SELEKSI GENOTIPE JAGUNG (
Zea mays
L.)
GENERASI S
1DAN S
2DI DUA LOKASI
UMI SALAMAH
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Seleksi Genotipe Jagung (Zea mays L.) Generasi S1 dan S2 di Dua Lokasi adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
RINGKASAN
UMI SALAMAH. Seleksi Genotipe Jagung (Zea mays L.) Generasi S1 dan S2 di Dua Lokasi. Dibimbing oleh HAJRIAL ASWIDINNOOR dan WILLY BAYUARDI SUWARNO.
Peningkatan produksi jagung dinilai penting dalam rangka memenuhi kebutuhan pangan, pakan, dan industri saat ini dan masa depan. Salah satu upaya peningkatan produksi yang dapat dilakukan adalah melalui penggunaan varietas jagung unggul. Pembentukan varietas hibrida jagung unggul baru memerlukan galur-galur murni potensial dengan daya gabung yang baik. Tujuan penelitian ini adalah (1) mengidentifikasi galur-galur S1 dan S2 yang memiliki keragaan agronomi yang baik, (2) menduga ragam genetik dan heritabilitas untuk peubah agronomi dan daya hasil, (3) menduga korelasi antar karakter tanaman, (4) menduga tingkat ketidakmiripan genetik antar genotipe uji dan galur-galur murni yang dievaluasi, dan (5) mengevaluasi tingkat inbreeding depression yang terimbangi dengan seleksi dari generasi S1 ke generasi S2.
Penelitian dilaksanakan selama dua musim, masing-masing untuk generasi S1 dan S2. Percobaan musim ke-1 dilaksanakan pada bulan Juni sampai September 2014, sedangkan percobaan musim ke-2 dilakukan pada bulan Januari sampai Mei 2015. Penelitian ditiap musim dilaksanakan di dua lokasi yaitu Kebun Percobaan IPB Leuwikopo dan Kebun Percobaan IPB Cikabayan, Dramaga, Bogor. Percobaan ditiap musim-lokasi menggunakan rancangan augmented kelompok lengkap teracak dengan menggunakan 72 genotipe uji, 3 galur murni, dan 5 varietas hibrida F1 sebagai pembanding.
Hasil percobaan menunjukkan bahwa heritabilitas kategori tinggi generasi S1 dan S2 terdapat pada karakter tinggi tanaman, diameter tongkol, jumlah baris biji, jumlah biji per baris, bobot 100 biji, bobot tongkol per tanaman, bobot biji per tongkol, dan hasil. Heritabilitas kategori sedang adalah umur berbunga betina dan jumlah daun, sedangkan heritabilitas kategori rendah terlihat pada persentase tidak rebah akar dan persentase tidak rebah batang. Terdapat beberapa populasi dasar yang galur-galurnya termasuk 10 terbaik pada generasi S1 dan S2 yaitu P27xNK6326, PERTIWI3xP31, NK6326xP31, P31xNK6326, dan P31xNK33. Karakter produksi generasi S1 dan S2 berkorelasi nyata terhadap karakter tinggi letak tongkol, tinggi tanaman, panjang tongkol, dan diameter tongkol. Pengaruh inbreeding depression terlihat pada karakter hasil namun tidak pada tinggi tanaman. Ketidakmiripan genetik antar ketiga galur murni (Mr4, Mr14, dan Nei9008) relatif besar berdasarkan karakter-karakter kuantitatif yang diamati. Beberapa genotipe uji mengelompok pada masing-masing galur murni yang berbeda (Mr4, Mr14 dan Nei9008). Galur-galur terseleksi dinilai potensial untuk pengembangan varietas jagung unggul baru.
SUMMARY
UMI SALAMAH. Selection of S1 and S2 maize genotypes in two locations. Supervised by HAJRIAL ASWIDINNOOR and WILLY BAYUARDI SUWARNO.
Increasing maize productivity is of importance to meet the need for food, fuel, and industry nowadays and in the future. One approach for increasing yield potential is through utilization of improved maize varieties. Hybrid maize breeding relies on the availability of good performing inbred lines with superior combining ability. The objectives of this study were to (1) identify good performing S1 and S2 maize genotypes, (2) estimate genetic variance and heritability for agronomic and yield traits, (3) estimate correlations of yield and agronomic traits, (4) estimate dissimilarities among the test genotypes and the inbred lines evaluated, and (5) estimate inbreeding depression levels that were balanced with selection from S1 to S2 generations.
The S1 trial was conducted from June to September 2014, and followed by the S2 trial from January to May 2015. Each trial was performed in two locations, namely Cikabayan and Leuwikopo experimental stations of IPB, Dramaga, Bogor. Each trial was arranged in an augmented randomized complete block design with three blocks, consisting of 72 test genotypes, 3 inbred lines, and 5 commercial hybrid varieties as checks.
The results showed that in both S1 and S2 generations, the heritability estimates were high for plant height,ear height, number of kernel rows, number of kernels per row, 100-grain weight, ear weight per plant, grain weight per ear, and grain yield. The heritability estimate were moderate for days to silking and number of leaf, and were low for stem and root lodging. The best performing genotypes in S1 and S2 generations were originated from the base populations of P27xNK6326, PERTIWI3xP31, NK6326xP31, P31xNK6326, and P31xNK33. Grain yield was significantly correlated with ear height, plant height, ear length, and ear diameter. The S2 generation exhibited inbreeding depression for grain yield, but not for plant height. Genetic dissimilarities were relatively large among the three inbred lines (Mr4, Mr14 and Nei9008) based on the quantitative traits observed. The selected genotypes are considered potential for development of new maize varieties.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SELEKSI GENOTIPE JAGUNG (
Zea mays
L.)
GENERASI S
1DAN S
2DI DUA LOKASI
UMI SALAMAH
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PRAKATA
Puji syukur penulis panjatkan kepada Allah SWT atas segala nikmat dan karunia-Nya sehingga penulis dapat menyelesaikan tesis yang berjudul “Seleksi Genotipe Jagung (Zea mays L.) Generasi S1 dan S2 di Dua Lokasi”.
Penulis menyampaikan terima kasih kepada Bapak Dr Ir Hajrial Aswidinnoor, MSc dan Bapak Dr Willy Bayuardi Suwarno, SP MSi selaku pembimbing telah sabar membimbing dan memotivasi penulis dengan sangat luar biasa. Penulis mengucapkan terima kasih kepada Bapak Prof Dr Muhammad Syukur SP MS selaku dosen penguji luar komisi. Penulis ucapkan terima kasih kepada Ibu Dr Ir Yudiwanti Wahyu EK, MS selaku ketua program studi yag telah membantu banyak selama ini terutama saat-saat menjelang ujian.
Penulis mengucapkan terima kasih kepada orang tua yang begitu berpengaruh besar. Untuk Ibu Sumini yang telah memberikan doa yang tiada henti, nasihat, dan semangat yang tulus, terima kasih telah menjadi penegar dalam hidup. Untuk Almarhum Bapak Yakub yang hanya ditemui lewat doa, terima kasih telah menghadirkan ke dunia. Untuk Bapak Bejo yang telah menjadi penuntun kehidupan hingga sekarang, terima kasih atas kesabaran yang tidak dapat diukur. Ketiga orang hebat tersebut yang membuat penulis mampu melalui semuanya dengan baik.
Penulis ucapkan terima kasih kepada dosen pengajar Pemuliaan dan Bioteknologi Tanaman yang telah memberikan ilmunya selama perkuliahan. Penulis ucapkan terima kasih kepada teknisi Kebun Percobaan Cikabayan dan Kebun Percobaan Leuwikopo Institut Pertanian Bogor yang telah membantu terlaksannya penelitian ini. Penulis ucapkan terima kasih yang sebesar-besarnya kepada Direktorat Jenderal Pendidikan Tinggi, Kementrian Pendidikan dan Kebudayaan yang telah memberikan kesempatan melanjutkan studi melalui beasiswa BPP-DN 2013. Penulis ucapkan terima kasih kepada keluarga besar Pemuliaan dan Bioteknologi Tanaman 2013 yang telah membersamai hingga beberapa tahun ini. Penulis ucapkan terima kasih kepada teman-teman yang telah membantu selama penelitian Rony Ramdani dan Rudiansyah yang berkecimpung di lahan selama penelitian. Penulis ucapkan terima kasih kepada sahabat seperjuangan Dia Novita Sari dan Eny Rolenti Togatorop yang telah bersama-sama berjuang dengan keyakinan. Penulis ucapkan terima kasih kepada teman-teman Baju Daerah yaitu Andi Adriani Wahditya, Ratna Ningsih Tarrafanur, Aqlima, Rahmi Hendayani, Desi Ratnasari dan Yudia Azmi serta Gerland Akhmadi yang telah bersam-sama mengisi banyak waktu disini. Penulis ucapkan terima kasih kepada adik-adik kosan An-nur yang telah menjadi saudara. Penulis ucapkan terima kasih kepada Sucipto, Yuli Susanti, dan Tria Ningsih atas diskusi dan pertemuan kembali di sini setelah sekian lama. Dan semua pihak yang terlibat. Semoga karya ilmiah ini bermanfaat.
DAFTAR ISI
DAFTAR TABEL vi
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN viii
1 PENDAHULUAN 1
Latar Belakang 1
Tujuan Penelitian 2
Hipotesis 2
2 TINJAUAN PUSTAKA 3
Asal Usul dan Deskripsi Tanaman Jagung 3
Pemuliaan Tanaman Jagung 4
3 BAHAN DAN METODE 7
Waktu dan Tempat 7
Prosedur Percobaan 7
Analisis Data 11
4 HASIL DAN PEMBAHASAN 14
Keragaan Karakter-Karakter Agronomi Genotipe Jagung S1 dan S2 14
Pendugaan Komponen Ragam dan Heritabilitas Arti Luas 18
Korelasi Antar Karakter Agronomi dan Hasil 24
Pengelompokan Genotipe Generasi S1 dan S2 26
Pendugaan Tingkat Inbreeding Depression yang Diimbangi dengan
Seleksi dari Generasi S1 ke S2 30
5 SIMPULAN 32
DAFTAR PUSTAKA 33
LAMPIRAN 39
DAFTAR TABEL
1 Pengujian sidik ragam rancangan augmented kelompok lengkap
teracak genotipe jagung di dua lokasi 12
2 Rekapitulasi sidik ragam gabungan karakter-karakter genotipe jagung
generasi S1 16
3 Rekapitulasi sidik ragam gabungan karakter-karakter genotipe jagung
generasi S2 17
4 Nilai duga komponen ragam genetik (VG), ragam lingkungan (VE),
ragam interaksi (VGxE) dan heritabilitas h2(bs) gabungan dua lokasi
genotipe jagung generasi S1 19
5 Nilai duga komponen ragam genetik (VG), ragam lingkungan (VE),
ragam interaksi (VGxE), dan heritabilitas (h2bs) gabungan dua lokasi
genotipe jagung generasi S2 20
6 Nilai tengah hasil dan komponen hasil sepuluh genotipe jagung
terbaik generasi S1 21
7 Nilai tengah komponen vegetatif berdasarkan hasil sepuluh genotipe
jagung terbaik tanaman jagung generasi S1 22
8 Nilai tengah hasil dan komponen hasil sepuluh genotipe jagung
terbaik generasi S2 23
9 Nilai tengah komponen vegetatif berdasarkan hasil sepuluh genotipe
jagung terbaik generasi S2 23
10 Koefisien korelasi antar karakter genotipe jagung generasi S1 24
11 Koefisien korelasi antar karakter genotipe jagung generasi S2 25
12 Lima genotipe jagung pembanding (hibrida) generasi S1 ke S2
menggunakan uji t 30
13 Pendugaan inbreeding depression 72 genotipe uji jagung generasi S1
DAFTAR GAMBAR
1 Bentuk ujung daun tanaman jagung 10
2 Lidah daun (ligule) (A) dan bunga jantan jagung (tassel) (B) 10
3 Tipe tassel (A) dan bagian tassel jagung (B) 10
4 Posisi biji dalam baris tongkol jagung 11
5 Dendogram genotipe jagung generasi S1 27
6 Dendogram genotipe jagung generasi S2 28
DAFTAR LAMPIRAN
1 Nama genotipe jagung generasi S1 dan S2 41
2 Pengacakan genotipe jagung dengan rancangan augmented kelompok
lengkap teracak musim 1 Cikabayan 43
3 Pengacakan genotipe jagung dengan rancangan augmented kelompok
lengkap teracak musim 1 Leuwikopo 44
4 Pengacakan genotipe jagung dengan rancangan augmented kelompok
lengkap teracak musim 2 Cikabayan 45
5 Pengacakan genotipe jagung dengan rancangan augmented kelompok
lengkap teracak musim 2 Leuwikopo 46
6 Karakter kualitatif fase vegetatif genotipe jagung generasi S1 47
7 Karakter kualitatif fase generatif genotipe jagung generasi S1 49
8 Karakter kualitatif fase vegetatif genotipe jagung generasi S2 51
9 Karakter kualitatif fase generatif genotipe jagung generasi S2 53
10 Sepuluh genotipe jagung terpilih generasi S1 (musim 1) 55
11 Sepuluh genotipe jagung terpilih generasi S2 (musim 2) 56
12 Tiga galur murni genotipe jagung 57
1 PENDAHULUAN
Latar Belakang
Jagung (Zea mays L.) merupakan salah satu tanaman yang paling penting di dunia. Jagung memiliki posisi unggul karena pemanfaatannya yang sangat luas dalam berbagai bidang, seperti pangan, pakan ternak, bahan bakar, dan industri (Troyer dan Wellin 2009). Produksi jagung pada tahun 2012 sebesar 19 387 022 ton pipilan kering menurun menjadi 18 506 287 ton pada tahun 2013 (BPS 2014). Produksi tahun 2012 lebih tinggi dibandingkan tahun 2013 tetapi, tetap dilakukan impor mencapai 1 797 876 ton. Rendahnya produksi jagung tersebut disebabkan karena berbagai faktor diantaranya adalah penggunaan benih unggul di kalangan petani masih rendah, teknik budidaya yang tidak sesuai dengan anjuran, serangan biotik, dan cekaman abiotik. Peningkatan produksi menjadi sangat penting dalam rangka meningkatkan potensi untuk memenuhi kebutuhan pangan di masa depan memerlukan kompetisi yang kuat, namun kebutuhan sumber daya yang ada masih terbatas (Meng et al. 2013).
Varietas hibrida yang unggul biasanya dicirikan dengan nilai heterosis dan daya hasil yang tinggi (Yustiana et al. 2013). Perakitan varietas unggul jagung banyak dilakukan dengan berbagai cara seperti pendekatan biologi molekuler yang telah berhasil dilakukan dengan mengunakan elektroporasi (D’Halluin et al. 1992), penembak biolistik (Frame et al. 2000), dan Agrobacterium tumefaciens (Negrotto et al. 2000; Frame et al. 2002; Utomo 2005). Namun, pemuliaan tanaman konvensional masih memiliki peran besar dalam pengembangan kultivar tanaman jagung (Campos et al. 2004; Kumar 2002).
Metode yang digunakan dalam program pemuliaan tanaman meliputi pemilihan tetua, hibridisasi, seleksi, evaluasi, dan pengujian daya adaptasi. Pentingnya mengetahui sifat unggul dan kualitas dari suatu tanaman agar dapat mempelajari jenis aksi gen dan transfer sifat. Hal ini akan lebih mudah untuk mempelajari cara kerja gen dan pola pewarisan sifat (Ali et al. 2014). Informasi-informasi populasi yang potensial diperlukan untuk merakit jagung hibrida yang unggul. Populasi yang unggul dibangun dari sumber plasma nutfah yang memiliki keragaman genetik. Adanya jarak genetik antar tetua akan memberikan peluang dalam menghasilkan kultivar baru dengan variabilitas genetik luas (Tenda et al. 2009). Genotipe yang diperoleh dari berbagai sumber untuk menghasilkan varietas yang baik perlu diseleksi pada beberapa lingkungan. Variabilitas genetik yang luas merupakan modal dasar dalam program pemuliaan tanaman (Babic et al. 2011). Penampilan hibrida tergantung pada latar belakang genetik galur murni (Suwarno 2014). Pembentukan galur murni yang potensial perlu dilakukan kegiatan penyerbukan sendiri (selfing) selama beberapa generasi saat tahap seleksi. Menurut Syukur (2012) penyerbukan sendiri cenderung menurunkan persentase heterozigositas dan menurunkan sifat-sifat suatu tanaman dalam populasi yang dikenal dengan inbreeding depression. Tingkat inbreeding depression pada jagung varietas Azam yang tinggi terdapat pada bobot 100 biji (Ahmad et al. 2010).
Hibrida silang ganda diperoleh dari empat galur murni yang berbeda misalnya A, B, C, dan D dapat ditulis sebagai (A x B) x (C x D). Penelitian di Brazil menunjukkan bahwa sekitar 20% dari benih komersil melibatkan kultivar hibrida silang ganda. Hal ini untuk mempertahankan heterosis antar sepasang hibrida silang tunggal. Sobrinho et al. (2002) melaporkan bahwa hibrida silang ganda pada tanaman jagung mampu memberikan hasil yang tinggi dari hibrida silang tunggal terbaik. Catenallos (2011) melaporkan bahwa hibrida silang ganda dapat memperkecil tingkat inbreeding depression. Balestre et al. (2011) menyatakan bahwa informasi mikrosatelit memberikan kemampuan prediksi yang relatif baik pada hibrida silang ganda, dengan korelasi antara nilai-nilai fenotipik yang diamati dan nilai-nilai genotipe berkisar antara 0.27–0.54.
Menurut Ahmad et al. (2010), penyerbukan sendiri (selfing) pada tanaman menyerbuk silang memberikan peluang alel tetua identik yang akan diwariskan kepada turunannya (fillial). Jagung merupakan tanaman dengan tingkat heterozigositas yang tinggi (Ganal et al. 2011), sehingga untuk memperkecil keragaman dilakukan seleksi agar meningkatkan persentase homozigositas. Homozigositas pada tanaman menyerbuk silang akan menghasilkan galur murni. Hibrida yang dihasilkan dari persilangan dari hasil inbreeding dikenal dengan galur murni atau inbred. Menurut Guo et al. (2013) Untuk mendapatkan galur murni terbaik dari hibrida superior perlu dilakukan seleksi dan selfing yang berulang untuk setiap generasi. McClosky et al. (2013) melaporkan bahwa kemajuan seleksi yang diikuti dengan selfing sebesar >70% pada populasi 100-500 tanaman.
Balitsereal telah mengeluarkan 27 varietas jagung hibrida, di antaranya BIMA-17, BIMA-18, BIMA 19-URI (STJ107), dan Bima-20 URI (Balitsereal 2014). Upaya perakitan varietas unggul jagung dengan daya hasil tinggi terus dilakukan, termasuk oleh Balai Penelitian Serealia (Balitsereal) dan Institut Pertanian Bogor. Varietas-varietas jagung baru diharapkan dapat membantu meningkatkan produksi jagung nasional.
Tujuan Penelitian
Tujuan penelitian ini adalah sebagai berikut :
1. Mengidentifikasi galur-galur S1 dan S2 yang memiliki keragaan agronomi
yang baik.
2. Menduga ragam genetik dan heritabilitas untuk peubah agronomi dan daya hasil.
3. Menduga korelasi antar karakter tanaman.
4. Menduga tingkat ketidakmiripan genetik antar genotipe uji dan galur-galur murni yang dievaluasi.
5. Mengevaluasi tingkat inbreeding depression yang terimbangi dengan seleksi dari generasi S1 ke generasi S2.
Hipotesis
2
TINJAUAN PUSTAKA
Asal Usul dan Deskripsi Tanaman Jagung
Jagung (Zea mays L.) merupakan tanaman penting di dunia. Ada beberapa teori tentang asal-usul tanaman jagung. Teori tentang jagung berasal dari Meksiko kemudian menyebar ke utara ke Kanada dan ke selatan sampai Argentina (Brown et al. 1985). Hal ini didukung oleh penemuan fosil tepung sari dan tongkol jagung dalam gua, dan kedua spesies mempunyai keragaman genetik yang luas. Teori lain mengatakan bahwa tanaman jagung yang ada di wilayah Asia diduga berasal dari Himalaya. Hal ini ditandai oleh ditemukannya tanaman keturunan jali (Coix spp.). Tanaman jagung berasal dari dataran tinggi Andean Peru, Bolivia, dan Ekuador. Hal ini didukung oleh hipotesis bahwa jagung berasal dari Amerika Selatan dan jagung Andean mempunyai keragaman genetik yang luas, terutama di dataran tinggi Peru (Bonavia 2013).
Daerah yang memproduksi jagung terbesar adalah Amerika Serikat, Cina, Brasil, dan Meksiko dengan 70% dari produksi dunia. India menyumbang 5% dari areal jagung dan memberikan kontribusi sebesar 2% dari produksi di dunia. Di negara maju, jagung dikonsumsi terutama dalam bentuk daging, telur dan produk susu. Di negara-negara berkembang, jagung dikonsumsi langsung dan berfungsi sebagai makanan diet. Selain itu, jagung diolah menjadi makanan sereal untuk menu sarapan. Namun, dalam bentuk olahan juga jagung dapat dimanfaatkan sebagai bahan bakar (etanol) dan pati (Plessis 2003).
Selain sebagai sumber karbohidrat, jagung juga merupakan sumber protein yang penting dalam menu masyarakat Indonesia. Kandungan gizi utama jagung adalah pati (72–73%), dengan nisbah amilosa (25–30%) dan amilopektin (70– 75%), akan tetapi pada jagung pulut (waxy maize) nisbah amilosa 7% dan amilopektin sebesar 93–100%. Kadar gula jagung (glukosa, fruktosa, dan sukrosa) berkisar 13%. Protein jagung sebesar 8–11% terdiri atas lima fraksi, yaitu albumin, globulin, prolamin, glutelin, dan nitrogen nonprotein (Suarni 2007).
Pemuliaan Tanaman Jagung
Secara alami jagung adalah tanaman menyerbuk silang dengan persentase 99.9%. Tanaman jagung bersifat monoesius, namun bunga jantan jagung matang lebih dahulu dibandingkan bunga betinanya (protandri) (Fehr, 1987). Polen mengalami kematangan (anthesis) selama 4-10 hari sebelum munculnya rambut tongkol (silk). Adanya selang waktu ini memungkinkan polen dari satu tanaman untuk menyerbuki tongkol tanaman yang lain, sedangkan tongkolnya sendiri diserbuki oleh polen tanaman lain.
Informasi lain dalam pembentukan varietas unggul jagung yaitu tentang ketahanan serangan biotik dan cekaman abiotik. Beberapa penelitian tentang ketahanan terhadap serangan biotik yaitu ketahanan terhadap kumbang Maize weevil (Sitophilus zeamais Motschulsky.) pada tanaman jagung (Dari et al. 2010) dan cekaman abiotik yaitu jagung toleran kekeringan (Ziyomo dan Bernado 2012), toleran suhu tinggi (Li et al. 2013), jagung toleran terhadap setres rendaman air (Papathanasiou et al. 2015).
Menurut Allard (1960) untuk memperoleh genotipe baru, keragaman menjadi modal utama untuk perbaikan varietas tanaman. Keberhasilan suatu program pemuliaan tanaman pada hakekatnya sangat bergantung pada keragaman genetik yang diturunkan (Poehlman, 1983). Dengan demikian semakin tinggi keragaman genetik semakin tinggi pula peluang untuk mendapatkan sumber gen bagi karakter yang akan diperbaiki. Populasi yang memiliki keragaman rendah hingga sedang digolongkan sebagai populasi dengan variabilitas genetik sempit sedangkan populasi yang memiliki keragaman cukup tinggi dan tinggi digolongkan dalam variabilitas genetik luas. Menurut Zen dan Bahar (2001), karakter yang memiliki nilai variabilitas genetik luas dapat digunakan dalam perbaikan genotipe, mampu meningkatkan potensi genetik karakter pada generasi selanjutnya sehingga seleksi terhadap karakter tersebut dapat berlangsung secara efektif. Karakter yang memiliki nilai koefisien keragaman genetik rendah sampai sedang menunjukkan bahwa perbedaan genetik dari karakter tersebut masih memiliki keragaman kecil atau dapat dikatakan bahwa keragaman tersebut memiliki genetik yang hampir seragam. Ruchjaningsih et al. (2002) menyatakan bahwa bila suatu keragaman genetik yang dimiliki tanaman bervariabilitas sempit, maka setiap individu dalam populasi tersebut hampir seragam sehingga tidak mungkin dilakukan perbaikan keragaman genetik melalui seleksi.
Evaluasi persilangan antar galur murni merupakan tahapan penting dalam
pengembangan varietas hibrida jagung (Yustiana et al. 2013). Dalam
pembentukan hibrida jagung, pembentukan galur murni dilakukan tahapan penyerbukan sendiri (selfing). Terlepas dari pentingnya penyerbukan sendiri dalam pemuliaan jagung, di Brasil relatif mengalami penurunan hasil akibat depresi tangkar dalam (inbreeding depression) (Hallauer et al 1988). Inbreeding
depression terjadi akibat tanaman yang menyerbuk silang terus menerus
mengalami penyerbukan sendiri yang mungkin sangat membatasi potensi dari frekuensi galur yang potensial dari suatu populasi (Crow dan Kimura 1970). Varietas jagung dengan penyerbukan sendiri pada galur superior atau inferior dapat diidentifikasikan potensi genetiknya. Inbreeding depression dapat diamati pada karakteristik seperti tinggi tanaman, bobot, hasil, komponen hasil, dan sifat-sifat kematangan (Ahmad et al. 2010). Menurut Pacheco et al. (2002) pengurangan hasil yang ditemukan pada populasi jagung generasi S0 ke S1
bervariasi dari 34% menjadi 59% dengan rata-rata penurunan hasil berkisar 49.1%. Inbreeding menawarkan banyak keuntungan tetapi juga menyajikan beberapa kendala untuk perbaikan genetik seperti pada tanaman singkong (Manihot esculenta Crantz) (Rojas et al 2009).
Seleksi adalah memilih serta mencari keturunan pertama tanaman yang memiliki sifat atau karakter yang baik, yang berguna untuk meningkatkan hasil serta mutunya. Karakter-karakter suatu tanaman ditentukan oleh genotipe tetapi ekspresinya dipengaruhi oleh lingkugan. Efektifitas suatu seleksi untuk memperbaiki suatu sifat sangat bergantung pada heritabilitas, intensitas seleksi, dan ragam fenotipe dari populasi seleksi. Karakter dengan heritabilitas tinggi merupakan respon seleksi. Karakter dengan heritabilitas tinggi memberikan respon seleksi yang tidak sama untuk setiap siklus dengan karakter berbeda (Allard 1960).
3 BAHAN DAN METODE
Waktu dan Tempat
Penelitian dilaksanakan di Kebun Leuwikopo Percobaan Institut Pertanian Bogor dengan ketinggian 250 m dpl dan Kebun Percobaan Cikabayan Bawah Percobaan Institut Pertanian Bogor dengan ketinggian 180 m dpl Kabupaten Bogor, Provinsi Jawa Barat. Penelitian dilaksanakan selama dua musim masing-masing untuk percobaan generasi S1 dan S2. Percobaan musim ke-1 dilakukan
pada bulan Juni sampai September 2014, sedangkan percobaan musim ke-2 dilakukan pada bulan Januari sampai Mei 2015.
Alat dan Bahan
Materi genetik yang digunakan dalam penelitian ini adalah 72 galur S1
berasal dari hibrida silang ganda untuk musim ke-1 dan 72 galur S2 hasil selfing
S1 untuk musim ke-2. Setiap percobaan menggunakan 3 galur murni koleksi
Balitsereal (Mr4, Mr14 dan Nei9008) dan 5 varietas hibrida F1 sebagai
pembanding (NK33, NK6326, Pertiwi3, P21, dan P27). Percobaan ditiap lokasi-musim disusun berdasarkan rancangan augmented kelompok lengkap teracak dengan 75 genotipe uji tidak diulang dan lima varietas pembanding (check) diulang tiga kali. Areal percobaan dibuat dalam tiga blok, pada masing-masing blok terdapat 30 plot percobaan. Genotipe uji ditempatkan secara acak pada seluruh blok dan kelima genotipe pembanding ditempatkan pada masing-masing blok.
Prosedur Percobaan
Persiapan tanam dilakukan dengan mengolah tanah menggunakan mesin bantu traktor. Luas lahan yang digunakan adalah 500 m2. Jarak tanam yang digunakan adalah 70 cm antar baris tanaman dan 20 cm antar tanaman dalam baris. Setiap baris tanaman diberikan label percobaan sesuai dengan genotipe yang digunakan. Penanaman dilakukan dengan terlebih dahulu melubangi sedalam 3 cm menggunakan tugal. Jumlah benih yang ditanam sebanyak 2 benih per lubang tanam.
disiangi secara manual menggunakan alat pemotong rumput. Pengendalian hama penyakit dilakukan dengan menggunakan pestisida berbahan aktif karbofuran 17
kg ha-1. Penyakit yang menyerang tanaman jagung adalah bulai
(Peronosclerospora maydis dan P. turcicum). Tanaman yang telah di serang
penyakit bulai dilakukan dengan mencabut dan dijauhkan dari lahan percobaan. Hal ini dilakukan agar tidak menyerang tanaman yang lainnya. Pemanenan dilakukan setelah tanaman berumur 110 hari. Tanaman yang sudah memasuki masa panen memiliki ciri-ciri yaitu warna kelobot telah mongering, biji mengkilap, dan terlihat ada lapisan hitam (black layer) pada pangkal yang menempel pada tongkol di biji serta tingkat kehijauan tanaman menurun.
Peubah pengamatan sebagian besar diamati pada 10 tanaman contoh yang diambil secara acak di setiap satuan percobaan (plot). Peubah pengamatan yang akan diamati berdasarkan Descriptors for Maize (CIMMYT 1991) adalah sebagai berikut :
A. Peubah Kuantitatif
Peubah kuantitatif yang akan diamati adalah sebagai berikut :
1. Tinggi tanaman (cm). Pengamatan dilakukan setelah stadia pembungaan pada bunga jantan dan betina menggunakan meteran, diukur dari permukaan tanah sampai dasar bunga jantan (tassel).
2. Tinggi letak tongkol. Diukur bersamaan dengan pengukuran tinggi tanaman, diukur dari permukaan tanah hingga pangkal tongkol utama (tongkol yang paling atas).
3. Jumlah daun. Pengamatan dilakukan setelah fase pembungaan.
4. Jumlah daun di atas daun tongkol (termasuk teratas). Daun tongkol adalah daun yang berada tepat di bawah tongkol, seringkali agak menutupi tongkol. Pengamatan dilakukan setelah tahap susu (milky stage).
5. Panjang daun (cm). Diukur dari ligule (lidah daun) ke ujung daun. Dipilih dari daun yang berada tepat di bawah tongkol teratas. Pengamatan dilakukan setelah fase pembungaan.
6. Umur berbunga jantan (days to tasseling) (hst). Pengamatan dilakukan pada fase pembungaan saat anthesis (munculnya kotak sari) dengan persentase 50% tanaman dalam setiap baris.
7. Umur berbunga betina (days to silking) (hst). Pengamatan dilakukan pada fase pembungaan saat keluar rambut dengan persentase 50% tanaman dalam setiap baris.
8. Panjang tassel (cm). Pengamatan dilakuakan setelah tahap susu (milky stage) setelah tongkol keluar rambut.
9. Jarak cabang tassel (cm). Jarak antara cabang primer pertama dan terakhir. Pengamatan dilakuakan setelah tahap susu.
10.Jumlah cabang utama tassel. Pengamatan dilakuakan setelah tahap susu (milky stage).
11.Persentase tidak rebah akar (%). Jumlah tanaman tidak rebah akar dibagi jumlah tanaman seluruhnya di dalam plot. Diamati dua minggu sebelum panen (sekitar umur 96 hst).
13.Diameter tongkol (cm). Pengamatan dilakukan setelah panen dengan menggunakan jangka sorong diukur tepat di tengah-tengah tongkol.
14.Panjang tongkol (cm). Pengamatan dilakukan setelah panen menggunakan penggaris diukur dari pangkal tongkol hingga ke ujung tongkol.
15.Jumlah baris biji dalam tongkol. Pengamatan diukur dengan menghitung baris biji dalam setiap tongkol.
16.Jumlah biji dalam baris. Pengamatan dilakukan dengan menghitung jumlah biji dalam satu baris per tongkol.
17.Panjang biji (mm). Rata-rata dari 10 biji yang diambil berturut-turut dari satu baris di tengah tongkol, diukur menggunakan jangka sorong.
18.Lebar biji (mm). Diukur pada 10 biji seperti pada panjang biji.
19.Tebal biji (mm). Diukur pada 10 biji seperti pada panjang biji.
20.Bobot tongkol per tanaman (g). Pengamatan dengan menimbang tongkol utama (tongkol paling besar) yang sudah dibuka klobotnya tetapi belum dipipil.
21.Bobot biji per tongkol (g). Pengamatan dengan menimbang biji dari tongkol yang telah dipipil.
22.Bobot 100 biji (g). Pengamatan dilakukan dengan menimbang bobot 100 biji.
23.Rendemen biji (%). Pengamatan dilakukan dengan mengukur persentase bobot biji per tongkol terhadap bobot tongkol.
24.Kadar air biji panen (%). Biji hasil pipilan dicampur kemudian diukur kadar airnya dengan menggunakan digital seed moisture tester.
25.Produksi (ton ha-1). Dihitung menggunakan rumus berikut:
Hasil (ton ha-1)
=
x B x JT x 0.7Keterangan :
KA = Kadar air biji kering
JT = Jumlah tanaman per ha (71 000 tanaman)
B = Bobot biji per tongkol (asumsinya, satu tanaman memiliki satu tongkol produktif)
B. Peubah Kualitatif
Peubah kualitatif yang akan diamati adalah sebagai berikut :
1. Warna antosianin pada pelepah daun. Pengamatan dilakukan pada daun pertama dengan skor penilaian: (1) tidak ada atau sangat lemah (3) lemah (5) sedang (7) kuat (9) sangat kuat.
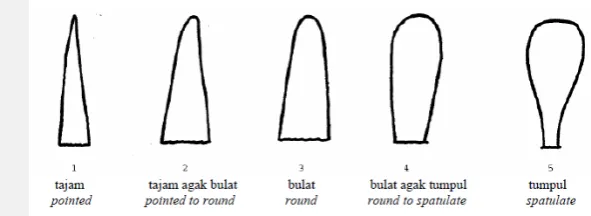
2. Bentuk ujung daun. Pengamatan dilakukan pada daun pertama dengan skor runcing dengan skor penilaian: (1) tajam (2) tajam agak bulat (3) bulat (4) bulat agak tumpul (5) tumpul.
3. Warna antosianin batang. Skor pengamatan dengan (1) tidak ada atau sangat lemah (3) lemah (5) sedang (7) kuat sangat (9) kuat.
Gambar 1 Bentuk ujung daun tanaman jagung
5. Orientasi daun. Pengamatan dilakukan setelah fase berbunga. Skor pengamatan: (1) tegak (2) terkulai.
6. Lidah daun (leaf ligule). Pengamatan dilakukan setelah fase berbunga. Skor pengamatan: (1) ada (0) tidak ada.
(A) (B)
Gambar 2 Lidah daun (ligule) (A) dan bunga jantan jagung (tassel) (B)
7. Urat daun. Diamati pada daun yang berada di bawah tongkol. Skor pengamatan: (1) ada (3) tidak ada.
8. Tipe tassel. Pengamatan dilakukan pada tahap susu. Skor pengamatan (1) primer (2) primer-sekunder (3) primer-sekunder-tersier (lihat Gambar. 3) 9.
(A) (B)
Gambar 4 Posisi biji dalam baris tongkol jagung
10.Ukuran tassel. Pengamatan dilakukan setelah tahap susu (milky stage). Skor pengamatan (3) kecil (5) sedang (7) besar.
11.Tipe biji. Pengamatan dilakukan setelah panen. Skor pengamatan: (1) mutiara (flint) (2) intermediet, di antara mutiara dan gigi kuda (dent), tetapi lebih dekat dengan dent (3) gigi kuda (dent).
12.Warna tongkol. Pengamatan dilakukan saat panen. Skor pengamatan: (1) putih (2) merah (3) coklat (4) ungu (5) variegata (6) lainnya.
13.Warna pericarp. Skor pengamatan: (1) tidak berwarna (2) putih keabu-abuan (3) merah (4) coklat (5) lainnya.
14.Warna endosperma. Skor pengamatan: (1) putih (2) kuning muda (3) kuning tua (5) oranye.
Analisis Data
Analisis Ragam Gabungan
Analisis ragam gabungan dilakukan dengan uji F untuk melihat pengaruh utama genotipe, lokasi, dan pengaruh interaksi genotipe x lokasi. Analisis dilakukan dengan menggunakan software SAS 9.1. Model linier untuk analisis gabungan antar lokasi memiliki persamaan sebagai berikut, dan garis besar sidik ragamnya ditampilkan pada Tabel 1.
Yijk = µ + Li + B(L)ji + Gk + GLki + εijk
Keterangan :
Yijk = Nilai pengamatan pada lingkungan ke-i, kelompok ke-j, dan genotipe ke-k
µ = Nilai umum
Li = Pengaruh lingkungan ke-i, dimana i=1,2
B(L)ji = Pengaruh kelompok ke-j dalam lingkungan ke-i, dimana j=1,2,3
Gk = Pengaruh genotipe ke-k, dimana k =1,2,3,....80
GLki = Pengaruh interaksi genotipe ke-k dan lingkungan ke-i
Tabel 1 Sidik ragam rancangan augmented kelompok lengkap teracak pengujian
E(KT) = harapan (kuadrat tengah); l = jumlah lokasi, r = jumlah ulangan, g = banyaknya genotipe uji, c = banyaknya genotipe kontrol.
Pendugaan Komponen Ragam dan Nilai Heritabilitas
Nilai heritabilitas tanaman yang dihitung merupakan heritabilitas dalam arti luas yaitu perbandingan ragam genetik dan ragam fenotipe (Syukur et al. 2012). Pendugaan komponen ragam yaitu ragam genotipe, ragam lingkungan, dan interaksi genetik x lingkungan berdasarkan nilai harapan kuadrat tengah pada Tabel 1 dihitung dengan SAS PROC GLM. Nilai heritabilitas disajikan dalam rumus sebagai berikut:
KTG = Kuadrat tengah genotipe
KTgalat = Kuadrat tengah galat
KTGxE = Kuadrat tengah interaksi (genotipe x lokasi)
Kriteria nilai heritabilitas arti luas (h2bs) dikelompokkan menurut Stanfield
(1983): 0.50 < h2bs < 1.00 = tinggi, 0.20 < h2bs < 0.50 = sedang, dan h2bs < 0.20 =
Seleksi Genotipe Terbaik Generasi S1 dan S2
Genotipe generasi S1 dan S2 diseleksi berdasarkan rataan produksi dan
diseleksi sebanyak 10 genotipe. Jika terdapat perbedaan nilai, dilakukan uji lanjut BNT 0.05 + rataan genotipe uji.
Korelasi Antar Peubah Agronomi dan Daya Hasil
Nilai korelasi antar peubah agronomi dan daya hasil dapat diduga dengan menggunakan rumus sebagai berikut :
r , = Cov
V . V
Keterangan:
r(X1,X2) = Korelasi antara X1 dan X2
Cov.X1.X2 = Kovarian antara X1 dan X2
V(X1) = Ragam X1
V(X2) = Ragam X2
Pengelompokan Genotipe
Pengelompokan genotipe dilakukan dengan menggunakan analisis gerombol. Analisis gerombol dilakukan untuk mengetahui koefisien ketidakmiripan genetik antar genotipe menggunakan metode Gower. Data yang digunakan dalam pengelompokan adalah data kuantitatif. Perhitungan jarak gerombol dilakukan dengan menggunakan metode average linkage menggunakan software R 3.1.1.
Eevaluasi Tingkat Inbreeding Depression yang diimbangi dengan Seleksi dari
Generasi S1 ke Generasi S2
Pendugaan inbreeding depression pada tanaman jagung dari generasi S1 ke
generasi S2 dilakukan melalui uji t, menggunakan perangkat lunak SAS 9.1.
Pengaruh musim dipelajari melalui uji t antara nilai tengah pembanding di musim pertama (S1) dengan nilai tengah pembanding di musim kedua (S2). Jika pengaruh
musim nyata dilakukan perhitungan nilai tengah rataan genotipe terseleksi
(adjusted entry means). Perhitungan ini dilakukan dengan mengurangi nilai
4 HASIL DAN PEMBAHASAN
Keragaan Karakter-Karakter Agronomi Genotipe Jagung S1 dan S2
Karakter merupakan cerminan setiap individu tanaman. Setiap individu terbentuk dapat dilihat dari seberapa besar karakter yang diamati. Hasil analisis ragam gabungan dua lokasi menunjukkan bahwa setiap karakter memiliki pengaruh yang berbeda-beda pada lokasi, kelompok, genotipe, dan lokasi x genotipe. Partisi lokasi x genotipe meliputi galur (line), hibrida pembanding (check), interaksi galur x pembanding, interaksi lokasi x pembanding, interaksi lokasi x galur, dan interaksi lokasi x (pembanding vsgalur) (Tabel 2 dan Tabel 3).
Pengaruh Lokasi dan Blok (lokasi) Terhadap Keragaan Karakter-Karakter
Agronomi Genotipe Jagung S1 dan S2
Pengaruh lokasi nyata untuk sebagian besar karakter yang diamati pada musim pertama, kecuali karakter rendemen biji, bobot 100 biji, dan kadar air sedangkan pengaruh blok (lokasi) menunjukkan tidak berbeda nyata pada sebagian besar karakter. Pengaruh blok (lokasi) yang sangat nyata terdapat pada karakter umur silking, umur anthesis, dan diameter tongkol sedangkan pengaruh blok (lokasi) yang nyata terdapat pada karakter tinggi letak tongkol, panjang tassel, persentase tidak rebah akar, bobot biji per tongkol, panjang biji, rendemen biji, dan hasil (Tabel 2).
Pengaruh lokasi nyata untuk sebagian besar karakter yang diamati pada musim kedua, kecuali karakter tidak rebah batang, panjang biji, lebar biji, rendemen biji, bobot 100 biji, dan kadar air. Pada musim kedua, blok (lokasi) sangat berpengaruh nyata pada karakter umur berbunga betina, umur berbunga jantan, jumlah daun di atas tongkol, dan produksi, berpengaruh nyata terhadap karakter tinggi letak tongkol, panjang daun, panjang biji, tebal biji, bobot tongkol per tanaman, bobot biji per tongkol, rendemen biji, dan bobot 100 biji (Tabel 3).
Pengaruh Genotipe Terhadap Keragaan Karakter-Karakter Agronomi
Genotipe Jagung S1 dan S2
Pengaruh genotipe musim kedua menunjukkan nyata pada beberapa karakter kecuali pada karakter jarak tinggi tanaman cabang tassel, persentase tidak rebah akar, persentase tidak rebah batang, panjang tongkol, diameter tongkol, lebar biji, dan kadar air. Pengaruh genotipe nyata dan ketiga partisi nyata terdapat pada karakter jumlah daun, jumlah daun di atas tongkol, panjang daun, panjang tassel, jumlah cabang utama tassel, jumlah baris biji, panjang biji, bobot tongkol per tanaman, dan rendemen biji. Pengaruh genotipe nyata karena pengaruh galur dan pengaruh rata-rata pembanding dan rata-rata galur nyata terdapat pada karakter umur berbunga betina, umur berbunga jantan, tinggi letak tongkol, jumlah biji per baris, bobot 100 biji, dan hasil. Pengaruh genotipe tidak nyata tetapi pengaruh rata-rata pembanding dan rata-rata galur nyata terdapat pada karakter jarak cabang tassel, panjang tongkol, dan diameter tongkol (Tabel).
Pengaruh Interaksi Lokasi x Genotipe Terhadap Keragaan
Karakter-Karakter Agronomi Genotipe Jagung S1 dan S2
Adanya interaksi genotipe dan lingkungan memerlukan evaluasi genotipe berbagai lingkungan untuk menemukan genotipe yang diinginkan (Zali et al. 2008). Interaksi lokasi x genotipe musim pertama berpengaruh nyata pada karakter umur berbunga, panjang daun, panjang tassel, dan lebar biji berpengaruh sangat nyata terhadap karakter jumlah daun, jarak cabang tassel, jumlah cabang utama tassel, diameter tongkol, panjang tongkol, jumlah baris biji, panjang biji, dan rendemen biji. Interaksi genotipe x lokasi terdiri dari tiga partisi meliputi lokasi x galur, lokasi x pembanding, dan lokasi x (galur vs pembanding) (Tabel 2).
Interaksi lokasi x genotipe nyata yang hanya dipengaruhi oleh interaksi lokasi x galur yaitu terdapat pada karakter jumlah daun per tanaman, panjang daun, panjang tassel, jarak cabang tassel, lebar biji, dan rendemen biji. Hal ini menunjukkan bahwa interaksi genotipe x lokasi dipengaruhi oleh interaksi rata-rata galur yang berbeda antar lokasi. Interaksi lokasi x genotipe nyata dengan pengaruh interaksi lokasi x galur dan pengaruh interaksi lokasi x (galur vs pembanding) nyata yaitu terdapat pada karakter umur berbunga betina, umur berbunga jantan, dan panjang biji. Hal ini menunjukkan bahwa interaksi lokasi x genotipe dipengaruhi oleh interaksi galur dan lokasi dengan rata-rata galur dan pembanding yang berbeda antar lokasi. Interaksi lokasi x genotipe nyata dengan pengaruh interaksi lokasi x pembanding dan pengaruh interaksi lokasi x galur nyata terdapat pada karakter jumlah cabang utama tassel, bobot 100 biji, dan diameter tongkol (Tabel 2).
Tabel 2 Rekapitulasi sidik ragam gabungan karakter-karakter genotipe jagung generasi S1
Peubah
** = Berbeda sangat nyata pada taraf 1% uji F, * = Berbeda nyata pada taraf 5% uji F, tn = tidak berbeda nyata pada taraf 5% uji F; KK = Koefisien keragaman.
karakter panjang daun dan panjang biji. Interaksi lokasi x genotipe nyata dipengaruhi lokasi x galur dan lokasi x (galur vs pembanding) terdapat pada karakter tinggi tanaman, tebal biji, dan rendemen biji. Hal ini menunjukkan bahwa interaksi lokasi x genotipe dipengaruhi oleh interaksi galur dan lokasi dengan rata-rata galur dan pembanding yang berbeda antar lokasi. Interaksi lokasi x genotipe nyata yang hanya dipengaruhi oleh interaksi lokasi x (galur vs pembanding) terdapat pada karakter bobot tongkol per tanaman. Hal ini menunjukkan bahwa interaksi lokasi x genotipe nyata yang hanya dipengaruhi oleh rataan pembanding dan galur yang berbeda antar lokasi. Interaksi lokasi x genotipe tidak nyata maka, akan diikuti oleh lokasi x galur, lokasi x pembanding, dan lokasi x (galur vs pembanding) yang tidak nyata juga.
Interaksi GxL harus dipertimbangkan oleh pemulia tanaman untuk mengembangkan varietas dengan daya hasil tinggi karena respon genotipe tidak sama di tiap lokasi (Lestari et al. 2010). Sejalan dengan penelitian Yustiana et al. (2013) bahwa nilai kuadrat tengah ragam interaksi GxE berbeda nyata untuk semua karakter yang diamati kecuali panjang tongkol dan potensi hasil. Pengaruh yang tidak nyata pada interaksi GxE menunjukkan bahwa genotipe-genotipe yang memberikan hasil yang tinggi di satu lokasi juga menunjukkan hasil yang relatif sama di lokasi lain (Rifianto et al. 2013). Menuru Boer (2011), dalam pendugaan komponen ragam, nilai keragaman yang timbul akibat adanya pengelompokan berfungsi sebagai lokal kontrol dalam memperkecil galat percobaan atau memperkecil nilai keragaman yang terjadi akibat adanya kesalahan percobaan sehingga ragam akibat perbedaan kultivar menjadi tidak berbias. Koefisien keragaman (KK) pada semua karakter kedua musim menunjukkan kisaran 2%– 30%. KK tertinggi pada karakter bobot tongkol per tanaman pada musim 1 dan bobot biji per tongkol pada musim kedua. Sedangkan KK terendah terdapat pada karakter umur berbunga pada musim pertama dan rendemen biji pada musim kedua.
Pendugaan Komponen Ragam dan Heritabilitas Arti Luas
Pendugaan komponen ragam fenotipik, genotipik, dan lingkungan akan menentukan nilai duga heritabilitas genotipe jagung. Heritabilitas sangat penting dalam menentukan metode seleksi. Nilai heritabilitas yang tinggi menandakan besarnya pengaruh genotipe terhadap karakter yang diamati (Ordas et al. 2012).
Heritabilitas kategori tinggi generasi S1 terdapat pada karakter umur bunga
jantan, tinggi tanaman, tinggi letak tongkol, panjang daun, panjang tassel, jarak cabang tassel, jumlah cabang primer tassel, diameter tongkol, jumlah baris biji, jumlah biji per baris, bobot 100 biji, bobot tongkol per tanaman, bobot biji per tongkol, lebar biji, dan hasil. Heritabilitas yang tinggi menunjukkan bahwa faktor genetik lebih dominan dibandingkan lingkungan. Heritabilitas tinggi terdapat pada bobot 100 biji, hasil, tinggi tanaman, jumlah baris biji, jumlah daun, bobot biji per tongkol, jumlah baris biji, panjang tassel, dan panjang tongkol (Hijria et al. 2012; Prado et al. 2013; Sultan et al. 2014). Heritabilitas yang tinggi akan memiliki kemampuan dalam perbaikan genetik dari setiap karakter dalam populasi (Ordas et al. 2012). Karakter yang memiliki nilai heritabilitas arti luas kategori sedang pada generasi S1 adalah umur bunga betina, jumlah daun, jumlah daun di atas
memiliki nilai heritabilitas kategori rendah generasi S1 yaitu karakter persentase
tidak rebah akar, persentase tidak rebah batang, dan panjang biji.
Tabel 4 Nilai duga komponen ragam genetik (VG), ragam lingkungan (VE), ragam
interaksi (VGxE) dan heritabilitas h2(bs) gabungan dua lokasi genotipe
jagung generasi S1
Karakter VE VG VGXE h2(bs) Kriteria
Umur berbunga betina (hst) 2.53 1.16 4.13 26.49 Sedang
Umur berbunga jantan (hst) 2.44 2.57 2.87 50.25 Tinggi
Tinggi letak tongkol (cm) 132.60 110.13 28.08 59.71 Tinggi
Tinggi tanaman (cm) 249.02 182.68 46.09 57.28 Tinggi
Jumlah daun 0.75 0.39 1.95 23.05 Sedang
Jumlah daun di atas tongkol 0.22 0.09 0.02 44.60 Sedang
Panjang daun (cm) 22.62 28.92 35.56 50.76 Tinggi
Panjang tassel (cm) 4.90 10.30 6.74 64.80 Tinggi
Jarak cabang tassel (cm) 0.78 2.42 1.58 67.83 Tinggi
Jumlah cabang primer tassel 1.05 6.40 2.15 80.51 Tinggi
Persentase tidak rebah batang (%) 186.01 3.48 -33.59 (0)# 3.95 Rendah Persentase tidak rebah akar (%) 130.22 -1.64 (0)# 7.23 0.00 Rendah
Jumlah baris biji 0.77 4.80 2.27 76.38 Tinggi
Jumlah biji per baris 7.49 12.58 5.53 67.08 Tinggi
Panjang tongkol (cm) 1.35 1.34 6.47 25.84 Sedang
Diameter tongkol (cm) 0.02 0.08 0.08 63.16 Tinggi
Bobot 100 biji (g) 17.15 34.11 11.29 71.73 Tinggi
Bobot tongkol per tanaman (g) 938.69 902.61 74.20 66.06 Tinggi
Bobot biji per tongkol (g) 531.23 579.80 -85.59 (0)# 70.59 Tinggi
Panjang biji (mm) 0.27 0.82 (0)# 47.64 0.00 Rendah
Lebar biji (mm) 0.42 0.53 0.58 52.29 Tinggi
Tebal biji (mm) 0.31 0.15 0.06 46.55 Sedang
Rendemen biji (%) 12.21 11.70 58.49 25.17 Sedang
Kadar air (%) 4.20 0.96 -0.89 (0)# 33.57 Sedang
Hasil (ton ha-1) 1.02 1.31 -0.03 (0)# 73.87 Tinggi
#
= nilai minus untuk ragam dianggap nol
Karakter yang memiliki nilai heritabilitas minus (dianggap nol) dikategorikan kedalam heritabilitas rendah seperti pada karakter persentase tidak rebah akar dan panjang biji. Heritabilitas yang rendah disebabkan karena pengaruh lingkungan yang tinggi (Sadras dan Slafer 2012). Dalam penelitian jagung manis, Sujiprihati et al. (2005) juga mendapati nilai nol pada varian genetik dan heritabilitas dalam arti luas untuk sifat diameter tongkol. Menurut Allard (1960) dalam perhitungan nilai ragam nol disebabkan karena nilai kuadrat tengah genotipe lebih kecil dibandingkan dengan nilai kuadrat tengah interaksi genotipe dan lingkungan, sehingga perhitungan ragam genetik tersebut merupakan hasil perhitungan pendugaan dari kuadrat nilai tengah harapan.
Heritabilitas kedua musim sebagian besar menunjukkan kategori yang sama pada generasi S1 dan S2 terutama pada heritabilitas kategori tinggi.
Heritabilitas generasi S2 kategori tinggi terdapat pada karakter tinggi tanaman,
tassel, diameter tongkol, jumlah baris biji, jumlah biji satu baris, bobot 100 biji, bobot tongkol per tanaman, bobot biji per tongkol, tebal biji, dan hasil.
Tabel 5 Nilai duga komponen ragam genetik (VG), ragam lingkungan (VE), ragam
interaksi (VGxE), dan heritabilitas (h2bs) gabungan dua lokasi genotipe
jagung generasi S2
Karakter VE VG VGXE h2(bs) Kriteria
Umur berbunga betina (hst) 10.01 7.02 17.17 34.83 Sedang
Umur berbunga jantan (hst) 9.10 5.70 10.99 37.16 Sedang
Tinggi letak tongkol (cm) 110.03 149.52 37.20 68.54 Tinggi
Tinggi tanaman (cm) 1.02 1.31 -0.03 (0)# 73.86 Tinggi
Jumlah daun 0.40 0.43 1.08 37.18 Sedang
Jumlah daun di atas tongkol 0.14 0.16 0.06 62.94 Tinggi
Panjang daun (cm) 28.24 35.12 48.24 48.73 Sedang
Panjang tassel (cm) 5.77 15.24 4.63 75.53 Tinggi
Jarak cabang tassel (cm) 6.13 2.17 -2.71 (0) 43.76 Sedang
Jumlah cabang utamatassel 1.28 3.58 1.12 75.80 Tinggi
Persentase tidak rebah batang (%) 29.77 -1.67 (0)# -1.80 (0)# 0.00 Rendah
Persentase tidak rebah akar (%) 36.08 1.95 19.47 6.94 Rendah
Jumlah baris biji 0.87 1.28 0.23 71.46 Tinggi
Jumlah biji per baris 11.54 28.01 -0.27 (0)# 84.22 Tinggi
Panjang tongkol (cm) 8.03 3.48 -4.09 (0)# 48.80 Sedang
Diameter tongkol (cm) 28.73 16.18 -4.10 (0)# 55.33 Tinggi
Bobot 100 biji (g) 9.35 13.03 7.04 62.63 Tinggi
Bobot tongkol per tanaman (g) 372.72 1659.37 490.67 80.00 Tinggi
Bobot biji per tongkol (g) 446.85 895.01 230.45 73.76 Tinggi
Panjang biji (mm) 0.30 0.70 1.19 48.82 Sedang
Lebar biji (mm) 0.37 0.12 -0.02 (0)# 41.02 Sedang
Tebal biji (mm) 0.14 0.23 0.22 57.55 Tinggi
Rendemen biji (%) 12.96 4.80 44.21 14.65 Rendah
Kadar air (%) 6.33 0.23 -1.01 (0)# 7.28 Rendah
Hasil (ton ha-1) 0.85 1.98 0.56 74.81 Tinggi
#
= nilai minus untuk ragam dianggap nol.
Heritabilitas yang tinggi menunjukkan bahwa faktor genetik lebih tinggi dibandingkan lingkungan. Sejalan dengan penelitian Badu-Apraku et al. (2012); Prado et al. (2013); Rafiq et al. (2013) menyatakan heritabilitas tinggi terdapat pada karakter bobot 100 biji, hasil, tinggi tanaman, jumlah baris biji, dan bobot biji. Menurut Herawati et al. (2009) menyatakan bahwa seleksi akan lebih efektif jika karakter yang menjadi target seleksi memiliki nilai heritabilitas yang tinggi. Karakter yang memiliki nilai heritabilitas arti luas kategori sedang pada generasi S2 adalah umur bunga betina, umur bunga jantan, jumlah daun, panjang daun,
jarak cabang tassel, panjang tongkol, panjang biji, dan lebar biji. Karakter yang memiliki nilai heritabilitas kategori rendah generasi S2 yaitu karakter persentase
tidak rebah batang, persentase tidak rebah akar, rendemen biji, dan kadar air. Karakter yang memiliki nilai heritabilitas minus (dianggap nol) dikategorikan kedalam heritabilitas rendah seperti pada karakter persentase tidak rebah batang.
Seleksi Berbagai Karakter Terhadap Genotipe Uji Jagung S1 dan S2
generasi S1 berdasarkan karakter hasil. Wardyn et al. (2009) menyatakan bahwa
genotipe terseleksi terbaik pada tanaman jagung diurutkan berdasarkan hasil dan tinggi tanaman. Sepuluh genotipe terseleksi pada generasi S1 adalah
P31xNK6326-93B, P27xNK6326-39A, PERTIWI3xNK33-84B, P31xNK33-44A, NK6326xP31-43D, NK6326xP31-71A, P27xPERTIWI3-35B, P27xNK6326-23A, NK6326xNK6326-102A dan PERTIWI3xP31-13A. Terdapat genotipe uji pada beberapa karakter yang menunjukkan nyata lebih besar dibandingkan dengan rataan genotipe uji. Genotipe P27xNK6326-39A, PERTIWI3xNK33-84B, P31xNK33-44A dan P27xPERTIWI3-35B memiliki nilai diameter tongkol yang lebih besar dibandingkan rataan genotipe (Tabel 6).
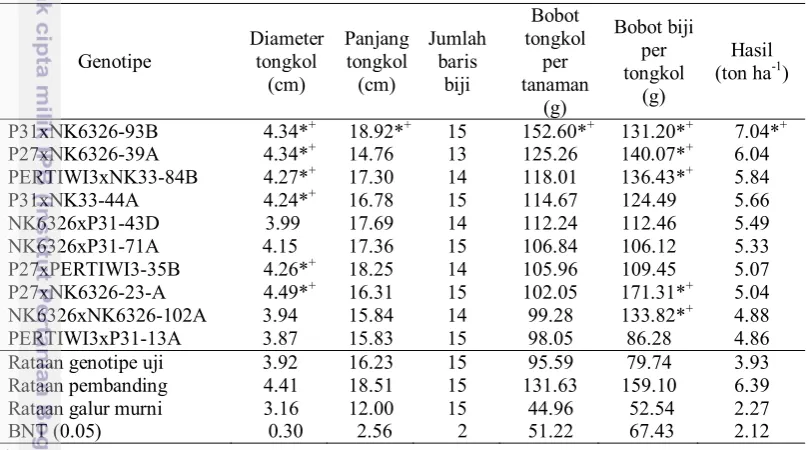
Tabel 6 Nilai tengah hasil dan komponen hasil sepuluh genotipe jagung terbaik generasi S1
PERTIWI3xNK33-84B 4.27*+ 17.30 14 118.01 136.43*+ 5.84
P31xNK33-44A 4.24*+ 16.78 15 114.67 124.49 5.66
NK6326xP31-43D 3.99 17.69 14 112.24 112.46 5.49
NK6326xP31-71A 4.15 17.36 15 106.84 106.12 5.33
P27xPERTIWI3-35B 4.26*+ 18.25 14 105.96 109.45 5.07
P27xNK6326-23-A 4.49*+ 16.31 15 102.05 171.31*+ 5.04
NK6326xNK6326-102A 3.94 15.84 14 99.28 133.82*+ 4.88
PERTIWI3xP31-13A 3.87 15.83 15 98.05 86.28 4.86
Rataan genotipe uji 3.92 16.23 15 95.59 79.74 3.93
Rataan pembanding 4.41 18.51 15 131.63 159.10 6.39
Rataan galur murni 3.16 12.00 15 44.96 52.54 2.27
BNT (0.05) 0.30 2.56 2 51.22 67.43 2.12
*+
= Lebih besar dari rataan genotipe uji + BNT.
Genotipe P27xNK6326-23A menunjukkan nilai karakter diameter tongkol dan bobot biji per tongkol nyata lebih besar dibandingkan dengan rataan genotipe uji. Genotipe P31xNK6326-93B menunjukkan nilai terbaik pada karakter diameter tongkol, panjang tongkol bobot tongkol per tanaman dan hasil lebih tinggi dibandingkan dengan rataan genotipe pada uji BNT 0.05. Tidak terdapat genotipe terseleksi pada karakter jumlah baris biji yang menunjukkan nyata lebih besar dibandingkan dengan rataan genotipe uji. Pengaruh genetik merupakan pengaruh keturunan yang dimiliki oleh setiap galur sedangkan pengaruh lingkungan adalah pengaruh yang ditimbulkan oleh habitat dan kondisi lingkungan (Kuruseng dan Kuruseng, 2008).
Tabel 7 Nilai tengah komponen vegetatif berdasarkan hasil sepuluh genotipe jagung terbaik tanaman jagung generasi S1
Genotipe
PERTIWI3xNK33-84B 56 55 111.70*++ 202.00 13 37.70
P31xNK33-44A 59 57 104.65*+ 211.85 14 37.33
NK6326xP31-43D 61 58 98.33 191.00 14 36.60
NK6326xP31-71A 59 58 105.75*++ 210.40 15 35.48
P27xPERTIWI3-35B 55- 56 101.65*+ 201.05 14 34.85
P27xNK6326-23-A 57 56 103.10*+ 210.10 15 36.13
NK6326xNK6326-102A 59 59 94.05 197.60 15 38.40
PERTIWI3xP31-13A 58 56 99.85 193.35 14 43.60*+
Rataan genotipe uji 60 58 97.92 192.80 14 36.95
Rataan pembanding 57 56 104.05 211.66 14 41.07
Rataan galur murni 62 60 76.07 147.62 13 30.81
BNT (0.05) 4 3 3.44 34.73 2 4.87
*+
= Lebih besar dari rataan genotipe uji + BNT; *++ = Lebih besar dari rataan genotipe uji + 2BNT.
Tabel 8 menunjukkan bahwa terdapat karakter vegetatif yang mendukung besarnya nilai karakter hasil. Genotipe P31xNK6326-93B nyata lebih besar dibandingkan dengan rataan genotipe uji pada karakter tinggi letak tongkol dan panjang tassel. Genotipe PERTIWI3xNK33-84B dan genotipe NK6326xP31-71A menunjukkan nilai lebih tinggi dibandingkan dengan rataan genotipe uji + 2 kali nilai BNT serta genotipe P31xNK33-44A, dan P27xNK6326-23-A menunjukkan nilai lebih tinggi dibandingkan dengan rataan genotipe uji pada karakter tinggi letak tongkol. Genotipe P27xPERTIWI3-35B menunjukkan nilai lebih tinggi dibandingkan dengan rataan genotipe uji pada karakter umur silking dan tinggi letak tongkol. Genotipe PERTIWI3xP31-13A menunjukkan nilai lebih tinggi dibandingkan dengan rataan genotipe uji pada karakter panjang tassel.
Tabel 8 menunjukkan bahwa sepuluh genotipe terbaik generasi S2 berbeda
dan panjang tongkol. Kedua karakter ini tidak terdapat genotipe uji yang memiliki nyata lebih besar dibandingkan dengan rataan genotipe uji.
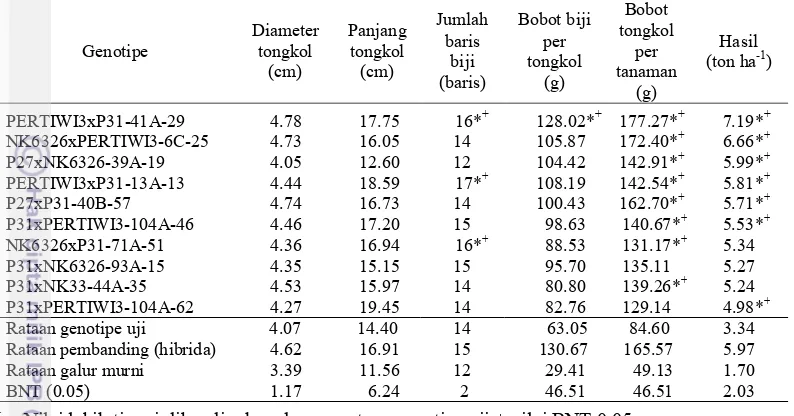
Tabel 8 Nilai tengah hasil dan komponen hasil sepuluh genotipe jagung terbaik generasi S2
P31xPERTIWI3-104A-62 4.27 19.45 14 82.76 129.14 4.98*+ Rataan genotipe uji 4.07 14.40 14 63.05 84.60 3.34 Rataan pembanding (hibrida) 4.62 16.91 15 130.67 165.57 5.97 Rataan galur murni 3.39 11.56 12 29.41 49.13 1.70 BNT (0.05) 1.17 6.24 2 46.51 46.51 2.03
+
= Nilai lebih tinggi dibandingkan dengan rataan genotipe uji + nilai BNT 0.05.
Tabel 9 Nilai tengah komponen vegetatif berdasarkan hasil sepuluh genotipe jagung terbaik generasi S2
+
= Nilai lebih tinggi dibandingkan dengan rataan genotipe uji + nilai BNT.
Rataan 72 genotipe uji menunjukkan hasil sebesar 3.34 ton ha-1, rataan hasil hibrida sebesar 5.97 ton ha-1 dan rataan galur murni pada hasil 1.70 ton ha-1. Rataan genotipe uji pada karakter hasil lebih tinggi dibandingkan dengan rataan hasil galur murni (Tabel 8). Penelitian tanaman pangan lain yaitu hasil gabah pada padi rata berkisar dari 0.53 ton ha-1–11.00 ton ha-1 (Lestari et al (2010).
Genotipe NK6326xP31-71A-51 menunjukkan nyata lebih besar dibandingkan dengan rataan genotipe uji pada karakter tinggi letak tongkol, tinggi tanaman, dan panjang tassel. Genotipe P31xPERTIWI3-104A-62 menunjukkan
Genotipe
nilai lebih besar dibandingkan dengan rataan genotipe uji + nilai BNT pada karakter tinggi letak tongkol dan panjang tassel. Genotipe P27xP31-40B-57 menunjukkan nilai lebih tinggi dibandingkan dengan rataan genotipe uji + nilai BNT pada karakter tinggi letak tongkol. Genotipe P31xPERTIWI3-104A-62 57 menunjukkan nyata lebih besar dibandingkan dengan rataan genotipe uji pada karakter pada panjang tassel.
Terdapat beberapa populasi dasar yang galur-galurnya termasuk 10 terbaik pada generasi S1 dan S2 yaitu P27xNK6326, PERTIWI3xP31, NK6326xP31,
P31xNK6326 dan P31xNK33. Hal tersebut mennjukkan bahwa genotipe-genotipe tersebut memiliki faktor genetik yang baik untuk diwariskan ke generasi selanjutnya.
Korelasi Antar Karakter Agronomi dan Hasil
Analisis korelasi digunakan untuk melihat kekuatan hubungan antara dua peubah melalui koefisien korelasi. Dalam seleksi, koefisien korelasi memiliki arti penting antarkarater terhadap daya hasil jagung. Semakin besar koefisien nilai korelasi semakin erat hubungan kekerabatan antar karakter. Nilai korelasi berkisar antara 1 dan -1 (Walpole 1982). Seleksi jagung generasi S1 menunjukkan bahwa
secara umum semua karakter berkorelasi antar karakter kecuali jumlah baris biji tidak berkorelasi pada semua karakter yang diamati. Hasil berkorelasi positif sangat nyata terhadap karakter tinggi tanaman, tinggi letak tongkol, diameter tongkol, panjang tongkol dan berkorelasi negatif sangat nyata terhadap umur anthesis dan umur silking. Ogunniyan et al. 2014 melaporkan bahwa hasil berkorelasi positif dengan tinggi tanaman, tinggi letak tongkol, jumlah daun per tanaman, dan luas daun. Pengetahuan tentang keterkaitan antara hasil dan komponen-komponennya berkontribusi akan meningkatkan efisiensi program pemuliaan (Zarei et al. 2012). Inamullah et al. (2011) melaporkan bahwa panjang tongkol digunakan sebagai penanda seleksi untuk melihat karakter hasil yang terbaik, sehingga panjang tongkol berkorelasi terhadap hasil didukung karakter bobot tongkol pertanaman, bobot biji per tongkol dan bobot 1000 biji.
Tabel 10 Koefisien korelasi antar karakter genotipe jagung generasi S1
UA US TT TLT DT PT JBB HSL diameter tongkol; PT = panjang tongkol; JBB = jumlah baris biji; HSL = hasil; * = berkorelasi nyata pada taraf 5%; ** = berkorelasi nyata pada taraf 1%; tn = tidak berkorelasi nyata.
Generasi S2 genotipe jagung menunjukkan bahwa hasil berkorelasi positif
Pada generasi S2, dari semua karakter yang berkorelasi karakter tinggi letak
tongkol dan tinggi tanaman berkorelasi positif dan sangat nyata antar semua karakter. Hal ini didukung oleh penelitian Ziyomo dan Bernado (2013) menyatakan bahwa tinggi tanaman jagung berkorelasi nyata dan positif terhadap hasil. Karakter hasil generasi S1 dan S2 berkorelasi nyata terhadap karakter tinggi
letak tongkol, tinggi tanaman, panjang tongkol dan diameter tongkol.
Tabel 11 Koefisien korelasi antar karakter genotipe jagung generasi S2
UA US TT TLT DT PT JBB HSL
UA = umurberbunga jantan; US = umur berbunga betina; TT = tinggi tanaman; TLT = tinggi letak tongkol; DT = diameter tongkol; PT = panjang tongkol; JBB = jumlah baris biji; HSL = hasil; * = berkorelasi nyata pada taraf 5%; ** = berkorelasi nyata pada taraf 1%; tn = tidak berkorelasi nyata.
Genotipe yang memiliki diameter tongkol yang besar cenderung memiliki tongkol yang panjang dan hasil yang tinggi. Sejalan dengan penelitian (Zarei et al. 2012; Mohammadi et al. 2003) mengatakan bahwa panjang tongkol, panjang biji, bobot 100 biji, jumlah biji per tongkol, dan persentase kelobot tongkol berkorelasi positif dan nyata terhadap hasil. Korelasi antara hasil dan komponen hasil dijumpai juga pada komoditas tanaman pangan lainnya seperti padi di mana hasil berkorelasi positif dan nyata terhadap jumlah gabah isi, hasil gabah dengan jumlah gabah total per malai, dan hasil gabah dengan jumlah malai per rumpun pada 12 genotipe padi sawah (Sutaryo2014).
Peubah Kualitatif Jagung S1 dan S2
Karakter kualitatif dikendalikan oleh satu atau dua gen saja dan sedikit sekali dikendalikan oleh lingkungan. Karakter kualitatif merupakan wujud fenotipik yang saling berbeda antara satu dengan yang lainnya dan masing-masing dapat dikelompokkan dalam bentuk kategori, misalnya warna hijau, putih, dan merah (Suryo 2004). Karakter fase vegetatif generasi S1 adalah karakter bentuk
Peubah kualitatif pada generasi jagung S1 diamati pada fase vegetatif dan
generatif. Karakter fase generatif genotipe jagung S1 adalah karakter kerusakan
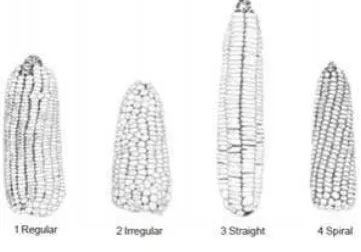
tongkol menunjukkan 1 genotipe dengan kerusakan banyak, 54 genotipe sangat sedikit dan 25 genotipe tidak ada kerusakan. Karakter warna batang menunjukkan warna hijau pada semua genotipe. Bentuk tongkol menunjukkan bahwa terdapat 14 genotipe bentuk kerucut, 8 silindris, dan 58 silindris mengerucut. Karakter warna permukaan biji menujukkan bahwa terdapat 33 genotipe kuning, 12 genotipe orange, 29 genotipe orange kuning, dan 6 genotipe putih kekuningan. Terdapat 9 genotipe lurus, 39 genotipe beraturan, 2 spiral, dan 30 tidak beraturan pada karakter posisi biji dalam baris. Warna endosperma putih pada semua genotipe. Terdapat 66 genotipe berwarna kuning dan 14 genotipe berwarna kuning tua pada warna pericarp. Terdapat 8 genotipe gigi kuda, 12 genotipe mutiara, dan 60 genotipe intermediet pada tipe biji (Lampiran 7).
Karakter fase vegetatif generasi S2 adalah karakter bentuk ujung daun,
warna batang, warna pelepah daun, orientasi daun, lidah daun, urat daun, tipe tassel dan ukuran tassel. Karakter bentuk ujung daun menunjukkan bahwa semua genotipe memiliki bentuk runcing. Begitu juga warna batang menunjukkan bahwa semua genotipe berwarna hijau. Karakter warna antosianin pelepah daun menunjukkan bahwa terdapat 13 genotipe kuat, 16 genotipe lemah, dan 51 genotipe sangat lemah. Karakter orientasi daun menunjukkan bahwa terdapat 59 genotipe tipe terkulai dan 21 genotipe tipe tegak. Terdapa karakter lidah daun pada semua genotipe. Tidak terdapat urat daun pada semua genotipe. Terdapat 7 genotipe primer, 41 genotipe tipe tassel primer-sekunder, dan 32 genotipe tipe primer sekunder tersier. Karakter ukuran tassel menunjukkan bahwa 24 genotipe ukuran besar, 18 genotipe ukuran sedang, dan 38 genotipe ukuran kecil (Lampiran 8).
Fase generatif genotipe jagung S2 adalah karakter kerusakan tongkol
menunjukkan terdapat 3 genotipe dengan kerusakan banyak, 26 genotipe sangat sedikit dan 51 genotipe tidak ada kerusakan. Bentuk tongkol menunjukkan bahwa terdapat 33 silindris dan 47 silindris mengerucut. Karakter warna tongkol menujukkan bahwa terdapat 34 genotipe kuning, 34 genotipe orange, dan 12 genotipe orange kuning. Karakter posisi biji dalam baris menunjukkan bahwa terdapat 11 genotipe lurus, 43 genotipe beraturan, dan 26 tidak beraturan. Warna endosperma putih pada semua genotipe. Terdapat 42 genotipe berwarna kuning, 3 kuning muda dan 35 genotipe berwarna kuning tua pada warna pericarp. Terdapat 6 genotipe gigi kuda, 4 genotipe mutiara, dan 70 genotipe intermediet pada tipe biji (Lampiran 9).
Pengelompokan Genotipe Generasi S1 dan S2
Gambar 5 Dendogram genotipe jagung generasi S1
Genoti
p
e
A
gg
lome
rativ
e C
oe
ff
icient =
0
Gambar 6 Dendogram genotipe jagung generasi S2
Genoti
p
e
A
gg
lome
rativ
e C
oe
ff
icient =
0
Dari hasil analisis gerombol pada metode Gower menunjukkan bahwa genotipe-genotipe generasi S1 secara keseluruhan terpisah menjadi dua bagian yaitu
mengelompok dan terpisah sendiri dengan koefisien ketidakmiripan sekitar 0.28. Genotipe yang terpisah sendiri adalah PERTIWI3xNK33-84A. Genotipe-genotipe yang mengelompok akan dilihat kedekatannya dengan ketiga galur murni (Mr4, Mr14, dan Nei9008).
Genotipe Mr4 dan Mr14 memiliki jarak genetik yang lebih dekat dibandingkan dengan Nei9008. Ketidakmiripan Mr4 dan Mr14 adalah sebesar ±0.16 sedangkan ketidakmiripan dengan Nei9008 adalah sebesar ±0.18. Genotipe pembanding yang cukup dekat dengan Mr4 adalah NK6323. Terdapat beberapa genotipe yang memiliki
kemiripan terhadap Mr4 diantaranya adalah P27xNK33-79A,
NK6326xPERTIWI3-76B, PERTIWI3xNK6326-48B, PERTIWI3xP31-41A, NK6326xP27-28B, P31xNK6326-26A, dan NK33xNK33-12A). Genotipe pembanding yang cukup dekat dengan Mr4 adalah P27. Beberapa contoh genotipe yang memiliki kemiripan terhadap Mr14 adalah genotipe NK6326xP31-112A, P31xNK33-108A, PERTIWI3xP27-90A,
NK6326xPERTIWI3-76A, NK33xPERTIWI3-68A, P27xNK6326-67A, PERTIWI3xP27-58B, NK6326xP27-53B, P31xPERTIWI3-52A,
NK6326xPERTIWI3-49B, P31xNK33-44A, NK6326xP31-43D, NK6326xP31-43B, 40A, P27xP31-40B, P27xNK6326-39A, P27xP27-37A, P27xPERTIWI3-35C, P27xPERTIWI3-35B, NK6326xP27-28A, NK6326xP31-24A, dan PERTIWI3xP31-13B. Hibrida sebagai genotipe pembanding yang memiliki kedekatan terhadap Nei9008 adalah PERTIWI3 dan untuk genotipe uji yang dekat dengan Nei9008 diantaranya genotipe P31xNK6326-93B, P27xP31-77A, 66B, NK33xP31-55A, PERTIWI3xNK6326-48A, NK6326xNK33-38A, NK33xPERTIWI3-36A, P31xNK6326-26B, dan P27xNK632623-A (Gamba 5).
Gambar 6 menunujukkan bahwa genotipe-genotipe generasi S2 secara
keseluruhan terbagi menjadi dua bagianyaitu mengelompok dan terpisah sendiri dengan koefisien ketidakmiripan sekitar 0.34. Genotipe yang terpisah sendiri adalah PERTIWI3xNK6326-10B-59. Genotipe-genotipe yang mengelompok akan dilihat kedekatannya dengan ketiga galur murni yaitu Mr4, Mr14, dan Nei9008. Seperti halnya generasi S1, genotipe Mr4 dan Mr14 memiliki jarak genetik yang lebih dekat
dibandingkan dengan Nei9008. Ketidakmiripan Mr4 dan Mr14 adalah sebesar ±0.16 sedangkan ketidakmiripan dengan Nei9008 adalah sebesar ±0.18. Terdapat beberapa genotipe generasi S2 yang memiliki kedekatan terhadap Mr4 diantaranya yaitu genotipe
NK6326xP31-43D-70, P31xPERTIWI3-19A-64, P27xP31-40A-54, NK6326xPERTIWI3-76B-49, PERTIWI3xNK6326-10B-42, P27xPERTIWI3-35A-40,
PERTIWI3xNK6326-48A-39, P27xP31-4B-38, NK6326xP31-112A-34, NK6326xP27-28A-23, P27xP31-40A-22, NK6326xP31-43D-18, PERTIWI3xP31-13A-13, dan P31xPERTIWI3-19A-11.
P31xNK33-108A-2, dan P31xNK6326-93B-1. Genotipe uji yang memiliki kedekatan terhadap Mr14 adalah NK6326. Genotipe uji yang memiliki kedekatan terhadap Nei9008 diantaranya adalah genotipe P31xNK6326-93B-63, NK6326xP31-71A-51, PERTIWI3xNK6326-10C-50, P31xNK33-44A-47, P31xNK6326-93B-43, PERTIWI3xNK33-84A-33, P27xPERTIWI3-35A-27, NK6326xP31-112A-12, dan NK6326xP31-24A-10.
Kedudukan hibrida sebagai pembanding dengan genotipe uji yang berasal dari hibrida tersebut memiliki hubungan yang dekat. Ketidakmiripan genetik antar galur murni pada generasi S1 dan generasi S2 menunjukkan nilai yang sama. Hal ini dapat
dilihat dari koefisien ketidakmiripan antara Mr4 dan Mr14 sebesar 0.16 dan ketidakmripan keduanya dengan Nei9008 sebesar 0.18. Kelompok genotipe yang terbentuk berpotensi untuk dijadikan grup heterotik, namun memerlukan penelitian lebih lanjut.
Pendugaan Tingkat Inbreeding Depression yang Diimbangi dengan Seleksi dari
Generasi S1 ke S2
Dalam tahapan pemuliaan, seleksi merupakan tahapan untuk memilih genotipe-genotipe terbaik dalam beberapa siklus. Seleksi yang diikuti selfing pada tanaman menyerbuk silang dalam beberapa generasi akan menyebabkan kondisi inbreeding depression. Kondisi inbreeding depression pada dua generasi genotipe uji jagung dapat menurunkan nilai karakter-karakternya.
Tabel 12 Pengaruh musim lima genotipe jagung pembanding (hibrida) generasi S1 ke
S2menggunakan uji t
Tinggi tanaman (cm) Hasil (ton ha-1)
Rata - rata S1 211.66 6.39
Rata - rata S2 175.69 5.97
Rata-rata S1 - S2 8.98** 1.19 tn
Nilai P (P-value) < 0.001 0.299
** = beda nyata pada taraf 1% pada uji t; tn = tidak nyata pada uji t.
Pengaruh musim pada percobaan ini diduga dari genotipe pembanding (hibrida) generasi S1 dan S2. Terdapat pengaruh musim yang tidak nyata pada karakter hasil
sedangkan pada karakter tinggi tanaman terdapat pengaruh musim yang nyata (Tabel 12). Dengan demikian karakter tinggi tanaman dilakukan adjusted mean untuk melihat kondisi inbreeding depression pada 72 genotipe uji jagung sedangkan tidak untuk karakter hasil.
Tabel 13 menunjukkan bahwa untuk karakter hasil terdapat perbedaan nyata antar generasi S1 dan S2. Hal ini menunjukkan bahwa galur-galur S2 terlihat mengalami
kondisi inbreeding depression dimana generasi S2 mengalami penurunan daya hasil dari
generasi S1. Penurunan rata-rata hasil generasi S1 ke S2 sebesar 0.73 ton ha-1. Ciri utama
Tabel 13 Pendugaan inbreeding depression 72 genotipe uji jagung generasi S1 ke S2
menggunakan uji t
Tinggi tanaman (cm) Hasil (ton ha-1)
Rata - rata S1 179.96 4.48
Rata - rata S2 170.58 3.34
Rata-rata S1 - S2 5.99 tn 0.73 **
Nilai P (P-value) 0.075 0.005
** = beda nyata pada taraf 1% pada uji t; tn = tidak nyata pada uji t.
Penurunan rata-rata tinggi tanaman generasi S1 ke S2 sebesar 5.99 cm. Tinggi
tanaman menunjukkan tidak berbeda nyata dari generasi S1 ke S2.. Hal ini menandakan