VARIASI SPASIAL EMISI GAS RUMAH KACA (GRK) PADA
EKOSISTEM MANGROVE DI PULAU KELONG, BINTAN
I WAYAN EKA DHARMAWAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis yang berjudul Variasi Spasial Emisi Gas Rumah Kaca (GRK) pada Ekosistem Mangrove di Pulau Kelong, Bintan adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Juni 2016
RINGKASAN
I WAYAN EKA DHARMAWAN. Variasi Spasial Emisi Gas Rumah Kaca (GRK) pada Ekosistem Mangrove di Pulau Kelong, Bintan. Dibimbing oleh NEVIATY P. ZAMANI dan HAWIS H. MADDUPPA.
Ekosistem mangrove dikenal dengan baik dalam mitigasi dampak perubahan iklim di wilayah pesisir dengan mampu menyerap CO2 dan menyimpannya dalam bentuk biomassa. Walaupun demikian, proses biokimia yang terjadi dalam sedimen menghasilkan gas rumah kaca (GRK) ke atmosfer. Mangrove Pulau Kelong bagian tenggara memiliki simpanan karbon yang sangat tinggi, dibandingkan dengan seluruh kawasan mangrove di Kawasan Konservasi Perairan Daerah (KKPD) Kabupaten Bintan. Tujuan dari penelitian ini adalah menganalisis kondisi ekosistem mangrove di Pulau Kelong bagian tenggara mencakup kondisi kajian vegetasi, dekomposisi maupun lingkungan pada tiap zona yang berbeda, yaitu zona dekat darat (L), zona tengah (M) dan zona dekat laut (S); Menghitung besaran laju emisi GRK secara spasial serta menganalisis hubungan kondisi komunitas mangrove, parameter lingkungan dan laju dekomposisi terhadap fluks GRK yang dihasilkan. Hasil penelitian diharapkan dapat digunakan untuk meningkatkan pelindungan kawasan untuk mitigasi perubahan iklim global.
Kondisi ekosistem mangrove di Pulau Kelong memiliki zonasi yang sangat jelas antara zona darat (L) dengan zona tengah (M) dan laut (S). Zonasi terbentuk berdasarkan sebaran jenis mangrove serta parameter lingkungan. Jenis Rhizophora apiculata, mendominasi penuh pada zona (S) dan mulai menurun pada zona (M). Pada zona L, Xylocarpus granatum lebih mendominasi dibandingkan dengan R. apiculata. Parameter suhu, pH, TOC, TP dan TN pada sedimen, menunjukkan kedekatan zona M dengan S dan berbeda nyata dengan L. Persentase tutupan kanopi mangrove pada seluruh zona adalah >75%, sehingga kondisi kesehatan komunitas mangrove masih sangat baik. Laju dekomposisi serasah juga tergolong tinggi (k = 0,025 ± 0,023), yaitu: 0,78 ±0,70 gr/m2/hari (S); 0,83 0,89 gr/m2/hari (M); dan 1,40 0,82 gr/m2/hari (L). Laju emisi GRK ke atmosfer menunjukkan nilai yang cukup rendah jika dibandingkan dengan penelitian lainnya. Tren laju emisi gas metana (CH4) meningkat dari zona L ke zona S dan berlawanan dengan laju CO2. Sementara itu, gas NO2 tidak memiliki pola emisi yang jelas secara spasial. Emisi gas CH4 dan N2O tidak berkorelasi nyata dengan kondisi lingkungan, namun laju emisi gas CO2 sangat dipengaruhi oleh pH dan TOC tanah serta TDS dan salinitas perairan. Cadangan karbon yang tinggi, emisi GRK yang rendah, kondisi komunitas masih alami menunjukkan bahwa kawasan mangrove bagian tenggara Pulau Kelong memiliki potensi mitigasi perubahan iklim yang sangat tinggi.
SUMMARY
I WAYAN EKA DHARMAWAN. Spatial Variation of Greenhouse Gases (GHGs) Emission on Mangrove Ecosystem in Kelong Island, Bintan, as one of perspective for mangrove ecosystem conservation. Supervised by NEVIATY P. ZAMANI and HAWIS H. MADDUPPA.
Mangrove ecosystem is well-known in climate change impact mitigation in coastal area in order to sequester CO2 and reform it to be biomasses. On the other hand, biochemical processess inner sediment produce greenhouse gases (GHGs) through atmosphere. Previous study found that mangrove area in south-easth Kelong Island has the highest carbon stock among other sites in Local Waters Conservation Area (KKPD) Bintan. Objectives of this research are: to analyze mangrove ecosystem condition in south-eastern Kelong island including community structure, decomposition rate and environmental parameters in three different zones; to quantify GHGs emission fluxes among those zones; and to analyze correlation between mangrove ecosystem to GHGs emission. Hopefully, resulted data might be used to upgrade the conserving status for global climate change mitigation.
Mangrove ecosystem in Kelong Island was clearly devided to be three zones, landward (L), middle zone (M) and seaward (S). It was performed based on species composition and environmental parameters. Domination of Rhizophora apiculata was total in seaward and decreased in middle zone. On landward area, Xylocarpus granatum had more domination than R. apiculata. Temperature, pH, TOC and TN of sediment elaborated M and S closeness and differed significantly to L. Canopy coverage analysis showed that all value was more than 75%, representing well standing condition of mangrove community. Decomposition rate leaf manure was also high (k = 0.025 ± 0.023), i.e. 0.78 ±0.70 gr/m2/day (S); 0.83 0.89 gr/m2/day (M); dan 1.40 0.82 gr/m2/day (L). GHGs emission rate showed the lower value comparing with other studies. Spatial pattern of methane (CH4) was increasing from landward to seaward where was contradictive with CO2 rate. Meanwhile, NO2 had irregular trend spatially. Emission rate of CH4 and N2O had no significant correlation with environmental parameters. On the other hand, soil’s pH-TOC and water’s TDS-salinity drove a significant effect for CO2 emission.High carbon stock, lower GHGs emission and pristine mangrove vegetation represented that mangrove in south-east Kelong Island has high benefit for climate change effect mitigation. Keywords: decomposition rate, global climate change, greenhouse gases, mangrove,
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Kelautan
VARIASI SPASIAL EMISI GAS RUMAH KACA (GRK)PADA
EKOSISTEM MANGROVE DI PULAU KELONG, BINTAN
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2016
PRAKATA
Puji dan syukur penulis panjatkan kepada Tuhan Yang Maha Esa, Ida Sang Hyang Widhi Wasa, atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema yang dipilih dalam penelitian yang dilaksanakan sejak bulan Maret 2015 ini adalah ekologi laut, dengan judul Variasi Spasial Emisi Gas Rumah Kaca (GRK) pada Ekosistem Mangrove di Pulau Kelong, Bintan.
Terima kasih penulis ucapkan kepada Ibu Dr. Ir. Neviaty Putri Zamani, M.Sc dan Bapak Dr. Hawis H. Madduppa, S.Pi., M.Si. selaku pembimbing, serta Bapak Prof. Dr. Ir. Dedi Soedharma, DEA yang telah banyak memberi saran. Di samping itu, penghargaan penulis sampaikan kepada rekan-rekan peneliti di UPT. Loka Konservasi Biota Laut, Biak – Papua dan Pusat Penelitian Oseanografi, Jakarta, Lembaga Ilmu Pengetahuan Indonesia (LIPI) serta Third Institute of Oceanography, Xiamen – China yang telah banyak memberikan sumbangan pemikiran dan penggunaan fasilitas laboratorium untuk menyelesaikan penelitian ini. Ungkapan terima kasih juga disampaikan kepada ayah, ibu, serta seluruh keluarga, atas segala doa dan kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juni 2016
DAFTAR ISI
DAFTAR TABEL x
DAFTAR GAMBAR x
DAFTAR LAMPIRAN x
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Hipotesis 3
Tujuan Penelitian 3
Manfaat Penelitian 3
Ruang Lingkup Penelitian 3
2 METODE 3
Waktu dan Lokasi Penelitian 3
Pengumpulan Data 4
Analisis Data 6
3 HASIL DAN PEMBAHASAN 7
Hasil Penelitian 7
Pembahasan 12
4 SIMPULAN DAN SARAN 15
Simpulan 15
Saran 16
DAFTAR PUSTAKA 16
LAMPIRAN 20
DAFTAR TABEL
1 Kerapatan pohon dan pancang; jumlah jenis; indeks nilai penting (INP) pohon dan persentase tutupan kanopi dari komunitas mangrove pada tiga zona penelitian di Pulau Kelong.
2 Perbandingan karakteristik antar tiga zona penelitian mangrove di Pulau Kelong.
3 Laju dekomposisi serasah (LD), koefisien dekomposisi (k), dan waktu paruh (t50) dari dekomposisi serasah daun di tiga zonasi mangrove Pulau Kelong. 4 Analisis korelasi Pearson antara emisi GRK dengan kondisi komunitas
mangrove, faktor lingkungan (sedimen dan air) serta laju dekomposisi. 5 Perbandingan aliran emisi GRK (µmol m-2 h-1) dari beberapa penelitian
DAFTAR GAMBAR
1 Diagram alir permasalahan yang ingin dipecahkan dalam penelitian. 2 Sebaran empat lokasi transek pada wilayah mangrove terpilih di Pulau
Kelong. Setiap lokasi transek dibagi menjadi tiga zona, yaitu: L (zona daratan); M (zona tengah) dan S (zona dekat laut).
3 Analisis MDS (multidimensional scaling) terhadap plot berdasarkan kondisi lingkungan in situ (air dan substrat) dan kelimpahan jenis pohon. 4 Berat kering (g) daun yang terdekomposisi dari hari ke-0 sampai hari ke-42. 5 Laju emisi CO2 (a); CH4 (b) dan N2O (c) dalam μmol m−2 h−1 yang diukur pada tiga zona yang berbeda di kawasan mangrove Pulau Kelong bagian tenggara.
DAFTAR LAMPIRAN
1 Publikasi ilmiah skala nasional yang sudah diterbitkan.2 Tulisan ilmiah skala internasional yang sedang dalam proses revisi.
3
4 9 9
11 8 8 10 10 13
1
PENDAHULUAN
Latar Belakang
Dalam satu dekade terakhir, dunia sedang dikhawatirkan dengan isu pemanasan global yang menyebabkan perubahan dan ketidakteraturan faktor iklim. Hal ini yang dipicu oleh peningkatan konsentrasi gas rumah kaca (GRK), yaitu karbon dioksida (CO2), metana (CH4) dan N2O (dinitrogen oksida) dalam atmosfer (IPCC 2001; IPCC 2015). Konsentrasi gas CO2 telah meningkat dari 278 ppm (1750) menjadi 391 ppm (2001) (IPCC 2015). Peningkatan konsentrasi GRK yang paling tinggi terjadi dalam dekade terakhir dan terus meningkat sampai saat ini. Di sisi lain, walaupun gas CH4 dan N2O cukup stabil dan konsentrasinya lebih rendah, namun gas – gas tersebut memiliki efek 298 dan 34 kali lebih tinggi dibandingkan dengan CO2 (Myhre et al. 2013).
Komunitas mangrove memiliki kemampuan dalam menyerap CO2, digunakan dalam proses fotosintesis dan disimpan dalam bentuk biomassa tubuh (Bouillon et al. 2008; Murdiyarso et al. 2015). Indonesia memiliki luasan dan keanekaragaman jenis mengrove paling tinggi di dunia (Sukardjo and Alongi 2012). Mangrove Indonesia juga memiliki cadangan karbon yang sangat signifikan dan berpotensi untuk digunakan dalam mitigasi pemanasan global (Murdiyarso et al. 2015). Geomorfologi dan topografi pantai secara spasial memungkinkan adanya gradien salinitas yang berbeda dari laut ke darat mempengaruhi proses biogeokimia sedimen serta zonasi dalam komunitas mangrove. Hal ini memungkinkan adanya variasi komposisi jenis, variasi faktor biotik dan abiotik yang teratur dari darat ke laut.
Selain mampu menyerap CO2, potensi emisi GRK dari ekosistem mangrove juga tidak bisa dikesampingkan. Setiap ekosistem mangrove merupakan salah satu penghasil GRK sebagai keluaran dari proses respirasi dan dekomposisi (Bouillon et al. 2008; Chen et al. 2010). Beberapa penelitian juga telah menunjukkan bahwa mangrove merupakan sumber dari emisi GRK (Allen et al. 2007; Chen et al. 2010; Chen et al. 2014a; Wang et al. 2016). Tingkat emisi GRK akan semakin meningkat seiring dengan peningkatan masukan nutrien melalui aliran sungai ke dalam ekosistem mangrove (Chen et al. 2010; Chen et al. 2014b). Penurunan luasan mangrove akibat aktivitas manusia sebagai penyimpan karbon juga berdampak pada peningkatan emisi CO2 ke dalam atmosfer (IPCC 2015). Informasi tentang emisi GRK dari ekosistem mangrove di Indonesia sangat jarang. Hanya penelitian Chen et al. (2014b) yang mengungkapkan laju emisi GRK yang rendah dari sedimen mangrove di Sulawesi Utara.
2
gangguan antropogenik. Diameter mangrovenya memiliki rata-rata paling tinggi dibandingkan dengan lokasi lainnya. Khusus wilayah Kelong bagian tenggara, mangrove mendistribusikan 42,08 – 101,03 gr/hari serasah ke laut dan memproduksi serasah 0,55 – 0,80 gr/hari. Estimasi serapan CO2 yang diperoleh dalam kawasan rata-rata sebesar ~28.027,40 gr CO2/ha/hari. Untuk melengkapi penelitian sebelumnya, kajian spasial tentang kondisi emisi GRK dari kawasan sangat diperlukan yang kemungkinan berkaitan dengan laju dekomposisi, parameter biogeokimia dan struktur komunitas dari zona darat ke laut.
Perumusan Masalah
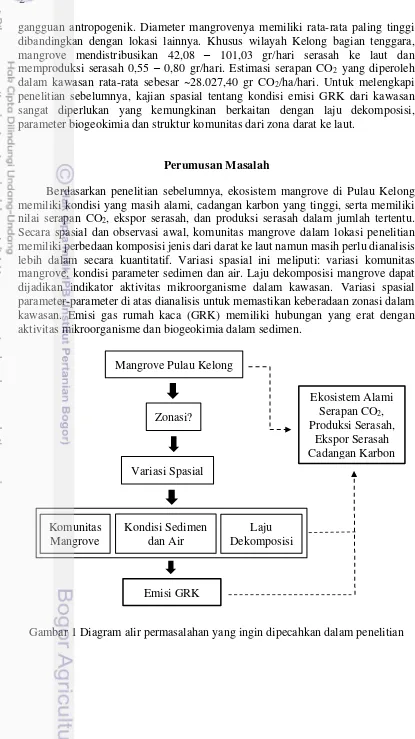
Berdasarkan penelitian sebelumnya, ekosistem mangrove di Pulau Kelong memiliki kondisi yang masih alami, cadangan karbon yang tinggi, serta memiliki nilai serapan CO2, ekspor serasah, dan produksi serasah dalam jumlah tertentu. Secara spasial dan observasi awal, komunitas mangrove dalam lokasi penelitian memiliki perbedaan komposisi jenis dari darat ke laut namun masih perlu dianalisis lebih dalam secara kuantitatif. Variasi spasial ini meliputi: variasi komunitas mangrove, kondisi parameter sedimen dan air. Laju dekomposisi mangrove dapat dijadikan indikator aktivitas mikroorganisme dalam kawasan. Variasi spasial parameter-parameter di atas dianalisis untuk memastikan keberadaan zonasi dalam kawasan. Emisi gas rumah kaca (GRK) memiliki hubungan yang erat dengan aktivitas mikroorganisme dan biogeokimia dalam sedimen.
Gambar 1 Diagram alir permasalahan yang ingin dipecahkan dalam penelitian Mangrove Pulau Kelong
Zonasi?
Emisi GRK Variasi Spasial
Ekosistem Alami Serapan CO2, Produksi Serasah,
Ekspor Serasah Cadangan Karbon
Kondisi Sedimen dan Air Komunitas
Mangrove
3 Hipotesis
Hipotesis dari penelitian ini adalah adanya variasi spasial dari konsidi komunitas mangrove dan variabel lingkungan dalam tiap zona memberikan pengaruh terhadap variasi fluks GRK dari darat ke laut
Tujuan Penelitian
Tujuan dari penelitian ini adalah untuk:
1. Menganalisis kondisi ekosistem mangrove di Pulau Kelong bagian tenggara mencakup kondisi kajian vegetasi, dekomposisi maupun lingkungan pada tiap zona yang berbeda, yaitu zona dekat darat (L), zona tengah (M) dan zona dekat laut (S).
2. Menghitung besaran laju emisi GRK ini dilakukan pada tiga zona tersebut. 3. Menganalisis hubungan kondisi komunitas mangrove, parameter
lingkungan dan laju dekomposisi terhadap emisi GRK yang dihasilkan.
Manfaat Penelitian
Penelitian ini diharapkan mampu memberikan informasi tentang laju emisi GRK secara spasial dari ekosistem mangrove yang masih alami yang memiliki cadangan karbon yang tinggi. Hasil yang diperoleh serta publikasi nasional dan internasional yang dihasilkan juga diharapkan bisa memberikan rekomendasi kepada pemerintah daerah Kabupaten Bintan tentang potensi mangrove dalam kawasan yang memiliki fungsi global dalam mitigasi perubahan iklim yang nyata.
Ruang Lingkup Penelitian
Batasan penelitian ini mencakup pada laju emisi GRK yang dipengaruhi oleh kondisi komunitas mangrove dan lingkungan di Pulau Kelong, bagian tenggara.
METODE
Waktu dan Lokasi Penelitian
4
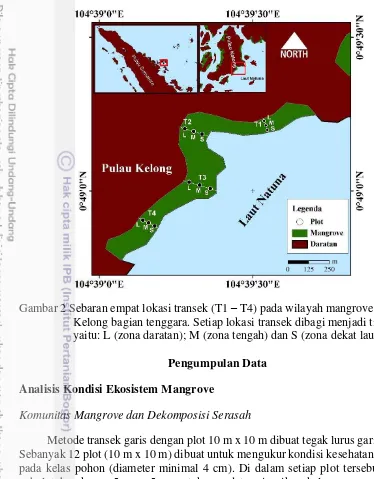
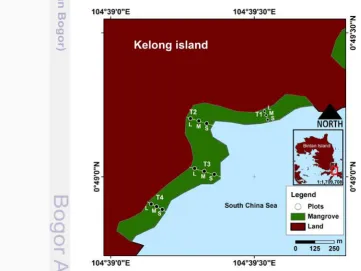
Gambar 2 Sebaran empat lokasi transek (T1 – T4) pada wilayah mangrove di Pulau Kelong bagian tenggara. Setiap lokasi transek dibagi menjadi tiga zona, yaitu: L (zona daratan); M (zona tengah) dan S (zona dekat laut).
Pengumpulan Data
Analisis Kondisi Ekosistem Mangrove
Komunitas Mangrove dan Dekomposisi Serasah
Metode transek garis dengan plot 10 m x 10 m dibuat tegak lurus garis pantai. Sebanyak 12 plot (10 m x 10 m) dibuat untuk mengukur kondisi kesehatan vegetasi pada kelas pohon (diameter minimal 4 cm). Di dalam setiap plot tersebut dibuat subplot berukuran 5 m x 5 m untuk mendeterminasikan kelas pancang/sapling (diameter lebih kecil dari 4 cm). Klasifikasi ini mengikuti Ashton and Macintosh (2002). Diukur dan dicatat keliling batang pada posisi dada 130 cm dan diidentifikasi jenisnya berdasarkan Giesen et al. (2006). Kerapatan serta indeks nilai penting jenis pada pohon dan sapling dihitung. Persentase tutupan kanopi mangrove didekati dengan metode hemispherical photography (Dharmawan and Pramudji 2014). Litter standing stock (stok serasah daun di permukaan tanah) pada setiap plot dihitung dalam plot kecil berukuran 1m x 1m. Serasah daun di oven pada suhu 80oC selama 1 minggu kemudian ditimbang (Robertson and Daniel 1989). Parameter Lingkungan
5 YSI EC200. Sampel tanah dikoleksi dari setiap plot pada kedalaman 5 cm dan disaring dalam mesh size 2 mm. pH tanah diukur pada perbandingan 1:5 antara tanah dan aquadesh menggunakan Hanna pH meter. Analisis Total Organik Karbon (TOC), Total Kjeldahl Nitrogen (TN) dan Total Fosfat (TP) dilakukan mengikuti Chen et al. (2014b). Analisis TOC dilakukan dengan metode loss on ignition (LOI) pada suhu 550 oC. Selanjutnya, TN dan TP dianalisis dengan metode flow injection analyzer (FIA).
Penghitungan Laju Dekomposisi
Laju dekomposisi dihitung berdasarkan metode termodifikasi dari Ashton et al. (1999) dan Dewiyanti (2010). Daun kuning yang masih segar pada pohon mangrove dikoleksi dari jenis mangrove yang mendominasi di setiap zona. Daun dibersihkan, didiamkan 24 jam, ditimbang sebanyak 10 gr (W1) dan dimasukkan ke dalam kantong jaring yang berukuran 10 cm x 10 cm dan lubang-lubang jaring (mesh) berukuran 1 mm2. Kantong daun diletakkan di atas permukaan sedimen mangrove dan diikatkan pada akar yang sejajar tanah untuk mencegah kantong terbawa arus pasang surut. Setiap zona dibuat empat kali ulangan, dan enam kali perlakuan waktu (hari ke-: 2, 4, 7, 14, 28, 42). Kantong dikoleksi pada hari yang sudah ditentukan dan dibawa ke laboratorium. Daun dibersihkan dan dimasukkan ke dalam oven suhu 60oC sampai berat konstan. Setiap sampel ditimbang dan dicatat berat keringnya (W2).
Faktor konversi (f) berat basah ke berat kering dihitung sebagai persentase kandungan biomassa dalam daun kuning. Koleksi daun kuning ditimbang berat basah (BB) kemudian dioven pada suhu 60oC sampai tidak ada perubahan berat. Daun kering ditimbang untuk mendapatkan berat kering (BK). Nilai f = ((BB –
BK)/BB *100%).
Pengukuran Emisi Gas Rumah Kaca (GRK)
Pengambilan sampel gas dilakukan dengan metode chamber tertutup pada waktu surut dengan durasi pengambilan 0, 10, 20 dan 30 menit (Chen et al. 2010; Chen et al. 2014b). Chamber gas terbuat dari aclyric berbentuk kubus terbuka pada satu sisi berukuran 20 cm x 20 cm x 25 cm. Dibuat dua katup berdiameter ± 0,5 cm pada bagian samping dan atas. Pengambilan sampel gas diawali dengan chamber gas diletakkan telungkup dengan kedua katup masih terbuka dan kemudian dibenamkan ± 2 cm. Selanjutnya, kedua katup ditutup untuk dimulai pengambilan sampel gas waktu ke-0. Gas dikoleksi dengan menggunakan syringe 10 ml secara hati-hati melalui katup atas, sedangkan katup samping dimasukan jarum syringe terbuka yang digunakan sebagai penyeimbang tekanan gas. Sampel gas dimasukkan ke dalam kantong gas aluminium foil dan dibawa ke laboratorium. Jarum syringe pada katup samping ditarik ketika pengambilan sampel selesai dilakukan setiap waktu durasi. Setiap zona dilakukan empat kali ulangan.
6
Setiap sampel diambil 1 ml kemudian diinjeksikan ke dalam GC. Konsentrasi gas metana dihitung dengan membandingkan luas peak/puncak sampel dengan kurva standar.
Analisis Data
Laju dekomposisi serasah (LD) dalam selang waktu tertentu (Δt), persentase
terdekomposisi (%D), koefisien laju dekomposisi (k) serta waktu yang dibutuhkan untuk mendekomposisi setengah sampel (t50) dihitung dengan menggunakan persamaan 1 – 4. Sedangkan analisis aliran emisi GRK (Fm) dilakukan mengikuti persamaan 5.
LD = [(W1 x f) – W2]/Δt …(1) %D = (W1– W2)/W1x 100% …(2)
K = ln(Xt/Xo)/Δt …(3)
t50 = ln 2/K …(4)
dimana:
LD = Laju dekomposisi (g.m-2.hari-1) W1 = berat basah daun kuning (gr)
W2 = berat kering daun setelah terdekomposisi dalam waktu tertentu (gr) f = faktor konversi berat basah – berat kering (%)
Δt = selang waktu pengamatan (hari) %D = persentase terdekomposisi k = koefisien dekomposisi
t50 = waktu yang dibutuhkan untuk mendekomposisi setengah sampel (hari)
Xt = persentase serasah daun setelah terdekomposisi dalam kurun waktu tertentu (t)
Xo = persentase serasah daun sebelum terdekomposisi. Fm = (V.ΔM/A.ρ).10-6 …(5) dimana :
Fm : aliran GRK (µmol m-2.h-1) V : volume chamber (m3)
ΔM : perubahan konsentrasi gas di dalam chamber (h-1). A : luas permukaan tanah (m2)
Ρ : konstanta volume setiap mol gas (m3.mol-1)
7 similaritas seluruh parameter digunakan untuk menentukan ordinasi setiap plot pengukuran dan menegaskan klasifikasi zonasi dengan menggunakan perangkat lunak PRIMER 6. Hubungan tiap parameter dengan laju emisi GRK dianalisis dalam korelasi Pearson.
HASIL DAN PEMBAHASAN
Hasil Penelitian
Ekosistem Mangrove Pulau Kelong Bagian Tenggara
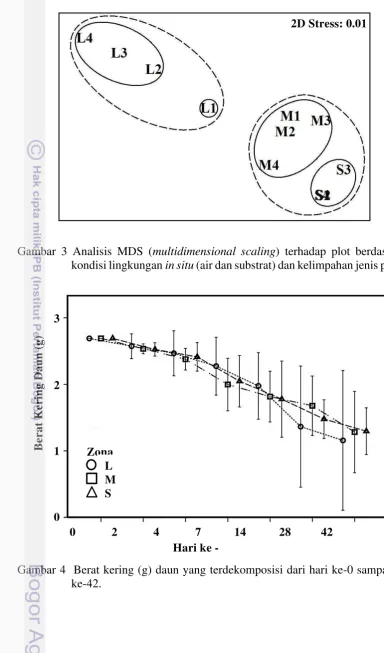
Komunitas mangrove dalam kawasan memiliki ketebalan maksimal 200 meter yang merupakan hutan primer. Kerapatan pohon pada lokasi penelitian tertinggi ditemukan di zona darat (L) sebesar 2.175 ± 222 pohon/ha yang berbeda nyata (P<0,05) dengan dua zona lainnya dimana zona dekat laut (S) memiliki kerapatan pohon yang paling rendah. Namun, pada kelas sapling (komunitas tegakan dengan diameter batang< 4 cm), zona S memiliki tegakan yang paling rapat dan berbeda nyata dibandingkan dengan dua stasiun lainya (Tabel 1). Dari segi keanekaragaman jenis, zona dekat laut hanya terdiri dari dua jenis dan paling rendah dibandingkan zona darat dan zona tengah. Kondisi kesehatan komunitas mangrove di dalam kawasan termasuk dalam kategori baik dengan rentang persentase tutupan tajuk, yaitu: 87,96 ± 6,88% (L) sampai 91,31 ± 1,14% (M) yang tidak berbeda nyata antar zona.
Hasil analisis parameter sedimen menunjukkan bahwa zona laut memiliki kecenderungan kesamaan dengan zona tengah dan berbeda dengan zona dekat darat (Tabel 2). Nilai pH sedimen secara spasial meningkat dari darat ke laut dengan kisaran 6,95±0,06 sampai 7,40±0,08. Tren tersebut juga diperoleh pada hasil analisis suhu (~28,12 - ~28,83 oC). Kecenderungan nilai TOC dan total N ditemukan berlawanan dengan suhu dan pH sedimen secara spasial. Sementara itu, kandungan P tidak memiliki pola yang teratur dari darat ke laut, yaitu: 0,21±0,05; 0,30±0,003 dan 0,27±0,03 mg/L.
8
Tabel 1 Kerapatan pohon dan pancang; jumlah jenis; indeks nilai penting (INP) pohon dan persentase tutupan kanopi dari komunitas mangrove pada tiga zona penelitian di Pulau Kelong.
NO PARAMETER ZONASI
L M S
1 Kerapatan Pohon (tree ha-1) 2175 ± 222a 1800 ± 141b 1550 ± 191b
Rhizophora apiculata 425 ± 544 1425 ± 126 1475 ± 171
Brugueira gymnorrhiza n/a 100 ± 82 n/a
Xylocarpus granatum 1750 ± 332 200 ± 82 n/a
X. mollucensis n/a 75 ± 50 75 ± 50
2 Kerapatan Pancang (id.ha-1) 975 ± 287a 925 ± 150a 1875 ± 50b
R. apiculata 325 ± 330 850 ± 191 1875 ± 50
B. gymnorrhiza n/a 200 ± 115 n/a
X. granatum 525 ± 171 75 ± 96 n/a
Aegiceras corniculatum 125 ± 96 n/a n/a
3 Jumlah Jenis 3 4 2
4 Indeks Nilai Penting (INP) Pohon*)
Tertinggi XG: 232,42% RA: 177,32% RA: 237,53%
Terendah RA: 67,58% XM : 30,60% XM: 62,47%
5 % tutupan kanopi 87,96 ± 6,88a 91,31 ± 1,14a 90,48 ± 0,54a
6 Stok serasah (gr DW/m2)
78,23 ± 14,16 81,61 ± 11,25 82,09 ± 19,37
*) XG = X. granatum; RA = R. apiculata; XM = X. mollucensis.
n/a = tidak ditemukan
ab menunjukkan hasil uji Tukey dimana huruf yang sama pada baris yang sama tidak memiliki
perbedaan nilai secara nyata.
Tabel 2 Perbandingan karakteristik sedimen dan air antar tiga zona penelitian mangrove di Pulau Kelong.
NO Parameter Dekat Darat (L) Tengah (M) Zona Dekat Laut (S)
1 Deskripsi Sedimen Berlumpur Berlumpur Pasir berlumpur 2 Dominansi jenis Tinggi
X. granatum
R. apiculata
dengan campuran jenis lainnya
Tinggi R. apiculata
3 pH tanah 6,95 ± 0,06a 7,08 ± 0,10a 7,40 ± 0,08b 4 Suhu tanah (oC) 28,13 ± 0,10a 28,75 ± 0,19b 28,83 ± 0,10b 5 TOC tanah (mg/L) 125,41 ± 1,10a 79,76 ± 1,18b 71,48 ± 17,28b 6 TP tanah (mg/L) 0,21 ± 0,05a 0,30 ± 0,003b 0,27 ± 0,03b 7 TN tanah (mg/L) 3,23 ± 0,31a 2,56 ± 0,16b 2,32 ± 0,41b 8 pH air 6,93 0,10a 7,10 0,08a 7,65 ± 0,13b
9 Suhu air (oC) 28,35 ± 0,06a 28,80 ± 0,14b 29,38 ± 0,26c 10 Salinitas air (ppt) 24,60 ± 0,68a 23,03 ± 2,48a 32,15 ± 0,13b 11 TDS air (g/L) 12,04 ± 0,33a 11,52 ± 1,08a 15,26 ± 0,04b
ab menunjukkan hasil uji Tukey dimana huruf yang sama pada baris yang sama tidak memiliki
9
Gambar 3 Analisis MDS (multidimensional scaling) terhadap plot berdasarkan kondisi lingkungan in situ (air dan substrat) dan kelimpahan jenis pohon.
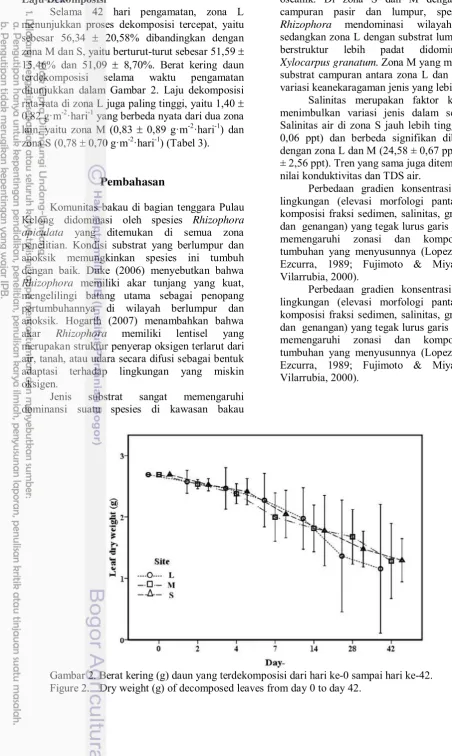
Gambar 4 Berat kering (g) daun yang terdekomposisi dari hari ke-0 sampai hari ke-42.
Berat K
er
in
g Daun
(g)
1 3
2
0
0 2 4 7 14 28 42 Hari ke -
Zona L M S
10
Dalam 42 hari pengamatan, L merupakan zona dengan proses dekomposisi yang paling tinggi (56,34 20,58%) dibandingkan dengan M dan S yang berturut-turut sebesar 51,59 15,46% dan 51,09 8,70% . Berat kering daun terdekomposisi selama waktu pengamatan ditunjukkan pada Gambar 4. Laju dekomposisi pada ekosistem mangrove pada zona darat paling tinggi dengan rata-rata 1,40 0,82 gr/m2/hari yang berbeda nyata dengan dua zona lainnya. Pada zona M dan S, laju dekomposisi rata-rata 0,83 0,89 gr/m2/hari dan 0,78 ± 0,70 gr/m2/hari secara berurutan (Tabel 3).
Tabel 3 Laju dekomposisi serasah (LD), koefisien dekomposisi (k), dan waktu paruh (t50) dari dekomposisi serasah daun di tiga zonasi mangrove Pulau Kelong.
Zona LD (g/m2/d) k (d-1) t50 (hari)
L 1,40 ± 0,82a 0,022± 0,012 45
M 0,83 ± 0,89b 0,027± 0,029 43
S 0,78 ± 0,70b 0,026± 0,024 44
Total 2,57 ± 2,20 0,025± 0,023 44
ab menunjukkan hasil uji Tukey dimana huruf yang sama pada kolom yang sama tidak
memiliki perbedaan nilai secara nyata.
Laju Emisi Gas Rumah Kaca (GRK)
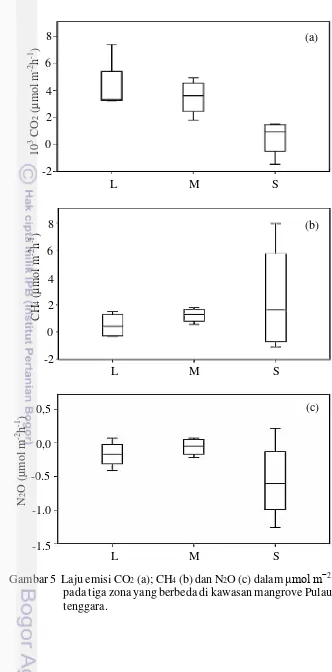
Secara umum, ekosistem mangrove di Pulau Kelong bagian tenggara memiliki produksi GRK yang rendah. Secara spasial, flux gas CH4 dan N2O tidak memiliki perbedaan yang signifikan antar zona. Walaupun demikian, gas CH4 bervariasi dari darat ke laut 0,52 0,92 - 2,54 4,17 μmol m−2 h−1, dan emisi gas N2O tidak memiliki pola yang jelas dan bernilai negatif. Sementara itu, emisi gas CO2 pada zona dekat laut berbeda signifikan dengan emisi yang terjadi pada zona darat (P < 0,05). Pola emisi CO2 berlawanan dengan pola emisi CH4 dimana meningkat dari laut ke darat 484,01 1.384,61 – 4.351,82 2.040,88 μmol m−2 h−1 (Gambar 5).
Hubungan Kondisi Ekosistem dengan Emisi GRK
11
Gambar 5 Laju emisi CO2 (a); CH4 (b) dan N2O (c) dalam μmol m−2 h−1 yang diukur pada tiga zona yang berbeda di kawasan mangrove Pulau Kelong bagian tenggara.
(a) 8
6 4 2 0 -2
L M S
10
3 CO
2
(
µ
m
ol
m
-2 h -1 )
C
H4
(
µ
m
ol m
-2 h -1 )
L M S
8 6 4 2 0 -2
(b)
0,5 0,0 -0.5 -1.0
-1.5
N2
O (µ
m
ol m
-2 h -1 )
L M S
12
Tabel 4 Analisis korelasi Pearson antara emisi GRK dengan kondisi komunitas mangrove, faktor lingkungan (sedimen dan air) serta laju dekomposisi.
Parameter CO2 CH4 N2O
Air Sal. -0,686* 0,262 -0,489
TDS -0643* 0,266 -0,454
Sedimen pH -0,864** 0,509 -0,412
Temp -0,483 0,373 -0,162
TOC 0,735* 0,244 0,476
TN 0,566 0,497 0,488
TP -0,633 0,372 -0,024
Komunitas Mangrove TA 0,464 -0,094 0,498
* : korelasi nyata pada α = 0,05 ** : korelasi nyata pada α = 0,01
Pembahasan
Komunitas mangrove di kawasan tenggara Pulau Kelong didominasi oleh Rhizophora apiculata karena jenis ini ditemukan di semua zona penelitian. Kondisi substrat yang berlumpur dan anoksik memungkinkan jenis ini tumbuh dengan baik dalam kawasan. Duke (2006) menyebutkan bahwa Rhizophora memiliki akar khusus dan kuat, berupa akar tunjang yang mengelilingi batang utama sebagai penopang pertumbuhannya di wilayah berlumpur dan anoksik. Hogarth (2007) menambahkan bahwa akar Rhizophora memiliki lentisel yang merupakan struktur penyerap oksigen terlarut dari air atau tanah serta dari udara secara difusi sebagai bentuk adaptasi terhadap lingkungan yang miskin oksigen.
Jenis substrat sangat mempengaruhi dominansi suatu spesies pada kawasan mangrove oseanik. Pada zona S dan M dengan sedimen campuran pasir dan lumpur, jenis Rhizophora mendominasi wilayah tersebut. Sedangkan pada wilayah L yang didominasi oleh substrat lumpuran yang berstruktur lebih padat, didominasi oleh Xylocarpus granatum. Zona M, yang memiliki tipe substrat campuran antara zona L dan S, memiliki variasi keanekaragaman jenis yang lebih beragam. Salinitas merupakan faktor kunci yang menimbulkan variasi jenis dalam setiap zona. Salinitas air pada zona laut jauh lebih tinggi (32,05 ± 0,06 ppt) dan berbeda signifikan dibandingkan dengan zona L dan M (24,58 ± 0,67 dan 24,13 ± 2,56 ppt). Tren tersebut juga ditemukan pada hasil pengukuran nilai TDS air.
Zonasi pada mangrove disebabkan oleh perbedaan gradien konsentrasi faktor lingkungan secara tegak lurus garis pantai yang akan mempengaruhi komposisi jenis tumbuhan yang menyusunnya. Faktor lingkungan tersebut antara lain: elevasi morfologi pantai, gradien komposisi fraksi sedimen, salinitas, gradient hara serta genangan (Fujimoto and Miyagi 1993; Lopez-Portillo and Ezcurra 1989; Vilarrúbia 2000). Tidak ada pola pasti dari zonasi mangrove, namun secara umum jenis yang tahan terhadap salinitas tinggi seperti Avicennia dan Sonneratia akan menempati zona pantai. Pada penelitian ini, Rhizophora apiculata ditemukan dominan pada zona S yang memiliki salinitas tinggi (32,05 ppt). Jenis ini memiliki kemampuan adaptasi pada rentang salinitas yang sangat lebar sehingga ditemukan di seluruh zona penelitian. Ada tiga mekanisme yang dimiliki oleh Rhizophora untuk beradaptasi dengan kadar garam yang tinggi, yaitu: akar ultra filtrasi, jaringan penyimpan air dan mengalirkan garam ke daun tua (Chapman 1976). Selain
13 mekanisme adaptasi tersebut, kondisi substrat yang berlumpur juga merupakan tempat hidup yang sangat baik untuk jenis R. apiculata.
Laju dekomposisi serasah yang diperoleh dari kombinasi biomassa yang hilang dan litter standing stock dalam setiap zona menunjukkan nilai rata-rata dengan rentang 0,78 ± 0,70(S) – 1,40 ± 0,82 (L) gram/m2/hari. Pada zona S dan M yang didominasi oleh R. apiculata memiliki laju dekomposisi yang lebih rendah dibandingkan dengan kawasan LWD yang didominasi oleh jenis X. granatum. Nilai laju dekomposisi berdasarkan hasil perhitungan t50 rata-rata diperoleh pada 44 hari. Penelitian Ashton et al. (1999) menunjukkan pada kawasan mangrove yang masih alami, nilai t50 berkisar 43 hari sedangkan pada kawasan terbuka nilai t50 lebih besar, yaitu 76 hari. Dewiyanti (2010) juga memperoleh nilai t50 di zona dekat laut (63 hari) lebih rendah dibandingkan zona darat (67 hari). Dalam Tabel 4, laju dekomposisi ditunjukkan memiliki hubungan yang signifikan dengan suhu substrat dan salinitas air. Graca et al. (2005) menjelaskan bahwa peluruhan dari serasah daun didefinisikan sebagai kehilangan berat akibat beberapa proses fisio-kimia yang disebabkan oleh kondisi lingkungan.
Berdasarkan nilai k yang diperoleh dengan rata-rata 0,025 ± 0,023 per hari, seluruh kawasan memiliki laju dekomposisi yang tergolong tinggi. Penelitian Ananda et al. (2008) menggolongkan laju dekomposisi menjadi tiga kategori berdasarkan nilai k, yaitu tinggi (k>0,01); sedang (k = 0,005 – 0,01); rendah (k<0,005). Penelitian Dewiyanti (2010) pada kawasan hutan mangrove Pulau Weh, Sabang telah diperoleh koefisien dekomposisi yang lebih rendah, yaitu 0,010 ± 0,000577 per hari. Hasil penelitian Ashton et al. (1999) menghitung nilai k sebesar 0,00163 per hari pada kawasan hutan mangrove alami yang didominasi oleh jenis R. apiculata di hutan mangrove Peninsular, Malaysia.
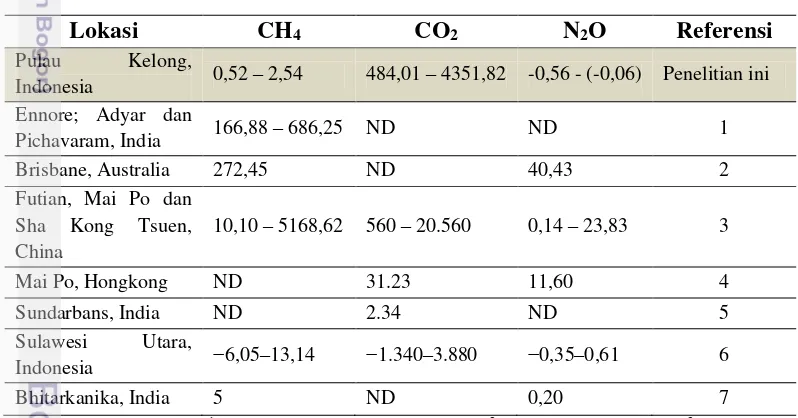
Tabel 5 Perbandingan aliran emisi GRK (µmol m-2 h-1) dari beberapa penelitian
Lokasi CH4 CO2 N2O Referensi
Pulau Kelong,
Indonesia 0,52 – 2,54 484,01 – 4351,82 -0,56 - (-0,06) Penelitian ini Ennore; Adyar dan
Pichavaram, India 166,88 – 686,25 ND ND 1 Brisbane, Australia 272,45 ND 40,43 2 Futian, Mai Po dan
Sha Kong Tsuen, China
10,10 – 5168,62 560 – 20.560 0,14 – 23,83 3
Mai Po, Hongkong ND 31.23 11,60 4 Sundarbans, India ND 2.34 ND 5 Sulawesi Utara,
Indonesia −6,05–13,14 −1.340–3.880 −0,35–0,61 6 Bhitarkanika, India 5 ND 0,20 7
14
Emisi gas CO2 dalam kawasan lebih tinggi dibandingkan dengan penelitian sebelumnya (Alongi et al. 2001; Chen et al. 2012; 2014b). Rata-rata flux CO2 dalam penelitian ini (2.778,61 µmol m−2 h−1) lebih rendah dibandingkan rata-rata flux CO
2 di 75 lokasi di dunia, yaitu 2.870 µmol m−2 h−1 (Alongi 2009). Selain itu, emisi dalam kawasan juga jauh lebih rendah dibandingkan dengan emisi di Australia (Lovelock 2008). Namun, variasi spasialnya menurun dari darat ke laut yang berlawanan dengan penelitian Chen et al. (2010).
Rata-rata aliran gas CH4 yang diperoleh (0,52 – 2,54 µmol m-2 h-1) lebih rendah dibandingkan dengan sebagian besar penelitian lainnya yang mengunakan metode closed-chamber (Tabel 5). Namun, nilainya lebih tinggi dibandingkan dengan emisi CH4 di Sai Keng, Hongkong; North Sulawesi, Indonesia and Sundarbans, India. Emisi CH4 yang tinggi di China Selatan dipicu oleh percobaan dengan menggunakan air limbah (Chen et al. 2010). Zona laut memiliki kemiripan emisi gas dengan emisi yang terjadi di persawahan (Lumbanraja et al. 1998). Flux gas metana pada zona S lebih tinggi dibandingkan dengan zona lainnya, sebuah pola yang mirip dengan penelitian Lu et al. (1998). Emisi gas metana yang lebih tinggi akan ditemukan pada kawasan estuari atau lagoon yang memiliki sedimen lumpur. Allen et al. (2007) menjelaskan bahwa sedimen berlumpur pada mangrove dalam aliran sungai memproduksi gas metana yang tinggi mencapai 1.087,50 µmol m-2 h-1.
Gas N2O merupakan produk sampingan yang dihasilkan dalam siklus nitrogen dalam sedimen yang melibatkan aktivitas mikroorganisme kompleks. Denitrifikasi, sebagai salah satu bagian dari siklus tersebut, dianggap sebagai tahapan yang penting dalam produksi N2O (Fernandes and Bharathi 2010). Dalam penelitian ini, nilai negatif pada emisi gas N2O menunjukkan bahwa ekosistem mangrove dalam kawasan memiliki kemampuan dalam menyerap N2O. Jika dibandingkan dengan hasil pengukuran di lokasi lain, rata-rata aliran emisi gas N2O (-0,26 µmol m-2 h-1) merupakan yang paling rendah dibandingkan dengan negara lain, seperti; India : 0,20 µmol m-2 h-1 (Chauhan et al. 2015), Hongkong : 11,60 µmol m-2 h-1 (Chen et al. 2012), and Australia : 40,43 µmol m-2 h-1 (Allen et al. 2007). Laju emisi gas N2O di kawasan hutan mangrove lainnya Indonesia, juga ditemukan lebih tinggi dibandingkan dengan penelitian ini (Chen et al. 2014b). Fluks GRK yang rendah mengindikasikan tingginya potensi mangrove di Pulau Kelong dalam mitigasi perubahan iklim ((Dharmawan et al. 2016b)
Reddy and Crohn (2014) menemukan bahwa salinitas memberikan pengaruh pada emisi CO2. Peningkatan salinitas menurunkan emisi gas CO2 sebesar 28-38%. Hal ini disebabkan oleh sensitivitas dari respirasi tanah terhadap peningkatan salinitas. Pola yang sama juga dapat dilihat pada pengaruh TDS pada air. Nilai TDS yang lebih tinggi, mengindikasikan aerasi antara partikel tanah lebih rendah. Hal ini menyebabkan tanah konsentrasi oksigen dalam tanah sebagai sumber respirasi mikroorganisme menjadi rendah.
15 surut. Dekomposisi dan respirasi yang terjadi dalam tanah bertanggung jawab dalam pemecahan karbon organik menjadi CO2. pH tanah mempengaruhi kandungan karbon dalam sedimen. Nursin et al. (2014) menunjukkan bahwa kandungan organik dalam sedimen akan menurun seiring dengan peningkatan pH dimana menurun dari 2,55% pada pH 6,1 menjadi 0,75% pada pH 7,2.
Berdasarkan analisis statistik, tidak ditemukan hubungan yang nyata antara parameter lingkungan dengan aliran emisi CH4 dan NO2. Namun, berdasarkan Poffenbarger et al. (2011), salinitas dan CH4 memiliki hubungan yang bernilai negatif dan signifikan pada ekosistem pesisir. Namun, analisis korelasi dalam penelitian ini memperlihatkan kondisi yang berlawanan. Couwenberg (2009) menjelaskan bahwa gas metana dengan mudah dihasilkan pada kondisi tanah anaerobik dan tinggi kandungan organik. Whalen (2005) juga menggambarkan bahwa suhu lingkungan juga mempengaruhi mekanisme produksi CH4 dalam sedimen. Lu et al. (1999) menambahkan parameter kelembaban substrat juga merupakan salah satu faktor penting dalam mengatur emisi gas CH4 ke atmosfer.
Analisis hubungan laju emisi gas N2O dengan faktor lingkungan memperlihatkan pH tanah yang memiliki hubungan yang paling kuat. Hal ini disebabkan oleh pH tanah mampu mengatur mekanisme denitrifikasi dan mempengaruhi produksi gas N2O. Penelitian Fernandes and Bharathi (2010) menunjukkan hubungan negatif antara denitrifikasi dengan pH tanah. Nilai emisi gas N2O yang tidak berbeda nyata antar zona menunjukkan bahwa tingkatan proses denitrifikasi yang sama dalam lokasi penelitian. Selain pH, salinitas juga memberikan pengaruh terhadap N2O dan proses denitrifikasi (Marton et al. 2012). Rendahnya nilai TOC dan TN pada zona dekat laut kemungkinan menyebabkan laju denitrifikasi yang rendah dan diikuti dengan rendahnya laju emisi N2O.
SIMPULAN DAN SARAN
Simpulan
Berdasarkan hasil penelitian yang telah diperoleh, maka dapat disimpulkan bahwa:
1. Komunitas mangrove di Pulau Kelong bagian tenggara memiliki zonasi yang cukup jelas dengan komposisi jenis, parameter lingkungan dan laju dekomposisi yang berbeda antar ketiga zona. Zona laut cenderung memiliki kemiripan dengan zona tengah lebih tinggi dibandingkan dengan zona darat. 2. Variasi spasial fluks CO2 meningkat dari darat ke laut dan berlawanan
dengan emisi gas CH4. Sementara itu, flux N2O bernilai negatif dan memiliki pola kecenderungan yang tidak teratur dari darat ke laut.
16
Saran
Kondisi ekosistem dan emisi GRK yang bervariasi secara spasial mengindikasikan perlunya dilakukan penelitian lebih mendalam tentang proses biogeokimia dan mikrobiologi dalam setiap zona. Estimasi tentang potensi kawasan dalam menyerap dan menghasilkan GRK dalam satuan luas juga menarik untuk dikaji lebih mendalam. Hasil penelitian ini dan beberapa penelitian sebelumnya dalam kawasan, menunjukkan bahwa kawasan memiliki potensi tinggi dalam mitigasi perubahan iklim, sehingga disarankan untuk mempertahankan kondisi ekosistem dengan memberikan perlakuan khusus terhapan kawasan. Kebijakan bisa diterapkan misalnya dengan menetapkan kawasan sebagai kawasan konservasi mangrove untuk mempertahankan fungsinya dalam mitigasi perubahan iklim.
DAFTAR PUSTAKA
Allen DE, Dalal RC, Rennenberg H, Meyer RL, Reeves S, Schmidt S. 2007. Spatial and temporal variation of nitrous oxide and methane flux between subtropical mangrove sediments and the atmosphere. Soil Biol Biochem. 39:622-631.
Alongi D. 2009. The Energetics of Mangrove Forests. Springer Netherlands, Amsterdam. doi:10.1007/978-1-4020-4271-3
Alongi DM, Wattayakorn G, Pfitzner J, Tirendi F, Zagorskis I, Brunskill GJ, Davidson A, Clough BF. 2001. Organic carbon accumulation and metabolic pathways in sediments of mangrove forests in southern Thailand. Mar Geol. 179:85-103.
Ananda K, Sridhar KR, Raviraja NS, Baerlocher F. 2008. Breakdown of fresh and dried Rhizophora mucronata leaves in a mangrove of Southwest India. Wetlands Ecol Manage. 112:73-81.
Ashton E, Hogarth, PJ , Ormond R. 1999. Breakdown of mangrove leaf litter in a managed mangrove forest in Peninsular Malaysia. Hydrobiologia. 413:77-88.
Ashton EC, Macintosh DJ. 2002. Preliminary assessment of the plant diversity and community ecology of the Sematan mangrove forest, Sarawak, Malaysia. Forest Ecol Manag. 166:111-129.
Bouillon S, Borges AV, Castañeda-Moya E, Diele K, Dittmar T, Duke NC, Kristensen E, Lee SY, Marchand C, Middelburg JJ et al. 2008. Mangrove production and carbon sinks: a revision of global budget estimates. Global Biochem Cy. 22:n/a-n/a.
Chanda A, Akhand A, Manna S, Dutta S, Das I, Hazra S, Rao KH, Dadhwal VK. 2013. Measuring daytime CO2 fluxes from the inter-tidal mangrove soils of Indian Sundarbans. Environ Earth Sci. 72:417-427.
17 Chauhan R, Datta A, Ramanathan AL, Adhya TK. 2015. Factors influencing
spatio-temporal variation of methane and nitrous oxide emission from a tropical mangrove of eastern coast of India. Atmos Environ. 107:95-106.
Chen GC, Tam NF, Ye Y. 2012. Spatial and seasonal variations of atmospheric N2O and CO2 fluxes from a subtropical mangrove swamp and their relationships with soil characteristics. Soil Biol Biochem. 48:175-181. Chen GC, Tam NFY, Ye Y. 2010. Summer fluxes of atmospheric greenhouse gases
N2O, CH4 and CO2 from mangrove soil in South China. Sci Total Environ. 408:2761-2767.
Chen GC, Tam NFY, Ye Y. 2014a. Does zinc in livestock wastewater reduce nitrous oxide (N2O) emissions from mangrove soils? Water Res. 65:402-413.
Chen GC, Ulumuddin YI, Pramudji S, Chen SY, Chen B, Ye Y, Ou DY, Ma ZY, Huang H, Wang JK. 2014b. Rich soil carbon and nitrogen but low atmospheric greenhouse gas fluxes from North Sulawesi mangrove swamps in Indonesia. Sci Total Environ. 487:91-96.
Couwenberg J. 2009. Methane emissions from peat soils (organic soils, histosols): facts, MRV-ability, emission factors. Wetlands International, Bonn
Dewiyanti I. 2010. Litter decomposition of Rhizophora stylosa in Sabang-Weh Island, Aceh, Indonesia; evidence from mass loss and nutrients. Biodiversitas. 11:139-144.
Dharmawan I, Zamani N, Madduppa H. 2016a. Laju dekomposisi serasah daun di ekosistem bakau Pulau Kelong, Kabupaten Bintan. Oseanol Limnol Indones. 1:1-10.
Dharmawan I, Zamani N, Madduppa H. 2016b. Spatial variation of greenhouse gases fluxes from mangrove soil in Kelong Island, Indonesia: A stepping-stone on recent mangrove conservation perspectives. Journal of Coastal Conservation. in Prep
Dharmawan IWE. 2014. Carbon cycle in Kelong Island. Research Center for Oceanography, Jakarta
Dharmawan IWE, Pramudji S. 2014. Panduan Monitoring Status Kesehatan Komunitas Mangrove. Pusat Penelitian Oseanografi, LIPI, Jakarta
Dharmawan IWE, Ulumudin YI, Afdal. 2014. Estimation of mangrove carbon stock in Easten Coast of Bintan Regency, Riau Islands. Oseanol Limnol Indones. 40:15.
Duke NC. 2006. Rhizophora apiculata, R. mucronata, R. stylosa, R. × annamalai, R. × lamarckii (Indo–West Pacific stilt mangrove). http://www.agroforestry.net/images/pdfs/Rhizophora-IWP.pdf. Accessed 12/01 2015
Fernandes SO, Bharathi PAL. 2010. Denitrification: an important pathway for nitrous oxide production in tropical mangrove sediments (Goa, India). J Environ Qual. 39:1507-1516.
Fujimoto K, Miyagi T. 1993. Development process of tidal-flat type mangrove habitats and their zonation in the Pacific Ocean: a geomorphological study. Vegetatio. 106:137-146.
18
Graca MAS, Baerlocher F, Gessner MO. 2005. A Practical Guide; Methods to Study Litter Decomposition. Springer Netherlands,
Hogarth PJ. 2007. The biology of mangroves and seagrasses. Oxford University Press,
[IPCC] Intergovernmental Panel on Climate Change. 2001. Climate Change 2001: Synthesis Report, Summary for Policymakers: An Assessment of the Intergovernmental Panel on Climate Change. Intergovernmental Panel on Climate Change, Wembley
[IPCC] Intergovernmental Panel on Climate Change. 2015. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Intergovernmental Panel on Climate Change, Geneva
Lopez-Portillo J, Ezcurra E. 1989. Zonation in Mangrove and Salt Marsh Vegetation at Laguna de Mecoacan, Mexico. Biotropica. 21:107-114. Lovelock CE. 2008. Soil Respiration and Belowground Carbon Allocation in
Mangrove Forests. Ecosystems. 11:342-354.
Lu C-y, Wong Y-s, Tam NFY, Ye Y, Cui S-H, Lin P. 1998. Preliminary studies on methane fluxes in Hainan mangrove communities. Chin J Oceanol Limnol. 16:64-71.
Lu CY, Wong YS, Tam NFY, Ye Y, Lin P. 1999. Methane flux and production from sediments of a mangrove wetland on Hainan Island, China. Mangr Salt Marsh. 3:41-49.
Lumbanraja J, Nugroho SG, Niswati A, Ardjasa WS, Subadiyasa N, Arya N, Haraguchi H, Kimura M. 1998. Methane emission from Indonesian rice fields with special references to the effects of yearly and seasonal variations, rice variety, soil type and water management. Hydrol Process. 12:2057-2072.
Marton JM, Herbert ER, Craft CB. 2012. Effects of Salinity on Denitrification and Greenhouse Gas Production from Laboratory-incubated Tidal Forest Soils. Wetlands. 32:347-357.
Morell FJ, Cantero-Martínez C, Lampurlanés J, Plaza-Bonilla D, Álvaro-Fuentes J. 2011. Soil Carbon Dioxide Flux and Organic Carbon Content: Effects of Tillage and Nitrogen Fertilization. Soil Sci Soc Am J. 75
Murdiyarso D, Purbopuspito J, Kauffman JB, Warren MW, Sasmito SD, Donato DC, Manuri S, Krisnawati H, Taberima S, Kurnianto S. 2015. The potential of Indonesian mangrove forests for global climate change mitigation. Nat Climate Change. 5:1089-1092.
Myhre G, Shindell D, Bréon F, Collins W, Fuglestvedt J, Huang J, Koch D, Lamarque J, Lee D, Mendoza B (2013) Anthropogenic and natural radiative forcing. In: Stocker T et al. (eds) Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of Intergovermental Panel of Climate Change. Cambridge University Press, Cambridge, UK and New York, pp 659-740
Nursin A, Wardah W, Yusran. 2014. Sifat kimia tanah pada berbagai zonasi hutan mangrove di Desa Tumpapa Kecamatan Balinggi Kabupaten Parigi Moutong Warta Rimba. 2:17-23.
19 Purvaja R, Ramesh R. 2000. Human impacts on methane emission from mangrove
ecosystems in India. Reg Environ Change. 1:86-97.
Reddy N, Crohn DM. 2014. Effects of soil salinity and carbon availability from organic amendments on nitrous oxide emissions. Geoderma. 235– 236:363-371.
Robertson AI, Daniel PA. 1989. The influence of crabs on litter processing in high intertidal mangrove forests in tropical Australia. Oecologia. 78:191-198. Sukardjo S, Alongi D. 2012. Mangroves of the South China Sea: Ecology and
Human Impacts on Indonesia s Forests. Nova Science Publishers Inc, New York
Vilarrúbia TV. 2000. Zonation pattern of an isolated mangrove community at Playa Medina, Venezuela. Wetlands Ecol Manage. 8:9-17.
Wang H, Liao G, D’Souza M, Yu X, Yang J, Yang X, Zheng T. 2016. Temporal
and spatial variations of greenhouse gas fluxes from a tidal mangrove wetland in Southeast China. Environ Sci Pollut R. 23:1873-1885.
Laju Dekomposisi Serasah Daun di Ekosistem Bakau Pulau Kelong, Kabupaten Bintan
Decomposition Rate of Leaf Litter in The Mangrove Ecosystem of Kelong Island, Bintan Regency
I Wayan E. Dharmawan1,2, Neviaty P. Zamani1 & Hawis H. Madduppa1
1Fakultas Perikanan dan Ilmu Kelautan, Institut Pertanian Bogor
2UPT. Loka Konservasi Biota Laut, Biak
Email: [email protected]
Submitted 28 May 2015. Reviewed 11 December 2015. Accepted 4 April 2016.
Abstrak
Hutan bakau di bagian tenggara Pulau Kelong memiliki cadangan karbon yang banyak dalam tegakan dan sedimen yang mengindikasikan produktivitas primer kawasan yang tinggi. Dekomposisi serasah daun, sebagai bagian dari produktivitas kawasan, menghasilkan unsur hara esensial sederhana yang dimanfaatkan untuk menopang pertumbuhan bakau. Tujuan penelitian ini adalah untuk mengetahui laju dekomposisi
serasah daun di setiap zona penelitian, yaitu zona darat (Landward, L), tengah (Middle zone, M), dan dekat
laut (Seaward, S) yang dibedakan berdasarkan kelimpahan jenis tumbuhan dan parameter in situ. Penelitian
dilaksanakan pada bulan Maret–Mei 2015 di kawasan bakau Pulau Kelong, Kabupaten Bintan. Proses dekomposisi diamati pada enam interval waktu, yaitu 2, 4, 7, 14, 28, dan 42 h ari dengan menggunakan kantong jaring yang berisi 10 g daun basah yang diletakkan di permukaan substrat. Hasil penelitian
menunjukkan bahwa L merupakan zona dengan persentase dekomposisi tertinggi (56,34 ± 20,58%)
dibandingkan dengan M dan S yang berturut-turut sebesar 51,59 ± 15,46% dan 51,09 ± 8,70%. Laju
dekomposisi serasah paling tinggi di zona L yaitu 1,40 ± 0,82 g·m-2·hari-1 yang berbeda nyata dari zona M
dan S, berturut-turut sebesar 0,83 ± 0,89 g·m-2·hari-1 dan 0,78 ± 0,70 g·m-2·hari-1. Analisis koefisien laju
dekomposisi (k) menunjukkan bahwa laju dekomposisi di dalam kawasan tergolong tinggi (k > 0,01). Laju
dekomposisi memiliki korelasi dengan kelimpahan spesies bakau, nilai salinitas air, dan suhu tanah.
Kata kunci: bakau, produktivitas primer, komposisi jenis, laju dekomposisi, serasah daun.
Abstract
The mangrove forests in the southeastern part of Kelong Island have abundant carbon reserves that indicate high primary productivity areas. Decomposition of leaf litter, as a part of regional productivity, produces simple essential nutrients utilized to sustain the growth of mangroves. The purpose of this study was to determine the decomposition rate of leaf litter in each zone of the study, the Landward (L), Middle zone (M), and Seaward (S) which is distinguished by the abundance of plant species and in situ environmental parameters. The experiment was conducted in March–May 2015 in the mangrove areas of
Dharmawan et al.
Island Kelong, Bintan regency. The decomposition process was observed at six time intervals, i.e. 2, 4, 7, 14, 28, and 42 days using net bag containing 10 g of wet leaves placed on the substrate surface. The results
showed that L was a zone with highest percentage of decomposition (56.34 ± 20.58%) compared to the M
and S, respectively for 51.59 ± 15.46% and 51.09 ± 8.70%. The highest rate of litter decomposition in zone L
was 1.40 ± 0.82 g·m-2·day-1 that was significantly different from the zones M and S, respectively of 0.83 ±
0.89 g·m-2·day-1 and 0.78 ± 0.70 g·m-2·day-1. Analysis of the coefficient of decomposition rate (k) indicated
that the rate of decomposition in this region was high (k > 0.01). The decomposition rate has a significant correlation with the abundance of mangrove species, the salinity value of the water, and soil temperature.
Keywords: mangrove, primary productivity, species composition, decomposition rate, leaf litter.
Pendahuluan
Bakau sebagai salah satu ekosistem pesisir merupakan komunitas tumbuhan tinggi yang mampu beradaptasi dengan baik di kawasan pasang-surut (Kathiresan & Bingham, 2001). Struktur tumbuhannya yang tinggi dan kokoh menjadikan hutan bakau berfungsi sebagai komponen penting dalam rantai makanan yang sangat kompleks dan potensial bagi kehidupan berbagai biota laut maupun terestrial, baik
mikroorganisme maupun makroorganisme
(Saenger et al., 2012; Abrantes et al., 2014). Tumbuhan bakau juga berperan sebagai sumber energi melalui guguran serasah atau bagian tumbuhan yang telah mati (Abrantes & Sheaves, 2009). Biota herbivora akan memecahnya menjadi bagian yang lebih kecil yang kemudian diuraikan oleh mikroorganisme melalui mekanisme dekomposisi (Camilleri, 1992; Robertson, 1998; Longonje & Raffaelli, 2015).
Dalam ekosistem bakau, dekomposisi merupakan proses katabolisme yang sangat penting bagi produktivitas primer kawasan. Pemecahan mekanik dan kimiawi yang dilakukan oleh organisme pengurai menghasilkan unsur hara esensial sederhana yang dapat dimanfaatkan untuk menopang pertumbuhan bakau dan biota lainnya. Kajian tentang dekomposisi sudah banyak dilakukan, baik di wilayah tropis maupun subtropis, antara lain oleh Ashton et al. (1999), Ananda et al. (2008), Li & Ye (2014), Tran (2014), dan Keuskamp et al. (2015). L etak geografis sangat penting dalam memengaruhi laju dekomposisi (Ainley & Bishop, 2015). Peningkatan aktivitas dekomposisi yang terjadi secara aerobik dapat meningkatkan emisi gas karbon dioksida ke dalam atmosfer (Cai, 2011). Hal ini disebabkan oleh kandungan organik yang tinggi dan faktor lingkungan in situ yang optimal bagi pertumbuhan organisme pengurai.
Serasah daun sebagai komponen utama dalam produtivitas primer bakau merupakan sumber karbon penting dalam proses
de-komposisi. Kualitas dan kuantitas serasah dalam ekosistem memberikan pengaruh kuat bagi aktivitas katabolisme organisme pengurai (Mooshammer et al., 2012). Kualitas serasah sangat dipengaruhi oleh jenis bakau yang memiliki kandungan nutrisi dan air yang berbeda-beda. Semakin tebal daun, maka semakin lama waktu yang dibutuhkan untuk menguraikannya (Gartner & Cardon, 2004). Luas dan keragaman bakau di Indonesia menimbulkan variasi yang cukup tinggi pada kuantitas serasah bakau yang
dihasilkan, yaitu berkisar 7,1–23,7 ton·ha-1·tahun-1
(Sukardjo & Yamada, 1992; Sukardjo et al., 2013). Oleh karena itu, penelitian ini penting untuk dilakukan di Indonesia.
Pulau Kelong terletak dalam gugusan kepulauan di bagian tenggara Kabupaten Bintan. Pulau ini dikelilingi oleh hutan bakau yang cukup lebat. Aktivitas penambangan bauksit juga banyak ditemui di kawasan ini yang dalam dekade terakhir semakin meningkat. Hasil interpretasi citra Landsat tahun 2004–2014 menunjukkan bahwa terjadi peningkatan luas lahan tambang secara horizontal. Secara vertikal, wilayah yang dulu merupakan bukit, sekarang hampir rata dengan tanah. Dharmawan et al. (2014) menjelaskan bahwa hutan bakau di Pulau Kelong memiliki cadangan karbon paling tinggi dengan kondisi vegetasi yang paling baik di antara sepuluh wilayah kajian di kawasan pesisir timur Kabupaten Bintan. Hutan bakau Pulau Kelong bagian tenggara memiliki cadangan karbon tiga kali lipat lebih besar dibandingkan dengan wilayah lain di Pulau Kelong (Dharmawan et al., 2014).
Oseanologi dan Limnologi di Indonesia 2016 1(1): 1-10
serasah daun di ekosistem bakau di Pulau Kelong bagian tenggara.
Metodologi
Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada Maret–Mei 2015 di kawasan hutan bakau Pulau Kelong bagian tenggara. Kawasan ini dipilih karena memiliki cadangan karbon tertinggi dibandingkan dengan wilayah lain di Pulau Kelong, maupun di kawasan pesisir timur Kabupaten Bintan (Dharmawan et al., 2014). Secara administratif, lokasi penelitian terletak di Kecamatan Bintan Timur yang berjarak 26 km dari pusat Kota Tanjung Pinang, Kabupaten Bintan.
Hutan bakau ini memiliki ketebalan maksimal 200 m yang merupakan hutan primer. Kawasan ini memiliki enam muara sungai kecil dengan lebar ± 1 m. Kawasan dibagi dalam tiga zona, yaitu zona dekat laut (Seaward, S), zona tengah (Middle zone, M), dan zona dekat daratan (Landward, L) yang dikaji dengan empat transek tegak lurus garis pantai (Gambar 1). Karakteristik setiap zona diperlihatkan dalam Tabel 1.
Substrat berbatu ditemukan di bagian pantai dengan ketebalan ekosistem bakau sekitar 304 m. Pasang surut air laut tipe campuran condong ke harian tunggal dengan rentang pasang tertinggi dan surut terendah sebesar 89 cm, serta kecepatan
surut maksimal 0,494 cm/menit dan pasang 0,466 cm/menit.
Analisis Kondisi Vegetasi Bakau
Penelitian ini menggunakan metode transek garis dengan plot berukuran 10 m x 10 m yang
ditarik tegak lurus garis pantai. Total plot
sebanyak 12 bua h (3 zona x 4 t ransek) dibuat untuk mengukur kondisi komunitas bakau
kategori pohon atau setinggi ± 130 cm dengan
diameter ≥ 4 cm dan kategori pancang (sapling)
dengandiameter < 4 cm. Di dalam plot 10 x 10 m2
dibuat lagi subplot berukuran 5 m x 5 m (Ashton & Macintosh, 2002). Identifikasi s pesies bakau dilakukan berdasarkan Noor et al. (1999) dan Giesen et al. (2006). Kerapatan serta indeks nilai penting jenis pada pohon dan pancang dihitung berdasarkan English et al. (1997). Persentase tutupan kanopi bakau didekati dengan metode
hemispherical photography (Dharmawan &
Pramudji, 2014). Parameter lingkungan in situ seperti suhu, pH, salinitas, konduktivitas, dan TDS dalam air dan sedimen diukur dengan
menggunakan pHmeter EZODO PH5011 dan
SCTmeter YSI EC200. Stok serasah daun (litter
standing stock) di permukaan tanah setiap plot dihitung menggunakan plot kecil berukuran 1 m x 1 m. Serasah daun dikeringkan pada suhu 80°C selama 1 minggu, kemudian ditimbang (Robertson & Daniel, 1989).
Gambar 1. Lokasi penelitian yang menunjukkan empat lokasi transek (T1–T4) serta zona daratan (L), zona tengah (M), dan zona laut (S).
Dharmawan et al.
Tabel 1. Perbandingan nilai rata-rata parameter lingkungan dalam substrat dan air di setiap zona transek (L, M, dan S) bakau di Pulau Kelong.
Table 1. Comparison of the average values of environmental parameters in each zone of mangrove transects (L, M, and S) of Kelong island.
Zone
L M S
Substrate Mud Mud Sandy mud
Species composition Xylocarpus Mostly
granatum
Different letters indicate statistically significant differences among sampling sites.
Penghitungan Laju Dekomposisi
Laju dekomposisi dihitung berdasarkan metode yang dimodifikasi dari Ashton et al. (1999) dan Dewiyanti (2010). Daun kuning yang masih segar dalam kelompok pohon dari spesies
yang dominan dikoleksi dari setiap zona, yaitu R.
apiculata dari zona S dan M, serta X. granatum
dari zona L. Daun dibersihkan, didiamkan selama
24 jam, ditimbang sebanyak 10 g (W1), lalu
dimasukkan ke dalam kantong jaring berukuran
10 x 10 c m2 dengan ukuran mata jaring (mesh
size) 1 mm2. Kantong daun diletakkan di atas
permukaan substrat bakau dan diikatkan pada akar sejajar dengan substrat untuk mencegah kantong terbawa arus pasang surut. Untuk setiap zona dibuat empat kali ulangan dan enam kali perlakuan waktu pengamatan (hari ke-2, 4, 7, 14, 28, 42). Kantong dikoleksi pada hari yang sudah ditentukan dan dibawa ke laboratorium. Daun dibersihkan dan dimasukkan ke dalam oven bersuhu 60°C selama 3–5 hari atau sampai beratnya konstan. Setiap sampel ditimbang dan
dicatat berat keringnya (W2).
Faktor konversi (f) berat basah ke berat kering dihitung sebagai persentase kandungan biomassa dalam daun kuning. Berat basah (BB) koleksi daun kuning ditimbang, kemudian dioven pada suhu 60°C selama 3–5 hari atau sampai tidak ada perubahan berat. Daun kering ditimbang untuk mendapatkan berat kering (BK).
f =BB−BK
selang waktu tertentu (Δt), persentase
terdekomposisi (D), koefisien laju dekomposisi
(k), dan waktu yang dibutuhkan untuk
mendekomposisi setengah sampel (t50) dihitung
dengan menggunakan persamaan (2)–(5)
berdasarkan Ashton et al. (1999), yaitu:
Oseanologi dan Limnologi di Indonesia 2016 1(1): 1-10
Xt = persentase serasah daun setelah
terdekomposisi dalam kurun waktu tertentu (t)
X0 = persentase serasah daun sebelum
terdekomposisi.
Analisis Data
Data univariat seperti vegetasi (kerapatan bakau kategori pohon dan pancang), persentase tutupan kanopi, stok serasah bakau, parameter lingkungan (pH, suhu, salinitas, konduktivitas, TDS dalam sedimen dan air), laju dekomposisi,
koefisien laju dekomposisi (k), dan t50 dianalisis
dengan ANOVA menggunakan perangkat lunak tidak berbayar R-studio. Analisis jarak (indeks
similaritas) Eucladian distance digunakan untuk
setiap parameter lingkungan dan komposisi jenis
pohon. Analisis multidimensional scaling (MDS)
terhadap indeks similaritas seluruh parameter digunakan untuk menentukan ordinasi setiap plot pengukuran dan menegaskan klasifikasi zonasi dengan menggunakan perangkat lunak PRIMER 5. Selain itu, indeks similaritas juga digunakan
untuk menentukan korelasi antarparameter dengan menggunakan analisis korelasi Spearman.
Hasil
Kondisi Komunitas Bakau
Kerapatan pohon di lokasi penelitian
tertinggi ditemukan di zona darat (Landward, L)
sebesar 2.175 ± 222 pohon/ha yang berbeda nyata (P < 0,05) dari dua zona yang lain. Zona dekat
laut (Seaward, S) memiliki kerapatan bakau
kategori pohon yang paling rendah. Namun, pada
kategori pancang, zona S memiliki tegakan yang
paling rapat dan berbeda nyata dibandingkan dua stasiun lain (Tabel 2). Dari segi keanekaragaman jenis, zona S hanya memiliki dua spesies bakau dan paling rendah dibandingkan zona darat dan zona tengah. Berdasarkan persentase tutupan tajuk, komunitas bakau di dalam kawasan termasuk dalam kategori baik, dengan nilai
persentase tutupan 87,96 ± 6,88% (L) sampai
91,31 ± 1,14% (M) yang tidak berbeda nyata antarzona.
Tabel 2. Kerapatan bakau kategori pohon dan pancang, jumlah jenis, indeks nilai penting (INP) pohon, dan persentase tutupan kanopi bakau serta kepadatan serasah di tiga zona (L, M, dan S) Pulau Kelong. Table 2. Density of mangrove for tree and sapling categories, number of species, importance value (IV)
index, percentage of canopy cover of mangrove, and litter stand in three zones (L, M, dan S) of Kelong Island.
Dharmawan et al.
Laju Dekomposisi
Selama 42 hari pengamatan, zona L menunjukkan proses dekomposisi tercepat, yaitu
sebesar 56,34 ± 20,58% dibandingkan dengan
zona M dan S, yaitu berturut-turut sebesar 51,59 ±
15,46% dan 51,09 ± 8,70%. Berat kering daun
terdekomposisi selama waktu pengamatan ditunjukkan dalam Gambar 2. Laju dekomposisi
rata-rata di zona L juga paling tinggi, yaitu 1,40 ±
0,82 g·m-2·hari-1 yang berbeda nyata dari dua zona
lain, yaitu zona M (0,83 ± 0,89 g·m-2·hari-1) dan
zona S (0,78 ± 0,70 g·m-2·hari-1) (Tabel 3).
Pembahasan
Komunitas bakau di bagian tenggara Pulau
Kelong didominasi oleh spesies Rhizophora
apiculata yang ditemukan di semua zona
penelitian. Kondisi substrat yang berlumpur dan anoksik memungkinkan spesies ini tumbuh dengan baik. Duke (2006) menyebutkan bahwa
Rhizophora memiliki akar tunjang yang kuat,
mengelilingi batang utama sebagai penopang pertumbuhannya di wilayah berlumpur dan anoksik. Hogarth (2007) menambahkan bahwa
akar Rhizophora memiliki lentisel yang
merupakan struktur penyerap oksigen terlarut dari air, tanah, atau udara secara difusi sebagai bentuk adaptasi terhadap lingkungan yang miskin oksigen.
Jenis substrat sangat memengaruhi dominansi suatu spesies di kawasan bakau
oseanik. Di zona S dan M dengan sedimen campuran pasir dan lumpur, spesies-spesies
Rhizophora mendominasi wilayah tersebut,
sedangkan zona L dengan substrat lumpuran yang berstruktur lebih padat didominasi oleh
Xylocarpus granatum. Zona M yang memiliki tipe
substrat campuran antara zona L dan S memiliki variasi keanekaragaman jenis yang lebih beragam. Salinitas merupakan faktor kunci yang menimbulkan variasi jenis dalam setiap zona. Salinitas air di zona S jauh lebih tinggi (32,05 ± 0,06 ppt) dan berbeda signifikan dibandingkan dengan zona L dan M (24,58 ± 0,67 ppt dan 24,13 ± 2,56 ppt). Tren yang sama juga ditemukan untuk nilai konduktivitas dan TDS air.
Perbedaan gradien konsentrasi parameter lingkungan (elevasi morfologi pantai, gradien komposisi fraksi sedimen, salinitas, gradien hara, dan genangan) yang tegak lurus garis pantai akan memengaruhi zonasi dan komposisi jenis tumbuhan yang menyusunnya (Lopez-Portillo & Ezcurra, 1989; Fujimoto & Miyagi, 1993; Vilarrubia, 2000).
Perbedaan gradien konsentrasi parameter lingkungan (elevasi morfologi pantai, gradien komposisi fraksi sedimen, salinitas, gradien hara, dan genangan) yang tegak lurus garis pantai akan memengaruhi zonasi dan komposisi jenis tumbuhan yang menyusunnya (Lopez-Portillo & Ezcurra, 1989; Fujimoto & Miyagi, 1993; Vilarrubia, 2000).