PENGEMBANGAN GALUR MANDUL JANTAN
DENGAN TIGA SUMBER SITOPLASMA
UNTUK PERAKITAN PADI HIBRIDA
INDRASTUTI APRI RUMANTI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Dengan ini saya menyatakan bahwa disertasi Pengembangan Galur Mandul Jantan dengan Tiga Sumber Sitoplasma untuk Perakitan Padi Hibrida adalah karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Januari 2012
Derived from Three Cytoplasm Sources and Its Use on Hybrid Rice Breeding. Under direction of BAMBANG S. PURWOKO as chairman, HAJRIAL ASWIDINNOOR and ISWARI S. DEWI as members of the advisory committee.
Hybrid rice is an alternative technology to increase lowland rice productivity. In Indonesia, hybrid rice was developed through three line system that involved three parental lines i.e. cytoplasmic male sterile lines (CMS), maintainer (B) and Restorer (R). CMS as seed parent should have high pollen sterility, be stable, easy to restore, good in agro-morphology and flower behavior, high in combining ability and resistant to pest and diseases.
Intensive use of single source of male sterile cytoplasm (i.e. wild abortive or WA) in developing rice hybrids might lead to genetic vulnerability associated with susceptibility to pests and diseases.A series of experiments were conducted to identify new CMS from three cytoplasm sources (WA, Gambiaca and Kalinga) with high pollen sterility, stable and easy to restore. CMS efectivity on seed production and hybrid rice breeding were studied.
The results showed that among doubled haploid population,panicle weight had thelargest contribution to grain weight per hill, as shown by high and positive direct effect value (0.7739) and indirect effect value through number of filled grain per panicle (0.6556), therefore the characters could be usedfor selection of good maintainer lines. H36-3-Mb, H36-3-Mc, H36-4-M, H45-3-Da, B1-2-Pb and B2-2-Pb were suitable to be used as new maintainer candidates related to their high number of filled grain and yield. Doubled haploid of B1-1-Mb, Db and B2-1-M were resistant to Bacterial Leaf Blight (BLB) strain III. B4-1-Dc line was very resistant (score 0) to BLB strain IV and VIII.
Six CMS of wild abortive types, one CMS of Gambiaca and three Kalinga types were bred through successive backcrosses. The three types of CMS lines were semi dwarf and having better flowering characters than the check line. The good flowering behavior of the new CMS lines improved seed set up to 25.90% compared to 2.98% of seed set in IR58025A. The new CMS were more effective and easier in its seed production. Phenotype obstacles such as low panicle exertion and short panicle length were improved through GA3 spraying. It increased the yield of seed.
The three types of CMS had spesific combining ability to many restorers and yielded high yielding hybrid combination with good heterosis. The results indicated that the yield of hybrids were controlled by overdominant, dominant x dominantor epistasis gene activity. The best hybrid was derived from WA cytoplasm i.e. BI485A/IR53942 (10.21 t/ha), while the best hybrid derived from Gambiaca cytoplasm was BI625A/SMD11 (9.88 t/ha). BI665A/IR53942 was the best hybrid derived from Kalinga cytoplasm, yielded up to 9.15 t/ha.
It can be concluded that new CMS derived from WA, Gambiaca and Kalinga cytoplasm were more efective on seed production and hybrid rice development because of their good flower behaviour, resistance to BLB and good specific combiner with restorers.
Tiga Sumber Sitoplasma untuk Perakitan Padi Hibrida.Dibimbing oleh BAMBANG S. PURWOKO sebagai ketua, HAJRIAL ASWIDINNOOR dan ISWARI S. DEWI sebagai anggota komisi pembimbing.
Teknologi padi hibrida merupakan salah satu alternatif dalam usaha peningkatan produktivitas padi sawah.Perakitan padi hibrida di Indonesia menggunakan sistem tiga galur yang melibatkan tiga galur tetua yaitu galur mandul jantan (GMJ), galur pelestari (B) dan galur pemulih kesuburan (R).GMJ merupakan kunci utama dalam pengembangan padi hibrida sistem tiga galur, karena berfungsi sebagai seed parent. GMJ yang baik harus memiliki sterilitas polen sempurna, stabil, mudah dipulihkan fertilitasnya, memiliki karakter agromorfologi dan bunga baik, memiliki daya gabung baik dan tahan terhadap satu atau lebih hama penyakit tropis.
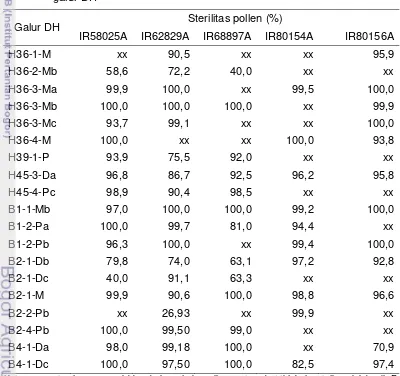
GMJ yang digunakan di Indonesia masih didominasi oleh GMJ dengan latar belakang sitoplasma wild abortive (WA). Penggunaan satu tipe GMJ dalam area luas dikhawatirkan dapat menimbulkan kerapuhan genetik, sehingga hibrida menjadi lebih rentan terhadap hama, penyakit maupun perubahan lingkungan yang ekstrim. Serangkaian penelitian dilakukan untuk memperoleh GMJ baru dengan tiga sitoplasma berbeda (WA, Gambiaca dan Kalinga) yang memiliki sterilitas polen sempurna, stabil dan mudah dipulihkan. Efektivitas GMJ dalam produksi benih dan perakitan padi hibrida menjadi target berikutnya dalam penelitian ini.
Hasil penelitian menunjukkan bahwa pada populasi galur dihaploid calon pelestari, bobot malai mempunyai kontribusi terbesar terhadap hasil biji per rumpun, terlihat dari nilai pengaruh langsungnya yang positif dan besar (0,7399) dan pengaruh tidak langsungnya melalui jumlah gabah isi per malai yang juga tinggi dan positif (0,6556), sehingga dua karakter ini dapat digunakan sebagai karakter seleksi galur pelestari yang baik. Galur haploid ganda Mb, H36-3-Mc, H36-4-M, H45-3-Da, B1-2-Pb dan B2-2-Pb sesuai untuk digunakan sebagai calon galur pelestari karena mempunyai jumlah gabah isi per malai dan bobot hasil yang tinggi.Galur DH B1-1-Mb, B2-1-Db dan B2-1-M bereaksi tahan terhadap Hawar Daun Bakteri (HDB) strain III. Galur B4-1-Dc merupakan galur DH pelestari sangat tahan (skor 0) terhadap HDB strain IV dan VIII.
Transfer sifat mandul jantan dan silang balik berkelanjutan telah berhasil mengidentifikasi 10 GMJ baru dengan tiga latar belakang sitoplasma berbeda (WA, Gambiaca dan Kalinga). Sepuluh GMJ baru tersebut memiliki sterilitas polen yang sempurna dan stabil.GMJ tipe Gambiaca dan Kalinga lebih cepat mencapai 100% steril dibandingkan GMJ tipe WA.GMJ baru tersebut memiliki tinggi tanaman antara 66 – 107 cm, jumlah anakan kategori sedang dan umur genjah, serta didukung oleh persentase eksersi stigma yang baik. Satu GMJ tipe WA, Gambiaca dan Kalinga bereaksi tahan terhadap HDB strain III, berturut-turut yaitu BI703A, BI855A dan BI669A. Galur BI543A dan BI571A (WA) teridentifikasi tahan terhadap HDB strain VIII. BI665A (Kalinga) memberikan reaksi tahan terhadap dua strain sekaligus, yaitu IV dan VIII, yang merupakan strain paling virulen di Indonesia.
stigma (r = 0,44*), eksersi stigma (r = 0,54*) dan sudut membuka lemma dan palea galur mandul jantan (r = 0,42*). Seed set GMJ juga berkorelasi positif dengan panjang filamen (r = 0,47*) dan sudut pembukaan bunga galur pelestari (r = 0,57**).
Studi terhadap potensi produksi benih GMJ baru, menunjukkan bahwa GMJ baru tipe WA, Gambiaca dan Kalinga mampu menghasilkan bobot panen benih yang lebih tinggi dibandingkan IR58025A. Kelemahan GMJ baru berupa rendahnya persentase eksersi malai dan malai pendek dapat diatasi dengan penyemprotan 200 ppm GA3. Aplikasi GA3 meningkatkan hasil benih GMJ baru yaitu mencapai 1031,90 kg/ha (WA), 763,34 kg/ha (Kalinga) dan 457,45 kg/ha (Gambiaca).
Tiga tipe GMJ baru teridentifikasi memiliki daya gabung khusus dengan berbagai galur pemulih kesuburan, sehingga menghasilkan kombinasi hibrida dengan potensi hasil dan heterosis tinggi baik terhadap tetua terbaik, rata-rata tetua maupun varietas pembanding. Hal ini menunjukkan bahwa bobot hasil padi hibrida dikendalikan oleh aksi gen overdominan, dominan x dominan atau epistasis. Kombinasi hibrida turunan GMJ tipe WA terbaik adalah BI485A/IR53942 (10,21 t/ha), sedangkan hibrida turunan GMJ tipe Gambiaca terbaik adalah BI625A/SMD11 (9,88 t/ha). Hibrida turunan GMJ tipe Kalinga terbaik ditunjukkan oleh BI665A/IR53942 (9,15 t/ha)
Dari rangkaian penelitian ini dapat disimpulkan bahwa GMJ baru tipe WA, Gambiaca maupun Kalinga efektif digunakan dalam produksi benih dan perakitan padi hibrida karena memiliki biologi dan perilaku bunga baik, tahan terhadap HDB dan memiliki daya gabung baik terhadap galur pemulih kesuburan yang ada. Diperoleh kombinasi hibrida dengan hasil dan heterosis tinggi dari ketiga latar belakang genetik GMJ.
© Hak Cipta milik IPB, tahun 2012
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang wajar IPB.
PENGEMBANGAN GALUR MANDUL JANTAN
DENGAN TIGA SUMBER SITOPLASMA
UNTUK PERAKITAN PADI HIBRIDA
INDRASTUTI APRI RUMANTI
Disertasi
sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertutup: Dr. Ir. Satoto, MS Dr. M Syukur, SP., M.Si.
Sumber Sitoplasma untuk Perakitan Padi Hibrida
Nama
: Indrastuti Apri Rumanti
NIM
: A263074011
Disetujui
Komisi Pembimbing
Prof. Dr. Ir.Bambang S. Purwoko, M.Sc.
Ketua
Dr. Ir.Hajrial Aswidinnoor, M.Sc.
Dr.Ir. Iswari S. Dewi
Anggota
Anggota
Mengetahui
Ketua Program Studi
Dekan Sekolah Pascasarjana
Pemuliaan dan Bioteknologi Tanaman
Dr. Ir.Trikoesoemaningtyas, M.Sc.
Dr. Ir. Dahrul Syah, M.Sc. Agr.
Alhamdulillaahirabbil’aalamiin.
Atas rahmat dan karunia Allah SWT,
disertasi ini berhasil diselesaikan dengan baik. Adanya dominasi
sitoplasma secara luas di Indonesia menyebabkan kualitas dan hasil padi
hibrida hampir sama antar varietas. Terlebih lagi, kekhawatiran terjadinya
kerapuhan genetik pada GMJ seperti yang terjadi pada jagung
Texas
,
maka penulis memilih tema
Pengembangan Galur Mandul Jantan
dengan Tiga Sumber Sitoplasma untuk Perakitan Padi Hibrida.
Terimakasih penulis sampaikan kepada Prof. Dr. Bambang S.
Purwoko MSc., Dr. Ir. Hajrial Aswidinnoor MSc. dan Dr. Ir. Iswari S. Dewi
selaku komisi pembimbing yang dengan ikhlas memberikan masukan,
arahan, bimbingan, dorongan dan motivasi dalam penulisan disertasi ini.
Penulis sampaikan penghargaan sebesar-besarnya kepada Dr. Satoto
dan Dr. M Syukur selaku penguji luar komisi dalam ujian tertutup, serta Dr.
Trikoesoemaningtyas dan Dr. Suwarno yang telah berkenan menjadi
penguji luar komisi dalam ujian terbuka.
Penulis juga menyampaikan terimakasih kepada Badan Litbang
Pertanian atas dukungan dana beasiswa dan dana penelitian melalui
kerjasama Badan Litbang Pertanian& IPB dalam program KKP3T 2007
–
2011 a.n. Prof. Bambang S. Purwoko dkk. Penulis juga menyampaikan
terimakasih kepada BB Biogen atas pemberian materi hasil kultur antera
dan fasilitas melalui Dr. Iswari S. Dewi sebagai bahan penelitian, serta
Pak Iman dan Mbak Yeni yang dengan ikhlas telah membantu penulis
selama bekerja di laboratorium dan Rumah Kaca. Penghargaan dan
terimakasih, penulis sampaikan kepada Kepala Balai Besar Penelitian
Padi (BB Padi) dan ketuatim penelitian padi hibrida BB Padi, Dr. Satoto,
atas pemberian materi penelitian, ijin, dukungan dan masukannya. Juga
teman-teman peneliti dan teknisi: Murdani, Yuni Widyastuti, Sudibyo,
Suwarto, Wasidi, Cecep, Sarmadi, Suardi, Firman, Dede, Nita, Bayu dan
Bowo untuk dukungan doa dan tenaganya. Teman-teman seperjuangan di
PBT 2007, 2008, 2009 dan 2010 untuk persahabatannya.Terimakasih
khusus penulis ucapkan untuk Indria Mulsanti dan keluarga, semoga
silaturahmi terus terjaga.
Kepada suami tercinta, Didik Ariyanto, Bapak Ibu Wahyuno, Ibu
Siswantinah dan ananda terkasih Zaki dan Kayla, juga kakak dan adik
tersayang, disampaikan terimakasih atas cinta, doa, dorongan dan kasih
sayang yang telah menguatkan penulis dalam melaksanakan semua
penelitian dan penulisan disertasi ini.
Tiada yang sempurna pada karya manusia, tetapi besar harapan
penulis, karya ini dapat bermanfaat dalam pengembangan padi,
khususnya padi hibrida di Indonesia.
Bogor, Januari 2012
Penulis dilahirkan di Banyumas pada tanggal 27 April 1977 dari
Bapak Wahyuno dan Ibu Ambarwati, sebagai anak pertama dari dua
bersaudara.Penulis memiliki satu orang adik laki-laki bernama Prastoto
Indratmoko. Pada tahun 2000, penulis menikah dengan Didik Ariyanto,
S.Pi, M.Si dan dikaruniai dua putra putri, yang diberi nama Zaki Najmudin
Furqon Ariyanto dan Kayla Putri Ariyanto.
Penulis lulus dari Fakultas Pertanian, Universitas Jendral Soedirman,
Program Studi Pemuliaan Tanaman pada tahun 1999. Selanjutnya tahun
2000 penulis mulai bekerja di Balai Besar Penelitian Tanaman Padi pada
Kelompok Peneliti Pemuliaan Padi dan sejak saat itu penulis intensif
melakukan penelitian bersama tim padi hibrida. Penulis turut aktif dalam
pelepasan varietas padi hibrida sejak tahun 2002 hingga 2011. Sejumlah
artikel ilmiah sebagai wujud penyebaran informasi juga telah penulis
publikasikan di jurnal maupun prosiding bersama tim peneliti padi hibrida
BB Padi.
Halaman
DAFTAR TABEL ... xv
DAFTAR GAMBAR ... xvi
PENDAHULUAN ... 1
Latar Belakang... 1
Tujuan Penelitian ... 4
Hipotesis ... 4
Diagram Alur Penelitian ... 5
TINJAUAN PUSTAKA ... 7
Asal dan Taksonomi Padi ... 7
Padi Hibrida ... 9
Sistem Mandul Jantan pada Padi ... 12
Uji Daya Gabung ... 14
EVALUASI GALUR-GALUR DIHAPLOID UNTUK DIGUNAKAN SEBAGAI CALON GALUR PELESTARI ……… 17
Abstrak ... 17
Abstract... 18
Pendahuluan ... 19
Bahan dan Metode ... 21
Hasil dan Pembahasan... 26
Kesimpulan ... 38
TRANSFER SIFAT MANDUL JANTAN DAN PEMBENTUKAN GALUR MANDUL JANTAN MELALUI SILANG BALIK ... 39
Abstrak ... 39
Abstract... 40
Pendahuluan ... 41
Bahan dan Metode ... 42
Hasil dan Pembahasan... 44
Kesimpulan ... 52
KARAKTER BUNGA YANG MENDUKUNG KEMAMPUAN MENYERBUK SILANG GALUR MANDUL JANTAN 53 Abstrak ... 53
Bahan dan Metode ... 56
Hasildan Pembahasan... 58
Kesimpulan ... 72
POTENSI PRODUKSI BENIH GALUR MANDUL JANTAN BARU TIPE WILD ABORTIVE, GAMBIACA DAN KALINGA ... 73
Abstrak ... 73
Abstract... 74
Pendahuluan ... 75
Bahan dan Metode ... 76
Hasil dan Pembahasan ... 80
Kesimpulan ... 85
EVALUASI DAYA GABUNG DAN HETEROSIS HIBRIDA TURUNAN GMJ TIPE WILD ABORTIVE, GAMBIACA DAN KALINGA MENGGUNAKAN ANALISIS LINI X TESTER ... 87
Abstrak ... 87
Abstract... 88
Pendahuluan ... 89
Bahan dan Metode ... 90
Hasil dan Pembahasan ... 92
Kesimpulan ... 114
PEMBAHASAN UMUM ... 117
KESIMPULAN DAN SARAN ... 127
Kesimpulan ... 127
Saran ... 128
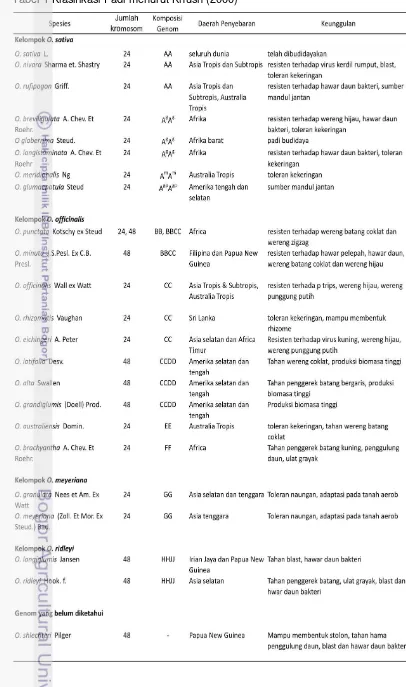
Halaman 1 Klasifikasi padi menurut Khush (2000) ... 8
2 Galur calon pelestari DH2 hasil kultur antera dari 6 kombinasi
persilangan B x B ……….
21
3 Skor warna daun berdasarkan SES (IRRI 2002) ……..………. 22
4 Skala sudut daun bendera berdasarkan SES (IRRI 2002) …... 22
5 Skala eksersi malai berdasarkan SES (IRRI 2002) ……… 22
6 Skor keberadaan bulu pada gabah ……….. 23
7 Kriteria penilaian ketahanan terhadap hawar daun bakteri
(HDB) ……….……… 24
8 Analisis ragam (anova) rancangan acak kelompok ……… 25 9 Karakter morfologi dan karakter bunga 19 calon galur
pelestari ……….………... 27
10 Nilai rata-rata dan parameter genetik dari delapan karakter
pada 19 galur dihaploid calon galur pelestari ……….. 28 11 Koefisien korelasi pearson antar 7 karakter pada galur-galur
dihaploid padi ... 30
12 Pengaruh langsung dan tidak langsung karakter vegetatif dan komponen hasil terhadap hasil biji per rumpun pada
galur-galur dihaploid padi ………….……… 31
13 Nilai rata-rata karakter agronomis dan komponen hasil 19
calon galur pelestari ……… 34
14 Hasil uji ketahanan galur dihaploid calon pelestari terhadap
hawar daun bakteri ……….. 35
15 Skor eksersi malai berdasarkan SES (IRRI 2002) ... 44
16 Skor eksersi stigma berdasarkan SES (IRRI 2002) ... 44
17 Sterilitas polen tanaman F1 hasil persilangan dari 5 GMJ
generasi F1 - F1BC5………. 47 19 Keragaan karakter agronomis, eksersi malai dan eksersi
stigma tiga tipe galur mandul jantan baru ……… 49
20 Ketahanan galur mandul jantan baru terhadap Hawar Daun
Bakteri ……….. 51
21 Eksersi malai, eksersi stigma, panjang dan lebar stigma, panjang stilus dan warna putik dari tiga tipe galur mandul
jantan baru ... 60
22 Panjang antera, filamen, skor anthesis dan warna antera
galur pelestari ……… 64
23 Umur berbunga, durasi dan sudut pembukaan bunga serta seed set yang terbentuk pada galur mandul jantan baru dan
pelestarinya ………. 68
24 Korelasi fenotipik sejumlah karakter dan perilaku bunga galur mandul jantan terhadap persentase terbentuknya biji (seed
set) ……… 69 25 Korelasi fenotipik sejumlah karakter dan perilaku bunga galur
pelestari terhadap persentase terbentuknya biji (seed set) ….. 70 26 Karakter agromorfologi dan umur berbunga 50% tiga galur
mandul jantan baru dan pembanding (IR58025A) pada dua
konsentrasi GA3 selama produksi benih galur mandul jantan 80
27 Sterilitas polen, eksersi stigma, eksersi malai, persentase pengisian biji (seed set) dan produktivitas galur mandul jantan baru dan pembanding (IR58025A) pada dua konsentrasi GA3
selama produksi benih galur mandul jantan ……… 82 28 Daftar galur mandul jantan dan restorer yang digunakan
dalam pembentukan hibrida ... 90 29 Struktur tabel analisis ragam untuk analisis lini × tester ……… 92 30 Nilai kuadrat tengah hasil analisis varians daya gabung pada
beberapa karakter padi ……….. 93
31 Proporsi kontribusi lini, tester dan interaksinya terhadap
varians total ………. 95
32 Efek daya gabung umum tetua hibrida baru pada berbagai
karakter ……….
96
33 Efek daya gabung khusus karakter agromorfologi, komponen hasil dan hasil 75 hibrida baru ……….
hasil tetua jantan dan betina yang digunakan dalam perakitan
hibrida ……….
102
35 Heterobeltiosis karakter agromorfologi dan komponen hasil
75 hibrida baru …….……… 103
36 Heterosis mid parent karakter agromorfologi dan komponen
hasil 75 hibrida baru ………. 106
37 Heterobeltiosis, mid parent heterosis dan standar heterosis
karakter bobot hasil 75 hibrida baru ……….………….. 111 38 Karakter agronomi dan standar heterosis karakter hasil
Halaman 1 Diagram alur penelitian ... 5
2 Lintasan evolusi dua species padi budidaya (Khush 2000) ... 9
3 Skema sistem galur mandul jantan pada padi ... 13
4 Sidik lintas dan korelasi antar karakter pada galur-galur
dihaploid padi ………. 32
5 Penampilan DH calon galur pelestari yang sangat tahan (kiri) dan rentan (tengah) serta lesi yang diakibatkan HDB pada
tanaman rentan (kanan) ……… 35
6 Ukuran polen steril (kiri atas), polen fertil yang terwarnai oleh IKI 1% (kiri bawah) dan polen steril yang tidak terwarnai oleh
IKI 1% (kanan); pembesaran 40x pada mikroskup binokuler .. 48
7 Penampilan Galur Mandul Jantan Baru yang tahan terhadap hawar daun bakteri, isolat terisolir pada ujung bekas
pengguntingan (lingkaran) ……… 51
8 Eksersi malai galur pelestari (kiri) dan galur mandul jantan
(kanan) ……… 59
9 Eksersi stigma yang tinggi pada Galur Mandul Jantan Baru
(B) dibandingkan dengan eksersi stigma pada IR58025A (A) 63
10 Proses keluarnya polen dari antera saat antesis. (Sumber:
Matsui et al. 1999) ……….. 65
11 Kriteria ideal galur pelestari, meliputi filamen panjang, antera kuning dan besar, antera menggugurkan sebagian besar
polennya setelah berada di luar bunga ……… 67 12 Petak percobaan produksi benih 1 pasang GMJ ………... 77 13 Morfologi stigma GMJ selama reseptif polen ……….. 83 14 Durasi reseptif putik tiga galur mandul jantan baru dan
pembanding (IR58025A) pada 0 ppm GA3 (A) dan 200 ppm
GA3 (B) selama produksi benih galur mandul jantan …………. 84 15 Bobot benih GMJ hasil produksi benih pada konsentrasi GA3
0 dan 200 ppm ……… 84
PENDAHULUAN
Latar Belakang
Kebutuhan terhadap pangan khususnya beras, semakin meningkat sejalan dengan pertambahan jumlah penduduk, sedangkan usaha diversifikasi pangan berjalan lambat. Jumlah penduduk pada tahun 2010, 2015 dan 2020 diprediksikan akan mencapai 235 juta, 249 juta dan 263 juta jiwa. Oleh karena itu laju produksi padi nasional harus terus ditingkatkan. Pada tahun 2009 produksi padi nasional mencapai 64,4 juta ton gabah kering giling (GKG) atau meningkat 6,83% dibandingkan tahun 2008. Namun pada tahun 2010, peningkatan produksi padi nasional lebih rendah, hanya mencapai 2,45% dibandingkan tahun 2009 (BPS 2010). Hal ini disebabkan banyaknya bencana kekeringan, banjir dan juga tingkat serangan hama dan penyakit yang tinggi.
Teknologi padi hibrida yang memanfaatkan gejala heterosis merupakan salah satu cara untuk meningkatkan produktivitas padi di Indonesia. Padi hibrida menghasilkan pertambahan produktivitas mendekati 15-20% lebih tinggi dibanding varietas padi komersial terbaik. China telah sukses mengaplikasikan teknologi padi hibrida. Area yang ditanami padi hibrida di China saat ini terhitung lebih dari 50% dari total area padi. P
enggunaan padi hibrida di China mampu
meningkatkan produksi padi total dari 128 juta ton menjadi 189 juta ton,
walau terjadi penurunan luas lahan padi dari 36,5 juta ha pada 1975
menjadi 29,2 juta ha pada 2007. Peningkatan produktivitas pada periode
tersebut sangat besar yaitu dari 3,5 t/ha menjadi 6,35 t/ha (FAO 2008).
Budidaya padi hibrida juga mulai dilakukan pada skala luas di banyak negara Asia, seperti India dan Vietnam (You et al. 2006).
terutama GMJ yang merupakan tetua betina (seed parent) dan menjadi kunci dalam komersialisasi padi hibrida.
Galur mandul jantan (galur A/GMJ) dapat diperoleh secara spontan di antara galur hasil pemuliaan yang berasal dari persilangan kerabat jauh (interspesifik) atau melalui mutagenesis (Hanson & Bentolila 2004). Ada beberapa tipe galur mandul jantan sitoplasma, antara lain Wild-Abortive (WA), Gambiaca, Kalinga, Dissy, Indonesian-type dan lain-lain. Galur mandul jantan tipe WA merupakan galur yang banyak digunakan dalam pengembangan varietas padi hibrida. Jing et al. (2001) menyebutkan bahwa 90% padi hibrida yang ditanam di Cina menggunakan GMJ tipe WA sebagai tetua betinanya. GMJ tipe WA merupakan GMJ yang dikembangkan pada padi indica dengan sitoplasma yang berasal dari populasi padi liar Oryza rufipogon Griff. (Eckardt 2006). Di Indonesia, GMJ tipe WA tersebut juga digunakan untuk merakit beberapa F1 hibrida unggul (Suwarno et al. 2003). GMJ yang digunakan dalam produksi benih padi hibrida tersebut berasal dari IRRI, seperti IR58025A, IR62829A, IR68886A, IR68888A, dan IR68897A. Namun penggunaan satu tipe sitoplasma yang sama untuk kepentingan komersial secara intensif, dikhawatirkan akan menimbulkan kerapuhan genetik, seperti kasus Texas cytoplasm pada jagung. Kerapuhan genetik disebabkan oleh sempitnya latar belakang genetik, sehingga tanaman menjadi rentan terhadap penyakit, hama maupun faktor-faktor iklim yang tidak menguntungkan (Pradhan & Jachuck 1999).
memiliki ketahanan terhadap hawar daun bakteri (HDB) dan wereng batang coklat (WBC) (Satoto et al. 2008).
Peneliti di BB Padi melaporkan bahwa dari sekitar 1500 tanaman F1 (GMJ x galur-galur elit) yang dievaluasi tingkat fertilitasnya di pertanaman testcross, persentase tanaman dengan sterilitas 100% ternyata sangat kecil, yaitu hanya sekitar 1%, 3% F1 mempunyai fertilitas tinggi, sedangkan 96% lainnya adalah parsial fertil atau parsial steril (Rumanti et al. 2005). Hal ini menunjukkan bahwa transfer sifat mandul jantan tidak mudah, karena sifat mandul jantan dikendalikan oleh interaksi antara sitoplasma yang steril dengan gen-gen male sterile resesif yang ada di dalam nukleus. Perakitan galur mandul jantan memerlukan galur elite yang berpotensi untuk digunakan merakit GMJ. Galur tersebut adalah galur pelestari (maintainer/galur B) yang efektif melestarikan sifat mandul jantan pada GMJ dan sudah beradaptasi dengan lingkungan target. Menurut Virmani et al. (1997), frekuensi ditemukannya galur pelestari diantara galur-galur elite indica (galur indica mendominasi di Indonesia) sangat kecil, yaitu bervariasi antara 0-10%. Pembentukan dan perbaikan galur pelestari dapat dilakukan dengan persilangan B x B yang keduanya mempunyai sifat baik, kemudian dilanjutkan dengan seleksi terhadap rekombinan terbaik pada generasi F2 – F5. Untuk memperoleh mandul jantan baru dari pelestari perlu dilakukan silang balik sebanyak minimal 5 generasi (Virmani et al. 1997). Teknik ini bila dilakukan dengan program pemuliaan biasa, perlu waktu yang sangat lama untuk mendapatkan calon galur pelestari dan galur mandul jantan. Oleh karena itu, kultur antera telah diterapkan untuk pembentukan galur murni yang lebih cepat, efektif dan akurat (Dewi & Purwoko 2011). Dewi et al. (2005) telah memperoleh galur-galur dihaploid hasil kultur antera yang dapat diuji efektivitasnya dalam melestarikan GMJ.
mendukung perolehan nilai heterosis yang tinggi pada padi hibrida. Serangkaian kegiatan penelitian tersebut disajikan pada Gambar 1.
Tujuan Penelitian Tujuan Umum
Penelitian ini bertujuan untuk memperoleh GMJ baru dengan tiga tipe sitoplasma yang memiliki karakter bunga baik dan tahan terhadap penyakit Hawar Daun Bakteri untuk pengembangan padi hibrida.
Tujuan Khusus
Penelitian ini bertujuan untuk :
(1) Mengevaluasi karakter agromorfologi, komponen hasil, hasil dan ketahanan terhadap HDB pada galur dihaploid (DH2) calon galur pelestari
(2) Memperoleh Galur Mandul Jantan tipe WA, Gambiaca dan Kalinga beserta pelestari pasangannya yang tahan terhadap hawar daun bakteri
(3) Mendapatkan GMJ baru yang memiliki morfologi dan perilaku bunga mendukung kemampuan menyerbuk silang alami
(4) Memperoleh informasi potensi produksi benih galur mandul jantan tipe WA, Kalinga dan Gambiaca
(5) Memperoleh informasi efektivitas tiga tipe galur mandul jantan sitoplasmik baru untuk perakitan padi hibrida
(6) Mendapatkan hibrida baru dengan heterosis tinggi
Hipotesis
1. Galur mandul jantan baru dapat diperoleh dari populasi dihaploid turunan kultur antera dari persilangan B x B dengan sumber sitoplasma yang berbeda (WA, Kalinga atau Gambiaca)
2. Galur mandul jantan dengan karakter bunga tertentu, akan meningkatkan potensi hasil pada produksi benih GMJ
3. Terdapat GMJ dengan sterilitas polen 100% dan stabil yang mempunyai daya gabung umum dan khusus yang tinggi terhadap galur pemulih kesuburan, sehingga mampu menghasilkan kombinasi hibrida berheterosis tinggi.
TINJAUAN PUSTAKA
Asal dan Taksonomi Padi
Padi (Oryza sativa L.), seperti halnya gandum, jagung dan barley termasuk dalam famili Graminae (Poaceae) atau rumput-rumputan. Genus Oryza terdiri atas 23 spesies, 21 diantaranya adalah padi liar (wild relatives) yang memiliki genom diploid atau tetraploid, sedangkan 2 lainnya merupakan padi budidaya. Studi variasi karakter morfologi terhadap 16 spesies padi, telah dilaporkan oleh Morishima dan Oka (1960) yang menyatakan bahwa spesies Oryza dapat dibagi ke dalam tiga grup utama, yaitu (1) Oryza sativa dan kerabat dekatnya, (2) Oryza officinalis dan kerabatnya serta (3) spesies kerabat jauh lainnya.
Oryza sativa terdiri atas banyak subspesies dan telah terdistribusi ke seluruh dunia. Dikenal dua grup varietas padi yaitu indica dan japonica, kedua grup ini memiliki banyak perbedaan karakter dibandingkan antar varietas tipikalnya, tetapi variasinya saling overlap. Kedua grup ini telah dikarakterisasi berdasarkan resistensinya, panjang bulu di apikulus dan reaksinya terhadap phenol. Morinaga (1954) dalam Kush (2000) mengusulkan adanya grup ketiga yang terdiri atas varietas bulu dan gundil dari Indonesia. Grup ini disebut javanica, dan Galszmann (1987) dalam Kush (2000) menyatakan bahwa javanica berada di antara grup japonica berdasarkan analisis isozym. Saat ini javanica dikenal sebagai tropical japonica, sedangkan typical japonica disebut sebagai temperate japonica (Gambar 2).
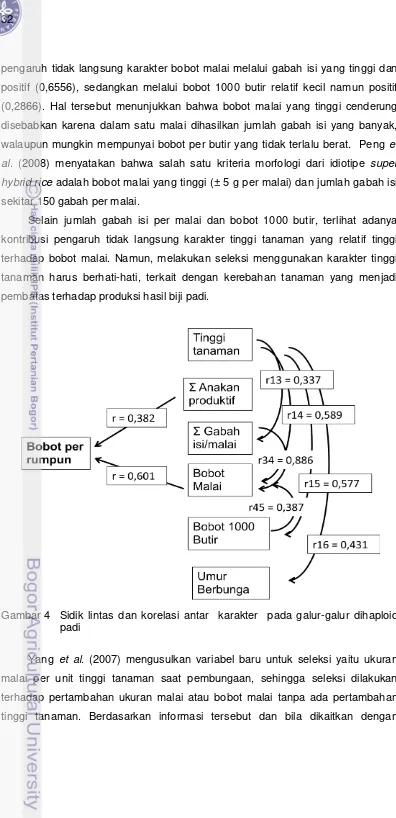
Gambar 2 Lintasan evolusi dua species padi budidaya (Kush 2000)
Padi Hibrida
(You et al. 2006). Oleh karena itu, untuk mendapatkan varietas padi hibrida yang baik dengan sifat-sifat yang diinginkan seperti daya hasil tinggi (ditunjukkan oleh nilai heterosis yang tinggi), dan tahan terhadap hama dan penyakit utama, maka penelitian padi hibrida diutamakan pada proses perbaikan galur-galur tetua padi hibrida serta proses pembentukan kombinasi persilangan yang menghasilkan produksi dan heterosis tinggi.
Padi hibrida dikembangkan dengan memanfaatkan adanya fenomena heterosis yang menyebabkan produksi yang lebih tinggi dibanding galur murni. Beberapa hipotesis telah diajukan untuk menjelaskan dasar genetis dari heterosis. Hipotesis dominansi (Bruce 1910 diacu dalam You et al. 2006) menjelaskan faktor dominan salah satu tetua menutupi gangguan mutasi resesif tetua lain pada populasi F1 heterosigos. Sebaliknya hipotesis overdominan (Shull 1908 diacu dalam You et al. 2006) percaya bahwa heterosigositas pada lokus tunggal lebih superior dibanding yang homosigos. Kedua hipotesis tersebut telah diverifikasi melalui penelitian biologi molekuler (Stuber et al. 1992; Xiao et al. 1995). Hipotesis ketiga menduga bahwa heterosis mungkin akibat terjadinya epistasi antara alel-alel pada lokus yang berbeda (Yu et al. 1997; Goodnight 1999). Beberapa hasil penelitian menunjukkan bahwa epistasis merupakan penyebab utama heterosis secara genetik (Li et al. 2001; Luo et al. 2001).
kerentanan GMJ China terhadap hama dan penyakit (Virmani & Kumar 2004). Oleh karena itu IRRI menitikberatkan penelitian padi hibrida terutama pada perakitan galur-galur tetua padi hibrida dan telah banyak menghasilkan kombinasi hibrida unggul yang telah dilepas sebagai varietas di banyak negara, termasuk Indonesia.
Suksesnya penelitian dan pengembangan padi hibrida di China dan IRRI, mendorong Indonesia mulai mengintensifkan pemuliaan padi hibrida pada tahun 1998, dengan menguji bahan pemuliaan introduksi dan perakitan galur tetua serta membentuk berbagai kombinasi hibrida sendiri (Badan Litbang 2007). Indonesia, dalam hal ini Balai Besar Penelitian Tanaman Padi (BB Padi) berhasil melepas varietas hibrida pertama pada tahun 2002, yaitu Maro dan Rokan. Kedua varietas tersebut merupakan hasil introduksi dari IRRI. Varietas pertama yang menggunakan tetua jantan lokal Indonesia pertama dilepas pada tahun 2007, yaitu Hipa6 Jete. Sampai tahun 2010, di Indonesia telah dilepas 69 varietas hibrida dan 17 diantaranya dilepas oleh BB Padi, sedangkan varietas lainnya dilepas oleh perusahaan swasta nasional maupun multinasional yang ada di Indonesia seperti PT BISI, Kondo, Bangun Pusaka, KNB Mandiri, Makmur Sejahtera, Bayer, DuPont, Primasid, Tri Usaha Sari Tani, Sang Hyang Seri, Sumber Alam Sutra, Biogene Plantation, Metahelix dan Advanta Indonesia. Semua varietas hibrida yang telah dilepas menunjukkan heterosis pada karakter hasil terhadap IR64 dan Ciherang, tetapi sebagian besar varietas tersebut tidak memiliki ketahanan terhadap hama dan penyakit padi di negara tropis.
Selain sistem tiga galur, terdapat dua sistem yang dapat digunakan dalam pengembangan padi hibrida, yaitu sistem dua galur dan sistem satu galur. Sistem satu galur adalah penggunaan sistem apomiktik (tanpa pembuahan), namun sampai saat ini masih dalam skala penelitian. Sistem dua galur melibatkan penggunaan galur mandul jantan TGMS/Temperature-sensitive Genic Male Sterility dan galur fertil. Keuntungan sistem ini adalah (1) tidak memerlukan galur pelestari dalam produksi benih TGMS, (2) semua galur fertil dengan sifat-sifat yang baik dapat digunakan sebagai tetua jantan dalam produksi benih hibrida, dan (3) potensi heterosisnya lebih besar dibanding sistem tiga galur (Virmani et al. 2003).
(Liu et al. 1998), dan di Indonesia sering sekali terjadi pergantian temperatur yang sangat ekstrim, karena itu sistem tiga galur masih merupakan sistem yang dapat diterapkan.
Sistem Mandul Jantan pada Padi
Mandul jantan sitoplasmik adalah kondisi dimana tanaman tidak mampu memproduksi polen/tepung sari fungsional. Mandul jantan merupakan karakter yang diwariskan secara maternal, dan biasanya terkait dengan adanya gangguan pada open reading frame (ORF) di genom mitokondria (Hanson & Bentotila 2004). Pada banyak kasus, termasuk padi, kesuburan mandul jantan dapat dipulihkan oleh gen-gen nukleus yang mengkode fertility restoration (gen Rf) yang terdapat pada galur pemulih kesuburan (Restorer/galur R). Sistem mandul jantan sitoplasmik terjadi karena adanya interaksi antara genom nukleus dan mitokondria. Dalam hal ini, sterilitas akibat gen mitokondrial menyebabkan disfungsi sitoplasmik, sedangkan gen-gen nukleus akan menekan disfungsi sitoplasmik tersebut (Ekcard 2006).
Penggunaan galur mandul jantan merupakan prasyarat untuk eksploitasi heterosis pada padi. Cytoplasmic-genetic male sterility (CgMS/CMS) telah digunakan secara luas untuk mengembangkan padi hibrida. Pistil galur mandul jantan tumbuh normal dan dapat memproduksi biji bila diserbuki oleh polen normal. Jika faktor genetik yang menginduksi kemandulan tersebut tidak ada dalam sitoplasma maka tanaman menjadi normal (male fertile).
Jika terdapat tanaman normal (male fertile) yang memiliki faktor pengendali kemandulan pada sitoplasma dan gen inti yang memulihkan kesuburan bersifat resesif, maka tanaman tersebut dapat mempertahankan sifat mandul jantan. Tanaman atau galur tersebut disebut galur pelestari (maintainer line) yaitu suatu galur yang mempunyai sitoplasma normal tetapi gen inti yang berkaitan dengan pemulihan kesuburan resesif, galur ini berfungsi untuk melestarikan keberadaan galur mandul jantan pasangannya. Adanya gen restorer dominan di nukleus pada suatu galur mengakibatkan galur tersebut mampu memulihkan kesuburan pada hibrida hasil persilangan antara galur tersebut dan galur CMS. Galur ini disebut sebagai galur pemulih kesuburan atau restorer.
crossing) untuk produksi benih skala kecil, maupun melalui persilangan alami pada plot terisolasi untuk produksi benih dalam skala besar. Galur mandul jantan dan galur pelestari selalu sama secara morfologi, hanya galur mandul jantan steril sedangkan galur pelestari fertil. Namun kadang-kadang, kedua galur tersebut juga menunjukkan beberapa sifat morfologi dan agronomis yang berbeda karena dipengaruhi oleh faktor-faktor yang ada di dalam sitoplasma yang menginduksi mandul jantan (Virmani et al. 1997). Restorer yang mempunyai gen pemulih kesuburan (Rf) dominan, ketika disilangkan dengan GMJ akan menghasilkan F1 hibrida yang fertil. Perbanyakan benih galur pelestari dan pemulih kesuburan dilakukan seperti perbanyakan padi biasa, karena kedua galur ini fertil dan mempunyai bunga sempurna.
Gambar 3 Skema sistem galur mandul jantan pada padi
sitoplasma Boro-type termasuk dalam tipe ini; (2) berdasarkan pola pelestarian-pemulih kesuburan dari GMJ, terdapat tiga tipe galur mandul jantan yaitu WA, Honglian dan Boro type (BT); dan (3) berdasarkan morfologi polen, GMJ digolongkan ke dalam tipe typical abortion, spherical abortion, dan stained abortion. GMJ tipe typical abortion mempunyai bentuk polen tidak beraturan dan pengguguran polen biasanya terjadi pada fase uninukleat, sedangkan tipe spherical abortion mempunyai polen berbentuk agak lonjong dan polen gugur kira-kira saat fase binukleat. Terakhir, bentuk polen tipe stained abortion juga agak lonjong tetapi agak lebih kecil dibanding polen normal dan polen gugur saat fase trinukleat (Yuan et al. 2003).
Galur mandul jantan yang berkembang di Indonesia hampir seluruhnya mempunyai sumber sitoplasma yang sama, yaitu Wild Abortive (WA). Apabila galur ini digunakan secara terus-menerus, ada kekhawatiran akan mengakibatkan kerapuhan genetik padi hibrida terhadap hama dan penyakit padi yang kemudian akan menyebabkan ledakan populasi hama dan penyakit, sama seperti yang pernah terjadi pada jagung hibrida (kasus Texas CMS). IRRI telah mengembangkan beberapa galur mandul jantan dengan sumber sitoplasma yang berbeda antara lain Gambiaca, Dissi, ARC, Kalinga, Hong Lian, Indonesian Paddy (IP) dan beberapa sumber sitoplasma dari padi liar seperti Oryza rufipogon dan O. perennis. Namun di Indonesia sumber-sumber sitoplasma di atas belum digunakan dalam pembentukan dan produksi padi hibrida, karena belum teridentifikasi galur pemulih kesuburan yang cocok berkombinasi dengan GMJ dengan sumber-sumber sitoplasma di atas.
Uji Daya Gabung
Keberhasilan setiap program pemuliaan tanaman tergantung pada pemilihan genotipe tetua yang tepat untuk program persilangan, disesuaikan dengan target pemuliaan. Studi daya gabung tetua-tetua yang digunakan dalam program pemuliaan memberikan informasi yang dapat membantu pemulia melakukan seleksi terhadap tetua yang lebih baik untuk digunakan dalam program pemuliaan, sehingga program tersebut menjadi lebih efektif.
dengan penampilan superior. Nilai masing-masing galur terletak pada kemampuannya untuk menghasilkan keturunan unggul bila dikombinasikan dengan galur-galur lain (Allard 1960).
Daya gabung umum (general combining ability) adalah nilai rata-rata galur-galur dalam seluruh kombinasi persilangan bila disilangkan dengan galur-galur-galur-galur lain. Daya gabung umum (DGU) menggambarkan peranan dan interaksi yang kuat antar efek genetik aditif pada populasi dasar, sedangkan efek daya gabung khusus memberi ilustrasi mengenai variasi non-aditif (Falconer 1983). Nilai DGU yang tinggi menunjukkan tetua atau galur yang bersangkutan mempunyai kemampuan berkombinasi dengan baik, sedangkan nilai DGU yang rendah menunjukkan bahwa tetua tersebut mempunyai kemampuan berkombinasi yang lebih buruk dibanding tetua lain. Nilai DGU dapat positif atau negatif, tergantung pada karakter yang diamati dan cara menilainya. Nilai daya gabung dapat memberikan informasi mengenai kombinasi-kombinasi yang dapat menghasilkan turunan yang berpotensi hasil tinggi, yang ditunjukkan oleh diperolehnya heterosis dan nilai daya gabung yang besar.
Daya gabung khusus (specific combining abillity atau DGK) adalah penampilan kombinasi pasangan persilangan tertentu. Nilai daya gabung khusus baik apabila nilai pasangan persilangan tertentu lebih tinggi dibanding nilai rata-rata keseluruhan persilangan yang terlibat (Poehlman & Sleper 1990). Persilangan-persilangan yang memiliki nilai daya gabung khusus tinggi sangat berpotensi untuk dieksploitasi efek heterosisnya dan dapat dikembangkan sebagai varietas hibrida. Evaluasi persilangan antar galur inbred merupakan langkah penting dalam pengembangan padi hibrida. Secara ideal, proses ini sebaiknya dilakukan melalui evaluasi terhadap semua kemungkinan persilangan yang terjadi, sehingga dapat diketahui kombinasi yang sangat sesuai untuk disilangkan agar dapat menghasilkan suatu varietas yang baik.
Abstrak
Perakitan galur mandul jantan sangat penting perannya dalam pembentukan varietas padi hibrida berheterosis tinggi. Untuk memperoleh galur mandul jantan baru, perlu dilakukan seleksi calon galur pelestari dan silang balik berkelanjutan terhadap donor sitoplasmik. Penelitian ini bertujuan untuk (1) mengevaluasi karakter agromorfologi, komponen hasil dan hasil serta karakter bunga galur-galur dihaploid calon galur pelestari hasil kultur antera dan (2) mengevaluasi ketahanan galur-galur dihaploid calon galur pelestari terhadap penyakit hawar daun bakteri. Sembilan belas galur dihaploid generasi kedua (DH2) diuji menggunakan rancangan acak lengkap yang diulang 4 kali. Hasil penelitian menunjukkan bahwa karakter bobot malai, jumlah gabah isi per malai dan hasil biji per rumpun mempunyai nilai KVG tertinggi. Nilai heritabilitas arti luas kategori tinggi ditunjukkan oleh karakter tinggi tanaman, umur berbunga, bobot malai, jumlah gabah isi per malai, bobot 1000 butir dan hasil biji per rumpun. Dari seluruh karakter yang diuji, bobot malai mempunyai kontribusi terbesar terhadap hasil biji per rumpun, terlihat dari nilai pengaruh langsungnya yang positif dan besar (0,7399) dan pengaruh tidak langsungnya melalui jumlah gabah isi per malai yang juga tinggi dan positif (0,6556). Jumlah anakan produktif juga memberikan korelasi yang positif dan tinggi secara langsung terhadap hasil biji per rumpun. Karakter jumlah anakan produktif, bobot malai dan gabah isi per malai dapat digunakan sebagai karakter seleksi galur pelestari yang baik. Semua galur dihaploid yang diuji kecuali B4-1-Da dan B4-1-Dc sesuai untuk digunakan sebagai calon galur pelestari berdasarkan bobot malai yang tinggi (≥ 2,99 g), jumlah gabah isi per malai (>125 butir/malai) dan bobot hasil yang tinggi (>19 g/rumpun). Pada percobaan inokulasi menggunakan inokulum hawar daun bakteri patotipe III, terdapat tiga galur DH pelestari bereaksi tahan (skor 1), diantaranya adalah B1-1-Mb, B2-1-Db dan B2-1-M. Galur B4-1-Dc merupakan galur DH pelestari yang bereaksi sangat tahan (skor 0) terhadap Xoo patotipe IV, dan dua galur DH pelestari menunjukkan reaksi yang sangat tahan terhadap Xoo patotipe VIII, yaitu H36-4-M dan B4-1-Dc (skor 0). Karakter sembilan belas galur DH pelestari bervariasi, sebagian besar memiliki morfologi yang mendukung perolehan hasil tinggi seperti warna daun yang hijau tua, eksersi malai sempurna dan posisi daun bendera yang tegak.
Abstract
Breeding of male sterile lines is important in hybrid rice development. It is done through selection of maintainer candidate lines and successive backcrosses to cytoplasmic donors. The aims of this research were to evaluate several agronomic characters, yield components and yield of doubled haploid maintainer candidate lines, and to study their resistance to three pathotypes of bacterial leaf blight pathogens (Xanthomonas oryzae pv. oryzae (Xoo)). Nineteen dihaploid lines (DH2) were tested using completely randomized design with 4 replications. The results showed that panicle weight, filled grain per panicle and grain weight per plant had the highest genotype coefficient of variance (GCV). High broad sense heritability value was shown by plant height, flowering time, panicle weight, 1000 grain weight and grain per plant. The direct correlation value of panicle weight to yield per plant was high and positive (0.7399). Indirect correlation through filled grain per panicle was also positive and high (0.6556). The number of panicle per plant also gave positive and high correlation directly to grain weight per plant. It was shown that panicle number per plant, panicle weight and filled grain per panicle had the highest contribution to yield per plant, therefore the characters could be used as selection criteria of good maintainer. All of dihaploid lines except B4-1-Da and B4-1-Dc can be used as maintainer candidate lines, based on their panicle weight (≥ 2.99 g), high filled grain per panicle (> 125 grain/panicle) and yield per plant (> 19 g/plant). There were three dihaploid lines resistant to Xoo pathotype III (score 1), i.e. B1-1-Mb, B2-1-Db and B2-1-M. B4-1-Dc were dihaploid lines highly resistant to BLB pathotype IV (score 0), and two lines were highly resistant to pathotype VIII, i.e. H36-4-M and B4-1-Dc (score 0). Morphology of nineteen maintainer candidate lines were varied and suitable for supporting high yield, such as dark green leaves, good panicle exertion and erect flag leaf.
Pendahuluan
Teknik kultur antera merupakan teknologi yang banyak digunakan untuk percepatan perakitan galur murni yang homozigos. Tanaman-tanaman dihaploid yang dihasilkan melalui kultur antera bersifat homozigos penuh dan breed true, karena kedua kopi informasi genetik pada tanaman-tanaman tersebut identik. Individu tanaman yang dihasilkan oleh mikrospora yang sama akan mempunyai karakter agromorfologi yang sama pada generasi selanjutnya (Dewi & Purwoko 2001).
Tanaman homozigos atau galur murni dengan sifat-sifat yang unggul sangat diperlukan dalam pemuliaan tanaman. Tanaman homozigos akan memudahkan seleksi fenotipe terhadap karakter-karakter kuantitatif tanpa disulitkan oleh hubungan dominan resesif seperti pada tanaman heterozigos. Tercapainya galur murni pada keturunan pertama menyebabkan siklus pemuliaan dapat lebih singkat karena dapat menghilangkan sebagian besar kegiatan seleksi per generasi (6-8 generasi) yang umum pada pemuliaan konvensional (Dewi et al. 1996).
Dewi & Purwoko (2011) menyatakan bahwa sangat mudah untuk membedakan tanaman haploid dari dihaploid pada generasi awal (DH0), yaitu melalui perbedaan dalam bentuk tanaman (stature), warna daun, bentuk daun, perkembangan anakan dan akar, pembentukan biji, serta jumlah dan ukuran stomata. Oleh karena itu, evaluasi terhadap karakter target pada tanaman dihaploid hasil kultur antera dapat dilaksanakan dengan mudah menggunakan metode seleksi konvensional. Kriteria agromorfologi dalam pemilihan galur pelestari antara lain malai keluar sempurna dari seludang daun bendera, filamen harus panjang sehingga antera menjulur keluar sempurna dari bunga, antera harus besar dan gemuk dengan butir tepung sari/polen yang banyak, antera harus menggugurkan butir tepungsarinya ketika antera sudah keluar dari bunga (Virmani et al. 2003).
menentukan ukuran populasi yang harus dipelihara saat melakukan seleksi dan menentukan metode alternatif dalam seleksi (Roy 2000).
Kearsey & Pooni (1996) menyatakan bahwa heritabilitas dapat diprediksi menggunakan perhitungan statistika terhadap komponen ragam. Secara umum, heritabilitas merupakan proporsi dari ragam fenotipik total hasil kontribusi dari efek gen. Terdapat dua tipe heritabilitas, yaitu heritabilitas arti luas dan heritabilitas arti sempit. Heritabilitas arti luas merupakan rasio antara nilai variasi genetik total terhadap variasi fenotipik total. Heritabilitas ini menjelaskan mengenai efek-efek dominan dan epistasis. Heritabilitas arti sempit merupakan proporsi dari variasi yang terjadi akibat gen-gen aditif dalam suatu populasi spesifik. Heritabilitas ini memiliki peran yang lebih penting dalam suatu program seleksi karena dapat mengukur nilai estimasi hasil pemuliaan dari suatu populasi (breeding value) dan respon terhadap seleksi sangat tergantung pada varians genetik aditif (Wray & Visscher 2008). Aksi gen memegang peranan penting dalam menentukan nilai heritabilitas. Karakter-karakter yang ekspresinya dipengaruhi oleh gen aditif akan memiliki nilai heritabilitas yang lebih tinggi dibandingkan karakter yang dikontrol oleh gen non-aditif (Falconer 1988).
Hawar daun bakteri merupakan penyakit utama padi yang merusak klorofil daun, sehingga fotosintesis terhambat dan tanaman tidak mampu menghasilkan malai. Salah satu pengendalian yang ramah lingkungan adalah menggunakan varietas tahan. Sebagian besar padi hibrida yang ada di Indonesia masih belum memiliki ketahanan terhadap hawar daun bakteri. Oleh karena itu, skrining terhadap galur-galur tetua perlu dilakukan untuk mengetahui ketahanannya terhadap patogen Xoo.
Percobaan ini bertujuan untuk (1) mengevaluasi karakter agromorfologi, komponen hasil dan hasil serta karakter bunga galur-galur dihaploid calon galur pelestari hasil kultur antera dan (2) mengevaluasi ketahanan galur-galur dihaploid calon galur pelestari terhadap penyakit hawar daun bakteri.
Bahan dan Metode Tempat dan Waktu
Bahan dan Metode
Bahan yang digunakan adalah benih 19 galur tanaman dihaploid generasi dua (DH2) hasil kultur antera F1 turunan dari persilangan antara galur pelestari (B) dengan Galur/Varietas Donor resisten Hawar Daun Bakteri (Tabel 2).
Evaluasi Karakter Agromorfologi Galur Dihaploid Calon Galur Pelestari
[image:45.595.111.503.211.781.2]Percobaan dilakukan mengikuti Rancangan Acak Kelompok yang diulang 4 kali. Perlakuan terdiri atas 19 galur dihaploid generasi kedua (DH2) calon pelestari (Tabel 2). Satu satuan unit percobaan adalah 3 ember, masing-masing berisi dua rumpun tanaman untuk setiap galur. Benih tanaman uji disemai dalam bak berisi tanah sawah. Bibit berumur 21 hari ditanam dalam ember yang berisi tanah sawah. Setiap ember diisi 12 kg campuran tanah sawah dan pupuk kandang, dengan perbandingan 3:1.
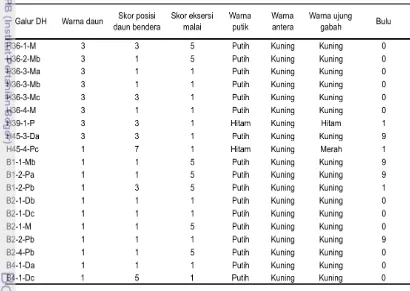
Tabel 2 Galur calon pelestari DH2 hasil kultur antera dari 6 kombinasi persilangan B x B
No No Kode TC Keterangan
Lap.
1 B-1 H36-1-M Kultur antera F1 hasil persilangan
2 B-2 H36-2-Mb H36 : IR72080B-1/Memberamo
3 B-3 H36-3-Ma H39 : IR71562B-1/IR74
4 B-4 H36-3-Mb H45 : IR72080B-20/IR71031
5 B-5 H36-3-Mc
6 B-6 H36-4-M
7 B-8 H39-1-P
8 B-9 H45-3-Da
9 B-10 H45-4-Pc
10 B-11 B1-1-Mb Kultur antera F1 hasil persilangan
11 B-12 B1-2-Pa B1 : IR58025B-W1/Memberamo
12 B-13 B1-2-Pb B2 : IR58025B-W2/Barumun
13 B-14 B2-1-Db B4 : IR68897B-W2/Barumun
14 B-15 B2-1-Dc
15 B-16 B2-1-M
16 B-17 B2-2-Pb
17 B-18 B2-4-Pb
18 B-19 B4-1-Da
19 B-20 B4-1-Dc
pada saat tanaman berumur 40 hari setelah tanam (HST). Pengendalian hama dan penyakit dilakukan saat munculnya gejala serangan hama dan penyakit.
Pengamatan dilakukan terhadap karakter agromorfologi dan karakter bunga calon galur pelestari, antara lain :
A. Karakter agromorfologi tanaman, meliputi:
1. Tinggi tanaman, diukur dari permukaan tanah sampai ujung malai terpanjang (cm); diamati saat stadia masak susu atau menjelang panen.
2. Jumlah anakan, diamati saat stadia bunting.
3. Warna daun, diamati pada stadia vegetatif sampai heading; data berupa skor (1-7) (Tabel 3).
Tabel 3 Skor warna daun berdasarkan SES (IRRI 2002)
Skor Warna
1 Hijau muda/terang
2 Hijau
3 Hijau tua/gelap
4 Hijau dengan ujung daun berwarna ungu 5 Hijau dengan strip ungu di pinggir daun 6 Bercak ungu (ungu bercampur dengan hijau)
7 Ungu
4. Sudut daun bendera, diukur di pangkal daun bendera (dekat collar); data berupa skor (1-7) (Tabel 4).
Tabel 4 Skala sudut daun bendera berdasarkan SES (IRRI 2002) Skor Kriteria
1 Tegak
3 Intermediet
5 Horisontal
7 Terkulai
5. Eksersi malai, diamati posisi kedudukan leher malai terhadap daun bendera; data berupa skoring (1-9), diamati pada stadia masak susu atau menjelang panen (Tabel 5).
Tabel 5 Skala eksersi malai berdasarkan SES (IRRI 2002)
Skala Kriteria
1 Keluar sempurna
3 Keluar (moderate)
5 Agak keluar (leher malai berada tepat pada pangkal daun bendera)
7 Keluar sebagian
9 Tertutup
B. Karakter komponen hasil dan hasil, meliputi:
1. Jumlah gabah per malai, dihitung total keberadaan gabah dalam satu malai, baik hampa maupun bernas.
2. Jumlah gabah isi per malai, dihitung dari jumlah gabah yang bernas dalam satu malai.
3. Jumlah gabah hampa per malai, dihitung jumlah gabah kosong hampa dalam satu malai.
4. Bobot 1000 biji, diamati dengan menimbang 1000 butir gabah bernas yang berkadar air 13 – 14% (gram).
5. Panjang malai, diamati dari pangkal malai sampai ujung malai (cm) 6. Bobot malai, diamati dengan menimbang malai utama (gram).
C. Karakter bunga, meliputi:
1. Warna putik, diamati saat bunga mekar yaitu sekitar jam 9 – 10 pagi (udara cerah)
2. Warna antera, diamati saat bunga mekar yaitu sekitar jam 9 – 10 pagi (udara cerah)
3. Warna ujung gabah, diamati saat menjelang panen;
4. Umur berbunga, diamati jumlah hari saat tanaman berbunga 50%
5. Keberadaan bulu pada ujung gabah, data berupa skoring (1-9) (Tabel 6).
Tabel 6 Skor keberadaan bulu pada gabah
Skor Kriteria
0 Tidak ada
1 Pendek dan kadang berbulu
5 Pendek dan banyak bulu
7 Panjang dan kadang berbulu
9 Panjang dan banyak bulu
D. Bobot produksi per rumpun, diamati dengan menimbang total gabah isi (kadar air 14%) dalam satu rumpun (gram).
Evaluasi Ketahanan Galur-Galur Dihaploid Calon Galur Pelestari terhadap Hawar Daun Bakteri
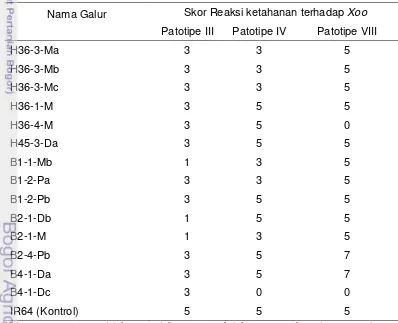
menggunakan gunting yang telah dicelupkan ke dalam larutan bakteri patotipe III, IV dan VIII. Setiap patotipe diujikan pada sembilan rumpun tanaman, yang setiap rumpunnya digunting sebanyak lima helai daun, sehingga jumlah contoh seluruhnya meliputi 45 helai daun. Pada pengujian ini digunakan empat belas galur dihaploid calon pelestari. Keempat belas galur dihaploid tersebut telah teridentifikasi mempunyai kemampuan melestarikan kemandulan galur mandul jantan. Pembanding rentan digunakan IR64 dan sebagai pembanding tahan digunakan Angke dan Conde. Pengamatan dilakukan dua minggu setelah inokulasi, menggunakan skala sesuai kriteria pada Standard Evaluation System for Rice (IRRI 2002) (Tabel 7).
Tabel 7 Kriteria penilaian ketahanan terhadap hawar daun bakteri (HDB).
Nilai Skala Gejala Serangan Tingkat ketahanan
0 Tidak ada gejala Sangat Tahan (ST)
1 keparahan 1-5% Tahan (T)
3 keparahan >5 – 12% Agak Tahan (AT)
5 keparahan >12 – 25% Agak Rentan (AR)
7 keparahan > 25-50% Rentan (SR)
9 keparahan > 50-100% Sangat Rentan
Berdasarkan skala keparahan penyakit tersebut kemudian dihitung indek penyakit HDB dengan rumus sebagai berikut :
n(1)+n(2)+n(3)+n(4)+n(5)+ n(6)+n(7)+n(8)+n(9) Di = ---
ZN Keterangan:
Di = indeks penyakit HDB
n = jumlah tanaman terserang HDB dengan skala tertentu Z = total rumpun yang diamati
N = skala keparahan tertinggi
Analisis Data
Sebelum dilakukan analisis data, guna melengkapi data maka dilakukan perhitungan sederhana terhadap data gabah isi menjadi satu karakter baru, yaitu persentase gabah isi per malai (spikelet fertility) yang dihitung menggunakan rumus sebagai berikut:
jumlah gabah isi per malai
Selanjutnya, semua data pengamatan dianalisis menggunakan bantuan SAS 9.0 untuk menghitung:
1. Analisis ragam (anova) yang dilanjutkan dengan uji Duncan Multiple Range Test (DMRT)
Tabel 8 Analisis ragam (anova) rancangan acak kelompok
Sumber keragaman
Derajat bebas Kuadrat tengah Estimasi kuadrat tengah
Ulangan (r) r – 1 M1 σ2e + gσ2r
Genotipe (g) g – 1 M2 σ2e + rσ2eg
Galat (e) (r - 1)(g - 1) M3 σ2e
2. Parameter genetik dan nilai heritabilitas dalam arti luas untuk mengetahui besarnya sumbangan pengaruh genetik terhadap fenotipe. Pendugaan parameter genetik dan nilai heritabilitas dihitung berdasarkan analisis ragam menurut metode yang dikemukakan oleh Singh & Chaudhary (1979). Berdasarkan sumber keragaman pada Tabel 8, maka parameter genetik dan nilai heritabilitas arti luas dihitung menggunakan rumus sebagai berikut:
√
√
Keterangan: : ragam genotipik, : ragam fenotipik, KVG: koefisien keragaman genotipik; KVP= koefisien keragaman fenotipik, h2(bs) : heritabilitas arti luas
Kriteria nilai heritabilitas arti luas ditetapkan berdasarkan Stansfield (1983), yaitu:
0,50 < h2≤ 1,00 = tinggi
0,20 ≤ h2≤ 0,50 = sedang
3. Analisis Sidik Lintas (Path Coefficient Analysis) untuk mengetahui pengaruh langsung dan tidak langsung dari karakter morfologi dan komponen hasil terhadap hasil. Analisis lintas dihitung berdasarkan persamaan simultan yang dikemukakan oleh Singh & Chaudhary (1979), sebagai berikut:
[
] [ ] [
]
Rx Ci Ry
Berdasarkan persamaan di atas, nilai Ci (pengaruh langsung) dapat dihitung menggunakan rumus:
Keterangan:
Rx = matriks korelasi antara peubah bebas, = invers matriks Rx
Ci = vektor koefisien lintasan yang menunjukkan pengaruh langsung setiap peubah bebas yang telah dibakukan terhadap peubah tak bebas Ry = vektor koefisien korelasi antar peubah bebas Xi (i = 1, 2, ..., p).
Hasil dan Pembahasan
Karakter Agromorfologi dan Bunga Galur Dihaploid Calon Galur Pelestari
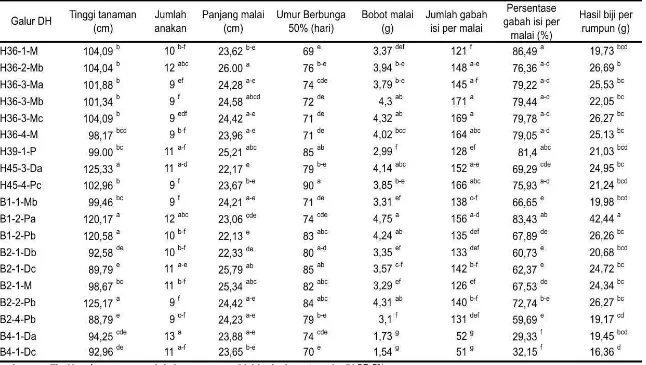
Karakter agromorfologi dan karakter bunga semua calon galur pelestari ditampilkan pada Tabel 9. Warna daun 19 calon galur pelestari terbagi ke dalam dua kelompok yaitu hijau muda dan hijau tua. Posisi daun bendera terhadap batang atau malai merupakan komponen karakter morfologi yang penting, sesuai idiotype dari galur-galur padi yang mendukung fotosintesis. Peng et al. (2008) mengemukakan bahwa posisi daun bendera yang terbaik adalah tegak (erect). Calon galur pelestari didominasi oleh galur-galur yang masuk dalam kategori tersebut. Posisi daun bendera demikian dapat meningkatkan besarnya intersepsi cahaya matahari pada tiga daun teratas yang akan mendukung proses fotosintesis dan akumulasi asimilat, sehingga akan meningkatkan proses pengisian biji.
malai yang terbungkus oleh daun bendera dengan kisaran 11-25% atau skor 5, sedangkan 12 genotipe lainnya mempunyai malai yang muncul sempurna dari pelepah daun bendera.
[image:51.595.110.520.309.600.2]Warna putik dan ujung gabah dapat digunakan sebagai salah satu karakter seleksi saat produksi benih padi. Sebagian besar genotipe uji memiliki putik yang berwarna putih, hanya dua genotipe yang memiliki putik berwarna hitam yaitu H39-1-P dan H45-4-Pc. Warna antera yang kuning dan bentuknya yang gemuk (plumpy) menunjukkan bahwa antera genotipe-genotipe tersebut mengandung banyak polen fertil yang siap digunakan dalam penyerbukan, baik untuk pembentukan benih dirinya sendiri maupun produksi benih galur mandul jantan.
Tabel 9 Karakter morfologi dan karakter bunga 19 calon galur pelestari
Keterangan: skor warna daun: 1= hijau terang, 3= hijau gelap; skor posisi daun bendera: 1= tegak, 3= agak tegak, 5= horizontal, 7= terkulai; skor eksersi malai: 1= keluar penuh, 5= berada tepat di pangkal pelepah daun bendera; skor bulu: 0= tidak ada, 1= pendek dan kadang berbulu, 9= panjang dan banyak bulu.
Parameter genetik
Pendugaan komponen ragam dan heritabilitas dilakukan untuk mengetahui proporsi keragaman yang disebabkan oleh faktor genetik dan lingkungan. Heritabilitas sangat menentukan keberhasilan seleksi untuk lingkungan yang sesuai, karena heritabilitas dapat memberikan gambaran suatu sifat dipengaruhi oleh faktor genetik atau lingkungan.
Nilai pendugaan parameter genetik tanaman ditampilkan pada Tabel 10. Nilai koefisien variasi genotipik (KVG) dan fenotipik (KVP) tanaman berkisar antara 3,19-23,63% dan 3,97-27,58%. Dari nilai KVG absolut 0-23,63% ditetapkan nilai relatifnya. Nilai absolut 23,63% sebagai nilai relatif 100%. Kriteria KVG relatif adalah rendah (0 < x < 25%), agak rendah (25% < x < 50%), cukup
tinggi (50% < x < 75%), dan tinggi (75% < x ≤ 100%) (Moedjiono & Mejaya 1994). Jadi nilai absolut kriteria tersebut adalah rendah (0,0% < x < 5,91%), agak rendah (5,91% < x < 11,82%), cukup tinggi (11,82% < x < 17,72%) dan tinggi (17,72% < x ≤ 23,63%). Kriteria nilai absolut untuk KVP adalah rendah (0 < x < 6,90%), agak rendah (6,90% < x < 13,79%), cukup tinggi (13,79% < x < 20,69%) dan tinggi (20,69% < x ≤ 27,58%).
Tabel 10 Nilai rata-rata dan parameter genetik dari delapan karakter pada 19 galur dihaploid calon galur pelestari.
Karakter
Rata-rata
h2 (bs) (%)
KVG (%)
KVP (%)
Tinggi tanaman (cm) 103,33 126,22 139,63 90,40 10,87 11,44
Jumlah Anakan 10,13 1,15 3,25 35,50 10,60 17,78
Umur berbunga (hari) 78,19 37,58 39,21 95,85 7,84 8,01
Panjang malai (cm) 24,05 0,77 2,63 29,18 3,64 6,75
Bobot malai (g) 3,57 0,66 0,82 80,18 22,77 25,42
Jumlah gabah isi/malai 135,05 1018,15 1281,16 79,47 23,63 26,50
Bobot 1000 butir (g) 21,58 0,47 0,73 64,59 3,19 3,97
Hasil biji per rumpun (g) 23,80 25,28 43,10 58,65 21,13 27,58
Keterangan: = ragam genotipik; = ragam fenotipik; h2 (bs)= heritabilitas arti luas; KVG=
koefisien keragaman genotipik; KVP= koefisien keragaman fenotipik
digolongkan sebagai karakter variabilitas genetik luas (Murdaningsih et al. 1990). Berdasarkan kriteria tersebut, terdapat lima karakter dengan koefisien variasi genotipik tergolong rendah dan agak rendah, yaitu tinggi tanaman, jumlah anakan, umur berbunga, panjang malai dan bobot 1000 butir, sedangkan tiga karakter tergolong tinggi yaitu bobot malai, jumlah gabah isi per malai dan hasil biji per rumpun. Dengan demikian, terdapat lima karakter variabilitas rendah dan tiga karakter variabilitas luas. Hal ini berarti terdapat peluang perbaikan genetik terhadap karakter hasil melalui karakter bobot malai, jumlah gabah isi per malai, dan hasil biji per rumpun. Hal ini sejalan dengan hasil penelitian Lestari & Nugraha (2007) yang menyebutkan bahwa peluang perbaikan genetik terhadap hasil biji galur padi hasil kultur antera dapat dilakukan melalui seleksi antara lain terhadap karakter jumlah gabah per malai dan hasil. Variabilitas genetik luas berarti bahwa seleksi terhadap karakter tersebut berlangsung efektif dan mampu meningkatkan potensi genetik karakter pada generasi selanjutnya (Zen & Bahar 2001). Seleksi dapat dilakukan lebih leluasa pada karakter yang mempunyai variabilitas genetik luas dan dapat digunakan dalam perbaikan genotipe.
Nilai duga heritabilitas arti luas terhadap karakter yang diamati berkisar antara 29,18% untuk panjang malai dan 95,85% untuk umur berbunga. Berdasarkan kriteria Stanfield (1983), nilai heritabilitas karakter jumlah anakan dan panjang malai tergolong sedang, sedangkan karakter tinggi tanaman, umur berbunga, bobot malai, jumlah gabah isi per malai, bobot 1000 butir dan hasil biji per rumpun tergolong tinggi. Tidak satupun karakter yang diuji mempunyai nilai heritabilitas rendah. Karakter yang mempunyai nilai heritabilitas tinggi menunjukkan bahwa faktor genetik lebih dominan terhadap karakter yang ditampilkan tanaman karena faktor genetiknya memberi sumbangan yang lebih besar dibanding faktor lingkungan dan seleksi terhadap karakter ini efektif dilakukan pada generasi awal (Wicaksana 2001; Lestari & Nugraha 2007). Seleksi akan lebih efektif bila ditunjang oleh nilai KVG dan heritabilitas yang tinggi (Zen 2002). Pada penelitian ini, karakter bobot malai, jumlah gabah isi per malai dan hasil biji per rumpun mempunyai nilai heritabilitas tinggi dan variabilitas luas. Keadaan demikian sangat menunjang keberhasilan dan kemajuan seleksi yang efektif (Bahar et al. 2000).
bila ditambahkan sejumlah teknologi budidaya seperti pemupukan dan pengaturan jarak tanam.
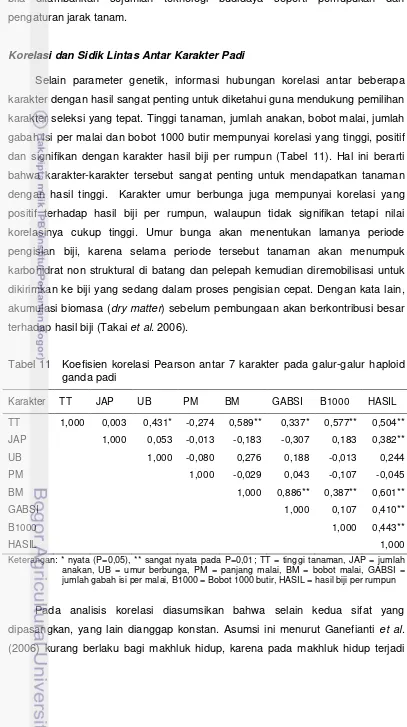
Korelasi dan Sidik Lintas Antar Karakter Padi
Selain parameter genetik, informasi hubungan korelasi antar beberapa karakter dengan hasil sangat penting untuk diketahui guna mendukung pemilihan karakter seleksi yang tepat. Tinggi tanaman, jumlah anakan, bobot malai, jumlah gabah isi per malai dan bobot 1000 butir mempunyai korelasi yang tinggi, positif dan signifikan dengan karakter hasil biji per rumpun (Tabel 11). Hal ini berarti bahwa karakter-karakter tersebut sangat penting untuk mendapatkan tanaman dengan hasil tinggi. Karakter umur berbunga juga mempunyai korelasi yang positif terhadap hasil biji per rumpun, walaupun tidak signifikan tetapi nilai korelasinya cukup tinggi. Umur bunga akan menentukan lamanya periode pengisian biji, karena selama periode tersebut tanaman akan menumpuk karbohidrat non struktural di batang dan pelepah kemudian diremobilisasi untuk dikirimkan ke biji yang sedang dalam proses pengisian cepat. Dengan kata lain, akumulasi biomasa (dry matter) sebelum pembungaan akan berkontribusi besar terhadap hasil biji (Takai et al. 2006).
Tabel 11 Koefisien korelasi Pearson antar 7 karakter pada galur-galur haploid ganda padi
Karakter TT JAP UB PM BM GABSI B1000 HASIL
TT 1,000 0,003 0,431* -0,274 0,589** 0,337* 0,577** 0,504**
JAP 1,000 0,053 -0,013 -0,183 -0,307 0,183 0,382**
UB 1,000 -0,080 0,276 0,188 -0,013 0,244
PM 1,000 -0,029 0,043 -0,107 -0,045
BM 1,000 0,886** 0,387** 0,601**
GABSI 1,000 0,107 0,410**
B1000 1,000 0,443**
HASIL 1,000
Keterangan: * nyata (P=0,05), ** sangat nyata pada P=0,01; TT = tinggi tanaman, JAP = jumlah anakan, UB = umur berbunga, PM = panjang malai, BM = bobot malai, GABSI = jumlah gabah isi per malai, B1000 = Bobot 1000 butir, HASIL = hasil biji per rumpun
[image:54.595.77.484.93.820.2]berbagai proses yang saling berkaitan antara satu dengan yang lain, sehingga analisis korelasi tidak dapat digunakan untuk menggambarkan besarnya sumbangan dari suatu peubah terhadap peubah yang lain. Hal ini dapat diatasi dengan menggunakan analisis lintasan (sidik lintas), karena masing-masing sifat yang dikorelasikan dengan hasil dapat diurai menjadi pengaruh langsung dan tidak langsung.
Tabel 12 Pengaruh langsung dan tidak langsung karakter vegetatif dan komponen hasil terhadap hasil biji per rumpun pada galur-galur dihaploid padi
Keterangan: TT= tinggi tanaman, JA= jumlah anakan, UB= umur berbunga 50%, PM= panjang malai, BM= bobot malai, GABSI= jumlah gabah isi per malai, B1000B= bobot 1000 butir