactiae di induk ikan nila pada pemijahan kedua
Perlakuan Keterangan
K Induk diinjeksi PBS pada pemijahan kedua
A Induk diinjeksi vaksin satu kali pada pemijahan kedua
B Induk diinjeksi vaksin dua kali dengan selang waktu satu bulan pada pemijahan kedua
Prosedur Penelitian
Penyediaan Vaksin Uji
Vaksin yang digunakan adalah vaksin gabungan antara sel utuh dan produk ekstraselular (ECP) S. agalactiae. Metode pembuatan vaksin mengacu pada Hardi
et al., (2013). Preparasi masing-masing vaksin dijelaskan sebagai berikut.
Preparasi vaksin sel utuh: Bakteri dikultur dalam media brain heart infusion
4
Preparasi vaksin produk ekstraselular (ECP): Bakteri dikultur dalam media
brain heart infusion (BHI, BD BactoTM) cair dengan masa inkubasi 72 jam pada suhu 28-30 °C. Media yang telah ditumbuhi bakteri ditambahkan 3% neutral buffered formalin dan diinkubasi selama 24 jam. Kemudian suspensi disentrifugasi 7.000 rpm selama 30 menit pada suhu 4 °C. Vaksin ECP diperoleh dengan menyaring supernatan dengan filter saring 0,22 µm. Protein di dalam supernatan kemudian dianalisis dengan sodium dodecyl sulfate polyacrylamide gel electrophoresis (SDS-PAGE) menggunakan metode pewarnaan Coomassie Brilliant Blue R-250 (Bio Rad) untuk mengetahui bobot molekul protein yang terkandung di dalamnya. Hasil analisis SDS-PAGE menunjukkan ECP yang didapat memiliki kandungan beberapa protein sebesar 51,8 kDa; 55,8 kDa; dan 62,3 kDa (Lampiran 2). Vaksin ECP kemudian diuji keamanannya dengan cara menyuntik vaksin ECP pada lima ekor ikan nila berukuran 10 gram sebanyak 0,1 mL/ekor. Jika hingga jam ke-72 tidak muncul kematian dan gejala streptococcosis maka vaksin ECP aman untuk digunakan.
Vaksinasi dan Pemijahan Induk Ikan Nila
Induk yang akan divaksinasi diambil sampel untuk dilihat tingkat kematangan gonadnya dengan metode kanulasi yaitu memasukkan selang kateter berdiameter 1 mm ke dalam lubang genital sedalam 2 – 4 cm lalu dihisap dan dicabut secara perlahan-lahan. Telur pada tingkat kematangan gonad dua selanjutnya dilakukan histologi jaringan (Lampiran 3). Vaksinasi induk diberikan pada fase tingkat kematangan gonad dua (TKG2). Vaksin diberikan melalui injeksi intraperitoneal. Vaksin diberikan ke induk ikan nila dengan dosis 0,4 mL/kg ikan. Vaksin sel utuh diberikan dengan konsentrasi 109 CFU/mL. Kombinasi vaksin sel utuh dan ECP adalah 50:50% (v/v) dari dosis penyuntikan, sedangkan kontrol diinjeksi dengan PBS. Sebelum divaksinasi induk ikan dipingsankan terlebih dahulu dengan bius. Interval waktu vaksinasi pertama dan kedua yaitu satu bulan. Pemijahan dilakukan dalam hapa berukuran 1×1×1 m3 dengan perbandingan satu jantan dan tiga betina. Pemijahan dilakukan secara alami. Jarak antara waktu vaksinasi dan pemijahan yaitu tiga minggu.
Penetasan Telur dan Pemeliharaan Benih
Ikan yang telah memijah dihitung fekunditas dan daya tetas telurnya. Telur yang telah dibuahi diperoleh dengan membuka mulut induk betina. Telur ikan nila kemudian ditetaskan dalam wadah akuarium berukuran 60×35×30 cm3 dan diberi aerasi yang cukup. Setelah telur menetas benih diberi pakan alami berupa cacing sutra secara ad libitum yang dimulai pada hari ke-7 (setelah kuning telur habis). Suhu pemeliharaan benih dijaga pada temperatur 28-29 °C; pH 6,85-7,68; dan OD 4,1-6,1 ppm. Pergantian air dilakukan setiap hari.
Uji Tantang Benih
5 Preparasi Sampel Serum dari Induk, Telur, dan Benih
Induk dan benih ikan dipingsankan dahulu menggunakan pembius ikan. Darah dari induk dikumpulkan melalui pembuluh vena dipangkal ekor. Darah disimpan dalam suhu ruang selama 1 jam dan selanjutnya disimpan semalaman pada suhu 4 °C. Serum dikumpulkan dengan cara disentrifugasi 5000 rpm selama 10 menit pada suhu 4 °C. Serum kemudian dipisahkan ke dalam tabung mikro dan disimpan pada suhu -20 °C untuk digunakan pada pengujian antibodi dan lisosim.
Telur dikumpulkan sesaat setelah induk memijah, sedangkan sampel benih diambil pada hari ke-5, ke-10, ke-15, dan ke-20 setelah menetas. Telur dan benih yang terkumpul dihomogenkan dalam larutan PBS-T (PBS+0,05% Tween-20) dengan perbandingan 1:4. Selanjutnya disentrifugasi 5000 rpm selama 10 menit pada suhu 4 °C. Supernatan kemudian dipisahkan ke dalam tabung mikro dan disimpan pada suhu -20 °C untuk digunakan pada pengujian antibodi dan lisosim.
Parameter Penelitian
Parameter yang diamati pada induk yaitu level antibodi, aktivitas lisosim, gambaran darah (total eritrosit, total leukosit, hemoglobin, hematokrit, aktivitas fagositik) dan fekunditas telur. Parameter yang diamati pada telur dan benih meliputi daya tetas telur, level antibodi, aktivitas lisosim, mortalitas benih setelah uji tantang, relative percent survival (RPS) benih, dan patologi anatomi mikroskopis jaringan otak benih ikan.
Total Eritrosit
Total eritrosit induk ikan diamati satu minggu setelah vaksinasi. Total eritrosit dihitung menggunakan metode Blaxhall dan Daisley (1973) yaitu darah sampel dihisap dengan pipet yang berisi bulir pengaduk warna merah sampai skala 0,5. Selanjutnya ditambah larutan Hayem’s sampai skala 101. Darah dalam pipet diaduk dengan cara menggoyangkan pipet membentuk angka delapan selama 3-5 menit sehingga darah tercampur rata. Dua tetes pertama larutan darah dalam pipet tersebut dibuang, selanjutnya larutan darah tersebut diteteskan di atas hemasitometer yang telah diletakkan gelas penutup di atasnya. Berikut ini adalah rumus perhitungan total eritrosit :
∑Eritrosit (sel/mm3) = ∑sel terhitung ×
× faktor pengenceran
Total Leukosit
Total leukosit induk ikan diamati satu minggu setelah vaksinasi. Total Leukosit dihitung menggunakan metode Blaxhall dan Daisley (1973). Darah dihisap menggunakan pipet hemasitometer berbulir merah sampai skala 0,5 lalu diencerkan dengan larutan Turk’s sampai skala 11. Kedua ujung ditutup sejajar kemudian digoyangkan membentuk angka delapan selama 3-5 menit. Setelah itu, larutan pada bagian ujung pipet yang tidak teraduk dibuang sebanyak dua tetes. Tetesan berikutnya dimasukkan ke dalam hemasitometer yang telah dilengkapi dengan kaca penutup. Berikut ini adalah rumus perhitungan total leukosit:
∑Leukosit (sel/mm3) = ∑sel terhitung ×
6
Kadar Hemoglobin
Kadar hemoglobin induk ikan diamati satu minggu setelah vaksinasi. Kadar hemoglobin diukur menurut metode Sahli (Wedemeyer dan Yasutake, 1977) yaitu dengan mengisi tabung sahlinometer dengan larutan HCl 0,1 N sampai garis skala 10 merah, kemudian ditempatkan diantara 2 tabung warna standar. Darah ikan dari tabung mikro diambil dengan pipet Sahli sebanyak 0,02 mL dan dimasukkan ke tabung Sahli, kemudian didiamkan selama 3 menit. Selanjutnya ditambahkan akuades dengan pipet tetes sedikit demi sedikit dan diaduk sampai berubah warna tepat sama dengan warna standar. Kadar hemoglobin dinyatakan dalam g/dL pada skala kuning.
Hematokrit
Hematokrit diamati satu minggu setelah vaksinasi induk ikan. Hematokrit diukur menggunakan metode Anderson dan Siwicki (1993) yaitu darah dihisap dengan tabung kapiler (mikrohematokrit) hingga ¾ bagian tabung, lalu ujung tabung ditutup dengan crytoceal. Setelah itu, tabung mikrohematokrit yang berisi darah disentrifugasi pada kecepatan 3000 rpm selama 5 menit. Perhitungan hematokrit dilakukan dengan cara membandingkan panjang endapan darah (a) terhadap panjang total seluruh darah (b). Berikut ini adalah rumus perhitungan kadar hematokrit:
Hematokrit (%) = × 100 Aktivitas Fagositik
Aktivitas fagositik induk ikan diamati satu minggu setelah vaksinasi. Aktivitas fagositik dihitung berdasarkan metode Anderson dan Siwicki (1993). Darah sebanyak 50µL dimasukkan ke dalam tabung mikro, ditambahkan 50µL suspensi bakteri Staphylococcus aureus dalam PBS, dihomogenkan dan diinkubasi pada suhu 28 °C selama 20 menit. Selanjutnya dari campuran tersebut diambil sebanyak 5 µL untuk dibuat preparat ulas. Preparat ini difiksasi dengan metanol selama 5 menit dan dikeringkan, kemudian direndam dalam larutan giemsa selama 15 menit. Preparat lalu dicuci dalam air mengalir dan dikeringkan. Pengamatan dilakukan menggunakan mikroskop. Aktivitas fagositik didasarkan pada persentase dari 100 sel fagosit yang menunjukkan aktivitas fagositik. Berikut adalah rumus untuk menghitung aktivitas fagositik :
Aktivitas Fagositik (%) =
× 100
Level Antibodi
Level antibodi diamati pada induk, telur, dan benih. Level antibodi induk ikan diamati dua minggu setelah vaksinasi, sedangkan level antibodi benih diamati pada hari ke-5, ke-10, ke-15, dan ke-20 setelah menetas. Pengukuran level antibodi dilakukan dengan menggunakan metode indirect enzyme-linked imunosorbent assay (ELISA) (Shelby et al., 2001). Sebanyak 100 μL antigen yang telah diencerkan 1:1 pada PBS (pH 7,2) dimasukkan ke dalam sumur
7 inkubasi 25 °C selama 1 jam, kemudian dicuci dengan PBS-T. Sampel serum induk nila diencerkan 1:50 pada PBS-T sedangkan untuk telur dan benih diencerkan 1:16 pada PBS-T, ditambahkan ke dalam microtiter plates sebanyak 100 μL dengan 3 kali ulangan. Microtiter plates diinkubasi 25 °C selama 1 jam kemudian dicuci dengan PBS-T. Imunoglobulin anti-nila rantai panjang untuk poliklonal antibodi spesifik diencerkan 1:200 pada PBS-T dan 100 μL dan ditambahkan ke setiap sumur microtiter plates. Microtiter plates diinkubasi pada suhu 25 °C selama 1 jam kemudian dicuci menggunakan PBS-T. Tambahkan 100 μL peroxidase-conjugated goat anti rabbit (Sigma) yang diencerkan menjadi 1:5000 pada PBS-T ke setiap sumur microtiter plates. Microtiter plates diinkubasi pada suhu 25 °C selama 1 jam kemudian dicuci dengan PBS-T. Tambahkan pada setiap sumur microtiter plates 100 μL One-Step Ultra TMB-ELISA (Sigma) (TMB 5 mg + 10 µL H2O2 38% dalam 5 mL asetat buffer pH 5). Untuk blanko tambahkan 50 μL H2SO4 3 M ke dalam sumur pertama dari microtiter plates dan diamkan selama 20 menit kemudian tambahkan 100 μL substrat TMB-ELISA. Setelah 20 menit, reaksi ELISA dihentikan dengan menambahkan 50 μL H2SO4 3 M, dan lakukan pembacaan optical density (OD) pada panjang gelombang 405 nm dengan Microplate Reader (Kayto RT-2100C).
Aktivitas Lisosim
Aktivitas lisosim diamati pada induk, telur, dan benih. Lisosim induk diamati satu minggu setelah vaksinasi, sedangkan lisosim benih diamati pada hari ke-5, ke-10, ke-15, dan ke-20 setelah menetas. Aktivitas lisosim diukur dengan metode yang digunakan oleh Hanif et al., (2004). Sampel (100 μL) ditambahkan suspensi cair bakteri Micrococcus lysodeikticus (Sigma) (sebanyak 100 μL (0,4 mg/mL dalam 0,1 M Phosphate buffered saline pH 6,2 pada suhu 25 oC. Dilakukan dua kali pembacaan adsorpsi pada panjang gelombang 450 nm dengan
Microplate Reader (Kayto RT-2100C) selama 30 detik pencampuran dan 30 menit pencampuran. Unit aktivitas lisosim akan dibatasi sejumlah enzim yang menyebabkan penurunan absorbans 0,001/menit. Berikut merupakan rumus perhitungan lisosim:
Nilai fekunditas dilakukan dengan cara menghitung jumlah telur yang dihasilkan dari satu ekor induk ikan nila. Selanjutnya daya tetas telur dihitung Tingkat mortalitas benih dihitung menggunakan rumus sebagai berikut:
Mortalitas (%) =
8
Relative Percent Survival (RPS)
Relative percent survival benih dihitung untuk mengetahui efektivitas transfer kekebalan maternal dari induk yang divaksin setelah uji tantang dengan
S. agalactiae. Nilai RPS dihitung pada akhir uji tantang. Berikut merupakan rumus perhitungan RPS:
RPS (%) = (1 –
) × 100
Patologi Anatomi Mikroskopis
Pengamatan patologi anatomi mikroskopis yang diamati yaitu perubahan jaringan otak benih ikan nila setelah uji tantang S. agalactiae. Pengamatan dilakukan dengan membuat preparat histologi dari jaringan otak benih ikan normal dan terserang S. agalactiae. Organ otak diambil dan difiksasi dalam larutan neutral buffered formalin 10% selama 24 jam yang selanjutnya dipindahkan dalam alkohol 70% sampai dilakukan proses pembuatan preparat histologi dengan menggunakan pewarnaan hematoxylin dan eosin (Geten et al., 2009).
Analisis Data
Penelitian akan dilakukan menggunakan rancangan acak lengkap (RAL) dengan tiga perlakuan dan tiga ulangan. Data yang diperoleh ditabulasi dengan program MS. Office Exel 2007. Data dianalisis menggunakan ANOVA melalui program Minitab versi 16 dengan tingkat selang kepercayaan 95%, jika signifikan maka akan diuji lanjut dengan uji lanjut Tukey’s. Histologi jaringan otak benih ikan nila di analisis secara deskriptif.
3
HASIL DAN PEMBAHASAN
Hasil
Gambaran Darah Induk Ikan Nila
9 Tabel 2 Gambaran darah induk ikan nila setelah vaksinasi pada perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B): total eritrosit, total leukosit, hemoglobin (Hb), hematokrit (Hc), dan aktivitas fagositik (AF)
Perlakuan ∑ Eritrosit (×106 sel/mm3)
∑ Leukosit
(×104 sel/mm3) Hb (g/dL) Hc (%) AF (%) K 2,03±0,27a 7,97±0,86c 7,33±0,81a 30,76±3,45a 24,51±2,02c A 2,11±0,19a 12,03±0,57b 7,47±0,70a 31,43±1,82a 49,75±3,24b B 2,35±0,08a 14,27±0,55a 8,33±0,40a 32,78±1,26a 69,02±1,53a
Standar
Keterangan: Huruf superskrip yang berbeda pada kolom yang sama menunjukkan pengaruh perlakuan yang berbeda nyata (Uji Tukey’s; P<0,05).
Level Antibodi Induk Ikan Nila
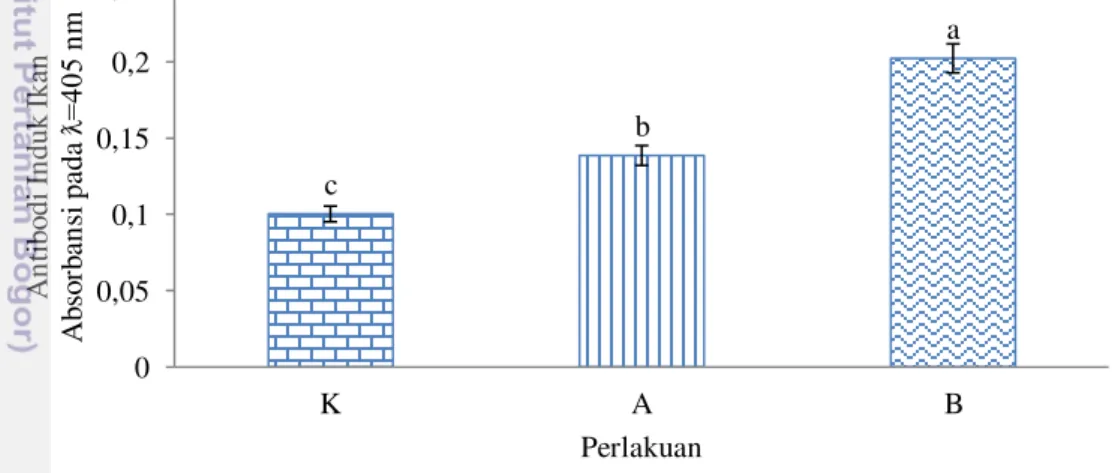
Level antibodi induk ikan nila setelah vaksinasi berbeda signifikan pada semua perlakuan. Level antibodi perlakuan induk B (OD: 0,20) signifikan lebih tinggi dari pada perlakuan lainnya. Level antibodi perlakuan induk A (OD: 0,14) signifikan lebih tinggi (P<0,05) dari pada perlakuan induk K (OD: 0,10). Level antibodi induk ikan nila setelah vaksinasi disajikan pada Gambar 1.
Gambar 1 Level antibodi induk ikan nila setelah vaksinasi pada perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Huruf yang berbeda di setiap bar menunjukkan pengaruh perlakuan yang berbeda nyata (Tukey’s P<0,05)
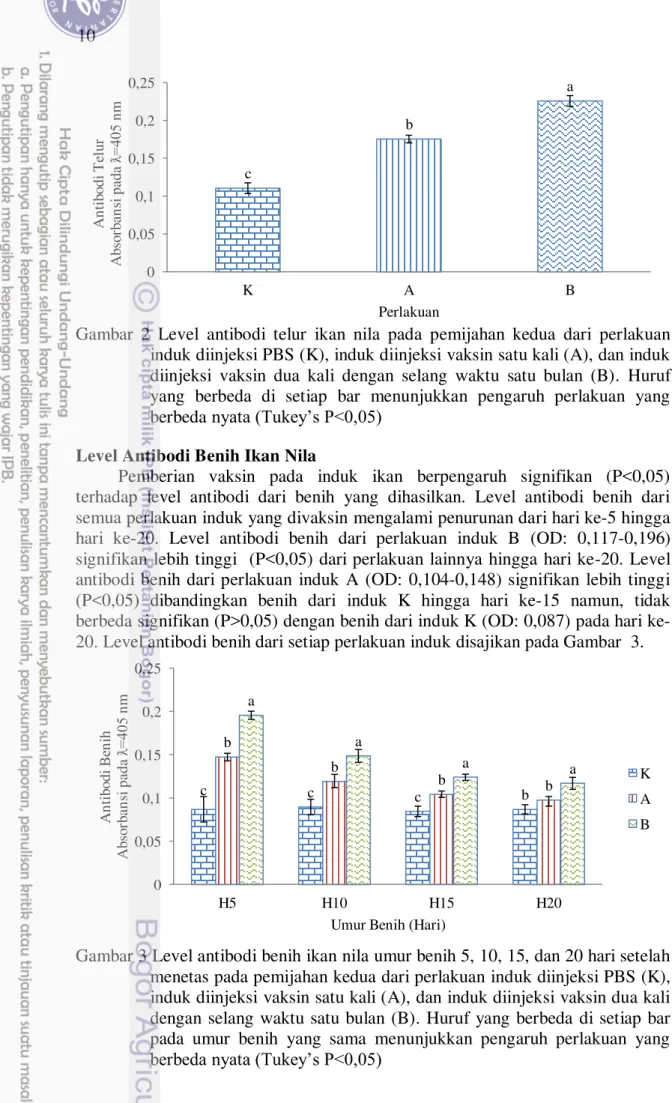
Level Antibodi Telur Ikan Nila
10
Gambar 2 Level antibodi telur ikan nila pada pemijahan kedua dari perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Huruf yang berbeda di setiap bar menunjukkan pengaruh perlakuan yang berbeda nyata (Tukey’s P<0,05)
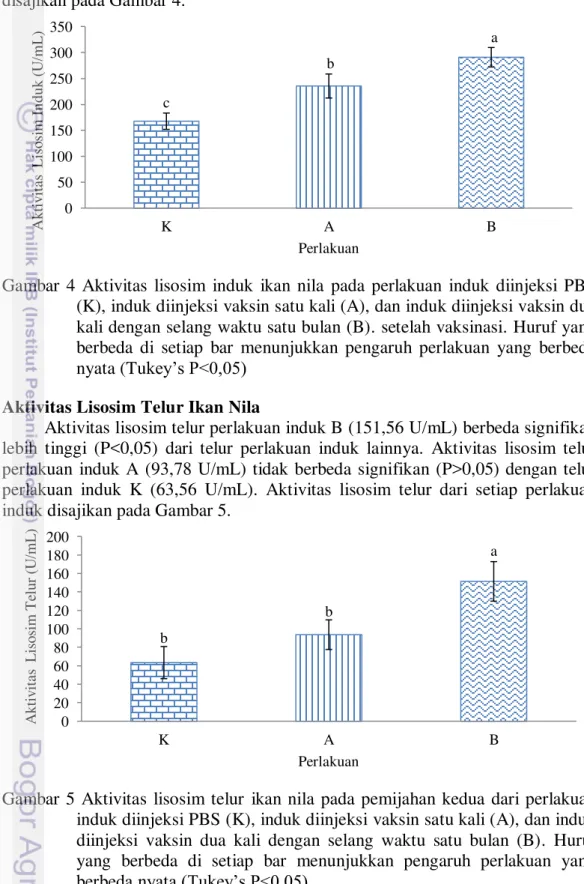
Level Antibodi Benih Ikan Nila
Pemberian vaksin pada induk ikan berpengaruh signifikan (P<0,05) terhadap level antibodi dari benih yang dihasilkan. Level antibodi benih dari semua perlakuan induk yang divaksin mengalami penurunan dari hari ke-5 hingga hari ke-20. Level antibodi benih dari perlakuan induk B (OD: 0,117-0,196) signifikan lebih tinggi (P<0,05) dari perlakuan lainnya hingga hari ke-20. Level antibodi benih dari perlakuan induk A (OD: 0,104-0,148) signifikan lebih tinggi (P<0,05) dibandingkan benih dari induk K hingga hari ke-15 namun, tidak berbeda signifikan (P>0,05) dengan benih dari induk K (OD: 0,087) pada hari ke-20. Level antibodi benih dari setiap perlakuan induk disajikan pada Gambar 3.
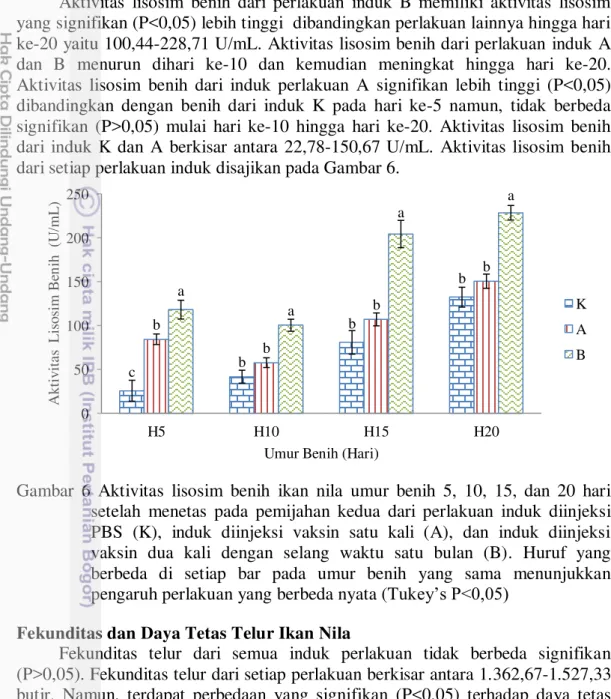
11 Aktivitas Lisosim Induk Ikan Nila
Aktivitas lisosim perlakuan induk B (291,11 U/mL) signifikan lebih tinggi (P<0,05) dari perlakuan induk lainnya. Aktivitas lisosim perlakuan induk A (235,67 U/mL) signifikan lebih tinggi (P<0,05) dibandingkan dengan perlakuan induk K (167,78 U/mL). Aktivitas lisosim induk ikan nila setelah vaksinasi disajikan pada Gambar 4.
Gambar 4 Aktivitas lisosim induk ikan nila pada perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). setelah vaksinasi. Huruf yang berbeda di setiap bar menunjukkan pengaruh perlakuan yang berbeda nyata (Tukey’s P<0,05)
Aktivitas Lisosim Telur Ikan Nila
Aktivitas lisosim telur perlakuan induk B (151,56 U/mL) berbeda signifikan lebih tinggi (P<0,05) dari telur perlakuan induk lainnya. Aktivitas lisosim telur perlakuan induk A (93,78 U/mL) tidak berbeda signifikan (P>0,05) dengan telur perlakuan induk K (63,56 U/mL). Aktivitas lisosim telur dari setiap perlakuan induk disajikan pada Gambar 5.
Gambar 5 Aktivitas lisosim telur ikan nila pada pemijahan kedua dari perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Huruf yang berbeda di setiap bar menunjukkan pengaruh perlakuan yang berbeda nyata (Tukey’s P<0,05)
12
Aktivitas Lisosim Benih Ikan Nila
Aktivitas lisosim benih dari perlakuan induk B memiliki aktivitas lisosim yang signifikan (P<0,05) lebih tinggi dibandingkan perlakuan lainnya hingga hari ke-20 yaitu 100,44-228,71 U/mL. Aktivitas lisosim benih dari perlakuan induk A dan B menurun dihari ke-10 dan kemudian meningkat hingga hari ke-20. Aktivitas lisosim benih dari induk perlakuan A signifikan lebih tinggi (P<0,05) dibandingkan dengan benih dari induk K pada hari ke-5 namun, tidak berbeda signifikan (P>0,05) mulai hari ke-10 hingga hari ke-20. Aktivitas lisosim benih dari induk K dan A berkisar antara 22,78-150,67 U/mL. Aktivitas lisosim benih dari setiap perlakuan induk disajikan pada Gambar 6.
Gambar 6 Aktivitas lisosim benih ikan nila umur benih 5, 10, 15, dan 20 hari setelah menetas pada pemijahan kedua dari perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Huruf yang berbeda di setiap bar pada umur benih yang sama menunjukkan pengaruh perlakuan yang berbeda nyata (Tukey’s P<0,05)
Fekunditas dan Daya Tetas Telur Ikan Nila
Fekunditas telur dari semua induk perlakuan tidak berbeda signifikan (P>0,05). Fekunditas telur dari setiap perlakuan berkisar antara 1.362,67-1.527,33 butir. Namun, terdapat perbedaan yang signifikan (P<0,05) terhadap daya tetas telur. Daya tetas telur perlakuan induk B (94,52%) signifikan lebih tinggi (P<0,05) dari perlakuan induk lainnya. Daya tetas telur perlakuan induk A (84,64%) tidak berbeda signifikan dengan perlakuan induk K (73,72%). Fekunditas dan daya tetas telur dari setiap perlakuan induk disajikan pada Tabel 3. Tabel 3 Fekunditas dan daya tetas telur ikan nila pada pemijahan kedua dari
perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B). Perlakuan Fekunditas Telur (Butir) Daya Tetas Telur (%)
K 1.362,67±123,84a 73,72±6,38b A 1.372,33±148,50a 82,08±3,92b
B 1.527,33±80,16a 94,07±2,60a
Keterangan: Huruf superskrip yang berbeda pada kolom yang sama menunjukkan pengaruh perlakuan yang berbeda nyata (Uji Tukey’s; P<0,05).
13 Tingkat Mortalitas dan Relative Percent Survival (RPS) Benih Ikan Nila
Tingkat mortalitas benih perlakuan induk B signifikan lebih rendah (P<0,05) dari perlakuan lainnya hingga hari ke-20. Namun, mortalitas benih perlakuan induk B tidak berbeda signifikan (P>0,05) dengan perlakuan benih dari induk A pada hari ke-5. Nilai RPS benih dari perlakuan induk B tidak berbeda signifikan (P>0,05) dengan perlakuan benih dari induk A pada hari ke-5. Namun, nilai RPS benih dari induk B signifikan lebih tinggi (P<0,05) dibandingkan perlakuan benih dari induk A mulai hari ke-10 hingga hari ke-20. Nilai RPS benih ikan nila disajikan pada Tabel 4.
Tabel 4 Tingkat mortalitas setelah infeksi S. agalactiae dan relative percent survival benih ikan nila pada pemijahan kedua dari perlakuan induk diinjeksi PBS (K), induk diinjeksi vaksin satu kali (A), dan induk diinjeksi vaksin dua kali dengan selang waktu satu bulan (B)
Umur Benih (Hari)
Mortalitas(%) RPS(%)
K A B A B
5 61,11±5,09a 8,89±3,33b 1,11±1,92b 85,60±7,75a 98,33±2,89a 10 70,00±3,33a 22,22±5,09b 6,67±3,33c 68,43±5,88b 90,61±4,32a 15 76,67±3,33a 37,78±6,94b 17,78±5,09c 50,86±7,84b 81,20±1,80a 20 77,78±5,09a 62,22±3,85b 35,56±5,09c 19,93±6,18b 54,17±7,53a Keterangan: Huruf superskrip yang berbeda pada umur benih dan parameter yang sama menunjukkan pengaruh perlakuan yang berbeda nyata (Uji Tukey’s; P<0,05).
Patologi Anatomi Mikroskopis
Hasil histologi organ otak benih ikan setelah uji tantang menunjukkan adanya perubahan patologi anatomi mikroskopis pada semua perlakuan. Otak ikan yang terinfeksi S. agalactiae terlihat mengalami nekrosis, sedangkan tidak terlihat pada jaringan otak ikan normal. Hasil histologi jaringan otak benih ikan nila yang terserang S. agalactiae disajikan pada Gambar 7.
Gambar 7 Patologi anatomi mikroskopis pada jaringan otak benih ikan nila setelah infeksi S. agalactiae: A) Jaringan otak normal; B) Jaringan otak terserang S. agalactiae mengalami nekrosis (Ne) dan Degenerasi (De)
Pembahasan
Hasil penelitian menunjukkan bahwa pemberian vaksin ulang pada induk ikan nila pada pemijahan kedua memberikan pengaruh yang signifikan lebih baik terhadap transfer imunitas maternal dari induk ke anak ikan nila. Huang et al.,
14
uji tantang (Angelidis, 2006; Ispir & Dorucu, 2010; Zhang et al., 2014; Ito & Maeno, 2015).
Parameter gambaran darah telah digunakan sebagai indikator status kesehatan dibeberapa spesies ikan (Santos et al., 2009; Adedeji, 2010; Akinrotimi
et al., 2012; Sayed & Moneeb, 2015). Penelitian vaksinasi ikan terkait total eritrosit, hemoglobin, dan hematokrit pada ikan nila yang divaksin telah dilaporkan tidak berpengaruh nyata terhadap status kesehatan ikan (Sukenda et al., 2014). Namun, terdapat pengaruh terhadap total leukosit dan aktivitas fagositik ikan yang divaksin (Hardi et al., 2013; Sukenda et al., 2015a). Hasil gambaran darah induk ikan setelah vaksinasi masih menunjukkan nilai yang normal. Akan tetapi, kesehatan induk ikan perlakuan B memiliki status yang lebih baik dari perlakuan lainnya. Hal tersebut dapat dilihat dari nilai total leukosit dan aktivitas fagositik induk pada perlakuan B yang memiliki nilai yang signifikan lebih tinggi dibandingkan dengan perlakuan lainnya. Hardi et al., (2013) dan Amrullah et al., (2014) melaporkan terjadi adanya peningkatan total leukosit dan aktivitas fagositik pada ikan nila yang diberi vaksin. Sel leukosit seperti neutrofil dan monosit merupakan sel-sel fagosit yang aktif ketika ada benda asing yang masuk ke dalam tubuh sebagai pertahanan nonspesifik (Katzenback & Belosevic, 2009).
Antibodi ditemukan dalam konsentrasi yang tinggi pada serum ikan (Magnadottir, 2010). Hasil pengukuran level antibodi perlakuan induk B lebih tinggi dibandingkan induk perlakuan lainnya. Hal ini menunjukkan pemberian vaksin ulang pada induk (B) pada pemijahan kedua dapat meningkatkan level antibodi pada induk, telur, dan benih ikan nila. Antibodi induk A dan B secara signifikan masih lebih tinggi dibandingkan dengan induk tanpa vaksinasi (K). Pemberian vaksin gabungan sel utuh dan ECP pada ikan nila mampu menginduksi tubuh untuk memproduksi antibodi spesifik (Hardi et al., 2013).Hasil fraksinasi protein melalui SDS-PAGE diketahui bahwa ECP bakteri S. agalactiae tipe non-hemolitik memiliki protein dengan bobot molekul protein yaitu 51,8; 55,8 dan 62,3 kDa. Hasil tersebut sama dengan yang dilakukan oleh Hardi et al., (2013) dan bersifat imunogenik. Menurut Pasnik et al., (2005) kandungan protein pada ECP bakteri S. agalactiae yang digunakan sebagai vaksin pada ikan berkisar antara 47-75 kDa. Sukenda et al., (2015a) melaporkan bahwa induk yang divaksin dengan sediaan vaksin gabungan sel utuh dan ECP mampu meningkatkan level antibodi induk dan benih yang dihasilkan. Selain itu menurut Zang et al., (2014) menjelaskan bahwa level antibodi ikan meningkat ketika diberi vaksin ulang.
15 mengakibatkan terbentuknya antibodi lebih cepat dan lebih banyak (Campbell et al., 2004).
Pada proses pematangan gonad, sinyal lingkungan diterima oleh sistem syaraf pusat dan diteruskan ke hipotalamus. Hipotalamus akan melepaskan
Gonadotropin Releasing Hormone (GnRH) yang bekerja di kelenjar hipofisis. Selanjutnya hipofisis melepaskan FSH yang bekerja pada sel teka dan mensintesis testosteron. Di lapisan granulosa, enzim aromatase akan mengubah testosteron menjadi estradiol-17β yang merangsang hati mensintesis vitelogenin (bakal kuning telur). Vitelogenin dibawa oleh aliran darah dan diserap oleh folikel oosit dan oosit membesar sampai ukuran maksimum yang dikenal dengan proses vitelogenesis. Jalur proses vitelogenesis ini memerantarai imunitas spesifik (antibodi) dan non-spesifik (lisosim) untuk masuk ke dalam kuning telur yang akan ditransfer ke anaknya (Swain dan Nayak, 2009; Wang et al., 2011).
Level antibodi telur dari induk perlakuan B memiliki nilai yang lebih tinggi dibandingkan dengan telur dari induk perlakuan lainnya. Hal ini dikarenakan induk yang diberi vaksin ulang memiliki level antibodi paling tinggi dibandingkan perlakuan lainnya, sehingga antibodi dari induk B ditransfer lebih banyak ke telurnya pada proses vitelogenesis. Penelitian pada beberapa spesies ikan juga menunjukkan bahwa antibodi dapat ditransfer dari induk ke anaknya melalui pembentukan kuning telur (Picchietti et al., 2004; Hanif et al., 2004; Dalmo, 2005; Swain et al., 2006).
Nilai level antibodi spesifik terhadap S. agalactiae pada benih secara perlahan mengalami penurunan hingga hingga hari ke-20 diseluruh perlakuan. Antibodi benih dari perlakuan B memiliki nilai yang lebih tinggi dari perlakuan ditransfer melalui induk secara alami ikut termetabolisme bersama dengan kuning telur. Hanif et al., (2004) membuktikan bahwa benih dari induk ikan sea bream
Sparus auratus yang diberi vaksin mengalami penurunan secara perlahan hingga akhir pengamatan yaitu 14 hari setelah telur menetas.
Induk yang divaksin ulang (B) memiliki aktivitas lisosim lebih tinggi dibandingkan dengan perlakuan induk lainnya. Beberapa penelitian menjelaskan bahwa pemberian vaksin bukan hanya meningkatkan respon imun spesifik tetapi juga meningkatkan respon imun non-spesifik (Hanif et al., 2004; Mulero et al.,
2007; Swain dan Nayak, 2009; Wang et al., 2013). Selain itu, telur dan benih hasil pemijahan dari induk yang divaksinasi ulang (B) menunjukkan aktivitas lisosim yang lebih tinggi dibandingkan dengan telur dan benih dari induk perlakuan lainnya. Lisosim muncul di awal pengembangan ikan, sebelum atau segera setelah menetas dan keberadaannya sangat penting terhadap kekebalan benih ikan (Magnadottir, 2006).
16
dari perlakuan induk B memiliki peningkatan yang lebih cepat dibandingkan dengan benih dari induk perlakuan lainnya. Perkembangan lisosim ini terkait dengan perkembangan organ-organ limfoid seperti ginjal, timus, dan limpa (Magnadottir et al., 2005). Lisosim mengandung enzim muramidase yang dapat merusak ikatan antara asam N-asetilglukosamin dengan N-asetilmuramic dalam dinding sel bakteri gram positif (Marsh dan Rice, 2010). Pada tahap awal, lisosim adalah salah satu protein penting yang terlibat dalam pertahanan non-spesifik (Hanif et al., 2005; Huttenhuis et al., 2006; Wang & Zhang, 2010).
Hasil pengukuran fekunditas telur dari semua induk perlakuan memiliki nilai yang sama. Akan tetapi, terjadi perbedaan pada nilai daya tetas telur. Daya tetas telur dari induk perlakuan B lebih tinggi dari perlakuan induk lainnya. Hal ini menunjukkan telur dari induk yang divaksin ulang terjadi peningkatan daya tetas telur, namun terjadi penurunan pada perlakuan induk A yang secara signifikan tidak berbeda dengan perlakuan induk K. Hal ini menunjukkan komponen lisosim dan antibodi yang ditransfer oleh induk telah memberikan pertahanan antibakteri untuk telur dan meningkatkan keberhasilan penetasan dan sintasan (Saino et al., 2002; Hanif et al., 2004; Hanif et al., 2005).
Pemberian vaksin pada induk ikan telah diketahui mampu meningkatkan proteksi benih yang dihasilkan dari serangan patogen (Hanif et al., 2004; Nur et al., 2004; Wang et al., 2012; Sukenda et al., 2015a). Hasil uji tantang benih dengan S agalactiae, diketahui benih dari induk perlakuan B memiliki nilai RPS yang lebih tinggi dibandingkan dengan benih dari induk A dari hari ke-10 hingga hari ke-20. Hal ini menunjukkan bahwa benih yang dihasilkan dari induk yang diberi vaksin ulang (B) memiliki tingkat proteksi lebih tinggi dibandingkan benih dari induk yang diberi vaksin satu kali (A). Nilai RPS benih dari setiap perlakuan menurun hingga hari ke-20 setelah menetas. Hal tersebut disebabkan antibodi spesifik yang ditransfer dari induk ke anaknya semakin berkurang seiring dengan bertambahnya umur benih.
Tingginya nilai RPS benih dari perlakuan induk B disebabkan oleh tingginya antibodi dan lisosim yang ditransfer oleh induk ikan nila B. Imunoglobulin seperti IgM dapat mengopsonin bakteri, sehingga dapat didegradasi dan diberantas oleh sel-sel fagosit (Schroeder & Cavacini, 2010; Rauta et al., 2012). Transfer antibodi maternal ke telur dan embrio juga dilaporkan dapat bertindak sebagai opsonin yang dapat memfasilitasi proses fagositosis bakteri oleh sel fagosit serta mengaktifkan komplemen (Magnadottir et al., 2005). Antibodi maternal dari induk ikan dapat mengurangi resiko kematian selama masa perkembangan benih ikan (Grindstaff, 2008). Hal ini menunjukkan bahwa antibodi spesifik memainkan peran penting dalam perlindungan ikan terhadap infeksi patogen.
17
4
SIMPULAN DAN SARAN
Simpulan
Pemberian vaksin ulang di induk nila pada pemijahan kedua dapat menstimulasi imunitas dan transfer imunitas maternal ke anaknya untuk pencegahan streptococcosis.
Saran
Pemberian vaksin ulang pada induk nila pada pemijahan kedua perlu dilakukan dalam manajemen teknik produksi benih ikan nila. Selain itu perlu dilakukan vaksinasi pada benih nila umur 20 hari untuk meningkatkan imunitas spesifik benih terhadap penyakit streptococcosis.
DAFTAR PUSTAKA
Adedeji OB. 2010. Acute effect of diazinon on blood plasma biochemistry in the African catfish Clarias gariepinus. Journal of Clinical Medicine & Research. 2: 1–6.
Akinrotimi OA, Agokei EO, Aranyo AA. 2012. Changes in blood parameters of tilapia guineensis exposed to different salinity levels. Journal Of Environmental Engineering & Technology. 1: 4–12.
Amrullah, Sukenda, Harris E, Alimuddin, Lusiastuti AM. 2014. Immunogenecity of the 89 kDa toxin protein from extracellular products of Streptococcus in
Oreochromis niloticus. Journal of Fisheries & Aquatic Science. 9: 176-186. Anderson DP, Siwicki AK. 1993. Basic haematology and serology for fish health
programs. Paper Presented in Second Symposium on Diseases in Asia Aquaculture “Aquatic Animal Health and The Environmental”. Phuket
Thailand. 25-29th October 1993.
Angelidis P. 2006. Immersion booster vaccination effect on sea bass
Dicentrarchus labrax L. juveniles. Journal of Animal Physiology & Animal Nutrition. 90: 46–49.
Blaxhall PC, Daisley KW. 1973. Reutine haemotologycal methods for use with fish blood. Journal Fish Biology. 5: 577–581.
Bowden TJ, Cook P, Rombout JHWM. 2005. Development and function of the thymus in teleosts. Fish & Shellfish Immunology. 19: 413-427.
Campbell NA, Reece JB, Mitchell LG. 2004. Biologi Edisi Kelima-Jilid III. Jakarta (ID): Erlangga.
Chen M, Li LP, Wang R, Liang WW, Huang Y, Li J, Lei AY, Huang WY, Gan X. 2012. PCR detection and PFGE genotype analyses of streptococcal clinical isolates from tilapia in China. Veterinary Microbiology. 159: 526–530. Dalmo RA. 2005. Ontogeny of the fish immune system. Fish & Shellfish
Immunology. 19. 395–396.
non-18
hemolitik pada ikan nila Oreochromis niloticus. Jurnal Akuakultur Rawa Indonesia. 2: 105-116.
Evans JJ, Klesius PH., Shoemaker CA, Fitzpatrick BT. 2005. Streptococcus agalactiae Vaccination and Infection Stress in Nile Tilapia, Oreochromis niloticus. Journal of Applied Aquaculture. 16: 105-115.
Evans JJ, Klesius PH, Shoemaker CA. 2006. An overview of Streptococcus in warmwater fish. Aquaculture Health International. 7: 10-14.
Filho CI, Muller EE, Pretto-Giordano LG, Bracarense AP. 2009. Histological finding of experomental Streptococcus agalactiae infection in nile tilapia
Oreochromis niloticus.BrazilianJournal of Veterinary Pathology. 2: 12-15. Genten F, Terwinghe E, Danguy A. 2009. Atlas of Fish Histology. New
Hampshire US: Science Publishers.
Grindstaff JL. 2008. Maternal antibodies reduce costs of an immune response during development. Journal of Experimental Biology. 211: 654-660.
Gudding R, Willem B, Muiswinkel V. 2013. A history of fish vaccination Science-based disease prevention in aquaculture. Fish & Shellfish Immunology. xxx: 1-6. doi.org/10.1016/j.fsi.2013.09.031.
Hanif A, Bakopoulos V, Dimitriadis GJ. 2004. Maternal transfer of humoral specific and non-specific immune parameters to sea bream Sparus aurata larvae. Fish & Shellfish Immunology. 17 : 411-435.
Hanif A, Bakopoulos V, Dimitriadis GJ. 2005. The effect of sea bream Sparus aurata broodstock and larval vaccination on the susceptibility by
Photobacterium damsela subsp. piscicida and on the humoral immune parameters. Fish & Shellfish Immunology. 19: 345-361.
Hardi EH, Sukenda, Harris E, Lusiastuti AM. 2011. Karakteristik dan patogenisitas Streptococcus agalactiae tipe β-hemolitik dan non-hemolitik pada ikan nila. Jurnal Veteriner. 12: 151-164.
Hardi EH, Sukenda, Harris E, Lusiastuti AM. 2013. Kandidat vaksin potensial
Streptococcus agalactiae untuk pencegahan penyakit streptococcosis pada ikan nila Oreochromis niloticus. Jurnal Veteriner. 14: 408-416.
Hirano M, Das S, Guo P, Cooper MD. 2011. The evolution of adaptive immunity in vertebrates. Advances in Immunology. 109: 125–157.
Huang LY, Wang KY, Xiao D, Chen DF, Geng Y, Wang J, Hea Y, Wang EL, Huang JL, Xiao GY. 2014. Safety and immunogenicity of an oral DNA vaccine encoding sip of Streptococcus agalactiae from nile tilapia
Oreochromis niloticus delivered by live attenuated Salmonella typhimurium. Fish & Shellfish Immunology. 38: 34-41.
Huttenhuis HBT, Grou CPO, Taverne-Thiele AJ, Taverne N, Rombout JHMW. 2006. Carp Cyprinus carpio L. innate immune factors are present before hatching. Fish & Shellfish Immunology. 20: 586-596.
Ispir U, Dorucu M. 2010. Effect of immersion booster vaccination with Yersinia ruckeri extracellular products (ECP) on rainbow trout Oncorhynchus mykiss.
International Aquatic Research. 2: 127-130.
Ito T, Maeno Y. 2015. Effect of booster shot and investigation of vaccination efficacy period against herpesviral haematopoietic necrosis (HVHN) in goldfish Carassius auratus. Veterinary Microbiology. 175: 139–144.
19 and red tilapia, Oreochromis sp., of Thailand. Journal of the World Aquaculture Society. 45: 392-402.
Katzenback BA, Belosevic M. 2009. Isolation and functional characterization of neutrophil-like cells, from goldfish Carassius auratus L. kidney.
Developmental & Comparative Immunology. 33: 601–611.
Magnadottir B, Lange S, Gudmundsdottir S, Bogwald J, Dalm RA. 2005. Ontogeny of humoral immune parameters in fish. Fish & Shellfish Immunology. 19: 429–439.
Magnadottir B. 2006. Innate immunity of fish: overview. Fish & Shellfish Immunology. 20: 137-151.
Magnadottir B. 2010. Immunological control of fish diseases. Journal of Marine Biotechnology. 12: 361–379.
Marsh MB, Rice CD. 2010. Development, characterization, and technical applications of a fish lysozyme-specific monoclonal antibody (mAb M24-2).
Comparative Immunology, Microbiology & Infectious Diseases. 33: 15–23. Martins ML, Mourino JLP, Amara GV, Vieira FN, Dotta G, Jatoba AMB, Pedrotti
FS, Jeronimo GT. 2008. Haematological changes in nile tilapia experimentally infected with Enterococcus sp. Journal Biology. 68: 657-661. Mian GF, Godoy DT, Leal CAG, Yuhara TY, Costa GM, Figueiredo. 2009.
Aspect of the natural history and virulence of Streptococcus agalactiae
infection in nile tilapia. Journal of Veterinary Microbiology. 136: 180-183. Mingming H, FuHong D, Zhen M, Jilin L. 2014. The effect of vaccinating turbot
broodstocks on the maternal immunity transfer to offspring immunity. Fish & Shellfish Immunology. xxx: 1-7. doi: org/10.1016/j.fsi.2014.03.010. Moyle PB, JJ Cech Jr. 1988. Fishes: An Introduction to Ichthyology. Prentice
Hall, NJ: Englewood Cliffs Press.
Muiswinkel WBV. 2008. A history of fish immunology and vaccination I. The early days. Fish & Shellfish Immunology. 25: 397-408.
Mulero I, García-Ayala A, Meseguer J, Mulero V. 2007. Maternal transfer of immunity and ontogeny of autologous immunocompetence of fish: minireview. Aquaculture. 268: 244–250.
Nur I, Sukenda, Dana D. 2004. Ketahanan benih ikan nila gift Oreochromis niloticus L. dari hasil induk yang diberi vaksin terhadap infeksi buatan
Streptococcus iniae. Jurnal Akuakultur Indonesia. 3: 37-43.
Pasnik DJ. Evans JJ. Klesius PH. 2005. Duration of protective antibodies and correlation with survival in nile tilapia Oreochromis niloticus following
Streptococcus agalactiae vaccination. Journal of Fish Disease. 66: 129-134. Picchietti S, Taddei AR, Scapigliati G, Buonocore F, Fausto AM, Romano N. 2004. Immunoglobulin protein and gene transcripts in ovarian follicles throughout oogenesis in the teleost Dicentrachus labrax. Cell & Tissue Research. 315: 259–270.
Rauta PR, Bismita N, Das S. 2012. Immune system and immune responses in fish and their role in comparative immunity study: A model for higher organisms. Immunology Letters. 148: 23–33.
20
Santos AA, Ranzani-Paiva MJT, Veiga ML, Faustino L, Egami MI. 2009. Hematological parameters and phagocytic activity in fat snook
Centropomus parallelus bred in captivity. Fish & Shellfish Immunology. 33: 953-961.
Sayed AEH, Moneeb RH. 2015. Hematological and biochemical characters of monosex tilapia Oreochromis niloticus Linnaeus 1758 cultivated using methyltestosterone. The Journal of Basic & Applied Zoology. 72: 36–42. Schroeder HW JR, Cavacini L. 2010. Structure and function of immunoglobulins.
Journal of Allergy & Clinical Immunology. 125: S41–52.
Secombes CJ. 2011. Fish immunity: the potential impact on vaccine development and performance. Aquaculture Research. 42: 90-92.
Sheehan B, Labrie L, Lee Y, Lim W, Wong F, Chan J. 2009. Streptococcal diseases in farmed tilapia. Aquaculture Asia Pacific. 5: 26–29.
Shelby RA, Shoemaker CA, Evans JJ, Klesius PH. 2001. Development of an indirect ELISA to detect humoral response to Streptococcus iniae infection of nile tilapia Oreochromis niloticu. Journal of Applied Aquaculture. 11: 35-44.
Sommerset I, Krossoy B, Biering E, Frost P. 2005. Vaccines for fish in aquaculture. Review Expert Review of Vaccines. 4: 89-101.
Sukenda, Febriansyah TR, Nuryati S. 2014. Efikasi vaksin sel utuh Streptococcus agalactiae pada ikan nila Oreochromis niloticus melalui perendaman.
Jurnal Akuakultur Indonesia. 13: 83-93.
Sukenda, Rahman, Nisaa K, Hidayatullah D. 2015a. The efficacy of Streptococcus agalactiae vaccine preparations on tilapia broodstock and maternal immunity transferred to offspring in preventing streptococcosis. Journal of Fisheries & Aquatic Science. Submitted.
Sukenda, Rusli, Nuryati S, Hidayatullah D. 2015b. Durasi proteksi vaksin
Streptococcus agalactiae untuk pencegahan streptococcosis pada ikan nila.
Jurnal Akuakultur Indonesia. Submitted.
Svobodova Z, Vyukusova B. 1991. Diagnostik, Prevention and Therapy of Fish Disease and Intoxication. Vodnany: Research Institute of fish Culture and Hydrobiology.
Swain P, Dash S, Bal J, Routray P, Sahoo PK, Sahoo SK, Saurabh S, Gupta SD, Meher PK. 2006. Passive transfer of maternal antibodies and their existence in eggs, larvae and fry of Indian major carp, Labeo rohita. Fish & Shellfish Immunology. 20: 519-527.
Swain P, Nayak S.K. 2009. Role of maternally derived immunity in fish. Fish & Shellfish immunology. 27: 89-99.
Takashima F, Hibiya T. 1995. An Atlas of Fish Histology: Normal and Pathogical Features. 2nd ed. Tokyo: Kodansha Ltd.
Taukhid, Purwaningsih U, Lustiastuti AM. 2012. Pengambangan vaksin inaktiv bakteri Streptococcus agalactiae: Penentuan teknik aplikasi dan dosis efektif vaksin melalui perendaman untuk pencegahan penyakit streptococcosis pada ikan nila Oreochromis niloticus. Prosiding seminar.
Bogor (ID). Balai Riset Perikanan Budidaya Air Tawar.
21 Wang SH, Wang Y, Ma J, Ding YC, Zhang SC. 2011. Phosvitin plays a critical role in the immunity of zebrafish embryos via acting as a pattern recognition receptor and an antimicrobial effector. Journal of Biological Chemistry.
286: 22653–22664.
Wang H, Ji D, Shao J, Zhang S. 2012. Maternal transfer and protective role of antibodies in zebrafish Danio rerio. Molecular Immunology. 51: 332– 336. Wang N, Yang Z, Zang M, Liu Y, Lu C. 2013. Identification of Omp38 by
immunoproteomic analysis and evaluation as a potential vaccine antigen against Aeromonas hydrophila in Chinese breams. Fish & Shellfish Immunology. 34: 74-81.
Wedemeyer IWT, Yasutake WT. 1977. Clinical methods for the assessment of the effect on environmental stress on fish health. Journal fish & wildlife service.
89: 1-17.
Yang W, Li A. 2009. Isolation and characterization of Streptococcus agalactiae
from diseased Acipenser schrenckii. Aquaculture. 294: 14-17.
Ye X, Li J, Lu MX, Deng GC, Jiang XY, Tian YY, Quan YC, Jian Q. 2011. Identification and molecular typing of Streptococcus agalactiae isolated from pond-cultured tilapia in China. Fisheries Science. 77: 623-632.
Yi T, Li WY, Liu L, Xiao XX, Li AX. 2014. Protection of nile tilapia
Oreochromis niloticus L. against Streptococcus agalactiae following immunization with recombinant FbsA and α-enolase. Aquaculture. 428– 429: 35–40.
Zapata A, Díez B, Cejalvo T, Gutiérrez-de CF, Cortés A. 2006. Ontogeny of the immune system of fish. Fish & Shellfish Immunology. 20: 126–136.
Zhang S, Wang Z, Wang H. 2013. Maternal immunity in fish: Review.
Developmental & Comparative Immunology. 39: 72–78.
22
LAMPIRAN
Lampiran 1 Karakterisasi bakteri S. agalactiae menggunakan KIT API 20 Strep
Tests Active ingredients Hasil
VP Sodium pyruvate +
HIP Hippuric acid +
ESC Esculin ferric citrate - PYRA Pyroglutamic acid-β-naphthylamide -
αGAL 6-bromo-2-naphthyl-αD-galactopyranoside -
βGUR Naphthol ASBI-glucuronic acid -
βGAL 2-naphthyl-βD- galactopyranoside -
PAL 2-naphthyl phosphate +
LAP L-leucine-β- naphthylamide +
ADH L-arginine +
RIB D-ribose +
ARA L-arabinose -
MAN D-mannitol -
SOR D-sorbitol -
LAC D-lactose (bovine origin) -
TRE D-trehalose -
INU Inulin -
RAF D-raffinose -
AMD Strach (2) -