KARAKTERISASI DAN PENINGKATAN JUMLAH BUNGA
HERMAPRODIT TANAMAN JARAK PAGAR (
Jatropha
curcas
) ANDROMONOECIOUS
DASUMIATI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa disertasi berjudul “Karakterisasi dan Peningkatan Jumlah Bunga Hermaprodit Tanaman Jarak Pagar (Jatropha curcas) Andromonoecious” adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juli 2014
Dasumiati
RINGKASAN
DASUMIATI. Karakterisasi dan Peningkatan Jumlah Bunga Hermaprodit Tanaman Jarak Pagar (Jatropha curcas) Andromonoecious. Dibimbing oleh MIFTAHUDIN, TRIADIATI, dan ALEX HARTANA.
Jarak pagar (Jatropha curcas L.) berpotensi sebagai bahan baku biodiesel terbaharukan. Perkembangan buah dan biji yang merupakan bahan baku biodiesel pada jarak pagar sangat bergantung pada karakteristik bunga. Pembungaan pada jarak pagar cukup unik karena menghasilkan bunga jantan, betina, dan atau hermaprodit. Jarak pagar andromonoecious menghasilkan bunga jantan dan hermaprodit pada infloresen bunga yang sama dan masih sangat jarang ditemukan, sedangkan jarak pagar monoecious yang umum ditemukan dan menghasilkan bunga jantan dan betina pada infloresen bunga yang sama.
Penelitian ini bertujuan untuk menganalisis karakter tanaman jarak pagar andromonoecious dan meningkatkan jumlah bunga hermaprodit. Untuk mencapai tujuan ini lima tahap penelitian telah dilakukan. Tahap pertama, melakukan eksplorasi terhadap tipe seks pada jarak pagar dan menganalisis kestabilannya. Tahap kedua, menganalisis karakter morfologi dan perkembangan jarak pagar andromonoecious. Tahap ketiga, menganalisis karakter bunga dan fenologi jarak pagar andromonoecious. Tahap keempat, mengembangkan marka molekuler untuk karakter tipe seks pada jarak pagar andromonoecious. Tahap kelima, peningkatan bunga hermaprodit pada jarak pagar andromonoecious.
Eksplorasi tipe seks jarak pagar dilakukan pada dua perkebunan jarak pagar, yaitu Kebun Industri Bukit Indah (KIBI) di Cikampek dan Kebun Induk Jarak Pagar (KIJP) di Pakuwon. Tiga tipe seks jarak pagar ditemukan pada kedua perkebunan tersebut, yaitu monoecious, andromonoecious, dan trimonoecious. Monoecious merupakan tipe seks jarak pagar yang umum ditemukan. Tipe seks andromonoecious dan monoecious stabil pada tanaman hasil stek batang dan penyerbukan terbuka. Jarak pagar trimonoecious tidak stabil karena bunga hermaprodit, betina, dan jantan tidak selalu dihasilkan oleh infloresen yang sama, individu yang sama dan pada musim yang sama.
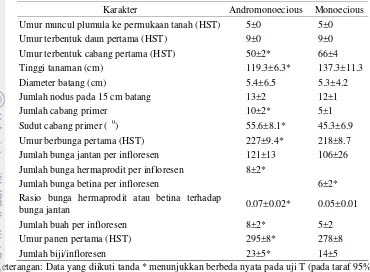
Karakter morfologi diamati pada jarak pagar andromonoecious dan monoecious aksesi Dompu. Perkembangan tanaman jarak pagar diamati pada tanaman hasil penyerbukan sendiri kedua jarak pagar tersebut. Karakter yang khas dan membedakan jarak pagar andromonoecious dengan monoecious adalah jumlah cabang, sudut cabang, jarak antar nodus, jumlah infloresen per tanaman, persentase bunga menjadi buah (fruit set), dan berat total biji. Karakter-karakter ini lebih besar pada jarak pagar andromonoecious dibandingkan monoecious kecuali jarak antar nodus. Pertumbuhan kedua jarak pagar ini hampir sama kecuali pada umur terbentuknya cabang pertama dan umur berbunga pertama. Jarak pagar andromonoecious membentuk cabang lebih cepat dibandingkan monoecious, tetapi andromonoecious berbunga pertama lebih lambat dari monecious.
cabang sekunder dan tersier. Setiap cabang ditutup oleh bunga hermaprodit. Jumlah cabang infloresen pada jarak pagar andromonoecious lebih banyak dibandingkan monoecious. Bunga hermaprodit memiliki diameter bunga, sepal, dan tangkai bunga lebih besar dibandingkan dengan bunga jantan dan betina. Perkembangan bunga jarak pagar andromonoecious terbagi menjadi 4 fase, yaitu fase inisiasi (0-3 hsi = hari setelah inisiasi), fase kuncup kecil (3-7 hsi), fase kuncup besar (7-25 hsi), dan fase mekar (20-29 hsi). Bunga hermaprodit mekar sempurna bersamaan dengan antera merekah yaitu pukul 06.00-06.59 WIB yang menyebabkan penyerbukan sendiri telah terjadi pada bunga hermaprodit. Rata-rata viabilitas polen pada saat mekar 96.3%, dan 88.8% nya masih bertahan sampai hari berikutnya.
Pengembangan marka molekuler bertujuan untuk mencari marka jarak pagar andromonoecious sebagai pembeda dari jarak pagar monoecious pada aksesi Dompu. Lima pasang primer didisain berdasarkan EST (Expressed Sequence Tag) dari gen AGAMOUS, APETALA 3, dan SEPALLATA. Namun hanya satu pasang primer dari APETALA 3 yang memperlihatkan pola pita polimorfisme pada jarak pagar andromnoecious dan jarak pagar monoecious. Hasil analisis dengan EST
diperoleh dua pita DNA (marka) yaitu 100 bp pada jarak pagar monoecious dan 500 bp pada jarak pagar andromonoecious. Untuk melihat pola pewarisan kedua marka ini disilangkan jarak pagar monoecious dan andromonoecious. Kedua marka diwariskan pada tanaman F1 dan F2, namun tidak mengikuti pola segregasi monohibrid dan dihibrid Mendel. Marka 500 bp berkorelasi dengan banyaknya cabang, sedangkan marka 100 bp berkorelasi dengan tipe seks.
Zat pengatur tumbuh kinetin, GA3, dan IAA dengan tiga konsentrasi yang
berbeda (0, 50, dan 100 ppm) digunakan dalam peningkatan bunga hermaprodit pada jarak pagar andromonoecious. Percobaan dilakukan dengan Rancangan Acak Kelompok (RAK) dengan 9 perlakuan dan 4 ulangan. Zat pengatur tumbuh disemprotkan ke daun bagian pucuk setiap minggu selama 10 minggu dimulai sejak inisiasi bunga pertama. Kinetin, GA3, IAA dan 50 dan 100 ppm
meningkatkan jumlah bunga hermaprodit per tanaman berturut-turut sebesar, 163 dan 103% (kinetin), 220 dan 254% (GA3), 276 dan 183% (IAA). Perlakuan
kinetin 50 ppm, GA3 100 ppm, atau IAA 50 ppm berturut-turut meningkatkan
jumlah buah per tanaman sebesar 163, 257,dan 302%.
Karakter jarak pagar andromonoecious, seperti tipe seks yang stabil, jumlah cabang batang yang banyak, jarak antar nodus yang dekat, berbunga sepanjang tahun, fruit set yang tinggi, dan menyerbuk sendiri dapat meningkatkan produksi biji pada jarak pagar. Selain itu, marka molekuler yang dimiliki oleh jarak pagar andromonoecious untuk menduga gen penyandi jumlah cabang dan tipe seks berpotensi untuk ditindaklanjuti.
SUMMARY
DASUMIATI. Characteriation and the Increasing Number of Hermaphrodite Flower of Andromonoecious Jatropha curcas. Supervised by MIFTAHUDIN, TRIADIATI, and ALEX HARTANA.
Jatropha curcas L. seeds have potential to be used as a source for biodisel. Fruit and seed set in Jatropha depend on the characteristic of flower, which uniquely consist of male, female, and or hermaphrodit flowers. Andromonoecious Jatropha, which is rarely to be found, produces male and hermaphrodit flower in the same inflorescence flower, while monoecious, a common Jatropha plants, produces male and female flowers in the same inflorescence flower.
This research aimed to characterize andromonoecious Jatropha and to increase its hermaphrodit flower numbers. The research consisted of five activities, i.e.: 1. Exploration of sex types of Jatropha, 2. Analysis of morpholo-gical characteristic and the developing of andromonoecious Jatropha, 3. Analysis of flower characteristic and phenology of andromonoecious Jatropha, 4. Development of molecular marker for andromonoecious Jatropha, and 5. Hormon application to increase number of hermaphrodit flowers of andromonoecious Jatropha.
Exploration for sex type of Jatropha was conducted in two Jatropha plantations: Kebun Industri Bukit Indah (KIBI) Cikampek and Kebun Induk Jarak Pagar (KIJP) Pakuwon. Three sex types of Jatropha were found in both Jatropha plantations: monoecious, andromonoecious, and trimonoecious. Monoecious is a sex type of Jatropha that commonly found. Andromonoecious and monoecious sex type were stable in Jatropha plant that propagated with stem cutting as well as in open pollinated plants. Trimonoecious Jatropha plants were found to be unstable, since hermaphrodite, female, and male flowers were not always found in the same infloresence, in the same individual plant, and in the same season.
Andromonoecious and monoecious Jatropha Dompu accession were used to analyze morphological and developmental characters of andromonoecious Jatropha. The plants that were propagated from stem cutting were used to analyze morphological characters, while self pollinated plants were used to analyze the development of Jatropha plants. Number of branch, angle of branch, distance of internode, number of inflorescence per plant, number of fruit set, and total weight of seeds were observed in this research. Andromonoecious Jatropha was superior for those characters comparing to monoecious Jatropha, except for the distance of internode. The development of andromonoecious and monoecious Jatropha plants were almost the same. Andromonoecious Jatropha plants produced branch earlier than that of monoecious Jatropha plants, but andromonoecious flowers later than that of monoecious.
Flower characteristics and phenology of andromonoecious Jatropha are important to know before developing andromonoecious Jatropha. The part of the research observed flower characteristic, phenology, number and viability of pollen of Jatropha flowers. Inflorescence type of andromonoecious Jatropha was
Jatropha. The diameter, sepal and footstalk sizes of hermaphrodite flower were larger than that in male and female flowers. The development of andromonoe-cious Jatropha flowers consisted of 4 phases, i.e.: the initiation (0-3 dai = days after initiation), small bud (3-7 dai), large bud (7-20 dai), and flower blooming phases (20-39 dai). The anthesis and anther dehiscence of hermaphrodite flowers occured at the same time between 06:00 to 06:59 am suggesting that self-pollination occurred at the hermaphrodite flowers. The average pollen viability at the anthesis phase was 96.3%, and 88.8% of them will survive until the next day.
The development of molecular markers related to andromonoecious Jatropha sex type was carried out using Dompu accession. Five pairs of primer were designed based on EST (Expressed Sequence Tag) from AGAMOUS, APETALA 3, and SEPALLATA. But, only one pair of primer from APETALA 3
showed polimorphism between andromonoecious and monoecious Jatropha. A marker with the size of 100 bp was found specifically in monoecious and another marker with the size of 500 bp was found only in andromonoecious Jatropha. To analyze the inheritance pattern of both markers crosses between monoecious and andromonoecious Jatropha were conducted. Both markers inherited to F1 and F2 plants, but it did not follow the segregation pattern of mono and dihybrid segregation pattern of Mendel's laws. Further analysis suggested that the 500 bp marker might be linked to the number of branch, while the 100 bp marker might be linked to sex type of Jatropha.
To increase the number of hermaphrodite flowers in andromonoecious Jatropha, three different concentrations (0, 50, and 100 ppm) of kinetin, GA3, and
IAA were applied in this experiment. The kinetin, GA3, and IAA were sprayed
separately to leaf bud every week for 10 weeks since the first initiation. The 50 and 100 ppm of hormon increased the number of hermaphrodite flower per plant up to163 and 103% (kinetin), 220 and 254% (GA3), 276 and 183% (IAA). While
50 ppm kinetin, 100 ppm GA3, or 50 ppm IAA increased the number of fruit up to
163, 257,and 302%, respectively.
The andromonoecious Jatropha characters, such as stable sex type, high number of branch, and continous flower production along the year, self pollinated, and high fruit set could increase seed production. Molecular markers of 100 and 500 bp were suggested to be linked to number of branches and sex type characters in andromonoecious Jatropha.
© Hak Cipta Milik IPB, Tahun 2011
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Disertasi
sebagai salah satu syarat untuk memperoleh gelar Doktor pada
Program Studi Biologi Tumbuhan
KARAKTERISASI DAN PENINGKATAN JUMLAH BUNGA
HERMAPRODIT TANAMAN JARAK PAGAR (
Jatropha
curcas
) ANDROMONOECIOUS
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2014
Penguji pada Ujian Tertutup: 1. Prof Dr Ir Memen Surahman, MScAgr 2. Dr Ir Tatik Chikmawati, MSi
Judul Disertasi : Karakterisasi dan Peningkatan Jumlah Bunga Hermaprodit Tanaman Jarak Pagar (Jatropha curcas) Andromonoe-cious
Nama : Dasumiati
NIM : G363090021
Disetujui oleh Komisi Pembimbing
Dr Ir Miftahudin, MSi Ketua
Dr Dra Triadiati, MSi Prof Dr Ir Alex Hartana, MSc Anggota Anggota
Diketahui oleh
Ketua Program Studi Biologi Tumbuhan Dekan Sekolah Pascasarjana
Dr Ir Miftahudin, MSi . Dr Ir Dahrul Syah, MScAgr .
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini dapat diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Mei 2010 ini ialah pembungaan, dengan judul Karakterisasi dan Peningkatan Jumlah Bunga Hermaprodit Tanaman Jarak Pagar (Jatropha curcas) Andromonoecious.
Terima kasih penulis ucapkan kepada Dr Ir Miftahudin, MSi, Dr Dra Triadiati, MSi, dan Prof Dr Ir Alex Hartana, MSc selaku pembimbing yang banyak memberi saran. Ungkapan terima kasih juga penulis sampaikan kepada bapak, amak, suami, anak-anak ku, kakak-kakak dan adik-adik ku, ponakan-ponakan ku, paman, serta seluruh keluarga atas segala doa dan kasih sayangnya. Terima kasih juga penulis ucapkan pada Kementerian Pertanian melalui program KKP3T tahun 2010 a/n Triadiati yang telah membiayai sebagian dari penelitian ini.
Topik pengembangan marka molekuler untuk tipe seks pada jarak pagar (Jatropha curcas) telah diseminarkan di Kasetsart University, Thailand pada acara I-KUSTARS tanggal 27-28 Maret 2014. Semoga semua topik dalam karya ilmiah ini bermanfaat.
Bogor, Juli 2014
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN viii
1 PENDAHULUAN 11
Latar Belakang 1
Perumusan Masalah 2
Tujuan Penelitian 3
Manfaat Penelitian 3
Ruang Lingkup Penelitian 4
2 TINJAUAN PUSTAKA 5
Botani Tanaman Jarak Pagar 5
Potensi Tanaman Jarak Pagar sebagai Biodiesel 6
Karakterisasi Tanaman 8
Determinasi Seks pada Tanaman 10
3 TIPE SEKS PADA BUNGA TANAMAN JARAK PAGAR (Jatropha
curcas) 15
Abstract 15
Pendahuluan 15
Bahan dan Metode 16
Hasil 17
Pembahasan 21
Simpulan 22
4 KARAKTER MORFOLOGI DAN PERKEMBANGAN JARAK PAGAR
(Jatropha curcas) ANDROMONOECIOUS 23
Abstract 23
Pendahuluan 23
Bahan dan Metode 24
Hasil 25
Pembahasan 31
Simpulan 34
5 KARAKTERISASI BUNGA DAN FENOLOGI JARAK PAGAR (Jatropha
curcas) ANDROMONOECIOUS 35
Abstract 35
Pendahuluan 35
Bahan dan Metode 36
Hasil 37
Pembahasan 43
6 PENGEMBANGAN MARKA MOLEKULER UNTUK KARAKTER TIPE SEKS PADA JARAK PAGAR (Jatropha curcas) ANDROMONOE-
CIOUS 48
Abstract 48
Pendahuluan 48
Bahan dan Metode 49
Hasil 51
Pembahasan 56
Simpulan 58
7 PENINGKATAN BUNGA HERMAPRODIT DENGAN PEMBERIAN ZAT PENGATUR TUMBUH PADA JARAK PAGAR (Jatropha curcas)
ANDROMONOECIOUS 60
Abstract 60
Pendahuluan 60
Bahan dan Metode 62
Hasil 63
Pembahasan 67
Simpulan 72
8 PEMBAHASAN UMUM 73
9 SIMPULAN DAN SARAN 78
Simpulan 78
Saran 78
DAFTAR PUSTAKA 79
LAMPIRAN 87
DAFTAR TABEL
3.1 Tipe seks individu tanaman koleksi jarak pagar di perkebunan
KIBI Cikampek dan KIJP Pakuwon 18
3.2 Kestabilan tipe seks dan jumlahnya pada tanaman hasil perbanyakan vegetatif jarak pagar monoecious, andromonoecious,
dan trimonoecious selama musim berbunga pertama 19 3.3 Tipe seks dan percabangan pada tanaman hasil perbanyakan
generatif jarak pagar andromonoecious, monoecious, dan
trimo-noecious 21
4.1 Perbandingan karakter organ vegetatif pada jarak pagar
andromo-noecious dengan moandromo-noecious 26
4.2 Perbandingan karakter organ generatif pada jarak pagar
andromo-noecious dengan moandromo-noecious 28
4.3 Karakter perkembangan tanaman jarak pagar andromonoecious
dan monoecious 31
5.1 Karakteristik bunga jarak pagar andromonoecious dan monoecious 39 5.2 Persentase bunga mekar pada interval waktu tertentu berdasarkan
proses mekarnya pada jarak pagar andromonoecious dan
monoecious 42
6.1 Sekuen primer yang digunakan dalam penelitian pengembangan
marka molekular terkait karakter tipe seks pada jarak pagar 51 6.2 Hasil analisis menggunakan BLAST dari pita hasil amplifikasi
DNA jarak pagar andromonoecious dan monoecious 52 6.3 Tipe seks, jumlah cabang, dan pola pita hasil amplifikasi DNA
jarak pagar monoecious, andromonoecious, serta tanaman F1 hasil
persilangan dan resiprokalnya 53
6.4 Tipe seks, jumlah cabang, dan pola pita hasil amplifikasi DNA tanaman F2 hasil persilangan jarak pagar monoecious dan
andromonoecious 54
6.5 Uji khi-kuadrat pola segregasi pita 100 dan 500 bp pada tanaman F2 hasil persilangan jarak pagar monoecious dan
andromonoe-cious 55
7.1 Korelasi karakter produksi dengan jumlah cabang pada jarak pagar andromonoecious yang diberi perlakuan ZPT (kinetin, GA3,
dan IAA) 68
8.1 Nilai skor karakter-karakter kuantitatif dan kualitatif dari jarak
DAFTAR GAMBAR
1.1. Bagan alur penelitian karakterisasi dan peningkatan jumlah bunga
hermaprodit tanaman jarak pagar andromonoecious 4 2.1. Konsep gen model ABC dalam meregulasi pembentukan struktur
dasar bunga 11
3.1 Bunga dan tipe seks tanaman hasil perbanyakan vegetatif jarak pagar andromonoecious, monoecious, trimonoecious,
trimonoe-cious yang menampilkan monoetrimonoe-cious 19
3.2 Profil tanaman hasil perbanyakan vegetatif jarak pagar andromo-noecious, moandromo-noecious, trimonoecious aksesi Lampung,
trimonoe-cious aksesi Banten umur 5 bulan 20
4.1 Profil jarak pagar andromonoecious dengan percabangannya dan
profil jarak pagar monoecious dengan percabangannya 26 4.2 Kecenderungan karakter vegetatif pada jarak pagar
andromonoe-cious dan monoeandromonoe-cious 27
4.3 Infloresen, rangkaian buah, buah, dan biji jarak pagar
andromonoecious, serta rangkaian buah jarak pagar monoecious 28 4.4 Jumlah infloresen yang dihasilkan jarak pagar andromonoecious
dan monoecious setiap bulan pada dua musim berbunga (Juli
2010-Juni 2011) 29
4.5 Proses perkecambahan biji jarak pagar andromonoecious 3 HST, 5
HST, 7 HST, 9 HST, 11 HST, dan 15 HST 30
4.6 Percabangan pada jarak pagar andromonoecious umur 1.5 bulan dan 1 tahun, serta percabangan pada jarak pagar monoecious umur
1.5 bulan dan 1 tahun 30
5.1 Infloresen bunga jarak pagar andromonoecious dan monoecious 38 5.2 Struktur penyusun bunga hermaprodit, bunga betina, bunga jantan
jarak pagar 39
5.3 Fase perkembangan bunga jarak pagar andromonoecious dan
ana-tomi kuncup bunga umur 3 hari setelah inisiasi 40
5.4 Tahapan mekar bunga hermaprodit 41
5.5 Polen viabel dan polen yang mulai berkecambah pada bunga hermaprodit sebelum antera merekah, anatomi bunga hermaprodit
sebelum antesis, dan anatomi bunga betina setelah antesis 43 6.1 Hasil amplifikasi DNA jarak pagar monoecious,
andromonoe-cious, F1 dan F2 hasil persilangan monoecious dengan
andromo-noecious menggunakan primer JA3 52
6.2 Percabangan pada tanaman tetua monoecious, andromonoecious,
dan F1 setelah terbentuknya cabang pertama dan umur satu tahun 56 7.1 Pengaruh kinetin, GA3, dan IAA terhadap jumlah infloresen pada
jarak pagar andromonoecious selama 10 minggu pengamatan 63 7.2 Pengaruh perlakuan ZPT terhadap percabangan infloresen bunga
7.3 Pengaruh kinetin, GA3, dan IAA terhadap jumlah bunga
hermaprodit per infloresen, jumlah bunga hermaprodit per tanaman, jumlah bunga jantan per infloresen, dan rasio bunga hermaprodit terhadap bunga jantan per infloresen selama 10
minggu pengamatan 65
7.4 Pengaruh kinetin, GA3, dan IAA terhadap buah per infloresen,
jumlah buah per tanaman, berat biji per infloresen, dan berat 100
biji selama 10 minggu pengamatan 66
7.5 Buah jarak pagar andromonoecious dengan 3 ruang buah dan 4
ruang buah 66
DAFTAR LAMPIRAN
1 Data iklim di Dramaga, Bogor, Jawa Barat pada tahun 2011 87 2 Alur pembuatan preparat permanen dengan metode parafin 88
3 Alur pewarnaan safranin-fast green 89
4 Foto beberapa serangga pengunjung pada bunga tanaman jarak
pagar selama pengamatan 90
5 Data iklim di Bambu Apus, Kecamatan Pamulang, Kota
1
PENDAHULUAN
Latar Belakang
Krisis energi yang sudah terjadi sejak tahun 1970-an dapat berupa keterbatasan bahan baku dan dampak penggunaannya. Keterbatasan bahan baku merupakan keterbatasan sumber daya energi fosil. Dampak penggunaan energi fosil meningkatkan polusi lingkungan dan perubahan kondisi iklim (Srivastava et al. 2011). Kedua hal ini merangsang upaya pengembangan minyak nabati sebagai sumber energi terbaharukan (renewable energy) yang dapat mendukung upaya pelestarian lingkungan hidup. Berdasarkan konsepnya, teknologi energi baru dan terbarukan bukan merupakan sesuatu yang baru, karena sumber-sumber energi ini berupa biomassa, terutama biomassa produk-produk tumbuhan.
Tujuan utama pengembangan energi minyak nabati adalah untuk pemanfaatan tanaman penghasil minyak untuk pembangunan ekonomi pedesaan dan lingkungan yang berkelanjutan, serta untuk membuat daerah pedesaan mandiri energi, khususnya bahan bakar cair. Tujuan ini diharapkan dapat dicapai tanpa menggusur tanaman pertanian lain dalam penggunaan lahan. Jarak pagar (Jatropha curcas) merupakan salah satu tanaman yang dapat terpilih sebagai penghasil minyak nabati utama (Openshaw 2000).
Jarak pagar sebagai sumber Bahan Bakar Nabati (BBN) yang potensial, mempunyai banyak keunggulan dibandingkan sumber nabati lainnya. Tanaman ini memiliki kemampuan adaptasi yang baik terhadap berbagai lingkungan (Heller 1996), toleran terhadap kekeringan, tahan terhadap hama dan penyakit, pertumbuhannya cepat, serta masih dapat berproduksi pada lahan dengan kesuburan rendah seperti lahan marginal (Openshaw 2000, Maes et al. 2009, Mishra 2009, Yi et al. 2010) sehingga dapat menunjang usaha konversi lahan. Oleh karena itu tanaman jarak pagar sangat berpeluang dikembangkan di Indonesia, karena tersedia 49.53 juta ha sesuai untuk jarak pagar yang terdiri dari 14.28 juta hasangat sesuai, 5.53 juta ha cukup sesuai, dan sesuai marginal 29.72 juta ha yang belum dimanfaatkan dan dikelola dengan baik (Syakir 2010).
Pengembangan BBN sangat didukung oleh pemerintah. Berdasarkan Perpres No. 5 tahun 2006 tentang Kebijakan Energi Nasional, dan Instruksi Presiden No. 1 tahun 2006 yang antara lain menugaskan Kementerian Pertanian untuk mendorong penyediaan tanaman termasuk memfasilitasi penyediaan benih dan bibit, penyuluhan, serta pengintegrasian kegiatan pengembangan dan kegiatan pasca panen. Organisasi nasional yang telah dibentuk adalah Tim Nasional Pengembangan BBN dan kelompok-kelompok kerjanya yang disertai Keppres No. 10 tahun 2006 (http://www.esdm.go.id/regulasi/perpres/cat_view/64-regulasi/ 229-peraturan-presiden/232-tahun-2006.html).
2
untuk perkebunan jangka panjang dalam memproduksi minyak (Heller 1996). Akan tetapi pengembangan jarak pagar belum didukung oleh ketersediaan benih bermutu dengan potensi produksi yang tinggi (Yoga et al. 2007). Produksi biji pada jarak pagar sangat terkait dengan pembungaan.
Jarak pagar memiliki pembungaan yang cukup menarik, yaitu dapat menghasilkan bunga jantan dan betina atau hermaprodit pada infloresen yang sama. Tanaman yang menghasilkan bunga jantan dan betina pada individu tanaman yang sama disebut monoecious, sedangkan tanaman yang menghasilkan bunga jantan dan hermaprodit pada individu tanaman yang sama disebut andromonoecious (Dellaporta & Urrea 1993). Monoecious pada tumbuhan hanya sekitar 10% (Lebel-Hardenack & Grant 1997), tetapi tanaman jarak pagar pada umumnya adalah monoecious (Alam et al. 2011). Tipe seks andromonoecious yang menghasilkan bunga hermaprodit dan jantan masih jarang ditemukan, dan belum tereksplorasi pada tanaman jarak pagar.
Jarak pagar yang dikembangkan selama ini adalah jarak pagar monoecious. Jumlah bunga betina per infloresen dan penyerbukan sendiri pada jarak pagar monoecious menjadi kendala dalam pengembangan tanaman jarak pagar. Jumlah bunga betina yang dihasilkan berkisar 5 bunga dalam satu infloresen (Wijaya et al. 2009) dengan rasio terhadap bunga jantan sekitar 1:29 (Raju & Ezradanam 2002). Tanaman jarak pagar monoecious cenderung menyerbuk silang (cross pollinated) (Hartati 2009), sehingga terjadi keragaman benih (terjadi rekombinasi gen) dan tanaman yang dihasilkan tidak sama dengan tetuanya (Cholid et al. 2012). Penyerbukan silang sangat bergantung pada keberadaan polinator, akibatnya keberhasilan pembentukan buah juga sangat bergantung pada polinator. Jika polinator tidak ada, maka bunga tidak berkembang menjadi buah.
Jarak pagar andromonoecious menghasilkan bunga jantan dan hermaprodit pada infloresen yang sama. Bunga hermaprodit yang dihasilkan jarak pagar andromonoecious ini lebih berpeluang melakukan penyerbukan sendiri (self pollination) (Hartati 2009). Penyerbukan sendiri berpeluang menghasilkan benih yang lebih seragam dan tanaman yang memiliki karakter yang sama dengan tetuanya. Berdasarkan penelitian pendahuluan persentase bunga menjadi buah (fruit set) pada jarak pagar andromonoecious dapat mencapai 100%. Pada musim kemarau, tanaman jarak pagar andromonoecious dapat memproduksi bunga hermaprodit dan buah mencapai 15 buah per infloresen, sedangkan tanaman monoecious sebagian besar tidak berbunga.
Perumusan Masalah
Pengembangan tanaman jarak pagar dalam menghadapi krisis energi dapat menjadi sumber energi alternatif BBN yang ramah lingkungan dan meningkatkan perekonomian masyarakat. Ada beberapa kendala dalam pengembangan jarak pagar terutama yang berhubungan dengan pembungaan untuk menghasilkan produksi tinggi dan stabil, serta benih yang seragam.
3 dalam hal produksi karena jumlahnya per infloresen rendah dan penyerbukan sangat bergantung pada polinator sehingga cenderung mengalami penyerbukan silang yang menyebabkan keragaman benih.
Bunga hermaprodit yang dimiliki jarak pagar andromonoecious sangat berpeluang untuk mengatasi kendala pengembangan jarak pagar, terutama yang terkait dengan tingkat produksi dan penyediaan benih. Untuk itu perlu dilakukan eksplorasi terhadap tanaman jarak pagar berdasarkan tipe pembungaannya. Tanaman jarak pagar yang menghasilkan bunga hermaprodit (jarak pagar andromonoecious) perlu diteliti lebih lanjut tentang karakter morfologi dan molekuler, konsistensi berbunga hermaprodit, pewarisan sifat berbunga hermaprodit, biologi bunga, sampai upaya peningkatan jumlah bunga hermaprodit.
Tujuan Penelitian
Tujuan penelitian ini adalah untuk :
1. Mengeksplorasi jarak pagar berdasarkan tipe seks bunga pada populasi jarak pagar yang ditanam di Kebun Industri Bukit Indah (KIBI) Cikampek and Kebun Induk Jarak Pagar (KIJP) Pakuwon dan menganalisis kestabilan tipe seks tersebut.
2. Menganalisis karakter (karakterisasi) jarak pagar andromonoecious melalui pendekatan morfologi dan pertumbuhan.
3. Menganalisis karakter bunga dan fenologi jarak pagar andromonoecious. 4. Mengembangkan marka molekuler terkait dengan tipe seks pada jarak pagar
menggunakan marka molekuler expressed sequence tag (EST).
5. Mendapatkan teknologi peningkatan pembentukan bunga hermaprodit dengan menggunakan zat pengatur tumbuh pada tanaman jarak pagar andromo-noecious.
Manfaat Penelitian
Hasil penelitian ini dapat menjadi dasar bagi penelitian selanjutnya dalam hal pengembangan tanaman jarak pagar untuk menunjang pengembangan BBN. Karakter-karakter spesifik pada tiap tipe seks jarak pagar dapat digunakan untuk mendeteksi tipe seks yang dihasilkan oleh tanaman jarak pagar lebih dini. Tanaman yang menghasilkan bunga hermaprodit (andromonoecious) sudah dapat diketahui sejak masih anakan dan dapat dikembangkan lebih lanjut untuk memperoleh produksi yang tinggi. Pembentukan biji dapat dipelajari melalui pengamatan fenologi bunga, sehingga proses untuk mendapat benih yang berkualitas dapat diketahui dan dikembangkan sesuai dengan proses pembentukannya. Marka molekuler yang membedakan antara jarak pagar andromonoecious dengan monoecious dapat dikembangkan untuk menganalisis kandidat gen yang ikut mengendalikan tipe seks pada jarak pagar dan dapat digunakan dalam pemuliaan tanaman jarak pagar. Selain itu, dari penelitian ini dapat diperoleh teknik peningkatan bunga hermaprodit pada tanaman jarak pagar andromonoecious.
4
andromonoecious yang masih belum ada informasinya yaitu karakter morfologi dan fenologi. Kedua, penemuan jarak pagar andromonoecious yang memiliki tipe seks yang stabil. Ketiga, teknik peningkatan bunga hermaprodit pada jarak pagar andromonoecious.
Ruang Lingkup Penelitian
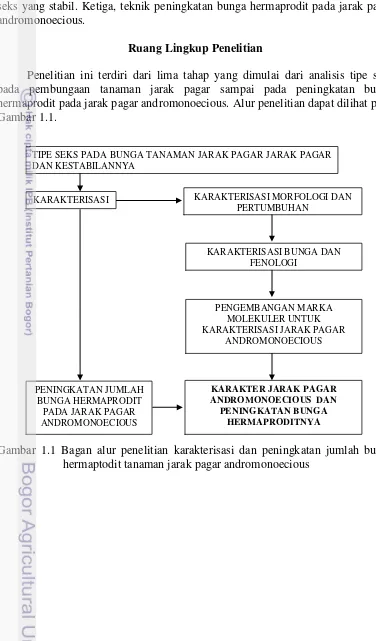
Penelitian ini terdiri dari lima tahap yang dimulai dari analisis tipe seks pada pembungaan tanaman jarak pagar sampai pada peningkatan bunga hermaprodit pada jarak pagar andromonoecious. Alur penelitian dapat dilihat pada Gambar 1.1.
Gambar 1.1 Bagan alur penelitian karakterisasi dan peningkatan jumlah bunga hermaptodit tanaman jarak pagar andromonoecious
TIPE SEKS PADA BUNGA TANAMAN JARAK PAGAR JARAK PAGAR DAN KESTABILANNYA
KARAKTERISASI MORFOLOGI DAN PERTUMBUHAN
PENGEMBANGAN MARKA MOLEKULER UNTUK KARAKTERISASI JARAK PAGAR
ANDROMONOECIOUS KARAKTERISASI BUNGA DAN
FENOLOGI
PENINGKATAN JUMLAH BUNGA HERMAPRODIT
PADA JARAK PAGAR ANDROMONOECIOUS KARAKTERISASI
KARAKTER JARAK PAGAR ANDROMONOECIOUS DAN
5
2 TINJAUAN PUSTAKA
Botani Tanaman Jarak Pagar
Jarak pagar (J. curcas L.) termasuk ke dalam famili Euphorbiaceae. Jarak pagar merupakan tanaman tropis asli Meksiko dan Amerika Tengah (Mishra 2009). Jarak pagar banyak dibudidayakan di Amerika Tengah, Amerika Selatan, Asia Tenggara, India dan Afrika (Gubitz et al. 1999). Tanaman ini mungkin didistribusikan oleh pelaut Portugis melalui Kepulauan Tanjung Verde dan Guinea Bissau ke negara-negara lain di Afrika dan Asia (Heller 1996). Di Indonesia, tanaman ini juga dikenal dengan nama jarak budeg, jarak gundul, atau jarak Cina.
Jarak pagar merupakan tanaman berkayu dengan batang berbentuk bulat dan mengandung banyak getah. Tinggi tanaman dapat mencapai 6 meter dan mampu hidup sampai 50 tahun. Kulit batang bewarna coklat pucat, tipis, dan cenderung mudah terkelupas. Batang memiliki percabangan yang tidak teratur dan berkembang ke arah atas. Daun tunggal dengan kedudukan berselang seling sepanjang batang. Daun memiliki tangkai (petiolus) dengan panjang 2-20 cm. Helaian daun berbentuk palmate atau menjari dengan sisi berlekuk-lekuk sebanyak 3-5 buah, lebar dengan ukuran 12.5-18 cm × 11-16 cm. Apeks (ujung) daun berbentuk acuminate dan margin (pinggiran) bergelombang (Jones & Csurhes 2008).
Bunga tanaman jarak pagar adalah bunga majemuk berbentuk malai dengan warna kuning kehijauan. Bunga berkelamin tunggal (unisexual), kadang-kadang hermaphrodit. Bunga betina memiliki ovarium yang terdiri dari 3 lokus, berbentuk ellips dengan diameter 1.5-2 mm. Sepal bunga betina lebih besar dari bunga jantan, yaitu sampai 18 mm. Bunga jantan memiliki 10 stamen (benang sari) tersusun dalam dua lingkaran. Jumlah bunga jantan 4-5 kali lebih banyak daripada bunga betina atau bahkan lebih. Buah berbentuk buah kendaga atau kapsul, oval atau bulat telur, diameter 2-4 cm. Permukaan kulit buah tidak berbulu (gundul), berwarna hijau ketika masih muda dan bewarna kuning kecoklatan setelah tua. Buah terbagi menjadi 3 ruang yang masing-masing memiliki 1 biji. Biji berbentuk bulat lonjong, berwarna coklat kehitaman dengan ukuran panjang 2 cm, tebal 1 cm, dan berat 0.4-0.6 gram/biji (Jones & Csurhes 2008). Tanaman ini mulai menghasilkan buah pada usia 4-5 bulan dan mencapai produksi biji maksimum sekitar tiga tahun (Heller 1996).
6
terpenting adalah minyak kental yang dikandung oleh biji jarak pagar dapat digunakan sebagai pengganti solar atau minyak tanah, serta untuk pembuatan sabun dalam industri kosmetik (Openshaw 2000). Aplikasi komersial pertama jarak pagar dilaporkan dari Lisbon, minyak jarak pagar impor dari Cape Verde digunakan untuk produksi sabun dan lampu. Bahkan saat ini, jarak pagar terutama dibudidayakan untuk produksi minyak sebagai pengganti bahan bakar (Gubitz et al. 1999).
Tanaman jarak pagar tidak memerlukan syarat tumbuh yang khusus. Tanaman ini secara luas ditanam di daerah tropis sebagai pagar hidup di sekitar ladang dan desa-desa. Jarak pagar mudah beradaptasi di lingkungan termasuk pada kondisi lingkungan yang kritis dan marginal, tanaman ini juga bisa ditanam untuk reboisasi wilayah terkikis (Heller 1996). Jarak pagar mampu hidup pada ketinggian 0-2000 m dpl, curah hujan 300-1200 mm per tahun, dan suhu berkisar antara 18-30oC. Pada daerah dengan suhu rendah (<18oC) dapat menghambat pertumbuhan, sedangkan pada suhu tinggi (>35oC) dapat menyebabkan gugur daun dan bunga, buah menjadi kering sehingga produksi tanaman ini menurun. Jarak pagar dapat tumbuh pada daerah yang kurang subur tetapi harus memiliki drainase yang baik, tidak tergenang, dan pH tanah 5.0-6.5 (Prihandana & Hendroko 2006). Selain itu, tanaman ini dapat tumbuh dengan baik pada lahan kering. Aksesi-aksesi jarak pagar yang tahan terhadap kekeringan tidak menunjukkan perbedaan pertumbuhan dengan penyiraman yang berbeda pada interval waktu penyiraman mulai dari 10 sampai 30 hari sekali (Behera et al. 2010).
Potensi Tanaman Jarak Pagar sebagai Biodiesel
Kenaikan harga bahan bakar minyak (BBM) yang berbasis fosil pada tahun 2005 sempat menyentuh angka lebih dari 70 US dollar per barel. Berbagai prediksi memperkirakan bahwa kenaikan harga tersebut bukanlah akhir dari episode kenaikan BBM fosil, melainkan tahap awal yang akan terus diikuti oleh kenaikan harga di tahun-tahun berikutnya, seperti yang terjadi pada pertengahan tahun 2013. Kenaikan harga BBM disebabkan oleh cadangan dan bahan baku BBM yang kian menipis atau bahkan habis. Peristiwa ini menjadi momentum penting bagi pengembangan energi alternatif pengganti BBM (Tim Nasional Pengembangan BBN 2002).
Bahan bakar nabati memiliki perbedaan yang cukup jauh dengan BBM, terutama dari sumber dan dampak penggunaannya. Bahan bakar nabati membutuhkan bahan baku biomassa tumbuhan sehingga lebih berbasis pada industri perkebunan dan pertanian, sedangkan BBM berasal dari fosil yang terbentuk dari tanaman serta hewan mikroskopis selama ratusan juta tahun. Basis BBN lebih ditekankan pada budidaya energi (energy farming), bukan berburu energi (energy hunting) seperti halnya yang dilakukan pada pengolahan BBM.
7 energi hijau (green energy), seperti budidaya tanaman jarak pagar untuk bahan baku BBN (Tim Nasional Pengembangan BBN 2002).
Berdasarkan hasil penelitian Badan Pengkajian dan Penerapan Teknologi tahun 2006, jarak pagar merupakan salah satu jenis tanaman di Indonesia yang dapat digunakan untuk pengembangan BBN. Minyak yang diperoleh setelah transesterifikasi dapat digunakan sebagai biofuel dalam mesin diesel. Biji jarak pagar dengan testanya mengandung 20-40% minyak nabati, namun bagian inti biji tanpa testa dapat mengandung 45-60% minyak kasar. Berdasarkan analisis terhadap komposisi asam lemak dari 11 provenans jarak pagar, diketahui bahwa asam lemak yang dominan adalah asam oleat, asam linoleat, asam stearat, dan asam palmitat. Komposisi asam oleat dan asam linoleat bervariasi, asam stearat dan asam palmitat merupakan asam lemak jenuh dengan komposisi yang relatif tetap (Heller 1996).
Minyak dari biji jarak pagar memiliki prospek menghasilkan biodiesel. Bahan tersisa setelah ekstraksi minyak (press cake) juga berpotensi sebagai pakan ternak terutama jika menggunakan kultivar yang tidak beracun, dan proses detoksifikasi murah telah berhasil dikembangkan. Tanaman ini juga telah diusulkan sebagai komoditas yang dapat diperdagangkan untuk kredit karbon. Nilai ekonomi potensial dari suatu industri perkebunan jarak pagar dibandingkan dengan spesies lain seperti Pongamia pinnata, merupakan biofuel yang potensial dengan potensi keuntungan industri mencapai $337-$396 juta per tahun (Jones & Csurhes 2008).
Beberapa negara yang miskin sumber daya BBM, seperti India, Tanzania dan Gambia telah lama mengembangkan tanaman jarak pagar sebagai pengganti kerosin (minyak tanah) untuk kompor dan lampu. Indonesia dalam targetnya pada tahun 2010 sudah siap memasuki generasi kedua pemakaian BBM dan BBN, yaitu pemakaian BBM campur BBN (1:1) (Tim Nasional Pengembangan BBN 2002).
Pengembangan pemanfaatan tanaman jarak pagar sebagai sumber bahan bakar alternatif terbarukan sangat potensial, karena tanaman ini mempunyai banyak keunggulan dibandingkan sumber nabati lainnya. Selain memiliki kandungan minyak yang tinggi, jarak pagar relatif mudah dibudidayakan. Jarak pagar tumbuh dengan baik pada kondisi kering dengan curah hujan rendah, pada lahan dengan kesuburan rendah yaitu lahan marginal atau lahan kritis sehingga dapat menunjang usaha konversi lahan. Oleh karena itu budidaya tanaman jarak pagar tidak berkompetisi dalam pemanfaatan lahan jika dibandingkan dengan tanaman pangan dan perkebunan (kelapa sawit atau tebu) (Sharma et al. 2009). Sifat ini memberikan peluang yang besar untuk dikembangkan, karena Indonesia masih memiliki lahan marginal yang cukup luas dan belum dimanfaatkan sebagai lahan pertanian.
8
Karakterisasi Tanaman
Karakterisasi merupakan proses mencari ciri spesifik yang dimiliki oleh tumbuhan serta digunakan untuk membedakan antar jenis dan antar individu dalam satu jenis suatu tumbuhan. Identitas atau identifikasi suatu tumbuhan dapat dilakukan dengan cara karakterisasi. Mengidentifikasi tumbuhan berarti mengungkapkan atau menetapkan identitas atau jati diri suatu tumbuhan, yang nantinya akan ditentukan namanya yang benar dan tempatnya yang tepat dalam suatu sistem (Tjitrosoepomo 1993). Dari hasil identifikasi banyak karakter yang bisa dijadikan sumber informasi untuk membedakan antar jenis atau spesies serta tipe seks tumbuhan. Sumber informasi yang dapat digunakan dalam membedakan antara satu jenis tumbuhan dengan jenis lain adalah informasi struktur, informasi kimia, informasi kromosom, dan informasi sistem penangkaran atau breeding
(Hartono 1989).
Informasi struktur. Informasi struktur yang dapat digunakan mengenal keragaman tumbuhan meliputi struktur organ vegetatif (anatomi dan morfologi) dan struktur organ generatif (anatomi dan morfologi). Pada organ vegetatif dapat digali informasi, seperti bentuk habit, bentuk batang, sistem perakaran, sistem percabangan, bentuk daun, jaringan pembuluh, bentuk stomata pada daun, bentuk dan jumlah lapisan sel epidermis, bentuk trikom, jumlah lapisan sel tunika-korpus, susunan sel sklerenkim. Informasi pada organ generatif meliputi bunga, buah dan biji, seperti jenis bunga, bentuk bunga, kelengkapan bunga, dan karakter-karakter dari bagian-bagian bunga (calyx, corolla, epicalyx) dan kelamin bunga (androecium dan gynoecium), bentuk buah, jenis buah, tipe plasenta, jumlah ruang buah, bentuk biji, bentuk dan kedudukan embrio, jumlah dan kedudukan kotiledon, dan sebagainya (Hartono 1989).
Karakterisasi secara morfologi telah banyak dilakukan pada tanaman jarak pagar monoecious. Karakterisasi morfologi sering dikaitkan dengan pencarian
candidate plus tanaman jarak pagar, seperti yang dilakukan Sunil et al. (2008) dan Mishra (2009) yang membuat kriteria morfologi tanaman jarak pagar dan memberi nilai atau skor untuk masing-masing kriteria tersebut. Karakter yang dinilai adalah tinggi tanaman, diameter kanopi, jumlah cabang, dan karakter-karakter produksi seperti jumlah buah, jumlah biji yang dihasilkan. Kegiatan ini dilajutkan oleh Sunil et al. (2013) dengan melakukan deskripsi minimal untuk karakterisasi dan evaluasi plasma nutfah jarak pagar dalam perbaikan tanaman. Hartati et al. (2009) telah mengamati keragaman morfologi jarak pagar di kebun percobaan Pakuwon Sukabumi.
Informasi kimia. Informasi kimia dikenal juga dengan kemotaksonomi tumbuhan, kemosistematik atau fitokimia. Informasi kimia meliputi hasil metabolisme primer, metabolisme sekunder, DNA, RNA, dan protein. Hasil metabolisme primer adalah lemak, protein, karbohidrat, gula, dan asam amino. Hasil metabolisme sekunder seperti vitamin, zat pengatur tumbuh endogen, senyawa alkaloid, fenolik, glukosin, terpen, asam amino tertentu, dan lain-lain. Hasil metabolisme sekunder ini dapat berupa ikatan yang tidak dijumpai pada semua jenis tumbuhan (Hartono 1989).
9 pada kandungan minyak biji, diantaranya Openshaw (2000), Sunil et al. (2008), dan Mishra (2009). Selain itu, analisis kandungan toksin (Tanya et al. 2011), fisikokimia (Parthiban et al. 2011), dan alelopati (Rejila & Vijayakumar 2011) pada jarak pagar telah dilakukan.
Informasi kimia untuk mengidentifikasi tumbuhan meliputi DNA, RNA, protein, dan genetik. Perbedaan genetik tentu saja akan menyebabkan perbedaan tumbuhan baik dari morfologi maupun jenisnya. Perubahan genetik dapat terjadi akibat adanya perubahan lingkungan, perubahan ini menyebabkan mutasi yang dapat diekspresikan secara fenotipik atau tidak. Pendekatan molekuler merupakan pendekatan yang tepat untuk menganalisis keragaman, di mana penanda ini langsung berintegrasi dengan genetik dan menggambarkan genom yang sesungguhnya, sebagai dasar penanda adalah polimorfisme protein dan DNA. Penanda molekuler dapat mendeteksi variasi genetik pada tingkat jaringan atau seluler dan polimorfisme tanpa dipengaruhi oleh faktor lingkungan (Tanskley 1983). Terdapat berbagai macam penanda DNA yang digunakan dalam analisis keragaman tanaman jarak pagar, diantaranya Random Amplified Polymorphyc DNA (RAPD) (Basha et al. 2007, Gangopadhyay et al. 2007, Gupta et al. 2008, Surahman et al. 2009), Inter Simple Sequence Repeats (ISSR) (Kumar et al. 2009), dan Simple Sequence Repeats (SSR) (Whankaew et al. 2011).
Pengembangan marka molekuler selalu dilakukan oleh para peneliti untuk mengidentifikasi keragaman tanaman jarak pagar, diantaranya pengembangan marka molekuler EST-SSR. Expressed sequence tag (EST) merupakan sekuen nukleotida pendek yang digunakan untuk mengidentifikasi sebuah gen yang diekspresikan. Meskipun sekuen EST hanya 200 hingga 500 nukleotida, tetapi cukup untuk mengidentifikasi DNA komplementer (cDNA). Wen et al. (2010) mengembangkan marker ETS-SSR dan genomic-SSR untuk mengakses diversitas genetik jarak pagar. Yadav et al. (2011) mengembangkan marker EST-derived
SSR dalam mengkaji perkembangan, karakterisasi, polimorfisme, dan
transferability pada spesies atau genus jarak pagar. Namun penggunaan EST belum dikembangkan dalam mengidentifikasi tanaman jarak pagar berdasarkan tipe seks.
Informasi kromosom. Informasi kromosom sebagai informasi penting dalam sistematika tumbuhan diantaranya adalah jumlah kromosom, bentuk kromosom, sifat kromosom, serta struktur kromosom. Informasi ini dapat digunakan untuk membedakan antar sepesies dalam satu genus atau di bawah spesies. Sasikala dan Paramathma (2010) melakukan studi terhadap kromosom genus Jatropha, dari 10 spesies yang digunakan 7 spesies memiliki jumlah kromosom 2n=22 sedangkan 3 spesies lainnya memiliki jumlah kromosom 2n=20. Carvalho et al. (2008) meneliti ukuran genom, komposisi dasar dan karyotipe J. curcas. Kariotipe J. curcas terdiri dari 22 kromosom metasentrik dan submetasentrik relatif kecil yang ukurannya berkisar 1.71-1.24 pM.
10
Beberapa penelitian terhadap penyerbukan dan persilangan jarak pagar juga telah dilakukan. Tanaman jarak pagar yang umumnya memiliki tipe seks monoecious dengan bunga betina dan jantan pada infloresen yang sama memungkinkan terjadinya penyerbukan sendiri atau penyerbukan silang tergantung pada polinatornya. Kaur et al. (2011) mengamati sistem breeding tanaman jarak pagar di India dan melihat pengaruhnya pada fruit set. Ahoton dan Quenum (2012) meneliti potensi hibridisasi pada sembilan aksesi jarak pagar yang berasal dari tiga benua (Afrika, Asia, dan Amerika).
Determinasi Sex pada Tanaman
Determinasi atau penentuan seks pada tanaman berbunga adalah proses perkembangan fundamental yang penting secara ekonomis (Guo et al. 2010). Secara umum, ada tiga cara penentuan seks pada angiospermae yaitu pada individu bunga, individu tanaman, dan populasi tanaman. Berdasarkan individu bunga, tipe seks bunga adalah hermaprodit (biseksual), betina, dan jantan. Berdasarkan individu tanaman, tipe seks adalah hermaprodit, monoecious, dioecious, gynoecious, androecious, gynomonoecious, andromonoecious, dan trimonoecious (polygamous). Berdasarkan populasi tanaman, tipe seks adalah hermaprodit, monoecious, dioecious, gynodioecious, androdioecious, dan trioecious (subdioecious) (Dellaporta & Urrea 1993).
Tipe seks andromonoecious dihipotesiskan merupakan suatu mekanisme yang memungkinkan tanaman untuk mandiri dalam mengalokasikan fungsi betina dan jantan. Jika produksi bunga jantan merupakan suatu mekanisme untuk mengatur alokasi untuk fungsi betina (misalnya produksi buah), maka spesies andromonoecious memiliki peluang yang besar untuk menghasilkan buah. Karakter bagian-bagian bunga (ukuran bunga, lebar ovarium, massa buah) berhubungan dengan perubahan ekspresi seksual andromonoecious (Miller & Diggle 2007).
Pada tanaman jarak pagar ditemukan bermacam-macam tipe seks bunga pada satu infloresen, yaitu bunga jantan, betina, dan hermaprodit. Pada satu indvidu tanaman ditemukan bunga jantan dan hermaprodit (andromonoecious), bunga jantan dan betina (monoecious), dan bunga jantan, betina, serta hermaprodit (trimonoecious). Tanaman jarak pagar yang umum ditemukan adalah monoecious, sedangkan andromonoecious dan trimonoecious masih sangat jarang ditemukan.
Pada tumbuhan, sistem penentuan seks berhubungan erat dengan proses evolusi tipe seks dan kromosom seks. Sistem penentuan seks telah berevolusi berkali-kali mulai dari leluhur hermaprodit, lalu tanaman monoecious dengan bunga jantan dan betina terpisah pada individu yang sama. Demikian juga dengan sistem kromosom pada evolusi tanaman berbunga. Konsisten dengan model teoritis untuk transisi evolusioner dari hermaphrodit ke monoecious, pada penentuan tipe seks terlibat beberapa gen termasuk faktor-faktor mandul jantan (male-sterility) dan mandul betina (female-sterility) (Charleswort 1997).
11 rekombinasi antara gen yang menentukan seks jantan dan seks betina, keduanya merupakan gen yang independen, sehingga dioecious bukan berasal dari hermaprodit (Ainsworth et al. 1995).
Evolusi seks juga terlihat pada tanaman jarak pagar. Jarak pagar dikenal dengan spesies yang monoecious dan uniseksual. Pada setiap infloresen terdapat 5-7 cabang infloresen yang menghasilkan bunga jantan dan betina. Distribusi bunga betina diatur oleh center of principle, dan proses pembentukan bunga-bunga jantan dan betina berbeda. Penentuan seks pada jarak pagar sudah dapat dilihat pada tahap awal perkembangan bunga, pemanjangan ujung meristem kelopak akan memperlihatkan perbedaan tipe seks bunga. Untuk itu dikemukakan hipotesis-hipotesis bisexual site origin dan female factors (Irwanto 2006).
Tipe seks pada tumbuhan dipengaruhi oleh beberapa faktor. Penentuan seks dikontrol oleh ekspresi gen seks, hormon tanaman, serta faktor lingkungan (seperti panjang hari dan suhu) yang dapat memodulasi ekspresi gen penentuan seks (Meagher 2007).
Faktor genetik. Pembentukan organ kelamin pada tanaman terkait dengan konsep model ABC dalam pembentukan struktur dasar bunga (sepal, petal, stamen, dan karpel). Keempat organ dasar ini diekspresikan oleh satu atau beberapa gen yang saling berinteraksi. Gen yang terlibat dalam pembentukan sepal adalah tipe A (APETALA2), gen yang terlibat dalam pembentukan petal adalah tipe A dan tipe B (APETALA3/PISTILATA), gen yang terlibat dalam pembentukan stamen adalah tipe B dan tipe C (AGAMOUS), sedangkan gen yang terlibat dalam pembentukan karpel adalah tipe C saja (Robles & Pelaz 2005, Taiz & Zeiger 2010). Gen yang mengendalikan pembentukan organ seks tumbuhan adalah kelompok gen B dan C. Gen B ikut mengendalikan pembentukan stamen atau organ seks jantan, sedangkan gen C ikut mengendalikan pembentukan karpel atau organ seks betina. Jika semua gen ini (gen A, B, dan C) dimiliki oleh tanaman maka dihasilkan bunga lengkap atau bunga hermaprodit (Gambar 2.1).
Gambar 2.1 Konsep gen model ABC dalam meregulasi pembentukan struktur dasar bunga (Taiz & Zeiger 2010)
12
ovule, sedangkan tipe E dikendalikan oleh gen SEPALLATA1-4 (SEP1-4) yang terkait dengan meristem floral yang memfasilitasi loci original dari ABC (Robles & Pelaz 2005, Taiz & Zeiger 2010).
Berdasarkan penelitian-penelitian tentang penentuan seks pada banyak jenis tanaman, telah teridentifikasi keragaman faktor genetik dan epigenetik yang terlibat dalam penentuan tipe seks bunga atau individu. Gen-gen yang mempengaruhi ekspresi seks dapat diketahui dengan cara identifikasi mutasi. Dengan mempelajari interaksi antar gen, dapat diketahui bahwa sinyal seks ikut menentukan dalam mengekspresikan atau menekan ekspresi gen spesifik jantan atau betina (Juarez & Banks 1998).
Variasi bentuk seks mulai dari hermaprodit primitif sampai monoecious sudah diamati pada tanaman sayuran Cucurbita secara ekstensif. Variasi fenotipe seks mentimun ditentukan oleh tiga lokus utama, yaitu lokus F, M, dan A. Masing-masing lokus memiliki penyandian yang berbeda. Lokus F mempengaruhi tingkat kebetinaan (FF>Ff>ff), sedangkan lokus M menentukan uniseksual (M_) atau biseksual (mm) bunga. Kondisi lokus A meningkatkan kecenderungan jantan jika tanaman homozigot resesif aa dan ff. Interaksi antara lokus ini menghasilkan tipe seks dasar yang ditemukan dalam mentimun (Galun 1961).
Analisis penentuan seks pada berbagai tanaman juga telah dilakukan melalui marka molekuler. Deputy et al. (2002) mengembangkan marka molekuler untuk penentuan seks pada pada tanaman pepaya (Carica papaya L.). Pepaya memiliki tiga tipe seks, yaitu jantan, hermaprodit, dan betina. Pengembangan marka molekuler ini terkait dengan Sex1 yang merupakan gen penentu tipe seks. Berdasarkan sequens primer SCAR, SCAR T12 dan SCAR W11 diperoleh produk PCR pada tanaman hermaprodit dan jantan, tetapi jarang pada betina. SCAR T1 menghasilkan produk pada semua tipe seks pepaya. SCAR T12 dan SCAR W11 tidak menunjukkan rekombinasi pada populasi 182 tanaman F2 hasil persilangan
SunUp dengan Kapoho. Berdasarkan hasil PCR untuk menentukan jenis seks pepaya secara cepat dan akurat dikembangkan W11 atau T12 untuk mendeteksi hermaprodit atau alel jantan dan T1. Teknik sexing dengan SCAR T12 dan SCAR T1 sebagai kontrol positif, digunakan untuk memperkirakan tanaman pepaya hermaprodit pada populasi bibit dengan akurasi keseluruhan sebesar 99.2%.
Marka molekuler ISSR juga banyak digunakan untuk mempelajari penentuan seks pada tanaman. Sharma et al. (2008) telah melakukan deteksi jantan dan betina terhadap tanaman jojoba (Simmondsia chinensis) menggunakan marka ISSR. Thatte dan Deothar (2012) melakukan studi pembungaan dan penentuan seks pada Garcinia indica menggunakan marka ISSR. Paolucci et al. (2010) mempelajari peta hubungan genetik dari Populus alba L. dan analisis pemetaan komparatif penentuan seks pada seluruh spesies Populus menggunakan marka AFLP, SSR, RAPD, dan marka spesifik gen, sedangkan Prasanthi et al. (2010) menggunakan marka RAPD dan SCAR untuk menganalisis penentuan seks Simarouba glauca.
13 memberikan pengaruh seks yang berbeda dan juga berbeda pada setiap jenis tumbuhan.
Penelitian pada mentimun (Cucumnis sativus L) dan beberapa spesies lainnya menunjukkan bahwa aplikasi zat pengatur tumbuh mempengaruhi ekspresi seks tanaman. Pada penelitian ini dihipotesiskan bahwa auksin berfungsi sebagai regulator alami ekspresi seks. Kandungan auksin tanaman hermaprodit lebih besar daripada tanaman andromonoecious. Hal ini kompatibel dengan hipotesis yang menyatakan bahwa auksin endogen adalah pengatur ekspresi seks melalui kontrol gen (Galun et al. 1964).
Makwana et al. (2010) melaporkan pengaruh aplikasi GA3 eksogen yang
dapat memodifikasi seks pada J. curcas. Ada peningkatan dalam jumlah bunga betina oleh aplikasi GA3. Pada konsentrasi yang lebih rendah (10 dan 100 ppm),
terjadi peningkatan jumlah bunga betina dan buah yang dihasilkan proporsional dengan konsentrasi GA3 yang digunakan, tetapi pada konsentrasi yang lebih tinggi
(1000 ppm) peningkatan jumlah bunga betina tidak diikuti oleh peningkatan jumlah buah.
Giberelin (GA3) juga dapat menginduksi pembentukan bunga dalam
beberapa hari pada tanaman hari panjang, dan GA3 eksogen dapat menggantikan
perlakuan cahaya pada tanaman hari pendek. Berkenaan dengan LEAFY di
Arabidopsis thaliana, GA3 mempromosikan ekspresi CmFL, sebuah gen
FLORICAULA/LEAFY yang homolog dari krisan, dan upregulation CmFL memerlukan GA3 (Sumitomo et al. 2009). Pada Rhododendron, aplikasi inhibitor
biosintesis giberelin dapat menghambat pembungaan yang merupakan bukti bahwa giberelin menginduksi pembungaan (Sharp et al. 2010). Hal yang sama juga sudah ditemukan Brooking dan Cohen (2002) pada Zantedeschia, bahwa peningkatan konsentrasi dan durasi dari GA3 dapat meningkatkan proporsi tunas
pembungaan. Peningkatan konsentrasi GA3 juga meningkatkan proporsi inisiasi
bunga dan proporsi tunas aksiler yang menghasilkan bunga. Tanaman yang diperlakukan GA3 konsentrasi tinggi dapat berbunga lebih awal dengan jumlah
daun yang lebih rendah. GA3 konsentrasi tinggi mendorong inisiasi bunga lebih
cepat dibandingkan kensentrasi yang lebih rendah. Selain berperan dalam inisiasi bunga, giberelin juga berperan dalam perkembangan bunga.
Sitokinin adalah hormon tanaman dengan fungsi ganda, sebagian besar terkait dengan stimulasi pembelahan sel dan pertumbuhan (Dobrev et al. 2002). Sitokinin mengatur perkembangan bunga melalui aktivasi gen yang mengendalikan aktivitas meristem apeks pucuk (Lindsay et al. 2006). Sitokinin juga terlibat dalam pengaturan buah dan pengisian biji-bijian (Xu et al. 2007). Pada tanaman Vitis vinifera, sitokinin dapat memberikan perkembangan positif dan dapat mempengaruhi ukuran buah (Zabadal & Bukovac 2006).
14
pembentukan tunas aksilar dan terminal bunga. Ethoxyvinylglycine 2-amino (AVG) dan 2,5-norbornadiene (NBD) merupakan inhibitor kerja etilen, aplikasinya membalikkan efek penghambatan ABA pada pembungaan.
Produksi etilen pada setiap bagian bunga berbeda sesuai dengan perkembangan bunga. Produksi etilen pada bunga Eustoma grandiflorum terutama di putik dan kelopak meningkat dengan terjadinya penuaan bunga. Pada saat mekar, bunga tidak peka terhadap etilen, namun menjadi lebih sensitif dengan terjadinya penuaan, sehingga etilen dianggap terlibat dalam penuaan bunga
Eustoma (Ichimura et al. 1998).
Faktor lingkungan. Faktor lingkungan dapat mempengaruhi tipe seks suatu makhluk hidup termasuk tumbuhan. Penentuan tipe seks yang dipengaruhi oleh lingkungan merupakan salah satu hal yang langsung menyebabkan rasio seks zuriat sangat bervariasi (Stehlik et al. 2008). Determinasi seks lingkungan adalah suatu mekanisme seks individu ditentukan setelah pembuahan, sehingga seks suatu individu sesuai dengan lingkungannya (Bull 1981).
Pengaruh faktor lingkungan terhadap penentuan seks tanaman dapat dilihat pada beberapa tanaman, seperti faktor suhu pada mentimun dan glukosa pada paku ekor kuda (Equisetum). Guillon dan Raquin (2002) mempelajari diferensiasi seksual gametofit Equisetum yang berada di bawah pengaruh kondisi lingkungan (pengaruh glukosa atau gula). Namun, isyarat lingkungan yang bertanggung jawab dalam penentuan jenis kelamin gametofit baik secara in vitro ataupun di alam liar masih sulit untuk dipahami. Perbedaan rasio seks diperoleh ketika gametofit ditanam pada media dengan atau tanpa gula. Gametofit jantan melebihi betina pada kondisi eksperimental sukrosa 60-120 mM dan 120 mM glukosa, sedangkan pada ketiadaan gula gametofit betina melebihi gametofit jantan. Pada penelitian lain, aspek-aspek lingkungan induk dapat mempengaruhi rasio seks turunan pada persilangan tanaman dioecious Rumex nivalis (Stehlik et al. 2008). Unsur hara pada media pertumbuhan tanaman paku Woodwardia radicans dapat mempengaruhi ekspresi seks atau pembentukan archegonia dan atheridia (Desoto
15
3 TIPE SEKS PADA BUNGA TANAMAN JARAK PAGAR
(
Jatropha curcas
)
(Flower Sex Type of Jatropha curcas)
Abstract
Jatropha curcas L. produces male, female, and hermaprodit flowers depending on the sex type of plant. The combination of those flowers when present in one plant caused the sex differentiation and determination in Jathropa. This study aimed to explore the sex types of flowers and individual on Jatropha, as well as to see the stability of the sex types after propagation through stem cuttings. Survey was conducted to the population of Jatropha in Kebun Industri Bukit Indah (KIBI) Cikampek and Kebun Induk Jarak Pagar (KIJP) Pakuwon. We found three sex types in both plantations, i.e.: monoecious that produces male and female flowers, andromonocious that produces male and hermaphrodite flowers, and trimonoecious that produces male, female, and hermaphrodite flowers in the same individual plant. Andromonoecious and monoecious Jatropha has stable sex type, but trimonoecious Jatropha has unstable sex type because hermaphrodite flower not necessarily be generated each time flowering. The andromonoecious and monoecious sex types were consistent in four flowering seasons as well as in open pollinated plant. The stability of sex type was in conjuction with number of branch and distance between nodes.
Keywords: andromonoecious, monoecious, trimonoecious
Pendahuluan
Tanaman jarak pagar (Jatropha curcas L) merupakan salah satu anggota dari famili Euphorbiaceae. Tanaman ini berasal dari Meksiko dan Amerika Tengah dan menyebar ke daerah-daerah lain dibawa oleh pelaut Portugis (Heller 1996). Saat ini jarak pagar sudah tersebar di berbagai daerah di Indonesia.
Seiring dengan pengembangan jarak pagar sebagai bahan baku biodiesel, Pusat Penelitian dan Pengembangan Perkebunan telah melakukan eksplorasi tanaman jarak pagar ke beberapa daerah di Indonesia sejak tahun 2005. Eksplorasi dilakukan ke Sumatera Barat, Lampung, Banten, Jawa Barat, Jawa Tengah, Jawa Timur, Nusa Tenggara Barat, Nusa Tenggara Timur, dan Sulawesi. Bahan tanaman hasil eksplorasi tersebut dikoleksi dan ditanam di beberapa Kebun Induk Jarak Pagar (KIJP), yaitu di Pakuwon, Jawa Barat seluas 33.9 ha, di Muktiharjo, Jawa Tengah seluas 13 ha, dan di Asembagus, Jawa Timur seluas 10 ha (Hasnam 2006). Selain Pusat Penelitian dan Pengembangan Perkebunan, perusahaan yang juga ikut mengembangkan tanaman jarak pagar adalah perusahaan-perusahaan yang tergabung di Kawasan Industri Bukit Indah (KIBI) di Cikampek.
16
dan Jawa Barat. Pembungaan tanaman jarak pagar pada kedua perkebunan ini sangat menarik karena ditemukan jarak pagar yang menghasilkan bunga hermaprodit. Bunga hermaprodit merupakan bunga yang jarang ditemukan pada jarak pagar (Heller 1996).
Jarak pagar mempunyai tiga tipe seks bunga yaitu bunga jantan, betina, dan hermaprodit. Ketiga tipe seks bunga tersebut dapat dihasilkan oleh individu tanaman yang sama atau berbeda, maka satu individu tanaman dapat memiliki beberapa tipe seks, yaitu hermaprodit, monoecious, dioecious, gynoecious, androecious, gynomonoecious, andromonoecious, atau trimonoecious. Tanaman hermaprodit adalah tanaman yang menghasilkan bunga hermaprodit saja. Tanaman monoecious adalah tanaman yang menghasilkan bunga jantan dan betina, sedangkan tanaman dioecious menghasilkan bunga jantan dan betina pada individu tanaman yang berbeda. Tanaman gynoecious adalah tanaman yang menghasilkan bunga betina saja, tanaman androecious menghasilkan bunga jantan saja. Tanaman yang menghasilkan bunga betina dan hermaprodit disebut gynomonoecious, sedangkan andromonoecious adalah tanaman yang menghasilkan bunga jantan dan hermaprodit, sedangkan tanaman yang menghasilkan bunga jantan, betina, dan hermaprodit disebut trimonoecious (Dellaporta & Urrea 1993, Miller & Diggle 2007).
Tipe seks individu tanaman yang telah ditemukan pada jarak pagar adalah monoecious, andromonoecious, dan trimonoecious. Penelitian ini bertujuan untuk mengidentifikasi tipe seks dan kestabilannya pada pembungaan jarak pagar di perkebunan KIBI Cikampek dan di KIJP Pakuwon.
Bahan dan Metode
Bahan Tanaman
Bahan tanaman yang diteliti adalah koleksi tanaman jarak pagar di perkebunan KIBI Cikampek, Jawa barat dan KIJP Pakuwon Sukabumi Jawa Barat yang masing-masing mempunyai 5 aksesi jarak pagar (Tabel 3.1). Kestabilan tipe seks diamati pada tanaman hasil stek batang (perbanyakan vegetatif) dari tanaman jarak pagar yang berbunga dari kedua perkebunan dan tanaman hasil penyerbukan terbuka (perbanyakan generatif) dari tanaman yang berbunga pada tanaman hasil stek batang.
Tipe Seks pada Pembungaan Jarak Pagar
Tipe seks pada pembungan jarak pagar diidentifikasi pada bulan Januari-Maret 2010 di perkebunan KIBI Cikampek, sedangkan di perkebunan KIJP Pakuwon Sukabumi diidentifikasi pada bulan April-Juli 2010. Tipe seks diamati pada individu bunga dan individu tanaman. Aksesi jarak pagar yang berbunga diperbanyak melalui stek batang untuk mengetahui kestabilan tipe seksnya.
Kestabilan Tipe Seks
17 diperbanyak melalui stek batang (3 tanaman jarak pagar andromonoecious aksesi Dompu, 3 tanaman jarak pagar monoecious aksesi Dompu, 1 tanaman jarak pagar trimonoecious aksesi Lampung, dan 1 tanaman jarak pagar trimonoecious aksesi Banten). Batang tanaman jarak pagar dipotong-potong sepanjang 20 cm dan ditanam dalam polibag diameter 10 cm dengan media tanaman campuran tanah dan kompos (1:1 v/v). Setiap aksesi dan tipe seks dibuat lebih kurang 70 stek batang. Stek batang dipelihara dengan baik sampai terbentuk akar dan beberapa daun, selanjutnya dipindah tanam ke lapangan dengan jarak tanam 2 m x 2 m. Tanaman hasil perbanyakan secara vegetatif dari jarak pagar di perkebunan KIBI Cikampek ditanam di Dramaga Bogor pada bulan Maret 2010, sedangkan tanaman jarak pagar dari KIJP Pakuwon tetap ditanam di perkebunan tersebut pada bulan Juli 2010. Karakter yang diamati adalah kestabilan tipe seks bunga atau tipe seks tanaman, jumlah dan sudut cabang primer, jumlah dan sudut cabang sekunder. Biji hasil penyerbukan terbuka dari tanaman hasil stek batang kembali ditanam untuk pengamatan kestabilan tipe seks melalui perbanyakan generatif.
Kestabilan tipe seks melalui perbanyakan generatif. Tanaman yang dihasilkan dari penyerbukan terbuka berbagai tipe seks pada kebun jarak pagar di Dramaga Bogor dan KIJP Pakuwon ditanam di Bambu Apus, Pamulang, Tangerang Selatan. Masing-masing diamati 30 tanaman. Karakter yang diamati adalah kestabilan tipe seks bunga atau tipe seks tanaman, jumlah dan sudut cabang primer, jumlah dan sudut cabang sekunder.
Hasil
Tipe Seks pada Pembungaan Jarak Pagar
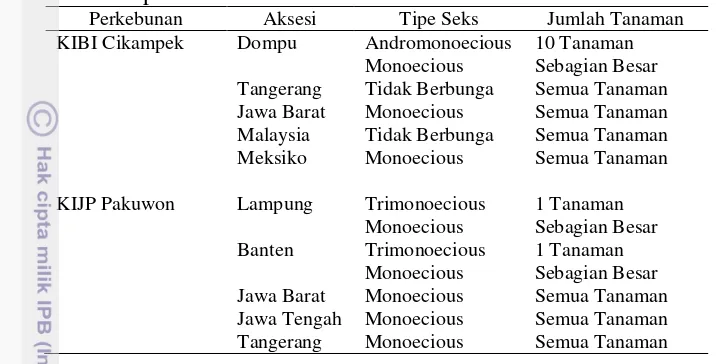
Tipe seks pada pembungaan jarak pagar berdasarkan individu bunga ditemukan bunga jantan, betina dan hermaprodit, sedangkan berdasarkan individu tanaman juga ditemukan tiga tipe seks yaitu monoecious, andromonoecious, dan trimonoecious. Pada koleksi jarak pagar di perkebunan KIBI Cikampek ditemukan dua tipe seks individu tanaman yaitu monoecious dan andromo-noecious. Jarak pagar monoecious menghasilkan bunga jantan dan bunga betina pada infloresen yang sama, sedangkan jarak pagar andromonoecious menghasilkan bunga jantan dan bunga hermaprodit pada infloresen yang sama. Tipe seks yang umum ditemukan pada koleksi jarak pagar di perkebunan KIBI Cikampek adalah monoecious, sedangkan tipe seks andromonoecious hanya 10 tanaman dari aksesi Dompu. Selain itu juga ditemukan dua aksesi jarak pagar yang tidak berbunga sampai berakhir penelitian ini, yaitu aksesi Tangerang dan Malaysia (Tabel 3.1).
18
tersebut memiliki tipe seks trimonoecious yang menghasilkan bunga jantan, bunga betina, dan bunga hermaprodit pada infloresen atau individu yang sama.
Tabel 3.1 Tipe seks individu tanaman koleksi jarak pagar di perkebunan KIBI Cikampek dan KIJP Pakuwon
Perkebunan Aksesi Tipe Seks Jumlah Tanaman KIBI Cikampek Dompu Andromonoecious 10 Tanaman
Monoecious Sebagian Besar Tangerang Tidak Berbunga Semua Tanaman Jawa Barat Monoecious Semua Tanaman Malaysia Tidak Berbunga Semua Tanaman Meksiko Monoecious Semua Tanaman KIJP Pakuwon Lampung Trimonoecious 1 Tanaman
Monoecious Sebagian Besar Banten Trimonoecious 1 Tanaman
Monoecious Sebagian Besar Jawa Barat Monoecious Semua Tanaman Jawa Tengah Monoecious Semua Tanaman Tangerang Monoecious Semua Tanaman
Jarak pagar tipe seks monoecious dan andromonoecious aksesi Dompu di perkebunan KIBI Cikampek cukup berbeda jika dilihat dari percabangan dan profil atau perawakannya. Jarak pagar andromonoecious memiliki percabangan yang banyak dan tersusun rapat pada bagian pangkal batang dan bagian tempat terbentuknya cabang tersier. Jarak pagar monoecious memiliki jumlah cabang yang lebih sedikit dibandingkan andromonoecious. Posisi percabangan pada jarak pagar monoecious ini berjauhan antara satu cabang dengan cabang lainnya, bahkan cabang terbentuk sekitar 30 cm dari pangkal batang. Akibat percabangan yang dihasilkan oleh tanaman ini maka profil jarak pagar andromonoecious lebih rimbun dibandingkan monoecious.
Kestabilan Tipe Seks pada Pembungaan Jarak Pagar
Kestabilan tipe seks melalui perbanyakan vegetatif. Tipe seks diamati pada tanaman jarak pagar hasil perbanyakan vegetatif (jarak pagar monoecious aksesi Dompu 60 tanaman, andromonoecious aksesi Dompu 60 tanaman, dan trimonoecious aksesi Lampung dan Banten masing-masing 60 tanaman). Tipe seks monoecious dan andromonoecious stabil pada semua tanaman jarak pagar yang diperbanyak melalui vegetatif. Namun, tanaman dari jarak pagar trimonoecious menampilkan tipe seks yang bervariasi (Gambar 3.1, Tabel 3.2).
[image:36.595.95.453.151.333.2]19 andromonoecious, dan 73.3% tanaman menampilkan tipe seks trimonoecious. Pada akhir musim berbunga pertama hanya 3.3% tanaman menampilkan tipe seks andromonoecious dan 96.7% tanaman menampilkan tipe seks trimonoecious (Tabel 3.2).
Gambar 3.1 Bunga dan tipe seks tanaman hasil perbanyakan vegetatif jarak pagar andromonoecious (A), monoecious (B), trimonoecious (C), trimo-noecious yang menampil