ISOLASI DAN UJI ANTAGONIS Fusarim sp. PADA TANAMAN ANGGREK MENGGUNAKAN BAKTERI KITINOLITIK LOKAL
SKRIPSI
Diajukan untuk memenuhi syarat mencapai gelar Sarjan Sains
SITI MAIMUNAH 060805001
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM UNIVERSITAS SUMATERA UTARA
PERSETUJUAN
Judul : ISOLASI DAN UJI ANTAGONIS Fusarim sp. PADA TANAMAN ANGGREK MENGGUNAKAN BAKTERI KITINOLITIK LOKAL
Kategori : SKRIPSI
Nama : SITI MAIMUNAH
Nomor Induk Mahasiswa : 060805001
Program Studi : SARJANA (S1) BIOLOGI
Departemen : BIOLOGI
Fakultas : MATEMATIKA DAN ILMU PENGETAHUAN ALAM
(FMIPA) UNIVERSITAS SUMATERA UTARA
Diluluskan di Medan, Januari 2010 Komisi Pembimbing :
Pembimbing II Pembimbing I
Drs. Kiki Nurtjahja, M.Sc. Prof. Dr. Dwi Suryanto, M.Sc NIP.196212111998031001 NIP. 196404091994031003
Diketahui / Disetujui oleh
Departemen Biologi FMIPA USU
PERNYATAAN
ISOLASI DAN UJI ANTAGONIS Fusarim sp. PADA TANAMAN ANGGREK MENGGUNAKAN BAKTERI KITINOLITIK LOKAL
SKRIPSI
Saya mengakui bahwa skripsi ini adalah hasil kerja saya sendiri, kecuali beberapa kutipan dan ringkasan yang masing-masing disebutkan sumbernya.
Medan, Desember 2010
PENGHARGAAN
Puji dan syukur penulis panjatkan kepada Allah SWT yang Maha Pengasih lagi Maha Penyayang atas rahmat, kekuatan dan kemurahan-Nya sehingga penulis dimampukan menyelesaikan skripsi yang berjudul, ”ISOLASI DAN UJI ANTAGONIS FUNGI PATOGEN TANAMAN ANGGREK MENGGUNAKAN BAKTERI KITINOLITIK LOKAL” dalam waktu yang ditetapkan.
Ucapan terima kasih penulis sampaikan kepada Prof. Dr. Dwi Suryanto, M.Sc selaku Dosen Pembimbing I dan kepada Bapak Drs. Kiki Nurtjahja, M.Sc sebagai Dosen Pembimbing II yang telah banyak memberikan bimbingan dan arahan, waktu dan perhatiannya yang besar terutama saat memulai penulisan hingga penyusunan skripsi ini. Ucapan terima kasih juga saya sampaikan kepada Ibu Dra. Nunuk priyani, M.Sc dan Ibu Dra. Suci Rahayu, M.Sc sebagai Ketua dan Sekretaris Penguji yang telah banyak memberikan arahan dan saran dalam penyempurnaan penulisan skripsi ini.
Ucapan terima kasih juga saya tunjukkan kepada Bapak Prof. Dr. H. Erman Munir, M.Sc. selaku Dosen Akademik saya dan juga kepada Bapak Prof. Dr. Dwi Suryanto, M.Sc Ketua Deapartemen Biologi FMIPA USU dan Ibu Dra. Nunuk Priyani, M.Sc selaku Sekretaris Departemen Biologi FMIPA USU. Dr. Sutarman, M.Sc selaku Dekan FMIPA USU. Bapak Prof. Dr. H. Erman Munir, M.Sc. selaku kepala Laboratorium Mikrobiologi. Staf pengajar di Departemen Biologi FMIPA USU. Bapak Alm. Sukirmanto, Ibu Nurhasni Muluk, Ibu Roslina Ginting, dan Bang Erwin selaku Staf Pegawai Departemen BIOLOGI FMIPA USU.
Ungkapan terima kasih yang tidak ternilai juga penulis ucapkan kepada keluargaku yang saya sayangi, kedua orang tua (Ayahanda Supri dan Ibunda Arsih) yang sudah memberikan banyak dukungan baik dari segi moril maupun materiil sehingga penulis bisa menyelesaikan perkuliahan ini, dan kepada yang paling saya sayangi saudara kandung saya Abangda Mhd. Syafi’i, Mhd. Jupri dan Kakanda Siti Laila, Siti Zaleha, Siti Patimah serta kepada Adindaku Siti Sarah, Siti Hamidah, Siti Nurjanah, Nining dan Mhd Irmam penulis ucapkan ribuan terima kasih atas segala cinta, kasih sayang, pengorbanan moril maupun materil, motivasi, kesabaran serta do’a yang tidak akan pernah bisa penulis balas sampai kapanpun.
terlibat langsung maupun tidak langsung yang tidak dapat disebutkan satu persatu atas dukungan, perhatian, dan bantuannya kepada penulis.
Penulis menyadari bahwa skripsi ini masih jauh dari kesempurnaan. Oleh karena itu penulis mengharapkan kritik dan saran yang bersifat membangun dalam melengkapi kekurangan serta penyempurnaan laporan hasil penelitian ini. Akhir kata semoga skripsi ini bermanfaat bagi kita semua.
ISOLASI DAN UJI ANTAGONIS Fusarim sp. PADA TANAMAN ANGGREK MENGGUNAKAN BAKTERI KITINOLITIK LOKAL
SITI MAIMUNAH 060805001
ABSTRAK
Penelitian mengenai Isolasi dan Uji Antagonis Fusarium sp. pada Tanaman Anggrek Menggunakan Bakteri Kitinolitik Lokal, telah dilaksanakan di Laboratorium Mikrobiologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara, Medan dari Bulan Mei sampai Desember 2010. Penelitian ini bertujuan untuk mengetahui potensi bakteri kitinolitik yang berasal dari Bangka, Langkat dan Deli Serdang, hasil penelitian sebelumnya, dalam menghambat pertumbuhan Fusarium sp 1., yang terdapat pada tanaman anggrek Cattelya sp, Fusarium sp 2 yang terdapat pada tanaman anggrek Vanda sp., Fusarium sp 3., Fusarium sp 4., dan Fusarium sp 5., yang terdapat pada tanaman Phalaenopsis sp yang diisolasi dari Perkebunan anggrek di Tiga Dolok, Kabupaten Simalungun, Kecamatan Dolok Panribuan, dan untuk memastikan adanya fungi patogen dengan mencatat simptomnya. Isolasi dilakukan dengan teknik direct plating dengan meletakkan irisan akar, daun, dan batang sepanjang 3 cm dari tanaman anggrek yang sakit pada medium PDA steril yang telah ditambah kloramfenikol dan yeast ekstrak dalam cawan Petri steril, kemudian diinkubasi pada suhu ruangan selama 72 jam. Koloni-koloni diambil dan dikultur dengan menggunakan cawan Petri steril. Fungi yang diperoleh kemudian dibiakkan dalam media PDA miring dan disimpan untuk uji berikutnya. Identifikasi fungi yang diduga patogen dilakukan secara makroskopis dan mikroskopis. Hasil isolasi fungi, diambil gambarnya dan dideterminasi dengan menggunakan kunci determinasi. Hasil penelitian menunjukkan efektivitas paling tinggi dalam menghambat pertumbuhan Fusarium sp. 5, adalah isolat KM15, dengan jari-jari zona hambat sebesar 2,82 cm. Sedangkan efektifitas terendah ditunjukkan oleh isolat KM13, dengan jari-jari zona hambat sebesar 0,42 cm. Hifa abnormal dicirikan pada Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3, Fusarium sp. 4, dan Fusarium sp. 5, mengalami lisis, menggulung dan melilit setelah dilakukan uji antagonis.
ISOLATION AND THE ANTAGONISTIC TEST OF Fusarium sp. ISOLATED FROM SOME ORCHID PLANTS AGAINST LOCAL CHITINOLITIC
BACTERIA
SITI MAIMUNAH 060805001
ABSTRAK
An Assay On Isolation And The Antagonistic Test Of Fusarium sp. Isolated From Some Orchid Plants Against Local Chitinolitic Bacteria, has been conducted in Laboratory of Microbiology, Department of Biology, Faculty of Matematics and Natural Sciences, Sumatera Utara University, Medan from May to December 2010. The purpose of the research was to study the ability of bacteria isolated from Bangka, Langkat and Deli Serdang (from previous study) to inhibit the growth of Fusarium sp 1., isolased from Cattelya sp; Fusarium sp. 2 from Vanda sp.; Fusarium sp. 3, Fusarium sp. 4, and Fusarium sp. 5, from Phalaenopsis sp on isolation from orchid plantation Tiga Dolok, District Simalungun, Sub-District Dolok Panribuan, and the existence of fungal pathogens by noting the simptom. Isolation of pathogenic fungi was conducted by direct plating method placing 3 cm slices of diseased orchid of roots, leaves and stems on sterile PDA medium containing chloramphenicol and yeast extract. The isolates then were incubated at room temperature for 72 hours. Colonies were cultured on sterile Petri dishes to ensure the existence of fungal pathogens. The obtained fungi were isolated and identified using identifiction book of fungi. Based on the antaonistic test, it was shown that the most effective bacteria in inhibiting the growth of Fusarium sp. 5, was BK15, with inhibition zone radius of 2.82 cm. Whereas, the least effective bacteria was BK13, with inhibition zone radius of 0,42 cm.. And for abnormal hyphae Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3, Fusarium sp. 4, dan Fusarium sp. 5, in experienced lysis, rolled and twisted after the test antagonistic.
DAFTAR ISI Halaman Persetujuan Pernyataan Penghargaan Abstrak Daftar isi Daftar Gambar Daftar Lampiran ii iii iv v viii ix x
Bab 1 Pendahuluan 1
1.1 Latar Belakang 1
1.2 Permasalahan penelitian 5
1.3 Tujuan Penelitian 5
1.4 Hipotesis Penelitian 5
1.5 Manfaat Penelitian 5
Bab 2 Bahan dan Metode 6
2.1 Waktu dan Tempat 6
2.2 Alat dan Bahan 6
2.3 Isolat Bakteri dan Fungi 6
2.4 Isolasi dan Identifikasi Fungi Mikroskopikdari Tanaman Anggrek 7
2.5 Uji Patogenitas Dengan Bakteri Kitinolitik 7
2.6 Pengamatan Struktur Hifa Fungi Setelah Uji Patogenitas 8
Bab 3 Hasil dan Pembahasan 9
3.1 Karakteristik Fusarium sp., yang terdapat pada Tanaman Anggrek Cattelya sp, Vanda sp., dan Phalaenopsis sp. 9
3.2 Uji Antagonisme In Vitro Bakteri Kitinolitik dengan Fusarium sp., pada Tanaman Anggrek Jenis Cattelya sp., Vanda sp., dan Phalaenopsis sp . 12
3.3 Pengamatan Struktur Hifa Abnormal Fusarium sp., pada Tanaman Anggrek setelah Uji Patogenitas. 16
Bab 4 Kesimpulan dan Saran 18
4.1 Kesimpulan 18
4.2 Saran 18
Daftar Pustaka 19
DAFTAR GAMBAR
Halaman Gambar 1. Tiga genus anggrek yang terserang Fusarium sp. (a)
Cattelya sp. (b) Vanda sp. dan (c) Phalaenopsis sp.
9
Gambar 2. Hasil isolasi fungi patogen pada tanaman anggrek yang terkena penyakit, masing-masing berasal dari (a) Fusarium sp. 1, (C2) yang menyerang batang Cattelya sp., (b) Fusarium sp. 2, (V1) yang menyerang daun Van da sp., (c) Fusarium sp. 3, (P7) dan Fusarium sp. 4, (P5) yang menyerang daun Phalaenopsis sp., dan (d) Fusarium sp. 5, (P8) yang menyerang pada daun Phala enopsis sp. Dengan masa inkubasi 2 X 24 jam pada suhu 30oC.
10
Gambar 3. Biakan murni dan mikroskopik (a) Fusarium sp. 1, (b) Fusarium sp.2, (c) Fusarium sp. 3, (d) Fusarium sp. 4 (e) Fusarium sp. 5, pada media PDA suhu 30oC, beru mur 3 hari (perbesaran 400).
11
Gambar 4. Uji Antagonis Bakteri Kitinolitik (a) PP01 sp. 2, terha dap Fusarium sp. 4, dari Phalaenopsis sp., dan (b) LK 08 terhadap Fusarium sp. 1, pada Vanda sp., dengan masa inkubasi 10 hari pada suhu 30oC.
16
Gambar 5. Bentuk hifa abnormal Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3, Fusarium sp. 4, dan Fusarium sp. 5 (a) menggulung, (b) lisis, (c) menyatu, (d) melilit masa inkubasi 10 hari (Perbesaran 400).
17
DAFTAR LAMPIRAN
Halaman
Lampiran 1. Komposisi Medium 22
Lampiran 2. Alur Kerja Karakterisasi Sifat Morfologi Dan Biokimia Isolat
24 Lampiran 3. Isolasi dan Identifikasi Mikroskopis Jamur Patogen Tanaman Anggrek
25 Lampiran 4. Alur Kerja Asai Antagonime in vitro 26 Lampiran 5. Pembuatan Koloidal Kitin dengan Cara hidrolisis Parsial
(Rodriquez-Kabana et al, 1983).
ISOLASI DAN UJI ANTAGONIS Fusarim sp. PADA TANAMAN ANGGREK MENGGUNAKAN BAKTERI KITINOLITIK LOKAL
SITI MAIMUNAH 060805001
ABSTRAK
Penelitian mengenai Isolasi dan Uji Antagonis Fusarium sp. pada Tanaman Anggrek Menggunakan Bakteri Kitinolitik Lokal, telah dilaksanakan di Laboratorium Mikrobiologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara, Medan dari Bulan Mei sampai Desember 2010. Penelitian ini bertujuan untuk mengetahui potensi bakteri kitinolitik yang berasal dari Bangka, Langkat dan Deli Serdang, hasil penelitian sebelumnya, dalam menghambat pertumbuhan Fusarium sp 1., yang terdapat pada tanaman anggrek Cattelya sp, Fusarium sp 2 yang terdapat pada tanaman anggrek Vanda sp., Fusarium sp 3., Fusarium sp 4., dan Fusarium sp 5., yang terdapat pada tanaman Phalaenopsis sp yang diisolasi dari Perkebunan anggrek di Tiga Dolok, Kabupaten Simalungun, Kecamatan Dolok Panribuan, dan untuk memastikan adanya fungi patogen dengan mencatat simptomnya. Isolasi dilakukan dengan teknik direct plating dengan meletakkan irisan akar, daun, dan batang sepanjang 3 cm dari tanaman anggrek yang sakit pada medium PDA steril yang telah ditambah kloramfenikol dan yeast ekstrak dalam cawan Petri steril, kemudian diinkubasi pada suhu ruangan selama 72 jam. Koloni-koloni diambil dan dikultur dengan menggunakan cawan Petri steril. Fungi yang diperoleh kemudian dibiakkan dalam media PDA miring dan disimpan untuk uji berikutnya. Identifikasi fungi yang diduga patogen dilakukan secara makroskopis dan mikroskopis. Hasil isolasi fungi, diambil gambarnya dan dideterminasi dengan menggunakan kunci determinasi. Hasil penelitian menunjukkan efektivitas paling tinggi dalam menghambat pertumbuhan Fusarium sp. 5, adalah isolat KM15, dengan jari-jari zona hambat sebesar 2,82 cm. Sedangkan efektifitas terendah ditunjukkan oleh isolat KM13, dengan jari-jari zona hambat sebesar 0,42 cm. Hifa abnormal dicirikan pada Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3, Fusarium sp. 4, dan Fusarium sp. 5, mengalami lisis, menggulung dan melilit setelah dilakukan uji antagonis.
ISOLATION AND THE ANTAGONISTIC TEST OF Fusarium sp. ISOLATED FROM SOME ORCHID PLANTS AGAINST LOCAL CHITINOLITIC
BACTERIA
SITI MAIMUNAH 060805001
ABSTRAK
An Assay On Isolation And The Antagonistic Test Of Fusarium sp. Isolated From Some Orchid Plants Against Local Chitinolitic Bacteria, has been conducted in Laboratory of Microbiology, Department of Biology, Faculty of Matematics and Natural Sciences, Sumatera Utara University, Medan from May to December 2010. The purpose of the research was to study the ability of bacteria isolated from Bangka, Langkat and Deli Serdang (from previous study) to inhibit the growth of Fusarium sp 1., isolased from Cattelya sp; Fusarium sp. 2 from Vanda sp.; Fusarium sp. 3, Fusarium sp. 4, and Fusarium sp. 5, from Phalaenopsis sp on isolation from orchid plantation Tiga Dolok, District Simalungun, Sub-District Dolok Panribuan, and the existence of fungal pathogens by noting the simptom. Isolation of pathogenic fungi was conducted by direct plating method placing 3 cm slices of diseased orchid of roots, leaves and stems on sterile PDA medium containing chloramphenicol and yeast extract. The isolates then were incubated at room temperature for 72 hours. Colonies were cultured on sterile Petri dishes to ensure the existence of fungal pathogens. The obtained fungi were isolated and identified using identifiction book of fungi. Based on the antaonistic test, it was shown that the most effective bacteria in inhibiting the growth of Fusarium sp. 5, was BK15, with inhibition zone radius of 2.82 cm. Whereas, the least effective bacteria was BK13, with inhibition zone radius of 0,42 cm.. And for abnormal hyphae Fusarium sp. 1, Fusarium sp. 2, Fusarium sp. 3, Fusarium sp. 4, dan Fusarium sp. 5, in experienced lysis, rolled and twisted after the test antagonistic.
BAB 1
PENDAHULUAN
1.1Latar Belakang
Jenis-jenis flora yang ada di Indonesia masih banyak yang belum dimanfaatkan dan dimasyarakatkan. Eksplorasi dan inventarisasi untuk menyelamatkan plasma nutfah tanaman anggrek ini diterapkan agar tanaman anggrek tidak terancam punah. Tetapi permasalaham penyakit timbul ketika memelihara anggrek di rumah kaca atau di kebun (Munawaroh, 2001). Penyakit yang meninfeksi anggrek antara lain dapat diakibatkan oleh bakteri dan fungi. Kedua penyakit ini berbeda dari cara serangannya, gejalanya, penyebarannya, penanggulangannya, dan pencegahannya
Menurut Hasanuddin (2003) yang mengatakan bahwa, saat ini sudah menjadi satu pengetahuan bahwa pengendalian hayati memainkan peranan penting dalam bidang pertanian, karena penggunaan bahan kimia sebagai pestisida dapat menyebabkan toksik pada manusia, hewan dan tumbuhan. Kebiasaan para petani menyemprot pestisida secara sembarangan menyebabkan timbulnya strain baru dari fungi-fungi patogen pada tanaman, yang ditunjukkan adanya kekebalan fungi yang tumbuh pada tanaman terhadap fungisida sintetis tertentu atau dosis efektif, fungisida sintetis dapat mencapai dua kali dosis anjuran. Untuk menghindari kondisi yang lebih parah, tindakan yang perlu dilakukan adalah mengganti fungisida yang biasa dipakai dengan fungisida yang berbeda bahan aktif dan cara kerjanya.
tertinggalnya mikroorganisme seperti fungi di areal tanaman anggrek tersebut (http:// www.warintekjogja.com/. Diakses pada tanggal 16 April 2010).
Penyakit bercak hitam pada tanaman anggrek merupakan penyakit yang cepat menular melalui akar dan alat yang tidak steril, gejalanya timbul warna cokelat kehitaman pada bagian tanaman yang terserang. Mulai dari daun ke atas sampai ke tunas dan ke bawah hingga ujung akar. Tanaman terlambat tumbuh, kerdil dan mengakibatkan kematian. Penyakit busuk dengan gejalanya terdapat bintil-bintil kecil berwarna cokelat pada bagian tanaman yang terkena penyakit. Penyakit bercak cokelat dengan gejalanya bercak cokelat pada permukaan daun, lalu menyebar keseluruh bagian tanama 2010).
Busuk lunak, busuk pucuk batang yang disebabkan oleh Erwinia carotovora pada anggrek menyerang pangkal batang, tunas baru, mata tunas, dan akar. Cara serangan masuk ke dalam jaringan tanaman melalui lubang tanaman, luka pada tanaman, dan bekas potongan (Semangun, 1996). Gejalanya berupa jaringan tanaman yang terserang berubah warna menjadi kuning kemudian cokelat, biasanya mengeluarkan cairan lendir berbau busuk. Penyebaran penyakit disebabkan oleh percikan air hujan yang jatuh ke tanah kemudian dipantulkan ke atas kembali, angin, tangan manusia, serangga, gesekan pakaian manusia, alat-alat pemotong. Pengendaliannya mengkarantina tanaman yang terserang, menjauhkan dari tanaman sehat, membakar tanaman, membersihkan alat pemotong dengan dibakar atau menggunakan alkohol 70% sebelum digunakan
genangan air sisa penyiraman di atas permukaan daun. Gejalanya daun berubah warna kemudian melunak seperti berair di dalam jaringan (Semangun, 1996).
Gejala yang biasanya menyerang Phalaenopsis sp., dan Paphiopedilum sp., terdapat bercak cokelat, lunak dan berair pada permukaan daun, lalu menyebar keseluruh bagian tanaman (Semangun, 1996). Pengendaliannya dengan memotong semua bagian yang terinfeksi dengan alat yang steril. Merendam tanaman dengan natriphene atau physan selama kira-kira 1 jam, menyemprotkan fungisida antibiotik Streptomycin atau physan 20 (http://www.duniahobi.com. Diakses pada tanggal 27 April 2010).
Busuk hitam merupakan penyakit paling merusak yang menyerang tanaman anggrek, Cattleya sp., adalah yang terbanyak. Penyebabnya Pythium ultimum, gejalanya menginfeksi daun, kemudian menjalar sampai akhirnya ke akar. Daerah daun yang terinfeksi terdapat bercak berwarna cokelat dan dikelilingi oleh warna kuning, kontras dengan bagian daun lainnya yang berwarna hijau. Daerah pangkal daun yang terinfeksi berwarna kehitaman lalu melunak, jika ditekan akan mengeluarkan cairan dan proses pembusukannya cepat. Pada akar awalnya menjadi keriput, kemudian warnanya berubah menjadi cokelat tua dan akhirnya putus (Semangun, 1996). Untuk penanggulangan, bagian tanaman yang terinfeksi dipotong sampai batas area yang bebas dari penyakit dengan alat yang steril setelah itu merendam tanaman pada fungisida dan dikeringkan sebelum di tanam kembali. Menghindari busuk hitam dengan menjaga agar tanaman dan sekelilingnya tidak terlalu basah. Pemberian pupuk dengan kalsium yang tinggi akan mencegah busuk hitam pada daun. Untuk pengendaliannya dapat menggunakan fungisida (http://www.duniahobi.com/. Diakses pada tanggal 27 April 2010).
membusuk dan menjalar ke akar dan batang, daun dan batang menguning, berkeriput, tipis dan bengkok mengakibatkan tanaman menjadi kerdil dan tidak sehat (Semangun, 1996).
Beberapa isolat bakteri kitinolitik berpotensi untuk dikembangkan sebagai agen pengendali hayati fungi patogen tanaman. Uji pendahuluan yang dilakukan untuk mengetahui kemampuan bakteri kitinolitik menunjukkan bahwa terdapat isolat bakteri kitinolitik yang mampu menghambat pertumbuhan fungi uji. Meski demikian, kemampuan menghambat pertumbuhan fungi uji bervariasi (Suryanto et al., 2009a; Suryanto et al., 2009b; Suryanto et al., 2009e). Hal ini menunjukkan bahwa spesifisitas masing-masing bakteri berbeda. Menurut Irawati (2008) mengatakan bahwa bakteri kitinolitik dengan kode BK08, LK08, KR05, BK07 dan BK09, memiliki kemampuan dalam menghambat Ganoderma boninense, Fusarium oxyforum, dan Penicillium citrinum. Perbedaan tersebut dapat berasal dari kemampuan yang berbeda dalam menghasilkan enzim-enzim yang mampu mendegradasi dinding sel fungi (Suryanto et al., 2009e).
Kitinase dan kitin deasetilase adalah enzim yang aktif mengkatalisis degradasi
kitin. Kitinase dapat menghidrolisis kitin secara acak pada ikatan glikosidiknya,
sedang kitin deasetilase menghidrolisis kitin menjadi produk yang mempunyai daya
guna yang lebih tinggi seperti kitosan (Saleh et al., 1994). Banyak jenis
mikroorganisme dapat memproduksi enzim pendegradasi kitin dan kitosan, baik
bakteri seperti Bacillus megaterium (Pelleter et al., 1990), Streptomyces spp.,
(Boucher et al., 1992), Bacillus circulans (Alam et al., 1996), dan Vibrio alginolyticus
(Ohishi et al., 1996), Pseudomonas aeruginosa (Wang dan Chang, 1997),
Streptococcus lydicus (Crawford dan Mahadevan, 1997). Beberapa jenis fungi seperti
Aspergillus carneus (Sherief et al., 1991), Mucor rouxii (Bouriotis et al., 1993;
Kafetzoupoulos et al., 1994; Martinou et al., 1995) Aspergillus nidulans (Alfonso et
al., 1995; Reves et al., 1995), Absidia coerulea (Gao et al., 1995), dan Trichoderma
1.2Permasalahan Penelitian
Kitinase digunakan dalam pertanian sebagai pengendali fungi patogen tanaman dan hama serangga (Patil et al., 2000). Namun hingga saat ini belum dilakukan penelitian pada fungi yang menyerang tanaman anggrek.
1.3Tujuan Penelitian
Untuk mengetahui potensi bakteri kitinolitik sebagai agen pengendali hayati terhadap fungi patogen pada tanaman anggrek Cattleya sp., Vanda sp., dan Phalaenopsis sp.
1.4Hipotesis Penelitian
Bakteri kitinolitik mampu menghambat pertumbuhan fungi patogen yang diisolasi dari tanaman anggrek Cattleya sp., Vanda sp., dan Phalaenopsis sp.
1.5 Manfaat Penelitian
BAB 2
BAHAN DAN METODE
2.1 Waktu dan Tempat
Penelitian ini dilakukan dari bulan Mei sampai Desember 2010 bertempat di Laboratorium Mikrobiologi, Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam, Universitas Sumatera Utara, Medan, untuk isolasi,determinasi dan uji antagonis, sedangkan isolat fungi diperoleh dari tanaman anggrek yang terdapat di perkebunan tanaman anggrek daerah Tiga Dolok, Kabupaten Simalungun, Kecamatan Dolok Panribuan.
2.2 Alat dan Bahan
Alat yang digunakan dalam penelitian ini adalah cawan Petri, gunting tanaman, benang wol, mikroskop, buku identifikasi, cutter, Erlenmeyer, hotplate, objek glass, cover glass, ose bengkok, bunsen, handspray, aluminium foil, kertas label, thermometer, hygrometer, kamera digital, gunting, pipet serologi, propipet, tabung reaksi, pipet tetes, gelas ukur, timbangan analitik, oven, autoklaf, inkubator fungi, corkborer, neraca, label tempel, chingwrap, indikator pH, spatula, dan inkubator bakteri. Sedangkan bahan yang digunakan adalah akar, batang, dan daun dari tanaman anggrek Cattelya sp., Vanda sp., dan Phalaenopis sp., yang diduga terserang fungi patogen, PDA (potato dextrose agar), serbuk kitin, larutan garam, agar, spirtus.
2.3 Isolat Bakteri dan Fungi
pada pH 6,8 dan diinkubasi pada suhu 30oC. Sedangkan isolat fungi diperoleh dari isolasi langsung dari tanaman anggrek spesies Cattelya sp., Vanda sp., dan Phalaenopsis sp.
2.4 Isolasi dan Identifikasi Fungi Mikroskopik dari Tanaman Anggrek
Isolasi dilakukan dengan teknik direct plating (Malloc, 1997) dengan meletakkan irisan akar, daun, dan batang sepanjang 3 cm dari tanaman anggrek yang sakit pada medium PDA steril yang telah ditambah kloramfenikol dan yeast ekstrak dalam cawan Petri steril, kemudian diinkubasi pada suhu ruangan selama 72 jam. Koloni-koloni diambil dengan menggunakan cawan Petri steril dan untuk memastikan adanya fungi patogen dengan mencatat simptomnya. Fungi yang diperoleh kemudian dibiakkan dalam media PDA miring dan disimpan untuk uji berikutnya (Tsao, 1983). Identifikasi fungi yang diduga patogen dilakukan secara makroskopis dan mikroskopis. Hasil isolasi fungi yang berupa biakan murni, diambil gambarnya berdasarkan morfologi mikroskopisnya dan dideterminasi dengan menggunakan kunci determinasi fungi hingga pada marga dan jenisnya (Barnett dan Hunter, 1972; Malloch, 1997; Barnes dan Ervin, 1997).
2.5 Uji Antagonis Fungi Patogen pada Tanaman Anggrek Dengan Bakteri Kitinolitik lokal
Pengukuran penghambatan bakteri kitinase terhadap pertumbuhan fungi patogen dilakukan dengan cara mengukur pertumbuhan fungi yang terhambat dikurang dengan batas akhir pertumbuhan fungi patogen yang tidak terhambat kemudian dibagi dua (Martorejo, 2001).
2.6 Pengamatan Struktur Hifa Fungi setelah Uji Antagonis
BAB 3
HASIL DAN PEMBAHASAN
3.1 Karakteristik Fusarium sp., yang terdapat pada Tanaman Anggrek Cattelya sp, Vanda sp., dan Phalaenopsis sp.
Isolasi terhadap fungi patogen dilakukan dari tanaman anggrek yang sakit. Tiga genus anggrek yaitu Cattelya sp., Vanda sp., dan Phalaenopis sp., yang terserang Fusarium sp., dapat dilihat pada Gambar. 1 berikut ini:
a b c
Gambar 1. Tiga genus anggrek yang terserang Fusarium sp. (a) Cattelya sp. (b) Vanda sp. dan (c) Phalaenopsis sp.
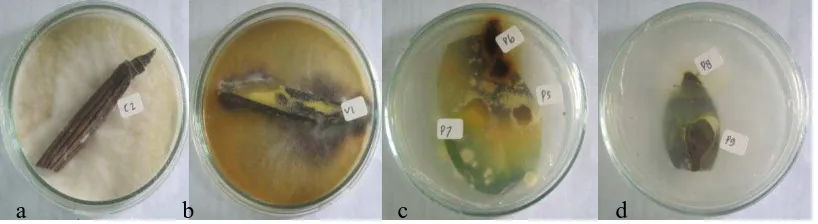
tanaman anggrek lainnya yang ada disekitarnya, namun hal ini tidak efesien. Fusarium sp. 1, memiliki miselium seperti kapas, berwarna putih. Fusarium sp. 2, memiliki miselium seperti kapas, berwarna abu-abu dan biasanya agak keunguan yang terlihat pada permukaan medium. Fusarium sp. 3, seperti kapas, berwarna putih dan tebal. Fusarium sp. 4, miselium seperti kapas, berwarna putih dan tipis. Miselium dari Fusarium sp. 5, ini tidak begitu tipis dan tidak begitu tebal, berwarna putih dan jika dilihat pada bagian bawah terlihat berawarna putih kekuningan berbeda dengan Fusarium sp. 3, dan Fusarium sp. 4.
a b c d
Gambar 2. Hasil isolasi fungi patogen pada tanaman anggrek yang terkena penyakit, masing-masing berasal dari (a) Fusarium sp. 1, (C2) yang menyerang batang Cattelya sp., (b) Fusarium sp. 2, (V1) yang menyerang daun Vanda sp., (c) Fusarium sp. 3, (P7) dan Fusarium sp. 4, (P5) yang menyerang daun Phalaenopsis sp., dan (d) Fusarium sp. 5, (P8) yang menyerang pada daun Phalaenopsis sp. Dengan masa inkubasi 2 X 24 jam pada suhu 30oC.
a b c
d e
Gambar 3. Biakan murni dan mikroskopik (a) Fusarium sp. 1, (b) Fusarium sp. 2, (c) Fusarium sp. 3, (d) Fusarium sp. 4, (e) Fusarium sp. 5, pada media PDA suhu 30oC, berumur 3 hari (perbesaran 400).
Mehrotra (1983) mengatakan bahwa hifa dari fungi Fusarium sp., terdapat di bagian sel dan antar sel jaringan tanaman inang. Jumlah hifa banyak pada seluruh pembuluh, kemudian menyebar dengan sistem beragam dan akhirnya menginfeksi pada bagian pangkal akar. Konidium Fusarium oxysporum f.sp. cubense berkembang menjadi klamidospora. Pada tanah yang terinfeksi berat dan berdrainasi buruk penyakit lebih cepat berkembang dibandingkan pada tanah yang berdrainasi baik. Pemupukan yang tepat serta drainasi yang baik dapat menekan perkembangan penyakit (Stover, 1970).
Alexopoulos dan Mimms (1979) setelah berkecambah miselium Fusarium sp., akan menghasilkan konidia dalam waktu 6-8 jam, sedang klamidospora terbentuk dalam waktu 2-3 hari. Di dalam jaringan pembuluh tanaman, fungi tumbuh dan masuk kejaringan parenkim yang berdekatan dan menghasilkan sejumlah besar konidia dan klamidospora. Konidia ini dapat berkembang menjadi klamidospora yang dapat kembali masuk ke dalam tanah ketika jaringan yang terinfeksi mati dan membusuk. Klamidospora ini tetap hidup dan bertahan dalam jangka waktu yang cukup lama di dalam tanah. Siklus penyakit akan berulang bila klamidospora ini berkecambah dan tumbuh kembali baik sebagai saprofit atau menyerang tanaman inang.
3.2 Uji Antagonisme In Vitro Bakteri Kitinolitik dengan Fusarium sp., pada Cattelya sp., Vanda sp., dan Phalaenopsis sp.
Tabel 1. Uji Anatgonisme antara Bakteri Kitinolitik dengan Fusarium sp., pada Cattelya sp., Vanda sp., dan Phalaenopsis sp.
Isolat
bakteri Fungi Zona hambatan (cm) hari ke-
6 7 8 9 10
BK07 Fusarium sp. 1 1,15 1,59 1,59 1,4 1,92 Fusarium sp. 2 0,39 0,04 0,66 1,79 1,00 Fusarium sp. 3 1,19 1,75 1,95 1,4 2,29 Fusarium sp. 4 0,74 0,86 1,14 1,5 2,19 Fusarium sp. 5 1 1,35 1,66 1,55 2,30 BK08 Fusarium sp. 1 0,66 1,5 1,54 1,95 0,49 Fusarium sp. 2 0,36 1,04 149 1,45 1,61 Fusarium sp. 3 0,94 1,99 2 1,33 2,11 Fusarium sp. 4 1,34 1,3 1,36 1,6 1,81 Fusarium sp. 5 1,22 1,43 1,55 1,45 2,1 BK09 Fusarium sp. 1 1,54 1,61 1,81 1,51 2,21 Fusarium sp. 2 0,04 0,14 0,3 0,6 0,69 Fusarium sp. 3 0,85 1,27 1,3 1,75 1,76 Fusarium sp. 4 1,39 1,36 1,35 1,7 2,49 Fusarium sp. 5 0,73 0,84 1,02 1,1 1,2 LK08 Fusarium sp. 1 0,73 1,16 1,6 1,6 1,7 Fusarium sp. 2 0,24 0,36 0,51 0,84 0,99 Fusarium sp. 3 0,36 1,56 1,55 1,65 2,30 Fusarium sp. 4 1,65 1,95 1,97 1,56 2,22 Fusarium sp. 5 1,43 1,99 0,87 1,69 1,33 KR05 Fusarium sp. 1 0,66 1,5 1,54 1,95 0,49 Fusarium sp. 2 0,36 1,04 1,49 1,45 1,61 Fusarium sp. 3 0,94 1,99 2 1,33 2,11 Fusarium sp. 4 1,34 1,3 1,36 1,6 1,81 Fusarium sp. 5 1,22 1,43 1,55 1,45 2,1 BK13 Fusarium sp. 1 0,34 0,41 0,6 0,6 2,63 Fusarium sp. 2 0,44 0,65 1,79 0,8 0,42 Fusarium sp. 3 0,91 1,4 1,44 1,45 1,46 Fusarium sp. 4 1,54 1,66 2,09 1,85 2,51 Fusarium sp. 5 0,83 1,78 2,26 1,95 2,49 BK14 Fusarium sp. 1 1,23 1,59 1,90 1,76 2,29 Fusarium sp. 2 0,49 0,04 0,7 1,09 1,29 Fusarium sp. 3 1,3 1,26 1,6 1,39 1,9 Fusarium sp. 4 1,15 1,22 1,4 1,2 1,81 Fusarium sp. 5 0,65 1,41 1,41 1,81 2,09
Pada pengamatan hari kesepuluh dari isolat bakteri. BK07 memiliki efektivitas penghambatan Fusarium sp. 5, dengan jari-jari zona hambat 2,3 cm adalah yang tinggi, sedangkan terendah adalah Fusarium sp. 2, dengan jari-jari zona hambat 1 cm. BK08 memiliki efektivitas penghambatan Fusarium sp. 3, adalah yang tinggi dengan jari-jari zona hambat 2,11 cm, sedangkan terendah adalah Fusarium sp. 1, dengan jari-jari zona hambat 0,49 cm. BK09 memiliki efektivitas penghambatan Fusarium sp. 4, adalah yang tinggi dengan jari-jari zona hambat 2,49 cm, sedangkan terendah adalah Fusarium sp. 2, dengan jari-jari zona hambat 0,69 cm. LK08 memiliki efektivitas penghambatan Fusarium sp. 3, adalah yang tinggi dengan jari-jari zona hambat 2,22 cm, sedangkan terendah adalah Fusarium sp. 2, dengan jari-jari zona hambat 0,99 cm. KR05 memiliki efektivitas penghambatan Fusarium sp. 3, adalah yang tinggi dengan jari-jari zona hambat 2,11 cm, sedangkan terendah adalah Fusarium sp. 1, dengan jari-jari zona hambat 0,49 cm.
BK13 memiliki efektivitas penghambatan Fusarium sp. 1, adalah yang tinggi dengan jari-jari zona hambat 2,63 cm, sedangkan terendah adalah Fusarium sp 2, dengan jari-jari zona hambat 0,42 cm. BK14, memiliki efektivitas penghambatan Fusarium sp. 1, adalah yang tinggi dengan jari-jari zona hambat 2,29 cm, sedangkan terendah adalah Fusarium sp. 2, dengan jari-jari zona hambat 1,29 cm. BK15, memiliki efektivitas penghambatan Fusarium sp. 5, adalah yang tinggi dengan jari-jari zona hambat 2,82 cm, sedangkan terendah adalah Fusarium sp. 2, dengan jari-jari zona hambat 0,9 cm. BK16, memiliki efektivitas penghambatan Fusarium sp. 5, adalah yang tinggi dengan jari-jari zona hambat 2,61 cm, sedangkan terendah adalah Fusarium sp. 2, dengan jari-jari zona hambat 0,46 cm. Untuk BK17, memiliki efektivitas penghambatan Fusarium sp. 2, adalah yang tinggi dengan jari-jari zona hambat 2,4 cm, sedangkan terendah adalah Fusarium sp. 4, dengan jari-jari zona hambat 1,7 cm.
menguraikan kitin yang ada pada dinding sel fungi. Menurut Muharni (2009) kitinase merupakan enzim yang mendegradasi kitin menjadi N-asetilglukosamin, degradasi kitin dapat dilakukan oleh organisme kitinolitik dengan melibatkan enzim kitinase. Organisme pendegradasi kitin umumnya berasal dari kelompok mikroorganisme, diantaranya adalah dari kelompok bakteri. Bakteri yang dilaporkan memiliki aktivitas kitinolitik seperti, Vibrio furnissi, Serratia marcescens, Bacillus circulans dan Pseudomonas aeruginosa. Bakteri kitinolitik pada bidang pertanian berfungsi sebagai biokontrol terhadap fungi patogen maupun serangga hama yang umumnya memiliki komponen kitin pada dinding selnya (Chernin et al., 1998).
Menurut El-Katatny et al., (2000) satu kelompok organisme yang memiliki potensi sebagai agen pengendali hayati fungi berasal dari kelompok mikroba penghasil kitinase. Pengendalian hayati fungi dengan menggunakan mikroba kitinolitik didasarkan pada kemampuan mikroba menghasilkan kitinase dan β-1,3-glukanase yang dapat melisiskan sel fungi. Bakteri lain yang juga digunakan sebagai pengendali hayati komersial seperti P. syringae, Burkholderia cepacia, B. subtilis, Agrobacterium radiobacter, Enterobacter cloacae, dan Streptomyces griseoviridis (Fravel et al., 1998; McQuilken et al., 1998). Bakteri kitinolitik seperti A. hydrophila, A. caviae, Pseudomonas maltophila, Bacillus licheniformis, B. circulans, Vibrio furnissii, Xanthomonas spp., dan Serratia marcescens memainkan peranan penting dalam
pengendalian hayati fungi patogen tanaman (Gohel et al., 2003). Penguraian kitin pada dinding sel fungi Fusarium sp., dapat dilihat pada Gambar. 4 berikut ini:
Gambar 4. Uji antagonis bakteri kitinolitik (a) BK14, terhadap Fusarium sp. 4, pada Phalaenopsis sp., dan (b) LK08 terhadap Fusarium sp. 1, pada Vanda sp., dengan masa inkubasi 10 hari pada suhu 30oC.
Shahidi et al., (1999) mengatakan bahwa kitin merupakan polimer alami kedua yang paling banyak tersedia di alam setelah selulosa, merupakan polimer aminoglukan dari N-asetil-D-glukosamin yang tidak larut air. Mikroorganisme kitinolitik mempunyai aktivitas antagonisme yang kuat terhadap fungi patogen dengan mekanisme hiperparasitismenya dan antibiotiknya sehingga efektif dalam menghambat pertumbuhan fungi patogen tanaman dengan mendegradasi dinding selnya. Beberapa enzim kitinolitiknya toksik pada fungi patogen penyebab penyakit tanaman budidaya tetapi tidak pada mikroorganisme lain dalam tanah dan tumbuhan inang (Kloepper et al., 1989). Menurut Oku (1994), peranan kitinase dalam pertahanan tanaman terhadap serangan
patogen terjadi melalui dua cara, yaitu: (1) menghambat pertumbuhan fungi dengan
secara langsung menghidrolisis dinding miselia dan (2) melalui pelepasaan elisitor
endogen oleh aktivitas kitinase yang kemudian memicu reaksi ketahanan sistematik pada
inang.
3.3 Pengamatan Struktur Hifa Abnormal Fusarium sp., pada Tanaman Anggrek setelah Uji Antagonisme
Dari uji antagonis yang dilakukan dengan menggunakan Fusarium sp., yang berasal dari tanaman anggrek yang diduga terserang fungi dengan sepuluh isolat bakteri kitinolitik, ternyata hifa Fusarium sp. 1, Fusarium sp. 3, dan Fusarium sp. 5, melilit atau keriting dan lisis, sedangkan pada Fusarium sp. 2, hifanya menyatu seperti menggulung-gulung dan pada Fusarium sp. 4, hifanya melilit atau keriting, lisis dan menyatu seperti menggulung-gulung.
et al., 1995b). Keanekaragaman, spesifitas organ dan perkembangan dan pola ekspresi yang berbeda menunjukkan bahwa β-1,3-glukanase memiliki fungsi biologis dalam pertumbuhan dan perkembangan tanaman dalam mekanisme pertahanan pada tanaman (Jin et al., 1999). Gambar. 5 berikut ini merupakan bentuk-bentuk hifa abnormal fungi patogen yang terdapat pada tanaman anggrek:
[image:30.612.111.509.198.302.2]
a b c d
BAB 4
KESIMPULAN DAN SARAN`
4.1 Kesimpulan
Dari hasil penelitian yang telah dilakukan dapat diambil kesimpulan bahwa BK15 memiliki efektivitas penghambatan paling tinggi yaitu pada Fusarium sp. 5, dengan jari-jari zona hambat sebesar 2,82 cm. Sedangkan BK13 memiliki efektivitas penghambatan paling rendah yang terdapat pada Fusarium sp. 2, dengan jari-jari zona hambat sebesar 0,42 cm. Untuk hifa abnormal dalam pengamatan mikroskopik Fusarium sp., yang ada pada Cattelya sp., Phalaenopsis sp., dan Vanda sp., mengalami lisis, menggulung dan melilit setelah dilakukan uji antagonis.
4.2 Saran
Perlu dilakukan penelitian lebih lanjut mengenai variasi dari jenis tanaman anggrek lainnya yang terserang penyakit, identifikasi isolat serta pemanfaatan isolat untuk tujuan pengendalian hayati.
DAFTAR PUSTAKA
Alam, Md. M., Mizutani, T., Isono, M., Nikaidou, N. and Atanabe, T. 1996. Three Chitinase Genes (chiA, chiB, and chiC) Comprise The Chitinase System of Bacillus circulans WL-12. J. Ferment Bioeng. 82(1): 28-36.
Alexopoulos, C. J and C. W. Mimms. 1979. Introductory Mycology. Third edition. New York: John Wiley and Sons, Inc. 296.
Alfonso,C., Neuro, O.M., Santamaria, F. and Reves, F. 1995. Purification of Heat Stable Chitin Deacetylase From Aspergillus nidulans and Its Role in Cell Wall Degradation. J. Curr. Microbiol. 30(1): 49-54.
Barnes, Ervin H. 1997. Atlas and Manual of Plant Pathology. Appleton-Century-Crofts. New York. 126-130.
Barnett, H.L. and B.B. Hunter. 1972. Illustrated Genera of Imperfect Fungi. Co. Minneapolis.
Boucher, I, Dupuy, A., Vidal, P., Neugebauer, W.A. and Brzezinski, R. 1992. Purification and Characterization of Chitosanase From Streptomyces N-174. J. Appl. Microbiol
and Biotechnol. 38(2): 188-193.
Bouriotis, V., Kafetzopoulos, D. and Vournakis, J. 1993. Process for Isolating and Preparing Purified Chitin Deacetylase. Inst. Mol. Biol. Biotechnol. US Patent No. 5219749.
Crawford D. L. and Mahadevan, B. 1997. Properties of The Chitinase of the Antifungal Biocontrol Agent Streptomyces lydicus WYEC-108. Enzyme Microb. Technol. 20(7): 489-493.
Donzelli B, Lorito M, Scala F, and Harman G. 2001. Cloning, Sequence and Structure of a Gene Encoding an Antifungal Glucan 1,3-β-Glucosidase from Trichoderma atroviride (T. harzianum).
De la Cruz J, Pintor-Toro JA, Benitzer T, and L1obel A. 1995. Purification and Characterization of An Endo-β-1,6-Glucanase from Trichoderma harzianum that is Related to Its Mycoparasitism. Journal of Biotecnology. 177: 64-71.
Fravel DR, Connick Jr, WJ, and Lewis JA. 1998. Formulation of Microorganisms to Control Plant Diseases. In Formulation of Microbial Biopesticide. Ed. Burges HD. Kluwer Academic Press. Dordrecht. 187-202.
Gao, X. D., Katsumoto, T., and Ondera, K. 1995. Purification and Characterization of Chitin Deacetylase from Absidia coerulea. J. Biochem. 117(2): 257-263.
Gohel V, Singh A, Vimal M, Ashwini D, and Chatpar HS. 2003. Bioprospecting and Antifungal Potential of Chitinolytic Microorganism. African J Biotechnol. 5: 54-72.
Hasanuddin. 2003. Peningkatan Peranan Mikroorganisme Dalam Sistem Pengendalian Penyakit Tumbuhan Secara Terpadu. Jurusan Hama dan Penyakit Tumbuhan. Fakultas Pertanian. Universitas Sumatera Utara. Digitilized by USU digital library.
Hocking and J. L. Pitt. 1997. Fungi and Food Spoilage. Blackie Academic & Professional: Australia.
Di akses tanggal 16 April 2010.
http://www.duniahobi.com/forum/viewtopic.php?f=81&t=840. Diakses tanggal 27 April 2010.
Irawati, N. 2008. Potensi Bakteri Kitinolitik Lokal Dalam Pengendalian Fungi Patogen Tumbuhan. Skripsi. USU. Medan.
Jin W, Horner H, Palmer R, and Shoemaker R. 1999. Analysis and Mapping of Gene Families Encoding β-1,3-Glucanase of Soybean. Genetic. 153: 445-452.
Kafetzoupoulos, D., Martinou, A., Tsigos, I., Christodoulidou,A., Kavelaki, P. and Bouriotis, V. 1994. Chitin Deacetylation by Enzymic Means. J. Prog. Biotechn o
l. 9(1): 291-294.
Malloch, D. 1997. Moulds Isolation, Cultivation, Identification, Mycology. Toronto: Departement of Botany, University of Toronto.
Martorejo, T., C. Sumardiyono dan E. H. Astuti. 2001. Kajian Pengendalian Hayati Kapang Hijau pada Buah Jeruk Dengan Trichoderma sp. Prosding Seminar PFI. IPB. Bogor. 354-356.
McQuilken, M., h. G. Powell, S. P. Budge and J. M. Wipps. 1997. Effect of Storage on the Survival and Biocontrol Activity of Pythium oligandrum in Pelleted Sugar Beet Seed. Biocon. Sci and Tech. 8: 237-241.
Morikawa, Y., Nogawa, M., Takahashi, H., Kashigawa, A., Oshima, K and Okada, H. 1998. Purification and characterization of exo-β- D-glucosaminidase from Sellulotic Fungus Trichoderma reesei PC-37. J. Appl. Environ. Microbiol. 64(3): 890-895.
Munawaroh, E. 2001. Eksplorasi dan Inventarisasi Tumbuhan Dalam Rangka Konservasi Keanekargaman Jenis Tumbuhan. Sehari hasil-hasil penelitian bidang ilmu hayat. Bogor. Balai Pengembangan Kebun Raya-LIPI.
Muharni. 2009. Isolasi dan Identifikasi Bakteri Penghasil Kitinase dari Sumber Air Panas Danau Ranau Sumatera Selatan. Jurusan Biologi FMIPA, Universitas Sriwijaya, Sumatera Selatan, Indonesia. Jurnal Penelitian Sains. Edisi Khusus Desember. 09: 12-15.
Ohishi, K., Yamagishi, M., Okta, T., Suzuki, M. and Izumida, H. 1996. Purification and Properties of Two Chitinases from Vibrio alginolyticus H-8. J. Ferment. Bioeng. 82(6): 598-600.
Oku, H. 1994. Plant Pathogenesis and Disease Control. London: Lewis Pub.
Patil, R. S., V. Ghormade and M. V., Deshpande. 1999. Chitinolytic Enzymes AnEx ploration. J. Enzyme and Microbial Technol. 26: 473-483.
Patil, R. S., V. Ghormade and M. V., Deshpande. 2000. Chitinolytic Enzymes: An Exploration. Enz. Microb. Tech. Rev. 26: 473-483.
Pelletier, A. and Sygusch, J. 1990. Purification and Characterization of Three Chitosan ase Activities from Bacillus megaterium PI. J. Appl. Environ. Microbiol. 6(4): 844-848.
Reves, F., Alfonso, C., Neuro, O.M. and Santamaria, F. 1995. Purification of Heat-Stable Chitin Deacetylase from Aspergillus nidulans and Its Role in Cell Wall Degradation. Current Microbiol. 30(1) : 49-54.
Saleh, M., Abdilah, R., Suherman, M., Basmal, J. dan Indriati, N. 1994. Pengaruh Suhu, Waktu, dan Konsentrasi Pelarut pada Ekstraksi Kitosan dari Limbah Pengolahan Udang Beku terhadap beberapa Parameter Mutu Kitosan. Jurnal Pen. Pasca
Semangun, H. 1996. Ilmu Penyakit Tanaman. Yogyakarta. Gadjah Mada University Press. hlm 96.
Sherief, A. A., El-Sawah, M. M., and El-Naby, M. A. Abd. 1991. Some Properties of Chitinase Produced by a Potent Aspergillus carneus Strain. J. Appl. Microbiol.
Biotech. 35(2): 228-230.
Suryanto, D., Fakhrullah., Napitupulu, D., Munir, E. 2009e. Assay of Three Chitinolytic Bacterial Isolates of Taman Nasional Gunung Leuser Tangkahan to Inhibit Plant Pathogenic Fungi. Microbiol Indones Press.
Tsao, P. H. 1983. Factors Affecting Isolation & Quantitation of Phytophthora from soil. In D. C. Erwin, S. B. Garcia dan P. H. Tsao. Phytophthora its Biology, Taxonomy and Ecology. The American Phytopatological Society. St. Paul. 219-236.
Wang, San-Lang and Chang, Wen-Tsu, 1997. Purification and Characterization of Two Bifunctional Chitinases Lysozymes Extracellularly Produced by Pseudomonas
Lampiran 1: Komposisi Medium Medium MGMC padat
• K2HPO4 0,7 g
• KH2PO4 0,3 g
• MgSO4.7H2O 0,5 g
• FeSO4.7H2O 0,01 g
• ZnSO4 0,001 g
• MnCl2 0,001 g
• Koloidal kitin 12,5% (b/v) 72,7 ml
• Agar 20 g
• pH 6,8
Cara Pembuatan
Lampiran 2 : Alur Kerja Karakterisasi Sifat Morfologi Dan Biokimia Isolat Isolat Bakteri kitinolitik
Karakteristik
Morfologi Pewarnaan Gram Uji Biokimia Sederhana
Bentuk koloni Uji Katalase
Warna koloni Uji Sitrat
Elevasi Koloni UJi Hidrogen Sulfida
Bentuk Sel Uji Motilitas
Lampiran 3. Isolasi dan Identifikasi Mikroskopis Jamur Patogen Tanaman Anggrek
Akar, batang, daun dan bunga tanaman anggrek yang sakit
Disimpan di dalam kantong plastik yang disemprot alkohol 70 % dan dibawa ke Laboratorium Mikrobiologi USU
Dipotong 3 cm Diinkubasi selama 2-5 hari
Ditumbuhkan ke dalam cawan petri steril berisi media PDA (potato dextrose agar) padat.
Diinkubasi selama 2-5 hari Isolat Fungi
Diamati di bawah mikroskop dengan metode block square
Diidentifikasi dengan panduan buku identifikasi fungi Hocking and J. L Pitt (1997)
Lampiran 4. Alur Kerja AsaiAntagonime in vitro
Biakan (Fusarium sp 1., Fusarium sp 2., Fusarium sp 3., Fusarium sp 4., Fusarium sp 5)
Diinokulasikan pada media MGMC (Suryanto, 2001) pada jarak 3,5 cm dari cakram tempat inokulum bakteri.
Diinkubasi selama 10 hari Biakan Fungi
Dibuat cakram dengan diameter 0,5 cm dibagian pinggir media
Dibuat 2 titik pengulangan
Setelah 24 jam kemudian, sebanyak 30 µ l (≈ 108 sel/ml) sel bakteri diinokulasikan ke dalam sumur berdiameter 2 mm pada bagian tengah jamur yang telah diinkubasi.
Diinkubasi pada suhu 30oC dan dilakukan pengamatan selama 10 hari
Diamati zona hambat terhadap miselia fungi patogen tanaman anggrek yang terbentuk mulai dari hari ke-2 sampai ke-10
Hasil Pengamatan
Pengamatan Struktur Hifa Jamur Patogen Tanaman Anggrek Setelah Uji Patogenitas.
Isolat Fungi
Lampiran 5. Pembuatan Koloidal Kitin dengan Cara hidrolisis Parsial (Rodriquez-Kabana et al, 1983).
20 gr serbuk kitin
Dihaluskan
Dilarutkan dalam 180 ml 10 N HCl sambil diaduk dengan pengaduk magnetik selama 1,5- 2 jam.
Suspensi Kitin
Dituang ke dalam erlenmeyer yang berisi 2 liter air
Dibiarkan semalam kemudian bagian yang bening dipisahkan dari endapannya.
Endapan Bagian Bening
Dicuci sampai pH suspensi kitin antara 5,5 – 6
Diambil 10 ml kemudian dikeringkan pada suhu 80o C Dihitung berat keringnya