PADA SISTEM INDOOR DAN OUTDOOR
YOGA TRISWANTO
SKRIPSI
DEPARTEMEN ILMU DAN TEKNOLOGI KELAUTAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
Dengan ini Saya menyatakan bahwa Skripsi yang berjudul:
KULTIVASI DIATOM PENGHASIL BIOFUEL JENIS
Skeletonema costatum, Thalassiosira sp., DAN Chaetoceros gracilis
PADA SISTEM INDOOR DAN OUTDOOR
adalah benar merupakan hasil karya sendiri dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Semua sumber data dan informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan oleh penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir Skripsi ini.
Bogor, Januari 2011
costatum, Thalassiosira sp., dan Chaetoceros gracilis pada Sistem Indoor dan
Outdoor. Dibimbing oleh MUJIZAT KAWAROE dan ADRIANI SUNUDDIN. Penelitian dengan topik kultivasi diatom penghasil biofuel pada sistem indoor
dan outdoor ini dilakukan pada bulan Mei–Juli 2010 di Laboratorium PT. Suri Tani Pemuka (Japfa), Unit Hatchery Udang Vannamei, Jalan Raya Gilimanuk km 35, Desa Pemutaran, Kecamatan Gerokgak, Kabupaten Singaraja, Propinsi Bali. Penghitungan kelimpahan diatom menggunakan haemocytometer dan mikroskop. Parameter yang diukur meliputi kelimpahan diatom, suhu salinitas, derajat keasaman (pH), dan intensitas cahaya. Analisis yang digunakan meliputi penghitungan
kelimpahan, laju pertumbuhan spesifik dan maksimum (µ dan µmaks), serta uji validitas Pearson.
Kultivasi pada sistem indoor memperlihatkan bahwa Chaetoceros gracilis
memiliki µmaks (0,520) dan kelimpahan maksimum tertinggi sebesar 7,3×106 sel/mL diikuti dengan Skeletonema costatum yang memiliki µmaks 0,515 dengan kelimpahan maksimum sebesar 1,97×106 sel/mL. Thalassiosira sp. memiliki µmaks 0,482 dengan kelimpahan maksimum terendah yaitu 0,9×106 sel/mL.
Kultivasi pada sistem outdoor memperlihatkan bahwa Skeletonema costatum
memiliki µmaks (0,506) yang lebih tinggi dibandingkan dua spesies lainnya dengan kelimpahan maksimum sebesar 1,78×106 sel/mL yang lebih rendah dibandingkan
Chaetoceros gracilis sebesar 2,79×106 sel/mL. Chaetoceros gracilis memiliki µmaks (0,268) yang lebih rendah dibandingkan Skeletonema costatum dan kelimpahan maksimum tertinggi dibandingkan dua spesies diatom lainnya. Thalassiosira sp. memilki µmaks (0,205) terendah dengan kelimpahan maksimum (0,47×106 sel/mL) terendah dibandingkan dua spesies diatom lainnya.
Skeletonema costatum, Thalassiosira sp., dan Chaetoceros gracilis pada sistem
indoor memiliki µmaks lebih besar dibandingkan pada sistem outdoor. Kelimpahan tertinggi pada kultivasi ketiga spesies tersebut juga dijumpai pada perlakuan sistem
indoor. Diatom yang dianggap paling efektif dikultur dalam sistem indoor adalah
Chaetoceros gracilis, sedangkan yang efektif dikultur dalam sistem outdoor adalah
© Hak Cipta milik IPB, tahun 2011
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau
menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan yang
wajar IPB
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis
PADA SISTEM INDOOR DAN OUTDOOR
YOGA TRISWANTO
SKRIPSI
sebagai salah satu syarat untuk memperoleh gelar Sarjana Ilmu Kelautan pada
Departemen Ilmu dan Teknologi Kelautan
DEPARTEMEN ILMU DAN TEKNOLOGI KELAUTAN FAKULTAS PERIKANAN DAN ILMU KELAUTAN
Judul Skripsi: KULTIVASI DIATOM PENGHASIL BIOFUEL JENIS Skeletonema costatum, Thalassiosira sp., DAN
Chaetoceros gracilis PADA SISTEM INDOOR DAN
OUTDOOR
Nama Mahasiswa: Yoga Triswanto Nomor Pokok: C54061023
Departemen: Ilmu dan Teknologi Kelautan
Menyetujui,
Dosen Pembimbing
Dr. Ir. Mujizat Kawaroe, M.Si Adriani Sunuddin, S. Pi., M. Si Utama Anggota
Mengetahui,
Tanggal Lulus: 3 Januari 2011
vi
rahmat dan karunianya, penulis dapat menyelesaikan skripsi ini. Skripsi yang
berjudul ”KULTIVASI DIATOM PENGHASIL BIOFUEL JENIS
Skeletonema costatum, Thalassiosira sp., DAN Chaetoceros gracilis PADA SISTEM INDOOR DAN OUTDOOR” diajukan sebagai salah satu syarat untuk mendapatkan gelar sarjana.
Penulis mengucapkan terima kasih sebesar-besarnya kepada kedua orang tua dan keluarga. Tidak lupa ucapan terima kasih diucapkan kepada Ibu Mujizat Kawaroe selaku dosen pembimbing utama, Ibu Adriani Sunuddin selaku
pembimbing anggota, Bapak Agus Somamihardja selaku pembimbing lapang dan juga yang telah memberikan izin tempat untuk melakukan kegiatan penelitian, Pak Irza, Pak Petyanto, Ilmi, Adi, Aris, Yafril, Dwi, Agung, Ugi, Vicky, Romdon, Heru, Mas Hadi, Mbak Ika, Mbak Ni luh, keluarga besar ITK khususnya angkatan 43, staf karyawan PT. Suri Tani Pemuka (Japfa) Unit Hatchery Bali-Banyuwangi, serta semua pihak yang telah membantu dalam penyusunan skripsi penelitian ini.
Penulis menyadari skripsi ini jauh dari kesempurnaan, oleh karena itu saran dan kritik sangat diharapkan demi kesempurnaan skripsi ini. Akhir kata penulis berharap agar skripsi ini dapat berguna bagi diri sendiri maupun orang lain dan dapat dikembangkan untuk penelitian selanjutnya.
Bogor, Januari 2011
vii
4.1 Kelimpahan dan Laju Pertumbuhan Spesifik Diatom pada Sistem Indoor ... 32
4.1.1 Kelimpahan dan Laju Pertumbuhan Spesifik Skeletonema costatum ... 33
4.1.2 Kelimpahan dan Laju Pertumbuhan Spesifik Thalassiosira sp. ... 34
4.1.3 Kelimpahan dan Laju Pertumbuhan Spesifik Chaetoceros gracilis ... 36
4.2 Kelimpahan dan Laju Pertumbuhan Spesifik Diatom pada Sistem Outdoor ... 38
viii
Chaetoceros gracilis. ... 42
4.3 Perbandingan Kelimpahan dan Laju Pertumbuhan Spesifik pada Kedua Sistem ... 44
4.3.1 Skeletonema costatum ... 44
4.3.2 Thalassiosira sp. ... 46
4.3.3 Chaetoceros gracilis ... 49
4.4 Kualitas Air ... 52
4.4.1 Kualitas Air pada Sistem Indoor ... 52
4.4.2 Kualitas Air pada Sistem Outdoor ... 57
5. KESIMPULAN DAN SARAN ... 63
5.1 Kesimpulan ... 63
5.2 Saran ... 64
DAFTAR PUSTAKA ... 65
ix
1. Alat dan bahan yang digunakan ... 22
2. Kelimpahan (×106 sel/ml) dan laju pertumbuhan spesifik (µ) diatom pada sistem indoor ... 32
3. Kelimpahan (×106 sel/ml) dan laju pertumbuhan spesifik (µ) diatom pada sistem outdoor ... 38
4. Kelimpahan dan laju pertumbuhan spesifik Skeletonema costatum pada sistem indoor ... 72
5. Kelimpahan dan laju pertumbuhan spesifik Thalassiosira sp. pada sistem indoor ... 73
6. Kelimpahan dan laju pertumbuhan spesifik Chaetoceros gracilis pada sistem indoor ... 74
7. Kualitas air Skeletonema costatum pada kultivasi sistem indoor .... 75
8. Kualitas air Thalassiosira sp. pada kultivasi sistem indoor ... 76
9. Kualitas air Chaetoceros gracilis pada kultivasi sistem indoor ... 77
10. Kelimpahan dan laju pertumbuhan spesifik Skeletonema costatum pada sistem outdoor ... 78
11. Kelimpahan dan laju pertumbuhan spesifik Thalassiosira sp. pada sistem outdoor ... 79
x
1. Bentuk sel Skeletonema costatum ... 8
2. Bentuk sel Thalassiosira sp. ... 10
3. Bentuk sel Chaetoceros gracilis ... 11
4. Grafik fase pertumbuhan mikroalga ... 17
5. Diagram alir prosedur penelitian kultivasi diatom ... 24
6. Diagram alir kultivasi diatom ... 28
7. Grafik kelimpahan diatom pada sistem indoor ... 33
8. Grafik kelimpahan diatom pada sistem outdoor ... 39
9. Kelimpahan Skeletonema costatum pada sistem indoor dan outdoor ... 45
10. Kelimpahan Thalassiosira sp. pada sistem indoor dan outdoor ... 47
11. Kelimpahan Chaetoceros gracilis pada sistem indoor dan outdoor .... 50
12. Suhu pada kultivasi sistem indoor ... 53
13. Salinitas pada kultivasi sistem indoor ... 54
14. Derajat keasaman (pH) pada kultivasi sistem indoor ... 55
15. Intensitas cahaya pada kultivasi sistem indoor ... 56
16. Suhu pada kultivasi sistem outdoor ... 57
17. Kisaran fluktuasi suhu dalam 1 hari ... 58
18. Salinitas pada kultivasi sistem outdoor ... 59
19. Derajat keasaman (pH) pada kultivasi sistem outdoor ... 61
xi
1. Penghitungan kelimpahan diatom ... 68
2. Penghitungan laju pertumbuhan spesifik dan laju pertumbuhan spesifik maksimum ... 68
3. Uji validitas Pearson dan uji lanjut regresi ... 69
4. Kelimpahan dan laju pertumbuhan spesifik diatom pada kultivasi sistem indoor ... 72
5. Kualitas air pada kultivasi sistem indoor ... 75
6. Kelimpahan dan laju pertumbuhan spesifik diatom pada kultivasi sistem outdoor ... 78
7. Kualitas air pada kultivasi sistem outdoor ... 81
8. Intensitas cahaya pada sistem kultivasi indoor dan outdoor ... 85
9. Komposisi kimiawi pupuk F2 dan pupuk teknis ... 86
10. Dokumentasi foto alat dan bahan penelitian ... 87
1
1.
PENDAHULUAN
1.1 Latar Belakang
Diatom merupakan kelompok mikroalga atau fitoplankton dari Divisi
Chrysophyta yang memiliki keunggulan dibandingkan kelompok lainnya karena memiliki distribusi yang luas, variatif dan dijumpai di hampir semua permukaan substrat (mampu merekam sejarah habitat), siklus hidup pendek, reproduksi cepat,
banyak spesies yang sensitif terhadap perubahan lingkungan, mampu
merefleksikan perubahan kualitas air dalam jangka pendek dan panjang, mudah
pencuplikan; pengelolaan dan identifikasinya. Diatom mempunyai kontribusi 40 - 45% produktivitas laut sehingga lebih produktif dibandingkan dengan hutan hujan di seluruh dunia. Oleh karena itu tidak mengherankan apabila diatom mempunyai
peranan yang sangat penting dalam siklus silika dan karbon di alam sehingga kesinambungan perikanan terjaga (Mann dan Lazier, 2006).
Pemanfaatan diatom sebagai bioindikator perairan telah banyak
diimplementasikan. Diatom juga berpotensi untuk menyerap gas buang CO2 yang
dihasilkan oleh proses pembakaran baik kendaraan, industri, respirasi, dan dekomposisi. Kultivasi diatom telah banyak dimanfaatkan sebagai pakan alami dalam bidang budidaya.
Beberapa spesies mikroalga kelompok diatom memiliki potensi untuk dijadikan salah satu solusi dalam mengatasi masalah krisis sumber daya minyak.
Hal ini dikarenakan adanya kandungan asam lemak pada diatom (Pratoomyot et al., 2005). Diatom dapat diekstraksi secara besar-besaran dalam menghasilkan
biofuel. Diatom dapat diproduksi dalam waktu yang sangat singkat karena siklus
Ada tiga jenis spesies diatom yang biasa yang digunakan sebagai pakan udang vaname yaitu Skeletonema costatum, Thalassiosira sp., dan Chaetoceros gracilis. Ketiga spesies diatom tersebut sudah dapat dilakukan kultur secara
besar-besaran namun hanya sebagai pakan saja. Kemudahan dalam kultur
memungkinkan untuk dilakukan penelitian terhadap pemanfaatan dalam penghasil
biofuel melalui analisis asam lemaknya.
Kultur mikroalga terdiri dari dua sistem yaitu sistem indoor dan sistem
outdoor. Kedua sistem kultivasi ini membutuhkan beberapa tahapan yang harus
diperhatikan untuk dapat memenuhi target produksi yang maksimal sehingga diperlukan pengetahuan dasar sifat biologi mikroalga seperti pertumbuhan. Oleh
karena itu sangat penting untuk membandingkan pertumbuhan tiga spesies diatom (Skeletonema costatum, Thalassiosira sp., dan Chaetoceros gracilis) pada kedua sistem tersebut untuk mengetahui jenis diatom dengan hasil kultivasi
maksimal yang dapat digunakan untuk ekstraksi menjadi biofuel.
1.2 Tujuan
Tujuan dari Penelitian ini adalah membandingkan laju pertumbuhan tiga
3
2. TINJAUAN PUSTAKA
2.1 Biologi, Morfologi, dan Habitat Diatom
Diatom merupakan mikroalga uniseluler yang distribusinya sangat universal
di semua tipe perairan. Diatom memiliki bentuk kehidupan yang kosmopolit dan merupakan penyusun utama mikroalga baik di ekosistem perairan tawar maupun laut dengan jumlah spesies terbesar dibandingkan komunitas mikroalga lainnya.
Diatom mempunyai keunikan dan sangat spesifik, karena arsitektur dan anatomi dinding selnya yang tersusun dari silika, menyebabkannya dapat tersimpan dalam
kurun waktu yang sangat lama di dalam sedimen (Soeprobowati dan Suwarno, 2009).
Diatom dibedakan dengan tumbuhan lain sebab dinding selnya terdiri dari
silikon (silikat) yang terdiri dari dua bagian utama masing-masing bagian disebut
valve, menyatu bagaikan kaleng pastiles. Masing-masing valve sering memiliki
hiasan halus dan bagus. Umumnya kromatofora berwarna coklat keemasan dan klorofil ditutupi oleh pigmen berwarna coklat (Basmi, 1999). Diatom sering
terlihat dalam bentuk buih kuning pada permukaan air dalam kolam dan selokan (Pandey dan Trivedi, 2005).
Diatom (Bacillariophyceae) merupakan pembagian kelas dari divisi
Chrysophyta (Basmi, 1999). Dinding sel diatom sangat unik dan tersusun secara simetris yang membuat kelompok ini dipisahkan dari alga lainnya (Pandey dan
Bentuk uniselular diklasifikasikan kedalam dua order berdasarkan bentuk selnya yaitu diatom Pennales (Pennales) dengan bentuk bilateral simetris dan diatom centris (Centrales) dengan bentuk radial simetris (lingkaran) apabila
dilihat dari atas (Basmi, 1999; Pandey dan Trivedi, 2005). Ordo Centrales hiasan
valve-nya berbentuk jari-jari lingkaran, sedangkan pada Pennales secara bilateral.
Pergerakan tidak pernah dijumpaipada jenis-jenis anggota Centrales, namun hanya pada pennales yang valvenya berbentuk memanjang (Basmi, 1999). Diatom Centrales biasanya berbentuk panjang dengan filamen berantai dan valve
berdempet, melalui berbagai mekanisme, sel saling berdekatan pada valve yang mengikuti pembelahan sel. Sel biasannya berisi banyak kloroplas kecil dan
sebuah vakuola yang besar. Diatom Centrales biasanya planktonik dan lebih sering ditemukan pada air laut dan payau dibandingkan air tawar (Darley, 1982). Frustule simetri pada diatom Pennales merupakan bentuk bilateral; setiap valve
berbentuk seperti kano dibandingkan sebuah lingkaran cakram. Diatom Pennales
biasanya berisi dua atau satu kloroplas yang besar, kloroplas berbentuk H dan
vakuola kecil pada ujung sel. Diatom Pennales melimpah di habitat air tawar dan air laut, serta pada beberapa jenis sering berasosiasi dengan substrat. Diatom
Pennales sering menempel pada tanaman, hewan, batuan atau butir pasir dengan
sebuah lapisan getah atau tangkai atau mungkin juga secara bebas bergerak pada permukaan air atau dalam sedimen yang berlumpur (Darley, 1982).
Edhy et al. ( 2003) menyebutkan diatom memiliki beberapa karakteristik yang di antaranya :
- Sel tunggal dengan dinding yang ditutupi silikat.
- Zat warna berupa klorofil-a dan c, β-karoten, fukoxantin, diatoxanthin dan diadinixanthin.
- Thalus disebut frustula yang terdiri dari valve (atas) dan girdle (bawah). - Reproduksi aseksual dengan pembelahan dan seksual dengan oogami dan
isogami.
Reproduksi diatom secara aseksual biasanya berlangsung dengan cara
sederhana, yaitu dengan melakukan pembelahan sel dimana setengah protoplasma (yaitu protoplasma di dalam epiteka) dan protoplasma setengah lainnya (yang berada di hipoteka) menjadi frustul diatom baru (individu baru). Teka yang
dibangun dalam proses pembentukan frustul baru selalu hipoteka, dan kelak individu baru tersebut membelah lagi seperti cara di atas, sehingga makin lama
terbentuklah frustul-frustul baru yang lebih kecil, sampai batas tertentu sehingga frustul terkecil tadi tidak mampu (secara alami) atau tidak sanggup membelah lagi. Dengan demikian, selama proses pembelahan sel tersebut selalu terbentuk
frustul (individu) baru yang sama besar dengan induk dan seterusnya.
Fase pembelahan terakhir (frustul terkecil), mereka tidak lagi melakukan
pembelahan seperti diatas, tetapi protoplasmanya membesar membentuk spora yang disebut auxospora, yang mendesak cangkang menjadi terbuka sehingga auxospora meninggalkan cangkang. Demikian pula halnya dengan frustul terkecil
terdahulu, lalu terbentuklah epiteka dan hipoteka, dan akhirnya terbentuk frustul baru (individu baru) yang bentuk, besar, dan sifat (karakternya) sama dengan nenek moyangnya dahulu dan pada pembelahan sel pertama bentuknya sama
dengan nenek moyangnya dahulu. Namun adapula bentuk frustul yang lebih besar dari moyangnya dahulu, dan pada pembelahan sel pertama bentuknya sama
dengan nenek moyang berikut hiasan-hiasan valve-nya. Demikian pula dengan pembelahan sel (frustul) menjadi dua bagian akibat protoplasmanya tumbuh membesar sehingga mendesak epiteka dan hipoteka samapai kedua teka tadi
terlepas (terpisah) satu sama lain, yang masing-masing membawa setengah protoplasma induk.
Ordo Centrales pembelahan sel sama seperti yang diuraikan di atas, yang akhirnya terbentuk auxospora, dan dua auxospora menyatu sehingga terbentuk individu baru. Pembesaran kembali sel diatom melalui formasi penyatuan dua
auxospora terjadi pada sejumlah spesies-spesies diatom. Meskipun demikian, nampaknya hal tersebut tidak berlaku secara universal. Dalam proses pembelahan
sel tersebut terbentuk sel yang sama besar dengan sel induk, dan ada pula sel-sel baru yang semakin kecil, namun auxospora ini jarang dijumpai sewaktu
mengambil sampel diatom diperairan alami (Basmi, 1999).
Dalam beberapa hal, reproduksi berlangsung diantara diatom, terutama untuk spesies-spesies yang hidup di laut di zona dekat pantai (neritik), juga pada
spesies-spesies yang hidup di air tawar, mereka dapat membentuk spora diam. Spora-spora diam ini biasanya lebih kecil dari sel-sel diatom aslinya, dan dibungkus oleh dinding sel tebal (berat), serta mempunyai karakteristik baik
membebaskan diri dari frustulnya, namun ada pula yang tetap berada di dalam frustul. Dalam kasus demikian, ada spora yang melanjutkan hidupnya sebagai diatom planktonis, namun ada juga yang jatuh ke dasar perairan menjadi diatom
benthik, di dasar perairan berlumpur. Spora diam ini dapat hidup bertahan dan tumbuh dalam kondisi buruk (musim dingin) dan pada kondisi lingkungan normal
kembali maka ia akan hidup normal seperti biasanya (Basmi, 1999).
Reproduksi seksual berlangsung baik melalui sel-sel yang sama seperti auxospora, atau melalui perkawinan antara dua mikrospora yang berflagella
(biasannya secara isogamet), yang kemudian kedua mikrospora tersebut mengalami fusi (peleburan) sehingga membentuk satu axospora, seperti yang
banyak terjadi pada anggota Pennales. Kedua auxospora yang mengalami fusi tersebut dibungkus oleh lendir dan kemudian membentuk dua gamet. Hasil fusi dua axospora lainnya juga membentuk dua gamet. Masing-masing gamet dari
bentukan hasil fusi auxospora yang berbeda tadi kemudian bertemu (kawin) yang akan melahirkan individu baru (Basmi, 1999).
Contoh dari spesies kelompok diatom diantarannya adalah Chaetoceros
gracilis, Skeletonema costatum, dan Thalassiosira sp. (Edhy et al., 2003). Ketiga spesies ini merupakan kelompok diatom yang memiliki pola bentuk sentris
2.1.1 Skeletonema costatum
Klasifikasi Skeletonema costatum menurut Bougis (1979) dalam Tjahjo et al. (2002) dan Edhy et al. (2003) adalah sebagai berikut:
Divisi: Chrysophyta
Classis: Bacillariophyceae
Ordo: Centrales
Family: Skeletonemoidae Genus: Skeletonema
Spesies: Skeletonema costatum
. Sumber: SBRC
Gambar 1. Bentuk sel Skeletonema costatum
Skeletonema costatum merupakan mikroalga bersel tunggal, dengan ukuran sel berkisar antara 4-15 µm. Alga ini dapat membentuk untaian rantai yang terdiri
dari epiteka pada bagian atas dan hipoteka pada bagian bawah, serta pada setiap sel dipenuhi oleh sitoplasma (Isnansetyo dan Kurniastuty, 1995). Bentuk sel
Skeletonema costatum dapat dilihat pada Gambar 1. Kamat (1976) dalam Tjahjo
frustula yang dapat menghasilkan skeletal external yang berbentuk silindris (cembung) dan mempunyai duri-duri yang berfungsi sebagai penghubung antar frustula yang satu dengan frustula yang lainnya sehingga membentuk filamen.
Skeletonema costatum merupakan diatom yang bersifat euritermal yaitu mampu tumbuh pada kisaran suhu 3–30 oC dan temperatur optimal adalah 25-27 oC
(Susetyowati, 1994 dalam Tjahjo et al., 2002). Daerah penyebarannya meliputi daerah tropis dan subtropis, terdapatnya mulai dari pantai sampai lautan, sebagai meroplankton dan benthos. Perkembangbiakan diatom Skeletonema hanya dapat
terjadi secara aseksual (Mudjiman, 1999 dalam Nugraheny, 2001). Bentuk sel
Skeletonema costatum dapat dilihat pada Gambar 1.
2.1.2 Thalassiosira sp.
Klasifikasi Thalassiosira sp.munurut Edhy et al. (2003)adalah sebagai berikut:
Spesies: Thalassiosira sp.
Cleve (1873) dalam Somers (1972) menjelaskan genus Thalassiosira
memiliki karakteristik berupa benang mukosa sentral halus yang menghubungkan
sentral mukosa yang sering disebut dengan single apiculus (Hendley, 1959 dan Hasle, 1968 dalam Somers, 1972).
Deskripsi morfologi umum yang telah disepakati sebelumnya untuk
Thalassiosira sp. meliputi bentuk rantai dan inmucilage yang menempel pada koloni, benang-benang kitin menghubungkan sel dalam rantai, bentuk sel terlihat
mengelilingi persegi dengan sebuah cekungan dalam pusat valve, sebuah
rimoportula besar diantara muka valve dan mantel, sebuah lingkaran kecil yang diam dan dua atau tiga lingkaran kecil fultoportulae dan susunan areola (Becerril
et al., 2009).
Sumber: SBRC
Gambar 2. Bentuk sel Thalassiosira sp.
Diatom laut, Thalassiosira sp. yang dikultivasi pada medium N:P:Si=
11:1:6 memberikan biomassa sebesar 0,067 g/mL, dengan kandungan karbohidrat sebesar 7,7%, kandungan protein sebesar 0,93% dan kandungan lemak sebesar 9,69% (Purba, 2008). Bentuk sel Thalassiosira dapat dilihat pada
2.1.3 Chaetoceros gracilis
Klasifikasi Chaetoceros gracilis menurut Zipcodezoo (2009) dan Edhy et al. (2003) adalah sebagai berikut:
Divisi: Chrysophyta
Classis: Bacillariophyceae
Ordo: Centrales
Family: Chaetocerotaceae Genus : Chaetoceros
Spesies: Chaetoceros gracilis
Sumber : reed-mariculture.com (2010 )
Gambar 3. Bentuk sel Chaetoceros gracilis
Lavens dan Sorgeloos (1996) dalam Suantika et al. (2009) menyatakan
Chaetoceros gracilis merupakan diatom sentrik yang soliter, organisme
Spesies ini dapat hidup pada temperatur 10– 20 oC dan spesies ini dapat dikultur secara masal pada air laut yang diperkaya dengan pupuk anorganik atau pupuk kandang (Tjahjo et al., 2002). Chaetoceros merupakan salah satu genus diatom
penting dalam mikroalga laut, karena Chaetoceros merupakan genus terbesar dengan jumlah spesies sekitar 400. Secara ekologi genus ini juga berperan
sebagai produsen primer serta merupakan makanan penting bagi biota lain, terutama udang (Von-Quillfedlt, 2001 dalam Setiawati, 2009; Panggabean, 2007 dalam Setiawati, 2009). Bentuk Sel Chaetoceros Gracilis dapat dilihat pada
Gambar 3.
2.2 Persyaratan Kultivasi Mikroalga
Sylvester et al. (2002) menyatakan bahwa kelangsungan hidup dan
pertumbuhan suatu jenis mikroalga sangat erat kaitannya dengan ketersediaan nutrien (unsur hara) serta dipengaruhi oleh kondisi lingkungan. Dengan demikian faktor-faktor yang menentukan keberhasilan budidaya mikroalga seperti
pemilihan lokasi yang tepat dan penggunaan media yang sesuai dengan segala persyaratannya adalah langkah awal yang perlu dilakukan dalam melaksanakan
budidaya mikroalga.
2.2.1 Nutrien
Mikroalga membutuhkan berbagai macam unsur anorganik, baik sebagai
makro nutrien maupun mikro nutrien. Unsur makro nutrient terdiri dari N, P, K, C, Si, S, dan Ca serta unsur mikro nutrient terdiri dari Fe, Zn, Cu, Mg, Mo, Co, B,
Unsur N merupakan komponen utama dari protein sel yang merupakan bagian dasar kehidupan organisme. Nitrogen yang dibutuhkan untuk media kultur terdiri dari beberapa substansi berikut : KNO3; NaNO3; NH4Cl; (NH2)CO (urea)
dan lain-lain. Unsur P sangat dibutuhkan dalam proses protoplasma dan inti sel. Fosfor juga merupakan bahan dasar pembentuk asam nukleat, fosfolipid, enzim,
dan vitamin. Dengan demikian fosfor sangat berperan nyata dalam semua aktifitas kehidupan mikroalga. Fosfor yang dibutuhkan pada kultur mikroalga dapat diperoleh dari KH2PO4; NaH2PO4;Ca3PO4; Ca3PO4 (TSP) dan lain-lain.
Unsur K selain berperan dalam pembentukan protoplasma juga berperan penting dalam kegiatan metabolisme dan aktifitas lainnya. Karbon juga
diperlukan dalam pertumbuhan mikroalga. Sumber karbon yang dimanfaatkan mikroalga sebagian besar bersifat anorganik dalam bentuk CO2 dan bicarbonat. Silika merupakan nutrien yang banyak dimanfaatkan oleh mikroalga jenis
Bacillariophyceae (diatom) sebagai salah satu sumber elemen untuk membentuk komposisi frustula pada lapisan sel Bacillariophyceae dalam proses asimilasi.
Sulfur juga merupakan salah satu elemen penting yang dibutuhkan dalam pembentukan protein. Sulfur juga merupakan salah satu elemen penting yang dibutuhkan dalam pembentukan protein. Sulfur untuk media kultur alga dapat
diperoleh dari NH4SO4 (ZA); CuSO4 dan lain-lain. Unsur Ca berperan dalam penyelarasan dan pengaturan aktifitas protoplasma dan kandungan pH di dalam
sel. Sumber Ca antara lain adalah CaCl2 dan Ca(NO3)2 ( Kurniastuty dan
Unsur Ferrum berperan penting dalam pembentukan kloroplas dan sebagai komponen esensial dalam proses oksidasi. Unsur besi juga merupakan bahan dasar sitokrom dan heme atau nonheme protein, kofaktor untuk beberapa enzim.
Pada kultur alga komponen besi dapat diperoleh dari FeCl3, FeSO4 dan
FeCaH5O7. Unsur Zn Juga dibutuhkan oleh mikroalga yang berasal dari sumber
mineral ZnCl2 dan ZnSO4. Unsur Cuprum yang dibutuhkan oleh mikroalga biasanya bersumber dari mineral yaitu CuSO4.5H2O. Unsur Magnesium merupakan kation sel yang utama dan bahan dasar klorofil. Kation sel utama,
kofaktor anorganik untuk banyak reaksi enzimatik berfungsi di dalam penyatuan substrat dan enzim. Unsur molibdenum dibutuhkan oleh mikroalga dalam bentuk
Na2MoO4.H2O. Unsur kobalt dibutuhkan oleh mikroalga dapat diperoleh dari CoCl2. Unsur boron dibutuhkan mikroalga dalam bentuk H3BO3 (Sylvester et al., 2002; Edhy et al., 2003; PT. Suri Tani Pemuka, 2005; Cahyaningsih, 2009).
2.2.2 Faktor-Faktor Lingkungan
Pertumbuhan suatu jenis fitoplanton atau mikroalga sangat erat kaitannya
dengan ketersediaan hara makro dan mikro serta dipengaruhi oleh kondisi lingkungan di dalam media kulturnya. Faktor-faktor lingkungan yang berpengaruh terhadap pertumbuhan mikroalga antara lain cahaya, suhu, pH,
kandungan CO2 bebas dan tekanan osmosis (salinitas) (Sylvester et al., 2002). Mikroalga merupakan organisme autotrof yang mampu membentuk
senyawa organik dari senyawa-senyawa anorganik melalui proses fotosintesis. Dengan demikian cahaya mutlak diperlukan sebagai sumber energi (Sylvester et al., 2002). Laju fotosintesis akan tinggi bila intensitas cahaya tinggi dan menurun
Budidaya mikroalga di dalam laboratorium, cahaya matahari dapat digantikan dengan sinar lampu TL dengan intensitas cahaya 5.000-10.000 lux. Intensitas cahaya adalah jumlah cahaya yang mengenai satu satuan permukaan.
Satuannya adalah footcandle atau lux. Kisaran optimum intensitas cahaya bagi pertumbuhan mikroalga adalah 2.000-8.000 lux (Sylvester et al., 2002).
Suhu secara langsung mempengaruhi efisiensi fotosintesis dan merupakan faktor yang menentukan dalam pertumbuhan mikroalga. Umumnya pada kondisi laboratorium, perubahan suhu air dipengaruhi oleh temperatur ruangan dan
intensitas cahaya. Pada kultivasi mikroalga skala massal yang dilakukan di luar ruangan, suhu sangat dipengaruhi oleh keadaan cuaca. Kisaran optimum bagi
pertumbuhan mikroalga umumnya adalah 25–32 oC (Sylvester et al., 2002). Kebanyakan sel termasuk mikroalga sangat peka terhadap derajat keasaman cairan yang menjadi media hidupnya. Batas pH untuk pertumbuhan jasad
merupakan suatu gambaran dari batas pH bagi kegiatan enzim. Jika suatu enzim menunjukan kegiatannya pada pH tertentu, kenaikan dan penurunan pH dapat
menyebabkan kegiatan enzim itu berubah. pH optimum untuk kultivasi diatom adalah kisaran 7-8 (Sylvester et al., 2002; Cahyaningsih, 2009).
Senyawa CO2 adalah gas atmosfer yang terdiri dari satu atom karbon dan
dua atom oksigen. Karbon dioksida dihasilkan oleh semua hewan, tumbuh-tumbuhan, fungi, dan mikroorganisme pada proses respirasi dan digunakan oleh
tumbuhan dan mikroalga pada proses fotosintesis. CO2 di dalam media kultur merupakan faktor penting untuk mikroalga, karena secara langsung dipakai sebagai bahan untuk membentuk molekul-molekul organik seperti karbohidat
Sebagai salah satu organisme yang hidup di dalam air, salinitas merupakan salah satu faktor pembatas bagi pertumbuhan dan perkembangan mikroalga. Fluktuasi salinitas secara langsung menyebabkan perubahan tekanan osmosis di
dalam sel mikroalga. Salinitas yang terlampau tinggi atau rendah dapat menyebabkan tekanan osmosis di dalam sel menjadi lebih rendah atau lebih
tinggi, sehingga aktifitas sel terganggu. Hal ini mempengaruhi pH protoplasma sel dan menurunkan kegiatan enzim di dalam sel. Umumnya mikroalga air laut hidup normal pada salinitas optimum 25-35‰ (Sylvester et al., 2002). Salinititas
optimum untuk diatom adalah 28-32‰ (Cahyaningsih, 2009).
2.3 Fase Pertumbuhan Mikroalga
Edhy et al. (2003) menjelaskan terdapat 4 fase dalam pertumbuhan
mikroalga yaitu fase lag (istirahat), fase logaritmik (pertumbuhan eksponensial), fase stasioner (pertumbuhan stabil), dan fase deklinasi (kematian). Fase-fase
pertumbuhan mikroalga tersebut dapat dlihat pada Gambar 4.
Fase lag merupakan fase ketika populasi mikroalga tidak mengalami
perubahan, tetapi ukuran sel pada fase ini meningkat. Fotosintesis masih aktif berlangsung dan organisme mengalami metabolisme tetapi belum terjadi pembelahan sel sehingga kepadatannya belum meningkat. Dalam perairan
tambak kondisi air masih bening atau remang-remang dengan transparansi >80 cm.
Fase logaritmik diawali dengan pembelahan sel dengan laju pertumbuhan yang terus menerus, pertumbuhan pada fase ini mencapai maksimal. Dalam perairan tambak ditandai dengan air yang mulai berwarna sampai warna pekat
Sumber : Edhy et al. (2003)
Gambar 4. Grafik fase pertumbuhan mikroalga
Fase stasioner merupakan fase dengan pertumbuhan yang mulai mengalami
penurunan dibandingkan fase logaritmik. Pada fase ini, laju reproduksi atau pembelahan sel sama dengan laju kematian dalam arti penambahan dan
pengurangan plankton relatif sama sehingga kepadatan plankton cenderung tetap. Dalam perairan tambak fase ini memperlihatkan warna yang cenderung stabil dan sebaiknya dipertahankan supaya tidak terjadi droping plankton.
Fase deklinasi merupakan fase ketika terjadi penurunan jumlah atau kepadatan mikroalga. Pada fase ini laju kematian lebih cepat dibandingkan laju
reproduksi. Laju kematian mikroalga dipengaruhi oleh ketersediaan nutrien, cahaya, temperatur, dan umur mikroalga itu sendiri. Dalam perairan tambak kematian mikroalga ditandai dengan meningkatnya transparansi, adanya
2.4 Biofuel dari Mikroalga
Biofuel yang dapat terbarukan dibutuhkan untuk menggantikan minyak yang dijadikan bahan bakar yang berkontribusi pada pemanasan global dan
ketersediaannya yang terbatas. Biodiesel dan bioethanol merupakan bahan bakar yang berpotensi dapat diperbarui yang menarik perhatian besar. Biodiesel dan
bioethanol diproduksi oleh tanaman pertanian menggunakan metode yang ada yang keberadaannya tidak dapat menggantikan minyak fosil yang dijadikan bahan bakar, tapi ada sebuah alternatif. Oleh karena itu dibutuhkan penemuan baru
bahan baku yang cocok untuk produksi bahan bakar yang tidak mengurangi ketersediaan minyak nabati yang dapat dikonsumsi manusia sebagai makanan.
Salah satu alternatifnya yaitu mikroalga. Mikroalga lebih memiliki potensi untuk dijadikan biodiesel dibandingkan biofuel (Chisti, 2008). Minyak dari mikroalga mengandung lipid yang cocok untuk esterifikasi atau transesterifikasi (Umdu et
al., 2008). Di antara berbagai jenis alga, mikroalga merupakan kelompok biota yang menjanjikan hasil lebih baik karena:
1. Memiliki laju pertumbuhan yang tinggi (Ritt-man, 2008 dalam Umdu et al., 2008).
2. Kandungan lipid dapat disesuaikan dengan mengubah komposisi media untuk
tumbuh (Naik et al., 2006 dalam Umdu et al., 2008).
3. Dapat dipanen lebih dari sekali dalam satu tahun (Schenk et al., 2008 dalam
Umdu et al., 2008).
4. Dapat menggunakan air laut atau air limbah (Schenk et al., 2008 dalam Umdu
5. Karbondioksida di atmosfer, merupakan sumber untuk pertumbuhan mikroalga (Schenk et al., 2008 dalam Umdu et al., 2008)
6. Biodiesel dari lemak alga merupakan non toksik dan bersifat biodegradable
secara cepat. (Schenk et al., 2008 dalam Umdu et al., 2008).
7. Mikroalga yang digunakan untuk biodiesel mampu berproduksi 15-300 kali
lebih cepat dibandingkan tanaman daratan (Chisti, 2007).
2.5 Teknik Kultivasi Mikroalga
Teknik kultivasi mikroalga terbagi menjadi tiga tahap yaitu skala
laboratorium (indoor), skala semi-massal (semi-outdoor) dan skala massal (outdoor). Biasanya ketiga tahapan tersebut tidak semuanya dapat dilaksanakan.
Mengingat diperlukannya tenaga, tempat dan biaya yang cukup besar. Untuk pembenihan skala kecil atau rumah tangga biasanya hanya melakukan kultur skala semi massal dan skala massal, sedangkan bibitnya diperoleh dari unit-unit
pembenihan besar lainnya atau dari instansi pemerintah (Anjar et al., 2002). Kultur mikroalga skala laboratorium (indoor) memerlukan kondisi
lingkungan yang stabil, sehingga perlu dilengkapi dengan AC agar suhu ruangan selalu terkendali dan ruangan terisolasi dengan lingkungan luar, selain itu ada beberapa jenis mikroalga tumbuh lebih baik pada suhu yang relatif rendah.
Sumber aerasi (pengudaraan) digunakan Hi-blower tersendiri dan dilengkapi dengan saringan untuk memperkecil terjadinya kontaminasi. Pupuk yang
digunakan pada skala laboratorium terbuat dari bahan kimia PA (pro analis) dengan dosis pemakaian 1 ml pupuk untuk 1 liter volume kultur. Jenis dan formula pupuk adalah yang sudah distandarkan dan umum digunakan yaitu
pertumbuhan mikroalga optimal sehingga didapatkan bibit (starter) yang bermutu tinggi untuk skala kultur selanjutnya. Dalam kultur skala laboratorium ada beberapa kegiatan yang umum dilakukan antara lain: sterilisasi alat; bahan dan air
media; isolasi mikroalga; kultur di media agar; kultur di media cair; pembuatan pupuk; penghitungan mikroalga dan penyimpanan (Isnansetyo dan Kurniastuty,
1995; Anjar et al., 2002).
Kultur skala semi massal dimulai dari volume 30 liter hingga 100 liter dalam wadah aquarium yang diletakkan di luar laboratorium. Air laut dengan
salinitas tertentu dimasukkan ke dalam akuarium, kemudian dimasukkan ke dalam akuarium, selanjutnya dimasukkan inokulum sekitar 1/10 bagian dari total volume
budidaya. Inokulum berasal dari kultur skala laboratorium. Pupuk yang digunakan sama dengan pupuk yang digunakan pada kultur skala laboratorium dan diberikan sesuai takaran yang dibutuhan. Pencahayaan hanya mengandalkan
cahaya matahari pada siang hari. Pada keadaan tertentu di mana cahaya matahari kurang memadai, dapat menggunakan lampu TL atau lampu sorot. Aerasi dijaga
jangan sampai mati, karena hal ini akan menghambat pertumbuhan mikroalga dan dapat menyebabkan kematian (Isnansetyo dan Kurniastuty, 1995). Menurut Cahyaningsih (2009), kultur semi massal dilakukan di outdoor (di luar
laboratorium) dengan penyinaran cahaya matahari yang tak langsung.
Kultur massal atau outdoor dimulai dari 1 ton sampai 20 ton atau lebih. Air
laut dengan salinitas tertentu dimasukkan ke dalam bak-bak kultur. Selanjutnya dilakukan pemupukan dan diberi aerasi. Inokulum yang berasal dari kultur semi massal dimasukkan sebanyak 1/10 bagian sebagai bibit. Pupuk yang digunakan
ZA, NPK dan KNO3 sebagai sumber nitrogen dan TSP, SP3, NPK sebagai sumber fosfatnya. Vitamin dan mikronutrien lainnya bisa ditambahkan sebagai
pelengkap. Umumnya pada kultur mikroalga dari kelas diatom perlu
ditambahkan silikat sekitar 5-20 ppm. Kultur skala massal menggunakan penyinaran matahari secara langsung menggunakan penyaring fiber dengan
22
3. BAHAN DAN METODE
3.1 Waktu dan Lokasi Penelitian
Penelitian ini dilakukan pada bulan Mei–Juli 2010 di Laboratorium PT. Suri Tani Pemuka (Japfa), Unit Hatchery Udang Vannamei, Jalan Raya Gilimanuk km
35, Desa Pemutaran, Kecamatan Gerokgak, Kabupaten Singaraja, Propinsi Bali.
3.2 Alat dan Bahan
Alat dan bahan yang digunakan pada penelitian ini disajikan pada Tabel 1.
Tabel 1. Alat dan bahan yang digunakan pada penelitian
Alat dan Bahan Spesifikasi Unit/Satuan
Aerator Indoor Tekno Takatsuki 1/ -
Autoklaf - 1/ Celcius
Filter bertingkat Hayward Pro Series 2/ -
Filter ozon - 1/ -
Haemocytometer Assistant (Neubauer) 25x10-4 mm2 1/ sel/mL
Hotplate Labinco L-32 1/ Celcius
Kran Aerator - 24/ -
Lampu TL Philips Lifemax TLD 36 watt 9/ - Luxmeter Konica Minolta T-10 Illuminance 1/ Lux Mikroskop Olympus CH-BI45-2 1:200x 1/ -
Oven Memmert 1/ Celcius
pH meter Eutech Cyberscan pH 310 1/ -
Tabel 1. Alat dan bahan yang digunakan pada penelitian (Lanjutan)
Alat dan Bahan Spesifikasi Unit/Satuan
Pipet mohr Pyrex 1 mL 3/ mL
Refraktometer Hand Refraktometer Atago 1/ -
Selang aerasi - 45 meter/ -
Thermometer Air Raksa (Hg) 1/ Celcius
Timbangan analitik AND EK-3000i 1/ gram
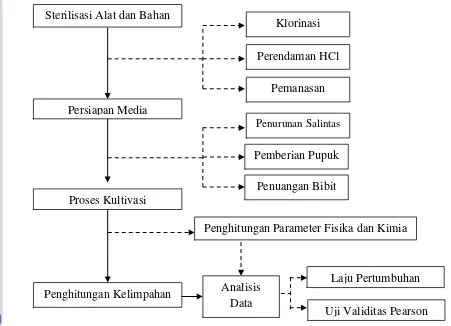
3.3 Prosedur Penelitian
Prosedur penelitian dilakukan secara bertahap untuk mencapai hasil yang baik dan benar. Prosedur penelitian yang dilakukan pada proses kultivasi secara
umum terdiri dari sterilisasi alat dan bahan, persiapan media, proses kultivasi, dan penghitungan kelimpahan. Prosedur penelitian yang dilakukan disajikan pada
Gambar 5.
Gambar 5. Diagram alir prosedur penelitian kultivasi diatom
3.3.1 Sterilisasi Alat dan Bahan
Sterilisasi merupakan suatu metode yang dilakukan untuk membersihkan
menggangu pertumbuhan mikroalga yang dikultivasi. Sterilisasi bertujuan untuk membersihkan alat serta bahan yang akan digunakan untuk isolasi maupun kultivasi mikroalga dari mikroorganisme ataupun bahan kimia yang dapat
mengganggu pertumbuhan mikroalga.
Sterilisasi dalam kultivasi mikroalga skala indoor terdiri atas sterilisasi
ruang, peralatan kultivasi, dan media cair. Sterilisasi ruang dilakukan dengan cara menyiapkan ruang dan rak yang akan digunakan sebagai laboratorium kultivasi. Ruangan dibersihkan dari debu dan kotoran lainnya dengan cara menyapu,
mengelap, dan mengepel ruangan, termasuk rak kultivasi hingga bersih dengan menggunakan larutan klorin 1%. Terakhir, ruangan yang sudah bersih dan kering
disemprot alkohol 70% menggunakan sprayer.
Peralatan kultivasi yang disterilisasi meliputi peralatan gelas, peralatan plastik, selang dan wadah kultivasi. Peralatan kultivasi mula-mula disterilisasi
dengan cara mencuci terlebih dahulu dengan tipol yang kemudian dibilas dengan air tawar. Kemudian dilakukan perendaman dengan HCl 0.2% selama 24 jam dan
dibilas kembali dengan air tawar. Sterilisasi alat-alat gelas dilanjutkan dengan menggunakan oven pada suhu 120 oC selama 1,5 jam. Sedangkan untuk sterilisasi
selang aerasi dan batu aerasi dilakukan pada autoklaf dengan suhu 121 oC selama 15 menit. Peralatan gelas yang telah dipanaskan kemudian dikeringkan dan dibungkus dengan alumunium foil. Sterilisasi peralatan yang tidak tahan panas
dan berukuran besar, misalnya galon dan toples plastik, dilakukan dengan merendamnya dalam larutan klorin 40 ppm.
µm, 5 µm, dan 2 µm. Sterilisasi media cair untuk fase test tube, Erlenmeyer, dan botol kaca menggunakan autoklaf dengan cara dipanaskan pada suhu 121 oC kurang lebih selama 1 jam untuk mensterilkan 10 L media cair. Kemudian Media
cair disimpan dalam wadah steril yang tertutup rapat. Sterilisasi media cair untuk fase toples dan galon juga dilakukan menggunakan filter bertingkat 50 µm, 10
µm, 5 µm, 2 µm namun hanya diklorinasi dengan konsentrasi 25 ppm dan diaerasi selama 24 jam. Kemudian media cair tersebut dinetralisir dengan natrium
tiosulfat 25 ppm untuk menghilangkan kandungan klorin pada media cair tersebut.
Pupuk yang digunakan pada kultivasi indoor disterilisasi dengan menggunakan autoklaf yang sama halnya dengan media cair yang digunakan pada fase test tube,
Erlenmeyer, dan botol kaca. Sterilisasi laboran dilakukan dengan menyemprotkan alkohol 70% pada kedua tangan untuk menghindari kontaminasi pada mikroalga ketika laboran berinteraksi dengan kultivan.
Sama halnya dengan sistem indoor, sterilisasi pada sistem outdoor terbagi menjadi sterilisasi ruang dan bak, sterilisasi peralatan kultivasi, dan sterilisasi
media cair. Sterilsasi mula-mula dilakukan dengan cara menyiapkan ruang dan bak yang akan digunakan. Ruangan dan bak dibersihkan dari debu dan kotoran
lainnya dengan cara mencuci, membilas, dan mengelap ruangan beserta bak yang akan digunakan untuk kultivasi hingga bersih menggunakan cairan klorin 1%. Ruang dan bak dikeringkan dengan cara dijemur langsung di bawah sinar
matahari selama satu hari.
Peralatan kultivasi yang disterilisasi meliputi peralatan gelas, peralatan
air tawar. Kemudian dilakukan perendaman dengan HCl 0,2% selama 24 jam dan dibilas kembali dengan air tawar. Sterilisasi alat-alat gelas dilanjutkan dengan menggunakan oven pada suhu 120 oC selama 1,5 jam. Sterilisasi selang dan batu
aerasi dilakukan dengan perendaman menggunakan larutan HCl 10% selama 24 jam. Peralatan gelas yang telah dipanaskan kemudian dikeringkan dan dibungkus
dengan alumunium foil. Sterilisasi peralatan yang tidak tahan panas dan berukuran besar misalnya boks stirofom dilanjutkan dengan melakukan perendaman menggunakan larutan klorin dengan konsentrasi 40 ppm.
Sterilisasi media cair dilakukan dengan menggunakan ozonisasi yang terlebih dahulu dilakukan filtrasi bertingkat 50 µm, 10 µm, 5 µm, dan 2 µm dan
diklorinasi dengan konsentrasi 50 ppm yang diaerasi selama 24 jam. Kemudian media cair tersebut dinetralisir dengan natrium tiosulfat 50 ppm untuk
menghilangkan kandungan klorin pada media cair tersebut. Pupuk yang
digunakan pada kultivasi outdoor juga disterilisasi dengan menggunakan autoklaf yang sama halnya seperti pupuk yang digunakan pada sistem indoor.
3.3.2 Persiapan Media
Media air untuk fase botol gelas 1 L dan Erlenmeyer 250 mL pada sistem
indoor dibuat dengan cara menurunkan salinitas air laut menjadi 28‰ dengan
menambahkan air tawar atau akuades terlebih dahulu. Kemudian dilanjutkan dengan sterilisasi menggunakan autoklaf pada suhu 121 oC selama 15 menit.
Setelah dingin, air dapat digunakan sebagai media kultivasi mikroalga. Media air untuk fase toples dan galon pada sistem indoor dipersiapkan
dan galon dan disterilisasi menggunakan klorin 25 ppm. Sebelum digunakan sebagai media kultivasi, air laut diaerasi selama 24 jam dan dinetralkan klorinnya dengan cara menambahkan natrium tiosulfat 25 ppm.
Media yang digunakan dalam sistem outdoor adalah air laut dengan salinitas 28‰. Air laut yang digunakan terlebih dahulu dilakukan penyaringan secara
bertingkat dan penembakan sinar UV. Kemudian, media disterilisasi dengan klorin sebanyak 50 ppm dan selanjutnya diaerasi, keesokan harinya dinetralkan
dengan natrium tiosulfat 50 ppm.
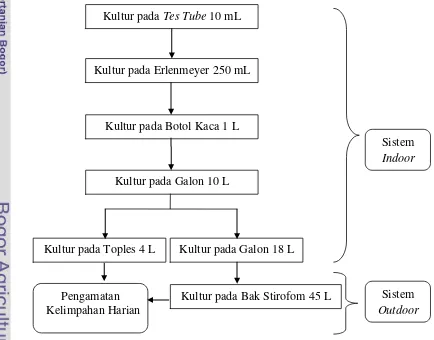
3.3.3 Proses Kultivasi
Kultivasi mikroalga pada penelitian ini disajikan pada Gambar 6.
Gambar 6. Diagram alir kultivasi diatom Kultur pada Tes Tube 10 mL
Kultur pada Erlenmeyer 250 mL
Kultur pada Botol Kaca 1 L
Kultur pada Galon 10 L
Kultur pada Toples 4 L Kultur pada Galon 18 L
Sistem
Indoor
Kultur pada Bak Stirofom 45 L Pengamatan
Kelimpahan Harian
Sistem
Proses kultivasi sistem indoor memiliki beberapa tahapan. Kultivasi pada Erlenmeyer dilakukan dengan cara menyiapkan Erlenmeyer ukuran 250 mL. Strain yang berasal dari test tube dipindahkan pada Erlenmeyer yang berukuran
250 mL sebanyak 10% dari volume media dan diisi air laut hingga mencapai 200 mL dan ditutup dengan alumunium foil. Setelah 7 hari strain dipindahkan ke
botol kaca yang berukuran 1 L dengan persentase 10% strain mikroalga dari botol kaca. Setelah 3 hari mikroalga siap dipindahkan ke galon dengan media sebanyak 10 L dan juga dengan persentase 10% dari media air pada galon. Setelah 3 hari,
strain pada media galon dipindahkan ke toples dengan volume media 4 L. Strain yang dituangkan sebanyak 10% dari media yang digunakan pada toples yaitu 400
mL. Volume inilah dilakukan pengamatan selama 15 hari dengan 3 kali pengulangan.
Proses kultivasi pada sistem outdoor merupakan tahapan lanjutan dari
sistem indoor. Kultivasi sistem outdoor diawali dengan penuangan strain sebanyak 10% dari ukuran bak yang digunakan pada kultivasi. Media air yang
akan digunakan pada sistem outdoor memiliki volume 45 L sehingga strain yang dituang adalah sebanyak 4,5 L. Strain mikroalga yang dituang berasal dari
kultivasi pada sistem indoor. Sistem outdoor juga dilakukan pengamatan selama 15 hari dan juga dilakukan sebanyak 3 kali pengulangan.
3.3.4 Penghitungan Kelimpahan Mikroalga (Jumlah Sel)
Monitoring kelimpahan dilakukan sebanyak 1 kali dalam 24 jam dan dimulai pada hari ke-0 (t0) hingga hari ke-15 (t15). Monitoring kelimpahan
dilakukan pada setiap sampel spesies dan pada kedua sistem (indoor dan outdoor).
pertumbuhan mikroalga dengan menghitung nilai kelimpahannya. Peralatan yang digunakan adalah mikroskop dan Haemocytometer. Monitoring kelimpahan dilakukan dengan melihat jumlah mikroalga pada kotak kecil Haemocytometer
yang dilakukan pada 5 titik pandang dan 3 kali ulangan. Adapun formulasi yang digunakan ada pada persamaan (1).
... (1)
Dimana N adalah jumlah sel mikroalga yang teramati
3.3.5 Pengukuran Parameter Fisika dan Kimia Media
Pengukuran parameter fisika dan kimia dilakukan pada toples yang berisi media air laut sebanyak 4 L dan pada bak stirofom yang berkapasitas 45 L.
Parameter fisika yang diukur adalah suhu, salinitas, dan cahaya, sedangkan parameter kimia yang diukur adalah pH. Pengukuran suhu, salinitas dan pH
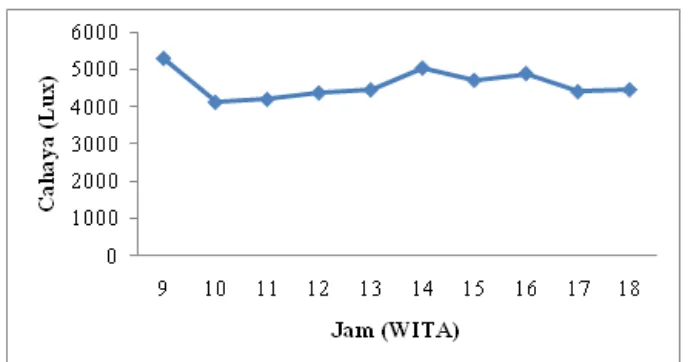
dilakukan dilakukan sebanyak 1 kali dalam 24 jam dan dimulai pada hari ke-0 (t0) hingga hari ke-15 (t15). Pengukuran cahaya yang dilakukan pada sistem indoor hanya 1 kali sedangkan pada sistem outdoor sebanyak 9 kali dalam waktu 24 jam,
dengan rentang waktu 1 jam pada pukul 09.00 – 18.00 WITA.
3.4 Analisis Data
Analisis data dilakukan dengan cara membandingkan kepadatan pada ketiga spesies diatom (Chaetoceros gracilis, Skeletonema costatum, dan Thalassiosira
sp.) dan juga pada kedua sistem (sistem indoor dan sistem outdoor. Perbandingan
menggunakan uji validitas Pearson untuk melihat korelasi yang terjadi dan uji lanjut regresi untuk melihat pengaruh parameter kualitas air terhadap kelimpahan dengan selang kepercayaan 0,05.
Laju pertumbuhan spesifik (µ) mikroalga dihitung dengan rumus menurut Krichnavaruk et al (2004), pada persamaan (2).
...(2)
Dimana Nt adalah kepadatan populasi pada waktu ke-t, N0 adalalah
kepadatan populasi sel pada waktu 0, T0 adalah waktu awal dan Tt adalah waktu pengamatan. Laju pertumbuhan spesifik maksimum (µmaks) dihitung dari kelimpahan pada saat awal kultivasi hingga puncak kelimpahan maksimum,
32
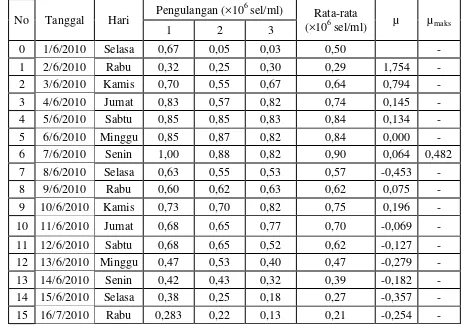
4.1 Kelimpahan dan Laju Pertumbuhan Spesifik Diatom Pada Sistem Indoor
Hasil penelitian menunjukan bahwa ketiga spesies diatom yang dikultur selama
lima belas hari pada sistem indoor memiliki pola pertumbuhan yang berbeda. Jumlah kelimpahan awal spesies diatom yang dikultivasi juga memiliki jumlah yang berbeda. Skeletonema costatum memiliki kelimpahan awal sebesar 0,15×106 sel/mL,
Thalassiosira sp. memiliki kelimpahan awal sebesar 0,05×106 sel/mL, dan
Chaetoceros gracilis memiliki jumlah kelimpahan awal sebesar 0,54×106 sel/mL.
Grafik kelimpahan dan laju pertumbuhan spesifik diatom pada sistem indoor dapat dilihat pada Tabel 2 dan Gambar 7.
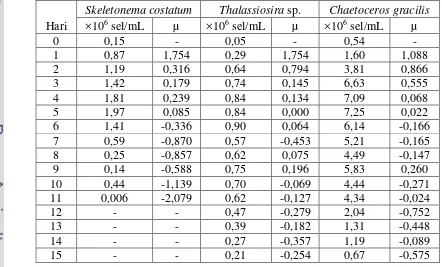
Tabel 2. Kelimpahan (×106 sel/ml) dan laju pertumbuhan spesifik (µ) diatom pada sistem indoor
Hari
Skeletonema costatum Thalassiosira sp. Chaetoceros gracilis
Gambar 7. Grafik kelimpahan diatom pada sistem indoor
4.1.1 Kelimpahan dan Laju Pertumbuhan Spesifik Skeletonema costatum
Pola pertumbuhan Skeletonema costatum hanya memiliki 1 buah puncak populasi pada hari ke-5 dengan nilai kelimpahan sebesar 1,97×106 sel/mL.
Skeletonema costatum memiliki daya adaptasi yang tinggi terhadap lingkungannya. Hal ini dapat dilihat dari nilai µ pada hari ke-1 sebesar 1,754. Laju pertumbuhan
spesifik maksimum (µmaks) yang dihitung dari awal kultur hingga puncak maksimum pada Skeletonema costatum adalah 0,515.
Fase lagSkeletonema costatum diduga terjadi kurang dari 24 jam. Keadaan
ini dapat dilihat dari kelimpahan dan nilai µ pada hari ke-1 yang semula 0,15×106 sel/mL meningkat secara drastis menjadi 0,87×106 sel/mL dalam waktu 24 jam yang
berarti Skeletonema costatum memiliki daya adaptasi yang tinggi terhadap
lingkungan yang baru memiliki kondisi yang sama pada lingkungan sebelumnya. Fase eksponensial diduga terjadi dalam waktu kurang dari 24 jam hingga hari ke-4.
Fase stasioner yaitu laju pertumbuhan yang relatif tetap diduga terjadi pada hari ke-4 hingga hari ke-5. Keadaan ini ditunjukan dengan adanya nilai µ yang semakin berkurang mendekati nilai 0 sebesar 0,085 pada saat memasuki hari ke-5. Fase
deklinasi diduga terjadi pada hari ke-5 hingga hari ke-11. Keadaan ini ditunjukan dengan nilai µ yang bernilai negatif sebesar -0,336 yang berarti terjadi penurunan
jumlah kelimpahan pada saat memasuki hari ke-6. Kelimpahan akhir yang didapat pada hari ke-11 adalah sebesar 0,006×106 sel/mL. Pengamatan kelimpahan pada hari ke-12 menunjukan populasi Skeletonema costatum yang berjumlah 0.
Turunnya laju pertumbuhan Skeletonema costatum dapat disebabkan oleh tiga hal, yaitu berkurangnya mikronutrien sebagai faktor pembatas karena telah
banyak dimanfaatkan selama fase eksponensial, adanya toksik yang dihasilkan oleh mikroalga itu sendiri sebagai hasil dari metabolisme yang meracuni mikroalga itu sendiri dan berkurangnya proses fotosintesis akibat bertambahnya jumlah sel
sehingga hanya bagian permukaan kultur saja yang memperoleh cahaya (Riley dan Chester, 1971 dalam Nugraheny, 2001).
4.1.2 Kelimpahan dan Laju Pertumbuhan Spesifik Thalassiosira sp.
Thalassiosira sp. memiliki pola pertumbuhan dengan dua buah puncak populasi yaitu hari ke-6 dengan nilai kelimpahan sebesar 0,90×106 sel/mL dan hari
gracilis. Keadaan ini dapat dilihat dari nilai µ pada hari ke-1 sebesar 1,754 dengan kelimpahan awal Thalassiosira sp sebesar 0,05×106 sel/mL yang meningkat menjadi
0,29×106 sel/mL. Laju pertumbuhan spesifik maksimum Thalassiosira sp. adalah sebesar 0,482.
Fase lagThalassiosira sp. diduga terjadi dalam waktu kurang dari 24 jam.
Keadaan ini dapat dilihat dari bentuk grafik dan nilai µ yang menunjukan bahwa
Thalassiosira sp. memiliki daya adaptasi yang tinggi sehingga waktu yang
dibutuhkan untuk beradapatasi terhadap lingkungan terlihat singkat yaitu kurang dari 24 jam. Fase eksponensial pada Thalassiosira sp. diduga terjadi pada waktu kurang dari 24 jam hingga hari ke-4 yang ditunjukan oleh adanya peningkatan drastis secara
eksponensial. Fase stasioner diduga terjadi pada hari ke-4 hingga hari ke-6. Keadaan ini dikarenakan laju pertumbuhan yang semakin menurun pada hari ke-4
hingga hari ke-6. Hal ini dapat dilihat dari nilai µ yang diperoleh pada hari ke-4 sebesar 0,134 yang menunjukan terjadinya penurunan laju pertumbuhan. Fase deklinasi dapat diduga terjadi pada hari ke-6 hingga hari ke-15. Namun pada hari
ke-8 terdapat kenaikan laju pertumbuhan kembali hingga hari ke-9. Hal ini diduga karena Thalassiosira sp. memasuki periode kriptik di mana sel-selyang masih hidup
memanfaatkan tambahan nutrisi dari sel-sel mikroalga yang lisis untuk
pertumbuhannya sehingga dapat meningkatkan populasinya kembali (Suantika, 2009).
kelimpahan ini juga dapat diduga karena berkurangnya mikronutrien sebagai faktor pembatas karena telah banyak dimanfaatkan selama fase eksponensial, adanya toksik
yang dihasilkan oleh mikroalga itu sendiri sebagai hasil dari metabolisme yang meracuni mikroalga itu sendiri dan berkurangnya proses fotosintesis akibat bertambahnya jumlah sel sehingga hanya bagian permukaan kultur saja yang
memperoleh cahaya (Riley dan Chester, 1971 dalam Nugraheny, 2001).
4.1.3 Kelimpahan dan Laju Pertumbuhan Spesifik Chaetoceros gracilis
Chaetoceros gracilis memiliki pola pertumbuhan dengan dua buah puncak
populasi yang lebih besar nilai kelimpahannya dibandingkan dua spesies lainnya yaitu hari ke-5 dengan kelimpahan sebesar 7,3×106 sel/mL dan hari ke-9 sebesar 5,83×106 sel/mL. Chaetoceros gracilis juga memiliki daya adaptasi yang tinggi
terhadap lingkungannya. Keadaan ini dapat dilihat dari nilai µ pada hari ke-1 sebesar 1,088. Hal ini dikarenakan lingkungan pada saat kultur memiliki kondisi
yang sama pada lingkungan sebelumnya. Laju pertumbuhan spesifik maksimum
Chaetoceros gracilis adalah sebesar 0,520.
Fase lagChaetoceros gracilis dapat diduga terjadi dalam waktu kurang dari 24
jam. Hal ini dapat dilihat dari nilai kelimpahan dan bentuk grafik yang terjadi pada hari ke-1 yang menunjukan pola pertumbuhan yang langsung memasuki fase
eksponensial. Fase eksponensial terjadi pada waktu kurang dari 24 jam hingga hari ke-3. Fase stasioner dapat diduga terjadi pada hari ke-3 hingga hari ke-5. Hal ini
hari ke-15. Keadaan ini dapat dilihat dari jumlah kelimpahan yang menurun pada hari ke-6. Kondisi ini juga dapat dilihat dari nilai µ (µ = -0,166) yang bernilai
negatif pada saat memasuki hari ke-6.
Hari ke-9 terjadi peningkatan kelimpahan kembali. Keadaan ini dapat diduga karena sel-sel Chaetoceros gracilis memasuki periode kriptik di mana sel-sel
Chaetoceros gracilis yang masih hidup memanfaatkan tambahan nutrisi dari sel-sel
Chaetoceros gracilis yang lisis untuk pertumbuhannya sehingga dapat meningkatkan
populasinya kembali (Suantika, 2009). Fase deklinasi berlanjut kembali hingga hari ke-15 dengan jumlah kelimpahan akhir sebesar 0,67×106 sel/mL. Fase deklinasi dapat terjadi karena nutrisi kultur telah habis dan terjadi akumulasi senyawa NH4+
dalam konsentrasi tinggi dan adanya produk ekstraseluler dari mikroalga yang meracuni diri sendiri sehingga dapat meningkatkan mortalitas Chaetoceros gracilis
(Fogg, 1965 dalam Panggabean, 2000 dan Suantika, 2009).
Hasil yang dapat disimpulkan dari µmaks dan kelimpahan maksimum ketiga spesies diatom pada sistem indoor adalah Chaetoceros gracilis memiliki laju
pertumbuhan yang tertinggi dengan µmaks sebesar 0,520 dan kelimpahan maksimum sebesar 7,25×106 sel/mL diikuti Skeletonema costatum dengan µmaks sebesar 0,515
dan kelimpahan maksimum sebesar 1,97×106 sel/mL. Thalassiosira sp. memiliki µmaks terendah sebesar 0,482, dan kelimpahan maksimum terendah sebesar 0,90×106 sel/mL. Chaetoceros gracilis memiliki µmaks dan kelimpahan tertinggi karena
4.2 Kelimpahan dan Laju Pertumbuhan Spesifik Diatom Pada Sistem Outdoor
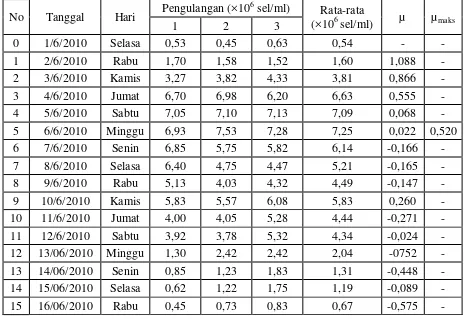
Kultur pada sistem outdoor yang dilakukan selama 15 hari memperlihatkan
pola pertumbuhan yang berbeda pada ketiga spesies diatom. Jumlah kelimpahan awal spesies diatom yang dikultivasi juga memiliki jumlah yang berbeda.
Skeletonema costatum memiliki kelimpahan awal sebesar 0,13×106 sel/mL,
Thalassiosira sp. memiliki kelimpahan awal sebesar 0,11×106 sel/mL, dan
Chaetoceros gracilis memiliki kelimpahan awal sebesar 0,43×106 sel/mL. Tabel 3
dan Gambar 8 menunjukan kelimpahan dan laju pertumbuhan spesifik Diatom yang dikultur pada sistem outdoor.
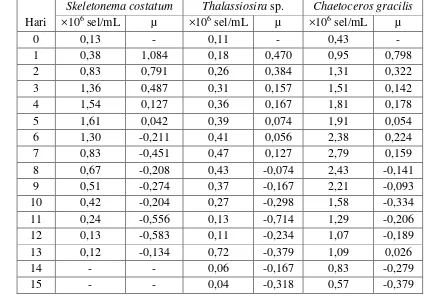
Tabel 3. Kelimpahan (×106 sel/ml) dan laju pertumbuhan spesifik (µ) diatom pada sistem outdoor
Hari
Skeletonema costatum Thalassiosira sp. Chaetoceros gracilis
Gambar 8. Grafik kelimpahan diatom pada sistem outdoor
4.2.1 Kelimpahan dan Laju Pertumbuhan Spesifik Skeletonema costatum
Pola pertumbuhan Skeletonema costatum pada sistem outdoor hanya memiliki satu buah puncak populasi pada hari ke-5 dengan nilai kelimpahan sebesar 1,61×106 sel/mL. Skeletonema costatum memiliki daya adaptasi yang tinggi terhadap
lingkungannya. Hal ini dapat dilihat dari nilai µ pada hari ke-1 yaitu sebesar 1,084. µmaks pada Skeletonema costatum adalah sebesar 0,506.
Fase lag pada Skeletonema costatum diduga terjadi kurang dari 24 jam. Keadaan ini dapat dilihat dari kelimpahan dan nilai µ pada hari ke-1 yang semula 0,13×106 sel/mL meningkat secara drastis menjadi 0,33×106 sel/mL dalam waktu 24
jam yang berarti bahwa Skeletonema costatum memiliki daya adaptasi yang tinggi terhadap lingkungan barunya sehingga mampu tumbuh dengan laju yang cepat pada
µ yang semakin berkurang mendekati nilai 0 sebesar 0,042 pada saat memasuki hari ke-5. Fase deklinasi diduga terjadi pada hari ke-5 hingga hari ke-13. Kelimpahan
akhir yang didapat pada hari ke-13 adalah sebesar 0,12×106 sel/mL. Keadaan ini ditunjukan dengan nilai µ yang bernilai negatif yaitu sebesar -0,211 yang artinya terjadi penurunan jumlah kelimpahan pada saat memasuki hari ke-6.
Pengamatan kelimpahan pada hari ke-14 menunjukan populasi Skeletonema costatum yang berjumlah 0. Turunnya laju pertumbuhan Skeletonema costatum
dapat disebabkan oleh tiga hal, yaitu berkurangnya mikronutrien sebagai faktor pembatas karena telah banyak dimanfaatkan selama fase eksponensial dan adanya toksik yang dihasilkan oleh mikroalga itu sendiri sebagai hasil dari metabolisme
yang meracuni mikroalga itu sendiri (Riley dan Chester, 1971 dalam Nugraheny, 2001).
4.2.2 Kelimpahan dan Laju Pertumbuhan Spesifik Thalassiosira sp.
Pola pertumbuhan Thalassiosira sp. pada sistem outdoor hanya memiliki 1 buah puncak kelimpahan pada hari ke-7 dengan nilai kelimpahan sebesar 0,47×106
sel/mL. Thalassiosira sp. memiliki daya adaptasi yang lebih rendah terhadap lingkungan dibandingkan 2 spesies lainnya. Keadaan ini dapat dilihat dari nilai µ
pada hari ke-1 sebesar 0,470 dengan kelimpahan awal (hari ke-0) sebesar 0,11×106 sel/mL meningkat menjadi 0,18×106 sel/mL. Hal ini mengindikasikan bahwa
Thalassiosira sp. memiliki daya adaptasi lingkungan yang rendah karena adanya
pertumbuhan spesifik maksimum Thalassiosira sp. adalah sebesar 0,205. Nilai µmaks pada spesies Thalassiosira sp. lebih rendah dibandingkan µmaks dua spesies lainnya.
Fase lagThalassiosira sp. diduga terjadi hingga hari ke-1. Keadaan ini dapat dilihat dari jumlah kelimpahan awal (hari ke-0) yang meningkat pada hari ke-1 namun jumlah kelimpahan pada hari ke-1 tidak mencapai dua kali lipat kelimpahan
awalnya. Fase eksponensial pada Thalassiosira sp. diduga terjadi pada hari ke-1 hingga hari ke-5 yang menunjukan kelimpahan meningkat drastis secara
eksponensial. Fase stasioner diduga terjadi pada hari ke-6 hingga hari ke-8. Keadaan ini dikarenakan laju pertumbuhan yang semakin menurun pada hari ke-6 hingga hari ke-8. Hal ini dapat dilihat dari nilai µ yang diperoleh pada hari ke-6
yaitu sebesar 0,056 yang menunjukan terjadinya penurunan laju pertumbuhan. Fase deklinasi dapat diduga terjadi pada hari ke-8 hingga hari ke-15. Hari ke-15,
Thalassiosira sp. memiliki kelimpahan akhir sebanyak 0,04×106 sel/mL. Fase
deklinasi ini ditentukan berdasarkan nilai µ yang bernilai negatif pada saat memasuki hari ke-8 yaitu sebesar -0,074.
Penurunan nilai kelimpahan dapat diduga karena berkurangnya mikronutrien sebagai faktor pembatas karena telah banyak dimanfaatkan selama fase eksponensial,
adanya toksik yang dihasilkan oleh mikroalga itu sendiri sebagai hasil dari metabolisme yang meracuni mikroalga itu sendiri dan berkurangnya proses fotosintesis akibat bertambahnya jumlah sel sehingga hanya bagian permukaan
4.2.3 Kelimpahan dan Laju Pertumbuhan Spesifik Chaetoceros gracilis
Chaetoceros gracilis pada sistem outdoor memiliki pola pertumbuhan dengan
dua buah puncak kelimpahan maksimum pada hari ke-7 dengan kelimpahan sebesar 2,79×106 sel/mL dan pada hari ke-13 sebesar 1,09×106 sel/mL. Chaetoceros gracilis
juga memiliki daya adaptasi yang tinggi terhadap lingkungannya. Hal ini dapat dilihat dari nilai µ pada hari ke-1 sebesar 0,798. Keadaan ini mengindikasikan
bahwa Chaetoceros gracilis memiliki kemampuan beradaptasi yang tinggi terhadap lingkungan barunya yang dipindahkan dari lingkungan indoor menuju outdoor. Selain itu, Chaetoceros gracilis memiliki laju pertumbuhan spesifik maksimum
sebesar 0,268.
Fase lagChaetoceros gracilis dapat diduga terjadi dalam waktu kurang dari 24
jam. Hal ini dapat dilihat dari nilai kelimpahan dan bentuk grafik pada Gambar 8 yang terjadi pada hari ke-1 yang menunjukan pola pertumbuhan yang langsung memasuki fase eksponensial. Fase eksponensial terjadi pada waktu kurang dari 24
jam hingga hari ke-7. Fase stasioner dapat diduga terjadi pada hari ke-7 hingga hari ke-8 dan dapat diduga terjadi dalam rentang waktu kurang dari 24 jam. Kondisi ini
terlihat dari bentuk grafik pertumbuhan yang langsung memperlihatkan penurunan nilai kelimpahan yang curam antara hari ke-7 dan ke-8 sehingga pendugaan fase stasioner terjadi dalam waktu kurang dari 24 jam. Fase deklinasi terjadi pada waktu
kurang dari 24 jam di hari ke-8 hingga hari ke-15. Kondisi ini juga dapat dilihat dari nilai µ (µ = -0,141) yang bernilai negatif pada saat memasuki hari ke-8
kriptik di mana sel-sel Chaetoceros gracilis yang masih hidup memanfaatkan tambahan nutrisi dari sel-sel Chaetoceros gracilis yang lisis untuk pertumbuhannya
sehingga dapat meningkatkan populasinya kembali (Suantika, 2009). Fase deklinasi berlanjut kembali hingga hari ke-15 dengan jumlah kelimpahan akhir sebesar
0,57×106 sel/mL. Fase deklinasi dapat terjadi karena nutrisi kultur telah habis dan
terjadi akumulasi senyawa NH4+ dalam konsentrasi tinggi dan adanya produk
ekstraseluler dari mikroalga yang meracuni diri sendiri sehingga dapat meningkatkan
mortalitas Chaetoceros gracilis (Fogg, 1965 dalam Panggabean, 2000 dan Suantika, 2009).
Hasil yang dapat disimpulkan pada penelitian sistem outdoor adalah
Skeletonema costatum memiliki laju pertumbuhan spesifik maksimum yang lebih tinggi dibandingkan dua spesies lainnya yaitu 0,506 dengan kelimpahan maksimum
sebesar 1,78×106 sel/mL. Chaetoceros gracilis memiliki laju pertumbuhan spesifik maksimum sebesar 0,268 dengan nilai kelimpahan maksimum sebesar 2,79×106 sel/mL. Thalassiosira sp. memilki nilai µmaks sebesar 0,205 dengan kelimpahan
sebesar 0,47×106 sel/mL. Laju pertumbuhan spesifik maksimum yang dihitung dari awal kultur hingga puncak maksimum tertinggi terdapat pada Skeletonema costatum
namun kelimpahan tertinggi terdapat pada spesies Chaetoceros gracilis. Walaupun nilai kelimpahan maksimum lebih besar namun belum tentu nilai µmaks lebih besar. Keadaan ini dipengaruhi oleh jumlah kelimpahan awal strain yang dituangkan pada
4.3 Perbandingan Kelimpahan dan Laju Pertumbuhan Spesifik pada Kedua Sistem
4.3.1 Skeletonema costatum
Pola pertumbuhan Skeletonema costatum pada sistem indoor memiliki satu
buah puncak populasi yaitu pada hari ke-5 dengan nilai kelimpahan sebesar 1,97×106 sel/mL. Pola pertumbuhan Skeletonema costatum pada sistem outdoor
juga memiliki satu buah puncak populasi pada hari ke-5 namun dengan nilai kelimpahan yang lebih kecil sebesar 1,61×106 sel/mL. Skeletonema costatum saat hari ke-1 pada sistem indoor memiliki daya adaptasi terhadap lingkungan yang lebih
tinggi (µ = 1,754) dibandingkan pada sistem outdoor (µ = 1,084). Hal ini diduga karena media strain sebelumnya memiliki lingkungan yang sama pada media strain
yang digunakan pada penelitian pada sistem indoor sedangkan pada media outdoor, media strain sebelumnya memiliki kondisi lingkungan yang berbeda dengan kondisi yang relatif terkontrol pada sistem indoor yang dipindahkan ke dalam media outdoor
yang tidak terkontrol lingkungannya. µmaks yang dihitung pada awal kultur hingga puncak kelimpahan maksimum pada spesies Skeletonema costatum sistem indoor
adalah sebesar 0,515 sedangkan pada sistem outdoor memiliki nilai µmaks lebih rendah yaitu 0,506. Hal ini menunjukan bahwa laju pertumbuhan spesifik
maksimum pada sistem indoor lebih tinggi dibandingkan laju pertumbuhan spesifik
Gambar 9. Kelimpahan Skeletonema costatum pada sistem indoor dan outdoor
Fase lagSkeletonema costatum pada kedua sistem diduga terjadi kurang dari 24 jam. Keadaan ini dapat dilihat dari kelimpahan dan nilai µ pada hari ke-1 pada kedua sistem yang memiliki nilai yang cukup besar. Skeletonema costatum pada
kedua sistem diduga memiliki daya adaptasi yang cukup tinggi berdasarkan nilai µ yang dihitung. Fase eksponensial pada kedua sistem juga diduga terjadi dalam
waktu kurang dari 24 jam. Fase eksponensial pada sistem indoor terjadi hingga hari ke-4 dan sistem outdoor terjadi hingga hari ke-3. Hal ini memperlihatkan bahwa fase eksponensial pada sistem outdoor lebih cepat dibandingkan fase eksponensial
pada sistem indoor.
Fase stasioner pada sistem indoor diduga terjadi pada hari 4 hingga hari