ANGGUN WIDYA NINGGAR. Introduksi Gen Toleran Genangan (Sub1) pada
Padi Varietas Ciherang. Dibimbing oleh DJAROT SASONGKO HAMI SENO
dan TRI JOKO SANTOSO.
(Sub1) on Ciherang Rice Varieties. Under the direction of DJAROT SASONGKO
HAMI SENO dan TRI JOKO SANTOSO.
PENDAHULUAN
Upaya untuk meningkatkan produksi padi terus dilakukan seiring dengan bertambahnya penduduk dan meningkatnya kebutuhan beras. Peningkatan produksi padi dapat dilakukan dengan memperbaiki produktivitas pada daerah-daerah dimana cekaman abiotik merupakan kendala utama dalam budidaya tanaman padi.
Adanya perubahan iklim yang tidak menentu menuntut terciptanya padi yang toleran terhadap cekaman abiotik, seperti kekeringan, salinitas tinggi, maupun genangan atau banjir. Hasil panen rendah dan resiko kegagalan tanam akibat banjir umum ditemui pada areal rawa pasang surut, rawa lebak, dan tepian sungai
.
Stres karenacekaman rendaman akibat banjir akan menyebabkan petani guram yang diperkirakan menempati 15 juta hektar di Asia Selatan dan Asia Tenggara setiap tahunnya (Septinigsih et al. 2009). Di beberapa tempat, petani menanam padi yang toleran rendaman tetapi memiliki hasil yang rendah. Sementara ditempat lain, memiliki hasil yang tinggi tetapi tidak toleran terhadap rendaman. Baru-baru ini, cekaman rendaman meningkat secara ekstrim ketika terjadi hujan yang deras di beberapa tempat di Asia.
Di Indonesia potensi areal persawahan yang terkena areal banjir sekitar 13.3 juta ha terdiri atas 4.2 juta ha genangan dangkal, 6.1 juta ha genangan sedang, dan 3.0 juta genangan dalam (Nugroho et al. 1993). Luas areal pertanaman padi yang mengalami cekaman rendaman karena banjir diperkirakan akan semakin bertambah karena terjadi peningkatan curah hujan dan kenaikan permukaan air laut akibat terjadinya pemanasan global (CGIAR 2006). Solusi yang berkelanjutan dan permanen diperlukan untuk menanggulangi masalah ini. Salah satu solusinya adalah mengembangkan varietas dengan hasil tinggi yang toleran terhadap rendaman.
Penelitian ini dilakukan dalam rangka mengembangan padi Ciherang toleran genangan secara spesifik tanpa harus melalui metode rekayasa genetik. Perakitan varietas dilakukan secara persilangan terarah (site-directed crossing) untuk menghindari produk transgenik (Xu et al. 2004, Mackill et al. 2007). Regulasi GMO (genetically modified organisms) yang ketat perlu diantisipasi dengan menghindari metode-metode pengembangan yang menghasilkan produk tanaman transgenik.
Sebagai tetua pemulih (host) digunakan Ciherang. Padi Ciherang merupakan padi varietas unggul nasional yang telah memiliki beberapa kelebihan dibanding padi lainnya, seperti produktivitas yang tinggi, taham hama dan penyakit serta waktu tanam yang lebih pendek, dibandingkan padi IR64 dan Swarna yang merupakan padi varietas asli tahan genangan. Sebagai tetua donor digunakan padi BC2F1 Ciherang/IR64-Sub1 dan BC2F1 Ciherang/Swarna-Sub1 yang membawa gen toleransi genangan Submergence-1(Sub1) dan padi varietas lokal populer Ciherang sebagai tetua pemulih (host) akan disilangbalikkan (backcross) hingga dihasilkan tanaman BC3F1 yang mengandung gen Sub1. Analisis molekuler berbasis Polymerase Chain Reaction (PCR) marka Sub1 akan digunakan untuk melacak keberadaan alel gen Sub1.
Penelitian ini bertujuan memperoleh tanaman BC3F1 hasil persilangan balik BC2F1-Ciherang dan menyeleksi tanaman BC3F1 yang membawa gen Sub1 dengan menggunakan PCR
.
Hasil dari penelitian ini diharapkan akan didapatkan tanaman padi unggul dan komersial yang toleran genangan nontransgenik melalui metode persilangan terarah (site-directed crossing). Manfaat jangka panjang penelitian ini yaitu meningkatkan produktivitas tanaman padi secara berkelanjutan untuk menjaga kondisi ketahanan pangan nasional.TINJAUAN PUSTAKA
Tanaman Padi Toleran Genangan
Padi (Oryza sativa L.) diklasifikasikan dalam Divisi Spermatophyta, Sub Divisi Angiospermae, Class Monocotyledone, Ordo Poales/Glumiflorae, Famili Graminae, Genus Oryza, dan Spesies Oryza sativa (Tjitrosoepomo 1923). Secara umum padi dapat tumbuh di daerah tropis/subtropis pada 45° LU sampai 45° LS dengan curah hujan yang baik adalah 200 mm/bulan atau 1500-2000 mm/tahun. Di dataran rendah padi tumbuh pada ketinggian 0-650 m dpl dengan temperatur 22-27 °C sedangkan di dataran tinggi 650-1500 meter dpl dengan temperatur 19-23 °C (Siregar 1981).
PENDAHULUAN
Upaya untuk meningkatkan produksi padi terus dilakukan seiring dengan bertambahnya penduduk dan meningkatnya kebutuhan beras. Peningkatan produksi padi dapat dilakukan dengan memperbaiki produktivitas pada daerah-daerah dimana cekaman abiotik merupakan kendala utama dalam budidaya tanaman padi.
Adanya perubahan iklim yang tidak menentu menuntut terciptanya padi yang toleran terhadap cekaman abiotik, seperti kekeringan, salinitas tinggi, maupun genangan atau banjir. Hasil panen rendah dan resiko kegagalan tanam akibat banjir umum ditemui pada areal rawa pasang surut, rawa lebak, dan tepian sungai
.
Stres karenacekaman rendaman akibat banjir akan menyebabkan petani guram yang diperkirakan menempati 15 juta hektar di Asia Selatan dan Asia Tenggara setiap tahunnya (Septinigsih et al. 2009). Di beberapa tempat, petani menanam padi yang toleran rendaman tetapi memiliki hasil yang rendah. Sementara ditempat lain, memiliki hasil yang tinggi tetapi tidak toleran terhadap rendaman. Baru-baru ini, cekaman rendaman meningkat secara ekstrim ketika terjadi hujan yang deras di beberapa tempat di Asia.
Di Indonesia potensi areal persawahan yang terkena areal banjir sekitar 13.3 juta ha terdiri atas 4.2 juta ha genangan dangkal, 6.1 juta ha genangan sedang, dan 3.0 juta genangan dalam (Nugroho et al. 1993). Luas areal pertanaman padi yang mengalami cekaman rendaman karena banjir diperkirakan akan semakin bertambah karena terjadi peningkatan curah hujan dan kenaikan permukaan air laut akibat terjadinya pemanasan global (CGIAR 2006). Solusi yang berkelanjutan dan permanen diperlukan untuk menanggulangi masalah ini. Salah satu solusinya adalah mengembangkan varietas dengan hasil tinggi yang toleran terhadap rendaman.
Penelitian ini dilakukan dalam rangka mengembangan padi Ciherang toleran genangan secara spesifik tanpa harus melalui metode rekayasa genetik. Perakitan varietas dilakukan secara persilangan terarah (site-directed crossing) untuk menghindari produk transgenik (Xu et al. 2004, Mackill et al. 2007). Regulasi GMO (genetically modified organisms) yang ketat perlu diantisipasi dengan menghindari metode-metode pengembangan yang menghasilkan produk tanaman transgenik.
Sebagai tetua pemulih (host) digunakan Ciherang. Padi Ciherang merupakan padi varietas unggul nasional yang telah memiliki beberapa kelebihan dibanding padi lainnya, seperti produktivitas yang tinggi, taham hama dan penyakit serta waktu tanam yang lebih pendek, dibandingkan padi IR64 dan Swarna yang merupakan padi varietas asli tahan genangan. Sebagai tetua donor digunakan padi BC2F1 Ciherang/IR64-Sub1 dan BC2F1 Ciherang/Swarna-Sub1 yang membawa gen toleransi genangan Submergence-1(Sub1) dan padi varietas lokal populer Ciherang sebagai tetua pemulih (host) akan disilangbalikkan (backcross) hingga dihasilkan tanaman BC3F1 yang mengandung gen Sub1. Analisis molekuler berbasis Polymerase Chain Reaction (PCR) marka Sub1 akan digunakan untuk melacak keberadaan alel gen Sub1.
Penelitian ini bertujuan memperoleh tanaman BC3F1 hasil persilangan balik BC2F1-Ciherang dan menyeleksi tanaman BC3F1 yang membawa gen Sub1 dengan menggunakan PCR
.
Hasil dari penelitian ini diharapkan akan didapatkan tanaman padi unggul dan komersial yang toleran genangan nontransgenik melalui metode persilangan terarah (site-directed crossing). Manfaat jangka panjang penelitian ini yaitu meningkatkan produktivitas tanaman padi secara berkelanjutan untuk menjaga kondisi ketahanan pangan nasional.TINJAUAN PUSTAKA
Tanaman Padi Toleran Genangan
Padi (Oryza sativa L.) diklasifikasikan dalam Divisi Spermatophyta, Sub Divisi Angiospermae, Class Monocotyledone, Ordo Poales/Glumiflorae, Famili Graminae, Genus Oryza, dan Spesies Oryza sativa (Tjitrosoepomo 1923). Secara umum padi dapat tumbuh di daerah tropis/subtropis pada 45° LU sampai 45° LS dengan curah hujan yang baik adalah 200 mm/bulan atau 1500-2000 mm/tahun. Di dataran rendah padi tumbuh pada ketinggian 0-650 m dpl dengan temperatur 22-27 °C sedangkan di dataran tinggi 650-1500 meter dpl dengan temperatur 19-23 °C (Siregar 1981).
dan siap untuk dipanen hasilnya ± 25 hari setelah penyerbukan dan tergantung varietas. Umur padi antar varietas beragam, rata-rata umur padi 100-150 HST (Siregar 1981). Dari sekian banyak spesies padi, Oryza sativa L merupakan salah satu spesies yang dibudidayakan di Asia sedangkan Oryza glaberrima steund adalah salah satu yang dibudidayakan di Afrika (Manurung & Ismunadi 1999).
Padi jenis Ciherang merupakan kelompok padi sawah varietas unggul hasil beberapa kali persilangan. Padi jenis ini memiliki karakteristik umur tanamnya cukup singkat yaitu 116 hingga 125 hari, bentuk tanaman tegak, tingginya mencapai 107 hingga 115 cm, menghasilkan anakan produktif 14 hingga 17 batang, warna kaki hijau, warna batang hijau, warna daun hijau, posisi daun tegak, bentuk gajah panjang ramping, warna gabah kuning bersih, tekstur nasi pulen, rata-rata produksi 5 hingga 8.5 ton/ha, tahan terhadap bakteri hawar daun (HDB) strain III dan IV, tahan terhadap wereng coklat biotipe 2 dan 3. Padi Ciherang mulai diresmikan oleh menteri pertanian pada tahun 2000 dengan anjuran cocok ditanam pada musim hujan dan kemarau dengan ketinggian di bawah 500 meter di bawah permukaan laut (Hermanto 2006).
Tahun 2006 padi varietas IR64 oleh IRRI dikembangkan menjadi varietas padi toleransi genangan dengan mentransfer gen Sub1 dari varietas FR13A tahan genangan. Padi IR64-Sub1 (Inpara 5) memiliki karakteristik umur berbunga 83-86 hari, umur panen 112-116 hari, tinggi tanaman 90-95 cm, kadar amilosa 22 %, gabah isi per malai sebanyak 83 butir, tekstur nasi sedang, toleran terhadap rendaman penuh selama 14 hari, dan peka terhadap penyakit hawar daun bakteri (IRRI 2009). Varietas padi IR 64-Sub1 banyak digunakan di Asia terutama di Asia Tenggara termasuk Indonesia untuk berbagai penelitian. Padi varietas IR64-Sub1 ini juga telah digunakan petani di beberapa daerah yang rawan banjir untuk mengurangi resiko kegagalan panen pada saat terjadinya musim hujan akibat perubahan iklim yang tidak menentu (Septiningsih et al. 2009).
Padi Swarna-Sub1(Inpara 4) memiliki karakteristik umur berbunga 100-104 hari, umur panen 130-134 hari, tinggi tanaman 75-85 cm, kadar amilosa 27 %, gabah per malai sebanyak 131 butir, tekstur nasi pera, toleran terhadap rendaman penuh selama 14 hari, dan peka terhadap penyakit hawar daun bakteri (IRRI 2007). Pada tahun 2006-2007 varietas
ini telah diuji lebih lanjut oleh petani dan peneliti di International Rice Research Institute (IRRI) untuk digunakan dan dipublikasikan. Pengembangan varietas ini adalah contoh aplikasi ilmu pengetahuan molekular modern dengan memanfaatkan varietas lokal populer untuk sampai pada sebuah peningkatan produk yang toleransi terhadap genangan dan juga secara lokal dapat diterima.
Teknologi inovatif yang handal untuk meningkatkan produktivitas padi, baik melalui peningkatan potensi atau daya hasil tanaman maupun toleransi dan ketahanannya terhadap cekaman biotik maupun abiotik (Suprihatno et al. 2007). Berdasarkan dinamika masalah dan kendala produksi serta tuntutan konsumen, varietas-varietas unggul tersebut dikelompokkan menjadi dua, yaitu varietas yang diperuntukkan bagi peningkatan produktivitas yang melebihi ambang potensi hasil yang sudah melandai dan varietas unggul spesifik yang diperuntukkan bagi pencapaian stabilitas hasil (tahan/toleran cekaman biotik atau abiotik), peningkatan kualitas hasil (mutu rasa dan mutu gizi) serta umur genjah (Sunendar & Fagi 2000).
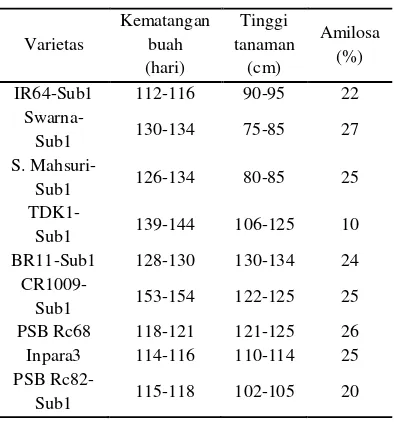
Beberapa padi toleran genangan telah teridentifikasi (Tabel 1), tetapi umumnya kemampuan kombinasi dan sifat agronominya (tanaman terlalu tinggi, sensitif penyakit dan hama serangga, produktivitas rendah) kurang memenuhi untuk kultivasi skala besar (Mohanty et al. 2000). Oleh karena itu sifat toleransi genangan perlu diintroduksi pada varietas padi populer yang
Tabel 1 Varietas padi tahan genangan (Mackill et al. 2011)
Varietas Kematangan buah (hari) Tinggi tanaman (cm) Amilosa (%) IR64-Sub1 112-116 90-95 22
Swarna-Sub1 130-134 75-85 27
S.
Mahsuri-Sub1 126-134 80-85 25
TDK1-Sub1 139-144 106-125 10 BR11-Sub1 128-130 130-134 24
CR1009-Sub1 153-154 122-125 25 PSB Rc68 118-121 121-125 26 Inpara3 114-116 110-114 25 PSB
lebih produktif. FR13A merupakan kultivar yang paling banyak digunakan sebagai sumber plasma nuftah dalam pengembangan varietas baru toleran genangan (Sarkar et al. 2006).
Aspek Fisiologis dan Morfologis Tanaman Padi terhadap Cekaman Rendaman
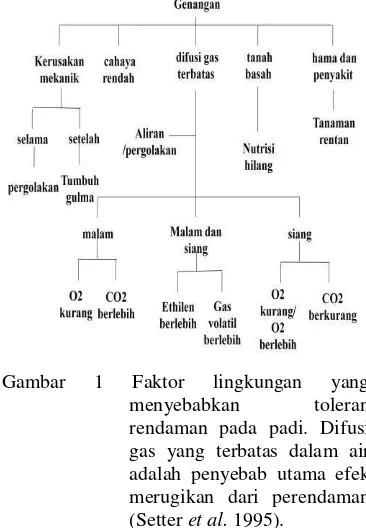
Ada dua perubahan lingkungan yang terjadi saat rendaman, yaitu aerobik ke anaerobik dan sebaliknya dari anaerobik ke aerobik setelah air berkurang. Faktor kunci untuk adaptasi dari aerobik ke anaerobik adalah suplai energi. Asimilasi karbon selama terendam akan dipengaruhi oleh beberapa faktor seperti suplai CO2, radiasi matahari,
kapasitas fotosintesis di bawah permukaan air yang dilemahkan oleh klorosis. Efisiensi penggunaan energi selama rendaman juga penting untuk adaptasi pada lingkungan anaerob (Kawano et al. 2008).
Sarkar et al. (2006) mengatakan bahwa toleransi rendaman merupakan adaptasi tanaman dalam merespon proses anaerob yang memampukan sel untuk mengatur atau memelihara keutuhannya sehingga tanaman mampu bertahan hidup dalam kondisi hipoksia tanpa kerusakan yang berarti. Sebuah evaluasi terhadap padi yang toleran dan tidak toleran menunjukkan bahwa bibit padi yang toleran memiliki 30-50% cadangan karbohidrat nonstruktural dibandingkan kultivar rentan. Karbohidrat ini dimanfaatkan selama terendam untuk mensuplai energi yang dibutuhkan untuk pertumbuhan dan mengatur metabolisme.
Tanah yang terendam air merupakan cekaman abiotik yang mempengaruhi komposisi spesies dan produktivitas pada berbagai tanaman. Pada tanaman padi, rendaman dimanipulasi sedemikian rupa untuk mendapatkan hasil yang maksimal. Beberapa spesies tanaman kelebihan air merupakan faktor penghambat produksi pada beberapa tempat dan situasi (Jackson 2004). Banjir terutama berpengaruh
terhadap hasil biji (Setter & Waters 2003). Hambatan utama yang disebabkan adanya rendaman pada spesies yang tidak bisa beradaptasi terhadap kekurangan oksigen adalah difusi oksigen di air lebih lambat 104 kali dibanding dengan di udara (Armstrong & Drew 2002). Hal lainnya adalah adanya perubahan level hormon etilen dan beberapa produk metabolisme anaerobik oleh mikroorganisme tanah seperti Mn2+, Fe2+, S2-, H2S dan asam karboksilat (Jackson & Colmer
2005). Lebih lanjut jika tanaman terendam
secara total akan mengakibatkan kekurangan karbondioksida, cahaya, dan oksigen sehingga dapat mengakibatkan kematian tanaman (Jackson & Ram 2003) (Gambar 1).
Pada saat tanaman padi tergenang, variasi konsentrasi O2 0.0-0.6 mol m–3
(kesetimbangan di udara 0.24 mol m–3 pada 30°C) dan CO2 0.004-0.020 mol m–
3
(kesetimbangan di udara 0.01 mol m–3) (Sarkar et al. 2006). Tanaman menderita kekurangan O2 parsial (hipoksia) atau tidak
mendapatkan sama sekali (anoksia) (Mohanty et al. 2000). Penurunan difusi gas ini mengakibatkan terhambatnya pertumbuhan, metabolisme, dan daya tahan tanaman (Mohanty et al. 2000, Sarkar et al. 2006). Berkurangnya suplai O2 menghambat
respirasi, kurangnya suplai CO2 menghambat
fotosintesis, dan terhambatnya difusi etilen mendorong klorosis dan perpanjangan daun berlebih pada kultivar yang intoleran. Klorosis adalah keadaan pada jaringan tanaman yaitu daun yang kekurangan klorofil sehingga tidak berwarna hijau melainkan kuning atau putih pucat (Jackson et al. 1987, Jackson & Ram 2003).
Sedangkan aspek morfologis padi adalah saat terjadi rendaman total sampai pada bagian daun paling atas, fotosintesis menjadi terhambat akibat kurangnya karbondioksida ekternal dan adanya semacam naungan (shading). Pada beberapa tanaman adanya etilen membuat stimulasi untuk memanjangkan batang (shoot elongation),
seperti pada padi, sehingga dapat melakukan escape dari cekaman rendaman. Salah satu mekanisme tanaman yang biasa hidup dalam keadaan terendam adalah memiliki jaringan aerankim (Seago et al. 2005). Aerenkim merupakan ruangan interselular yang terbentuk dari kombinasi pertumbuhan sel dan pembelahan sel (expansigeny). Proses masuknya gas dari atmosfer melalui aerenkim sebagian besar terjadi karena difusi. Namun demikian, aliran masa dapat pula terjadi jika alur jalan aerenkim membentuk tahanan yang rendah untuk dapat memasukan gas. Mommer dan Visser (2005) mengidentifikasi tampilan morfologi daun yang melakukan fotosintesis dibawah air. Penelitian ini menyimpulkan adanya hubungan kecepatan mengabsorbsi karbondioksida berhubungan erat dengan pembentukan lapisan kutikula dan lamina yang semakin menipis. Armstrong & Armstrong (2005) menggunakan external palarographic oxygen-sensing elektroda yang ditempatkan pada akar (rhizospher) untuk mengetahui proses keracunan sulfida pada akar tanaman padi. Mereka mengamati adanya reaksi reduksi dengan cepat terjadi karena berkurangnya oksigen di sekitar akar, sehingga mengakibatkan tekanan terhadap pemanjangan akar dan kemampuan mengambil air. Kondisi ini berlanjut sampai berhari-hari sulfida akan menghancurkan akar diikuti dengan tumbuhnya akar lateral. Sulfida juga dapat menghalangi aerenkim dan jaringan pembuluh tanaman. Kekurangan oksigen pada akar juga disebabkan oleh nitrat yang dibebaskan dari bahan organik sehingga menghalangi oksigen untuk diserap oleh akar (Kirk & Kronzucker 2005).
Gen Pengendali Toleransi Genangan
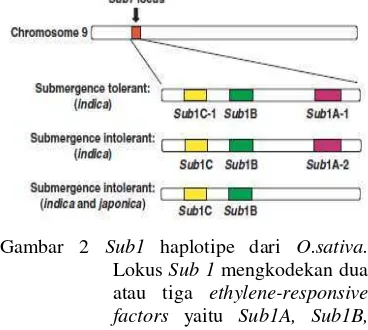
Saat tanaman tergenangi air, secara otomatis mereka memberikan respon untuk meningkatkan pertahanannya. Namun, jika terlalu lama tergenangi, maka tanaman akan layu dan mati. Padi juga tidak berbeda, meskipun padi ditanam dalam air, tanaman muda seringkali terpengaruh oleh banjir tahunan di lahan pertanian dataran rendah. Namun, beberapa kultivar sangat toleran serta dapat bertahan hidup sampai dua minggu penggenangan sempurna berkaitan dengan tempat percobaan kuantitatif utama yang ditunjuk sebagai Submergence 1 (Sub1) seperti pada FR13A (Xu et al. 2004). Lokus Sub1 terpetakan pada kromosom 9, berukuran 200 kb (Gambar 2), dan berperan dalam variasi toleransi genangan kultivar padi (Xu
Gambar 2 Sub1 haplotipe dari O.sativa. Lokus Sub 1 mengkodekan dua atau tiga ethylene-responsive factors yaitu Sub1A, Sub1B, dan Sub1C. Hanya lokus yang berisi alel Sub1A-1 yang dapat menyebabkan padi tahan terhadap rendaman (Fukao et al. 2006, Xu et al. 2006). & Mackill 1996, Xu et al. 2004, Perata & Voesenek 2006).
Penelitian lebih lanjut menunjukkan bahwa Sub1 pada FR13A mengkode tiga faktor transkripsi (Sub1A, Sub1B, dan Sub1C) (Gambar 2) yang termasuk kelompok B-2 subgrup ethylene response factor (ERFs) (Perata & Voesenek 2006). Regulasi transkripsi Sub1A dan Sub1C didapatkan meningkat akibat genangan. Peningkatan Sub1C berkurang dengan adanya Sub1A, mengindikasikan adanya represi Sub1C oleh Sub1A. ERF ketiga, Sub1B hanya sedikit terpengaruh oleh genangan (Fukao & Bailey-Serres 2008, Perata & Voesenek 2006).
Sub1A merupakan respon etilen mirip gen yang mengendalikan toleransi terhadap genangan pada padi (Xu et al. 2004). Sub1A yang pertama kali ditemukan merupakan suatu variabel tetapi dibutuhkan untuk toleransi terhadap genangan. Ketika terekspresi lebih dalam, gen Sub1A menyebabkan varietas padi toleran genangan di air. Suatu penyimpangan, Sub1A-1 hanya ditemukan dalam padi toleran genangan, sementara Sub1A-2 dengan suatu perubahan nukleotida tunggal merupakan versi ketidaktoleransi dari gen tersebut. Ketika dimasukkan kedalam varietas padi swarna tidak toleran genangan, yang tidak ada gen Sub1A, ilmuwan menemukan bahwa hasilnya tidak hanya toleran genangan air, tetapi juga produksi tinggi dan keuntungan lainnya.
al. 2004). Transformasi Sub1A-1 pada varietas Japonica intoleran genangan menghasilkan tanaman transgenik yang toleran genangan (Fukao & Bailey-Serres 2008, Perata & Voesenek 2006). Introgresi Sub1 (haplotipe Sub1A-1, Sub1B-1, Sub1C-1) pada intoleran kultivar japonica M202 mendapatkan tanaman yang lebih toleran terhadap genangan, lambat penurunan pati dan solubel karbohidratnya, kecil mRNA -amilase dan sukrosa sintasenya, tinggi aktivitas piruvat dekarboksilase (Pdc) dan alkohol dehidrogenase (Adh), kecil produksi etilennya, dan berkurang transkripsi gen ekspansinya (Fukao & Bailey-Serres 2008). Data-data fisiologi ini mendukung teori bahwa strategi survival terhadap genangan berlangsung melalui konservasi karbohidrat, represi elongasi sel, dan peningkatan kapasitas fermentasi (Perata & Voesenek 2006, Fukao & Bailey-Serres 2008).
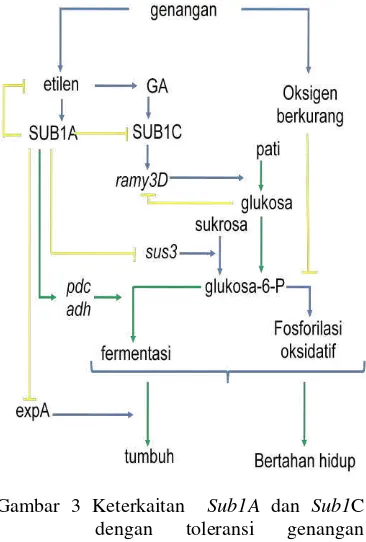
Keterkaitan Sub1 dalam survival tanaman terhadap genangan seperti disajikan pada Gambar 3 (Perata & Voesenek 2006, Fukao & Bailey-Serres 2008). Genangan mengakibatkan akumulasi etilen yang kemudian menginduksi transkripsi gen Sub1A sehingga terjadi akumulasi protein Sub1A. Selanjutnya Sub1A menghambat ekspansi A (ExpA) dan sukrosa sintase (Sus 3) sehingga menghambat pertumbuhan. Sub1A meningkatkan transkripsi gen yang berkaitan dengan fermentasi sehingga terjadi akumulasi mRNA dan peningkatan aktivitas Pdc dan Adh. Kondisi Fermentasi akan membuat glikolisis dapat berlanjut sehingga menghasilkan ATP untuk survival. Namun, laju produksi etanol tidak jauh berbeda dengan genotip yang tidak mengandung gen Sub1A, mengindikasikan bahwa induksi Pdc dan Adh tidak terlalu krusial. Sub1A menghambat gen yang berkaitan dengan elongasi sel dan katabolisme karbohidrat.
Degradasi pati menghasilkan sumber glukosa untuk glikolisis dan pertumbuhan. Padi yang mengekspresi Sub1A, laju elongasi rendah, pati dan karbohidrat yang terkumpul dapat digunakan untuk mempertahankan perlambatan sintesis ATP melalui fermentasi. Sub1C yang mengontrol gen -amilase (Ramy3D) dihambat oleh Sub1A. Gibberellins (GA) terlibat dalam regulasi ekspresi Sub1C. Namun, efek tersebut terhadap ekspresi Ramy3D bersifat tidak langsung, mengingat promotor gen ini yang tidak mengandung elemen GARE diperlukan untuk regulasi GA. Peningkatan regulasi Ramy3D oleh kandungan gula didapatkan
pada Sub1A-defisien. SUB1A juga bertanggung jawab pada restriksi feedback produksi etanol.
Gambar 3 Keterkaitan Sub1A dan Sub1C dengan toleransi genangan (Fukao et al. 2006, Perata & Voesenek 2007, Fukao & Bailley- Serres 2008).
Metode Analisis Toleransi Genangan
deteksi dengan metode PCR dengan marka Sub1.
Metode Persilangan Terarah (Site Directed
Crossing)
Tanaman transgenik akan dihambat dengan adanya regulasi GMO, sehingga sedapat mungkin menghindari metode-metode pengembangan yang menghasilkan produk tanaman transgenik. Metode site-directed crossing (Gambar 4) merupakan alternatif yang dapat digunakan untuk mengintroduksi sifat tertentu secara spesifik tanpa harus melalui rekayasa genetik yang menghasilkan produk tanaman transgenik (Xu et al. 2004, Mackill et al 2007).
Pada metode ini, introgresi donor dapat diminimalisasi, hanya sifat yang diinginkan yang terintroduksi pada host, sehingga dapat dipertahankan sifat-sifat yang baik pada padi host. Pembentukan populasi hingga BC5F1 akan menghasilkan turunan dengan sifat yang mendekati hampir 100 % host (retensi host maksimal) (Mackill et al. 2007), sehingga hanya sifat yang diinginkan (misalnya aromatik, toleransi genangan, dsb.) yang terintroduksi ke host. Selfing (pembentukan BC5F2) hanya diperlukan untuk introduksi sifat yang progeni gennya bersifat resesif (misalnya sifat aromatik), sedangkan untuk yang dominan (misalnya toleransi genangan) cukup hingga BC5F1.
Analisis molekuler (misalnya PCR) sangat diperlukan untuk memastikan bahwa tanaman yang akan dibackcross telah mengandung gen yang ditargetkan (Mackill et al. 2007) serta membantu mempersingkat tahapan penelitian. Pada introduksi toleransi genangan yang sifatnya dominan, dapat langsung dilakukan uji genangan, tetapi
Gambar 4 Diagram metode site-directed crossing. Keterangan: backcross = silang balik, selfing = silang dengan dirinya sendiri.
dengan adanya analisis molekuler akan lebih memastikan dan secara saintifik lebih valid. Metode site-directed crossing sering digunakan dengan nama marker- assisted backcrossing atau PCR-assisted backcrossing (Mackill et al. 2007, Lang & Buu 2008). Namun, umumnya tidak secara menyeluruh hingga turunan BC5F1 atau BC5F2, biasanya acak hingga BC3F1, BC2F2, dsb. Selain itu, belum pernah digunakan untuk introduksi lebih dari satu sifat.
Marka Molekular
Marka molekuler merupakan metode penunjuk keberadaan rangkaian nukleotida atau lebih umum dikenal pasangan basa (DNA) yang tidak dikenal sebagai suatu fungsional genetik. Marka dapat juga memberikan informasi suatu rangkaian sekuen tertentu dalam menyandikan suatu sifat atau memberikan informasi tentang keberadaan posisi suatu sekuen konservasi di dalam genom
Gupta et al. (2002) mengklasifikasikan marka molekuler kedalam beberapa generasi, diantaranya generasi pertama berdasarkan fragmen restriksi (Restriction Fragment Length Polymorphisms-RFLP) yang telah dilaporkan pada genom manusia pada awal 1980 (Botstein et al. 1980) menjadikan marka molekuler berbasis fragmen DNA sangat populer. Disusul dengan marka generasi kedua pada tahun 1990 yang meliputi mikrosatelit (Simple Sequence Repeats-SSRs) dan AFLPs (Amplified Fragment Length Polymorphisms) berbasiskan fingerprinting. Marka generasi ketiga pun muncul dengan pada tingkat yang lebih spesifik pada penyandi terkait ekspresi (Expressed Sequence Tags-ESTs) dan SNPs (Single Nucleotide Polymorphisms) diakhir 1990.
DNA yang dibutuhkan sangat tinggi serta dibutuhkan laboratorium khusus menangani radioaktif. RFLP merupakan sebuah teknik baru untuk memonitor transfer gen dari sebuah persilangan dan memiliki kemampuan untuk mengklon gen produk yang belum diketahui (Lang & Buu 2008).
Kedua, marka berdasarkan polymerase chain reaction (PCR) yang dikelompokan dalam satu primer pengamplifikasi; RAPD merupakan marka yang mengamplifikasi genom dengan satu primer spesifik secara acak (Williams et al. 1990) dan pasangan primer spesifik. STSs (Sequence-Tagged Sites) merupakan sekuen unik yang pendek yang mengidentifikasi satu atau lebih loci dan dapat teramplifikasi dengan PCR. SCARs (Sequence Characterized Amplified Regions); seperti STS primer yang mengidentifikasi RFLP loci. AFLP merupakan DNA fingerprinting yang berbasis pada amplifikasi PCR pada suatu set fragmen restriksi yang telah diligasikan suatu sekuen diketahui yang akan teramplifikasi biasanya menggunakan MseI atau EcoRI. AFLP ini mempunyai sifat kodominan sehingga dapat digunakan dalam studi tingkat polimorfis dan lebih efisien dibandingkan dengan teknik lainnya (Powell et al. 1996) dan SSRs yang merupakan DNA fingerprinting yang mempunyai tingkat kepercayaan lebih tinggi dibandingkan dengan penanda DNA berbasis fingerprinting.
Ketiga, marka berdasarkan PCR diteruskan hibridisasi. MP-PCR merupakan penggabungan beberapa teknik dalam proses PCR dengan menggunakan primer spesifik dan diteruskan dengan proses hibridisasi dengan probe radioaktif. Terakhir, marka berdasarkan hasil sequen seperti SNPs yang merupakan marka yang lebih spesifik pada perbedaan satu pasang basa nukleotida. Marka ini memerlukan sekuen lengkap tanaman dari suatu sekuen spesifik tertentu yang mempunyai suatu fungsi tertentu.
Setiap jenis marka mempunyai keuntungan dan kerugian dalam penggunaannya dan prosedur yang akan digunakan untuk beberapa tujuan. Faktor yang banyak mempengaruhi dalam pemilihan jenis marka antara lain kualitas dan kuantitas genom DNA, konsentrasi dan tidak adanya mispriming pada pasangan primer yang digunakan, konsentrasi pereaksi, jenis tanaman yang digunakan karena beberapa marka tergantung pada lokasi ekstraksi DNA tersebut dan pembuatan marka itu sendiri yang membutuhkan biaya, waktu dan tenaga
kerja yang komplikasi. Diantara marka molekular tersebut, SSR banyak dipakai karena sifatnya yang relatif praktis (PCR base), akurat, tingkat polimorfisme yang tinggi, memungkinkan multiplex (pengamatan beberapa marka sekaligus), dan sebarannya merata di seluruh bagian genom padi.
PCR dengan Marka Sub1
PCR adalah salah satu metode yang paling praktis untuk menunjukkan keberadaan gen pada suatu organisme. Analisis molekuler seperti PCR sangat diperlukan untuk memastikan bahwa tanaman yang akan disilangbalik (backcross) telah mengandung gen yang ditargetkan serta membantu mempersingkat tahapan penelitian (Mackill et al. 2007). PCR dengan marka Sub1 merupakan metode yang paling praktis dibandingkan dengan metode RAPD, RFLP, atau AFLP. Septiningsih et al. (2009) melaporkan bahwa ukuran/panjang gen padi toleran genangan (Sub1) berbeda dengan padi nontoleran genangan. Hal ini menyebabkan adanya pola pita yang berbeda pada visualisasi hasil PCR dengan elektroforesis gel agarosa. Panjang gen dan susunan gen yang berbeda antara padi toleran genangan dengan padi nontoleran genangan dapat membantu amplifikasi DNA dengan primer yang spesifik.
Marka-marka yang spesifik terhadap toleran genangan sudah banyak teridentifikasi (Septiningsih et al. 2009). Pada penelitian ini digunakan marka AEX1F dan AEX1R serta kombinasi antara marka RM219F dan AEX1R. Marka tersebut secara spesifik dirancang untuk alel toleran (IR40931). Marka AEX1F sebagai forward
dengan sekuen 5’
AGGCGGAGCTACGAGTACCA 3’.
Primer AEX1F berukuran 231 bp dengan nilai Tm sebesar 62.2ºC. Posisi dari primer ini SNP fungsional untuk Sub1A. Primer AEX1R merupakan primer reverse yang spesifik untuk gen Sub1 dengan nilai Tm sebesar 62.4ºC. Sekuen dari primer AEXR
adalah 5’ GCAGAGCGGCTGCGA.
sangat laborius untuk pengukuran dan memerlukan DNA dalam jumlah yang banyak atau berkualitas tinggi. Oleh karena itu, diperlukan marka PCR berbasis kodominan seperti marka mikrosatelit.
Salah satu marka mikrosatelit yang sering digunakan adalah SSR. Berdasarkan penelitian Xu et al. (2004) primer SSR yang ideal untuk seleksi toleran genangan adalah RM 219 dan RM 464A. Namun, pada penelitian kali ini hanya digunakan marka RM219F saja yang dikombinasikan dengan AEX1R. Seleksi Sub1 dengan marka mikrosatelit ini dapat dipercaya karena ikatannya sangat kuat (3.4 cM dan 0.7 cM). Primer forward dari marka RM219F
memiliki sekuen 5’
CGTCGGATGATGTAAAGCCT 3’ (Xu et
al. 2004). Meskipun primer tersebut telah terbukti pada berbagai analisis molekular varietas padi di berbagai negara, tetapi marka-marka tersebut belum pernah digunakan untuk varietas padi Indonesia.
BAHAN DAN METODE
Bahan dan Alat
Bahan tanaman padi diperoleh dari BB-BIOGEN dan LIPI. Tanaman yang digunakan yaitu host (Ciherang), padi BC2F1 Ciherang/IR64-Sub1, padi BC2F1 Ciherang/Swarna-Sub1, padi IR64-Sub1, padi Swarna-Sub1, dan padi IR42. Reagen untuk isolasi DNA yaitu etanol 70 %, etanol 95 %, bufer ekstraksi (NaCl 5 M, Tris-HCl 1 M, EDTA 0,5 M, lauril sarkosin, dan urea), fenol kloroform isoamilalkohol (PCI), isopropanol, dan larutan TE. Bahan-bahan yang digunakan untuk menguji hasil isolasi DNA dengan PCR adalah bufer PCR 10x, MgCl2 50 mM,
dNTP mix 10 mM, primer AEX1, primer RM219, Taq polymerase, sampel DNA 50 ng/ µL, dan MQ H2O. Bahan-bahan yang
digunakan untuk elektroforesis yaitu loading dye, bufer TAE 1x, agarosa, DNA hasil isolasi atau hasil PCR, marker, etidium bromida, dan akuabides.
Alat-alat yang digunakan untuk pembentukan benih BC3F1 adalah cawan petri, kertas saring, bak plastik volume 100 liter, ember, penyedot vakum, kertas minyak, klip, lampu listrik, dan gunting. Alat-alat yang digunakan untuk isolasi DNA adalah ruang laminar air flow, waterbath, cool box, pinset, tabung mikro, mortar, tip, vorteks, microfuge(Backman rotor 12), autopipet, dan inkubator. Alat-alat yang digunakan untuk elektroforesis adalah neraca analitik,
autoklaf, gelas piala, labu Erlenmeyer, magnetic stirrer, microwave, tangki elektroforesis, dan kertas aluminium. Alat-alat lain yang digunakan adalah spektrofotometer (SmartSpecTM Plus Spectrophotometer, Biorad), kuvet, pH meter, mesin PCR PTC-100 (MJ Research, Inc), dan UV illuminator Chemidoc EQ Biorad.
Metode Penelitian
Penanaman dan Persilangan Tanaman
Padi (Soedyanto et al. 1978)
Materi yang digunakan adalah varietas unggul Ciherang sebagai tetua pemulih, dan padi donor BC2F1 Ciherang/Swarna-Sub1 dan BC2F1 Ciherang/IR64-Sub1. Untuk menghasilkan biji BC3FI dilakukan penanaman butir padi dari masing-masing varietas dikecambahkan dalam cawan petri. Setelah berkecambah, dipilih 5 kecambah yang pertumbuhannya baik dan dipindahkan ke bak pembenihan. Setelah berumur 2 minggu, tanaman dipindahkan ke ember dan dipelihara sampai berbunga. Tanaman tetua betina Ciherang dan tetua jantan BC2F1 Ciherang-Sub1 yang pembungaannya bersamaan disilangkan untuk mendapatkan BC3FI. Sehari sebelum dilakukan persilangan, dilakukan kastrasi atau emaskulasi pada bunga-bunga tanaman tetua betina. Kastrasi adalah membuang bagian tanaman yang tidak diperlukan. Kastrasi dilakukan sehari sebelum penyerbukan agar putik menjadi masak sempurna saat penyerbukan sehingga keberhasilan penyilangan lebih tinggi. Setiap bunga terdapat enam benang sari dan dua kepala putik yang menyerupai rambut tidak boleh rusak. Bunga pada malai yang akan dikastrasi dijarangkan hingga tinggal 15-50 bunga. Sepertiga bagian bunga dipotong miring menggunakan gunting kemudian benang sari diambil dengan alat penyedot vacuum pump. Bunga yang telah bersih dari benang sari ditutup dengan glacine bag agar tidak terserbuki oleh tepung sari yang tidak dikehendaki.
sangat laborius untuk pengukuran dan memerlukan DNA dalam jumlah yang banyak atau berkualitas tinggi. Oleh karena itu, diperlukan marka PCR berbasis kodominan seperti marka mikrosatelit.
Salah satu marka mikrosatelit yang sering digunakan adalah SSR. Berdasarkan penelitian Xu et al. (2004) primer SSR yang ideal untuk seleksi toleran genangan adalah RM 219 dan RM 464A. Namun, pada penelitian kali ini hanya digunakan marka RM219F saja yang dikombinasikan dengan AEX1R. Seleksi Sub1 dengan marka mikrosatelit ini dapat dipercaya karena ikatannya sangat kuat (3.4 cM dan 0.7 cM). Primer forward dari marka RM219F
memiliki sekuen 5’
CGTCGGATGATGTAAAGCCT 3’ (Xu et
al. 2004). Meskipun primer tersebut telah terbukti pada berbagai analisis molekular varietas padi di berbagai negara, tetapi marka-marka tersebut belum pernah digunakan untuk varietas padi Indonesia.
BAHAN DAN METODE
Bahan dan Alat
Bahan tanaman padi diperoleh dari BB-BIOGEN dan LIPI. Tanaman yang digunakan yaitu host (Ciherang), padi BC2F1 Ciherang/IR64-Sub1, padi BC2F1 Ciherang/Swarna-Sub1, padi IR64-Sub1, padi Swarna-Sub1, dan padi IR42. Reagen untuk isolasi DNA yaitu etanol 70 %, etanol 95 %, bufer ekstraksi (NaCl 5 M, Tris-HCl 1 M, EDTA 0,5 M, lauril sarkosin, dan urea), fenol kloroform isoamilalkohol (PCI), isopropanol, dan larutan TE. Bahan-bahan yang digunakan untuk menguji hasil isolasi DNA dengan PCR adalah bufer PCR 10x, MgCl2 50 mM,
dNTP mix 10 mM, primer AEX1, primer RM219, Taq polymerase, sampel DNA 50 ng/ µL, dan MQ H2O. Bahan-bahan yang
digunakan untuk elektroforesis yaitu loading dye, bufer TAE 1x, agarosa, DNA hasil isolasi atau hasil PCR, marker, etidium bromida, dan akuabides.
Alat-alat yang digunakan untuk pembentukan benih BC3F1 adalah cawan petri, kertas saring, bak plastik volume 100 liter, ember, penyedot vakum, kertas minyak, klip, lampu listrik, dan gunting. Alat-alat yang digunakan untuk isolasi DNA adalah ruang laminar air flow, waterbath, cool box, pinset, tabung mikro, mortar, tip, vorteks, microfuge(Backman rotor 12), autopipet, dan inkubator. Alat-alat yang digunakan untuk elektroforesis adalah neraca analitik,
autoklaf, gelas piala, labu Erlenmeyer, magnetic stirrer, microwave, tangki elektroforesis, dan kertas aluminium. Alat-alat lain yang digunakan adalah spektrofotometer (SmartSpecTM Plus Spectrophotometer, Biorad), kuvet, pH meter, mesin PCR PTC-100 (MJ Research, Inc), dan UV illuminator Chemidoc EQ Biorad.
Metode Penelitian
Penanaman dan Persilangan Tanaman
Padi (Soedyanto et al. 1978)
Materi yang digunakan adalah varietas unggul Ciherang sebagai tetua pemulih, dan padi donor BC2F1 Ciherang/Swarna-Sub1 dan BC2F1 Ciherang/IR64-Sub1. Untuk menghasilkan biji BC3FI dilakukan penanaman butir padi dari masing-masing varietas dikecambahkan dalam cawan petri. Setelah berkecambah, dipilih 5 kecambah yang pertumbuhannya baik dan dipindahkan ke bak pembenihan. Setelah berumur 2 minggu, tanaman dipindahkan ke ember dan dipelihara sampai berbunga. Tanaman tetua betina Ciherang dan tetua jantan BC2F1 Ciherang-Sub1 yang pembungaannya bersamaan disilangkan untuk mendapatkan BC3FI. Sehari sebelum dilakukan persilangan, dilakukan kastrasi atau emaskulasi pada bunga-bunga tanaman tetua betina. Kastrasi adalah membuang bagian tanaman yang tidak diperlukan. Kastrasi dilakukan sehari sebelum penyerbukan agar putik menjadi masak sempurna saat penyerbukan sehingga keberhasilan penyilangan lebih tinggi. Setiap bunga terdapat enam benang sari dan dua kepala putik yang menyerupai rambut tidak boleh rusak. Bunga pada malai yang akan dikastrasi dijarangkan hingga tinggal 15-50 bunga. Sepertiga bagian bunga dipotong miring menggunakan gunting kemudian benang sari diambil dengan alat penyedot vacuum pump. Bunga yang telah bersih dari benang sari ditutup dengan glacine bag agar tidak terserbuki oleh tepung sari yang tidak dikehendaki.
jantan diletakkan di atasnya. Tanaman dipelihara sampai panen. Biji-biji yang telah masak dipanen dan dikeringkan dalam oven selama 1 malam.
Pengujian Toleransi genangan
(Septiningsih et al. 2009)
Pengujian ini dilakukan pada tanaman padi Ciherang, IR42, IR64-Sub1 dan Swarna-Sub1 dengan padi BC3FI Ciherang/IR64-Sub1 dan BC3FI Ciherang/Swarna-Ciherang/IR64-Sub1 mengikuti prosedur dari Septiningsih (2009). Benih sebanyak 10 biji per fenotipe (BC3FI) Ciherang/IR64-Sub1, Ciherang/Swarna-Sub1, tetua dari hasil persilangan tersebut masing-masing sebanyak 10 biji serta padi IR42 disemai di cawan petri selama 7 hari. Padi varietas IR42 biasanya digunakan sebagai kontrol toleran genangan karena sensitif terhadap genangan sehingga dapat memudahkan mengetahui batas uji genangan yang dilakukan. Setelah bibit berumur 7 hari (seminggu) kemudian dipindah tanam ke pot percobaan di Rumah Kaca. Setiap bak kecil ditanam satu bibit dalam satu lubang pada jarak tanam 5 cm x 5 cm dengan luasan pot 50cm x 25cm = 1.25 m2, sehingga jumlah tanaman per pot adalah 50 tanaman dan tanaman dibiarkan tumbuh sekitar 14 hari untuk siap dilakukan uji genangan.
Seleksi Sub1 dari hasil persilangan untuk uji genangan disejajarkan dengan tetua masing-masing varietas. Padi IR42 sebagai kontrol juga ditanam sejajar dalam satu pot kecil bersama dengan padi BC3FI hasil persilangan dan tetua. Setelah padi berumur 14 hari dalam pot maka dilakukan uji genangan dengan memasukkan pot yang berisi tanaman padi yang telah disejajarkan tersebut ke dalam bak (tray) yang besar. Kemudian bak yang sudah berisi tanaman padi tersebut diisi dengan air (digenangi) sampai keseluruhan padi tersebut terendam. Ketika kontrol IR42 sudah menunjukkan kerusakan >50% biasanya setelah 14 hari maka genangan air dalam bak tersebut dibuang. Setelah itu tanaman padi yang bertahan tersebut secara keseluruhan dibiarkan selama 10-21 hari untuk melakukan recovery. Kemudian tanaman padi yang akan recovery ditumbuhkan sampai daunnya siap untuk isolasi DNA.
Isolasi DNA Padi (Doyle dan Doyle 1987)
DNA genom total tanaman padi hasil persilangan Ciherang dengan BC2F1 Ciherang/IR64-Sub1 dan Ciherang/Swarna-Sub1 di isolasi dari daun (Doyle & Doyle
1987). Isolasi DNA dilakukan empat tahap yang meliputi pemanenan, preparasi ekstrak sel, pemurnian DNA, dan pemekatan DNA. Tanaman padi yang berumur 3 minggu dipanen dan dimasukkan dalam tabung mikro. Pemecahan sel dibantu dengan cara penggerusan dalam mortar dan ditambah 1000 µL bufer ekstraksi CTAB hingga homogen. Suspensi diinkubasi di dalam water bath selama 15 menit suhu 65 ºC (setiap 5 menit dikocok dengan cara tabung dibolak-balik secara perlahan). Pemurnian DNA dari pengotor dihilangkan dengan penambahan 100 µL fenol kloroform isoamilalkohol (PCI) ke dalam tabung dan dikocok selama 20 detik hingga merata. Suspensi selanjutnya disentrifugasi dengan kecepatan 12 000 rpm selama 10 menit.
Pemekatan DNA dilakukan dengan menambahkan 500 µL isopropanol ke supernatan dan dicampur selama 5 menit. Sampel divorteks dan disentrifugasi kembali pada kecepatan 12 000 rpm selama 5 menit. Pelet yang diperoleh dicuci dengan 500 µL etanol 70%. Campuran disentrifugasi kembali selama 5 menit pada kecepatan 12000 rpm. Pelet selanjutnya dikeringkan di oven suhu 60 0C selama 10 menit. Pelet yang telah kering dilarutkan dengan larutan TE yang mengandung ribonuklease sebanyak 50-100µL. Kemudian pelet yang telah dilarutkan dengan TE diinkubasi suhu 37 °C selama 30 menit.
Pengukuran Konsentrasi dan Kemurnian
DNA (Sambrook et al. 1989)
Konsentrasi DNA ditentukan dengan spektrofotometer pada λ 260 nm. Sedangkan kemurnian DNA diukur pada λ 260/280 nm. Sampel DNA sebanyak 2µL dilarutkan dalam buffer Tris-EDTA sebanyak 198 µL ke dalam kuvet sehingga volume akhirnya 2 mL. Angka yang muncul pada layar merupakan konsentrasi dari DNA sampel yang diukur dan dicatat. DNA yang sudah diukur konsentrasinya diencerkan 10x sehingga mendapatkan konsentrasi yang seragam untuk digunakan dalam analisis PCR. Kemurnian larutan DNA dapat dihitung dengan menghitung hasil OD 260 nm dibagi OD 280 nm. Hasil perbandingan antara 1.8-2.0 menunjukkan kemurnian yang tinggi.
Seleksi PCR dengan Marka AEX1
(Septiningsih et al. 2009)
mM, 1 µL masing-masing primer AEX1F ukuran 231 bp sebagai forward dengan
sekuen 5’ AGGCGGAGCTACGAGTACCA 3’ dan primer AEX1R sebagai reverse
dengan sekuen 5’ GCAGAGCGGCTGCGA
3’ (Septiningsih et al. 2009), 0.16 µL Taq
polymerase, dan 12.84 µL MQ H2O.
Kemudian digunakan program SUB I pada mesin PCR dengan kondisi denaturasi awal 94 0C selama 5 menit denaturasi 94 0C selama 1 menit, penempelan primer 55 0C selama 1 menit, perpanjangan primer 72 0C selama 2 menit, dan perpanjangan primer akhir 72 0C selama 5 menit.
Seleksi PCR dengan Marka RM219F dan AEX1R
Seleksi keberadaan gen Sub1 juga dilakukan dengan menggunakan marka mikrosatelit RM219F dan AEX1R dengan campuran reaksi untuk PCR terdiri atas 2 µL DNA 50 ng/ µL, 2 µL bufer PCR 10x, 0.6 µL MgCl2 50 mM, 0.4 µL dNTP mix 50 mM, 1
µL masing-masing primer RM219F sebagai forward dengan sekuen 5’
CGTCGGATGATGTAAAGCCT 3’ dan
primer AEX1R sebagai reverse dengan
sekuen 5’ GCAGAGCGGCTGCGA 3’
(Septiningsih et al. 2009), 0.16 µL Taq polymerase, dan 12.84 µL MQ H2O.
Kemudian dilakukan amplifikasi PCR seperti pada marka AEX1.
Elektroforesis Produk PCR (Septiningsih
et al. 2009)
Ukuran produk PCR selanjutnya dapat dianalisis dengan elekroforesis gel agarose. Sebanyak 1-2% gel agarosa dan 1x buffer TAE (Tris Acetic Acid EDTA) dimasukkan ke dalam cetakan. Setelah gel agarosa memadat kemudian dimasukkan ke dalam tangki elektroforesis yang berisi 1x buffer TAE. Sebanyak 10 µl produk PCR dari masing-masing sampel ditambahkan dengan 2 µl loading dye dan dicampur sempurna, kemudian dimasukkan ke dalam sumur di dalam gel. Ukuran dari produk PCR ditentukan dengan mnyertakan DNA standar (100 bp ladder) sebagai pembanding. Sampel dielektroforesis dengan tegangan 80 volt selama kurang lebih 1.5 jam. Setelah itu, gel agarose diwarnai dengan larutan etidium bromida (EtBr) (10 mg/l) selama 10 menit dan dicuci dengan air selama 10-20 menit. Gel agarose kemudian divisualisasi dengan Chemidoc gel system (Biorad).
HASIL DAN PEMBAHASAN
Pembentukan Benih dan Populasi BC3F1
Ciherang/Swarna-Sub1 dan BC3F1
Ciherang/IR64-Sub1
Pembentukan populasi BC3F1 Ciherang-Sub1 dari penelitian ini merupakan penggabungan sifat yang dimiliki dari dua jenis tetua yang berbeda. Proses pembentukan BC3F1 dilakukan dengan melakukan persilangan antara benang sari dan putik sehingga akan dihasilkan embrio yang akan berkembang menjadi benih. Penggabungan sifat yang dihasilkan dari kedua tetua pada BC3F1 terjadi secara acak, sehingga kombinasi sifat yang dihasilkan bersifat lebih menguntungkan dari kedua tetuanya (Welsh & James 1981). Keturunan yang dihasilkan akan memiliki sifat baru yang berbeda dengan kedua induknya. Hasil persilangan antara Ciherang (host) dengan BC2F1 (Ciherang/Swarna-Sub1 dan Ciherang/IR64-Sub1) akan menghasilkan komposisi gen 93,75%:6,25% yang artinya gen dari kedua tetua telah bersegregasi dalam tanaman BC3F1 dengan komposisi paling banyak gen Ciherang pada populasi BC3F1 (Gambar 5).
Dalam pembentukan benih BC3F1 Ciherang-Sub1 digunakan metode persilangan terarah (site-directed crossing). Pada metode ini, introgresi donor dapat diminimalisasi, hanya sifat yang diinginkan yang terintroduksi pada host, sehingga dapat dipertahankan sifat-sifat yang baik pada padi host. Metode ini lebih dikenal dengan nama marker assisted backcrossing (Lu & Chang 1980, Mackill et al. 2007). Seperti yang sudah dijelaskan sebelumnya, metode ini merupakan alternatif yang dapat digunakan untuk mengintroduksi sifat tertentu secara spesifik tanpa harus melalui rekayasa genetik yang menghasilkan produk tanaman transgenik (Xu et al. 2004, Mackill et al. 2007). Persilangan ini dimaksudkan agar tanaman BC3F1 Ciherang/Sub1 yang dihasilkan memiliki sifat penuh yang dimiliki padi Ciherang. Selanjutnya turunannya akan disilang balik lagi sampai generasi BC5F1 agar komposisi gen mendekati seratus persen kembali ke Ciherang.
mM, 1 µL masing-masing primer AEX1F ukuran 231 bp sebagai forward dengan
sekuen 5’ AGGCGGAGCTACGAGTACCA 3’ dan primer AEX1R sebagai reverse
dengan sekuen 5’ GCAGAGCGGCTGCGA
3’ (Septiningsih et al. 2009), 0.16 µL Taq
polymerase, dan 12.84 µL MQ H2O.
Kemudian digunakan program SUB I pada mesin PCR dengan kondisi denaturasi awal 94 0C selama 5 menit denaturasi 94 0C selama 1 menit, penempelan primer 55 0C selama 1 menit, perpanjangan primer 72 0C selama 2 menit, dan perpanjangan primer akhir 72 0C selama 5 menit.
Seleksi PCR dengan Marka RM219F dan AEX1R
Seleksi keberadaan gen Sub1 juga dilakukan dengan menggunakan marka mikrosatelit RM219F dan AEX1R dengan campuran reaksi untuk PCR terdiri atas 2 µL DNA 50 ng/ µL, 2 µL bufer PCR 10x, 0.6 µL MgCl2 50 mM, 0.4 µL dNTP mix 50 mM, 1
µL masing-masing primer RM219F sebagai forward dengan sekuen 5’
CGTCGGATGATGTAAAGCCT 3’ dan
primer AEX1R sebagai reverse dengan
sekuen 5’ GCAGAGCGGCTGCGA 3’
(Septiningsih et al. 2009), 0.16 µL Taq polymerase, dan 12.84 µL MQ H2O.
Kemudian dilakukan amplifikasi PCR seperti pada marka AEX1.
Elektroforesis Produk PCR (Septiningsih
et al. 2009)
Ukuran produk PCR selanjutnya dapat dianalisis dengan elekroforesis gel agarose. Sebanyak 1-2% gel agarosa dan 1x buffer TAE (Tris Acetic Acid EDTA) dimasukkan ke dalam cetakan. Setelah gel agarosa memadat kemudian dimasukkan ke dalam tangki elektroforesis yang berisi 1x buffer TAE. Sebanyak 10 µl produk PCR dari masing-masing sampel ditambahkan dengan 2 µl loading dye dan dicampur sempurna, kemudian dimasukkan ke dalam sumur di dalam gel. Ukuran dari produk PCR ditentukan dengan mnyertakan DNA standar (100 bp ladder) sebagai pembanding. Sampel dielektroforesis dengan tegangan 80 volt selama kurang lebih 1.5 jam. Setelah itu, gel agarose diwarnai dengan larutan etidium bromida (EtBr) (10 mg/l) selama 10 menit dan dicuci dengan air selama 10-20 menit. Gel agarose kemudian divisualisasi dengan Chemidoc gel system (Biorad).
HASIL DAN PEMBAHASAN
Pembentukan Benih dan Populasi BC3F1
Ciherang/Swarna-Sub1 dan BC3F1
Ciherang/IR64-Sub1
Pembentukan populasi BC3F1 Ciherang-Sub1 dari penelitian ini merupakan penggabungan sifat yang dimiliki dari dua jenis tetua yang berbeda. Proses pembentukan BC3F1 dilakukan dengan melakukan persilangan antara benang sari dan putik sehingga akan dihasilkan embrio yang akan berkembang menjadi benih. Penggabungan sifat yang dihasilkan dari kedua tetua pada BC3F1 terjadi secara acak, sehingga kombinasi sifat yang dihasilkan bersifat lebih menguntungkan dari kedua tetuanya (Welsh & James 1981). Keturunan yang dihasilkan akan memiliki sifat baru yang berbeda dengan kedua induknya. Hasil persilangan antara Ciherang (host) dengan BC2F1 (Ciherang/Swarna-Sub1 dan Ciherang/IR64-Sub1) akan menghasilkan komposisi gen 93,75%:6,25% yang artinya gen dari kedua tetua telah bersegregasi dalam tanaman BC3F1 dengan komposisi paling banyak gen Ciherang pada populasi BC3F1 (Gambar 5).
Dalam pembentukan benih BC3F1 Ciherang-Sub1 digunakan metode persilangan terarah (site-directed crossing). Pada metode ini, introgresi donor dapat diminimalisasi, hanya sifat yang diinginkan yang terintroduksi pada host, sehingga dapat dipertahankan sifat-sifat yang baik pada padi host. Metode ini lebih dikenal dengan nama marker assisted backcrossing (Lu & Chang 1980, Mackill et al. 2007). Seperti yang sudah dijelaskan sebelumnya, metode ini merupakan alternatif yang dapat digunakan untuk mengintroduksi sifat tertentu secara spesifik tanpa harus melalui rekayasa genetik yang menghasilkan produk tanaman transgenik (Xu et al. 2004, Mackill et al. 2007). Persilangan ini dimaksudkan agar tanaman BC3F1 Ciherang/Sub1 yang dihasilkan memiliki sifat penuh yang dimiliki padi Ciherang. Selanjutnya turunannya akan disilang balik lagi sampai generasi BC5F1 agar komposisi gen mendekati seratus persen kembali ke Ciherang.
Gambar 5 Diagram metode persilangan terarah (site directed crossing) .
dilakukan pengaturan dan penghitungan jarak waktu tanam agar kedua tetua dapat berbunga secara bersamaan. Persilangan antara Ciherang dengan BC2F1 Ciherang IR64-Sub1 terjadi lebih awal dengan masa berbunga ± 86 hari, sedangkan persilangan antara Ciherang dengan BC2F1 Ciherang/Swarna-Sub1 memiliki masa berbunga ± 104 hari.
Selanjutnya adalah kastrasi. Kastrasi atau emaskulasi adalah membuang bagian tanaman yang tidak diperlukan. Kegiatan ini biasa disebut dengan pengebirian. Kastrasi dilakukan sehari sebelum penyerbukan agar putik menjadi masak sempurna saat penyerbukan sehingga keberhasilan penyilangan lebih tinggi (Harahap 1982). Dua kepala putik yang menyerupai rambut tidak boleh rusak (UPLB 1967) sehingga perlu hati-hati dalam melakukan kastrasi. Bunga pada malai yang akan dikastrasi dijarangkan hingga tinggal 15-50 bunga (Harahap 1982). Sepertiga bagian bunga dipotong miring menggunakan gunting kemudian benang sari diambil dengan alat penyedot vacuum pump. Bunga yang telah bersih dari benang sari ditutup dengan glacine bag agar tidak terserbuki oleh tepung sari yang tidak dikehendaki.
Proses penyerbukan dilakukan dengan cara menyalakan semua lampu di ruang persilangan sejak pagi hari agar suhu ruangan meningkat untuk mempercepat pemasakan tepung sari (Soedyanto et al. 1978). Bunga yang sudah diserbuki segera ditutup dengan kantong kertas transparan atau glacine bag (Soedyanto et al. 1978). Tanaman hasil penyerbukan dipelihara di rumah kaca sampai biji hasil persilangan masak. Setelah 3-4 minggu, malai dipanen kemudian dikeringkan
dengan cara dijemur atau dioven (Sadjad 1993). Biji yang sudah kering dirontok. Benih yang dihasilakan dari persilangan memiliki bentuk ramping, panjang tanpa kulit luar. Benih-benih tersebut kemudian ditumbuhkan untuk menghasilkan tanaman utuh BC3F1 (Gambar 6).
Seleksi Uji Toleransi Genangan
Padi umumnya memang tahan terhadap rendaman air, seperti banjir berebihan dan lain sebagainya. Tanaman akan mati bila terlalu lama terendam. Uji toleran genangan yang dilakukan pada penelitian ini dimaksudkan untuk mengidentifikasi tanaman padi BC3F1 yang tahan terhadap rendaman (Septiningsih et al. 2009) dan sekaligus mempunyai gen Sub1 (Gambar 7). Pengujian ini tidak dapat membuktikan secara spesifik tanaman BC3F1 mengandung gen Sub1 setelah diintrogesi dari padi Ciherang dengan BC2F1 Ciherang-Sub1. Namun, uji ini dilakukan untuk melihat dan menyeleksi tanaman padi mana yang tahan terhadap rendaman.
Galur padi yang mendapat perlakuan uji toleransi genangan yaitu masing-masing tetua host (Ciherang) dan donor Sub1(Swarna-Sub1 dan IR64-Sub1(Swarna-Sub1), tanaman BC3F1 Ciherang/IR64-Sub1 dan BC3F1 Ciherang/Swarna-Sub1 serta tanaman IR42. Padi varietas IR42 yang digunakan dalam uji genangan berfungsi sebagai kontrol toleran genangan karena sensitif terhadap genangan sehingga dapat menjadi patokan batas uji genangan yang dilakukan (Septiningsih et al. 2009).
Pemberian air dalam bak dilakukan dengan menggunakan selang dengan kecepatan air disesuaikan agar tanaman padi tidak rusak. Selama 14 hari uji rendaman, tanaman dalam bak harus selalu diamati karena ketinggian air akan berkurang akibat penguapan dan harus ditambah lagi sampai ketinggian semula. Respon tanaman padi terhadap lingkungan yang seperti itu akan melakukan dua proses pertahanan, yaitu pemanjangan sel tanaman dan toleran terhadap cekaman rendaman tersebut (Fukao & Bailey-Serres 2008). Pemanjangan sel tanaman di bawah kondisi tergenang merupakan hal yang tidak menguntungkan. Hal ini karena tanaman akan menghabiskan cadangan makanan untuk pemanjangan sel tersebut, sedangkan pada kondisi tergenang tanaman harus bertahan hidup tanpa
memperoleh asupan nutrisi dari lingkungan (Setter et al. 1997).
Padi yang terlihat melakukan pemanjangan sel adalah Ciherang dan beberapa padi BC3F1. Setelah hasil pengujian toleransi genangan selama dua minggu, dilakukan pengurangan air dari dalam bak secara perlahan dan masih menyisakan sedikit air di dalam bak agar tanaman tidak stress. Kemudian tanaman dibiarkan sepuluh hari untuk dapat recovery (tumbuh normal kembali) dengan kondisi daun yang tidak terlalu tegak (Septiningsih et al. 2009). Gambar 8 memperlihatkan hasil recovery selama 10 hari. Hasil yang didapat dari uji genangan yaitu sebagian tanaman BC3F1 mampu bertahan hidup dan dari hasil recovery selama 10 hari terdapat 7 tanaman varietas Ciherang/IR64-Sub1 dan 6 tanaman galur Ciherang/Swarna-Sub1. Pengurangan jumlah tanaman BC3F1 yang hidup dikarenakan sebagian tanaman BC3F1 tidak
terintroduksi gen Sub1. Tanaman BC3F1 yang berhasil lolos dari uji genangan ditumbuhkan sampai cukup umur (± 3 minggu) untuk dilakukan isolasi dan pengujian molekuler berbasis PCR dengan marka spesifik Sub1 untuk mendeteksi keberadaan gen tersebut.
Hasil Isolasi dan Karakterisasi DNA
Hasil isolasi DNA diuji secara kualitatif maupun kuantitatif. Uji kualitatif dilakukan dengan melihat DNA hasil isolasi pada agarosa, sedangkan uji kuantitatif dilakukan dengan spektrofotometer yang meliputi konsentrasi dan kemurnian DNA pada perbandingan panjang gelombang 260/280 nm. Uji ini bermanfaat untuk menjamin tidak terdapat kontaminasi protein dan polisakarida pada DNA yang diisolasi.
Proses isolasi DNA pada dasarnya harus melalui tahapan penghancuran sel (lysis cell), pemisahan DNA dari komponen lain, pengendapan dan pemanenan DNA (Sambrook et al. 1989). Pada penelitian ini digunakan metode CTAB (Cetyl Trimethyl Ammonium Bromide) (Doyle & Doyle 1987). Keuntungan menggunakan metode ini adalah tidak membutuhkan waktu isolasi yang lama serta tahapan metode yang relatif lebih mudah dan cepat. Isolasi DNA dilakukan melalui empat tahap yang meliputi pemanenan, preparasi ekstrak sel, pemurnian DNA, dan pemekatan DNA. Selain itu, metode ini merupakan metode yang paling umum dilakukan karena konsentrasi DNA yang diperoleh relatif lebih banyak jika dibandingkan dengan metode yang lain (Lumaret et al. 1998; Jose & Usha 2000).
Ada tiga langkah utama dalam ekstraksi DNA, yaitu perusakan dinding sel (lisis), pemisahan DNA dari bahan padat seperti selulosa dan protein, serta pemurnian DNA (Nicholl 1993; Surzycki 2000). Daun yang diambil adalah daun yang muda sehingga memudahkan untuk penggerusan karena daun muda memiliki tekstur yang lebih lunak dan sedikit serat. Ekstrak sel dihasilkan melalui penggerusan dalam mortar dan secara kimia menggunakan buffer CTAB. CTAB merupakan detergen yang dapat melisiskan membran sel dan mampu mengendapkan polisakarida serta senyawa fenolik yang terdapat pada tanaman padi. Bufer CTAB mengandung Tris-Cl, EDTA, NaCl, CTAB, PVP, dan merkaptoetanol. Tris-Cl berfungsi untuk mendenaturasi protein. NaCl berfungsi sebagai bahan penetral pada gula fosfat DNA. EDTA berfungsi sebagai penghancur sel dengan cara mengikat ion magnesium yang diperlukan oleh sel untuk menjaga keutuhan selubung sel secara keseluruhan. Larutan CTAB, PVP dan merkaptoetanol pada bufer ekstraksi berfungsi untuk mendegradasi senyawa-senyawa metabolit sekunder yang terdapat dalam tanaman. Selain itu, CTAB berfungsi untuk memisahkan DNA dan RNA dari pengotor (karbohidrat dan protein) (Nicholl 1993).
DNA padi dimurnikan dengan senyawa kloroform isoamilalkohol dan asam asetat untuk mendenaturasi protein. RNAse yang berfungsi untuk melisiskan RNA. Presipitasi DNA menggunakan isopropanol yang berfungsi mengendapkan DNA dan memisahkan DNA dari garam-garam mineral, serta melarutkan sisa CTAB (Nicholl 1993)
Etanol 70% berfungsi untuk mencuci pelet yang dihasilkan pada proses presipitasi isopropanol.
Pemurnian tersebut bertujuan untuk menghilangkan senyawa-senyawa yang dapat menghambat reaksi enzimatis pada proses PCR. DNA hasil isolasi diuji kualitasnya dengan spektrofotometer. Panjang gelombang yang dipakai adalah 260/280. Tabel 2 menunjukkan hasil uji kualitas DNA yang diisolasi dengan menggunakan metode CTAB.
Data menunjukkan bahwa dari seluruh sampel yang dihitung konsentrasi kemurniannya, secara umum memiliki konsentrasi yang relatif besar. Hal ini menunjukkan bahwa jumlah DNA yang diperoleh dari hasil isolasi jumlahnya banyak. Nilai konsentrasi ini sangat berpengaruh terhadap analisis selanjutnya yaitu analisis PCR. Konsentrasi masing-masing sampel
Tabel 2 Pengukuran konsentrasi dan kemurnian DNA daun padi
Varietas padi Konsentrasi
(µg/ml) A260/280 Ciherang (Cih) 2733,7 1,87 Swarna-Sub1 4630,5 1,92 IR64-Sub1 2834,2 1,98
F1Cih/IR64-Sub1 1 3126,9 1,96
F1Cih/IR64-Sub1 2 4353,6 1,9
F1Cih/IR64-Sub1 3 3314 1,97
F1Cih/IR64-Sub1 4 3520,3 1,94
F1Cih/IR64-Sub1 5 5021,2 1,9
F1Cih/IR64-Sub1 6 2727,5 1,94
F1Cih/IR64-Sub1 7 1913,2 1,93
F1Cih/Swarna-Sub1 1 3307,1 1,95
F1Cih/Swarna-Sub1 2 2200,8 1,91
F1Cih/Swarna-Sub1 3 2835,2 1,89
F1Cih/Swarna-Sub1 4 5599,9 1,86
F1Cih/Swarna-Sub1 5 3959,7 1,94
F1Cih/Swarna-Sub1 6 2219,2 1,95
harus disamakan dalam analisis PCR agar jumlah DNA yang teramplifikasi relatif sama. Pengenceran dilakukan sampai 50 µg/ml.
Sedangkan kemurnian DNA dapat dilihat dari rasio absorbansi 260/280. Semua molekul dapat diabsorbsi pada panjang gelombang yang spesifik untuk molekul tersebut dapat diserap dan menghasilkan pengukuran yang akurat. Nukleotida dan RNA akan diserap pada panjang gelombang 260 nm, sedangkan protein, fenol, dan kontaminan yang lain dapat diserap kuat pada panjang gelombang mendekati 280 nm. Sehingga kemurnian DNA dapat dilihat pada rasio 260/280 nm (William et al. 1997). Data menunjukkan bahwa DNA yang diperoleh sangat murni karena nilainya berkisar antara 1.8-2.0. Menurut Sambrook dan Russel (1989), sampel DNA yang memiliki nilai dibawah 1.8 menunjukkan sampel DNA mengalami kontaminasi protein sedangkan nilai diatas 2.0 menunjukkan adanya kontaminasi oleh RNA.
Hasil Seleksi BC3F1 Ciherang-Sub1
dengan analisis PCR
Marka Sub1A (AEX1)
Setelah diuji genangan, tanaman BC3F1 harus diuji secara spesifik apakah sudah tersisipi gen Sub1 atau belum. Marka yang digunakan pada PCR ini adalah marka AEX1. Marka ini dapat mendeteksi adanya gen Sub1A pada padi. Menurut Xu et al (2004), Sub1A merupakan respon etilen mirip gen yang mengendalikan toleransi terhadap genangan pada padi. Sub1A yang pertama kali ditemukan merupakan suatu variabel. Namun, dibutuhkan untuk toleransi terhadap genangan. Ketika terekspresi lebih dalam, gen Sub1A menyebabkan varietas padi toleran genangan di air. Marka AEX1 merupakan pengembangan primer dengan SNP pada ujung 3’ nya. Marka AEX1F sebagai forward dengan sekuen 5’
AGGCGGAGCTACGAGTACCA 3’. Primer
AEX1F berukuran 231 bp. Primer AEX1R merupakan primer reverse yang spesifik untuk gen Sub1. Sekuen dari primer AEXR
adalah 5’ GCAGAGCGGCTGCGA.
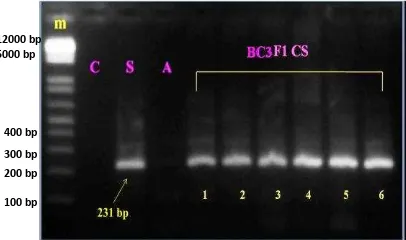
Hasil elektroforesis sampel DNA tanaman padi BC3F1 Ciherang/Swarna-Sub1 (Gambar 9) dan padi BC3F1 Ciherang/IR64-Sub1 (Gambar 10) memperlihatkan hasil yang positif keberadaan gen Sub1A. Hasil tersebut merupakan hasil PCR dengan visualisasi elektroforesis 2% sehingga terlihat jarak amplifikasi atau pita ukuran basa yang jelas antar spesies. Padi yang telah berhasil terintroduksi gen Sub1A memperlihatkan pita DNA pada ukuran 231 bp mengikuti tetua donornya masing-masing yaitu IR64-Sub1 dan Swarna-Sub1. Hasil ini sesuai dengan penelitian Septingsih et al. (2009). Dari hasil tersebut (Gambar 9 dan 10) terlihat bahwa padi yang digunakan sebagai kontrol negatif yaitu Ciherang tidak memperlihatkan adanya pita DNA sehingga terbukti bahwa Ciherang tidak toleran terhadap genangan karena tanaman tidak berhasil bertahan hidup pada waktu seleksi uji genangan. Sedangkan air yang digunakan untuk mendeteksi adanya kontaminan pada waktu elektroforesis juga tidak menunjukkan adanya pita sehingga hasil tersebut dinyatakan berhasil. Selanjutnya tanaman padi BC3F1 Ciherang-Sub1 yang telah positif mengandung gen Sub1 akan disilangbalikkan (backcross) kembali dengan padi Ciherang untuk menghasilkan tanaman BC4F1 Ciherang-Sub1.
Marka Gabungan RM219F dan AEX1R
Marka RM219 merupakan marka mikrosatelit (SSR atau Simple Sequence repeat) yang berupa urutan dinukleotida atau tetranukleotida yang berulang dan berurutan. Seperti yang dijelaskan sebelumnya, SSR
Gambar 9 Hasil elektroforesis padi BC3F1 Ciherang/IR64-Sub1 dengan marka AEX1F dan AEX1R. Padi Ciherang (C), donor IR64-Sub1 (S), sampel BC3F1 Ciherang-Sub1(F1 CS), dan Air (A) dengan marker (m) DNA 1 kb.
Gambar 10 Hasil elektroforesis padi BC3F1 Ciherang/Swarna-Sub1 dengan marka AEX1F dan AEX1R. Padi Ciherang (C), donor Swarna-Sub1 (S), sampel BC3F1 Ciherang-Sub1(F1 CS), dan Air (A) dengan marker (m) DNA 1 kb.
sering dipilih karena relatif praktis, efisien, dan memungkinkan pembuatan beberapa marka sekaligus dalam satu kali reaksi (multiplexing). Selain itu SSR juga cenderung bersifat polymorphic, codominant (mampu membedakan homozigot dan heterozigot), multi allelic serta akurat digunakan pada berbagai varietas padi (sebarannya merata di seluruh bagian genom padi) (McCouch et al. 2002).
Marka mikrosatelit RM219 merupakan marka alternatif yang digunakan untuk analisis kodominan padi BC3F1 Ciherang-Sub1. Analisis PCR dengan menggunakan marka mikrosatelit RM219 merupakan analisis lanjutan dari marka AEX1. Berdasarkan penelitian Marlina E (2011) dan Sinaga JR (2011) hasil amplifikasi PCR dengan marka RM219 dapat membedakan tetua Ciherang, Swarna-Sub1, IR64-Sub1, dan BC3F1 Ciherang-Sub1 dengan ukuran fragmen DNA yaitu 204 bp dan 194 bp. Akan tetapi, perbedaan fragmen DNA ini terlalu kecil hanya berkisar antar 5-10 bp sehingga dilakukan uji PCR dengan marka kombinasi RM219F dan AEX1R.
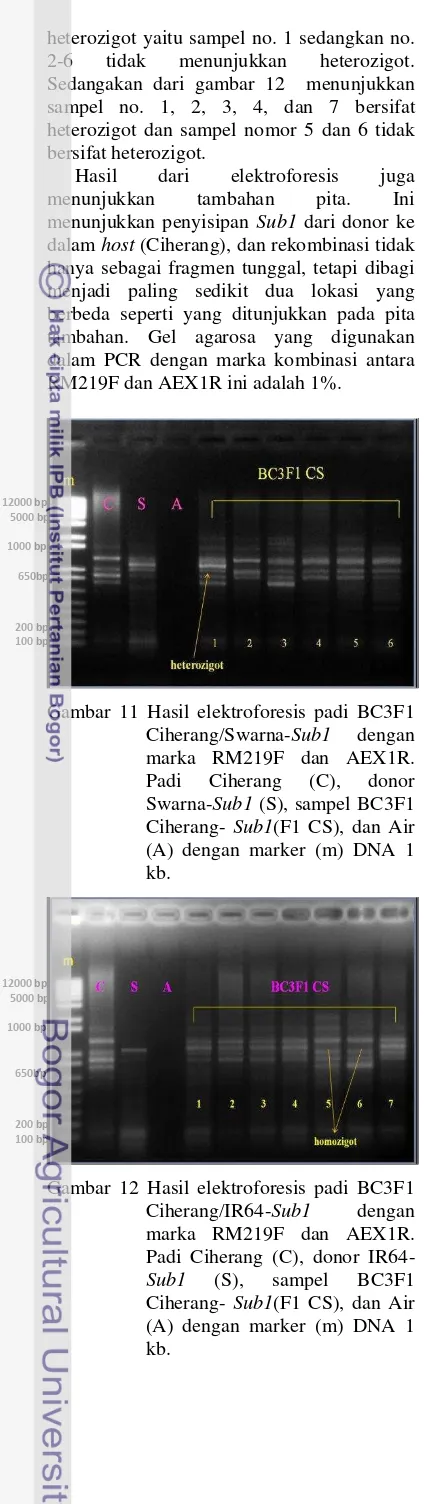
Hal ini dilakukan untuk mendeteksi keheterozigotan sifat padi BC3F1 agar lebih jelas pitanya. Hasil yang menarik dan konsisten (Gambar 11 dan 12) diperoleh saat RM219F dan AEX1R digunakan sebagai marker. Sampel donor Swarna dan IR64 menunjukkan pita yang tidak ada pada padi host (Ciherang). Sedangkan pita Ciherang dan pita Swarna/IR64 yang hadir dalam tanaman BC3F1 Ciherang-Sub1 menunjukkan bahwa padi tersebut bersifat heterozigot. Dari gambar 11 menunjukkan hanya satu sampel padi BC3F1 yang bersifat
heterozigot yaitu sampel no. 1 sedangkan no. 2-6 tidak menunjukkan heterozigot. Sedangakan dari gambar 12 menunjukkan sampel no. 1, 2, 3, 4, dan 7 bersifat heterozigot dan sampel nomor 5 dan 6 tidak bersifat heterozigot.
Hasil dari elektroforesis juga menunjukkan tambahan pita. Ini menunjukkan penyisipan Sub1 dari donor ke dalam host (Ciherang), dan rekombinasi tidak hanya sebagai fragmen tunggal, tetapi dibagi menjadi paling sedikit dua lokasi yang berbeda seperti yang ditunjukkan pada pita tambahan. Gel agarosa yang digunakan dalam PCR dengan marka kombinasi antara RM219F dan AEX1R ini adalah 1%.
Gambar 11 Hasil elektroforesis padi BC3F1 Ciherang/Swarna-Sub1 dengan marka RM219F dan AEX1R. Padi Ciherang (C), donor Swarna-Sub1 (S), sampel BC3F1 Ciherang- Sub1(F1 CS), dan Air (A) dengan marker (m) DNA 1 kb.
Gambar 12 Hasil elektroforesis padi BC3F1 Ciherang/IR64-Sub1 dengan marka RM219F dan AEX1R. Padi Ciherang (C), donor IR64-Sub1 (S), sampel BC3F1 Ciherang- Sub1(F1 CS), dan Air (A) dengan marker (m) DNA 1 kb.
SIMPULAN DAN SARAN
Simpulan
Pembentukan populasi BC3F1 Ciherang-Sub1 telah berhasil dilakukan melalui persilangan terarah (Site Directed Crossing) dengan silang balik antara host (Ciherang) dan padi BC2F1 Ciherang-Sub1. Hasil elektroforesis PCR menunjukkan bahwa terdapat pita DNA gen Sub1 hasil amplifikasi dengan ukuran 231 bp, sedangkan yang menggunakan marka RM219F dan AEX1R menunjukkan adanya pola pita heterozigot.
Saran
Penelitian lanjutan perlu dilakukan hingga dihasilkan tanaman generasi BC5F1 Ciherang-Sub1 dan perlu analisis lanjutan PCR dengan marka yang dapat membedakan dengan jelas keheterozigotan gen hasil persilangan seperti kit berbasis nanomolecular beacon yang merupakan solusi alternatif.
DAFTAR PUSTAKA
Armstrong J, Armstrong W. 2005. Rice: sulfide-induced barriers to root radial oxygen loss, Fe2+ and water uptake, and lateral root emergence. Annals of Botany 96: 625–638.
Armstrong W, Drew MC. 2002. Root Growth and Metabolism Under Oxygen Deficiency. Di dalam: Waisel Y, Eshel A, Kafkafi U, editor. Plant roots: the hidden half. Ed ke-3. New York: Marcel Dekker. hlm 729–761.
CGIAR (Consultative Group on International Agriculture Research). 2006. Intensified Research Effort Yie