BI OD EGRAD ASI AEROBI K SEN YAW A H I D ROKARBON AROM ATI K M ON OSI KLI S OLEH BAKTERI
D W I SURYAN TO
Pr ogr a m St u di Biologi
Fa k u lt a s M a t e m a t ik a da n I lm u Pe n ge t a h u a n Ala m Un ive r sit a s Su m a t e r a Ut a r a
I . PEN D AH ULUAN
Perput aran k arbon di alam t ergant ung reak si k at abolik m ik roorganism e. Biodegradasi hidrokarbon ini m erupak an proses k om plek s, y ang aspek k uant it at if dan k ualit at ifny a t ergant ung k epada sifat alam i dan j um lah hidrokarbon t ersebut , k ondisi lingk ungan, dan k om ponen k om unit as m ik roba ( Leahy and Colw ell, 1990; Nicholson et al., 1992) . Kapasit as m ik roorganism e unt uk m endegradasi secara alam i bahan organik yang t elah dilakukan j ut aan t ahun, sekarang dit ant ang dengan bahan kim ia sint et ik yang dengan sengaj a at aupun t idak sengaj a dim asuk k an k e dalam lingk ungan ( Port ier, 1991; Sem ple and Cain, 1996) .
Biorem ediasi m erupakan t eknik yang pot ensial unt uk m em bersihkan daerah t erkont am inasi bahan pencem ar ( Blasco et al., 1997; Laine and Jorgensen, 1996) . Tek nologi biorem ediasi secara sederhana m erupak an usaha unt uk m engopt im alkan kem am puan alam i m ikroorganism e unt uk m endegradasi/ m endaur ulang dengan m em berikan reak t an anorganik esensial dan m em inim um k an t ek anan abiot ik ( Port ier, 1991) . Tek nologi ini sangat berguna dan dapat digunak an pada berbagai t ahapan perlak uan. Terdapat t iga prinsip dalam t ek nologi biorem ediasi, y ait u pelepasan langsung m ik roba k e lingkungan t erkont am inasi, peningkat an kem am puan m ikroba indigenous ( asli) , dan penggunaan m ik roba dalam reak t or k husus ( Port ier, 1991) .
Biodegradasi hidrokarbon oleh k om unit as m ik roba t ergant ung pada k om posisi k om unit as dan respon adapt if t erhadap k ehadiran hidrokarbon ( Leahy and Colw ell, 1990) . Laj u biodegradasi seny aw a hidrokarbon k om plek s dengan berat m olekul besar sepert i senyaw a arom at ik, resin, dan asfalt en lebih lam bat dibandingk an dengan seny aw a dengan berat m olek ul rendah. Meski dem ik ian beberapa st udi m enunj ukkan bahw a degradasi pada kondisi opt im um t erhadap senyaw a kom pleks m em iliki laj u yang t inggi ( Leahy and Colw ell, 1990) . Dem ik ian j uga dengan fenol dan klorofenol ( Nicholson et al., 1992) .
Salah sat u bahan pencem ar yang sering m enim bulkan m asalah adalah hidrokarbon arom at is. Hidrokarbon y ang sering dij um pai, t erut am a di perairan, adalah fenol dan deriv at ny a dari k arbonisasi bat ubara, bahan k im ia sint et ik, dan indust ri m iny ak ( Sem ple and Cain, 1996) . Seny aw a fenolik ini m erupak an polut an berbahay a ( Dong et al. 1992) . Fenol alam i dapat dij um pai di berbagai t anam an. Tanin m erupakan suat u kelom pok senyaw a polifenolik yang biasanya m erupakan kom ponen t um buhan, dan t erdiri dari 2 kelas ut am a, yait u yang t erkondensasi dan hidrolisat . Disam ping it u t um buhan m enghasilkan lignin yang m erupak an k elom pok polifenol sek erabat dengan t anin y ang sangat sulit didegradasi oleh bak t eri ( Gam ble et al., 1996) .
beberapa dek ade t erak hir, sering t erak um ulasi dalam sedim en anaerobik , t anah, dan lingk ungan perairan ( Kuo and Gent hner, 1996) . Klorofenol m isalny a t erdapat dalam lim bah cair pulp dan dari proses lain. Senyaw a ini dapat m engkont am inasi berbagai t anah dan air baw ah t anah ( Laine and Jorgensen, 1996; Mohn and Kennedy , 1992) .
Degradasi fenol dan hom ologny a dilak uk an oleh berbagai organism e berupa bak t eri, j am ur, k apang, ganggang, dan t um buhan t ungk at t inggi ( Sem ple and Cain, 1996) . Penget ahuan t ent ang j alur biot ransform asi m erupak an hal pent ing unt uk m elihat resik o pada daerah t erkont am inasi dan penerapan perlak uan biologi. Bagaim anapun, j alur y ang diam at i di laborat orium dengan organism e t anpa ak lim at isasi sering berbeda dengan y ang diobservasi di lapangan at au pada proses perlakuan dengan konsorsium m ikroba yang dipaparkan dalam kont am inan unt uk w akt u yang lam a ( Nicholson et al., 1992) .
Pada k asus k loroarom at ik , alasan lam bat ny a at au t idak adany a biodegradasi dalam lingk ungan disebabk an oleh j um lah y ang t idak m em adai dari m ikroba pendegradasi poliklorofenol ( Blasco et al., 1997; Miet hling and Karlson, 1996) dan inhibisi oleh k onsent rasi t ok sik seny aw a ini, at au oleh k ont am inan lain ( Heipieper et al., 1992; Miet hling and Karlson, 1996) . Nam un k adang- k adang m ikroba khusus yang diint roduksikan bekerj a t idak sesuai dengan harapan, karena fakt or sepert i ket ahanan ( sur vival) rendah, predasi, dan pengat uran k em am puan degradasi y ang t idak baik ( Blasco et al., 1997) .
I I . PROSES D EGRAD ASI
Degradasi seny aw a fenol dapat dilak uk an lebih m udah dibandingk an dengan senyaw a hasil sint et ik derivat at au hom olog arom at is. Hal ini lebih disebabk an k arena seny aw a ini t elah lebih lam a dik enali bak t eri pendegradasi sehingga bak t eri m am pu m endegradasi j auh lebih baik dibandingk an dengan dengradasi seny aw a deriv at sint et ikny a.
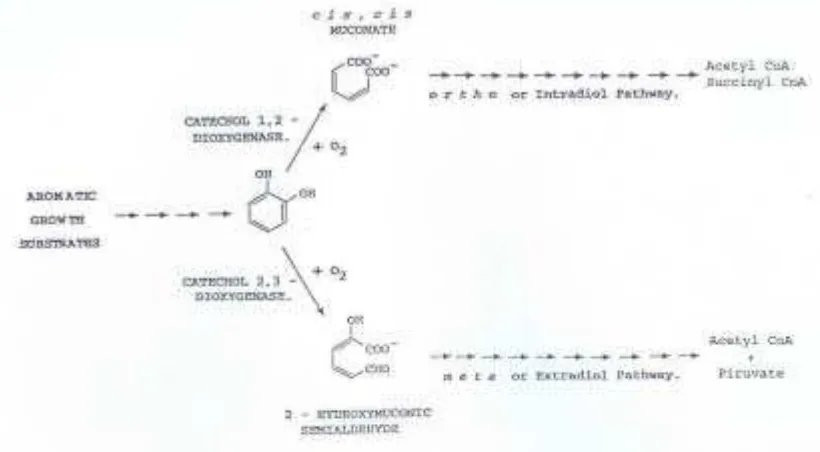
Proses pem ecahan fenol dan m ineralisasi dilakukan berbagai organism e m elalui dest abilisasi cincin arom at is fenol. Senyaw a fenol m engalam i oksidasi dengan bant uan enzim dioksigenase- cincin ( ring- dioxygenase) m enghasilkan dihidrodiol. Seny aw a k at ek ol ( dihy dric phenol) dihasilkan dari seny aw a dihidrodiol dehidrogenase. Melalui pem ecahan ort o dengan enzim k at ek ol 2,3-dioksigenase m enghasilkan cis- cis- m ukonat , at au pem ecahan m et a dengan enzim k at ek ol 2,3- dioksigenase, seny aw a k at ek ol diubah m enj adi hidroksi m uk onat sem ialdehid, dan pem ecahan lain. Hasil m et abolit ini dapat m asuk k e siklus TCA. Beberapa hom olog fenol j uga m em punyai j alur reaksi yang sam a sebelum m asuk siklus TCA ( Gam bar 1) .
Kem am puan degradasi m ik roba t erhadap seny aw a fenol dipengaruhi oleh beberapa fak t or sepert i j enis m ik roba, proses ak lim at isasi, seny aw a t ok sik, dan t oleransi m ik roba t erhadap seny aw a t ok sik. Beberapa m ik roba t ercat at m am pu m endegradasi fenol dengan baik. Ganggang euk aryot , Ochrom onas danica, m am pu t um buh pada fenol sebagai sat u- sat uny a sum ber k arbon. Ganggang ini m engoksidasi fenol dan m em ineralisasi fenol m enj adi kat ekol m elalui pem belahan m et a. Konversi fenol m enghasilkan CO2 sebany ak 60% , 15% t et ap
dalam m edium cair, dan sisany a dik onv ersi m enj adi biom assa ( Sem ple and Cain, 1996) . Jam ur Ceriporiopsis subverm ispora dan Cyat hus st ercoreus m am pu m endegradasi senyaw a t annin ( Gam ble et al., 1996) .
pencucian, akt ifit as gunung api, dan t ransform asi m ikroba) dan oleh kegiat an m anusia ( lim bah indust ri, pem buangan sam pah, dan pem bakaran bahan bakar fosil) m enyebabkan akum ulasi logam dalam relung lingkungan yang anaerobik ( Kuo and Gent hner, 1996) . Keadaan ini m em buat perluny a dik et ahui k em am puan m ik roba unt uk m endegradasi seny aw a arom at ik di daerah y ang j uga t ercem ar logam berat . Pert um buhan bersam a ant ara pereduk si Cr( VI ) , Escherichia coli ATCC 33456, dan pendegradasi fenol, Pseudom onas put ida DMP- 1, secara sim ult an m ereduk si Cr( VI ) dan m endegradasi fenol ( Shen and Wang, 1995) . Penam bahan Cr( VI ) sebany ak 0.01 ppm m eningk at k an biodegradasi fenol sam pai 179% dan benzoat sam pai 169% , sedang penam bahan Cd( I I ) dan Cu( I I ) sebany ak 0.01 ppm m eningk at k an laj u biodegradasi benzoat sam pai 185% dan 2- k lorofenol sam pai 168% . Unt uk Hg( I I ) 1.0- 2.0 ppm , 2- k lorofenol dan 3-k lorobenzoat t erdegradasi 133- 154% lebih cepat daripada 3-k ont rol set elah periode ak lim at isasiny a diperpanj ang ( Kuo and Gent hner, 1996) . Peningk at an t oleransi sel m elaw an subst rat beracun dapat m eningk at k an k em am puan degradasi bahan pencem ar oleh m ik roba t erkait . Perubahan k om posisi lem ak m em bran dari cis m enj adi t rans m enyebabkan peningkat an deraj at sat urasi lem ak m em bran. Modifikasi ini berhubungan dengan peningkat an t oleransi m em bran t erhadap seny aw a t ok sik, sepert i fenol dan k lorofenol ( Heipieper et al., 1992) .
Gam bar 1. Met abolism e senyaw a arom at ik um um nya m elalui kat ekol sebagai int erm ediet sent ral.
Beberapa deriv at arom at is at au hom olog fenol j uga m am pu didegradasi oleh m ikroba. St rain bakt eri MVI , suat u kelom pok bakt eri Gram - negat if dan basilus aerobik , y ang diisolasi dari lum pur y ang diperkay a y ang diam bil dari t em pat pengolahan air lim bah pabrik plast ik m em perlihat k an k em am puan m endegradasi bisfenol A. Sebanyak 60% bisfenol A t erm ineralisasi m enj adi CO2, dan 20%
[image:3.612.116.526.341.567.2]4-hidroksiaset ofenon unt uk kem udian dim ineralisasi dan diasim ilasi m enj adi karbon dalam sel. Dua puluh persen lainny a dihidroksilasi m em bent uk 2,2- bis( hidroksifenil) - 1- propanol, k em udian dit ransform asi m enj adi 2,3- bis( 4-hidroksifenil) - 1,2- propanediol. Sel y ang dit um buhk an pada bisfenol A t erny at a m am pu m endegradasi j uga bisfenol alk ana, asam benzoat t erhidrok silasi, dan aset ofenon t erhidroksilasi ( Lobos et al., 1992) . Selam a degradasi difenil et er yang dilakukan oleh bakt er i Sphingom onas sp. st rain SS3 t erbent uk int erm ediet fenol dan kat ekol yang kem udian m enuj u j alur 3- oksoadipat . Bakt eri ini j uga m am pu m enggunak an deriv at 4- floro, 4- k loro, dan sedik it 4- brom o dari difenil et er sebagai sat u- sat uny a sum ber k arbon dan energi. Langk ah inisiasi degradasi m engikut i m ekanism e 1,2- dioksigenase yang m enghasilkan fenolat hem iaset al y ang t idak st abil dari st ruk t ur difenil ( Schm idt et al., 1992) . Rhodobact er
capsulat us E1F1, bakt eri non sulfur ungu fot ot rofik yang m am pu
m em fot oasim ilasi nit rat at au nit rit , t um buh secara fot ot rofik pada m edium dengan m ono dan dinit rofenol dengan aset at sebagai sum ber k arbon. Pert um buhan t erbesar diperoleh pada kondisi m ikroaerobik ( Blasco and Cast illo, 1992) .
Pada kasus biodegradasi senyaw a arom at ik seringkali t erbent uk int erm ediet y ang lebih t ok sik dari seny aw a asli. Karena t ingk at k elarut an y ang t inggi m enyebabkan senyaw a ini m udah m enyebar. Oleh sebab it u, proses m ineralisasi harus m erupak an t uj uan ak hir dari degradasi seny aw a arom at is, bukan hanya sekedar t elah t erj adi konversi senyaw a ini ( Blasco et al., 1997; Laine and Jorgensen, 1996) .
Pada seny aw a k loroarom at is, m ineralisasi biasany a dilak uk an oleh enzim m elalui j alur klorokat ekol. Sayangnya hanya sedikit bakt eri yang m am pu m ent ransform asi klorofenol m enj adi klorokat ekol unt uk kem udian m enuj u proses m ineralisasi ( Blasco et al., 1997) . Reduk si dehalogenasi k elihat anny a m erupak an langkah inisiasi dalam degradasi anaerobik seluruh klorofenol ( Mohn and Kennedy , 1992; Nicholson et al., 1992) . Reduk si ini m em iliki nilai pent ing t erhadap lingkungan karena produk m et abolik yang lebih sedikit m engandung k lorin um um ny a k urang beracun dan lebih m udah didegradasi oleh bak t eri aerob dibandingkan dengan senyaw a induk yang m em iliki klorin lebih banyak ( Nicholson et al., 1992) . Nicholson et al. ( 1992) j uga m encat at bahw a reduk si dek lorinasi t erj adi pada lum pur buangan anaerobik y ang t idak diak lim at isasi dan yang diaklim at isasi, sedim en, t anah yang dit am bah dengan lum pur buangan, dan lingkungan perairan.
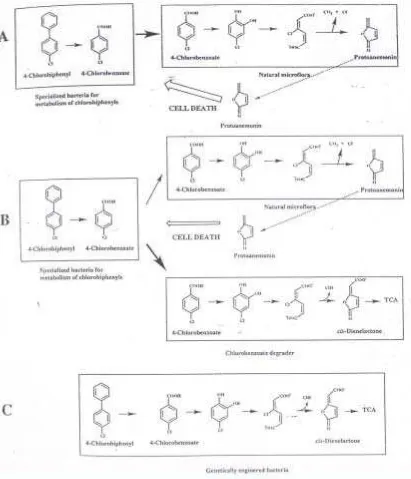
Jalur lain dalam degradasi hom olog fenol selain j alur klorokat ekol dapat saj a t erj adi m elalui pem belahan m et a dan 3- oksoadipat yang m enghasilkan prot oanem onin ( Gam bar 2) , suat u int erm ediet y ang lebih t ok sik daripada senyaw a induk. Prot oanem onin m erupakan suat u senyaw a ant ibiot ik spekt rum luas yang biasanya dihasilkan oleh t um buhan keluarga Ranunculaceae ( Blasco et
al., 1997) . Pem bent uk an prot oanem onin ini dibuk t ikan dengan percobaan
adanya akum ulasi senyaw a t oksik berupa prot oanem onin yang dibent uk oleh m ikroorganism e indigenous ( Blasco et al., 1997) .
[image:5.612.169.472.133.549.2]I nokulasi LB400 bersam a dengan Pseudom onas PS121 y ang m am pu m endegradasi 4- klorobenzoat m elalui 4- klorokat ekol dan j alur ort o ke dalam t anah t idak dist erilisasi m enunj ukkan t idak adanya penurunan viabilit as LB400. Hal y ang sam a j uga t erlihat pada k om binasi LB400 dengan P. put ida KT2442 yang m em iliki plasm id TOL yang m am pu m erubah 4- klorobenzoat m enj adi 5-k loro- 2- hidro5-ksim u5-k onat sem ialdehid, sehingga t ida5-k t erj adi a5-k um ulasi 4-klorokat ekol dan prot oanem onin ( Blasco et al., 1997) .
Beberapa m ikroba lain yang m am pu m engkonversi klorofenol t elah dilaporkan. Meski t idak m eny ebut k an secara spesifik, Mohn and Kennedy ( 1992) m elihat adany a beberapa m ik roba anaerob y ang m am pu m endegradasi k lorofenol dan m ungkin dapat digunakan pada lim bah yang m engandung klorofenol. Biodegradasi anaerobik m erupakan suat u pilihan yang m urah unt uk m engeluarkan bahan pencem ar organik in sit u dari lingk ungan ( Kuo and Gent hner, 1996) . Set elah ak lim at isasi pada 3,4 µM pent aklorofenol selam a 6 bulan, konsorsium m et anogen m am pu m engeluarkan klorin dari posisi ort o, m et a, dan para dari pent ak lorofenol dan produk reduk t if dek lorinasiny a. Pent ak lorofenol didegradasi m enj adi 2,3,4,5- t et rak lorofenol, 2,3,4,6-t e2,3,4,6-t rak lorofenol, dan 2,3,5,6- 2,3,4,6-t e2,3,4,6-t rak lorofenol. Proses dek lorinasi 2,3,4,5-t e2,3,4,5-t raklorofenol m enghasilkan 3,4,5- 2,3,4,5-t riklorofenol un2,3,4,5-t uk kem udian didegradasi m enj adi 3,4- diklorofenol dan 3,5- diklorofenol. Deklorinasi m elalui ort o dan m et a dari 2,3,4,6- t et rak lorofenol m enghasilkan 2,4,6- t riklorofenol dan 2,4,5-t riklorofenol, sedang 2,3,5,6- 2,4,5-t e2,4,5-t raklorofenol m enghasilkan 2,3,5- 2,4,5-t riklorofenol dilanj ut kan dengan pem bent ukan 3,5- diklorofenol. Degradasi 2,4,6- t riklorofenol m enghasilkan 2,4- diklorofenol, sedang deklorinasi 2,4,5- t riklorofenol pada dua posisi m enghasilkan 2,4- diklorofenol dan 3,4- diklorofenol. Dari t iga diklorofenol y ang dihasilkan hany a 2,4- dik lorofenol y ang dapat didegradasi dalam w ak t u relat if singkat unt uk m em bent uk 4- klorofenol ( Nicholson et al., 1992) . I nokulasi t anah dengan Sphingom onas chlorophenolica RA2 sebany ak 108 sel/ g m am pu m em perpendek secara m engesankan w akt u m ineralisasi 30 µg pent ak lorofenol dengan sek it ar 80% diubah m enj adi CO2. I nokulasi dengan Mycobact erium
chlorophenolicum PCP1 m eningk at k an m ineralisasi sedik it di at as bak t eri indigenous. Kem am puan yang buruk dari st rain ini m ungkin berhubungan dengan
sifat sensit ifnya t erhadap pent aklorofenol, j uga m ungkin karena kondisi t anah y ang sedik it asam ( Meit hling and Karlson, 1996) .
Penam bahan bahan t ert ent u yang m engandung inokulan ke dalam t anah t erkont am inasi k lorofenol dapat m em percepat proses degradasi k lorofenol. Set elah adapt asi dengan pent aklorofenol, kom pos j eram i m am pu m em ineralisasi 56% pent aklorofenol. Sedang t anah t erem ediasi ( rem ediat ed soil) yang t elah diperkay a m am pu m em ineralisasi 24% pent ak lorofenol ( Laino and Jorgensen, 1996) . Biodegradasi anaerobik seny aw a k lorofenol dan k lorobenzoat j uga t ergant ung k epada elekt ron y ang t ersedia dan posisi k lorin t ersubst it usi ( Haggblöm et al., 1993) .
I I I . APLI KASI D AN BI OTEKN OLOGI
Fenol dan hom ologny a sepert i k lorofenol m em erluk an suasana aerob dan anaerob agar dapat t erdegradasi. Reduk t if dehalogenasi dilak uk an dalam suasana anaerob, nam un t ahap pem bent uk an k at ek ol at au k lorokat ekol pada reaksi yang m enggunakan ring- dioxygenase dan ring- cleav age dioxy genase m em erluk an oksigen.
Reak t or degradasi, liquid solid cont act react or ( LSC) , m ungkin m erupakan
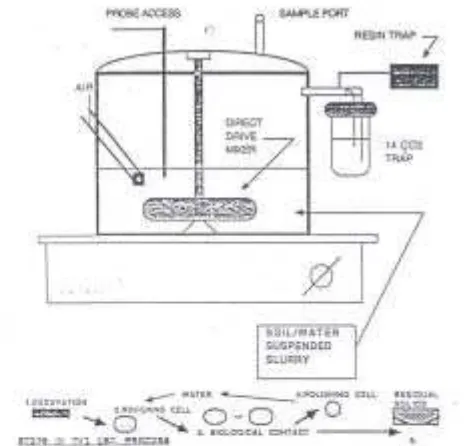
salah sat u alt ernat if reak t or biorem ediasi seny aw a fenol dan hom olog at au deriv at arom at is ( Gam bar 3) . Pada reak t or t erdapat suat u alat pengaduk dan aerat or dalam ruang t ert ut up yang m enerim a cont oh t anah dan air t anah secara t erus m enerus at au sem i- cont inuous ( Port ier, 1991) . Reak t or sej enis m am pu unt uk m em biorem ediasi t anah dan sedim en dengan bahan organik berbahaya y ang m elebihi 1% t ot al bahan organik ( Port ier, 1991) .
Gam bar 3. Sk em a unit reak t or liquid/ solid cont act ( LSC) ( dari Port ier, 1991) . Biorem ediasi dengan LSC didek at i dengan 2 langk ah proses perlak uan, y ait u t ahap pencam puran sel y ang di dalam ny a t anah t erkont am inasi dihom ogenasi dan diinokulasi dengan nut rien dan biom assa m ikroba, dan t ahap perlak uan biologi y ang di dalam ny a t erj adi proses m ineralisasi. Langk ah k et iga y ang m ungk in diperluk an y ait u air lim bah y ang dibuang dari reak t or j uga diberik an perlak uan biologi. Tanah residu y ang dik eluark an dapat dibuang di t anah pert anian at au dit em pat k an secara perm anen di suat u t em pat pem buangan ( Port ier, 1991) .
[image:7.612.204.438.243.466.2]4c) ( Blasco et al., 1997) . Selek si dan adapt asi j uga diperluk an dalam upay a lebih m eningk at k an laj u biodegradasi.
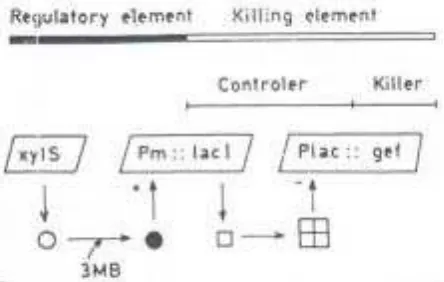
[image:8.612.116.527.157.636.2]Meskipun keunt ungan m em bersihkan lingkungan t ercem ar dengan m ikr oba r ekayasa t er lihat j elas, nam un t ingkah laku dalam w akt u lam a dar i m ikroba hasil rekayasa ini di lingkungan t erbuka belum diket ahui. Salah sat u upay a agar m ik roba ini dapat bert ahan hidup adalah dengan cara m em at ik anny a segera set elah aplikasi selesai, dengan cara m em beri m ereka kandungan biologi akt if ( act iv e biological cont ainm ent ) . Sist em act iv e biological cont ainm ent didasarkan pada pengendalian ekspresi fungsi let al. Sist em ini didasarkan pada sirkuit regulat or ekpresi j alur pem belahan m et a dari plasm id pWW0
Pseudom onas put ida, gen x ylS dengan prom ot or Pm dan gen gef dari Escherichia coli, y ang m eny andi prot ein m em bran y ang dapat m erusak pot ensial m em bran
sel dengan m em buat lubang pada m em bran. Model y ang dit unj uk k an pada Gam bar 5. m em predik si bahw a dengan adany a efekt or Xy lS, ekpresi prot ein Gef diham bat sehingga st rain dapat hidup dan m em bersihk an bahan pencem ar. Ket ik a seny aw a t arget habis, t idak adany a induk si Pm m engak ibat k an hilangny a LacI dalam sel inang, sehingga m enyebabkan t erekspresinya prot ein let al ( Molina et al. 1998) .
Gam bar 5. Model sist em act ive biological cont ainm ent ( dari Molina et al., 1998) .
I V . GEN ETI KA BAKTERI PEN D EGRAD ASI
Penget ahuan t ent ang genet ika m ikroba m enj adi sangat pent ing dalam penerapan biot eknologi unt uk m endegradasi senyaw a fenol dan hom olognya. Analisis biokim ia dan genet ik a degradasi aerob dilak uk an um um ny a pada
Pseudom onas ( Alt enschm idt et al. 1993; Dunaw ay - Mario and Babbit t , 1994;
Pow low ski and Shingler, 1994; William s and Sayers, 1994; Shield et al., 1995; de Sauza et al., 1995; Blasco et al., 1997; Fuenm ay or et al., 1998) . Degradasi seny aw a hidrokarbon arom at ik disandikan dalam plasm id at au k rom osom ( Haray am a et al., 1991; Jeffrey et al., 1992; Brenner et al., 1993) . Beberapa elem en loncat sert i Tn4651 dan Tn4653, t ransposon t oluena, dan Tn4655, t ransposon naft alena j uga m em baw a gen degradat if ( Wy ndham et al., 1994) . Shield et al. ( 1995) m elihat bahw a plasm id TOM, plasm id degradat if beruk uran 108 k b, bert anggung j aw ab t erhadap k at abolism e t oluena dan fenol. Plasm id ini m em iliki gen penyandi t oluene ort ho m onooxygenase dan cat echol
2,3-dioxy genase. Plasm id beruk uran besar y ang secara k olek t if disebut plasm id TOL
[image:9.612.210.432.292.433.2]Beberapa t elaah hom ologi gen- gen degradat if t elah dilak uk an. Kim et al. ( 1996) m elak uk an t elaah hom ologi gen degradat if pada Sphingom onas. Haray am a et al. ( 1991) m engam at i bahw a xylXYZ dari Pseum onas put ida and
benABC dari Acinet obact er calcoaset icus m em iliki induk yang sam a. Bundy et al.
( 1998) m elihat k esam aan ant ara ant ABC yang m enyandikan ant hranilat e
dioxy genase and benABC yang m enyandikan benzoat e dioxy genase dari Acinet obact er sp. st rain ADP1. Subst it usi ant C dari m ut an Acinet obact er dengan
benC ket ika dit um buhkan dalam ant ranilat m enunj ukkan bahw a BenC m em puny ai spesifit as subt rat y ang luas. Sebalikny a, benAB t idak dapat m ensubst it usi ant AB ( Bundy et al., 1998) m engindik asik an sem pit ny a subt rat unt uk BenAB ( Haray am a et al., 1991; Bundy et al., 1998) . Gen y ang bert anggung j aw ab m engkonversi naft alena m enj adi gent isat , nag, dari
Pseudom onas sp. st rain U2 yang diisolasi dari t anah t erkont am inasi m inyak t elah
disekuen. Perbandingan sekuen m enunj ukkan bahw a gen novel yang diw akili arket ipe unt uk st rain naft alena m enggunakan j alur gent isat ket im bang j alur m et a ( Fuenm ay or et al., 1998) .
Telaah perbandingan pada enzim y ang bert anggung j aw ab dalam degradasi seny aw a arom at ik dilak uk an oleh Dong et al. ( 1992) dan Neidle et al. ( 1991) . Cat echol 2,3- dioxy genase B. st earot herm ophilus m em punyai fungsi yang sam a dengan enzim yang disandikan x ylE dari P. put ida, m eskipun kest abilan panas dan hom ologiny a agak berbeda ( Dong et al., 1992) . Neidle et al. ( 1991) m enunj ukkan bahw a perbandingan sekuen deduksi asam am ino BenABC dari A.
calcoacet icus dengan sek uen sek erabat t erm asuk m ult ikom ponen t oluat e, t oluene, benzene, dan napht alene 1,2 dioxygenase m engindikasikan adanya
ukuran yang sam a dari sub- unit kom ponen hidroksilase yang dit urunkan dari induk yang sam a.
Klon gen- gen degradat if t elah dilaporkan. Kim and Oriel ( 1995) berhasil m engk lon pheA dan pheB dari B. st earot herm ophilus BR219 k e dalam E. coli. Gen ini m enyandikan konversi fenol m enj adi kat ekol dan kat ekol m enj adi 2-hidroksim uk onat sem ialdehida. Klon dan pem et aan gen pendegradasi fenol m elalui j alur m et a dari B. st earot herm ophilus FDTP- 3 ke dalam E. coli j uga t elah dilak uk an oleh Dong et al. ( 1992) . Springael et al. ( 1994) m elaporkan t ransfer gen degradat if k e dalam st rain resist en logam berat dari Alcaligenes eut rophus. Goy al and Zy lst ra ( 1996) m engk lon gen degradat if y ang berbeda dari gen k lasik
nah dari Com am onas t est ost eroni GZ39, m am pu m endegradasi hidrokarbon
arom at ik polisiklik. Klon dan sek uen sebagian dari gen degradasi at razin dari
Pseudom onas sp. st rain ADP t elah dilak uk an ( de Sauza et al., 1995) . Merek a
D a ft a r Pu st a k a
Alt enschm idt , U., B. Osw ald, E. St einer, H. Herrm ann, and G. Fuchs. 1993. New aerobic benzoat e oxidat ion pat hw ay v ia benzoyl- coenzy m e A and 3-hydr oxybenzoyl- coenzym e A in a denit r ifying Pseudom onas sp. J. Bact eriol. 175: 4851- 4858.
Assinder, S.J. and P.A. William s. 1990. The TOL plasm ids: det erm inant s of t he cat abolism of t oluene and t he xylenes. Adv. Microb. Physiol. 31: 1- 62.
Blasco, R. and F. Cast illo. 1992. Light - dependent degradat ion of nit rophenols by t he phot ot rophic bact erium Rhodobact er capsulat us E1F1. Appl. Environ. Microbiol. 58( 2) : 690- 695.
Blasco, R., M. Mallav arapu, R. Wit t ich, K.N. Tim m is, and D.H. Pieper. 1997. Evidence t hat form at ion of prot oanem onin from m et abolit es of 4-chloroby pheny l- com et abolizing m icroorganism s. Appl. Env iron. Microbiol. 63( 2) : 427- 434.
Brenner, V., B. S. Hernandez, and D. D. Focht . 1993. Variat ion in chlorobenzoat e cat abolism by Pseudom onas put ida P111 as a consequence of genet ic alt ernat ions. Appl. Environ. Microbiol. 59: 2790- 2794.
Bundy , B. M., A. L. Cam pbell, and E. L. Neidle. 1998. Sim ilarit ies bet w een t he
ant ABC- encoded ant hranilat e diox y genase and t he benABC- encoded
benzoat e diox y genase of Acinet obact er sp. st rain ADP1. J. Bact eriol. 180: 4466- 4474.
de Sauza, M.L., L.P. Wacket t , K.L. Boundy- Mills, T. Mandelbaum , and M.J. Sadow sk y . 1995. Cloning, charact erizat ion, and expression of a gene region from Pseudom onas st rain ADP involved in t he dechlorinat ion of at razine. Appl. Environ. Microbiol. 61: 3373- 3378.
Dunaway- Mariano, D. & P.C. Babbit t . 1994. On t he origins and funct ions of t he enzym es of t he 4- chlorobenzoat e t o 4- hydroxybenzoat e convert ing pat hway. Biodegrad. 5: 259- 276.
Dong, F., L. Wang, C. Wang, J. Cheng, Z. He, Z. Sheng, and R. Shen. 1992. Molecular cloning and m apping of phenol degradat ion genes from Bacillus
st earot herm ophilus FDTP- 3 and t heir expression in Escherichia coli. Appl.
Env iron. Microbiol. 58( 8) : 2531- 2535.
Fuenm ayor, S. L., M. Wild, A. L. Boyes, and P. A. William s. 1998. A gene clust er encoding st eps in conversion of napht halene t o gent isat e in Pseudom onas sp. st rain U2. J. Bact eriol. 1998. 180: 2522- 2530.
Gam ble, G.R., D.E. Ak in, H.P.S. Mak k ar, and K. Becker. 1996. Biological degradat ion of t annins in sericea lespedeza ( Lespedeza cuneat a) by t he w hit e rot fungi Ceriporiopsis subverm ispora and Cyat hus st ercoreus analy sed by sold- st at e 13C nuclear m agnet ic resonance spect roscopy. Appl. Env iron. Microbiol. 62( 10: 3600- 3604.
Haggblöm , M.M., M.D. Riv era, and L.Y. Young. 1993. I nfluence of alt ernat ive elect ron accept ors on t he anaerobic biodegradabilit y of chlorinat ed phenols and benzoat e acids. Appl. Env iron. Microbiol. 59( 4) : 1162- 1167.
Haray am a, S., M. Rek ik , A. Bairoch, E.L. Neidle, and L.N. Ornst on. 1991. Pot ent ial DNA slippage st ruct ures acquired during evolut ionary div ergence of Acinet obact er calcoacet icus chrom osom al benABC and Pseudom onas
put ida TOL pWW0 plasm id xylXYZ, genes encoding benzoat e
diox y genases. J. Bact eriol. 173: 7540- 7548.
Heipieper, H., R. Diefenbach, and H. Kew eloh. 1992. Conv ersion of cis unsat urat ed fat t y acids t o t rans, a possible m echanism for t he prot ect ion of phenol- degrading Pseudom onas put ida P8 from subst rat e t ox icit y . Appl. Env iron. Microbiol. 58( 6) : 427- 434.
Jeffrey, W. H., S. M. Cuskey, P. J. Chapm an, S. Resnick , and R. H. Olsen. 1992. Charact erizat ion of Pseudom onas put ida m ut ant s unable t o cat abolize benzoat e: Cloning and charact erizat ion of Pseudom onas genes involved in benzoat e cat abolism and isolat ion of a chrom osom al DNA fragm ent able t o subst it ut e for x ylS in act iv at ion of t he TOL low er- pat hw ay prom ot or. J. Bact eriol. 174: 4986- 4996.
Kim , I .C. and P.J. Oriel. 1995. Charact erizat ion of t he Bacillus st earot herm ophilus BR219 phenol hydroxylase gene. Appl. Environ. Microbiol. 61: 1252- 1256. Kim , E., P.J. Aversano, M.F. Rom ine, R.D. Schneider, and G.J. Zylst ra. 1996.
Hom ology bet ween genes for arom at ic hydrocarbon degradat ion in surface and deep- subsurface Sphingom onas st rains. Appl. Environ. Microbiol. 62: 1467- 1470.
Kuo, C. and B.R.S. Gent hner. 1996. Effect of added heav y m et al ions on biot ransform at ion and biodegradat ion of 2- chlorophenol and 3-chlorobenzoat e in anaerobic bact erial consort ia. Appl. Env iron. Microbiol. 62( 7) : 2317- 2323.
Laine, M.M. and K.S. Jorgensen. 1996. St raw com post and biorem ediat ed soil as inocula for t he biorem ediat ion of chlorophenol- cont am inat ed soil. Appl. Env iron. Microbiol. 62( 5) : 1507- 1513.
Leahy , J.G. and R.R. Colw ell. 1990. Microbial degradat ion of hy drocarbons in t he env ironm ent . Microbiol. Rev . 54( 3) : 305- 315.
Lobos, J.H., T.K. Leib, and T. Su. 1992. Biodegradat ion of bisphenol A and ot her bisphenols by Gram - Negat ive aerobic bact erium . Appl. Env iron. Microbiol. 58( 6) : 1823- 1831.
Miet hling, R. and U. Karlson. 1996. Accelerat ed m ineralizat ion of pent achlorophenol in soil upon inoculat ion w it h Mycobact erium
chlorophenolicum PCP1 and Sphingom onas chlorophenolica RA2. Appl.
Env iron. Microbiol. 62( 12) : 4361- 4366.
Mohn, W.W. and K.J. Kennedy . 1992. Lim it ed degradat ion of chlorophenols by anaerobic sludge granules. Appl. Env iron. Microbiol. 58( 7) : 2131- 2136. Molina, L., C. Ram os, M- C. Ronchel, S. Molin, and J.L. Ram os. 1998. Const ruct ion
of an efficient biologically cont ained Pseudom onas put ida st rain and it s suv iv al in out door assay s. Appl. Env iron. Microbiol. 64( 6) : 2072- 2078. Neidle, E.L., C. Hart net t , L.N. Ornst on, A. Bairoch, M. Rekik, and S. Harayam a.
Nicholson, F.D.K., S.L. Woods, J.D. I st ok , and D.C. Peeks. 1992. Reduct ive dechlorinat ion of chlorophenols by a pent achlorophenol- acclim at ed m et hanogenic consort ium . Appl. Env iron. Microbiol. 58( 7) : 2280- 2286. Port ier, R.J. 1991. Applicat ions of adapt ed m icro- organism s for sit e rem ediat ion
of cont am inat ed soil and ground w at er. I n Biological degradat ion of w ast es. Ed. A.M. Mart in. Elsev ier Applied Science. London. pp. 247- 259. Powlowski, J. and V. Shingler. 1994. Genet ics and biochem ist ry of phenol
degradat ion by Pseudom onas sp. CF600. Biodegrad. 5: 219- 236.
Schm idt , S.R. Wit t ich, D. Erdm ann, H. Wilkes, W. Francke, and P. Fort nagel. 1992. Biodegradat ion of diphenil et her and m onohalogenat ed deriv at iv es by Sphingom onas sp. st rain SS3. Appl. Env iron. Microbiol. 58( 9) 2744-2750.
Sem ple, K.T. and R.B. Cain. 1996. Biodegradat ion of phenols by t he alga
Ochrom onas danica. Appl. Env iron. Microbiol. 62( 4) : 1265- 1273.
Shen, H. and Y. Wang. 1995. Sim ult aneous chrom ium reduct ion and phenol degradat ion in a cocult ure of Escherichia coli ATCC 33456 and
Pseudom onas put ida DMP- 1. Appl. Env iron. Microbiol. 61( 7) : 2754- 2758
Shield, M.S., M.J. Reagin, R.R. Gerger, R. Cam pbell, and C. Som erville. 1995. TOM, a new arom at ic degradat ive plasm id from Burkholderia ( Pseudom onas) cepacea G4. Appl. Env iron. Microbiol. 61( 4) : 1352- 1356. Wy ndham , R.C., A.E. Cashore, C.H. Nak at su, and M.C. Peel. 1994. Cat abolic
t ransposons. Biodegrad. 5: 323- 342.
Werwat h, J., H. A. Arfm ann, D. H. Pieper, K. N. Tim m is, and R. Wit t ich. 1998. Biochem ical and genet ic charact erizat ion of a gent isat e 1,2- dioxygenase from Sphingom onas sp. st rain RW5. J. Bact eriol. 180: 4171- 4176.