PERBANDINGAN KARAKTER MORFOLOGI DAN
MOLEKULER ANTARA KARANG
Acanthophyllia deshayesiana
(Michelin, 1850) DENGAN
Cynarina lacrymalis
(Milne Edwards
& Haime, 1848)
ROBBA FAHRISY DARUS
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA*
Dengan ini saya menyatakan bahwa tesis berjudul Perbandingan Karakter Morfologi dan Molekuler Antara KarangAcanthophyllia deshayesiana (Michelin, 1850) dengan Cynarina lacrymalis (Milne Edwards & Haime, 1848) adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, April 2016
Robba Fahrisy Darus
RINGKASAN
ROBBA FAHRISY DARUS. Perbandingan Karakter Morfologi dan Molekuler Antara Karang Acanthophyllia deshayesiana (Michelin, 1850) dengan Cynarina lacrymalis (Milne Edwards & Haime, 1848). Dibimbing oleh NEVIATY P ZAMANI, SUHARSONO, dan DEDY DURYADI SOLIHIN.
Cynarina lacrymalis memiliki sinonim yaitu Acanthophyllia deshayesiana
sebagaimana terdaftar pada WoRMS (World of Registered Marine Species). Secara klasik, identifikasi karang menggunakan karakter morfologi dari kerangka (skeleton) karang untuk pengklasifikasian dalam dunia taksonomi. Perdebatan identifikasi dan pengklasifikasian berdasarkan karakter morfologi masih meragukan dan cenderung subjektif, sehingga adanya metode molekuler menjadi kajian baru untuk mengkonfirmasi identifikasi dan klasifikasi yang sudah ada. Sistematika pengklasifikasian karang secara klasik menggunakan morfologi kerangka karang dengan tujuan untuk mengetahui hubungan kekerabatan dan evolusinya.
Karakter morfologi yang digunakan terdiri dari karakter deskriptif dan morfometrik. Data karakter morfometrik dianalisa dengan analisis
Corresspondence Analysis of Principal Coordinates (CAP), dan klaster hierarki
Agglomerative Hierarchical Clustering (AHC) menggunakan perangkat lunak XLSTAT 2015. Kladogram berdasarkan karakter deskriptif dan morfometrik dianalisis menggunakan Unweighted Pair Group Method with Arithmetic Mean
(UPGMA) perangkat lunak PAUP 4. Data molekuler dihasilkan dari beberapa prosedur yaitu ekstraksi DNA total, amplifikasi PCR pada gen target ribosomal DNA dengan marka ITS dan sekuensing. Rekonstruksi pohon filogenetik menggunakan metode Neighbour Joining dengan model kimura 2-parameter 1000
bootstrap replikasi pada MEGA 5.2.
Berdasarkan kladogram karakter deskriptif, spesies A. deshayesiana dan C. lacrymalis dapat dibedakan dengan jelas pengelompokkannya secara terpisah, sedangkan pengelompokan berdasarkan karakter morfometrik tidak terlihat pengelompokannya dengan jelas dari hasil analisis CAP, AHC dan kladogram UPGMA. Hasil pohon filogenetik juga menunjukkan perbedaan antara kelompok karang C. lacrymalis dengan A. deshayesiana, hal ini dapat dilihat dari perbedaan jarak genetik sebesar 6,3 ± 4.3% yang berada diatas ambang batas jarak genetik sebesar ≤ 3%. Selain itu karakter molekuler dapat menemukan adanya variasi spesies yang dapat dimungkinkan sebagai subspesies dari kedua karang
Penelitian ini menyimpulkan bahwa pengelompokan berdasarkan karakter morfometrik masih tidak jelas, sedangkan karakter deskriptif, dan molekuler sudah dapat membedakan pengelompokan karang C. lacrymalis dan A. deshayesiana
secara terpisah. Secara genetik, kedua spesies tersebut memiliki keragaman intraspesies.
SUMMARY
ROBBA FAHRISY DARUS. Comparison of morphological and molecular characters between Acanthophyllia deshayesiana (Michelin, 1850) with Cynarina lacrymalis (Milne Edwards & Haime, 1848). Supervised by NEVIATY P ZAMANI, SUHARSONO, dan DEDY DURYADI SOLIHIN.
The Acanthophyllia deshayesiana is a synonym species of Cynarina lacrymalis that its registered in World of Registered Marine Species (WoRMS). Traditional identification of coral use morphological characters by skeleton for the coral taxonomy. Traditional identification based on morphological characters is distinct and subjective, so the molecular approach become a new study to confirm the identification and classification of coral species. Traditional systematic of coral use the skeleton to describe the relationship and evolution.
The morphometric and descriptive characters are used to described these coral. The morphometric data were analyzed by Correspondent Analysis of Principal Coordinates (CAP), and Agglomerative Hierarchical Clustering (AHC) to know the separation of species groups using XLSTAT 2015. The cladogram based morphometric and descriptive character were analyzed by Unweighted Pair Group Method with Arithmetic Mean (UPGMA) using PAUP 4. The molecular data were generated by a procedures that DNA extraction, PCR amplification (rDNA gen target and ITS markers) and sequencing. The reconstruction of phylogenetic tree used Neighbour Joining method with a kimura 2-parameter and 1000 bootstrap replicate by the MEGA 5.2.
The result of CAP, AHC, and UPGMA show that C. lacrymalis and A. deshayesiana can’t be grouped clearly based on morphometric characters, while
grouping with descriptive characters can differentiate clearly between C. lacrymalis
and A. deshayesiana. Phylogenetic tree show the difference between the group of C. lacrymalis with A. deshayesiana, it is known by the difference from genetic distance 6,3 ± 4.3%, which is above the threshold of genetic distance ≤ 3%. In addition, molecular characters can find a species variation which can be made as a subspecies both both these coral.
This study concludes that C. lacrymalis and A. deshayesianacan’t be grouped
based on morphometric characters clearly, while grouping with descriptive and molecular characters can defferentiate clearly between C. lacrymalis and A. deshayesiana. Based on genetic data, both of these species have intraspecies diversity.
© Hak Cipta Milik IPB, Tahun 2016
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
PERBANDINGAN KARAKTER MORFOLOGI DAN
MOLEKULER ANTARA KARANG
Acanthophyllia deshayesiana
(Michelin, 1850) DENGAN
Cynarina lacrymalis (Milne Edwards
& Haime, 1848)
ROBBA FAHRISY DARUS
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Ilmu Kelautan
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga karya ilmiah ini berhasil diselesaikan. Tema dalam penelitian yang dilaksanakan sejak Bulan Januari – November 2015 ini ialah Perbandingan Karakter Morfologi dan Molekuler antara Karang Acanthophyllia deshayesiana (Michelin, 1850) dengan Cynarina lacrymalis (Milne Edwards & Haime, 1848)
Terima kasih penulis ucapkan kepada semua pihak yang turut berperan dalam memberi arahan, dukungan , dan motivasi, mulai dari saat studi hingga penelitian dan penyusunan tesis ini, terutama kepada :
Dr Ir Neviaty P Zamani, M.Sc, Prof Dr Suharsono dan Dr Ir Dedy Duryadi Solihin, DEA selaku pembimbing
Bapak Agus Budiyanto dari Pusat Penelitian Oseanografi LIPI yang telah membantu dalam koleksi dan preservasi sampel
Gugus Kendali Mutu (GKM) FPIK IPB terutama kepada Dr Tri Prartono atas masukan dan arahannya dalam perbaikan naskah tesis ini
Direktorat Jenderal Pendidikan Tinggi (DIKTI) Kementerian Riset dan Pendidikan Tinggi, atas Beasiswa Pendidikan Pascasarjana Dalam Negeri (BPPDN) yang diberikan selama studi
Ayah, ibu, dan adik atas segala doa dan kasih sayangnya St Aisyah yang telah sabar menunggu selesainya studi ini
Rekan-rekan seperjuangan Alumni Fakultas Pertanian Universitas Trunojoyo Madura yang melanjutkan studi di IPB atas segala bantuan dan dukungannya Rekan-rekan di Laboratorium Biologi Molekuler PPSHB-IPB yang telah
memberi masukan dan berbagi ilmunya
Rekan-rekan Arafah yang telah memberikan semangat dan dukungan agar terselsaikannya studi ini
Rekan-rekan seperjuangan Mahasiswa Pascasarjana Ilmu Kelautan 2013 yang tetap setia menyemangati untuk segera menyelesaikan dan meluangkan waktu dalam seminar dan siding tesis
Penulis berharap karya ilmiah ini bermanfaat dalam perkembangan ilmu pengetahuan terutama dalam bidang Ilmu Kelautan.
Bogor, April 2016
DAFTAR ISI
PRAKATA x
DAFTAR ISI xi
DAFTAR TABEL xii
DAFTAR GAMBAR xii
DAFTAR LAMPIRAN xii
1 PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 3
Tujuan Penelitian 3
Manfaat Penelitian 3
Kerangka Pemikiran 4
2 METODE PENELITIAN 4
Waktu dan Tempat Penelitian 4
Koleksi Sampel 5
Data Morfologi 5
Data Molekuler 7
Analisa Data 8
3 HASIL DAN PEMBAHASAN 9
Analisis Morfologi 9
Perbandingan dan Kladogram morfologi antara A. deshayesiana
dengan C. lacrymalis 17
Analisis Molekuler 19
Implikasi Terhadap Taksonomi 28
SIMPULAN DAN SARAN 29
DAFTAR PUSTAKA 29
LAMPIRAN 33
DAFTAR TABEL
1. Kategori pengkelasan karakter deskriptif pada koralit karang 6 2. Karakter morfometrik yang digunakan dalam pengukuran koralit
karang 7
3. Perbandingan karakter morfometrik dan deskriptif karang C.
lacrymalis dan A. deshayesiana 18
4. Hasil blast basa nukliotida pada Gen Bank 20
5. Variasi situs nukliotida spesifik antara karang A. deshayesiana
dengan C. lacrymalis 24
6. Variasi situs nukliotida spesifik karang A. deshayesiana 25 7. Variasi situs nukliotida spesifik karang C. lacrymalis 25
DAFTAR GAMBAR
1. Diagram alur penelitian 4
2. Kondisi hidup karang: a) A. deshayesiana; b) C. lacrymalis 5 3. Uji morfologi pada kerangka karang C. lacrymalis dan A.
deshayesiana: 6
4. Perbandingan kerangka karang spesimen uji. A-H) C. lacrymalis, dan
I-O) A. deshayesiana 10
5. Bentuk gigi septa, septa dan pali, dentasi costae karang A.
deshayesiana dan C. lacrymalis. 11
6. Jumlah septa dan jumlah gigi septa karang A. deshayesiana. 12 7. Jumlah septa dan jumlah gigi septa karang A. deshayesiana. 13 8. Jumlah septa dan jumlah gigi septa karang C. lacrymalis. 14 9. Jumlah septa dan jumlah gigi septa karang C. lacrymalis 15 10.Pengelompokan karang C. lacrymalis dan A. deshayesiana
berdasarkan karakter morfometrik menggunakan analisis
Corresspondence Analysis of Principal Coordinates (CAP) 16 11.Dendogram karang C. lacrymalis dan A. deshayesiana berdasarkan
karakter morfometrik menggunakan Agglomerative Hierarchical
Clustering (AHC) 17
12.Kladogram morfologi menggunakan metode UPGMA 19
13.Hasil elektroforesis amplifikasi PCR 20
14.Hasil analisis data molekuler menggunakan Neighbour Joining model
Kimura 2-parameter 1000 bootstrap 27
DAFTAR LAMPIRAN
1. Karakter Morfometrik Karang Cynarina lacrymalis dan
Acanthophyllia deshayesiana 35
2. Matriks pengkelasan karakter deskriptif dan morfometrik 36
3. Matriks jarak genetik 37
4. Hasil analisis CAP menggunakan XL Stat 2015 38
1 PENDAHULUAN
Latar Belakang
Identifikasi karang lebih sulit dibandingkan dengan identifikasi tumbuhan dan ikan karena kunci identifikasi karang antar spesies, genus, dan famili berbeda-beda. Keberadaan ekosistem terumbu karang yang luas, perbedaan kondisi geografis, perubahan iklim, dan kondisi lingkungan menjadi faktor perbedaan jenis-jenis karang. Identifikasi jenis karang sudah banyak dilakukan tetapi masih memiliki permasalahan karena terjadi beberapa kemiripan karakter morfologi. Hal ini menyebabkan satu spesies karang memungkinkan memiliki kesamaan dengan spesies lain (sinonim). Sebagai contoh karang jenis Austrolomussa rowleyensis
mirip dengan Scolymia vitiensis, Cynarina lacrymalis mirip dengan Acantophyllia deshayesiana, Wellsophyllia radiata mirip dengan Trachyphyllia geoffroyi, dan
Symphyllia valenciennesi mirip dengan Symphyllia agaricia (Veron 2000, Suharsono 2010).
Kesamaan jenis karang berdampak pada pemanfaatan karang sebagai biota yang diperdagangkan. Karang yang diperdagangkan biasanya memiliki bentuk yang unik, menarik untuk dilihat dan warna yang mencolok (Giyanto 2007). Tahun 1970 menjadi awal terjadinya ekspor terumbu karang di Indonesia. Spesies sinonim dan penamaan pada karang yang belum selesai menjadi masalah bagi eksportir karang di Indonesia. Eksportir harus mengikuti permintaan importir sesuai nama dan jenis karang yang diminta meskipun harus melakukan pensortiran kembali. Penamaan jenis karang ini menjadi faktor penentu dalam keberlangsungan perdagangan dan pengawasan terhadap koloni karang. Pengawasan terhadap jenis karang yang diekspor harus sesuai dengan permintaan, jika tidak sesuai akan terjadi pengembalian dan pemberian denda kepada eksportir. Pendaftaran nama karang yang diperdagangkan sangat penting untuk menentukan harga, kuota spesies, dan kelancaran ekspor karang. Hal ini berkaitan juga dengan kerjasama bilateral kedua Negara dalam kegiatan ekspor impor. Namun demikian, dibutuhkan identifikasi ulang terkait jenis karang yang mirip dengan tujuan memperbaiki sistem klasifikasi karang.
Sistematika pengklasifikasian karang bertujuan untuk mengetahui hubungan kekerabatan dan evolusinya (Stobart 2000, Flot et al. 2008, Casebolt 2011), dan hingga saat ini masih didasarkan pada morfologi kerangka karang (Wolstenholme
et al. 2003, Schmidt-Roach et al. 2012, Arrigoni et al. 2014b). Namun demikian, karang memiliki tingkat plastisitas tinggi (Todd 2008) karena karang memiliki sifat responsif terhadap lingkungan (Hilbish 1985, Doebeli dan Dieckmann 2003, Prada
et al. 2008), sehingga karang memiliki morfologi yang beranekaragam. Hal ini menyebabkan pengklasifikasian karang menjadi rancu dan penuh keraguan, serta berdampak pada beberapa karang dikelompokkan dan diberi nama sama (Flot et al. 2008, Schmidt-Roach et al. 2012).
berdasarkan WoRMS (World of Registered Marine Species)1. Secara visual di lapangan, karang hias ini memiliki kesamaan karakter morfologinya. Wells (1937) mendeskripsikan bahwa karang A. deshayesiana memiliki kemiripan variasi morfologi kerangka koralit dengan C. lacrymalis. Best dan Hoeksema (1987) mendeskripsikan kesamaan antara kedua karang tersbut yaitu hidup soliter, memiliki diameter koralit 10 cm, tinggi koralit 8 cm, dentasi gigi septa dengan tinggi hingga 15 mm, epitheca berkembang dengan baik, korallumnya bergerigi dan sangat kuat. Holotipe karang C. lacrymalis terdapat di MNHN (Museum National
d’Histoire Naturelle, Paris, France) dengan karang asal Filipina sebagai tipe lokal
jenis ini. Namun, kedua spesies ini berada pada habitat yang berbeda, oleh karena itu perlu dilakukan observasi untuk mengkonfirmasi berdasarkan karakter morfologi yang menyeluruh yaitu deskriptif dan morfometrik.
Karakter morfologi merupakan dasar dari metode identifikasi karang secara klasik (Wolstenholme et al. 2003, Schmidt-Roach et al. 2012, Arrigoni et al. 2014b). Budd et al. (2012) menggunakan karakter mikromorfologi, makromorfologi, dan mikrustruktur untuk mengidentifikasi dan mengklsifikasikan Famili Mussidae. Benzoni et al. (2012) hanya menggunakan karakter makro dan mikromorfologi saja untuk melihat adanya family baru pada karang Genus
Craterastrea. Kongjandtre et al. (2012) menggunakan analisis morfometrik untuk melihat batasan-batasan spesies pada karang Genus Favia. Arrigoni et al. (2014a) dan Arrigoni et al. (2015) menggunakan karakter mikro dan makromorfologi untuk melihat taksonomi karang Genus Austrolomussa dan Parascolymia serta melihat adanya pemulihan Genus Sclerophyllia dari system taksonomi yang telah dilupakan. Perdebatan identifikasi dan pengklasifikasian berdasarkan karakter morfologi masih meragukan dan cenderung subjektif, sehingga adanya metode molekuler menjadi kajian baru untuk mengkonfirmasi identifikasi dan pengkalsifikasian yang sudah ada.
Sistematika karang baru-baru ini mengalami revolusi yang sangat cepat karena kemajuan ilmu pengetahuan berupa pendekatan filogenetik molekuler. Keunggulan filogenetik adalah karakter yang dibandingkan relatif stabil dan tidak dipengaruhi lingkungan seperti karakter morfologi. Beberapa penanda molekuler yang telah berhasil memilah spesies pada karang secara baik adalah marka genetik COI, ITS, dan H3 (Benzoni et al. 2012, Budd et al. 2012, Kongjandtre et al. 2012, Arrigoni et al. 2014a, Arrigoni et al. 2014b, Arrigoni et al. 2015). Analisis data molekuler menggunakan penanda genetik ITS pada gen target rDNA memilki keunggulan karena dapat mencirikan setiap individu dari satu spesies (Fukami et al. 2008, Huang et al. 2011, Arrigoni et al. 2012, Benzoni et al. 2012, Arrigoni
et al. 2014b). Selain itu, kombinasi antara karakter morfologi dan molekuler untuk mengidentifikasi, dan melihat pola evolusi pada karang. Arrigoni et al. (2014b) membuktikan bahwa dari data morfologi terjadi konsistensi dengan memanfaatkan analisis molekuler pada gen target COI dan ITS. Arrigoni et al. (2014a) melakukan pembukatian pada Genus Autrolomussa dan Parascolymia dengan menggunakan marka genetik COI, ITS, dan H3. Pembuktian karang genus Craterastea dengan memadukan data morfologi dan molekuler untuk sistematikanya (Benzoni et al. 2012). Stefani et al. (2008) dengan mencari karakter morfometrik untuk
1 WoRMS (World of Registered Marine Species) merupakan website resmi untuk klasifikasi
menyelesaikan batasan-batasan pada karang genus Psammocora yang dikombinasikan dengan karakter genotipiknya. Revisi spesies yang saling berkaitan pada karang jenis Pocillopora damicornis menggunakan data morfologi dan genetik (Schmidt-Roach et al. 2012). Wolstenholme et al. (2003) melakukan pencarian batasan-batasan spesies pada kelompok karang Acropora humilis yang memanfaatkan data morfologi dan mengkombinasi dengan data molekuler untuk menginterpretasi pola evolusinya. Identifikasi jenis menggunakan dinding koralit karang dan mikrostruktur septa sebagai pembanding dengan data molekuler pada kelompok Famili Faviidae telah dilakukan oleh Budd et al. (2012). Pengkajian kembali taksonomi Montipora digitata berdasarkan genetik dan morfometriknya (Stobart 2000). Pemanfaatan karakter morfologi dan molekuler dalam penelitian taksonomi serta filogeni sangat penting untuk menjawab beberapa permasalahan yang telah dipaparkan di atas. Pembuktian sinonim spesies dengan memanfaatkan data molekuler diharapkan dapat menyelesaikan keraguan dalam identifikasi dan sistematika taksonomi karang.
Perumusan Masalah
Berdasarkan uraian diatas, permasalahan yang menjadi dasar penelitian ini yaitu karang A. deshayesiana terdaftar sebagai C. lacrymalis, dan beberapa peneliti menganggap bahwa A. deshayesiana merupakan sinonim dari C. lacrymalis
berdasarkan klasifikasi dari karakter morfologinya.
Tujuan Penelitian
Tujuan penelitian ini untuk menganalisis morfologi dan molekuler untuk mendeskripsikan perbedaan spesies sinonim antara karang A. deshayesiana dengan
C. lacrymalis.
Manfaat Penelitian
Kerangka Pemikiran
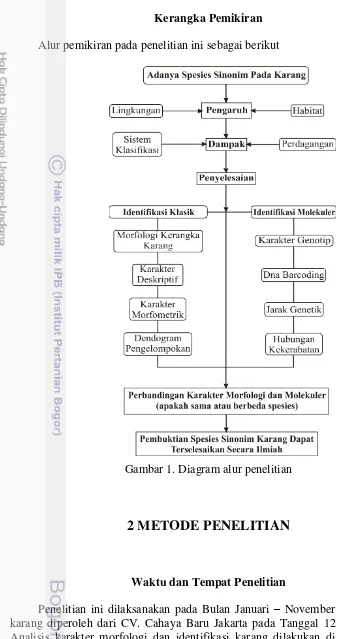
Alur pemikiran pada penelitian ini sebagai berikut
Gambar 1. Diagram alur penelitian
2 METODE PENELITIAN
Waktu dan Tempat Penelitian
Koleksi Sampel
Sampel yang digunakan adalah karang jenis C. lacrymalis (8 individu) dan A. deshayesaiana (7 individu) dari Famili Mussidae yang berasal dari Makassar (Gambar 2). Karang dari lapang terlebih dahulu diidentifikasi berdasarkan kunci identifikasi karang yang ditulis oleh Veron (2000) dan Suharsono (2010) dan pengambilan gambar sampel hidup serta pemberian label. Preservasi untuk ekstraksi DNA dilakukan dengan menambahkan etanol 96% teknis, dan sampel didiamkan selama ± 7 hari agar mudah dalam pengambilan daging (jaringan) karang dari kerangka (Oppen et al. 2000, Wolstenholme et al. 2003, Filatov et al. 2013, Kitano et al. 2014). Pelabelan dilakukan pada saat preservasi dan dilanjutkan kembali dengan pengambilan foto berlabel. Sampel karang yang akan dianalisis karakter morfologi, terlebih dahulu diputihkan dengan menggunakan larutan
Sodium hypoclorite, selanjutnya dilakukan pembilasan dengan air tawar dan dijemur. Sampel karang yang dinalisis karakter morfologi terdiri dari karakter deskriptif dan morfometrik yang saling berkaitan (Oppen et al. 2000, Wolstenholme et al. 2003, Stefani et al. 2008, Filatov et al. 2013, Kitano et al. 2014).
Gambar 2. Kondisi hidup karang: a) A. deshayesiana; b) C. lacrymalis
Data Morfologi
Karakter Deskriptif
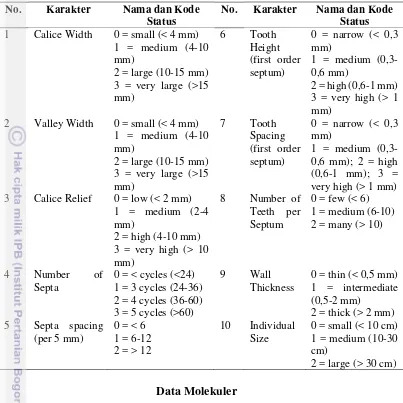
Pengamatan karakter desktiptif karang dilakukan dengan pengambilan gambar menggunakan kamera Canon powershoot D30. Karakter deskriptif yang diuji terdiri dari 11 karakter (Tabel 1 dan Gambar 3) (Budd dan Stolarski 2009, Casebolt 2011, Arrigoni et al. 2012, Benzoni et al. 2012, Budd et al. 2012, Arrigoni et al. 2014a, Arrigoni et al. 2014b).
Karakter Morfometrik
Tabel 1. Kategori pengkelasan karakter deskriptif pada koralit karang
Tabel 2. Karakter morfometrik yang digunakan dalam pengukuran koralit karang
Ekstraksi DNA menggunakan kit ekstraksi Qiagen DNaeasy Blood and Tissue kit, dimana prosedur ekstraksi mengikuti petunjuk dari perusahaan (Arrigoni
et al. 2012, Arrigoni et al. 2014a, Arrigoni et al. 2014b). Modifikasi juga dilakukan pada beberapa langkah untuk mempermudah mendapatkan DNA total karang. Hasil DNA total karang dilihat pada agar 1,2% (0,3 gr agarose, 50 ml buffer TAE 1X, ETBR 2,5 µl) dalam buffer TAE 1X, yang dialiri arus listrik 85 volt selama 30 menit (Hardja 2009, Haerul 2014).
Amplifikasi PCR dan Sekuensing
Hasil ekstraksi DNA total karang selanjutnya diamplifikasi dengan primer universal yaitu ITSZF forward (5’-TAAAAGTCGTAACAAGGTTTCCGTA–3’) dan ITSZR reverse (5’–CCTCCGCTTATTGATATGCTTAAAT–3’) (Hardja 2009, Haerul 2014). Panjang target gen ITS yang akan teramplifikasi pada penelitian ini yaitu ± 710 bp.
8,8 µl; 5 x Q5 Buffer 5 µl; 5 x Enhancher Q5 5µl; dNTP 1µl;ITSZF 10 pmol 1µl; ITSZR 10 pmol 1µl; template DNA 3 µl; dan Taq Hot Start Q5 0,2µl. Kondisi siklus PCR pada saat amplifikasi meliputi predenaturasi 95°C selama 3 menit; denaturasi 95°C selama 45 detik; annealing 51°C selama 1 menit; dan ekstensi 72°C selama 1 menit sebanyak 35 siklus, selanjutnya ditutup dengan ekstensi akhir 72°C selama 5 menit. Hasil PCR dilihat pada agar 1,2% (0,3 gr agarose, 50 ml buffer TAE 1X, ETBR 2,5 µl) dalam buffer TAE 1X, dialiri arus listrik 85 volt selama 60 menit (Hardja 2009, Benzoni et al. 2012, Budd et al. 2012, Arrigoni et al. 2014a, Haerul 2014, Reijnen et al. 2014). PCR produk yang baik jika terdapat pita pada saat dilakukan elektroforesis dan siap untuk dilakukan sekuensing. Sekuensing ini bertujuan untuk melihat runutan rantai basa nukliotida pada gen target. Sekuen produk PCR dikirim ke First Base Malaysia dengan pengiriman sekuen secara lengkap baik primer forward dan reverse serta komposisi primer yang digunakan sesuai pada saat amplifikasi.
Analisa Data
Analisis Morfologi
Data karakter morfometrik dianalisa dengan analisis Corresspondence Analysis of Principal Coordinates (CAP), dan klaster hierarki menggunakan
Agglomerative Hierarchical Clustering (AHC) untuk mengetahui pengelompokan spesies terhadap setiap karakter menggunakan perangkat lunak XLSTAT 2015 (Wolstenholme et al. 2003, Stefani et al. 2008). Data karakter deskriptif dan morfometrik dari hasil pengkelasan, selanjutnya dianalisis dengan metode
Unweighted Pair Group Method with Arithmetic Mean (UPGMA) menggunakan perangkat lunak PAUP 4 (Swofford 2002, Arrigoni et al. 2012, Benzoni et al. 2012, Arrigoni et al. 2014b) untuk menghasilkan kladogram berdasarkan karakter deskriptif dan morfometrik.
Analisis Molekuler
Hasil sekuensing yang terdiri dari forward dan reverse dilakukan pensejajaran, koreksi dan penggabungan menggunakan perangkat lunak MEGA 5.2. Sekuen yang telah disejajarkan selanjutnya dikoreksi dengan primer ITSZF dan ITSZR untuk memastikan bahwa primer menempel dengan sempurna. Sekuen dari setiap sampel dianalisis dengan melakukan pensejajaran menggunakan proses BLAST pada Gen Bank untuk mengetahui kedekatan terhadap sekuen lain yang tersimpan dalam Gen Bank. Kedekatan tersebut yang menjadi data sekuen sekunder untuk dianalisis sehingga menghasilkan pohon filogeni.
Sekuen yang digunakan pada penelitian ini yaitu 6 individu dari spesies
Acanthophyllia deshayesiana dan 7 individu dari spesies Cynarina lacrymalis. Selain itu, sekuen juga diunduh dari Gen Bank yaitu 2 spesies Symphyllia valenciennesii (HF954347.1 dan HF954349.1); 2 spesies Parascolymia vitiensis
(LK022368.1 dan LK022375.1); 1 spesies Australomussa rowleyensis
mengetahui matriks jarak genetiknya menggunakan jarak genetik model kimura 2-parameter. Rekonstruksi pohon filogenetik menggunakan metode Neighbour Joining dengan model kimura 2-parameter 1000 bootstrap replikasi pada MEGA 5.2. (Stefani et al. 2008, Arrigoni et al. 2012, Budd et al. 2012, Arrigoni et al. 2014a).
3 HASIL DAN PEMBAHASAN
Analisis Morfologi
Karakter Deskriptif
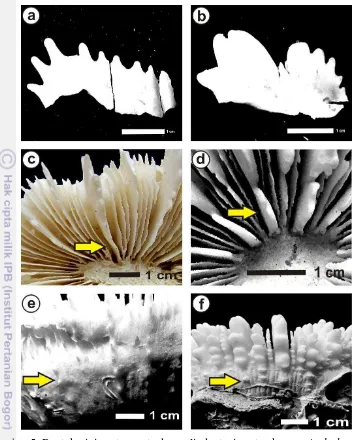
Bentuk koloni karang A. deshayesiana adalah soliter, sehingga tipe buddingnya tidak ada. Bentuk dindingnya synapticulothecal dengan struktur kolumellanya styloform (Gambar 4). Bentuk gigi pada septa karang ini kurus/kecil dan runcing, dan tidak memiliki pali, dimana semuanya adalah septa dari lembah kaliknya (Gambar 5). Karang A. deshayesiana memiliki septa yang berlanjut hingga costae dan memiliki pola costae sama, garis costea sama, dentasi pada costaenya pendek dan gemuk, serta coenesteumnya terlihat halus.
Bentuk koralit karang C. lacrymalis juga soliter dengan bentuk gigi pada setiap septa primer tebal, besar dan tumpul (Gambar 5). Pali pada karang ini berkembang dengan baik seperti yang dilaporkan dari hasil observasi Budd dan Stolarski (2009) bahwa karang C. lacrymalis memiliki septal lobe yang berkembang dengan baik. Tipe budding tidak ada karena bentuknya soliter, akan tetapi costaenya berlanjut, sehingga pola kolumellanya sama yang terlihat dari keberadaan garis costae, dan dentasi gigi pada costaenya runcing, kurus dan tajam (Gambar 5). Struktur dinding karang ini adalah synapticulothecal, struktur kolumellanya adalah styliform, dan memiliki coenesteum yang halus. Budd et al.
(2012) melaporkan bahwa karakter struktur dinding dengan bentuk synapticulothecal hanya dimiliki Genus Diploastrea.
Karakter Morfometrik
Karang A. deshayesiana memiliki tipe hidup soliter dengan kategori polip besar, sehingga ukuran individu pada 8 individu yang diobservasi berkisar 48-75 mm (Gambar 6, Gambar 7, dan Lampiran 1). Lebar kalik pada karang ini yaitu 42-62 mm, di mana nilai ini lebih besar dari nilai status karakter pada penelitian yang dilakukan oleh Budd et al. (2012). Besaran nilai lebar kalik mempengaruhi pada seluruh karakter, akan tetapi berbeda pada lebar lembah yaitu lebih kecil (6-9 mm), lebih kecil dari karang Parascolymia vitiensis (2,5 cm) (Arrigoni et al. 2014a), dan
sesuai dengan observasi pada karang spesies Parascolymia rowleyensis yaitu < 6 septa (Arrigoni et al. 2014a). Tinggi dan jarak antar gigi pada septa pertama sangat tinggi yaitu sebesar 3-5 mm, dan 4-6 mm. Jumlah septa yaitu 4 siklus (36-60) septa, sedangkan jumlah gigi pada septa primernya berkisar antara 6-12 gigi.
Gambar 5. Bentuk gigi septa, septa dan pali, dentasi costae karang A. deshayesiana
dan C. lacrymalis.
Karang C. lacrymalis memiliki ukuran individu yaitu berkisar 29-52 mm dengan dinding yang tebal yaitu 4-8 mm berdasarkan spesimen yang ada (Gambar 8 dan Gambar 9). Lebar kalik karang ini adalah berkisar 26-45 mm, yang memiliki kemiripan dengan identifikasi secara klasik di mana lebar kalik > 30 mm dapat dikatakan sangat lebar (Kongjandtre et al. 2012). Lebar lembah kalik medium dengan kisaran ukuran 5-9 mm, sedangkan relief kaliknya sangat tinggi (14-23 mm). Budd dan Stolarski (2009) melaporkan bahwa karang C. lacrymalis memiliki kalik relief yang tinggi (> 10 mm). Setiap 5 mm karang ini memiliki 3-4 septa dengan jumlah septanya 5 siklus. Arrigoni et al. (2015) melaporkan dari hasil penelitiannya bahwa jarak antar septa (per 5 mm) pada karang C. lacrymalis adalah lebar dan jumlah septanya 5 siklus. Jumlah gigi pada septa primernya yaitu 4-8 gigi, dengan jarak antar giginya sangat lebar yaitu 3-6 mm, dan tinggi gigi pada septa pertama yaitu 3-5 mm. Jarak antar gigi pada septa pertama dan tinggi gigi karang
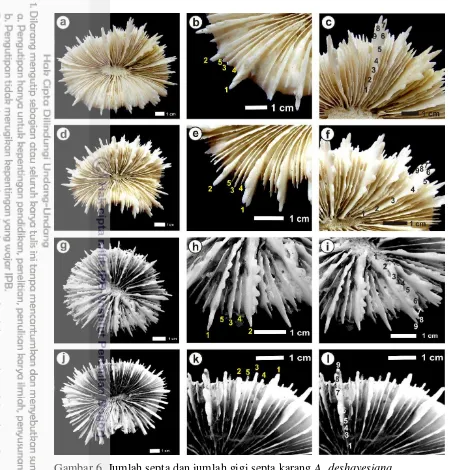
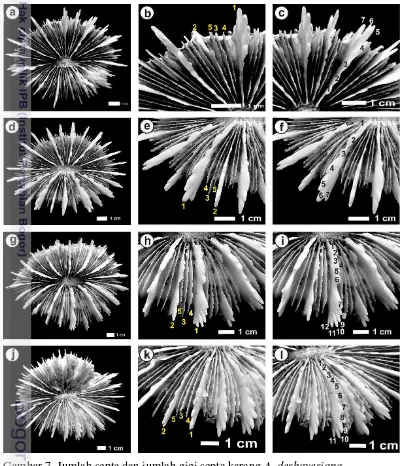
Gambar 6. Jumlah septa dan jumlah gigi septa karang A. deshayesiana.
Berdasarkan grafik simmetryc plot hasil CAP (Lampiran 4) dan AHC (Lampiran 5), terdapat 5 pengelompokan yang menginterpretasikan hubungan antara karakter morfometrik terhadap spesimen karang yaitu kelompok 1 (ACT3, ACT11, ACT7, ACT19, ACT21, ACT1, dan ACT41), kelompok 2 (ACT10, CYN40, CYN54, dan CYN56), kelompok 3 (ACT12, dan CYN50), kelompok 4 (CYN45), dan kelompok 5 (CYN59).
besaran nilai untuk lebar kalik dan lembah pada karang C. lacrymalis adalah > 30mm (Budd dan Stolarski 2009). Karang C. lacrymalis yang dikenal dengan karang polip besar merupakan karang yang memiliki lebar dan lembah kalik paling besar dalam Famili Mussidae dibandingkan Genus Favia yang berkisar antara 9-15 mm (Kongjandtre et al. 2012).
Kelompok 2 dikelompokkan berdasarkan kedalaman kalik/tinggi kalik (CR) (Gambar 10). Tinggi kalik yang dimiliki kelima karang ini lebih berhubungan dan memiliki nilai yang relatif sama yaitu 18,770 mm hingga 22,650 mm. Tinggi kalik pada karang C. lacrymalis masuk dalam kategori sangat tinggi yaitu > 10 mm (Budd dan Stolarski 2009). Bentuk kalik dipengaruhi oleh lebar kalik, dimana semakin lebar kalik akan semakin kecil tinggi kalik dan sebaliknya (Kongjandtre et al. 2012, Huang et al. 2014).
Kelompok 3 yang terdiri karang ACT12 dan CYN50 dikelompokkan berdasarkan karakter ukuran individu (IS) (Gambar 10). Hasil CAP menunjukkan bahwa posisi ACT12 sedikit menjauh, akan tetapi pada hasil AHC berada dalam satu kelompok dengan nilai kesamaanya 99,77% (Gambar 11). Ukuran individu berbanding lurus dengan lebar kalik dan lebar lembah. Selain itu, ukuran individu ini sangat dipengaruhi oleh umur dan pertumbuhan karang. Umur dan pertumbuhan karang yang semakin dewasa akan memiliki kalik dan lembah yang lebar juga.
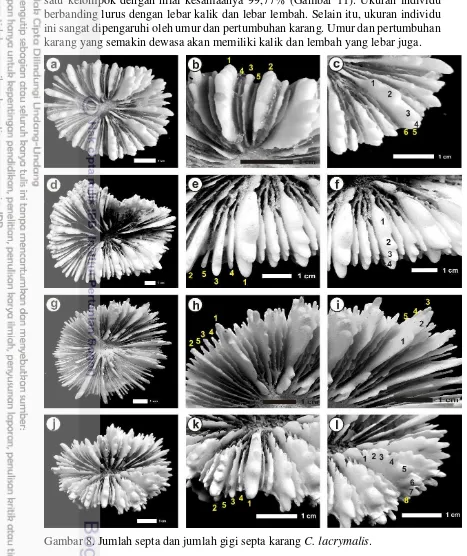
Gambar 9. Jumlah septa dan jumlah gigi septa karang C. lacrymalis
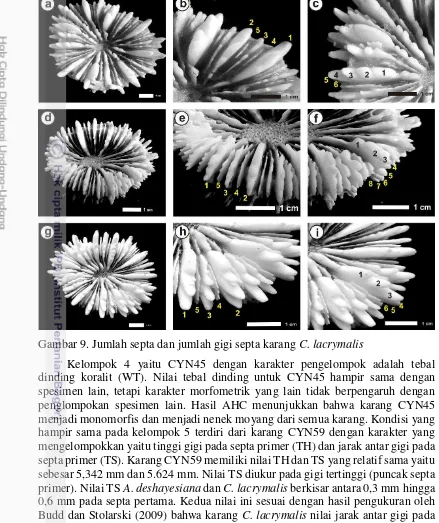
Kelompok 4 yaitu CYN45 dengan karakter pengelompok adalah tebal dinding koralit (WT). Nilai tebal dinding untuk CYN45 hampir sama dengan spesimen lain, tetapi karakter morfometrik yang lain tidak berpengaruh dengan penglompokan spesimen lain. Hasil AHC menunjukkan bahwa karang CYN45 menjadi monomorfis dan menjadi nenek moyang dari semua karang. Kondisi yang hampir sama pada kelompok 5 terdiri dari karang CYN59 dengan karakter yang mengelompokkan yaitu tinggi gigi pada septa primer (TH) dan jarak antar gigi pada septa primer (TS). Karang CYN59 memiliki nilai TH dan TS yang relatif sama yaitu sebesar 5,342 mm dan 5.624 mm. Nilai TS diukur pada gigi tertinggi (puncak septa primer). Nilai TS A. deshayesiana dan C. lacrymalis berkisar antara 0,3 mm hingga 0,6 mm pada septa pertama. Kedua nilai ini sesuai dengan hasil pengukuran oleh Budd dan Stolarski (2009) bahwa karang C. lacrymalis nilai jarak antar gigi pada septa < 6 mm. Hasil AHC membuktikan bahwa CYN59 menjadi monofiletik dari kelompok 1 meskipun kelompok ini memiliki nenek moyang dari karang CYN45.
kekeruhan rendah selama 8 bulan (Maret – November 2013). Perubahan kembali menjadi Pocillopora damicornis setelah 7 bulan (November 2013 – Juni 2014) yang dibuktikan dengan morfologi karang Pocillopora damicornis muncul kembali percabangan kecil. Karang dapat memodifikasi morfologinya untuk mengatasi perubahan lingkungan dengan variasi antara habitat dari waktu ke waktu (Prada et al. 2008). Perubahan lingkungan yang kuat di laut (seperti cahaya, pola arus, transport sedimen), memungkinkan seluruh organisme laut untuk melakukan adaptasi terhadap habitat (Hilbish 1985, Doebeli dan Dieckmann 2003).
Gambar 11. Dendogram karang C. lacrymalis dan A. deshayesiana berdasarkan karakter morfometrik menggunakan Agglomerative Hierarchical Clustering (AHC)
Perbandingan dan Kladogram morfologi antara A. deshayesiana dengan C. lacrymalis
Hasil pengamatan dan pengkelasan pada karakter deskriptif menunjukkan beberapa perbedaan antara karang A. deshayesiana dengan C. lacrymalis seperti bentuk gigi septa, pali atau septa, dan dentasi pada costae, sedangkan untuk karakter morfometrik yaitu jumlah gigi pada septa primernya (Tabel 3 dan Lampiran 2). Jumlah gigi pada karang C. lacrymalis lebih sedikit dibandingkan dengan A. deshayesiana yaitu 4-8 gigi dan 6-2 gigi pada septa pertamanya.
Karakter deskriptif yang membedakan antara karang C. lacrymalis dan A. deshayesiana adalah bentuk gigi septa (ST), septa atau pali (SPL), dan gigi costae (CD). Bentuk gigi septa (ST) pada seluruh karang A. deshayesiana berbentuk besar dan runcing, sedangkan C. lacrymalis lebih halus dan kecil (Tabel 3 dan Gambar 5). Gigi septa setiap genus hingga famili karang berbeda, hal ini sesuai pada penelitian untuk mencari karakter morfometrik karang Famili Favidae yang berbeda dan membingungkan (Budd dan Stolarski 2011). Perbedaan gigi septa juga ditemukan pada Famili Faviidae antara Atlantik dan Pasifik (Budd dan Stolarski 2009, Budd dan Stolarski 2011), sehingga sangat dimungkinkan lokasi yang berbeda mampu memberikan perbedaan pada morfologi karang.
Keberadaan pali atau septa (SPL) juga membedakan antara karang C. lacrymalis dengan A. deshayesaiana. Karang A. deshayesiana memiliki pali yang berkembang menjadi septa, sedangkan C. lacrymalis sangat jelas berbentuk pali. Budd dan Stolarski (2009) menunjukkan bahwa septa dan pali pada karang C. lacrymalis berkembang dengan baik. Secara umum, Famili Mussidae tidak memiliki pali seperti jenis Scolymia cubensis, Scolymia vitiensi, Favia speciosa,
dan Favia cf.lizardensis, akan tetapi terdapat pengecualian pada jenis C. lacrymalis
(Budd dan Stolarski 2009, Kongjandtre et al. 2012). Karakter yang membedakan antara C. lacrymalis dengan A. deshayesiana adalah dentasi/gigi pada costae (CD). CD sangat jelas membedakan antara A. deshayesiana dengan C. lacrymalis, yaitu bentuk gigi costae pada A. deshayesiana lebih halus dan kasar, sedangkan pada C. lacrymalis tidak memiliki gigi pada costae. Gigi costea pada karang jenis C. lacrymalis lebih membentuk tonjolan-tonjalan kecil (granule).
Tabel 3. Perbandingan karakter morfometrik dan deskriptif karang C. lacrymalis
dan A. deshayesiana
Karakter C. lacrymalis A. deshayesiana
Karakte
Calice Width Very Large, 26-45
mm
Very Large, 42-62 mm
Valley Width Medium, 5-9 mm Medium, 6-9 mm
Calice Relief Very high, 14-23
mm Overall Wall Thickness Thick, 4-8 mm Thick, 5-9 mm Individual Size Large, 29-52 mm Large, 48-74 mm
Karakte
r De
sk
rip
tif
Corallite Shape Solitary Solitary
Septal Teeth Large Acute or Small
Budding Type None None
Continuity of Costae Continous Continous
Septal and Paliform Lobes Pali Sometimes Septal
Columella Structure Styliform Styliform
Wall Structure Synapticulothecal Synapticulothecal
Costae Pattern Equal Equal
Costae Dentation Short Fine
Costae Alignment Present Present
Coenesteum Smooth Smooth
Kladogram morfologi ini berdasarkan pengkelasan pada karakter morfometrik dan deskriptif menggunakan metode Unweighted Pair Group Method with Arithmetic Mean (UPGMA) dengan model neighbour-joining. Kladogram morfologi membagi dua kelompok antara A. deshayesiana dengan C. lacrymalis (Gambar 12).
Berdasarkan kladogram morfologi, spesies A. deshayesiana dan C. lacrymalis
Lobophyllidae berdasarkan analisis molekuler dari marka cyt-B and CO-I. Hasil yang berbeda dilaporkan bahwa kelompok XIX yang didalamnya terdapat spesies
C. lacrymalis bukan lagi menjadi monofiletik, tetapi parafiletik karena dimasukkannya spesies Homophyllia australis dan menjadi saudara dari kelompok XXI (Budd et al. 2012). Perdebatan sering terjadi tentang monofiletik maupun parafiletik dari sebuah Famili jika dibandingkan dengan Famili yang lain. Budd et al. (2010) melaporkan 8 dari 24 Famili menggambarkan bahwa famili ini kuat (Robust) dan komplek (Complex), setiap famili yang monofiletik dapat masuk dan menggambarkan famili lain di kelompok yang berbeda. Penelitian yang dilakukan oleh Fukami et al. (2008) menunujukkan bahwa beberapa genus bukan monofiletik (Benzoni et al. 2007, Huang et al. 2009).
Gambar 12. Kladogram morfologi menggunakan metode UPGMA Analisis Molekuler
Hasil amplifikasi PCR karang C. lacrymalis dan A. deshayesiana yang dielektroforesis terlihat bersih yang menunjukkan bahwa gen ITS termaplifikasi dengan baik. Panjang gen target yang teramplifikasi ± 710 bp (Gambar 13). Selanjutnya dilakukan sekuensing dengan jumlah masing-masing sampel 30 µl. Hasil sekuensing berupa kromatogram yang terdiri dari sekuens forward dan
reverse masing-masing individu. Hasil pengeditan dan pengabungan sekuens masing-masing individu memiliki panjang basa nukliotida menjadi ± 686 bp.
(LK022368.1 dan LK022375.1); dan spesies Australomussa rowleyensis
(LK022359.1).
Karakteristik data sekuen karang dapat dilihat dari standarisasi dan koreksi dengan data Bank Gen di NCBI (National Center for Biotechnology Information). Standarisasi digunakan untuk mebuktikan apakah sekuen yang dimiliki berdekatan dengan data Bank Gen dan untuk mengetahui tingkat identifikasi nilai persentase kesamaannya. Berdasarkan hasil BLAST (Basic Local Alignmet Search Tool), menunjukkan perbedaan dan kesamaan pada beberapa sekuens dari masing-masing individu (Tabel 4). Kesamaan sekuen dari hasil blast terjadi pada karang C. lacrymalis dengan nilai kesamaan identifikasi pada karang C. lacrymalis terhadap data Gen Bank berkisar antara 96%-98% dengan query cover berkisar 93%-96%. Hasil yang berbeda dietmukan pada hasil blast karang A. deshayesiana, dimana nilai kesamaan identifikasinya mendekati karang Parascolymia vitiensis,
Austolomussa rowleyensis, dan Symphyllia valenciennesii. Hal ini dapat disebabkan ketersediaan data pada Bank Gen dan kedekatan terhadap ketiga jenis karang tersebut. Nilai identifikasi dari karang A. deshayesiana berkisar antara 96%-97% dengan query cover berkisar dari 97%-100%. Jenis karang yang memilki nilai kesamaan (Austrolomussa rowleyensis, Parascolymia viteinsis, dan Symphyllia valenciennesii) dilakukan pengunduhan untuk melihat kedekatannya dari hasil analisis filogenetik.
Gambar 13. Hasil elektroforesis amplifikasi PCR Tabel 4. Hasil blast basa nukliotida pada Gen Bank
No. Sampel Description Query Cover Ident
1 ACT_3 Australomussa rowleyensis genomic
DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate Z65785 (LK022359.1)
No. Sampel Description Query Cover Ident Parascolymia vitiensis genomic DNA
containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate HS2984 (LK022368.1)
97% 96%
2 ACT_11 Australomussa rowleyensis genomic
DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate Z65785 (LK022359.1)
100% 97%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate HS2984 (LK022368.1)
100% 96%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate PFB054 (LK022375.1)
100% 96%
3 ACT_7 Australomussa rowleyensis genomic
DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate Z65785 (LK022359.1)
100% 97%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate HS2984 (LK022368.1)
100% 97%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate PFB054 (LK022375.1)
100% 97%
4 ACT_10 Australomussa rowleyensis genomic
DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate Z65785 (LK022359.1)
100% 96%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate HS2984 (LK022368.1)
100% 96%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate PFB054 (LK022375.1)
100% 96%
5 ACT_12 Parascolymia vitiensis genomic DNA
containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate HS2984 (LK022368.1)
No. Sampel Description Query Cover Ident Australomussa rowleyensis genomic
DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate Z65785 (LK022359.1)
99% 97%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate PFB054 (LK022375.1)
99% 97%
6 ACT_19 Australomussa rowleyensis genomic
DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate Z65785 (LK022359.1)
100% 96%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate HS2984 (LK022368.1)
100% 96%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate PFB054 (LK022375.1)
100% 96%
7 ACT_21 Australomussa rowleyensis genomic
DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate Z65785 (LK022359.1)
98% 97%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate HS2984 (LK022368.1)
98% 97%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate PFB054 (LK022375.1)
98% 97%
8 ACT_1_ Australomussa rowleyensis genomic
DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate Z65785 (LK022359.1)
99% 96%
Parascolymia vitiensis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate HS2984 (LK022368.1)
99% 96%
Parascolymia vitiensis genomic DNA
No. Sampel Description Query Cover Ident
5.8S rRNA gene, ITS2 and 28S rRNA gene, isolate PFB054 (LK022375.1)
9 CYN_40
Cynarina lacrymalis genomic DNA containing partial 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and partial 28S rRNA gene, isolate NC674 (HE648552.1)
96% 97%
10 CYN_41
Cynarina lacrymalis genomic DNA containing partial 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and partial 28S rRNA gene, isolate NC674 (HE648552.1)
96% 97%
11 CYN_45
Cynarina lacrymalis genomic DNA containing 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2, 28S rRNA gene, strain MY011 (HF954288.1)
93% 98%
12 CYN_50
Cynarina lacrymalis genomic DNA containing partial 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and partial 28S rRNA gene, isolate NC674 (HE648552.1)
96% 97%
13 CYN_54
Cynarina lacrymalis genomic DNA containing partial 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and partial 28S rRNA gene, isolate NC674 (HE648552.1)
96% 97%
14 CYN_56
Cynarina lacrymalis genomic DNA containing partial 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and partial 28S rRNA gene, isolate NC674 (HE648552.1)
96% 97%
15 CYN_59
Cynarina lacrymalis genomic DNA containing partial 18S rRNA gene, ITS1, 5.8S rRNA gene, ITS2 and partial 28S rRNA gene, isolate NC674 (HE648552.1)
96% 96%
Keterangan: ACT (Acanthophyillia deshayesiana); CYN (Cynarina lacrymalis)
Variasai nukliotida pada karang A. deshayesiana (sampel), C. lacrymalis
(sampel dan Bank Gen), Parascolymia vitiensis, Austrolomussa rowleyensis, dan
CYN_45 T G G A G T C G G A T C A C G T C CYN_50 T G G A G T C G G A T C A C G T C CYN_54 T G G A G T C G G A T C A G G T C CYN_56 T G G A G T C G G A T C A G G T C CYN_59 T G G A G T C G G A T C A G G T C Hasil pengamatan dari variasi nukliotida di atas dapat diketahui perbedaan antara karang A. deshayesiana dengan C. lacrymalis berdasarkan nukliotida spesifik pada kode batang setiap individu dari masing-masing spesies. Secara umum, variasi nukliotida antara A. deshyesiana dengan C. lacrymalis dari sampel penelitian ini yaitu 87 situs, di mana terdapat 46 situs nukliotida spesifik yang sangat membedakan kedaunya (Tabel 5). Situs yang sangat membedakan antara A. deshyesiana dengan C. lacrymalis yaitu pada situs ke- 653 dan 654, di mana pada
A. deshayesiana terjadi delesi sehingga tidak memiliki kode batang (nukliotida), sedangkan C. lacrymalis memiliki kode batang pada kedua situs tersebut.
Tabel 6. Variasi situs nukliotida spesifik karang A. deshayesiana
Sampel
Tabel 7. Variasi situs nukliotida spesifik karang C. lacrymalis
moyang untuk individu pada sepesies yang sama dari sampel penelitian ini. Nenek moyang ini dapat dilihat dari nukliotida spesifik yaitu terdapat 12 situs (nukliotida ke-128, 154, 184, 216, 220, 539, 549, 661, 653, 654, 668, dan 671) (Tabel 7). Situs nukliotida ke-653 dan 654 merupakan variasi nukliotida spesifik pada spesies C. lacrymalis dari Bank Gen tidak memiliki kode batang dibandingkan C. lacrymalis
dari sampel penelitian ini.
Sekuen dari marka genetik ITS digunakan untuk analisis filogenetik dan menghasilkan pohon kekerabatan. Secara umum, metode yang digunakan untuk mendapatkan pohon kekerabatan dan jarak genetik (p-distance) yaitu Neighbour Joining dengan model kimura 2-parameter. Hasil analisis filogenetik menunjukkan bahwa kekerabatan antar individu dan spesies dapat dilihat pada Gambar 14. Pohon filogeni menunjukkan bahwa terbagi menjadi 2 kelompok besar. Kelompok 1 terdiri dari karang jenis A. deshayesiana, Parascolymia vitiensis, Austrolomussa rowleyensis, dan Symphyllia valenciennesii. Kelompok 2 terdiri dari karang hanya jenis C. lacrymalis Gen Bank dan C. lacrymalis dari sampel penelitian ini.
Kelompok I dibedakan lagi menjadi 2 kelompok kecil yaitu kelompok I.A (ACT10, ACT19, ACT12, ACT11, ACT7, dan ACT3) dan I.B (Symphyllia valenciennesii, Austrolomussa rowleyensis, dan Parascolymia vitiensis). Kelompok I.A dibedakan menjadi 2 kelompok yaitu kelompok 1.A.1 (ACT10 dan ACT19), I.A.2 (ACT12), I.A.3 (ACT11 dan ACT7), dan I.A.4 (ACT3). Kelompok I.B dibedakan kembali menjadi 2 kelompok yaitu I.B.1 (Symphyllia valenciennesii), dan I.B.2 (Autrolomussa rowleyensis, dan Parascolymia vitiensis). Kelompok II juga dibedakan lagi menjadi 2 kelompok kecil yaitu kelompok II.A yang terdiri dari karang C. lacrymalis (HE648552.1)Gen Bank , dan kelompok II.B C. lacrymalis
(HF954288.1)sampel penelitian ini (CYN40, CYN41, CYN45, CYN50, CYN59, CYN54, dan CYN56). Kelompok II.B dibagi kembali menjadi kelompok II.B.1 (C. lacrymalis (HF954288.1) Gen Bank, kelompok II.B.2 (CYN40, CYN41), Kelompok II.B.3 (CYN45), kelompok II.B.4 (CYN50 dan CYN59), dan II.B.5 (CYN54, dan CYN56).
Pohon filogeni menunjukkan bahwa karang A. deshayesiana tidak berada dalam satu cabang (kelompok) dengan karang C. lacrymalis. Hal ini menunjukkan bahwa karang C. lacrymalis tidak berkerabat dengan A. deshayesiana dengan perbedaan jarak genetik sebesar 6,3±4.3%. Secara umum, dalam penentuan spesies yang sama memiliki ambang batas jarak genetik sebesar ≤ 3% (Hebert et al. 2003, Barrett dan Hebert 2005), tetapi terjadi tumpang tindih pada karang, sehingga jarak genetik antar spesies dapat melebihi ambang batas. (Benzoni et al. (2012)) melaporkan bahwa jarak genetik antara Craterastrea levis dengan Leptoseris foliosa adalah 17,8 ± 1,4% karena berbeda Famili. Jarak genetik antara intrafamili
Craterastrea levis dengan taksa lainnya di kelompok famili yang sama (Famili Coscinaraeidae), yaitu 2,3 ± 0,4%.
sampel yang diunduh dari Bank Gen pada spesies S. valenciennesii, A. rowleyensis,
dan P. vitiensis memiliki nilai jarak genetik berkisar 0,1%-1,2%. Kelompok dari individu spesies A. deshayesiana mmemiliki nilai jarak genetik diatas ambang batas dengan kelompok individu dari Bank Gen (S. valenciennesii, A. rowleyensis, dan
P. vitiensis) yaitu 3,6 ± 0.6%. Dengan demikian, spesies A. deshayesiana tidak dapat dikatakan spesies yang sama dengan S. valenciennesii, A. rowleyensis, dan P. vitiensis tetapi berkerabat dekat dalam intra-Famili.
Gambar 14. Hasil analisis data molekuler menggunakan Neighbour Joining model Kimura 2-parameter 1000 bootstrap
Jarak genetik intraspesies A. deshayesiana sangat beragam, tetapi masih berada di bawah ambang batas. Jarak genetik antara ACT10 dengan ACT19 yaitu 1,8%, ACT12 dengan ACT10 dan ACT19 yaitu 2,3 ± 0.4%, ACT11 dengan ACT7 yaitu 0,3%, dan ACT3 dengan ACT11, ACT7, ACT10, ACT19, dan ACT12 yaitu 2.1 ± 0.8%. Kondisi yang tidak jauh berbeda juga ditemukan pada kelompok spesies
Jarak genetik spesies yang diunduh dari Bank Gen antara S. valenciennesii
(HF954347.1) dengan S. valenciennesii (HF954349.1) yaitu 0,2%, P. vitiensis
(LK022368.1) dengan P. vitiensis (LK022375.1) yaitu 0,2%, A. rowleyensis
(LK022359) dengan P. vitiensis (LK022368.1 dan LK022375.1) yaitu 0,1%. Jarak genetik antara spesies S. valenciennesii (HF954347.1 dan HF954349.1) dengan A. rowleyensis dan P. vitiensis (LK022359.1, LK022368.1 dan LK022375.1) yaitu 1.2 ± 0.9%, sehingga mereka dapat dikatakan berkerabat dekat dalam intra-Famili. Karang merupakan hewan spesial yang mampu melakukan perkawinan dengan spesies apa saja dan selalu terjadi tumpang tindih jarak genetik.
Implikasi Terhadap Taksonomi
Implikasi terhadap taksonomi pada penelitian ini sangat penting. Morfologi antara karang A. deshayesiana dengan C. lacrymalis berebeda. Morfologi kerangka yang membedakan kedua jenis tersebut terletak pada gigi septa, pali, gigi costae, dan jumlah septa pada septa primer. Karakter tersebut merupakan karakter baru yang dimunculkan pada penelitian ini dan berbeda dengan hasil penelitian dari Best dan Hoeksema (1987) bahwa karang A. deshayesiana dan C. lacrymalis sama pada diameter koralit, tinggi koralit, dentasi gigi septa, epitecha berkembang dengan baik. Hasil penelitian ini mempertegas perbedaan kedua jenis ini pada pali/epitecha bahwa A. deshayesiana tidak memiliki pali, sedangkan pali pada C. lacrymalis
berkembang dengan baik. Namun demikian, sejarah taksonomi kedua jenis ini sudah dimulai ketika Wells (1968) membedakan karang soliter pada Famili Mussidae dengan kunci identifikasi pada tingkat genus. Kunci identifikasi Genus
Cynarina berdasarkan pada septa primer yang tidak memiliki dentasi, sedangkan Genus Acantophyllia pada dentasi septa sangat lebar melebihi dinding kalik. Best dan Hoeksema (1987) melaporkan kesamaan kedua jenis ini dan hingga sekarang masih digunakan dalam taksonomi karang, tetapi hasil dari penelitian ini memperkuat kembali perbedaan kedua karang ini yang dilaporkan oleh Wells (1968). Kladogram morfologi juga memperkuat perbedaan kedua jenis ini (Gambar 12).
Perbedaan morfologi antara jenis pada karang memiliki keraguan karena termasuk kategori identifikasi secara klasik (Fukami et al. 2004, Benzoni et al. 2007, Todd 2008, Huang et al. 2011, Schmidt-Roach et al. 2012, Richards et al. 2013, Reijnen et al. 2014). Analisis molekuler digunakan untuk melihat konsistensi karater morfologi yang memedakan pada kladogram. Hasil analisis filogenetik mendukung perbedaan dari A. deshayesiana dengan C. lacrymalis. Pohon filogeni menunjukkan perbedaan yang jauh dengan jarak genetik 6,3 ± 4,3%. Arrigoni et al.
(2014a) menunjukkan bahwa berdasarkan pohon filogeni C. lacrymalis merupakan monofiletik dan berkerabat dekat dengan karang soliter lainnya seperti
Parascolymia vitiensis, Austrolomussa rowleyensis, dan Symphyllia valenceinnesii. Hasil pohon filogeni penelitian ini menunjukkan bahwa A. deshayesiana lebih berkerabat dekat dengan P. vitiensis, A. rowleyensis, dan S. valenciennesii karena berada dalam satu kluster dengan jarak genetik 3,6 ± 0,6%, sedangkan jarak genetik
Karang jenis A. deshayesiana dapat dimunculkan kembali menjadi jenis karang yang memiliki penciri karakter morfologi dan molekuler secara mandiri.
SIMPULAN DAN SARAN
Pengelompokan berdasarkan karakter morfometrik masih tidak jelas, sedangkan karakter deskriptif, dan molekuler sudah dapat membedakan pengelompokan karang C. lacrymalis dan A. deshayesiana secara terpisah. Secara genetik, kedua spesies tersebut memiliki keragaman intraspesies. Karakter deskriptif yang membedakan kedua jenis karang tersebut dilakukan analisis kembali untuk mebandingkan hasil pohon filogeninya, sehingga dapat ditemukan apakah karakter deskriptif menjadi karakter kunci pembeda keduanya.
DAFTAR PUSTAKA
Arrigoni R, Stefani F, Pichon M, Galli P, Benzoni F. 2012. Molecular phylogeny of robust clade (Faviidae, Mussidae, Merulinidae, and Pectiniidae): An Indian Ocean perspective. Mol. Phyl. and Evol. 65(1): 183-193.doi: 10.1016/j.ympev.2012.06.001.
Arrigoni R, Richards ZT, Chen CA, Budd AH, Benzoni F. 2014a. Taxonomy and phylogenetic relationships of the coral genera Autralomussa and
Parascolymia (Scleractiia, Lobophylliidae). Contrib. to Zoo. 83(3): 195-215.doi: 10.1016/j.ympev.2014.01.010.
Arrigoni R, Terraneo TI, Galli P, Benzoni F. 2014b. Lobophylliidae (Cnidaria, Scleractinia) reshuffled: pervasive non-monophyly at genus level. Mol. Phyl. and Evol. 7360-64.doi: 10.1016/j.ympev.2014.01.010.
Arrigoni R, Berumen ML, Terraneo TI, Caragnan A, Bouwmeester J, Benzoni F. 2015. Forgotten in the taxonomic literature: resurrection of the scleractinian coral genus Sclerophyllia (Scleractinia, Lobophylliidae) from the Arabian Peninsula and its phylogenetic relationships. Sys. and Biodiv. 13(2): 140-163.10.1080/14772000.2014.978915.
Barrett RDH, Hebert PDN. 2005. Identifying spiders through DNA barcodes.
Canad. J of Zoo. 83481–491
Benzoni F, Stefani F, Stolarski J, Pichon M, Mitta G, Galli P. 2007. Debating phylogenetic relationships of the scleractinian Psammocora: molecular and morphological evidences. Contrib. to Zoo. 76( 1): 35-54
Benzoni F, Arrigoni R, Stefani F, Stolarski J. 2012. Systematics of the coral genus
Craterastrea (Cnidaria, Anthozoa, Scleractinia) and description of a new family through combined morphological and molecular analyses. Sys. and Biodiv. 10(4): 417-433.10.1080/1472000.2012.744369.
Best MB, Hoeksema BW. 1987. New Observations on scleractinian coral from Indonesia: 1. Free-living species belonging to the Faviina. Zoo. Meded.
Budd AF, Stolarski J. 2009. Searching for new morphological characters in the systematics of scleractinian reef corals: comparison of septal teeth and granules between Atlantic and Pacific Mussidae. Acta Zoo. 90(2): 142-165.doi: 10.1111/j.1463-6395.2008.00345.x.
Budd AF, Romano SL, Smith ND, Barbeitos MS. 2010. Rethinking the Phylogeny of Scleractinian Corals: A Review of Morphological and Molecular Data.
Integ. and Comp. Biol. 50(3): 411-427
Budd AF, Stolarski J. 2011. Corallite Wall and Septal Microstructure in Scleractinian Reef Corals: Comparison of Molecular Clades Within the Family Faviidae. J of Morph. 272(1): 66-88.doi : 10.1002/jmor.10899. Budd AF, Fukami H, Smith ND, Knowlton N. 2012. Taxonomic classification of
the reef coral family Mussidae (Cnidaria: Anthozoa: Scleractinia). Zoo. J. of the Linn. Soc. of Lon. 166(3): 465-529.doi: 10.1111/j.1096-3642.2012.00855.x.
Casebolt SN. 2011. Phylogenetic analysis and quantitative assessment of micromorphology and microstructure in the coral family Mussidae (Scleractinia). Desertation, University of Iowa.
Doebeli M, Dieckmann U. 2003. Speciation along environmental gradients. Nature.
421(6920): 259-264.doi: 10.1038/nature01274.
Filatov MV, Frade PR, Bak RPM, A.Vermeij MJ, Kaandorp JA. 2013. Comparison between colony morphology and molecular phylogeny in the Carribean Scleractinian Coral Genus Madracis. PLoS ONE. 8(8): e71287.10.371/journal.pone.0071287.
Flot J-F, Magalon H, Cruaud C, Couloux A, Tiller S. 2008. Patterns of genetic structure among Hawaiian corals of the genus Pocillopora yield clusters of individuals that are compatible with morphology. Compt. Rend. Biol.
331(3): 239-247.doi: 10.1016/j.crvi.2007.12.003.
Fukami H, Budd AF, Paulay G, ´-Cava AS, Chen CA, Iwao K, Knowlton N. 2004. Conventional taxonomy obscures deep divergence between Pacific and Atlantic corals. Nature. 427832–835
Fukami H, Chen CA, Budd AF, Collins A, Wallace C, Chuang TY-Y, Chen C, Dai C-F, Iwao K, Sheppard C, Knowlton N. 2008. Mitochondria and nuclear genes suggest that stony corals are monophyletic but most Families of stony corals are not (Order Scleractinia, Class Anthozoa, Phylum Cnidaria). PLoS ONE 3(9): e3222.10.1371/journal.pone.0003222.
Giyanto A. 2007. Perdagangan karang hias: suatu ancaman terhadap ekosistem terumbu karang. Oseana. 4(32): 21-27
Haerul A. 2014. Karakterisasi Genetik Karang Genus Favites(Faviidae: Scleractinia) Di Perairan Kepulauan Spermonde, Sulawesi Selatan Tesis, Institut Pertanian Bogor.
Hardja JW. 2009. Karakterisasi Penanda Genetik mtDna COI Dan Daerah ITS rDna Karang Goniopora spp. (Cnidaria: Scleractinia) Dalam Upaya Pengelolaan Terumbu Karang Di Perairan Pulau Pramuka, Kepulauan Seribu. Tesis, Institut Pertanian Bogor.
Hilbish TJ. 1985. Demographic and temporal structure of an allele frequency cline in the mussel Mytilus edulis. Mar. Biol. 86(2): 163-171.doi : 10.1007/BF00399023.
Huang D, Meier R, Todd PA, Chou LM. 2009. More evidence for pervasive paraphyly in scleractinian corals: systematic study of Southeast Asian Faviidae (Cnidaria; Scleractinia) based on molecular and morphological data. Mol. Phyl. and Evol. 50(1): 102-116.doi: 10.1016/j.ympev.2008.10.012.
Huang D, Licuanan WY, Bird AH, Fukami H. 2011. Cleaning up the‘Bigmessidae’: Molecular phylogeny of scleractinian corals from Faviidae, Merulinidae, Pectiniidae and Trachyphylliidae. BMC Evol. Biol. 11(37): 1-13.10.1186/1471-2148-11-37.
Huang D, Benzoni F, Fukami H, Knowlton N, Smith ND, Budd AF. 2014. Taxonomic classification of the reef coral families Merulinidae, Montastraeidae, and Diploastraeidae (Cnidaria: Anthozoa: Scleractinia).
Zoo. J of the Linn. Soc. 171(2): 277-355.doi: 10.1111/zoj.12140.
Kitano YF, Benzoni F, Arrigoni R, Shirayama Y, Wallace CC, Fukami H. 2014. A phylogeny of the family Poritidae (Cnidaria, Scleractinia) based on molecular and morphological analyses. PLoS ONE. 9(S): e98406.doi: 10.1371/journal.pone.0098406.
Kongjandtre N, Ridgway T, Cook LG, Huelsken T, Budd AF, Hoegh-Guldberg O. 2012. Taxonomy and species boundaries in the coral genus Favia Milne Edwards and Haime, 1857 (Cnidaria: Scleractinia) from Thailand revealed by morphological and genetic data. Cor. Reefs. 31(2): 581–601.doi: 10.1007/s00338-011-0869-5.
Oppen MJHV, Willis BL, Vugt HWJAV, Miller DJ. 2000. Examination of species boundaries in the Acropora cervicornis group (Scleractinia, Cnidaria) using molecular DNA sequence analyses. Mol. Ecol. Resources. 9(9): 1363-1373.doi: 10.1046/j.1365-294x.2000.01010.x.
Paz-García DA, Hellberg ME, García-de-León FJ, Balart EF. 2015. Switch between Morphospecies of Pocillopora Corals. The Amer. Natur. 186(3): 434-440.doi: 10.1086/682363.
Prada C, Schizas NV, Yoshioka PM. 2008. Phenotypic plasticity or speciation? A case from a clonal marine organism. BMC Evol. Biol. 8(47): 10.1186/1471-2148-8-47.doi: 10.1186/1471-2148-8-47.
Reijnen BT, McFadden CS, Hermanlimianto YT, Ofwegen LPv. 2014. A molecular and morphological exploration of the genetic boundaries in the family Melithaeidae. Mol. Phyl. and Evol. 70383-401
Richards ZT, Miller DJ, Wallace CC. 2013. Molecular phylogenetics of geographically restricted Acropora species: Implication for threatened species conservation. Mol. Phyl. and Evol. 69837-851
Schmidt-Roach S, Lundragen P, Miller KJ, Gerlach G, Noreen AME, Andreakis N. 2012. Assesing hidden species diversity in the coral Pocillopora damicornis
from Eastern Australia. Cor. Reefs. 32(1): 161-172.doi: 10.1007/s00338-012-0959-z.
(Cnidaria; Scleractinia). Hydrobiologia. 596(1): 153-172.doi: 10.1007/s10750-007-9092-3.
Stobart B. 2000. A taxonomic reappraisal of Montipora digitata based on genetic and morphometric evidence. Zoo. stud. 39(3): 179-190
Suharsono 2010. Jenis-jenis Karang di Indonesia Jakarta (ID), Puslitbang Oseanologi-LIPI.
Swofford DL 2002. PAUP*. Phylogenetic Analysis Using Parsimony (*and Other Methods). Version 4, Sunderland, Massachusetts, Sinauer Associates. Todd PA. 2008. Morphological plasticity in scleractinian corals. Bio. Revie. 83315–
337.10.1111/j.1469-185X.2008.00045.x.
Veron J 2000. Corals Of The World, Australian (AUS), Australian Institute of Marine Science, PMB 3, Townsville MC.
Wells JW. 1937. Coral Studies. I Two new species of fossil corals. II Favie new genera of the Madreporaria. Bull. of Am. Paleo. 23(79): 238-250
Wells JW. 1968. The recent solitary mussid scleractinian corals. Zoo. Meded.
39(38): 375-384
Wolstenholme JK, Wallace CC, Chen CA. 2003. Species boundaries within the
Lampiran 1. Karakter Morfometrik Karang Cynarina lacrymalis dan
Acanthophyllia deshayesiana
SAMPEL
CW (mm)
VW (mm)
CR
(mm) NS SS
TH (mm)
TS
(mm) NT
WT (mm)
IS (mm)
ACT3 52.260 8.710 21.390 2.000 3.000 4.518 4.580 9.000 6.200 58.350 ACT11 46.670 8.080 16.930 4.000 4.000 4.916 4.416 6.000 8.230 57.080 ACT7 46.310 8.070 19.130 4.000 3.000 3.216 3.962 9.000 4.330 51.720 ACT10 42.300 6.150 22.380 4.000 3.000 4.354 4.192 7.000 6.750 47.890 ACT12 54.350 9.290 25.020 4.000 3.000 5.840 6.054 8.000 5.040 73.650 ACT19 55.440 7.740 22.070 4.000 3.000 5.470 4.462 11.000 7.500 59.130 ACT21 62.070 8.330 24.220 4.000 3.000 5.138 4.770 12.000 7.150 72.450 ACT1 45.000 8.920 20.330 4.000 3.000 3.584 5.302 8.000 9.170 53.710 CYN50 38.450 4.990 14.120 5.000 4.000 2.466 2.802 6.000 5.140 39.940 CYN54 46.820 6.250 14.960 5.000 4.000 4.206 3.528 5.000 7.610 48.310 CYN56 24.750 5.870 13.990 4.000 4.000 1.906 2.274 6.000 6.010 31.470 CYN40 44.560 7.040 22.650 3.000 3.000 4.712 5.518 4.000 8.440 52.470 CYN41 43.580 9.340 18.800 4.000 4.000 3.702 3.576 4.000 3.960 48.780 CYN45 26.220 4.540 16.580 4.000 4.000 3.210 4.150 7.000 4.790 29.310 CYN59 35.330 4.560 13.720 4.000 4.000 5.342 5.624 6.000 4.760 41.260 Keterangan :
Lampiran 3. Matriks jarak genetik
No. Sampel 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20
1 ACT_3_