Tradakfi eng&u f i t 6atiwa;4hh menumnt&an tiujan dari lbngit, &fu me&lir
di
dalbm tunahmnjadi
matu air? wm&n itu, tum6uti krenunya tanaman-tunamun yang 6ennacam-macam warnanya; Ibfu menjadilhyc
danbtting
&li&tannya, akfiimya&ring
dantiancur.
Sesunggutinya dalbm &C itu menjadi peringatan6agi
orang-orang yang 6erakaf (jk
-
ZumarZI)PERTUMBUHAN
DAN
PERKEMBANGAN
Scirpophaga innotata
WALKER
(LEPIDOPTERA. PYRALIDAE) PADA TANAMAN PAD1
DI
TANAH
DENCAN KANDUNGAN KALIUM BERBEDA
Oleh
TEDDY
SUPARNO
PROGRAM PASCASARJANA
INSTITUT
PERTANIAN
BOCOR
SUMMARY
TEDDY
SUP^.
The Growth and Development of Scirpophaga innotataWalker (Lepidoptera: Pyralidae) on Rice Plant Grown on Soil with Different
Potassium Contents (Under Guidance of SOEMARTONO SOSROMARSONO,
SIDARTO WARDOJO, SJAFRIDA M W O T O , AUNU RAW, and M.
SJARKAnrr MUSA).
The objective of the research is to study the growth and development of
the white stem borer (WSB), Scirpophaga innotata, on rice plant grown on soil
with a very low potassium content treated with potassium fertilizer to various
potassium levels.
Four experiments were conducted at the green house of Bogor Research
Institute for Food Crops, i. e (1) on the larval growth and development, and
fecundity of WSB on the rice plant, (2) on the damage of the rice plant caused by infestation of WSB, (3) on the physical changes of the larval mandible
resulted from the boring activity on the rice plant, and (4) on the diapause
incidence of WSB larvae feeding on the rice plant. All experiments were
conducted on soil with very low available potassium (50 ppm K) and high
available silica (141 ppm Si0J treated with potassium fertilizer. There were five
levels of potassium fertilizer treatment, i. e. 0, 75, 150,225 and 300 ppm K.
For the experiment on the diapause incidence additional soil treatments were
added, i. e. water saturated soil, soil water content at field capacity (26 per cent
cent water content set 80 days after planting). All experiments used
IR64
ricevariety.
The results showed that increasing rates of potassium fertilizer up to 150
ppm K, larval growth rate and development, fecundity and egg viability were
significantly increased and decreased slightly at 225 and 300 ppm K (high levels). However fecundity and egg fertility of WSB are still moderately high at 225 and
300 ppm K both in the vegetative as well as in the generative growth stage of the
rice plant, i. e. 233 k 31 eggs per female with 93
*
1.2 per cent of egg fertility and 312*
37.2 eggs per female with 94.2*
0.8 per cent egg fertility,respectively. Mortality rate of WSB were also still moderately high at 225 and
300 ppm K. High levels of potassium inhibited larval infestation significantly,
and reduced both early damage symptoms and maximum damage. However,
potassium treatment could not induce resistance to WSB in IR64 rice variety.
Rice plants fertilized with potassium at levels from 0 to 150 ppm had no
woming effect on the mandible teeths of first instar larvae. High levels of
potassium (225 and 300 ppm K) showed no significant effect of worning on the
mandible teeths of first instar, but the effect was significant on third instar larvae
which bored stems of the generative growth stage of the rice plant.
Rice plants fertilized with potassium at levels from 0 to 300 ppm K
had
no effect on WSB diapause incidence. Soil water content had significantly
affect WSB diapause incidence. The incidence of larval diapause on rice
plants grown in water saturated soil was very low (3.72
*
0.92 per cent), high(68.4
*
11.2 per cent) 00 rice plants grown in soil at field capacity, and veIyhigh (82.3 15.9 per cent) on rice plants grown in soil at half field capacity.
Abscisic acid (ABA) is assumed to play an important role in the process
leading to diapause. ABA content in the rice plant increased with the decrease
of the soil water content and thereby increasing the percentage of larvae
RINGKASAN
TEDDY SUPARNO. -~erturnbuhan dan Perkembangan Scirpophaga lnnoiata
Walker (Lepidoptera: Pyralidae) pada Tanaman Padi di Tanah dengan Kandungan Kalium Berbeda (Di b a w d bimbingan SOEMARTONO SOSROMARSONO
sebagai Ketua, SIDARTO WARDOJO, SJAFRIDA MANUWOTO, AUNU
RAUF dan M. SJARKANI MUSA sebagai Anggota).
Penelitian bertujuan untuk memahami perkembangan penggerek batang
padi putih Scirpophaga innnotata &lam tanaman padi yang ditanam pada tanah dengan kandungan kalium tersedia sangat rendah yang diberi tambahan pupuk
kalium dari dari dosis sangat rendah sampai tinggi.
Empat percobaan telah dilakukan yaitu (1) pertumbuhan dan perkem-
bangan larva serta keperidian betina PBPP dalam tanaman padi, (2) kerusakan
tanaman padi yang diinfestasi oleh larva PBPP, (3) perubahan kondisi fisik gerigi
mandibel larva PBPP yang menggerek batang padi, dan (4) kejadian 'diapause.
Untuk keempat percobaan itu digunakan tanah media tanam yang mengandung
kalium tersedia sangat rendah (50 ppm K) dan silikat tersedia tinggi (14 1 ppm Si0
3
yang diberi perlakuan pupuk kalium dengan dosis 0.75, 150,225, dan 300 ppm
K. Untuk percobaan kejadian diapause masih ditambah perIakuan kelengasan
tan& yaitu jenuh air (diberi air sampai setinggi 1
-
2 cm dari perrnukaan tanahlapang (13% air). Semua penelitian menggunakan varietas IR64.
Hasil penelitianmenunjukkan bahwa pada peningkatan pemberian pupuk
kalium sampai 150 ppm K, pertumbuhan larva, laju perkembangan lama,
keperidian, dan fertilitas telur nyata meningkat, kemudian menurun pada dosis 225
dm semakin menurun pada dosis 300 ppm K. Namun pemberian pupuk katium
pada dosis tinggi (225 ppm K dan 300 ppm K) itu menunjukkan angka keperidian
dan
fertilitas telur mash cukup tinggi, yaitu 200.2-
264.0 telur per betina dengan fertilitas telur 92.0-
94.0% untuk fase vegetatif dan 276.0-
349.2 telur per betina dengan fertilitas 93.5-
95.0% untuk fase generatif Mortalitas larva pada keduafase pertumbuhan padi yang diberi pupuk kalium dengan dosis tinggi adalah cukup tinggi, yaitu 35-60 %.
Pemberian
K
dengan dosis 0 sampai 150 ppmK
tidak berpengaruh nyatadalam menekan infestasi larva. Dosis tinggi (225 clan 300 ppm
K)
berpengaruhnyata dalam mengharnbat timbulnya gejala kerusakan awal
dan
menekan kerusakan (sundep dan beluk) maksimum tanaman padi. Tetapi pemberian pupuk kaliumsampai dosis tinggi tidak dapat menginduksi tanaman padi IR64 menjadi tahan
terhadap serangan PBPP.
Pemberian pupuk K &lam dosis 0 sampai 150 ppm K ti&k berpengaruh nyata terhadap kemakan gerigi mandibel larva instar kesatu. Pemberian pupuk
kalium dosis tinggi hanya berpengaruh nyata terhadap kerusakan gerigi mandibel
Pemberian pupuk kalium dengan dosis 0
-
300 ppm K tidak berpengaruh nyata terhadap penmu@ kejadian diapause PBPP. Kelengasan tanah berpengaruh nyata terhadap kejadian diapause. Kejadian diapause PBPP di dalam tanaman padiIR64 pada kondisi jenuh air, kelengasan tanah kapasitas lapang dan setengah
kapasitas lapang berturut-turut adalah sangat rendah (3.7
*
0.92%), cukup tinggi(68.4 h 11.2%), dan tinggi (82.3 15.9%).
Senyawa sekunder asam absisat
(ABA)
diduga sangat penting dalam proses menuju kondisi diapause. Kandungan ABA di dalam tanaman padi meningkatdengan t u m y a kandungan air tanah dan sejalan dengan itu persentase diapause
PERTUMBUHAN DAN PERKEMBANGAN Scirpophaga innotata WALKER
(LEPIDOPTERA: PYRALIDAE) PADA TANAMAN PAD1 DI TANAH
DENGAN KANDUNGAN KALIUM BERBEDA
Oleh
TEDDY SUPARNO
Disertasi sebagai salah satu syarat untuk memeperoleh gelar doktor dalam ilmu-ilmu pertanian
pada
Program Pascasarjana, Institut Pertanian Bogor
PROGRAM
PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul :
PERTUMBUHAN
DAN PERKEMBANGAN Scirpophaga innotota Walker (LEPIDOPTERA: PYRALIDAE) PADA TANAMAN PAD1 DI TANAH DENGAN KANDUNGAN KALIUM BERBEDANama Mahasiswa : Teddy Suparno
Nomor Pokok : 89 524
enyetujui
Prof. Ir. ~oemartono'~osrornarsono, Ph.D.
/ Ketua
Dr. Ir. Sidarto Wardojo Ir. j. Sjafrida Manuwoto, PhD
Anggota / Anggota
.I
Ir. Aunu Rauf, PhD Dr. Ir. M. Sjarkani Musa
Anggota Anggota
Ketua Program Studi
Ir. Aunu Rauf, Ph.D.
RlWAYAT HIDUP
Penulis addah anak ketiga dari delapan bersaudara dari ayah Soeroto
Soerowitono dan ibu Sockati, lahir tanggal 20 Mei 1954 di Boyolali. Ia menye-
lesaikan SMA Jurusan Pasti dan Alam pada tahun 1972 di Solo. Pada tahun
1978 memperoleh gelar Sarjana Muda Biologi dan pada tahun 1980
memperoleh gelar Sarjana Biologi dari Fakultas Biologi Universitas Kristen
Satyawacana Salatiga. Kemudian pa& tahun 1983 memperoleh gelar Magister
Sains Entomologi Kesehatan dari Fakultas Pascasajana, Institut Pertanian
Bogor. Tahun 1989 memperoleh kesempatan untuk mengikuti studi Program
Doktor di Institut Pertanian Bogor.
Tahun 1979
-
1980 bekerja sebagai staf edukatif Fakultas PertanianUniversitas Tidar Magelang. Tahun 1981
-
1986 bekerja di Direktorat Jenderalembera ant as an Penyakit Menular, Departemen Kesehatan R.I. Mulai tahun
1987 bekerja sebagai tenaga edukatif pada Laboratorium Hama dan Penyakit
Tumbuhan di Fakultas Pertanian Universitas Bengkulu hingga sekarang.
Penulis menikah dengan Yuniatmi Wahyuningsih puteri d a ~ i ayah
Soeharlan SoeroWdjojo dan ibu Siti Mariyah (Almarhumah) pada tanggal 14
Januari 1979 di Amabarawa, dan dikarunia tiga orang anak, Andang Siswo
Agung Raharjo (16 tahun), Mukti Dono Wilopo (12 tahun) dan Masitoh
UCAPAN TERIMA KASIH
Penulis sangat berterima kasih kepada Prof. Ir. Soemartono
Sosromarsono, Ph.D.. sebagai Ketua Komisi Pembimbing, atas saran,
bimbingan dan dorongannya daIam penyelesaian disertasi ini. Kepada Anggota-
Anggota Komisi Pembimbing, Dr. Ir. Sidarto Wardojo, Ir. Hj. Sjafrida
Manuwoto, Ph.D, Ir. Aunu Rauf, Ph.D., dan Dr. Ir. M. Sjarkani Musa yang
telah memberikan saran dan laitik yang sangat bermanfaat sejak awal penelitian
sampai selesainya disertasi ini diucapkan terima kasih.
Kepada Rektor Universitas Bengkulu, Direktur Program Pascasarjana
IPB dan Rektor IPB, Ketua Program Studi Entomologi dan Fitopalogi IPB,
yang telah memberikan kesempatan penulis untuk studi S3 di Program Pascasarjana IPB diucapkan terima kasih.
Ucapan terima kasih juga disampaikan kepada Drs. A. H. Razie Jachya,
Gubemur Propinsi Bengkulu (Periode 1989-1994) dan Drs. Usup Supriyadi,
Wagub Propinsi Bengkulu (Periode 1994-1999); Kepada Ir. Ratiq Ahmad, MS,
Kepala Pusat Penelitian dan Pengembangan Transmigrasi, Departemen Trans-
migrasi dan Permukiman Perambah Hutan; dan Yayasan Aji Dharma Bhakti
-
P.T. Ajino Moto Jakarta yang telah membantu dana penelitian dan penyelesaian
Penelitian ini tidak akan terlaksana dengan baik tanpa a n t u a n Kepala Bdai Penelitian Tanarnan Pangan, Ketua Kelompok Peneliti Entomologi
dan
Ketua Kelompok Peneliti Biokimia Balittan Bogor, dan Ketua kelompokPeneliti Bioteknologi Puslitbang Tanaman Pangan Bogor serta Kepala Pusat
Penelitian Botani Lembaga Biologi Nasional Bogor, untuk itu diucapkan terima
kasih.
Terima kasih yang sebesar-besamya ditujukan kepada Ir. M. Irnan,
Ph.D. yang telah mengizinkan nunah kacanya digunakan untuk penelitian
selama dua setengah tahun, membantu mencarikan bahan kirnia dan cara
penggunaan alat-alat di Laboratorium Entomologi dan Biokimia Balittan Bogor,
sehingga penulis dapat menyelesaikan penelitian disertasi ini.
Ucapan terirna kasih juga disampaikan kepada Lalu Soekamo, B.Sc,
yang telah banyak membantu menganalisis senyawa fitokimia contoh jaringan
tanarnan padi dengan menggunakan High Per$ormance Liquid
Chromatography (HPLC) di Laboratorium Biokimia Balittan Bogor.
Kepada Dr. Ir. Molide Rizal yang telah meminjamkan kornputer selama
penyelesaian disertasi ini, dan kepada Ir. Hermanu Triwidodo, Ph.D yang telah
memberikan fasilitas di Laboratorium Lapang, Panyingkiran, Karawang, untuk
melihat situasi seranganPBPP di lapang sebelum mengadakan penelitian ini dan
memberikan sebagian peralatan penelitian laboratorium, penulis ucapkan terima
Kepada Woro Edi Untari, karyawati P.T. Multirejeki
$its
Jakarta, atas usahanya mencarikan bahan standard Abscisic Acid (ABA) ke Industri KimiaMerck di Jerman, diucapkan terima kasih. Demikian juga ucapan terima kasih
disampakan kepada Tutom, laboran pada Kelompok Peneliti Entomologi di
Balai Penelitian Bioteknologi Tanaman Pangan, yang telah membantu dalam
pembiakan penggerek batang padi putih, sehingga penulis selalu memperoleh
larva penggerek batang padi putih sesuai dengan jadwal inokulasi yang telah
direncanakan.
Kepada Ir. Waluyo Wahyutomo yang telah m e m b e h fasilitas rumah
dari bulan Juli 1991 sampai September 1994, dan Dr. Ir. Hasnam yang telah
memberikan fasilitas rumah dari bulan September 1994 sampai selesai studi
S-3, penulis ucapkan terima kasih.
Akhimya kepada isteri dan anak-anak tercinta penulis mengucapkan
terima kasih atas cinta kasih, dorongan, pengertian, bantuan clan kesabaran sejak
DAFTAR TABEL
Nomor
Teks
1. Hasil determinasi sifat fisika dan kimia contoh tanah dari
Ciampea, Bogor
. . .
. . . .
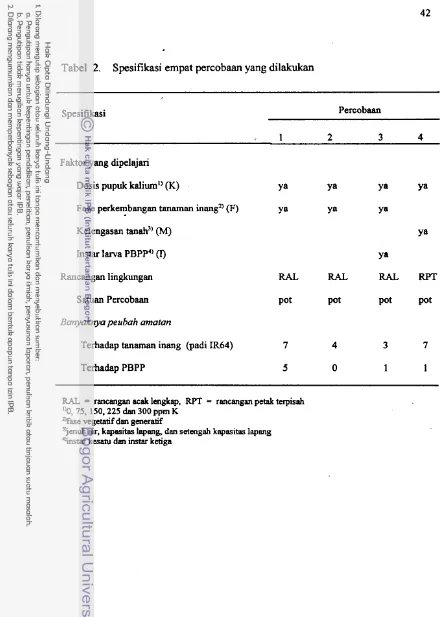
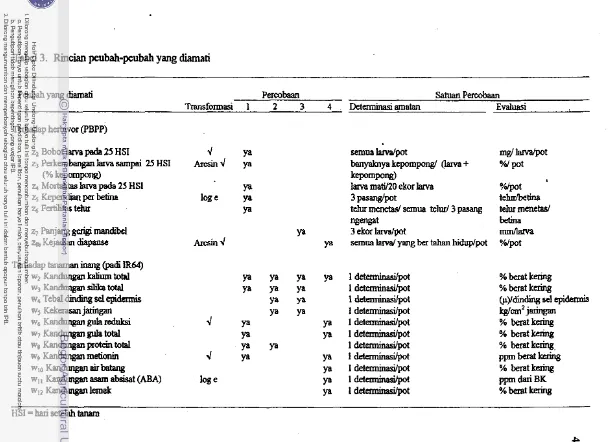
2. Spesifikasi empat percobaan yang dilakukan3. Rincian peubah-peubah yang diamati . . .
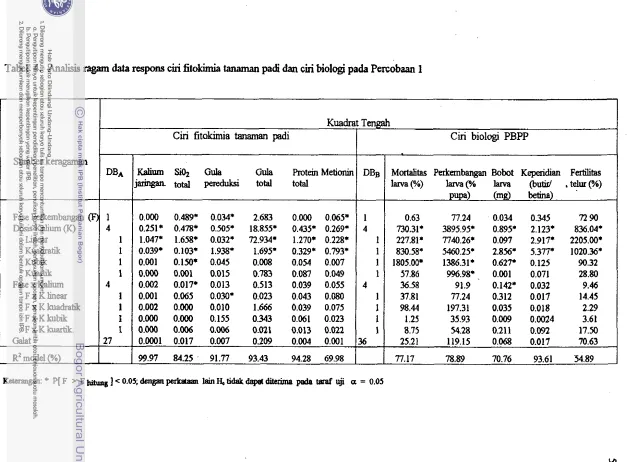
4. Analisis ragarn data respons ciri fitokimia clan biologi PBPP Percobaanl
. . .
5. Koefisien-koefisien regresi dari ubah respons pada dosis pupuk kalium (X) &lam ~ e r c o ~ 1
. . .
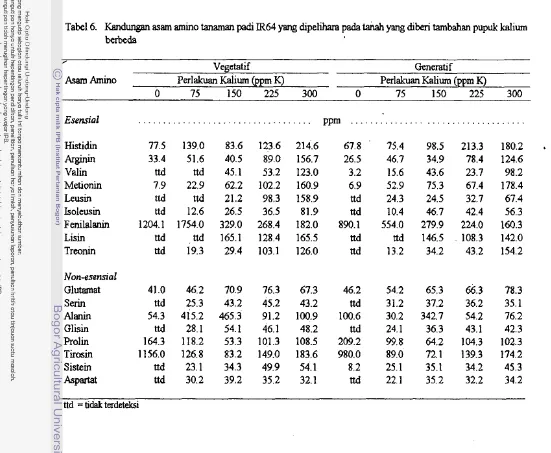
6. Kandungan asam amino tanaman pa& IR64 ang dipelihara pada tanah yang diberi tambahan pup& kazum berbeda
.
.
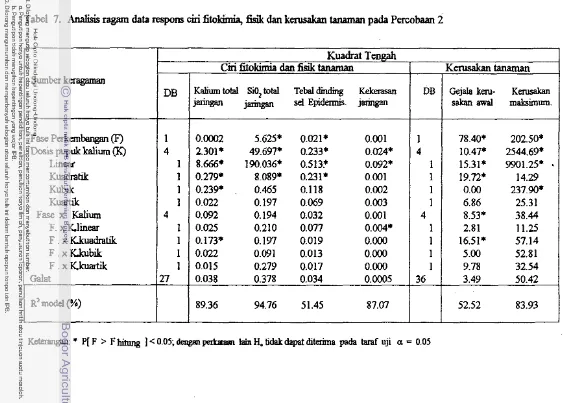
7. Analisis ragam data respons ciri fitokimia, fisik dan kerusak-
an tanaman pa& Percobaan 2
. . .
8. Koefisien-koefisien regresi dari ubah respons pa& dosis
kalium
(x)
dalam percoEan 2. . .
9. Analisis ragam data respons ciri fitokirnia, fisik tanaman padi
dan kondisl gerigi mandibel lava instar ketiga pada Perco- baan3
. . .
10. Koefisien-koefisien regresi dari peubah respons pada dosis
. . .
pupuk kalium (X) dalam Percobaan 311. Analisis ragarn data respons ciri fitokimia dan ciri biologi
. . .
padapercobaan 412. Koefisien-koefisien regresi dari peubah respons pada kele-
. . .
ngasan tanah (X) dalam Percobaan 4
13. Rataan kejadian diapause dan ngengat yang muncul dari
tanaman padi IR64 yang diberi perlakuan hara kalium dan
. . .
kelengasan tanah berbeda14. Rekapitulasi hasil pengamatan Percobaan 1
-
4. . .
Lampiran
Nomor
. . .
1. Diskripsi tanaman padi varietas IR64
2. Pengaruh dosis pupuk kalium terhadap atribut biologi pada
. . .
Percobaanl3. Pengaruh dosis pupuk kalium terhadap kandungan senyawa
. . .
fitokimia tanaman padi pa& Percobaan 1
4. Kerusakan tanaman padi IR64 yang diberi dosis pupuk kalium berbeda dan diinokulasi 20 larva per rumpun pada
. . .
Percobaan25 . Pengaruh dosis pupuk kalium terhadap kandungan senyawa fitokimia dan kondisi fisik tanaman padi pada Percobaan 2
. .
6 . Pengaruh dosis pupuk kalium terhadap kandungan senyawa fitokimia dm kondisi fisik jaringan tanaman padi dan panjang gerigi mandibel larva instar kesatu pa& Percobaan 3. . .
7. Pengaruh dosis pupuk kalium terhadap kandungan senyawa fitokimia dan kondisi fisik jaringan tanaman padi dan panjang gerigi mandibel larva instar ketiga pada Percobaan 3
. . .
8. Pengaruh dosis pupuk kalium terhadap kandungan senyawa fitokimia batang @ dan kejadian diapause pada tiga kelengasan tanahpadapercobaan 4
. . .
halaman
117
DAFTAR GAMBAR
Nomor Halaman
1. Gerigi mandibel yang diukur
. . .
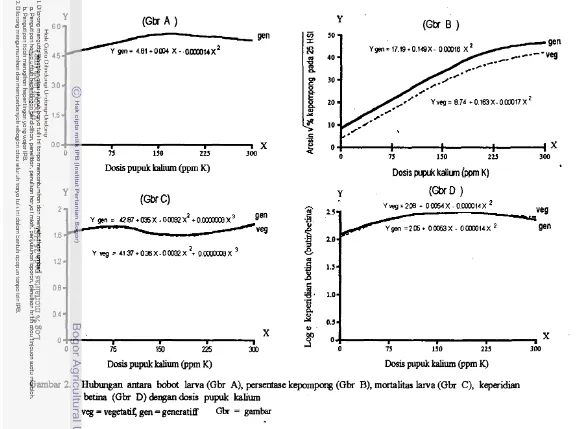
532. Hubungan antara bobot larva (Gbr A), persentase kepompong (Gbr B), mortalitas larva (Gbr C), dan keperidian betina (Gbr D) dengan
. . .
dosispupukkalium 62
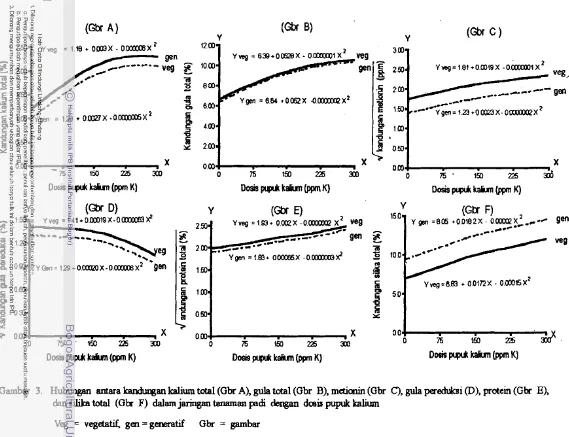
3. Hubungan antara kandungan kalium total (Gbr A), gula total (Gbr B), metionin (Gbr C), gula pereduksi (Gbr D), protein (Gbr E) dan silika total (Gbr F) dalam jaringan tanaman padi dengan dosis
. . .
pupukkalium 63
4. Hubungan antara sundep dan beluk maksimum (Gbr A), kandungan
kalium total (Gbr B), k a n d u m silika total (Gbr C) dan kekerasan
...
jaringan ( ~ b ; D) &man dengan dosis kalium 73
5 . Hubungan antara gigi mandibel terpanjang larva instar ketiga (Gbr A), kandungan kalium total (Gbr B), kekerasan jaringan (Gbr C), dan kandungan silika total (Gbr D) jaringan tanaman padi dengan
dosispupukkalium
. . .
80
6. Gerigi mandibel larva instar yang menggerek batang padi
IR64
.
.
82 7. Penampang membujur jaringan epidermis batang padi XR64. . .
848. Hubungan antara kejadian diapause (Gbr A), kandungan air (Gbr B), ABA (Gbr C) dan metionin (Gbr D)dalam jaringan batang padi
DAFTAR
IS1
DAFTARTABEL
. . .
DAFTARGAMBAR
. . .
PENDAHULMAN
. . .
. . . Latar Belakang
. . .
Rumusan Masalah
. . .
Tujuan Penelitian
. . .
Hipotesis
TMJAUAN PUSTAKA
. . .
Kalium
. . .
Kalium dan Fungsinya di dalam Tanaman
. . .
Hubungan Serangga Herbivora clan Tanaman yang Diberi Pupuk
. . .
Kalium Berbeda
Biologi Penggerek Batang Padi
. . .
Larva
. . .
Diapause
. . .
Kepompong. Ngengat dan Telur
. . .
BAHAN DAN METODE
. . .
Bahan . . .
MetodePercobaan
. . .
Percobaan 1
.
Pertumbuhan dan Perkembangan Larva. . .
serta Keperidian Betina
Percobaan 2
.
Kerusakan Tanaman Padi yang Diinfestasi. . .
oleh Larva PBPP
Percobaan 3
.
Perubahan Kondisi Fisik Gerigi Mandibel. . .
Larva PBPP yang Menggerek Tanaman Padi
. . .
Percobaan 4.
Kejadian DiapauseAnalisis Data
. . .
HASILDANPEMBAHASAN
. . .
Perhunbuhan clan Perkembangan Larva serta Keperidian Betina
Kemsakan Tanaman Padi yang Diinfestasi oleh Larva PBPP
. .
Perubahan Kondisi Fisik Gerigi Mandibel Larva PBPP yang
. . .
Menggerek Tanaman PadiKejadian Diapause
. . .
PembahasanUmum
. . .
KESIMPULAN DAN SARAN
. . .
. . .
KesimpulanSaran
. . .
DAFTAR PUSTAKA
. . .
PENDAHULUAN
Latar Belakang
Beras merupakan bahan pangan pokok sebagian besar penduduk Indonesia
dan bagi lebih dari setengah penduduk dunia. Untuk memenuhi permintaan yang
terus meningkat pemerintah terus bempaya meningkatkan produksi beras, melalui
program ekstensifikasi clan intensifikasi budidaya tanaman padi. Kedua usaha itu
membawa Indonesia mencapai swasembada beras pada tahun 1984 (Departemen
Pertanian, 1988).
Selama pembangunan jangka panjang tahap pertama (PJPT I) peningkatan
produksi bems melalui program intensifikasi lebih berarti dibanding dengan melalui
program ektensifikasi. Luasan panen yang pada tahun 1969 (awal PJPT I) 8 juta
hektar baru meningkat menjadi I l juta hektar pada tahun 1993 atau meningkat
sebesar 35%. Sedangproduksi meningkat dari 18.01 juta ton menjadi 48 2 juta ton
gabah kering giling atau rneningkat hampir sekitar tiga kali lipat. Peningkatan produksi
itu sebagian besar diperoleh melalui program intensifikasi (Warta Pertanian, 1994).
Luasan lahan intensifikasi pada awal PJPT I masih sangat sempit, yaitu dari
8.01 juta hektar luas panen yang telah intensifikasi baru rnencapai sekitar 2.1 juta hektar. Perkembangan luasan tanaman padi intensifikasi sangat pesat sehingga pada
tahun 199 1 luas lahan intensifikasi itu menjadi berbalik komposisinya, yaitu dari has-
an panen sekitar 10.19 juta hektar yang sudah menjadi lahan intensifikasi sekitar 8.64
berkembang.
Pelaksanaan usaha ektensifikasi dan intensifikasi itu menghadapi banyak
kendala, di antaranya adalah serangan hama dan penyakit. Terdapat sekitar 100
spesies serangga yang menyerang tanaman padi, 20 spesies di antamnya menyebabkan
kerusakan berarti. Penggerek batang padi tennasuk serangga hama penting yang
menyebabkan kerusakan ekonomis pada tanaman padi di Indonesia dan di negeri-
negeri Asia lainnya (Pathak, 1968; Soehardjan dan Iman, 1980)
Di Indonesia terdapat enam jenis penggerek batang padi, yaitu penggerek
batang padi kuning, Scirpophaga incertulas Walker (Lepidoptera: Pyralidae), penggerek batang padi putih, Scirpophaga innotata Walker (Lepidoptera: Pyralidae),
penggerek batang padi merah jambu, Sesamia fnferens Walker (Lepidoptera: Noctuidae), penggerek batang padi bergaris, Chilo suppressalis Walker (Lepidoptera:
Pyralidae), penggerek batang padi berkepala hitam, Chilo polych~sus Meyrick (Lepidoptera: Pyralidae) dan penggerek batang padi berkilat, Chilo auricilius Dudgeon (Lepidoptera: Pyralidae). Keenam spesies itu adalah hama penting pada
tanaman padi, namun penggerek batang padi putih merupakan hama yang paling
penting atau paling merusak di antara keenam spesies tersebut (van der Laan, 1959;
Soenardi, 1964; Kalshoven, 1981; Sosromarsono, 1990).
Penggerek batang padi putih (PBPP) menyerang tanaman padi pada semua
3
menyebabkan kerusakan tanaman @ di jalur pantai utara Jawa, sehingga penduduk di daerah itu menderita kelaparan. Teknik pengendalian dengan pengunduran masa
tanam di awal musim hujan, yang ditemukan oleh van der Goot (1925) efektif
mengendalikan ngengat yang muncul dari larva diapause. Namun pada permulaan tahun 1990 PBPP mengganas kembali di jalur pantai utara Jawa Barat, yaitu menye-
rang sekitar 75 ribu hektar dan menyebabkan kehilangan produksi padi sekitar 40%
atau senilai 60 milyar rupiah
(Oka,
1991; Priyanto dan Soenajo. 1992). Diperkirakanhama itu di masa-masa yang akan datang masih merupakan ancaman terhadap
kelestarian produksi padi di jalur pantai utara Jawa Barat.
Salah satu ciri usaha pertanian intensif adalah penggunaan pupuk yang cukup
banyak. Tiga unsur hara malcro yang sering digunakan dalam program intensifikasi
tanaman padi adalah nitrogen (N), fosfor (P), dan kalium (K). Pemberian ketiga
macam pupuk i& cenderung semakin meningkat.
Pupuk N dan P diketahui mempunyai pengaruh positif, sedang
K
diperkirakanmempunyai pengaruh negatif terhadap perkembangan penggerek batang padi.
Tanaman p d i yang dipupukN, jaringannya menjadi lebih sukulen dan lebih banyak
mengandung protein. Larva penggerek batang padi bergaris Chilo suppressalis dan
penggerek batang padi kuning Scirpophaga incertulas yang memakan tanaman itu
perkembangannya lebih baik dari pada tanaman yang tidak dipupuk. Larva lebih berhasil menggerek, bobot larva dan daya bertahan hidup larva lebih tinggi, masa
banyak (Israel, 1967; ~ u n a k a t a dan Okamoto, 1967; Man-, 1975).
Pemupukan N dosis tinggi dapat meningkatkan perkembangan penggerek
batang padi bergaris, dan dapat meningkatkan jumlah telur yang diletakkan sarnpai tujuh kali daripada yang tidak dipupuk N (Ishii, 1964; Pathak, 1968), sedang pemupukan P tidak berpengaruh nyata terhadap perkembangan larva penggerek batang pad^ (Wirano clan Ishii, 1959), tetapi berpengaruh baik terhadap serapan N oleh
tanaman padi (De Datta, 1981).
Berbeda dengan pengaruh pemberian pupuk N dan P, pemberian pupuk K yang cukup tinggi diperkirakan &pat melindungi tanaman padi dari serangan hama
dan penyakit, termasuk serangga penggerek batang padi (Soepardi, 1991). Keter-
sediaan hara
K
di lapisan olah tanah sawah sangat beragam. Hasil pemetaan status Ktanah sawah di Jawa dan Bali menunjukkan bahwa kandungan K terekstraksi atau tersedia berkisar dari sangat rendah (17 ppm K) sampai sangat tinggi (675 ppm
K)
(metode ekstraksi arnonium asetat 1 N) (Adiningsih, 1984; Soepartini et al., 1990).
LembagaPenelitian Tanah (1977) membagi status
K
tanah sawah terekstraksi(metode ekstraksi HC125%) atau tersedia ke dalam lima kelas yaitu 0
-
10, >10-
20, > 20-
40, >40-
60 dan > 60 mgK,O/
100 g tanah atau 0-
80,80-
160,160-
320,320
-
480, dan > 480 ppmK,
berturut-turut sebagai kandungan K tersedia sangatrendah, rendah, sedang, tinggi
dan
sangat tinggi. Status K tanah sawah di Jawa padatahun 1990 adalah 13.4% (489 524 ha) pada tingkat sangat rendah, 25.4% (964 374
5
ha) sangat tinggi (Soelaartini et al., 1990).
Serangan PBPP di daerah persawahan jalur pantai utara Jawa mulai musim
tanam 198911990 meliputi Kabupaten Bekasi, Karawang, Indramayu, Subang dan
Cirebon. Pemetaan status K yang dilakukan oleh Soepartini dan Widati (1 995) pada
keempat daerah persawahan itu menunjukkan bahwa K tersedia rendah masing-
masing dengan luasan 28 960, 41 690,2 638 dan 4 600 hektar (total seluas 78 ribu
hektar atau 16.83%); sedang dengan luasan 29 358, 19 919,47 456,25 594 dan
19 461 hektar (total seluas 141 788 hektar atau 30.63%), dan tinggi dengan luasan
28 475,46 541,39 540,89 561,
dan
39 096 hektar (total seluas 243 213 hektar atau52.54%). Pemberian pupuk
K
sebanyak 150 kg K,O per hektar sawah di sampingpupuk N dan P dapat meningkatkan produksi padi dari 4.6 menjadi 5.3 ton gabah
kering giling (Partohardjono et al., 1983)
Penelitian pengaruh K terhadap serangan PBPP telah dilakukan oleh van der Goot (1925) tetapi tidak dilaporkan hasihya Sehingga sampai saat ini belum tersedia
data yang cukup mengenai pengaruh ham K terhadap pertumbuhan dan perkernbang-
an penggerek batang padi putih (PBPP). Maka untuk itu perlu dilakukan penelitian
secara mendalam.
Rumusan Masalah
6
serapan K oleh tanaqan tergantung pada kandungan K tersedia bagi tanaman.
Semakin banyak kandungan K tersedia semakin banyak pula jumlah K yang diserap
oleh tanaman. Kecendemgan ini ia sebut sebagai konsumsi berlebih ( l m u y
conrumption). Menurut Grimme (1985) K diserap padi sawah lebih banyak dari N bahkan dapat mencapai lebih dari dua kali jumlah serapan N.
Kalium adalah unsur hara makro esensial bagi semuajenis tanaman. Di ddam sel-sel jaringan tanaman, K berfungsi sebagai aktivator enzim-enzim dalam reaksi
sintetik, meningkatkan permeabilitas rnembran sel, dan mengatur metabolisme air.
Dalam reaksi biosintetik, ion K di dalam jaringan tanaman berperan mening-
katkan sintesis gula dari senyawa anorganik (air dan asam arang) dalam proses
fotosintesis, gula sederhana menjadi polisakarida, sintesis asam amino, sintesis asam-
asam amino menjadi polipeptida atau protein, sintesis lignin dan selulose (Kock dan
Mengel, 1974; Kemler, 1980; Marschner, 1986).
Peningkatan kandungan K total dalam jaringan tanaman padi dapat mening-
katkan serapan silikat (Takijima et al., 1959; Mubekti, 1980). Tanaman padi yang
ditanam pda tanah yang dipupuk K dengan dosis berbeda diperkirakan akan meng-
hasilkan jaringan tanaman dengan kandungan silika total berbeda.
Dari dua macam peranan K di atas diketahui bahwa ion K yang diserap tanaman padi berpengaruh terhadap kandungan fitokimia yaitu gula pereduksi, gula
7
itu. Kandungan senyawa fitokimia dalam fase pertumbuhan vegetatif dan fase per-
tumbuhan generatif diperkirakan be*. Perbedaan kandungan senyawa fitokimia
tersebut itu diduga akan mempengaruhl pertumbuhan dan perkembangan PBPP yang memakan tanaman pa& tersebut. Oleh sebab itu, pengaruh makanan berupa tanaman
pa& yang berumur be* dan yang tumbuh pada tanah yang diberi pupuk K berbe-
da terhadap pertumbuhan dan perkembangan PBPP perlu dipelajari dengan seksama.
Selain itu ion K di dalarn jaringan tanaman dapat meningkatkan sintesis lignin
dan selulose, serta meningkatkan serapan silikat. Ketiga senyawa kimia itu dapat
mempengaruhi kekerasan jaringan tanaman padi. Pada kekerasan tertentu diper-
kirakan tanaman padi sulit digerek oleh larva PBPP. Dengan demikian pengaruh
pupuk K terhadap kekerasan jaringan tanaman padi perlu dipelajari.
Pada metabolisme air tanaman, K berperan &lam meningkatkan serapan air
melalui akar tanaman dan menghambat hilangnya air melalui transpirasi. Dalam
penghambatan hilangnya air melalui transpirasi, K berperan dalam meningkatkan
turgor sel penutup stomata. Pada kondisi tanaman kekurangan air, sel penutup akan menutup stomata (Kock dan Mengel, 1974; Kemler, 1980).
Kahat K, khususnya pada tanah media tumbuh yang kekurangan air menga-
kibatkan tidak terterkendalinya kehilangan air melalui transpimi. Pada kondisi demi-
kian terjadi p e n m a n kandungan air jaringan tanaman dan penurunan kandungan air
ABA dalam jaringan tanaman padi yang diakibatkan oleh kekuranp air dan
K
padatanah media tumbuhnya terhadap perkembangan
PBPP
belum diketahui dan perlu diteliti.Tujuan Penelitian
Penelitian bertujuan memahami perhmbuhan dan perkernbangan penggerek batang padi putih Scirpophaga innotata Walker yang makan dan hidup di dalam
tanaman padi yang ditamm pada tanah dengan kandungan K tersedia dalam kisaran
sangat rendah sarnpai tinggi.
Hipoteais
Tanaman padi yang diberi pupuk K dengan dosis berbeda
akan
mempunyaipengaruh berbeda terhadap perhmbuhan dan perkembangan penggerek batang @
TlNJAUAN PUSTAKA
Kalium
Kallum merupakan unsur hara yang sangat penting bagi tanaman. Di dalam tanah. K terdapt dalam bentuk langsung, cepat, lambat dan sangat lambat tersedia
tanaman, berturut-turut berupa K dalam larutan, K dapat ditukar,
K
terfiksasi,dan K dalam kisi-kisi mineral. Dua bentuk yang pertama disebut K segera tersedia
yang mudah diserap oleh tanaman. Dua bentuk yang terakhir juga &pat diserap
tanaman setelah berubah menjadi K tersedia, yang disebut bentuk cadangan
(Wiklander, 1954; Soepartini, 1995). Pemberian pupuk K &pat meningkatkan K tersedia dalam tanah (Tisdale dan Nelson, 1971).
Kalium diserap secara terus menerus oleh tanaman padi sampai akhu pertwnbuhan, dan tidak ditmnslokasi dari bagian vegetatif ke bagian panikel; sehingga
kaudungan K dalam jaringan batang lebih tinggi dibanding dalam biji (Tisdale dan
Nelson, 1971). Untuk pertumbuhan optimal, tanaman memerlukan K berkisar antara
dua sampai lirna persen dari berat keringnya (Marschnec, 1986). Kandungan K total
dalam batang padl tertinggi adalah 3.80% dari berat kering (De-Yin
dan
Bao, 1985).Kalium dan Fungsinya dalam Tanaman
Kalium adalah unsur hara makro esensial ketiga bagi tanaman setelah
tersedia dalam tanah semakin banyak pula jumlah K yang dapat diserap oleh
tanaman. Kecenderungan ini mereka sebut sebagai konsumsi berlebih (Iwrury
consumption) sebab peningkatan serapan melebihi jurnlah yang diperlukan untuk
produksi optimalnya.
Kalium yang diserap oleh tamman terdapt sebagai ion K+dalam cairan sel,
yaitu di dalam sitoplasma, vakuola dan inti sel. Unsur itu tidak menrpakan komponen bahan organik seperti N, P atau S, tetapi peranannya bersifat fungsional yang
penting dalam metabolisme. Peranan fungsional K dalam tanaman adalah mening- katkan serapan air dari tanah media tumbuh ke dalam jaringan tanaman melalui akar dan sebagai aktivator enzim dalam metabolisme primer maupun sekunder, mengatur
turgor sel dan transfer energi kimia.
Air yang telah diserap ke dalam jaringan tanaman berperan sebagai bahan
baku fotosintesis, pelarut dan media pengangkut hara mineral yang telah masuk ke
dalam akar menuju sel-sel jaringan, medium bagi berlangsungnya reaksi-reaksi
metabolisme, dan senyawa utama pembentuk protoplasma (Lehninger, 1982)
Terdapat lebih dari 40 jenis enzim metabolisme tanaman yang memerlukan ion K sebagai aktivatomya (Marschner, 1986). Enzimenzim teaebut berperan dalam proses biosintesis yang meliputi sintesis gula dalam proses fotosintesis, sintesis
glukosa menjadi p t i , sintesis protein, selulosa dan lignin (Evans dan Sorger, 1966).
Pada proses fotosintesis diperlukan bahan baku berupa air clan enzim ribulosa
bifosfat (RuBP) karboksilase, amilum sintetase, aldolase dan ATP (adenosin
dan berperan dalam mehngkatkan aktivitas keempat enzim itu (Bomer dan Varner, 1976; Marschner, 1986). Enzim RuBP-kabksilase sangat penting dalam biosintesis
dari bahan anorgatuk (CO, dan air) menjadi bahan organik 3-fosfogliseraldehid (3-
PGA) di dalam stroma dalam kloroplas. Enzim aldolase diperlukan dalam siklus
Calvin. Molekul3-PGA selanjutnya digunakan sebagai bahan sintesis karbohidrat
yang meliputi glukosa, sukrosa, amilum, selulosa dan lignin (Lehninger, 1982).
Dalam proses fotosintesis, selain te jadi proses sintesis bahan organik juga
te jadi proses transfer energi &ri energi sinar matahari menjadi bahan yang mengan-
dung energi kimia Bahan itu merupakan sumber energi utarna dalam metabolisme
setiap tanaman maupun hewan. Supaya dapat digunakan dalam proses metabolisme,
bahan tersebut umumnya diubah lebih dahulu ke dalam senyawa adenosin trifosfat
(ATP) (Lehninger, 1982) Dalam transfer energi, ion K diperlukan dalam sintesis
ATP baik pada proses fosforilasi fotosintetik dalam klorofil maupun pada proses
fosforilasi respirasi daIam mitokondria (Mengel dan Kirkby, 1982).
Pada tahap selanjutnya karbohidrat dari hasil proses fotosintesis digunakan
dalam metabolisme primer dan sekunder. Dalam metabolisme primer, karbohidrat itu digunakan untuk sintesis protein dan lemak, dan dalam metabolisme sekunder
digunakan untuk sintesis honnon tanaman (auksin, sitokinin, giberelin, asam absisat
atau ABA dan etilen), pigmen tanaman (antosianin, violaxantin, klorofil, sianofil),
1982; Marschner, 1986).
Sebelum digunakan untuk sintesis asam amino, protein dan Al3.4, karbohidrat mengalami proses glikolisis dan selanjutnya sebagian dari hasil proses glikolisis masuk
ke dalam siklus Kreb.
Pada
proses glikolisis, ion K diperlukan &lam kofaktor enzim6-fosfofruktokinase, piruvat kinase, dan asetil koenzim-A (Asetil-CoA) sintetase. Enzim 6-fosfofruktokinase adalah enzim yang berperan &lam sintesis fruktosa 6-
fosfat, enzim piruvat kinase berperan dalam sintesis asam piruvat,
dan
enzim asetil-CoA sintetase berperan dalam reaksi asam piruvat menjadi asetil-CoA (Borner dan
Vamer, 1976; Marschner, 1986). Berikutnya sebagian asetil-CoA digunakan dalam siklus Kreb, sebagian masuk ke dalam lintasan mevalonat, dan sebagian lainnya masuk ke lintasan malonat. Dalam siklus Kreb, ion K diperlukan sebagai aktivator enzim suksinil koenzim-A sintetase, yaituenzim yang berperan &lam sintesis asam a-ketoglutamt menjadi suksinil-CoA (Borner dan Varner, 1976; Marschner. 1986).
Pada sintesis protein, K diperlukan dalam meningkatkan sintesis protein dalarn ribosoma. Selain itu
K
diperlukan dalam sintesis enzim nitrat reduktase dan sebagaiaktivator enzim tersebut. Enzim nitrat reduktase beperan dalam reaksi reduksi nitrat menjadi amonium, dan kemudian senyawa amonium merupakan donor gugus amina
mengakibatkan terhambawa sintesis protein, yang ditandai dengan akumulasi
senyawa N berbobot molekul rendah, seperti asam amino bebas, amida-amida d m
nitrat (Kock dan Mengel, 1974).
Mengel dan Kirkby (1982) menyatakan bahwa asam-asam amino dan gula
pereduksi yang diakumulasi dalam jaringan tanaman yang tumbuh pada tanah kahat
K, adalah sebagai akibat langsung dari menurunnya aktivitas enzim. Hal itu dapat
terjadi karena suplai energi dari ATP tidak cukup. Mereka menerangkan bahwa kekurangan energi secara tidak langsung dapat mempengaruhi aktivitas enzim, di
antaranya adalah menunda sintesis protein Misalnya pada enzim nitrat reduktase dan
RuBP karboksilase.
Sebagian asetil-CoA yang masuk ke dalam lintasan mevalonat digunakan
sebagai bahan pembentuk senyawa terpenoid (monoterpenoid, seskuiterpenoid,
diterpenoid dan triterpenoid). Salah satu senyawa seskuiterpenoid adalah ABA
(Harborne, 1982; Vickery dan Vickery,l981).
Pada proses metabolisme sekunder, K meningkatkan aktivitas enzim selulosa
sintetase clan lignin sintetase dalam sintesis lignin, selulosa, fenolat dan menghambat
pembentukan asam absisat (ABA) (Kemler, 1971; Boyer, 1972; Kock dan Mengel,
1974; Saxena, 1985; Marschner, 1986).
Selain yang disebut di atas peningkatan kandungan K di dalam jaringan
tanaman dapat meningkatkan serapan silika, tetapi mekanismenya belum diketahui
(Kock dan Mengel, 1974; Mubekti, 1980). Kemampuan tanaman padi menyerap
14
(1976) kandungan silika tersedia dalam lapisan olah tanah yang rendah (di bawah 90
ppm Si02) menyebabkan tanaman padi yang tumbuh pada tanah itu mengandung
silika total rendah, sedang tanah dengan kandungan silika tersedia tinggi (di atas 130
ppm SiO,) menyebabkan tanaman padi yang tumbuh pada tanah itu mengandung
silika total tinggi.
Peranan K dalam jaringan tanaman yang telah diuraikan di atas adalah
meningkatkan serapan air dari air tanah ke dalarn jaringan tanaman, sintesis karbo-
hidrat dari bahan anorganik (H,O dan
Cod,
sintesis asam amino penyusun proteindan sintesis protein dalam metabolisme primer. Dalam metabolisme sekunder, K
berperan meningkatkan sintesis senyawa lignin, selulosa, fenolat dan menghambat
sintesis ABA. Selain itu dapat meningkatkan serapan silika.
Kahat K mengakibatkan akumulasi gula larut atau gula sederhana, sintesis pati
dan glikogea sangat terhambat, akumulasi asam amino, sintesis protein terhambat,
pemanfaatan substrat respiratori menjadi lambat, laju fosforilasi fotosintetik dan
oksidatif menurun (Heinrich, 1989).
Hubungan Serangga Herbivora den Tanaman yang Diberi Dosis Pupuk Kalium Berbeda
Serangga PBPP adalah serangga monofag, yang hanya mempunyai satu jenis
tanaman inang yaitu tanaman padi. Tanaman inang mempunyai kandungan fitokimia
yang mempunyai pengaruh terhadap pertumbuhan dan perkembangan sermgga
Pertumbuhan adalah proses peningkatan volume dan jumlah sel yang bersifat tidak dapat balik (rrreverslble). Secara sederhana pertumbuhan dapat diketahui dengan pertambahan bobot kering atau biomasa dalam rentang waktu tertentu.
Peningkatan biomasa adalah sebagai proses transformasi materi jaringan tanaman
sebagai makanan menjadi masa tubuh. Perkembangan melibatkan dua proses yang berbeda, yaitu pertumbuhan seperti yang telah diuraikan dan diferensiasi h g s i
organ. Diferensiasi adalah suatu proses pembahan pola aktivitas metabolisrne
(Sussman, 1960; Bursell, 1970).
Fitokimia adalah senyawa-senyawa kimia yang dihasilkan oleh metabolisme
tanaman baik metabolisme primer maupun metabolisme sekunder. Metabolit primer
adalah bahan nutrisi, sedang metabolit sekunder biasanya bekerja sebagai perangsang
atau penghambat makan, dan sebagian kecil saja yang mempunyai nilai nutrisi bagi
serangga herbivora (House 1965; Kogan, 1975; Vickery dan Vickery, 1981).
Bahan nutrisi bagi serangga herbivora adalah karbohidrat, protein, lemak,
vitamin dan air (Friend, 1958; House, 1965; Chapman 1971; Wigglesworth, 1972;
Scriber, 1984). Karbohidrat, protein, dan lemak adalah metabolit primer, dan
vitamin adalah metabolit sekunder (Vickery dan Vickery, 1981). Vitamin yang
tergolong senyawa sekunder itu adalah vitamin C (asam askorbat), E (a
-
tokoferol), K (filokuinon),p
-
karoten (prekursor vitamin A) dan ergosterol (prekursor vitaminD) (Ishii, 1964).
berbagai proses fisiolo&s. Protein merupakan senyawa nutrisi yang sangat penting
bagi tubuh, sebab selain sebagai surnber energi, protein berfimgsi sebagai senyawa
pembangun dan senyawa pengatur di dalam tubuh. Sebagai senyawa pembangum, fungsi utarnanya adalah untuk membentuk jaringan bam, misalnya pada proses embriogenesis pada pembentukan larva
dan
jaringan baru pada proses pertumbuhan larva (Chapman, 1976). Untuk menyusun protein dalam tubuh serangga diperlukanasam amino esensial (tidak dapat disintesis dalam tubuh serangga) dan asam amino non-esensial (dapat disintesis dalam tubuh serangga). Asam amino esensial terdiri
dari arginin, histidin, fenilalanin, metionin, treonin, lisin, isoleusin, leusin, triptofan
dan valin, dan asam amino non-esensial terdiri dari glisin, serin, sistin, tirosin, alanin,
glutamat, aspartat, sistein, prolin dan hidroksiprolin. Agar sintesis protein di dalam
tubuh berjalan lancar, asam amino esensial itu hasus ada dalam makanan dan apabila
semua asam amino esensial tidak ada dalam makanan menyebabkan larva p e n s r e k batang pad^ Chdo szppressalrs yang memakannya tidak d a p t tumbuh dan akan mati (Ishii, 1964). Kebutuhan asam-asam amino esensial dalam nutrisi serangga peng-
gerek batang padi adalah identik dengan hewan mamalia, misalnya tikus (Lipke dan
Fraenkel, 1956; Friend, 1958; House, 1961). Protein dalarn makanan yang mudah
dicema atau dihidrolisis oleh enzim-enzim pencemaan, serta mengandung ~ ~ a m - a ~ ~ m
amino esensial yang lengkap serta dalam jUmlah seimbang, merupakan protein
yang bemilai nutrisi tinggi (Muktadi, 1993).
jaringan yang telah diderna melalui usus tengah, dan selanjutnya digunakan untuk
memenuhi kebutuhan energi dan bahan penyusun tubuh.
Menurut Djamin dan Pathak (1967) kemampuan larva menggerek dipe-
ngaruhi oleh keadaan fisik tanaman padi. Tanaman padi yang mengandung silika
rendah (kurang dari 9% Si0J mudah digerek, sedang yang mengandung silika tinggi
(sekitar 13.9% Si0J sulit digerek dan gerigi mandibel larva yang menggerek tanaman
padi itu menjadi aus. Selanjutnya mereka menyatakan bahwa peningkatan kandungan
silika dalam jaringantanaman pad^ dapat menginduksi resistensi tanaman secam nyata
terhadap larva penggerek batang padi bergaris Chilo suppressalis Walker.
Tanaman padi merupakan tanaman silika, yaitu dapat menyerap silika dalam
jumlah banyak. Silika yang telah diserap oleh tanaman padi diendapkan di permu-
kaan tanaman dan di dalam jaringan tanaman. Silika yang diendapkan di permukaan
daun atau batang membentuk suatu polimer silika selulosa di antara lapisan kutikula
dan dinding sel epidermis bagian luar. Di dalam jaringan tanaman, silika diendapkan
dalam bentuk badan silika (srlica body) di dalam sel silika pada jaringan epidermis
Wchael, 1907; Soni et al., 1972).
Ketebalan dinding sel jaringan tanaman dapat menentukan derajat resistensi
beberapa Mtivar tanaman. Kultivar tebu dengan lapisan sel epidermis dan sklerenkirn
berdinding tebal sulit digerek oleh penggerek pucuk Scirpophoga nivela Linn. Batang
pad^ yang mempunyai dinding sel epidermis tebal dapat mencegah p e m a n oleh
keduanya dalam ~ m i t k 1989).
Pemberian pupuk K pada tanaman padi dengan dosis 162 kg KC1, (tidak
disebutkan berapa kandungan K tersedia dalam tanah media tumbuh), dapat mening-
katkan tebal dinding sel epidermis, p n k i m dan sklerenkim masing-masing dari
2.9 p menjadi 6.6 ,u, 2.0 ,u menjadi 4.2 F, dan 3.2 11 menjadi 6.8 ,u. Dalam
peningkatan tebal dinding sel, K berfungsi dalam sintesis lignin clan selulose. Kedua bahan itu adalah komponen pembentuk ran* dinding sel sekunder (Black, 1968;
Meyer dan Anderson, 1959; Steward dan Krikorian, 1971). Dinding sel epidermis
tanaman padi yang tebal dan permukaan sebelah luarnya mengandkg endapan
silikat dapat berfimgsi sebagai perisai dalam melindungi diri dari m g a n hama dan
penyakit (Soni et al., 1972).
Pemberian pupuk KC1 dalam dosis tinggi (200 dan 250 kg per hektar)
menurunkan kerusakan tanaman padi oleh serangan wereng hijau NephorertLr sp., wereng coklat Nilaparvata lugem Still, serangga penggulung dam Cnaphalocrocis medinalis Guinee, dan penggerek batang padi kuning Scirpophaga incertulas Walker (Subramanian dm Balasubramanian, 1976; John dan Thomas, 1980 kedua-
nya dalam Heinrich, 1989).
Tanaman padi yang berhasil digerek jaringan yang digerigit digunakan
sebagai makanan, yang kemudian ditelan dan dicerna di &lam usus tengah atau
dengan pertolongan enzim protease, karbohidrat menjadi glukosa dengan
pertolongan a-amilase clan P-glukosidase, lemak dirombak menjadi asam lemak dan gliserol dengan pertolongan enzirn lipse (Chapman, 1971 ; Rockstein, 1978). Enzim
protease, lipase, amilase, invertase
dan
maltase, biasa terdapat di dalam saluranpencemaan jenis-jenis serangga yang termasuk ordo Lepidoptera.
Ishii (1964) meneliti aktivitas enzirn-enzim yang berfungsi dalam mmcerna
karbohidrat dalam usus tengah larva penggerek batang padi bergaris. C.
suppressalis. Ia menggunakan makanan buatan yang diberi perlakuan beberapa macam gula. Gula yang digunakan terdiri dari monosakarida, disakarida, oligo- sakanida dan polisakarida. Sebagai sumber monosakarida adalah manosa, galaktosa,
sorbosa, fiuktosa, dan glukosa, sebagai sumber disakarida adalah sukrosa, trehalosa,
rnaltosa, sebagai sumber oligosakarida adalah melezitosa, dan rafinosa, dan sebagai
sumber polisakarida adalah amilum dan glikogen Dari hasil penelitiannya diketahui bahwa larva C. suppressalis lebih banyak menggunakan gula sederhana fruktosa, glukosa dan sukrosa sebagai sumber karbohidrat daripada bentuk yang lain.
Kemudian ia menyatakan bahwa aktivitas enzim invertase dalam rnesenteron larva
itu sangat kuat, sedang enzim amilasenya lemah.
Asam-asam amino hasil pencemaan protein di dalam usus yang telah diserap
ke dalam t u b a serangga digunakan untuk sintesis protein. Asam-asam amino yang
telah berhasil diserap oleh usus, sebagian digunakan untuk bahan sintesis protein
struktural dan sebagian digunakan sebagai bahan sintesis protein fkgsional.
protoplasma dan organel-organel dalam protoplasma, memb~an sel dari berbagai jaringan, rangka luar tubuh serangga, sutera pelindung kepompong. Sebagai bahan
fungsional, digunakan untuk sintesis protein enzim, protein pengangkut, histon yaitu
protein yang melindungi DNA (asam dioksiribonukleat), asam nukleat (DNA dan
RNA), dan pelindung hormon. Selain itu asam-asam amino digunakan dalam sintesis protein kuning telur atau vitelogenin pada ngengat betina dan protein sel sperma pada ngengat jantan (Chapman, 1971; Rockstein, 1978).
Asam amino yang digunakan sebagai komponen juvenile hormone (JH I , 11,
dan JII) adalah metionin. Sebelurn digunakan dalam sintesis honnon itu, metionin lebih dahulu diaktifkan oleh adenosil trifosfat (ATP) membentuk S-adenosil
metionin (SAM), yang selanjutnya SAM berfhgsi sebagai donor gugus metil ester
(Mem dan Beroza, 1972; McCurry, 1972; Rockstein, 1978). Senyawa JH
mempakan hormon penting dalam pertumbuhan, JH I dan JH I1 adalah honnon
morfogenetik, sedang JH
III
adalah honnon gonadotrofik (Lanzrein et al., 1975 datum Rockstein, 1978)).Proses pembentukan telur atau oogenesis pada ngengat betina d i p e m oleh kandungan protein cadangan makanan yang terbentuk selama pradewasa. Dalam
proses oogenesis itu terjadi pembentukan protein kuning telur ( j I R ) atau vitelogenin
dari bahan protein dan lipid. Protein ini berasal dari protein dalam darah yang
dipengaruhi oleh kualitas makanan; apabila protein yang diperoleh pada saat
pertumbuhan sedlkit maka pembentukan kuning telur menjadi terhambat, dan telur
generasi berikutnyamenjadi rendah (Chapman, 1971; Wigglesworth, 1974; Rockstein,
1978). Pembentdan telur lebih cepat, jumlahnya lebih banyak dan masa peneluran
lebih lama pada ngengat yang mempunyai cadangan makanan dengan kandungan
protein tinggi (Wigglesworth, 1974).
Pada ngengat jantan, te rjadi proses spermatogenesis yaitu proses pemben-
tukan sel-sel sperma, yang dimulai &ri germ cell yang membelah diri menjadi
spermatogonia, clan kemudian spermatogonia akan membentuk sel-sel sperma.
Apabila protein yang diperlukan dalam pembentukan sperma itu tidak cukup dan
kualitasnya kurang baik menyebabkan spermatogonia membentuk kista. Kista adalah spermatogonia yang tidak dapat berkembang (Chapman, 1971; Wigglesworth, 1974).
Peranan K dalam sintesis karbohidrat di dalam jaringan tanaman inang adalah sebagai aktivator enzim dalam reaksi penggabungan gula sederhana menjadi
polisakarida, dan dalam sintesis protein adalah sebagai aktivator enzim dalam sintesis
asam amino dan penggabungan asam-asam amino menjadi protein. Tanaman yang
cukup menyerap K menyebabkan banyak molekul gula sederhana yang disintesis
menjadi oligo- atau polisakarida; asam-asam amino disintesis menjadi protein.
Sebaliknya tanaman yang mengalami kahat K menyebabkan banyak molekul gula
sederhana, asam-asam amino dan yang terakumulasi di dalam jaringan tanaman.
Akumulasi gula sederhana dan asam-asarn amino dapat tejadi karena proses sintesis
senyawa-senyawa itu menjadi oligosakarida atau polisakarida dan protein terhambat
( ~ e m l e r , 1971; Boyer, 1972; Kock dan Mengel, 1974; Saxena, 1985). Tanaman
22
pad^ yang jaringannya lebih banyak mengakumulasi gula sederhana dan asam-asam
amino bebas lebih cocok sebagai sumber makanan bagi Chtlo suppressalis (Ishii, 1964).
Peranan air di dalam tubuh serangga adalah sebagai medium bagi berlang- sungnya reaksi-reaksi metabolisme dan sebagai pelarut dan media pengangkut m i n d
(Chapman, 1971).
Peranan K daIam metabolisme sekunder di dalam jaringan tanaman di
antaranya adalah menghambat pembentukan asam absisat (ABA) apabila tanaman
menderita kekurangan air. Peranan ABA dalam metabolisme serangga herbivora
belum banyak diteliti.
Menurut Addicott (1983) senyawa sekunder ABA dan analognya yang
terkandung dalam tanaman inang merupakan senyawa penting dalam hubungan
tanaman inang dengan serangga pemakannya (herbivora). Senyawa ABA adalab
hormon penghambat pertumbuhan dalam tanaman inang, yang selanjutnya berpe- ngaruh terhadap sistem neuroendokrin serangga yang memakan tanaman tersebut.
Pengaruh senyawa ABA terhadap sistem neuroendokrin serangga herbivora itu
menyebabkan pertumbuhan dan reproduksinya terhambat
P&gkatan kandungan ABA dalam jaringan tanaman &pat mempengaruhi
kandungan metabolit lain, contohnya adalah menurunkan kandungan fitosterol.
Telah diketahui bahwawaserangga tidak dapat mensintesis rangka steroid dalam meta- bolismenya, maka fitosterol itu hams ada dalam makanannya (Slama et a / . , 1976; Rockstein, 1978). Penurunan kandungan sterol dalam tanaman inang dapat meng-
akibatkan titer ekdisteroid atau ekdison dalam darah serangga pemakannya menjadi
rendah, yang akan menghambat perkembangannya (Kaplain et al., 1980 dalam Addicott, 1983).
Senyawa ABA yang telah masuk ke dalam tubuh serangga berpengaruh
terhadap perkembangannya. Ada beberapa peneliti yang telah mernpelaj& pengaruh
ABA terhadap perkembangan serangga. Eidt dan Little (1970) mempelajari pengaruh
ABA terhadap perkembangan Tenebrio molitor, hasilnya menunjukkan bahwa ABA yang diinjeksikan ke dalam tubuh larva serangga itu mengakibatkan perkembangan
larva menjadi kepompong menjadi sangat lambat. Selanjutnya de Man ei al.(1981)
melaporkan bahwa senyawa ABA yang diinjeksikan ke dalam tubuh Sarcophaga bullata mempunyai efek menghambat produksi vitellogenin, yaitu senyawa protein spesifik pada serangga sebagai penyusun protein kuning telur. Selain itu mereka telah
membuktikan bahwa ABA mempunyai efek menghambat sintesis ekdison dalam tubuh
serangga tersebut.
Addicott (1983) melaporkan bahwa ABA yang diinjeksikan ke dalam tubuh
Sarcophaga argvrosioma disebabkan oleh tidak adanya ekdisteroid atau ekdison
dalam sirkulasi darah.
Titer ekdisteroid dalam darah yang rendah pada larva Galleria mellonella
Linn. rnerupakan hasil dari inhibisi sintesis ekdisteroid dalam kelenjar protoraks Titer
ekdison yang rendah dalam darah telah diketahui sebagai respons terhadap sintesis
PTTH (prothoracicoh~oprc hormone) yang rendah dalam otak atau disebabkan oleh
inhibisi pelepasan PTTH dari otak (Pytel et al.. 1993). Mereka juga menyatakan bahwa penghambat pelepasan PTTH ke &lam darah larva G. mellonella yang
berdiapause adalah titer
JH
yang tinggi dalam darah.Senyawa sekunder ABA itu segolongan dengan JH, yaitu termasuk golongan
seskuiterpenoid dari derivat terpenoid, dan keduanya terbentuk melalui lintasan
mevalonat. Senyawa sekunder terpenoid hanya terdapat pada tumbuhan Sperma- tophyta atau tanaman berbunga (Vickery dan Vickery, 1981). Tanaman padi
termasuk di dalam spermatophyta tersebut.
Menurut sejarah pembentukannya atau berdasarkan hssil penelitian paleon-
tologi, senyawa terpenoid diketahui belum terdapat dalam tumbuhan sampai 200 juta
tahun yang laly dan terdapatnya senyawa itu bertepatan dengan munculnya tanaman berbunga clan kehidupan serangga modem (Harbome, 1982). Dalam pedalanan koevolusi antara serangga herbivora dan tumbuhan inangnya, yaitu pada awal perkembangan serangga oligofagi atau serangga modem diketahui bahwa terpenoid dalam tumbuhan yang ikut termakan serangga dapat digunakan untuk bahan sintesis
Kandungan ABA dalam jaringan tanaman padi sangat tergantung pada
kandungan air dalam jaringan tanaman tersebut. Kandungan ABA itu meningkat
secara linier sesuai dengan penurunan kandungan air atau turgor tanaman @ dan
mempunyai korelasi sangat nyata (Henson, 1982 dalam Suge. 1984).
Ion K" dalam tanaman berperan dalam mengatur turgor sel-sel tanaman
khususnya pada sel-sel mu&. Pa& kondisi kahat K (kurang dari 0.9% berat kering
jaringan) dan tanah media tanam kekurangan air mengakibatkan turgor sel menjadi rendah. Sebaliknya ion K yang cukup tinggi (lebih dari 2% berat kering jaringan) mengakibatkan turgor sel terpelihara dengan baik (Mengel dan Kirkby, 1982).
Ion K dalam jaringan tanaman yang ikut tennakan oleh serangga berperan
sebagai aktivator enzim pada reaksi pelepasan energi, sintesis protein dan glikogen,
untuk mempertahankan tekanan osmotik dan memelihara keseimbangan asam dan
basa di dalam tubuh. Selain itu, K berperanan dalam transmisi impuls syaraf dan
menstimulasi kontraksi otot (Chapman, 1971; Lehninger, 1982).
Biologi Penggerek Batang Padi
Lama
Terdapat lima instar selama periode perkembangan larva PBPP. Rataan
periode tiap instar dalam tanaman padi berumur panjang di Laboratorium Bogor
berturut-turut adalah 5, 5, 5, 7 dan 9 hari. Laju perkembangan larva itu tidak
berbdda dengan yang ada di lapangan (van der Goot, 1925). Perkembangan instar
Vietnam Selatan dan Malaysia Timur lebih singkat yaitu antara 19 sampai 3 1 hari (Feakin, 197 1).
Menurut van der Goot (1925) instar larva PBPP &pat dibedakan berdasarkan
jumlah kroset, ukuran dan m atubuh sebagai berikut. Larva intar kesatu yang baru keluar dari telur berwama kelabu, dengan kepala dan pronotum b e m a hitam kecoklatan, panjang tubuh 2.0
-
2.4 mm dan lebar sekitar 0.45mm,
serta tungkaipalsu mempunyai kroset sembilan buah. Larva instar kedua, kepala dan pronotum
b e m a hitam mengkilap, warna tubuh sama dengan instar satu, panjang tubuh 3.5
mm, serta tungkai palsu mempunyai kroset 1 1-13 buah. Larva instar ketiga yang baru
berganti Wit, kepala dan pronotum berwarna coklat mu&, panjang tubuh 6.5 mm
dan lebar 1.0 mm, tungkai palsu mempunyai kroset 17 buah. Larva instar keempat awal, wama tubuh masih sama instar ketiga, panjang tubuhnya 13 mm, yang lebih tua
benvarna putih kekuningan, dan tungkai palsu mernpunyai h s e t 30 - 3 1 buah. Larva
instar kelima, berwama putih kekuningan dengan saluran darah dorsal membayang kehitaman, panjang tubuh 21-25 mm, dan tungkai palsu mempunyai kroset 29-32 buah.
Larva yang baru keluar dari telur menggerek di antam pelepah dam. Seringkali larva itu turun dengan benang sutera ke bawah dan ikut terbawa aliran air
sampai ke tanaman lain. pula stadium ini mortalitas larva sangat tinggi. Pada tanaman
persemaian dan tanaman mu& larva menggerek ke dalam tunas ke bawah dan merusak titik tumbuh, sehingga bagian atas tanaman mati yang ditunjuklcan dengan
pindah ke tanaman di dekatnya. Pada tanaman yang sudah bemalai larva langsung menggerek ke bagian batang atau melalui bagian atas pelepah daun ke tangkai malai
dan terus menuju ke bawah. Padi yang terserang itu kemudian menmjukkan gejala
beluk (white head). Dalam tanaman mud& untuk seluruh perkembangan seekor
larva membutuhkan enam tanaman padi sedang pada tanaman tua cukup satu batang
tanaman (van der Goot, 1925).
Diapause
Diapause adalah strategi serangga menekan pertumbuhan dan menunda
perkembangannya dalam sementara waktu mtuk menghadapi keadaan lingkungan
yang buruk atau tidak menguntungkan (Beck, 1968; Chapman, 1971).
Diapause dapat terjadi pada setiap tingkat perkembangan serangga, yaitu
telur, larva, kepompong, pra kepompong atau ngengat. Serangga berdiapause pada
salah satu dari kelima tingkat perkembangan itu, yaitu tingkat perkembangan yang
paling tahan terhadap kondisi lingkungan yang tidak menguntungkan yang akan
dihadapi (Andrewartha, 1952). Serangga PBPP berdiapause dalam bentuk larva
instar akhir (van der Goot, 1925).
Diapause serangga dapat digolongkan menjadi dua, yaitu diapause obligat
dan fakultatif Diapause bersifat obligat apabila setiap individu dari setiap generasi
mengalami diapause, dan diapause obligat itu adalah bagian dari sejarah hidupnya.
Jenis diapause ini terjadi pada serangga univoltin, yaitu serangga yang hanya mem-
tidak te rjadi pada setiap generasi. Jenis diapause ini dijumpai pada serangga yang
mempunyai lebih dari dua generasi per tahun atau multivoltin (Beck, 1968; Chapman, 1971). Berdasarkan penggolongan itu, diapause larva PBPP dapat dimasukkan ke dalam diapause fakultatif Menurut Raufet al.(1993) larva PBPP mempunyai enam generasi per tahun dan yang mengalami diapause di jalur pantai utara Jawa Barat
adalah generasi keenam.
Diapause larva PBPP dilaporkan pertama kali oleh Dammerman (1915) di
jalur pantai utara Jawa pada awal abad ini, ketika padi ditanam sekali dalam setahun
pada musim hujan. Ia menyebut diapause tersebut sebagai tidur musim kering
(droogteslaap), yaitu larva yang berada dalam batang padi di bagian bawah yang sudah mengering setelah panen tidak berkembang lebih lanjut, dan dalam keadaan
hidup sebagai larva selama musim kering (antara bulan Mei dan Oktober). Larva
tersebut akan berkembang menjadi ngengat pada awal musim hujan berikutnya.
Dammerman berpendapat bahwa diapause itu disebabkan oleh keadaan lingkungan
yang kering karena adanya (1) pengeringan sawah pada waktu akan panen; (2)
berhentinya hujan setelah masa panen; dan (3) pengeringan bekas tanaman setelah
panen.
Diapause PBPP diteliti secara mendalam oleh van der Goot. Hasil penelitian
,
van der Goot (1925) menunjukkan bahwa larva penggerek batang padi putih juga mengalami diapause, meski keadaan sekelilingnya b a d . Larva yang menggerek pada
batang pad yang belum membentuk malai tidak berdiapause, sedang pada padi yang
29
diapause Iarva PBPP sebagai "tidur pematangan" (rijpings stuap). Ia menyatakan
bahwa diapause PBPP &rangsang oleh perubahan kompsisi nutrisi tanaman selama
proses penuaan. Selain itu juga dilaporkan bahwa setelah instar keempat, larva yang
akan berdiapause masih mengalami tiga sampai empat kali ganti kulit sebelum
memasuki ruas batang padi paling bawah.
Ganti kulit pada larva yang mengalami diapause tidak biasa tejadi pada
serangga yang termasuk dalam famili Pyralidae (Yagi dan Fukaya, 1974; Chippen-
dale, 1977, 1978). Selain larva PBPP, juga dilaporkan terjadi pada larva Dialraeu
grundiosella Dyar yaitu mengalami tambahan ganti kulit sebanyak tiga kali selama
perkembangan diapausenya. Pada setiap proses pergantian kulit pada larva D.
grandiosella itu kelenjar korpora alata tetap &if memproduksi hormon juvenil (JH) untuk mempertahankan titer JH cukup tinggi agar larva yang telah berganti kulit tetap
dalam bentuk larva.
Larva PBPP yang berdiapause secara morfologi berbeda dengan larva yang
tidak berdiapause. Perkembangan awal Iarva diapause (instar keenam) mempunyai
ukuran tubuh lebih besar, warna tubuhnya lebih kuning dan saluran darah dorsal lebih
jelas. Rataan lebar kapsul kepala larva yang hidup pada tanaman padi IR64 untuk
instar l , 2 , 3 , 4, 5 (larva tidak berdiapause) berturut-turut adalah 0.25, 0.40.0.60,
0.85, 1.05 mm; dan instar 6,
f,
8 (larva berdiapause) berturut-turut adalah 1.3, 1.8dan 2.1 mm (Triwidodo, 1993). Pada waktu 52 hari setelah inokulasi, larva yang
berdiapause telah ditemukan dalam ruas batang padi paling bawah atau ruas kesatu
berdiapause itu tidak aktif bergerak dan membungkus tubuhnya dengan sutera
(Triwidodo, 1993). Laju metabolisme pada serangga yang sedang berdiapause lebih
lambat dari pada yang tidak berdiapause (Tauber et al., 1983). Serangga itu
mempunyai adaptasi struktural dan fisiologis, sehingga lebih tahan terhadap keke-
ringan selama diapause. Ada dua mekanisme pertahanan yang dipunyai serangga
&lam menghadapi kekeringan, yaitu ( 1) menurunkan permeabilitas kutikula dengan
melapisi lilin pada bagian permukaan luarnya, dan (2) mencegah hilangnya uap air
dari dalam tubuh dengan hibemakulum (Chapman 1971; Denlinger, 1986). Selain
itu larva penggerek batang padi yang berdiapause melengkapi dengan lipid kutikular
yang tersusun dari rantai asam kaprilat atau kaprat, yang gunanya untuk melindungi
din dari serangan fungi (Koidsumi, 1957).
Hasil percobaan van der Goot yang dilakukan di laboratorium pada tahun
1922 dan 1923 menunjukkan bahwa makin lama waktu diapause sebelurn te ja d i
pelembaban tanah maki