HUBUNGAN KERAGAMAN GEN DGAT1 (
diacylglycerol
acyltransferase1
) TERHADAP PRODUKSI DAN PROFIL ASAM
LEMAK SUSU SAPI PERAH FRIESIAN HOLSTEIN
SANTIANANDA ARTA ASMARASARI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Hubungan Keragaman Gen DGAT1 (diacylglycerol acyltransferase1) terhadap Produksi dan Profil Asam Lemak Susu Sapi Perah Friesian Holstein adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2013
Santiananda Arta Asmarasari
RINGKASAN
SANTIANANDA ARTA ASMARASARI. Hubungan Keragaman Gen DGAT1 (diacylglycerol acyltransferase1) terhadap Produksi dan Profil Asam Lemak Susu Sapi Perah Friesian Holstein. Dibimbing oleh CECE SUMANTRI dan I WAYAN MATHIUS.
Komposisi asam lemak pada susu sangat dipengaruhi oleh faktor genetik dan lingkungan. Komposisi lemak susu dapat diubah melalui perbaikan nutrisi maupun perbaikan genetik yang dilakukan melalui seleksi keunggulan pada gen yang berasosiasi kuat terhadap sifat produksi susu dan persentase kadar lemak. Suatu studi pemetaan lokus sifat kuantitatif (QTL) pada sapi menghasilkan identifikasi polimorfisme (K232A) dalam pengkodean gen untuk acyl CoA:diacylgliserol acyltransferase1 (DGAT1), yang merupakan kunci enzim dalam sintesis trigliserida dan memiliki efek kuat pada persentase lemak susu dan karakteristik produksi susu lainnya. Pengaruh dari mutasi DGAT1 pada komposisi lemak susu sapi perah di Indonesia belum banyak dilakukan. Penelitian ini bertujuan untuk mengidentifikasi keragaman genetik gen DGAT1 pada sapi Friesian Holstein (FH) serta pengujian hubungan antara keragaman genotipe gen DGAT1 terhadap produksi dan profil asam lemak susu.
Penelitian terdiri atas dua tahap. Tahap pertama adalah identifikasi keragaman gen DGAT1 pada sapi perah FH dan tahap kedua adalah pengujian hubungan keragaman gen DGAT1 terhadap produksi susu dan profil asam lemak susu. Sampel darah yang digunakan untuk melihat keragaman genetik gen DGAT1 diambil dari 7 populasi di Jawa Tengah dan Jawa Barat dengan total 300 ekor, yaitu BBPTU Sapi Perah (SP) Baturraden (123), BET Cipelang (32), BPPT Cikole (36), peternakan rakyat Cilumber (34) dan peternakan rakyat Pasir Kemis (34). Aliran genetik dari pejantan terhadap betina laktasi dilihat dengan menggunakan sampel sapi pejantan FH yang berasal dari lokasi Balai Inseminasi Buatan (BIB) Lembang (16) dan Balai Besar Inseminasi Buatan (BBIB) Singosari (28). Sampel susu untuk melihat hubungan keragaman gen DGAT1 terhadap produksi dan profil asam lemak susu hanya digunakan sampel dari BBPTU SP Baturraden sebanyak 40 sampel. Sampel susu dikoleksi berdasarkan uji satu hari dengan menjumlahkan produksi pagi dan sore hari dari sapi laktasi dalam kisaran periode laktasi 1-6 dan bulan laktasi 1-12. Data asam lemak susu diperoleh dari hasil pengujian di labooratorium menggunakan metode Gas Chromatography.
Ektraksi DNA dilakukan dari sampel darah sapi FH mengikuti metode (Sambrook
et al. 1989) yang telah dimodifikasi.
Penentuan genotipe masing-masing individu dilakukan dengan pendekatan
Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (RFLP). Keragaman gen dilihat dengan menghitung frekuensi alel, frekuensi genotipe dan nilai heterozigositas. Hubungan antara varian genotipe gen DGAT1 dengan produksi susu dan profil asam lemak susu dihitung menggunakan General Linear Model (GLM) dengan bantuan software SAS 9.1.
dengan panjang fragmen 411 bp dan genotipe AK ditunjukkan dengan panjang fragmen 203, 208 dan 411 bp.
Frekuensi genotipe KA (73%) pada tujuh populasi sapi perah FH lebih tinggi dibanding dengan frekuensi genotip KK (27%) dan AA (0%). Fenomena tidak adanya genotipe AA dari sapi betina FH, baik di peternakan rakyat maupun di stasiun bibit salah satunya adalah dipengaruhi oleh sumber pejantan inseminasi buatan (IB) yang digunakan. Pejantan yang digunakan untuk IB pada betina laktasi umumnya berasal dari BIB Lembang dan BBIB Singosari. Berdasarkan hasil analisis pada sapi pejantan IB yang berasal dari BIB Lembang dan BBIB Singosari tidak ditemukan genotipe AA dan frekuensi alel A rendah pada pejantan yang diamati. Frekuensi alel sapi FH dari tujuh populasi diperoleh alel K (64%) lebih tinggi dibanding alel A (36%). Dari hasil penelitian ini gen DGAT1 pada sapi FH dari tujuh populasi yang diamati bersifat polimorfik karena ditemukan dua tipe alel, yaitu alel K dan alel A.
Nilai heterozigositas pengamatan (Ho) populasi sapi FH dari tujuh lokasi
berbeda berkisar antara 0.313-0.938 dan nilai heterozigositas harapan (He)
berkisar antara 0.264-0.498. Nilai heterozigositas pengamatan (Ho) yang tertinggi
adalah sapi FH dari BET Cipelang yaitu sebesar 0.938 dan nilai heterozigositas pengamatan (Ho) yang terendah adalah sapi FH di lokasi BIB Lembang (0.313).
Begitu pula nilai heterozigositas harapan (He) yang tertinggi adalah populasi sapi
FH dari Cipelang (0.498) dan yang terendah adalah dari BIB Lembang (0.264). Tingginya nilai heterozigositas pengamatan (Ho) menunjukkan adanya keragaman
alel dalam populasi tersebut.
Pengujian pengaruh varian genotipe gen DGAT1 terhadap rataan kadar protein susu menunjukkan bahwa sapi dengan genotipe KK menghasilkan kadar protein susu lebih tinggi (3.12%) dibandingkan sapi dengan genotipe AK (3.06%), meskipun pengaruh tersebut secara statistik tidak nyata (P>0.05). Sebaliknya sapi dengan genotipe AK cenderung menghasilkan produksi susu lebih tinggi dibandingkan KK (P>0.05). Sapi dengan genotipe KK menghasilkan kadar lemak susu lebih banyak (3.32%) dibanding sapi AK (3.20%). Sapi dengan genotipe KK juga bertendensi menghasilkan Bahan Kering Tanpa Lemak (BKTL) dan Berat Jenis (BJ) lebih tinggi dibandingkan AK (P<0.05). Nilai kadar protein, kadar lemak, Berat Jenis (BJ) dan Bahan Kering Tanpa Lemak (BKTL) pada sapi FH bergenotipe KK dan AK telah memenuhi standar SNI 01-3141-1998 mengenai syarat mutu susu segar (SNI, 1998).
Pengujian pengaruh varian genotipe gen DGAT1 terhadap profil asam lemak susu menunjukkan sapi dengan genotip AK menghasilkan kadar asam lemak nervonat (C24:1) lebih tinggi daripada sapi dengan genotip KK (P<0.05). Kata kunci : Diacylglycerol acyl transferase1, gen, Friesian Holstein, produksi
SUMMARY
SANTIANANDA ARTA ASMARASARI. The relationship of Diacylglicerol acyltransferas (DGAT1) Gene Diversity to Friesian Holstein Dairy Cattle‘s Milk Production and Fatty Acid Profile. Supervised by CECE SUMANTRI and I WAYAN MATHIUS.
Milk fatty acid composition is influenced by genetic and environmental factors. Milk fat composition can be changed by nutrition and genetic improvement. Genetic improvement can be done through selection to genes which have associated strongly with milk production and milk fat percentage. A study of quantitative trait locus mapping (QTL) in cattle has been resulted that from the identification of polymorphism (K232A) in the gene coding for acyl CoA is diacylgliserol acyltransferase1 (DGAT1). DGAT1 is a key enzyme in the synthesis of triglycerides and has a strong effect on milk fat percentage and milk production. The effect of DGAT1 mutations in milk fat composition of dairy cattle in Indonesia has not been done. This study aims to, firstly, to identify the genetic diversity of DGAT1 gene in cattle Friesian Holstein (FH), secondly, to test the relationship between genotype diversity DGAT1 gene with milk production and milk fatty acid profile.
The study consisted of two steps. The first was identification of DGAT1 gene diversity in FH dairy cows. The second was testing relationship of DGAT1 gene diversity to milk production and milk fatty acid profile. The identification of DGAT1 gene diversity used blood samples which collected from 7 populations in Central Java and West Java. There were 300 heads dairy cattles used in this research which taken from BBPTU Baturraden SP (123), BET Cipelang (32), BPPT Cikole (36), Cilumber farm (34) Pasir Kemis Farm (34). Genetic flow from the male to the lactation female viewed by using samples which derived from BIB Lembang (16) and BBIB Singosari (28). The identification of the relationship of DGAT1 gene diversity to milk production and milk fatty acid profile used milk samples which collected from BBPTU Baturaden’s Dairy Cattle (40). Milk samples were collected by a single test day in which milk production in the morning and in the afternoon is calculated. Such milk samples were collected from dairy cattle which has 1-6 lactation periods and 1-12 month lactation. The data of milk fatty acid obtained through a laboratorium test results which using Gas Chromatography method. DNA extraction has been done by using modified (Sambrook et al. 1989).
Determination of the genotype of each individual was conducted with Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP) approach. Gene diversity has been seen by counting the frequency of allele, genotype frequencies and heterozygosity values. The relationship between DGAT1 gene genotype variants with milk production and milk fatty acid profile was calculated based on the General Linear Model (GLM) method using SAS 9.1 software.
KA. KK genotype is shown with 411 bp fragment length and AK genotype is shown with 203, 208 and 411 bp fragment length.
Frequiency of AK genotype (73%) at seven FH dairy cows population was higher than the KK genotype (27%) and AA genotype (0%). The phenomenon of the lack of AA genotype of FH cows, either at the farm or at the seeds center institution, was influenced by the source of male cattle which used in artificial insemination (AI) activity. Males cattle were used for AI in lactating females were generally derived from BIB Lembang and BBIB Singosari. Based on the analysis result, AA genotypes were not found in bulls from BIB Lembang and BBIB Singosari and A allele frequencies in bulls was low. Allele frequencies of seven cattle populations of FH resulted allele K (64%). That was higher than A allele (36%). This research resulted that gene DGAT1 on observed FH dairy cattle from seven populations had polymorphic characteristic due to there were found two types of alleles ( K allele and A allele).
Observation heterozygosity values (Ho) of FH cattle population from
seven different locations ranged between 0313-0938. Expected heterozygosity values (He) of FH cattle population from seven different locations ranged between
0264-0498. The highest of observation heterozygosity values (Ho) was FH dairy
cattle from BET Cipelang (0.938) and the lowest was FH dairy cattle from BIB lembang (0.313). Whereas, the highest of heterozygosity values (He) was FH
dairy cattle population from Cipelang and the lowest one was came from BIB Lembang (0.264). A high value of the observation heterozygosity (Ho) shows the
diversity of alleles in the population.
Assessment of the effect of DGAT1 gene variant genotype to average protein content of milk showed that cows with KK genotype tended to produce higher milk protein content, namely 3.12%. Such milk protein content higher than if we compared with AK genotype (3.06%), although based on statistically analysis the effect was not significsntly different (P<0.05). FH dairy cattle with KK genotype also tended to produce more milk fat (3.32%) than AA cows (3.20%). Value of protein content, fat content, specific gravity and solid non fat (SNF) in FH dairy cattle with genotype KK and AK have apropriated with SNI 01-3141-1998 standard of quality requirements of fresh milk (SNI, 1998). DGAT1 gene does not significantly affected the unsaturated fatty acids (myristoleic, palmitoleic, oleic, eurat, linolenic, and arachidonat eicosatrinoat). Nevertheles, DGAT1 gene significantly affected (P<0.05)nervonat acid (C24: 1). Keywords: Diacylglycerol acyl Transferase,1Genes, Friesian Holstein, milk
© Hak Cipta Milik IPB, Tahun 2013
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada
Program Studi Ilmu Produksi dan Teknologi Peternakan
HUBUNGAN KERAGAMAN GEN DGAT1 (
diacylglycerol
acyltransferase1
) TERHADAP PRODUKSI DAN PROFIL ASAM
LEMAK SUSU SAPI PERAH FRIESIAN HOLSTEIN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2013
Judul Tesis : Hubungan Keragaman Gen DGAT1 (diacylglicerol
acyltransferase1) terhadap Produksi dan Profil Asam Lemak Susu Sapi Perah Friesian Holstein
Nama : Santiananda Arta Asmarasari
NIM : D151100111
Disetujui oleh Komisi Pembimbing
Prof Dr Ir Cece Sumantri, MAgrSc. Ketua
Prof Dr Ir I Wayan Mathius, MSc. Anggota
Diketahui oleh
Ketua Program Studi Mayor Ilmu Produksi dan Teknologi Peternakan
Prof Dr Ir Muladno, MSA
Dekan Sekolah Pascasarjana
Dr Ir Dahrul Syah, MScAgr
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga penulis mampu menyelesaikan tesis ini dengan judul “Hubungan Keragaman Gen DGAT1 (diacylglcerol acyltransferase1) terhadap Produksi dan Profil Asam Lemak Susu Sapi Friesian Holstein”. Tesis ini dibuat sebagai salah satu syarat untuk memperoleh gelar Magister Sains pada Mayor Ilmu Produksi dan Teknologi Peternakan.
Penulis menyampaikan penghargaan dan ucapan terima kasih setinggi-tingginya kepada Bapak Prof Dr Ir Cece Sumantri, MAgrSc selaku ketua komisi pembimbing dan Bapak Prof Dr Ir I Wayan Mathius, MSc selaku anggota komisi pembimbing atas segala curahan waktu, bimbingan serta ketulusan dan kesabarannya sehingga penulis bisa menyelesaikan tugas akhir ini. Kepada Bapak Dr. Jakaria, SPt, MSi. penulis menyampaikan terima kasih atas kesediaan dan masukannya sebagai penguji luar komisi pada ujian tesis.
Ucapan terima kasih juga disampaikan kepada Bapak Kepala Balai Besar Pembibitan Ternak Unggul (BBPTU) Sapi Perah Baturraden Purwokerto Jawa Tengah beserta staf dan jajarannya, atas segala bantuannya selama pengambilan darah sapi perah Friesian Holstein (FH) serta data produksi dan kualitas susu.
Penulis mengucapkan terima kasih kepada Ibu Dr Bess Tiesnamurti, MSc selaku Kepala Pusat Penelitian dan Pengembangan Peternakan serta Bapak Dr Nasrullah, MSc selaku Kepala Balai Penelitian Ternak, atas kesempatan yang diberikan untuk melanjutkan studi dan dukungan maupun dorongan yang tiada henti untuk segera menyelesaikan studi.
Kepada rekan sekantor yaitu ibu Dr Anneke Anggraeni, Dr Polmer Situmorang, Dr Eko Handiwirawan, penulis mengucapkan terima kasih atas bantuan dan saran-saran yang diberikan.
Kepada teman-teman seperjuangan di Laboratorium Genetika Molekuler Ternak (Eryk Andreas, Restu, Almira, Ibu Nena, Ibu Nungky, Ibu Hidayati, Ferdi, Rey, dan Sri) penulis mengucapkan terima kasih atas bantuan dan kerjasamanya selama penelitian berlangsung. Kepada rekan-rekan mahasiswa pascasarjana program studi ITP angkatan 2010 (Hearty, Sasa, Ibu Fifi, Andi, Imam, Awlia, Alwi, David, Rupi dan Rifah) terima kasih atas kebersamaan dan supportnya yang tiada henti.
Ungkapan terima kasih juga penulis sampaikan kepada orang-orang terdekat yang sangat penulis cintai, yaitu Ayahanda Herry Santoso dan Almh. Ibunda Yeni Kristianing serta suami Cecep Hidayat, yang selalu memberikan memberikan motivasi, doa dan dukungan moril. Tiada henti mereka memberikan dorongan untuk terus berupaya keras agar penulis segera menyelesaikan studi, meringankan beban di kala menghadapi kesulitan selama menjalani proses studi. Kepada buah hatiku Maryam Al Ghazelia Arta dan Ibrahim El Rumi Danendra, terima kasih atas pengertian dan keceriaan yang diberikan, semoga ini dapat menjadi tambahan motivasi dan teladan untuk terus menuntut ilmu lebih banyak.
Semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2013
DAFTAR ISI
Hubungan varian genetik gen DGAT1 dengan sifat produksi susu dan kualitas susu
10 Memodifikasi profil asam lemak susu melalui teknologi pakan 11
3 METODE 12
Waktu dan Lokasi Penelitian 12
Bahan dan Alat 12
Sampel darah sapi perah FH 12
Sampel susu 12
Amplifikasi ruas gen DGAT1 15
Elektroforesis produk PCR 15
Penentuan genotipe dengan pendekatan RFLP 16
Uji kualitas susu 16
Analisa profil asam lemak susu 16
Analisa Data 16
Frekuensi alel dan genotipe 16
Heterozigositas 17
Analisa hubungan genotipe gen DGAT1 terhadap produksi
susu dan profil asam lemak susu 17
4 HASIL DAN PEMBAHASAN 18
Frekuensi genotipe dan alel gen DGAT1 20
Heterozigositas 22
Pengaruh varian genotipe gen DGAT1 terhadap kualitas susu
segar sapi FH 23
5 SIMPULAN DAN SARAN 28
Simpulan 28
Saran 28
DAFTAR PUSTAKA 28
LAMPIRAN 33
DAFTAR TABEL
1 Karakteristik Sampel Darah Sapi Perah FH 12
2 Runutan primer yang digunakan 13
3 Frekuensi genotipe dan alel gen DGAT1 pada sapi Friesian Holstein 21 4 Frekuensi genotipe dan frekuensi alel pada beberapa bangsa sapi perah 22
5 Heterozigositas gen DGAT1pada sapi FH 22
6 Produksi dan kualitas susu segar sapi FH berdasarkan genotipe 24 7 Polimorfisme gen DGAT1 pada beberapa bangsa sapiperah 25 8 Profil asam lemak susu sapi perah Friesian Holstein 26
DAFTAR GAMBAR
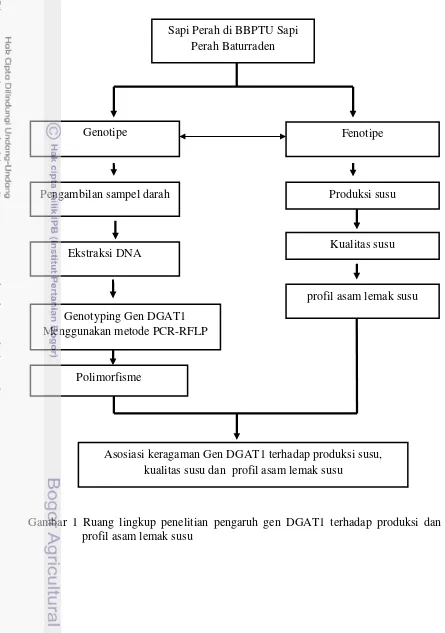
1 Ruang lingkup penelitian pengaruh gen DGAT1 terhadap produksi dan profil asam lemak susu
4 2 Kurva laktasi produksi susu dengan masa laktasi 330 hari dan
Kurva konsumsi bahan kering pakan (Dry Matter Intake=DMI) 6 3 Grafik hubungan produksi susu dan kadar lemak susu 7 4 Rekonstruksi struktur gen DGAT1 berdasarkan sekuens gen DGAT1
di GenBank (KodeAkses AY065621) 11
5 Visualisasi pita DNA hasil amplifikasi ruas Gen DGAT1 pada gel
agarose 1,5% 18
6 Visualisasi pita DNA Gen DGAT1 pada gel agarose 2% 19 7 Runutan nukleotida ruas gen DGAT1 (GenBank nomor akses No 20
AY065621)
8 Perbedaan sekuen alel K dan A gen DGAT1|EaeI 20
DAFTAR LAMPIRAN
1 Sekuen gen DGAT1 yang di akses di gene bank No AY065621 33
1
PENDAHULUAN
Latar Belakang
Susu sapi dikenal sebagai komponen penting dari diet manusia. Nilai gizi susu sapi sering dipromosikan sebagai minuman kesehatan dan terbukti memberikan pengaruh yang positif bagi kesehatan tubuh manusia. Susu sebagai makanan
pharmaceutical bermanfaat untuk mencegah penyakit terkait dengan sistem kekebalan tubuh, sistem endokrin, sistem saraf, sistem pencernaan dan sistem sirkulasi darah. Susu sapi juga berperan penting pada aktivitas anti-hipertensi, antivirus, antibakteri, antioksidatif, dan untuk kesehatan tulang (Cashman et al.
2006). Meskipun susu sapi memiliki banyak fungsi bagi kesehatan, namun komposisi asam lemak susu yang terkandung di dalamnya sering pula menjadi perhatian karena susu sapi memiliki proporsi asam lemak jenuh (saturated fatty acid/SFA) yang relatif tinggi, tetapi rendah proporsi asam lemak tak jenuh ganda atau (polyunsaturated fatty acid/ PUFA) (Mensink et al. 2003). Sebuah studi mengenai asam lemak ditemukan bahwa asupan asam lemak jenuh yang relatif tinggi (SFA) berpotensi meningkatkan kolesterol low density lipoprotein (LDL) dalam darah manusia sedangkan asupan asam lemak tak jenuh ganda dapat menurunkan kolesterol LDL (Mensink et al. 2003).
Saat ini susu digunakan sebagai makanan pharmaceutical. Diantaranya susu digunakan sebagai media terapi obesitas dan penyakit jantung. Susu yang digunakan untuk keperluan tersebut merupakan susu yang mengandung Conjugated Linoleic Acid (CLA). Asam lemak yang mengandung Conjungated Linoleic Acid (CLA), asam butirat dan spingomielin dalam susu dilaporkan mampu mencegah pertumbuhan dan perkembangan sel kanker. Asam lemak tidak jenuh yang baik bagi kesehatan dikenal dengan sebutan asam lemak tak jenuh Omega 3, Omega 6 dan Omega 9. Saat ini kebutuhan akan makanan pharmaceutical semakin meningkat seiring dengan bertambahnya kejadian penyakit degeneratif seperti jantung, tekanan darah tinggi, kencing manis dan obesitas.
Komposisi asam lemak pada susu sangat dipengaruhi oleh faktor genetik dan lingkungan. Lemak susu sapi perah biasanya terdiri dari 50-70% asam lemak jenuh (SFA), 20-40% asam lemak tidak jenuh tunggal (MUFA), dan sisanya dalam jumlah relatif kecil 1-5% asam lemak tidak jenuh ganda (PUFA). Meskipun persentase dari asam lemak tak jenuh dari susu sapi sangat dipengaruhi oleh faktor nutrisi dari pakan ternak (Salter et al. 2007), namun terdapat variasi yang cukup jelas antara individu dalam bangsa maupun antara bangsa sapi perah (Soyeurt et al. 2006).
2
memberikan kontribusi terhadap peningkatan persentase lemak susu dari 3,66% pada tahun 1950 menjadi 4,42% pada tahun 2005 (NRS 2006).
Seleksi pemuliaan secara tradisional membutuhkan recording fenotip yang lengkap. sebaliknya teknologi dengan penanda molekuler genetik memberikan manfaat untuk mempercepat kemajuan seleksi secara genetik. Seleksi berdasarkan marker genetik untuk penciri sifat tertentu yang bernilai ekonomis akan menjadikan seleksi berlangsung lebih awal. Beberapa penelitian berkaitan dengan usaha untuk mengubah komposisi asam lemak telah dilakukan melalui perbaikan nutrisi pakan (Hossein et al. 2011). Pada tingkat molekuler, identifikasi keragaman gen penyandi enzim yang berperan dalam pembentukan lemak dan asam lemak juga telah dilaporkan. Schennink et al.(2007) menyatakan bahwa nilai heritabilitas asam lemak susu tinggi untuk asam lemak jenuh rantai pendek sampai sedang (C4:0-C16:0), heritabilitas sedang untuk asam lemak jenuh dan tidak jenuh rantai panjang (C18).
Suatu studi pemetaan lokus sifat kuantitatif (QTL) pada sapi menghasilkan identifikasi polimorfisme (K232A) dalam pengkodean gen untuk asil
CoA:diasilgliserol 1 acyltransferase (DGAT1), yang merupakan kunci enzim dalam sintesis trigliserida dan memiliki efek kuat pada persentase lemak susu dan karakteristik produksi susu lainnya (Grisart et al. 2002; Winter et al. 2002). Berdasarkan penelitian Shorten et al. (2004) dilaporkan bahwa peningkatan produksi susu karena alel DGAT1 232K diduga akan mengakibatkan komposisi lemak lebih jenuh. Pengaruh dari mutasi DGAT1 pada komposisi lemak susu sapi perah di Indonesia belum banyak dilakukan. Oleh sebab itu, penggunaan gen DGAT1 sebagai marka gen perlu diverifikasi pada sapi perah di Indonesia.
Tujuan Penelitian
Tujuan dari penelitian ini antara lain untuk mengidentifikasi varian genetik dari gen DGAT1 pada sapi FH dan menguji pengaruh varian genotipe dari gen DGAT1 terhadap produksi susu dan profil asam lemak susu pada sapi Friesian Holstein.
Manfaat Penelitian
Peluang pemanfaatan gen DGAT1 sebagai salah satu gen pengontrol kadar lemak susu dapat digunakan sebagai marka dibantu seleksi (MAS) dalam menghasilkan susu dengan kandungan asam lemak tidak jenuh yang tinggi. Manfaat dalam jangka panjang dimungkinkan untuk membentuk breed sapi perah Friesian Holstein yang mampu menghasilkan susu yang menyehatkan.
Ruang Lingkup Penelitian
3
di stasiun bibit pemerintah (BBPTU Sapi Perah Baturraden, BET Cipelang, BPPT Cikole) dan sapi FH yang berasal dari peternakan semi intensif yaitu di peternakan rakyat Pasir Kemis dan Cilumber. Dua peternakan rakyat ini merupakan binaan KPSBU Lembang. Sumbangan pejantan hasil inseminasi buatan (IB) terhadap keragaman genetik gen DGAT1 pada betina turunannya, dilihat melalui analisa sampel DNA yang berasal dari Balai Inseminasi Buatan (BIB) Lembang dan Balai Besar Inseminasi Buatan (BBIB) Singasari.
4
Gambar 1 Ruang lingkup penelitian pengaruh gen DGAT1 terhadap produksi dan profil asam lemak susu
Sapi Perah di BBPTU Sapi Perah Baturraden
Fenotipe Genotipe
Produksi susu
Genotyping Gen DGAT1 Menggunakan metode PCR-RFLP
Asosiasi keragaman Gen DGAT1 terhadap produksi susu, kualitas susu dan profil asam lemak susu
Polimorfisme Pengambilan sampel darah
Kualitas susu Ekstraksi DNA
5
2 TINJAUAN PUSTAKA
Sapi Friesian Holstein
Data populasi dan produksi peternakan di Indonesia menunjukkan bahwa gambaran kondisi peternakan sapi perah di Indonesia masih dominan tersebar di Pulau Jawa seperti di Provinsi Jawa Timur, Jawa Barat dan Jawa Tengah, yang sebagian besar adalah bangsa sapi Friesian Holstein (FH). Populasi sapi perah pada tahun 2013 diperkirakan sebanyak 611.940 ekor lebih tinggi dibandingkan populasi pada tahun 2012 yaitu sekitar 597.210 ekor dengan tingkat pertumbuhan 2.41%. Produksi susu pada tahun 2013 sebesar 959.700 ton sedikit menurun dibandingkan pada tahun 2012 yaitu sebesar 974.690 ton (Direktorat Jenderal Peternakan 2013).
Sapi Friesian Holstein (FH) adalah bangsa sapi perah yang berasal dari Belanda dari propinsi Belanda Utara dan propinsi Friesland Barat (Schmidt et al. 1988). Sapi FH mempunyai karakteristik dengan warna bulu hitam dan putih dan memiliki beberapa keunggulan, diantaranya adalah mempunyai tubuh luas ke belakang, sistem dan bentuk perambingan yang baik, bentuk puting yang simetris, serta memiliki efisiensi pakan yang baik untuk dialihkan menjadi produksi susu (Blakely dan Bade 1998). Selain itu sapi FH memiliki temperamen tenang, kadar lemak susu 3.5-3.7% dengan warna lemak kuning membentuk butiran-butiran (globula) sehingga aman untuk konsumsi susu segar, bahan kering tanpa lemak (BKTL) 8.5% dan rata-rata produksi susu per tahun 5750-6250 kg. Adapun klasifikasi zoologi sapi FH menurut (Tyler dan Ensminger 2006) sapi perah termasuk ke dalam Divisi Animalia, Filum Chordata, Kelas Mamalia, Ordo Artyodactyla, Famili Bovidae, Genus Bos, Spesies Bos Taurus. Sapi yang berasal dari Bos Taurus yang banyak dikembangkan ada lima bangsa yaitu (Holstein, Brown Swiss, Ayshire, Guernsey dan Jersey.
Produksi Susu
Sapi FH memiliki produksi susu tertinggi dibandingkan bangsa-bangsa sapi perah lainnya dengan persentase kadar lemak susu paling rendah yaitu 3.65% dan memiliki produksi susu per laktasi sebesar 7245 kg (Schmidt et al. 1988). Produksi susu yang tinggi diakibatkan sapi FH mempunyai ambing yang besar sehingga mampu memproduksi susu lebih banyak (Webster 1993).
6
Payne (1990) menyatakan bahwa suhu nyaman untuk Sapi Friesian Holstein untuk berproduksi susu adalah 5ºC hingga 20ºC dan kelembaban 55-65%, dengan produksi optimal terjadi pada suhu sekitar 10ºC sedangkan suhu kritis untuk sapi FH adalah 27ºC (McDowell 1972).
Produktivitas sapi perah dapat dilihat dengan cara mengukur jumlah produksi susu selama satu masa laktasi. Masa laktasi adalah masa sapi sedang menghasilkan susu, yakni selama 10 bulan (305 hari) antara saat beranak dan masa kering kandang. Produksi susu total untuk setiap periode laktasi bervariasi, namun umumnya puncak produksi dicapai pada umur 6-7 tahun atau pada laktasi ketiga dan keempat. Menurut (Tyler dan Ensminger 2006) produksi susu terbanyak akan dicapai pada umur 7-8 tahun. Umur sapi juga mempengaruhi jumlah produksi susu yang dihasilkan. Semakin umur sapi bertambah akan diikuti dengan penurunan produksi secara bertahap. Produksi susu pada laktasi pertama (umur 2 tahun) adalah 77%, laktasi kedua (umur 3 tahun) 87%, laktasi keempat (umur 5 tahun) 94%, dan laktasi kelima (umur 6 tahun) 98%.
Sapi FH yang dipelihara di Indonesia mempunyai produksi susu lebih rendah dibanding potensi genetik di daerah asalnya, hal ini dikarenakan suhu dan kelembaban yang tinggi. Penelitian (Anggraeni et al. 2008) melaporkan rataan produksi susu lengkap sapi FH pada peternakan di BBPT-SP Cikole Lembang Jawa Barat berkisar 4.083-5.240 kg pada periode laktasi pertama sampai ke empat sedangkan menurut Wicaksono (2004) produksi susu sapi FH selama 305 hari per ekor pada periode laktasi ke 1, 2, 3, 4 dan 5 berturut-turut adalah 3438,23 kg, 3378,77 kg, 3527,79 kg, 3383,71 kg dan 3255,69 kg..
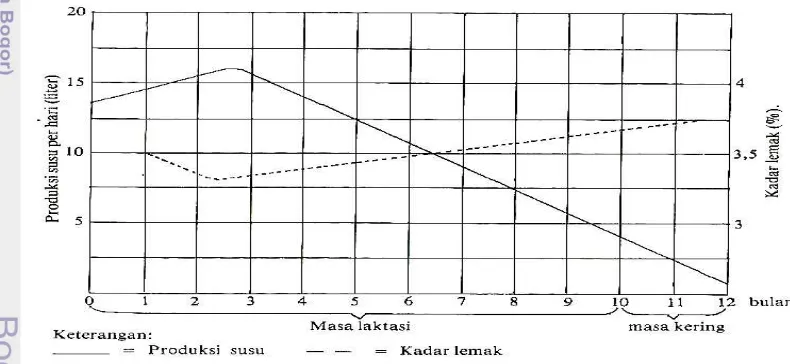
Tilman et al. (1986) menyatakan bahwa produksi susu sapi perah mengikuti pola yang teratur pada setiap laktasi. Produksi susu akan naik selama 45-60 hari setelah sapi beranak hingga mencapai puncak produksi dan kemudian turun secara perlahan-lahan hingga akhir laktasi (Gambar 2). Tingkat produksi susu dalam satu laktasi ikut mempengaruhi tingkat konsumsi bahan kering ransum, dimana terlihat bahwa produksi susu tinggi akan mengakibatkan konsumsi bahan kering juga menjadi tinggi.
7
Asam Lemak Susu
Lemak merupakan komponen yang paling penting dalam susu, yang memiliki bentuk butiran dan tersusun dalam susu sebagai emulsi lemak dalam air. Persentase lemak susu bervariasi antara 2,4% – 5,5%. Lemak susu terdiri atas trigliserida yang tersusun dari satu molekul gliserol dengan tiga molekul asam lemak melalui ikatan-ikatan ester. Lemak susu mengandung asam lemak jenuh sebesar 65-75%, asam lemak tidak jenuh sebesar 25-30%, dan asam lemak tidak jenuh ganda sebesar 4% (Buckle et al. 2007). Kandungan lemak pada susu berperan penting pada flavor, citarasa di mulut (mouthfeel) maupun kestabilan produk hasil olahan susu.
Sebagian lemak susu disintesis di dalam kelenjar ambing, yaitu 50% berasal dari asam lemak rantai pendek (C4-C14) berupa asetat dan beta hidroksi butirat yang dihasilkan oleh fermentasi selulosa di dalam rumen, sebagian lagi berasal dari asam lemak rantai panjang (C16-C18) dari makanan dan cadangan lemak tubuh (Palmquist 2006). Sumber pembentukan lemak susu ada tiga yaitu glukosa, triasilgliserol dari bahan makanan atau yang dibentuk oleh bakteri rumen dan asam lemak yang disintesis oleh kelenjar ambing (Tyler dan Ensminger 2006).
Hubungan produksi susu dengan kadar lemak susu ditunjukkan pada Gambar 3. Pada Gambar 3 terlihat bahwa kandungan lemak terlihat menurun ketika produksi susu meningkat. Begitu juga sebaliknya, kandungan lemak akan meningkat ketika produksi susu menurun.
Gambar 4. Grafik Produksi Susu dan Kadar Lemak (Siregar, 1995)
Gambar 3 Grafik hubungan produksi susu dengan kadar lemak susu
8
sisanya dalam jumlah relatif kecil 1-5% asam lemak tidak jenuh ganda (PUFA) (Salter et al. 2007). Meskipun persentase dari asam lemak susu sapi sangat dipengaruhi oleh faktor nutrisi dari pakan ternak (Salter et al. 2007) namun terdapat variasi yang cukup jelas antara individu dalam bangsa maupun antara bangsa sapi perah (Soyeurt et al. 2006).
Berdasarkan panjang rantai karbon, asam lemak digolongkan menjadi rantai pendek (C2-C6), rantai sedang (C8-C12) dan rantai panjang (C14-C24) (Doyle 1997). Berdasarkan derajat kejenuhannya, asam lemak terbagi menjadi asam lemak jenuh
(Saturated Fatty Acid/SFA), asam lemak tak jenuh tunggal (Mono Unsaturated Fatty Acid/MUFA) dan asam lemak tak jenuh ganda (PolyUnsaturated Fatty Acid/PUFA)
(Silalahi 2000). Berdasarkan isomer geometrik, asam lemak terbagi menjadi asam essensial dan non essensial. Asam lemak essensial tidak dapat disintesa oleh ternak sehingga asam lemak essensial dalam susu sapi berasal dari ransum. Asam lemak essensial dalam susu sapi merupakan asam lemak dengan jumlah karbon 18 atau lebih seperti asam stearat (18:0), asam oleat (18:1), asam linoleat (18:2), asam linolenat (18:3), EPA (20:5) dan DHA (22:6) yang terkandung dalam susu sapi.
Absorbsi asam lemak yang berasal dari ransum dalam plasma menunjukkan asam lemak dalam ransum yang terabsorbsi dan terinkorporasi dalam lemak, yang dihasilkan oleh sel intestinal dan dibawa dalam plasma ke jaringan mamari. Asam lemak dalam lemak plasma (asam lemak plasma) berasal dari asam lemak dalam ransum yang terabsorbsi dan terinkorporasi dalam lemak dalam sel enterosit. Asam lemak dalam plasma juga dapat berasal dari hasil perombakan cadangan lemak dalam
pool lemak atau jaringan adiposa. Selain itu, asam lemak dalam plasma berasal dari asam lemak dalam biomassa bakteri dan protozoa rumen yang dicerna dalam abomasal dan diabsorbsi oleh sel enterosit, lalu diesterifikasi menjadi lemak. Selanjutnya lemak ini bergabung dengan khilomikron dan VLDL dan dibawa oleh darah untuk ditranspor ke jaringan mamari (Rashid et al. 2007).
9
Fatty Acid/TUFA) tidak hanya meningkatkan kadar kolesterol LDL, tetapi secara bersamaan juga menurunkan kadar kolesterol HDL.
Asam lemak tidak jenuh ganda (Poly Unsaturated Fatty Acid/PUFA)
omega-3 adalah asam lemak yang mengandung dua atau lebih ikatan rangkap, dengan ikatan rangkap terakhir terletak pada atom karbon ketiga dari ujung metil rantai asam lemak. Asam alfa linolenik (ALA, 18:3), asam eikosapentaenoik (EPA, 20:5), dan asam dokosaheksaenoik (OHA, 22:6) adalah asam lemak omega-3 yang paling umum. Asam lemak Omega-3 sudah terbukti mempunyai dampak menguntungkan dalam pencegahan penyakit kardiovaskuler, kanker, Alzheimer dan schizoprenia. Khususnya, OHA memainkan peran penting dalam perkembangan otak dan retinal bayi. Sebagai lemak esensial bagi manusia, asam lemak omega-3 perlu ditambahkan pada makanan untuk memelihara tubuh dan otak dalam kondisi puncak.
Pada penyakit kardiovaskuler, asam lemak omega-3 telah menunjukkan berdampak menguntungkan pada sistim kardiovaskuler. Resiko dari terkena penyakit diabetes, obesitas, asma, dan lain-lain terbukti berkurang dengan meningkatkan konsumsi omega-3 (Simopoulos 2002). Studi terbaru menunjukkan bahwa asam lemak omega-3 bisa bergabung dengan membran sel jantung, sehingga berdampak kardioprotektif terhadap beberapa penyakit (Masson et al. 2007). Asam lemak Omega-3 mempunyai banyak manfaat kesehatan dan harus dimasukkan dalam diet manusia (Etherton dan Innis 2007).
Polimorfisme gen diacylglicerol acyltransferase1 (DGAT1)
Pada sapi Bos Taurus, gen Diacylglicerol acyltransferase1 (DGAT1) terletak pada kromosom 14 dan mengkode enzim asil KoA. Diasilglicerol-acyltransferase
memainkan peranan penting dalam metabolism sel diasilgliserol dalam proses fisiologis, seperti penyerapan lemak usus, penyerapan lipoprotein, pembentukan jaringan adiposa, dan laktasi, serta terlibat dalam metabolisme triacylgliserol pada eukariot yang lebih tinggi (Cases et al.1998).
Grisart et al. (2001) menegaskan bahwa gen kandidat yang kuat dalam mengkontrol kandungan lemak susu adalah diasilgliserol acyltransferase (DGAT1). Substitusi protein lisina/alanina (K232A) dikodekan oleh gen DGAT1 telah terbukti berhubungan langsung dengan kandungan lemak susu dari banyak bangsa sapi perah seperti Friesian Holstein, Fleckvieh, dan Jersey (Grisart et al. 2001; Winter et al. 2002). Untuk sapi perah, varian lisin (K) dari DGAT1 terkait dengan kandungan kadar lemak susu yang tinggi, sedangkan jika lisin digantikan oleh alanin akan terkait dengan susu kadar lemak rendah (Grisart et al. 2004). Selain itu, Thaller et al. (2003) menunjukkan bahwa alel lisin DGAT1 juga berpengaruh positif terhadap intramuskuler kadar lemak pada Charolais dan keturunan Holstein silangan.
10
pertama kali oleh Schennink (2007). Dalam hasilnya disebutkan bahwa 232K diasosiasikan dengan fraksi yang lebih besar dari C16:0; fraksi yang lebih kecil dari C14:0, asam lemak tak jenuh C18, dan CLA (Conjugated Linoleic Acid), dan rasio yang lebih tinggi SFA/UFA. DGAT1 mengkatalisis langkah terakhir dalam sintesis trigliserida dimana hasil esterifikasi dari lemak asil-KoA ke posisi sn-3 dari sebuah diasilgliserol.
Hubungan varian genetik gen DGAT1 dengan sifat produksi susu dan kualitas susu
Protein Diacylglicerol acyltransferase1 (DGAT1) adalah sebuah enzim yang mengkatalisis tahapan terakhir pada sintesis trigliserida. Sejumlah mutasi pada gen DGAT1 banyak ditemukan pada sapi tetapi hanya satu lokasi di exon 8 pada posisi basa 10433 dan 10434 yang menyebabkan substitusi lisin menjadi alanin (K232A), yang telah terbukti terkait dengan sifat produksi susu (Woszuk et al. 2008). Woszuk
et al. (2008) melaporkan keragaman gen DGAT1 menggunakan metode Polymerase Chain Reaction-Restriction Fragment Length Polymorphism (PCR-RFLP) dengan enzim pemotong CfrI pada sapi perah bangsa Polish Holstein Friesian (PHF). Ditemukan ada tiga genotipe KK, KA dan AA serta dua alel K dan A. Frekuensi genotipe KK, KA dan AA masing-masing adalah 27%, 55% dan 18% sedangkan frekuensi alel K dan A masing-masing adalah 54% dan 46%.
Oshima dan Serrano (2003) melaporkan bahwa interaksi antara gen DGAT1 dan gen regulator hormon pertumbuhan (Pit1) terhadap produksi susu pada sapi Baja California Holstein memiliki pengaruh yang signifikan pada total produksi susu. Substitusi alel A menjadi K pada lokus DGAT1 mengakibatkan peningkatan produksi susu sebanyak 263.22 kg. Hasil ini berbeda dengan studi awal yang dilaporkan Grisart et al. (2002) dan Spelman et al. (2002).
Lokus = AY065621
Panjang = 14117 bp
Gen = 693-887, 1465-1566, 3605-12168
Sekuen depan = 3604
Exon 1 = 3605 – 3795 = 190 bp Intron 1 = 3796 – 7412 = 3616 bp
Exon 2 = 7413 – 7500 = 87 bp Intron 2 = 7501 – 9443 = 1942 bp
Exon 3 = 9444 – 9484 = 40 bp Intron 3 = 9485 – 9563 = 78 bp
Exon 4 = 9564 – 9649 = 85 bp Intron 4 = 9650 – 9741 = 91 bp
Exon 5 = 9742 – 9794 = 52 bp Intron 5 = 9795 – 10009 = 214 bp
Exon 6 = 10010 – 10115 = 105 bp Intron 6 = 10116 – 10204 = 88 bp
11
Exon 8 = 10419 – 10493 = 74 bp Intron 8 = 10494 – 10563 = 69 bp
Exon 9 = 10564 – 10667 = 103 bp Intron 9 = 10668 – 10757 = 109 bp
Exon 10 = 10758 – 10796 = 38 bp Intron 10 = 10797 – 10862 = 65 bp
Exon 11 = 10863 – 10904 = 41 bp Intron 11 = 10905 – 10977 = 72 bp
Exon 12 = 10978 – 11022 = 44 bp Intron 12 = 11023 – 11096 = 73 bp
Exon 13 = 11097 – 11209 = 112 bp Intron 13 = 11210 – 11296 = 86 bp
Exon 14 = 11297 – 11362 = 65 bp Intron 14 = 11363 – 11448 = 85 bp
Exon 15 = 11449 – 11536 = 87 bp Intron 15 = 11537 – 11617 = 80 bp
Exon 16 = 11618 – 11680 = 62 bp Intron 16 = 11681 – 11752 = 71 bp
Exon 17 = 11753 – 11908 = 155 bp
Gambar 4 Rekonstruksi struktur gen DGAT1 berdasarkan sekuens gen DGAT1 di GenBank (Kode Akses AY065621) (Grisart et al. 2002)
Memodifikasi profil asam lemak susu melalui teknologi pakan
Lemak susu merupakan faktor penentu penting dari kualitas susu. Asam lemak jenuh dianggap menghasilkan efek negatif, sedangkan yang lain (18:01, 18:02 isomer, dan 18:03 n-3) memiliki potensi efek positif pada kesehatan manusia (Parodi, 2005). Profil asam lemak susu bisa diubah secara substansial dengan memanipulasi pola makan ternak (Bandara 1994). Peningkatan kadar asam oleat dan asam linoleat dengan mengurangi asam laurat, miristat dan palmitat dianggap paling diinginkan dari perspektif kesehatan manusia dan juga memberikan manfaat tambahan dalam menghasilkan produk susu (mentega) yang lebih lembut. Bandara (1997) menyatakan perubahan dalam profil asam lemak produk susu jika diterapkan di negara-negara Barat, merupakan strategi yang potensial untuk menurunkan risiko penyakit jantung koroner (PJK) tanpa merubah pola makan yang biasa dilakukan. Salah satu upaya untuk memodifikasi profil asam lemak susu adalah dengan menggunakan teknologi pakan. Suplementasi pada diet sapi perah dengan biji-bijian yang mengandung UFA (asam lemak tidak jenuh) bisa diberikan pada ternak sapi perah (Kennelley 1996). Sumber utama lemak tak jenuh pada tanaman di antaranya kedelai, kanola, dan minyak biji bunga matahari.
12
3 METODE
Waktu dan Lokasi Penelitian
Penelitian ini dilaksanakan selama sebelas bulan, yaitu dari bulan Maret 2012 sampai dengan Februari 2012. Penelitian ini dilaksanakan di Laboratorium Genetika Molekuler Ternak, bagian Pemuliaan dan Genetika, Departemen Ilmu Produksi dan Teknologi Peternakan, Fakultas Peternakan, Institut Pertanian Bogor.
Bahan dan Alat Sampel Darah Sapi Perah FH
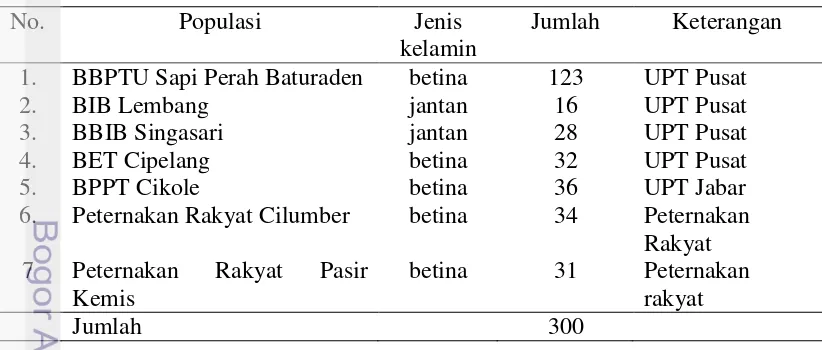
Sampel darah yang digunakan adalah 300 ekor diambil dari 7 populasi di Jawa Tengah dan Jawa Barat (BBPTU Sapi Perah Baturraden, BIB Lembang, BBIB Singasari, BET Cipelang, BPPT Cikole, peternakan rakyat Cilumber dan Pasir kemis). Sedangkan untuk melihat hubungan keragaman gen DGAT1 dengan produksi dan profil asam lemak susu hanya digunakan sampel dari BBPTU Sapi Perah Baturraden. Sampel darah sapi FH diambil melalui vena coccygea (ekor), menggunakan tabung vaccutainer yang mengandung antikoagulan. Sampel tersebut kemudian disimpan dalam termos es dan suhunya dipertahakan sekitar 4°C.
Tabel 1 Karakteristik sampel darah sapi perah FH
No. Populasi Jenis
kelamin
Jumlah Keterangan 1. BBPTU Sapi Perah Baturaden betina 123 UPT Pusat
2. BIB Lembang jantan 16 UPT Pusat
3. BBIB Singasari jantan 28 UPT Pusat
4. BET Cipelang betina 32 UPT Pusat
5. BPPT Cikole betina 36 UPT Jabar
6. Peternakan Rakyat Cilumber betina 34 Peternakan Rakyat 7 Peternakan Rakyat Pasir
Kemis
betina 31 Peternakan rakyat
Jumlah 300
Sampel Susu
13
produksi susu diperoleh berdasarkan uji satu hari dengan menjumlahkan produksi susu pagi dan sore hari dalam kisaran periode laktasi 1-6 dan bulan laktasi 1-9. Sedangkan jumlah sampel yang diambil untuk analisa profil asam lemak sebanyak 40 sampel yang berasal dari 40 ekor sapi perah periode laktasi.
Primer
Primer untuk mengamplifikasi ruas gen DGAT1 mengikuti (Grisart et al. 2002), dengan produk hasil amplifikasi sepanjang 411 pb. Runutan Primer Forward
dan Reverse dapat dilihat pada tabel 2. Tabel 2 Runutan primer yang digunakan
primer Sekuens primer Ukuran
produk
Target amplifikasi DGAT-1 F 5’-GCACCATCCTCTTCCTCAAG-3’ 411 pb Ekson 8 DGAT-1 R 5’-GGAAGCGCTTTCGGATG-3’
Bahan dan Alat untuk Pengambilan Sampel
Bahan yang digunakan untuk pengambilan sampel adalah alkohol 70%, es batu dan kapas. Alat yang digunakan antara lain jarum venoject, tabung vaccutainer 10ml yang mengandung EDTA dan termos.
Bahan dan Alat untuk Ekstraksi DNA
Bahan yang digunakan untuk ekstraksi DNA adalah sampel darah 200 µl, 800 µl lysis buffer, 40 µl SDS 10% (Sodium Dodecyl Sulfat), buffer TEN (10 mM Tris-HCl, 5 mM EDTA dan 10 mM NaCl), 10 µl enzim Proteinase K mg/ml, 350 µl 1 x STE (5 M NaCl, 2 M Tris HCl, 0.2 M EDTA), 400 µl phenol, 400 µl CIAA (Chloroform : Isoamyl alkohol = 24 :1), 40 µl NaCl 5 M, 800 µl etanol absolut, 800 µl etanol 70 %, 100 µl buffer TE 80% (Tris EDTA). Peralatan yang digunakan adalah tabung eppendorf 1,5 ml, satu set mikro pipet, tip pipet (100 ; 200 dan 1000 l), vortex, sentrifuge, nutating mixer, inkubator, refigerator dan freezer.
Bahan dan Alat untuk Reaksi Polymerase Chain Reaction (PCR)
Bahan yang digunakan dalam reaksi PCR adalah 1 µl sampel DNA, 0.5 µl primer DGAT1 (Forward dan Reverse), 0,5 µl dNTPs, 0,5 µl MgCl2, 1,5 µl 10 x
14
mesin PCR (Thermal Cycler GeneAmp PCR System 2400 dan 9600, Perkin Elmer),
showcase.
Bahan dan Alat untuk Elektroforesis
Bahan yang digunakan yaitu loading dye (bromthymol blue 0.01%, xylene cyanol 0.01% dan gliserol 50%) dan untuk membuat gel agarose 1.5% adalah agarose 0.45 g, 0.5 x TBE 30 ml dan 2.5 µl ethidium bromide (EtBr). Alat-alat yang digunakan adalah microwave, stirer, magnetik stirer, gelas ukur, tabung kimia, gel tray, pencetak untuk sumur (comb), sarung tangan, tip, mikropipet satu set alat elektroforesis gel agarose (MUPID) dan UV transilluminator.
Bahan dan Alat untuk Genotyping
Bahan-bahan yang digunakan yaitu enzim restriksi EaeI dengan buffernya,
loading dye bromthymol blue 0.01%, Xylene cyanol 0,01% dan gliserol 50%) dan untuk membuat gel agarose 2% adalah agarose 0.6 g, 0,5 x TBE 30 ml dan 2.5 µl
EtBr. Peralatan yang akan digunakan microwave, stirer, magnetik stirer, gelas ukur, tabung kimia, gel tray, pencetak untuk sumur (comb), power supply electrophoresis
100 volt, tip, mikropipet, sarung tangan, satu set alat elektroforesis gel agarose (MUPID) dan UV transilluminator.
Prosedur Ekstraksi DNA
Ektraksi DNA dilakukan dari sampel darah sapi FH mengikuti metode (Sambrook et al. 1989) yang dimodifikasi.
Preparasi Sampel. Sampel darah yang disimpan dalam tabung vaccutainer yang mengandung EDTA diambil sebanyak 200 µl dan dimasukan ke dalam tabung 1,5 ml, kemudian ditambahkan lysis buffer sebanyak 800 µl. Sampel kemudian dikocok kuat dengan vortex selama 15 detik dan didiamkan pada suhu ruang selama 5 menit setelah itu disentrifuge pada kecepatan 8000 rpm selama 5 menit sehingga terbentuk 2 lapisan, bagian supernatan dibuang.
Degradasi Protein. Sampel yang telah bersih dari alkohol ditambahkan 40 µl sodium dodesil sulfat 10%, 10 µl proteinase K 5 mg/ml, 1 x STE (sodium tris EDTA) sampai volume 350 µl, dan. Tabung eppendorf dikocok pelan sambil disentil menggunakan jari agar tidak terjadi endapan. Campuran dikocok pelan menggunakan nutating mixer di dalam inkubator pada suhu 55ºC selama 2 jam.
15
Presipitasi DNA. Molekul DNA yang larut dalam fase air dipisahkan dari fase
phenol dengan disentrifugasi pada kecepatan 12.000 rpm selama 5 menit.. Setelah terbentuk fase DNA, DNA tersebut diambil sebanyak 400 µl dan dipindahkan ke tabung 1,5 ml yang baru. Kemudian ditambahkan NaCl 5M sebanyak 40 µl dan etanol absolut sebanyak 800 µl .Tabung yang berisi DNA kemudian disimpan di
freezer pada suhu -20ºC selama semalam. Molekul DNA kemudian dipisahkan dari etanol absolut dengan cara disentrifugasi pada kecepatan 12.000 rpm selama 5 menit, kemudian supernatan yang diperoleh dibuang. Endapan DNA yang diperoleh kemudian dicuci dengan alkohol 70% sebanyak 800 µl dan disentrifugasi lagi dengan kecepatan 12.000 rpm selama 5 menit. Supernatan yang diperoleh kemudian dibuang sehingga didapatkan endapan molekul DNA. Tabung yang berisi DNA kemudian dikeringkan dalam ruang terbuka sampai alkohol benar-benar hilang, dengan posisi tabung terbalik. Endapan DNA yang telah bersih dari alkohol kemudian ditambahkan 100 µl TE (Tris EDTA) 80%. Sampel DNA disimpan pada suhu -20ºC dan siap untuk digunakan.
Amplifikasi Ruas Gen DGAT1
Amplifikasi ruas gen DGAT1 dilakukan dengan metode PCR (Polymerase Chain Reaction). Pereaksi yang digunakan untuk amplifikasi ruas gen target adalah1 µl sampel DNA, 0,5 µl primer DGAT1 (Forward dan Reverse), 0.5 µl dNTPs, 0.5 µl MgCl2, 1.5 µl 10 x buffer, 0.1 µl enzim taq polymerase, 10.9 µl Destilation Water
(DW) dalam larutan total 14 µl. Amplifikasi in vitro dengan mesin thermal cycler
dilakukan dengan kondisi denaturasi awal pada suhu 95ºC selama 5 menit, 35 siklus yang terdiri dari denaturasi pada suhu 95ºC selama 30 detik, penempelan primer (annealing)pada suhu 60ºC selama 1 menit dan pemanjangan DNA baru pada suhu 72ºC selama 1 menit, serta pemanjanganakhir pada suhu 72ºC selama 5 menit. Elektroforesis Produk PCR
16
Penentuan Genotipe dengan Pendekatan PCR-RFLP
Penentuan genotipe masing-masing individu dilakukan dengan pendekatan
polymerase chain reaction dan restriction fragment length polymorphism (RFLP) menggunakan enzim restriksi spesifik EaeI yang mengenali situs mutasi. Sebanyak 5 µl produk PCR dipindahkan ke dalam tabung 0.5 ml dan ditambahkan 2 µl enzim restriksi EaeI, 0.7 µl buffer 4 dan 0.8 µl DW. Campuran tersebut kemudian disentrifugasi dengan kecepatan 5000 rpm selama 10 detik dan diinkubasi pada suhu 370C selama 16 jam. Hasil PCR-RFLP divisualisasikan pada gel agarose 2% dengan bufer 0,5x TBE (tris borat EDTA) yang difungsikan pada tegangan 100 V selama 45 menit yang diwarnai dengan etidium bromida diatas UV trans illuminator.
Uji kualitas susu
Pengujian kualitas susu dilakukan menggunakan milk analyzer yang meliputi uji kadar lemak, protein, laktosa, bahan kering tanpa lemak (BKTL), berat jenis dan air.
Analisa profil asam lemak susu
Pengambilan sampel dilakukan dengan mengambil sampel susu segar yang berasal dari 40 ekor sapi FH di BBPTU Sapi Perah Baturraden. Sapi yang dipilih adalah sapi laktasi yang memiliki umur yang relatif sama tetapi memiliki periode dan bulan laktasi yang berbeda. Waktu pengambilan sampel adalah pagi hari pukul 04.00 WIB dan sore hari pukul 15.00 WIB. Susu diperah dari empat kuartir kemudian ditampung dalam kaleng susu kemudian susu diambil menggunakan gelas ukur sebanyak 100 ml kemudian susu dimasukkan dalam tabung dan diberi label. Susu selanjutnya disimpan dalam sterofom yang diberi es batu di dalamnya kemudian dibawa ke Laboratorium Penelitian dan Pengujian Terpadu (LPPT) UGM Yogyakarta. Pengukuran kandungan asam lemak pada sampel susu dilakukan menggunakan metode Gas Chromatography (AOAC, 2005).
Analisis Data Frekuensi Alel dan Genotipe
Frekuensi alel merupakan rasio suatu alel terhadap keseluruhan alel pada suatu lokus dalam populasi. Frekuensi alel dihitung dengan persamaan menurut Nei dan Kumar (2000) sebagai berikut :
17
Keterangan :
хi = frekuensi alel ke-i
nii = jumlah individu yang yang bergenotipe ii
nij = jumlah individu yang bergenotipe ij
n = jumlah sampel
Frekuensi genotipe dapat diperkirakan dengan menghitung perbandingan jumlah genotipe pada populasi. Dengan menggunakan asumsi sebelumnya, maka frekuensi genotipe AiAi(χii) dapat dihitung dengan persamaan sebagai berikut:
n nii ii
Heterozigositas
Heterozigositas merupakan salah satu indikator keragaman gen dalam populasi yang menunjukkan proporsi heterozigot pada individu yang diambil secara acak. Ho adalah frekuensi heterozigositas pengamatan, Nij adalah jumlah individu
heterozigositas pada lokus ke-1 dan N adalah jumlah individu yang dianalisa.
0
Heterozigositas harapan (He) dan ragam heterozigositas harapan dihitung menggunakan rumus Weir (1996).
He = frekuensi heterozigositas harapan
Pii2 = frekuensi alel ke-i pada lokus 1
n = jumlah alel pada lokus ke-1
Analisis keragaman genetik dan keseimbangan genetik gen DGAT1 diuji dengan menggunakan software PopGene32 versi 1.31.
Analisis Hubungan Genotipe dengan Produksi Susu dan Profil Asam Lemak Susu
18
lemak. Pengujian statistik dilakukan dengan pendekatan ANOVA (analysis of variance) menggunakan prosedur General Linear Model (GLM) dari SAS versi 9.1 (SAS Institute, Garry, NC, USA) dengan model (Mattjik dan Sumertawijaya, 2006):
Yijk = μ + Ai + eijk Keterangan :
Yijk = nilai pengamatan
μ = komponen aditif dari rataan Ai = pengaruh aditif gen DGAT1 €ijk = pengaruh galat perlakuan
4 HASIL DAN PEMBAHASAN
Amplifikasi Gen DGAT1 (Diacylglicerol Acyltransferase1)
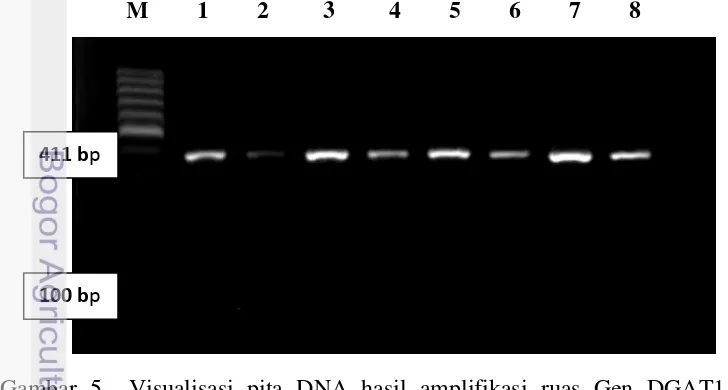
Ruas gen DGAT1 exon 8 sapi perah FH yang berasal dari sejumlah lokasi masing-masing dari BBPTU Sapi Perah Baturraden Purwokerto (123 sampel); BIB lembang (16 sampel); BBIB Singasari (28 sampel); BET Cipelang (32 sampel) dan BPPT Cikole (36 sampel); peternakan Cilumber (34 sampel); dan peternakan Pasir Kemis (31 sampel) telah berhasil diamplifikasi dengan menggunakan teknik
Polymerase Chain Reaction (PCR). Persentase keberhasilan amplifikasi gen DGAT1 dalam penelitian ini adalah 100%. Hasil amplifikasi gen DGAT1 pada gel agarose 1,5% disajikan pada Gambar 5.
M 1 2 3 4 5 6 7 8
Gambar 5 Visualisasi pita DNA hasil amplifikasi ruas Gen DGAT1 pada gel agarose 1,5% M: marker 100 bp, 1-8: nomor sampel
411 bp
19
Salah satu faktor yang menentukan tingkat keberhasilan amplifikasi diantaranya adalah suhu annealing, dimana suhu annealing adalah suhu yang memungkinkan terjadinya penempelan primer pada DNA cetakan selama proses PCR. Suhu annealing yang digunakan dalam penelitian ini adalah 600C selama 1 menit. Suhu ini sama dengan yang digunakan oleh penelitian Schennink et al. (2007). Lacorte et al. (2006) berhasil mengamplifikasi pada suhu annealing yang lebih rendah yaitu 560C selama 120 detik. Muladno (2002) menyatakan bahwa suhu penempelan primer (annealing) berkisar antara 350C sampai dengan 720C, namun suhu yang biasa digunakan adalah 50-600C.
Berdasarkan referensi pada Bank Gen nomor akses AY065621, didapatkan produk PCR hasil amplifikasi berukuran 411 pb (Gambar 5). Hasil penelitian (Lacorte et al. 2006) menunjukkan, jika pemotongan produk PCR 411 pb dengan enzim restriksi EaeI menghasilkan dua pita DNA berukuran 203 dan 208 pb, maka sapi diidentifikasi memiliki genotipe homozygote AA. Jika diperoleh tiga pita DNA berukuran 411, 203 dan 208 pb, diidentifikasi sebagai genotipe AK. Bila pita DNA tidak terpotong sehingga hanya diperoleh satu pita berukuran 411 pb, maka diperoleh genotipe homozigot KK
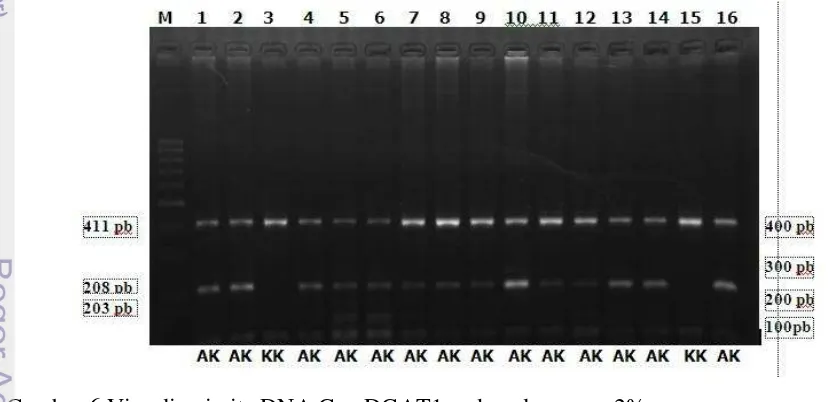
Namun dalam penelitian ini pendeteksian keragaman Gen DGAT1 dengan teknik PCR-RFLP menggunakan enzim EaeI yang dilakukan pada sapi perah Friesian Holstein (FH) hanya didapatkan dua varian genotipe yaitu genotipe KK dan AK (Gambar 6). Sapi FH dikatakan memiliki genotipe KK apabila memiliki satu pita DNA dengan panjang 411 pb sedangkan genotipe AK jika memiliki tiga pita DNA dengan panjang 411, 203 dan 208 pb.
Gambar 6 Visualisasi pita DNA Gen DGAT1 pada gel agarose 2%. M: marker 100bp, 1-16: nomor sampel
20
(mutasi K232A) (Grisart et al. 2002). Situs pemotongan enzim EaeI disajikan pada Gambar 7.
6601 tgggctccgt gctggccctg atggtctaca ccatcctctt cctcaagctg ttctcctacc
Primer Forward
6661 gggacgtcaa cctctggtgc cgagagcgca gggctggggc caaggccaag gctggtgagg 6721 gctgcctcgg gctggggcca ctgggctgcc acttgcctcg ggaccggcag gggctcggct 6781 cacccccgac ccgccccctg ccgcttgctc gtagctttgg caggtaagaa ggccaacggg
gc
6841 ggagctgccc agcgcaccgt gagctacccc gacaacctga cctaccgcgg tgaggatcct 6901 gccgggggct ggggggactg cccggcggcc tggcctgcta gccccgccct cccttccaga 6961 tctctactac ttcctcttcg cccccaccct gtgctacgag ctcaacttcc cccgctcccc 7021 ccgcatccga aagcgcttcc tgctgcggcg actcctggag atggtgaggc ggggcctcgc
Primer Reverse
Gambar 7 Runutan nukleotida ruas gen DGAT1 (GenBank nomor akses AY065621). Posisi primer (cetak tebal tanda panah), aggcca = situs pemotongan enzim
EaeI (cetak tebal bergaris bawah) dan aa = titik mutasi (cetak tebal bergaris bawah).
Perbedaan sekuen alel K dan alel A yang disebabkan mutasi pada asam amino ke 232 disajikan pada Gambar 8.
Alel K : 5’---GTAGCTTTGGCAGGTAAGAAGGCCAACGGG---3’ Alel A : 5’---GTAGCTTTGGCAGGTAAGGCGGCCAACGGG---3’
Gambar 8 Perbedaan sekuen alel K dan A gen DGAT1|EaeI. Situs potong enzim restriksi EaeI (cetak tebal) dengan titik mutasi (garis bawah).
Frekuensi Genotipe dan Alel Gen DGAT1
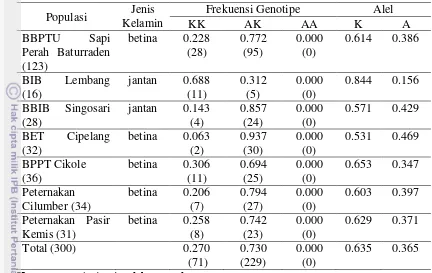
Hasil analisis frekuensi genotipe dan alel sapi FH dari lokasi BBPTU Sapi Perah Baturraden, BIB Lembang, BBIB Singosari, BET Cipelang, BPPT Cikole, Peternakan Cilumber, Peternakan Pasir Kemis disajikan pada Tabel 3.
21
Tabel 3 Frekuensi Genotipe dan Alel Gen DGAT1 pada sapi Friesian Holstein Populasi Jenis
Fenomena tidak adanya genotipe AA dari sapi betina FH, baik di peternakan rakyat maupun di stasiun bibit salah satunya adalah dipengaruhi oleh sumber pejantan inseminasi buatan (IB) yang digunakan. Pejantan yang digunakan untuk IB pada betina laktasi umumnya berasal dari BIB Lembang dan BBIB Singasari. Berdasarkan hasil analisis pada sapi pejantan IB yang berasal dari BIB Lembang dan BBIB Singasari tidak ditemukan genotipe AA dan frekuensi alel A rendah pada pejantan yang diamati.
Frekuensi alel adalah frekuensi relatif dari suatu alel dalam populasi atau jumlah suatu alel terhadap jumlah total yang terdapat dalam suatu populasi (Nei dan Kumar 2000). Dari hasil penelitian ini, gen DGAT1 pada sapi FH dari tujuh populasi yang diamati bersifat polimorfik karena ditemukan dua tipe alel, yaitu alel K dan alel A. Menurut Nei dan Kumar (2000) menyatakan bahwa suatu alel dapat dikatakan polimorfik jika memiliki frekuensi alel sama dengan atau kurang dari 0.99.
Keragaman genotipe gen DGAT1 sapi FH dari tujuh populasi dapat dibandingkan dengan beberapa penelitian sebelumnya. Penelitian ini tidak sesuai dengan Lacorte et al. (2006) yang melaporkan bahwa sapi Holstein di Brazil memiliki tiga varian genotipe yaitu KK (14%), AK (26%) dan AA (60%) dengan genotipe yang tertinggi adalah genotipe AA (60%). Frekuensi alel yang didapatkan yaitu alel K (27%) lebih rendah daripada alel A (73%). Perbedaan hasil genotyping
22
frekuensi genotipe AA kemungkinan disebabkan karena semen pejantan pada sapi Holstein Brazil berasal dari semen yang diimpor dari Amerka Serikat yang berasal dari gene pool yang sudah mengalami adanya seleksi tidak langsung, yang dilakukan secara intensif ke arah produksi susu. Perbedaan jumlah sampel juga mempengaruhi peluang untuk mendapatkan genotipe yang berbeda. Frekuensi alel dan genotipe pada beberapa bangsa sapi perah menurut penelitian sebelumnya dapat dilihat pada Tabel 4.
Tabel 4 Frekuensi genotipe dan frekuensi alel pada beberapa bangsa sapi perah Bangsa Frekuensi genotipe (%) Frekuensi alel (%)
KK AK AA K A
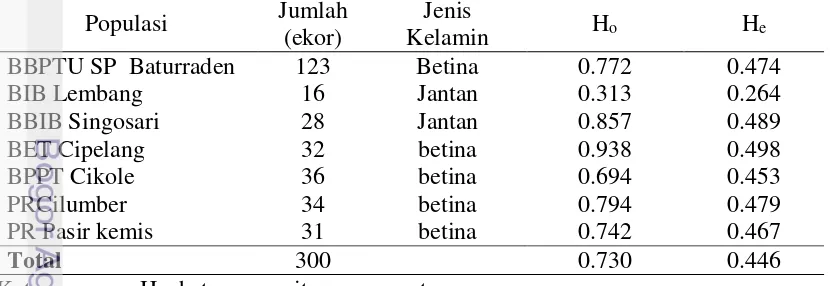
Pendugaan nilai heterozigositas memiliki arti pnting untuk diketahui, yaitu untuk mendapatkan gambaran variabilitas genetik (Marson et al. 2005). Heterozigositas disebut juga sebagai keragaman genetik. Nilai heterozigositas dipengaruhi oleh jumlah sampel, jumlah alel dan frekuensi alel. Hasil analisis heterozigositas populasi sapi FH pada lokasi yang berbeda ditampilkan pada Tabel 5. Tabel 5 Heterozigositas gen DGAT1 pada sapi FH
Populasi Jumlah
Keterangan : Ho=heterozogositas pengamatan
He=heterozigositas harapan
23
pengamatan (Ho) yang tertinggi adalah sapi FH dari BET Cipelang yaitu sebesar 0.938 dan nilai heterozigositas pengamatan (Ho) yang terendah adalah sapi FH di lokasi BBIB Lembang (0.313). Begitu pula nilai heterozigositas harapan (He) yang tertinggi adalah populasi sapi FH dari Cipelang dan yang terendah adalah dari BIB Lembang (0.264). Menurut Javanmard et al. (2005) nilai heterozigositas kurang dari 0.5 mengindikasikan rendahnya variasi gen dalam suatu populasi. Avise (1994) menyatakan bahwa semakin tinggi derajat heterozigositas suatu populasi, maka daya tahan hidup populasi tersebut akan semakin tinggi. Tingginya nilai heterozigositas pengamatan (Ho) juga menunjukkan adanya keragaman alel dalam populasi tersebut. Semakin tinggi nilai heterozigositas dalam populasi, maka semakin besar peluang untuk melakukan seleksi pada populasi tersebut.
Rendahnya nilai heterozigositas pada sampel yang berasal dari stasiun bibit BIB Lembang kemungkinan disebabkan karena sapi perah yang dipelihara merupakan sapi-sapi hasil seleksi terhadap pejantan dengan kualitas genetik yang unggul. BIB Lembang adalah stasiun bibit milik pemerintah dimana tugas pokoknya adalah melaksanakan produksi dan pemasaran semen beku benih unggul ternak serta pengembangan inseminasi buatan untuk kesejahteraan peternak.
Pengaruh varian genotipe gen DGAT1 terhadap produksi dan kualitas susu segar sapi FH
Produksi susu
Pengujian pengaruh varian genotipe gen DGAT1 terhadap produksi susu dilakukan pada stasiun bibit BBPTU Sapi Perah Baturraden, dimana pada stasiun ini menerapkan pola pemeliharaan intensif. Tujuan pengamatan dilakukan di stasiun ini adalah untuk meminimalisasi pengaruh lingkungan dan manajemen pemeliharaan terhadap tingkat produksi susu. Berdasarkan hasil analisis General Linear Model (GLM), genotipe gen DGAT1|EaeI tidak memberikan pengaruh terhadap produksi susu harian (P>0.05). Sapi dengan genotipe AK (12.33 L) menghasilkan produksi susu lebih tinggi dibandingkan KK (11.32 L). Produksi susu secara umum dikontrol oleh faktor luar (eksternal) dan faktor dalam (internal). Faktor eksternal adalah faktor yang berasal dari luar tubuh ternak seperti iklim, jumlah dan kualitas pakan, penyakit dan parasit (Indrijani 2001), sedangkan faktor internal adalah faktor genetik, periode laktasi, frekuensi pemerahan, umur dan ukuran tubuh ternak, masa kering, siklus estrus dan kebuntingan, ketosis dan milk fever (Sudono et al. 2003).
Kadar Protein
24
disintesis dalam sel epitel kelenjar ambing yang dikontrol oleh gen. Standar kadar protein susu sapi perah sesuai SNI susu segar adalah 2.80% (BSN 1998).
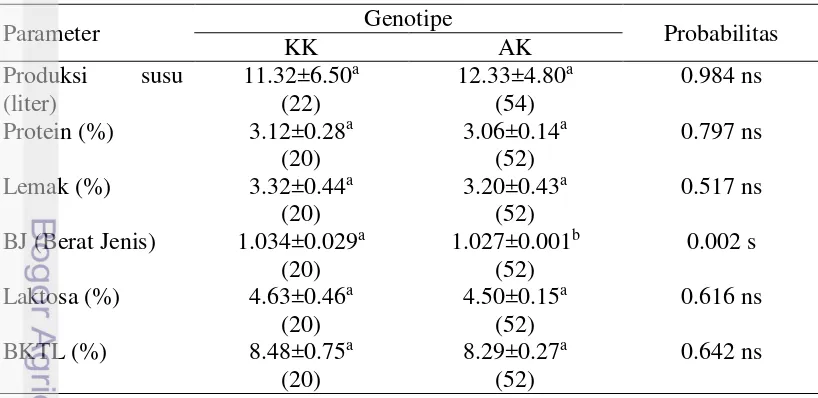
Kadar lemak
Sapi dengan genotipe KK cenderung menghasilkan kadar lemak susu lebih banyak (3.32%) dibanding sapi AK (3.20%).
Bahan Kering Tanpa Lemak (BKTL)
Sapi dengan genotipe KK juga bertendensi menghasilkan Bahan Kering Tanpa Lemak (BKTL) 8.48% lebih tinggi daripada sapi dengan genotip AK (8.295). Berat Jenis (BJ)
Berat Jenis (BJ) lebih tinggi dibandingkan AK (P<0.05). Nilai kadar protein, kadar lemak, Berat Jenis (BJ) dan Bahan Kering Tanpa Lemak (BKTL) pada sapi FH bergenotipe KK dan AK telah memenuhi standar SNI 01-3141-1998 mengenai syarat mutu susu segar (BSN 1998). Berat jenis susu dipengaruhi oleh zat-zat padatan yang terkandung didalam susu seperti lemak, protein, laktosa, vitamin dan mineral. Nilai berat jenis susu dipengaruhi oleh kadar lemak dan kadar padatan tanpa lemak, yang tidak lepas dari pengaruh makanan dan kadar air dalam susu. Menurut SNI mengenai susu segar, syarat minimum BJ pada sapi perah adalah 1.0270 (BSN 1998).
Menurut Ensminger dan Tyler (2006) komposisi susu terdiri atas air 87.2%, lemak 3.7%, bahan kering tanpa lemak (BKTL) 9.1%, protein 3.5%, laktosa 4.9% dan mineral 0.7%.
Tabel 6 Produksi dan kualitas susu segar sapi FH berdasarkan genotipe
Parameter Genotipe Probabilitas
S pada baris yang sama yang diikuti oleh huruf yang berbeda menunjukkan berbeda nyata pada taraf
uji 5% (uji selang berganda Duncan). ns = non signifikan; s = signifikan
25
Tabel 7 Polimorfisme gen DGAT1 pada beberapa bangsa sapi perah Populasi (N) Frekuensi kadar lemak susu dan kadar protein ;
Gautier et al. total,persentase lemak susu dalam suatu volume dan kadar protein; menurunkan produksi susu dan kadar protein total
Schennink et al. 2007
Pengujian pengaruh genotipe gen DGAT1 terhadap profil asam lemak tidak jenuh (unsaturated fatty acid) disajikan pada Tabel 8. Pengamatan komposisi asam lemak susu dilakukan di BBPTU SP Baturraden yang menerapkan pola pemeliharaan intensif. Hal ini bertujuan untuk meminimalkan pengaruh lingkungan dan manajemen pemeliharaan terhadap komposisi asam lemak. Berdasarkan analisa kualitatif menggunakan Gas Cromatography (GC), asam lemak susu yang teridentifikasi ada 22 jenis asam lemak, yang terdiri dari 13 jenis asam lemak jenuh (SFA), 5 jenis asam lemak tidak jenuh tunggal (MUFA), dan 4 jenis asam lemak tidak jenuh ganda (PUFA). Komposisi bruto asam lemak susu dalam penelitian ini berkisar 65.59-67.94% untuk asam lemak jenuh (SFA), 29.19-30.01% untuk asam lemak tidak jenuh tunggal (MUFA) dan 0.94-2.88% untuk asam lemak tidak jenuh ganda (PUFA). Hasil ini sejalan dengan report yang dilakukan oleh Mansson (2001), dimana dikatakan komposisi bruto lemak susu pada sapi Swedish adalah 69,4% asam lemak jenuh dan 30,6% asam lemak tak jenuh.
Asam Lemak Jenuh (saturated fatty acid)
26
bahwa polimorfisme gen DGAT1 dikaitkan lebih banyak asam jenuh dan fraksi yang lebih kecil dari C14.
Tabel 8 Profil asam lemak susu sapi perah Friesian Holstein berdasarkan genotipe Asam Lemak KK (n=6) AK (n=34) Probabilitas