PENAMBAHAN GLUTAMAT SEBAGAI PELINDUNG
TEKANAN OSMOSIS DALAM KONDISI PEMBIAKAN
ETANOL TINGGI PADA REKOMBINAN
Escherichia coli
ARI PERMANA PUTRA
DEPARTEMEN TEKNOLOGI INDUSTRI PERTANIAN FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR BOGOR
PERNYATAAN MENGENAI SKRIPSI DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa skripsi berjudul Penambahan Glutamat sebagai Pelindung Tekanan Osmosis dalam Kondisi Pembiakan Etanol Tinggi pada Rekombinan Escherichia coli adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir skripsi ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Desember 2014
ABSTRAK
ARI PERMANA PUTRA. Penambahan Glutamat sebagai Pelindung Tekanan Osmosis dalam Kondisi Pembiakan Etanol Tinggi pada Rekombinan Escherichia coli. Dibimbing oleh PRAYOGA SURYADARMA.
Peningkatan pertumbuhan sel rekombinan Escherichia coli pada kondisi etanol tinggi diinvestigasi dengan penambahan glutamat sebagai pelindung tekanan osmosis. Pengaruh glutamat tersebut juga didukung melalui analisis bobot sel kering, konsumsi glukosa, dan akumulasi asetat. Sel E. coli ditumbuhkan dalam kondisi aerob dengan penambahan etanol dan glutamat pada jam ke-6. Pada pembiakan dengan kondisi etanol tinggi (etanol 20 g/l) tanpa penambahan glutamat, bobot sel kering dan konsumsi glukosa berturut-turut mencapai 0.91±0.14 g/l dan 13.23±0.89 g/l. Nilai tersebut lebih rendah dibandingkan dengan kondisi pembiakan dengan penambahan glutamat 2 g/l (bobot sel kering 1.8±0.03 g/l dan konsumsi glukosa 17.75±0.43 g/l), dengan akumulasi asetat juga ditekan sebesar 3.29 g/l. Berdasarkan aliran karbon pada pusat metabolisme karbon, akumulasi asetat dihambat melalui penambahan glutamat karena aliran karbon beralih menuju siklus tricarboxylic acid (TCA) sebagai ganti menuju asetat. Akumulasi asetat juga menunjukkan sebagai hasil dari metabolisme berlebih (overflow metabolism) karena tingginya konsumsi glukosa dan level oksigenasi.
ABSTRACT
ARI PERMANA PUTRA. Glutamate Additions as Osmoprotectant under High Ethanol Culture Conditions in Escherichia coli Recombinant. Supervised by PRAYOGA SURYADARMA.
Enhancement of Escherichia coli recombinant cell growth in high ethanol culture conditions was investigated by additional glutamate as osmoprotectant. It was also supported by analysis of dry cell weight (DCW), glucose uptake, and acetate accumulation. Escherichia coli cells were grown under aerobic conditions with ethanol and glutamate addition in the culture time of t = 6 h. In high ethanol culture conditions (20 g/l ethanol) without glutamate acid additions, DCW and glucose uptake reached in 0.91±0.14 g/l and 13.23±0.89 g/l, respectively. These values were lower than culture conditions with 2 g/l glutamate acid additions (1.8±0.03 g/l DCW and 17.75±0.43 g/l glucose uptake), which also suppressed the accumulation of acetate in 3.29 g/l. Based on carbon flow in central carbon metabolism, the accumulation of acetate was suppressed by glutamate additions in the culture because the carbon flow diversion to tricarboxylic acid (TCA) cycle instead of entering the acetate. The accumulation of acetate also showed as a result of an overflow metabolism by high rate of glucose uptake and oxygenation level.
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Teknologi Pertanian
pada
Departemen Teknologi Industri Pertanian
PENAMBAHAN GLUTAMAT SEBAGAI PELINDUNG
TEKANAN OSMOSIS DALAM KONDISI PEMBIAKAN
ETANOL TINGGI PADA REKOMBINAN
Escherichia coli
ARI PERMANA PUTRA
DEPARTEMEN TEKNOLOGI INDUSTRI PERTANIAN FAKULTAS TEKNOLOGI PERTANIAN
INSTITUT PERTANIAN BOGOR BOGOR
Judul Skripsi : Penambahan Glutamat sebagai Pelindung Tekanan Osmosis dalam Kondisi Pembiakan Etanol Tinggi pada Rekombinan Escherichia coli
Nama : Ari Permana Putra NIM : F34100142
Disetujui oleh
Dr Prayoga Suryadarma, S.TP, MT NIP. 19741110 199903 1 001
Diketahui oleh
Prof Dr Ir Nastiti Siswi Indrasti NIP. 19621009 198903 2 001
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah subhanahu wa ta’ala atas segala karunia-Nya sehingga skripsi ini berhasil diselesaikan. Judul yang dipilih dalam penelitian ini adalah Penambahan Glutamat sebagai Pelindung Tekanan Osmosis dalam Kondisi Pembiakan Etanol Tinggi pada Rekombinan Escherichia coli.
Terima kasih penulis ucapkan kepada Bapak Dr Prayoga Suryadarma, S.TP, MT selaku dosen pembimbing dan seluruh civitas Departemen Teknologi Industri Pertanian atas segala ilmu yang diberikan. Penghargaan juga penulis sampaikan kepada pimpinan Pusat Penelitian Sumberdaya Hayati dan Bioteknologi (PPSHB) Pusat Antar Universitas IPB yang telah mengizinkan penulis belajar rekayasa genetika di Laboratorium Rekayasa Bioproses. Tidak lupa ungkapan terima kasih juga disampaikan penulis kepada ayah, ibu, keluarga, serta seluruh sahabat atas segala doa dan dukungannya, supervisor dan rekan-rekan “strategis” program beasiswa PPSDMS Nurul Fikri angkatan 6 regional Bogor atas bantuan dana, pembinaan karakter, dan persahabatannya, dan rekan-rekan TIN 47 yang selalu mendukung dan memberi semangat kepada penulis.
Semoga skripsi ini dapat bermanfaat bagi pembaca dan memberikan kontribusi dalam perkembangan ilmu pengetahuan.
Bogor, Desember 2014
DAFTAR ISI
DAFTAR TABEL vii
DAFTAR GAMBAR vii
DAFTAR LAMPIRAN vii
PENDAHULUAN 1
Latar Belakang 1
Perumusan Masalah 2
Hipotesis 2
Tujuan Penelitian 3
Manfaat Penelitian 3
METODE 4
Bahan 4
Alat 4
Tahapan Penelitian 4
HASIL DAN PEMBAHASAN 7
Pengaruh Penambahan Etanol Terhadap Pertumbuhan Sel 7
SIMPULAN DAN SARAN 13
Simpulan 13
Saran 13
DAFTAR PUSTAKA 13
LAMPIRAN 16
DAFTAR TABEL
1 Perkembangan penelitian produksi etanol 1
DAFTAR GAMBAR
1 Strategi penambahan glutamat untuk memperbaiki dinding sel akibat
tekanan osmosis oleh etanol tinggi 3
2 Tahapan penelitian 5
3 Penyisipan plasmid pTadhB-pdc dan pHfdh ke dalam galur E. coli
BW25113Δpta 6
4 Pengaruh penambahan etanol terhadap bobot sel kering E. coli
BW25113Δpta/pHfdh/pTadhB-pdc. Pembiakan dilakukan selama 24 jam dengan penambahan etanol dilakukan pada jam ke-6 8 5 Pengaruh penambahan glutamat terhadap bobot sel kering E. coli
BW25113Δpta/pHfdh/pTadhB-pdc. Pembiakan dilakukan selama 24 jam dengan penambahan etanol 20 g/l dan glutamat dilakukan pada jam
ke-6 9
6 Konsumsi glukosa oleh biakan E. coli BW25113Δpta /pHfdh/pTadhB-pdc. Pembiakan dilakukan selama 24 jam dengan penambahan etanol
20 g/l dan glutamat dilakukan pada jam ke-6 10
7 Produksi asetat sel E. coli BW25113Δpta/pHfdh/pTadhB-pdc dalam
media pembiakan selama 24 jam 11
8 Pusat metabolisme karbon pada E. coli BW25113Δpta /pHfdh/pTadhB-pdc. Tanpa penambahan glutamat (A). Dengan penambahan glutamat
(B) 12
DAFTAR LAMPIRAN
1 Prosedur transformasi metode kejut panas(Panja et al. 2006) 17 2 Prosedur pembuatan persediaan biakan (Lustig et al. 1996) 17
3 Prosedur prapembiakan (Ojima et al. 2012) 17
4 Prosedur pembiakan (Ojima et al. 2012) 18
5 Prosedur analisis bobot sel kering (Ojima et al. 2012) 18 6 Prosedur analisis glukosa (F-Kit 716251) (Suryadarma et al. 2012) 18
7 Prosedur analisis asetat (Stimson 2011) 19
PENDAHULUAN
Latar Belakang
Salah satu permasalahan yang dihadapi oleh berbagai negara adalah krisis energi. Krisis energi terjadi karena adanya peningkatan kebutuhan bahan bakar yang tidak diimbangi dengan ketersediaan yang mencukupi. Oleh sebab itu, diperlukan adanya bahan bakar alternatif untuk mengatasi masalah ini.
Menurut Hahn et al. (2006), solusi untuk menghadapi krisis energi dan dampak negatif gas rumah kaca dari bahan bakar fosil adalah etanol. Pada tahun 1900, Amerika pertama kalinya memproduksi etanol dari pati jagung dengan menggunakan Saccharomyces cerevisiae (Bothast dan Schlicher 2005). Namun, penggunaan pati jagung sebagai bahan baku etanol akan berdampak buruk terhadap ketersediaan kebutuhan pangan. Di sisi lain, bahan lignoselulosa dari limbah pertanian dan perhutanan yang jumlahnya tersedia melimpah di alam, memiliki kandungan gula pentosa yang sangat berpotensi untuk dikonversi menjadi etanol (Deanda et al. 1996; Wyman 2003). Selain itu, penggunaan limbah lignoselulosa juga tidak akan berdampak buruk terhadap kebutuhan pangan. Oleh sebab itu, diperlukan mikroorganisme yang mampu menghasilkan etanol dari limbah lignoselulosa. Tabel 1 menunjukkan perkembangan penelitian produksi etanol dari berbagai macam mikroorganisme.
Tabel 1 Perkembangan penelitian produksi etanol
Kemajuan rekayasa genetika telah membuktikan bahwa Escherichia coli mampu memproduksi etanol melalui transformasi gen pembentuk etanol dari Zymomonas mobilis (Ingram et al. 1987). Kemampuan dalam memproduksi etanol pada E. coli juga didukung dengan kemampuannya dalam mengkonsumsi berbagai macam gula sederhana. Escherichia coli mampu mengkonsumsi berbagai macam gula sederhana termasuk gula pentosa yang ada pada bahan lignoselulosa (Alterthum dan Ingram 1989). Akan tetapi, kemampuan tersebut
Mikroorganisme Kondisi Proses Etanol (g/l)
tidak dimiliki oleh Zymomonas mobilis dan Saccharomyces cerevisiae. Secara alami, kedua mikroorganisme tersebut hanya mampu mengkonsumsi glukosa (Hahn et al. 2006). Dengan demikian, melimpahnya limbah lignoselulosa menjadikan E. coli sangat berpotensi sebagai mikroorganisme penghasil etanol.
Tabel 1 telah menunjukkan kemampuan E. coli dalam memproduksi etanol dengan menggunakan berbagai macam gula sederhana. Namun, produksi etanol tersebut berlangsung pada kondisi anaerob. Hal ini mengakibatkan pertumbuhan sel menjadi lambat sehingga metabolisme E. coli terganggu. Selain itu, kondisi anaerob juga mengakibatkan terbentuknya produk samping seperti laktat oleh enzim laktat dehidrogenase (LDH) (Suryadarma et al. 2012). Dengan demikian, etanol yang terbentuk akan menjadi rendah.
Sementara pada kondisi aerob, aktifitas enzim LDH menjadi inaktif karena enzim tersebut sensitif terhadap oksigen. Hal ini mengakibatkan produksi laktat sebagai produk samping terhambat (Clark 1989). Kondisi aerob juga mampu mengubah aliran karbon menuju siklus TCA karena adanya penggunaan NADH untuk respirasi. Hal tersebut mengakibatkan pertumbuhan sel meningkat (Suryadarma et al. 2012). Dengan demikian, terhambatnya produksi laktat dan meningkatnya pertumbuhan sel pada kondisi aerob, mampu meningkatkan produksi etanol.
Perumusan Masalah
Pengaturan kondisi pembiakan E. coli telah diupayakan untuk meningkatkan produksi etanol dan mengurangi produk samping. Pengaturan kondisi aerob mampu meningkatkan aktifitas siklus TCA sehingga pertumbuhan sel meningkat. Dengan meningkatnya pertumbuhan sel, maka produksi etanol juga akan meningkat. Akan tetapi, produksi etanol yang tinggi bisa menjadi penghambat bagi E. coli. Konsentrasi etanol yang tinggi akan mengakibatkan tekanan osmosis pada dinding sel sehingga akan menurunkan ketahanan dinding sel (Strom 1998).
Adanya kerusakan dinding sel mengakibatkan pertumbuhan sel menjadi rendah. Terhambatnya pertumbuhan sel mengindikasikan adanya pemanfaatan lain dari glukosa yang dikonsumsi, seperti pembentukkan produk samping, asetat. Terbentuknya asetat juga mengindikasikan sedikitnya aliran karbon menuju siklus TCA sehingga mengakibatkan pertumbuhan sel rendah.
Oleh sebab itu, diperlukan strategi yang mampu mengatasi kerusakan dinding sel akibat tekanan osmosis. Adanya pelindung tekanan osmosis diharapkan mampu meningkatkan ketahanan dinding sel. Salah satu pelindung tekanan osmosis adalah glutamat yang merupakan asam amino penting penyusun dinding sel (Kim et al. 2007).
Hipotesis
Glutamat
untuk memperkuat ketahanan dinding sel terhadap tekanan osmosis (Lundqvist et al. 2007). Dengan adanya penambahan glutamat maka akan meningkatkan komponen D-glutamat pada peptidoglikan sehingga mampu meningkatkan ketahanan dinding sel terhadap etanol tinggi.
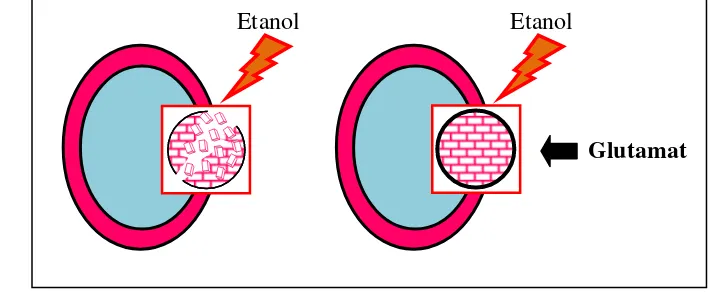
Gambar 1 menunjukkan strategi yang akan digunakan dalam penelitian ini. Penambahan etanol pada konsentrasi tinggi akan merusak dinding sel sehingga ketahanan dinding sel menurun. Hal ini akan mengakibatkan pertumbuhan sel menjadi rendah. Sementara itu, dengan adanya penambahan glutamat yang berperan sebagai penguat dinding sel, diperkirakan mampu memperbaiki kerusakan dinding sel sehingga meningkatkan ketahanan dinding sel. Dengan meningkatnya ketahanan dinding sel akibat penambahan glutamat, diharapkan pertumbuhan sel akan tetap tinggi.
Gambar 1 Strategi penambahan glutamat untuk memperbaiki dinding sel akibat tekanan osmosis oleh etanol tinggi
Tujuan Penelitian
Tujuan penelitian ini adalah untuk mengetahui pengaruh penambahan etanol sebagai penyebab tekanan osmosis dan peranan glutamat sebagai pelindung tekanan osmosis untuk meningkatkan pertumbuhan sel.
Manfaat Penelitian
Penelitian ini bermanfaat untuk memberikan informasi mengenai strategi mengatasi penurunan pertumbuhan sel akibat produksi etanol yang tinggi pada E. coli dalam kondisi aerob.
METODE
Bahan
Galur (strain) induk E. coli yang digunakan dalam penelitian ini adalah BW25113Δpta. Plasmid yang akan disisipkan ke dalam galur tersebut yaitu pTadhB-pdc dan pHfdh. Bahan yang digunakan dalam transformasi yaitu ampisilin, kloramfenikol, kanamisin, CaCl2 0.1 M, dimethyl sulfoxide (DMSO), dan 2xYT.
Media yang digunakan untuk prapembiakan mengandung 5 g ekstrak khamir, 10 g pepton, dan 10 g NaCl yang dilarutkan dalam 1 l aquades. Bahan yang digunakan untuk pembuatan persediaan biakan adalah gliserol 50%. Media yang digunakan untuk pembiakan mengandung bahan-bahan antara lain 40 g glukosa, 5 g ekstrak khamir, 10 g pepton, 10 g NaCl, dan 4 g sodium format yang dilarutkan dalam 1 l aquades. Selain itu juga ditambahkan isopropyl thiogalactoside (IPTG) 0.5 mM, ampisilin 50 mg/l, kloramfenikol 34 mg/l, kanamisin 15 mg/l, CaCO3 20 g/l, etanol 20 g/l, dan sodium glutamat 2 g/l.
Alat
Alat yang digunakan untuk transformasi yaitu clean bench, tabung reaksi, pipet mikro, spektrofotometer, kultur bergoyang 3000 rpm 4 °C, eppendorf 1.5 ml, cool box, lemari pendingin, penangas air, kultur bergoyang 250 rpm 37 °C, timbangan analitik, otoklaf 121 °C, cawan petri, erlenmeyer, bunsen, dan GeneJETTM miniprep kit. Alat yang digunakan untuk pembuatan persediaan biakan yaitu eppendorfdan pipet mikro.
Alat yang digunakan untuk prapembiakan yakni lain clean bench, erlenmeyer, gelas ukur, gelas piala, pH meter, timbangan analitik, otoklaf 121 °C, kultur bergoyang 120 rpm 37 °C, dan pipet mikro. Alat yang digunakan untuk pembiakan adalah clean bench, spektrofotometer Hach DR 2500, baffled conical flask 250 ml, stopper, pipet mikro, dan kultur bergoyang 250 rpm 37 °C. Alat yang digunakan untuk analisis yaitu eppendorf, syringe filter 0.2 µm, sentrifugator 10 000 rpm 4 °C, tabung ulir, spektrofotometer Hach DR 2500, pipet volumetrik 10 ml, pipet mikro, pH meter, spektrofotometer UV, vortex, HPLC, dan kolom ZORBAX SB-Aq 883975-914.
Tahapan Penelitian
Gambar 2 Tahapan penelitian Penyiapan Biakan
Pada penelitian ini, penyiapan biakan meliputi dua tahap yaitu transformasi dan pembuatan persediaan biakan. Transformasi dilakukan untuk memasukkan molekul DNA ke dalam sel bakteri. Metode transformasi yang dilakukan pada penelitian ini adalah kejut panas. Prinsip metode ini adalah dengan memberikan kejut panas sehingga pori membran sel akan terbuka.
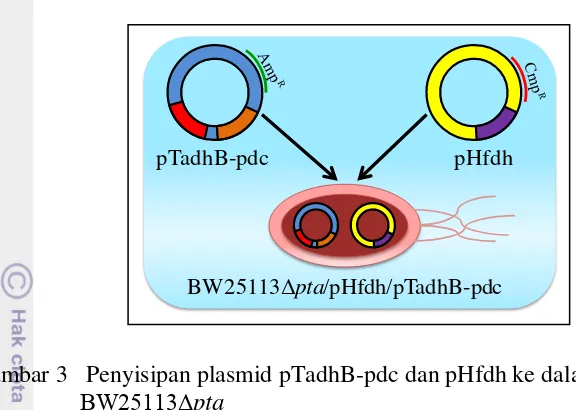
Metode transformasi ini diawali dengan pembuatan sel kompeten yaitu sel yang memiliki kemampuan untuk disisipi DNA dari luar. Penggunaan CaCl2 dalam pembuatan sel kompeten bisa mempengaruhi porositas membran sel sehingga membran menjadi tidak selektif lagi ketika ada lonjakan suhu. Pada penelitian ini, plasmid yang akan disisipkan ke dalam sel E. coli adalah plasmid pHfdh dan pTadhB-pdc (Gambar 3). Plasmid pTadhB-pdc tahan terhadap antibiotik ampisilin sedangkan plasmid pHfdh tahan terhadap antibiotik kloramfenikol. Plasmid pHfdh mengandung gen fdh yang bisa menghasilkan NADH. Sedangkan plasmid pTadhB-pdc mengandung gen adh dan pdc yang bisa menghasilkan etanol. Kedua plasmid tersebut didapatkan dari universitas Osaka, Jepang. Prosedur transformasi dengan metode kejut panas dapat dilihat pada Lampiran 1.
Pembuatan persediaan biakan bertujuan untuk menyimpan galur E. coli hasil transformasi. Dengan demikian, untuk melakukan prapembiakan tidak perlu mengulang pembuatan galur dari proses transformasi lagi. Prosedur pembuatan persediaan biakan dapat dilihat pada Lampiran 2.
Mulai
Penyiapan biakan
Prapembiakan
Pembiakan
Gambar 3 Penyisipan plasmid pTadhB-pdc dan pHfdh ke dalam galur E. coli
BW25113Δpta
Prapembiakan
Prapembiakan dilakukan untuk menyegarkan kembali sel E. coli sehingga pertumbuhannya bisa optimal ketika pembiakan. Pertumbuhan sel diukur dengan mengukur nilai kerapatan optik (OD) pada panjang gelombang 660 nm. Prosedur prapembiakan dapat dilihat pada Lampiran 3.
Pembiakan
Pembiakan dilakukan secara aerobik dengan laju agitasi 250 rpm selama 24 jam pada volume media 50 ml. Sel E. coli yang telah diprapembiakan hingga nilai OD660 1–1.5 kemudian diinokulasi ke dalam media pembiakan yang mengandung ampisilin, kloramfenikol, kanamisin, IPTG, sodium format, dan CaCO3. Penambahan sodium format bertujuan untuk meningkatkan NADH. Sementara itu, penambahan CaCO3 bertujuan untuk mencegah penurunan pH selama pembiakan.
Pada saat jam ke-6 dilakukan penambahan etanol 20 g/l dan glutamat. Penambahan glutamat dilakukan dengan konsentrasi 2, 4, dan 6 g/l. Lampiran 4 menunjukkan prosedur pembiakan.
Analisis
Analisis pada penelitian ini meliputi bobot sel kering, glukosa, dan asetat. Pengambilan sampel untuk analisis dilakukan pada jam ke-24. Sampel untuk masing-masing analisis disimpan dalam eppendorf. Sampel untuk analisis glukosa dan asetat terlebih dahulu disentrifus untuk memisahkan supernatan dari pelet kemudian difilter dengan syringe filter 0.2 µm.
Analisis bobot sel kering dilakukan untuk mengetahui pertumbuhan sel. Pertumbuhan sel dihitung dengan mengukur nilai OD660. Nilai tersebut kemudian dikonversi dengan cara mengalikannya dengan 0.36 yang merepresentasikan sebagai bobot sel kering (Ojima et al. 2012). Sebelum mengukur nilai OD660, sampel terlebih dahulu dicampur dengan HCl 1 M untuk melarutkan CaCO3 (Suryadarma et al. 2012). Prosedur analisis bobot sel kering dapat dilihat pada Lampiran 5.
pTadhB-pdc pHfdh
Analisis glukosa bertujuan untuk mengetahui jumlah glukosa yang dikonsumsi. Analisis ini dilakukan secara enzimatik menggunakan F-Kit 716251. Lampiran 6 menyajikan prosedur analisis glukosa.
Analisis asetat bertujuan untuk mengetahui jumlah asetat yang terbentuk. Asetat dianalisis menggunakan HPLC dengan kolom ZORBAX SB-Aq 883975-914. Suhu, fase bergerak, laju alir, dan panjang gelombang yang digunakan, berturut-turut yakni 35 ºC, NaH2PO4 20 mM dan ACN, 1 ml/menit, dan 210 nm. Prosedur analisis asetat bisa dilihat pada Lampiran 7.
Data yang diperoleh pada penelitian ini direpresentasikan sebagai nilai rata-rata±simpangan baku dengan pengulangan sebanyak tiga kali kecuali pada analisis asetat. Pengolahan data berupa uji beda dilakukan dengan menggunakan uji-T pada microsoft office excel 2007.
HASIL DAN PEMBAHASAN
Pengaruh Penambahan Etanol Terhadap Pertumbuhan Sel
Kemajuan rekayasa genetika telah membuktikan bahwa Escherichia coli mampu memproduksi etanol (Ingram et al. 1987). Akan tetapi, meningkatnya produksi etanol bisa menjadi penghambat bagi pertumbuhan E. coli (Ingram 1986). Penelitian ini diawali dengan menguji pengaruh penambahan etanol pada konsentrasi berbeda terhadap E. coli BW25113Δpta/pHfdh/pTadhB-pdc. Pengujian tersebut dilakukan untuk mengetahui besarnya konsentrasi etanol yang mampu menghambat pertumbuhan sel (Gambar 4).
0
BW25113Δpta/pHfdh/pTadhB-pdc. Pembiakan dilakukan selama 24 jam dengan penambahan etanol dilakukan pada jam ke-6
Penurunan pertumbuhan sel akibat penambahan etanol 20 g/l mengindikasikan bahwa pertumbuhan sel Escherichia coli terhambat dengan kehadiran etanol yang tinggi (20 g/l). Hal ini juga telah dibuktikan dalam penelitian sebelumnya, penambahan etanol 10 g/l mampu memberikan pengaruh terhadap penurunan pertumbuhan sel E. coli K 12 dalam kondisi anaerob (Huffer et al. 2011). Etanol merupakan molekul amfipatik karena molekulnya memiliki gugus polar (hidrofilik) dan gugus nonpolar (hidrofobik). Gugus hidrofilik etanol akan berikatan dengan air sehingga etanol larut dalam media. Kemudian etanol akan dibawa masuk ke dalam sel oleh protein pembawa. Gugus hidrofobik etanol akan berikatan protein pembawa untuk berpenetrasi menembus lapisan fosfolipid membran sel. Dengan demikian, etanol mampu masuk ke dalam sel. Masuknya etanol ke dalam sel mengakibatkan tidak terjadinya ikatan silang pada struktur peptidoglikan sehingga akan mengurangi kekuatan peptidoglikan (Ingram dan Vreeland 1980). Dengan demikian telah mengkonfirmasi bahwa konsentrasi etanol tinggi mampu mengganggu ketahanan dinding sel sehingga menghambat pertumbuhan sel E. coli pada kondisi aerobik.
Untuk mengatasi penghambatan pertumbuhan sel oleh etanol, maka dilakukan penelitian penambahan glutamat. D-glutamat merupakan salah satu asam amino penting penyusun peptidoglikan pada dinding sel yang dihasilkan dari konversi L-glutamat dengan bantuan enzim glutamate racemase (MurI) (Kim et al. 2007). D-glutamat yang terbentuk akan berikatan dengan UDP-N-acetylmuramyl-L-alanin melalui ikatan peptida. Selanjutnya, terjadi penggabungan dengan mes-diaminopimelic acid, D-alanil, dan D-alanin hingga membentuk monomer peptidoglikan UDP-NAM-pentapeptida (Ho et al. 1994). Adanya peranan glutamat dalam memperkuat dinding sel telah dibuktikan dengan terjadinya penurunan pertumbuhan sel pada penelitian sebelumnya. Sel E. coli WM335 pada kondisi anaerob mengalami penurunan pertumbuhan ketika dipindahkan dari media yang mengandung glutamat ke media tanpa glutamat.
*
0
Penurunan pertumbuhan ini terjadi karena peptidoglikan mengalami lisis akibat kekurangan glutamat (Doublet et al. 1992).
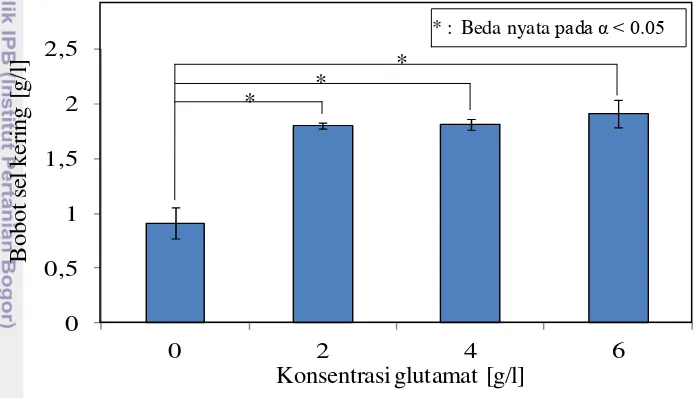
Pengaruh glutamat sebagai penguat peptidoglikan telah terbukti pada kondisi anaerob. Untuk mengetahui pengaruh penambahan glutamat terhadap tekanan osmosis akibat etanol 20 g/l pada kondisi aerob, maka dilakukan penelitian pengaruh penambahan glutamat terhadap pertumbuhan sel (Gambar 5). Peningkatan konsentrasi glutamat mampu meningkatkan pertumbuhan sel. Konsentrasi glutamat 2 g/l memberikan pengaruh yang berbeda nyata terhadap peningkatkan bobot sel kering yaitu hampir dua kali lipat dari perlakuan tanpa penambahan glutamat. Selanjutnya, peningkatan konsentrasi glutamat hingga 6 g/l tidak menunjukkan perbedaan bobot sel kering yang berbeda nyata terhadap konsentrasi sebelumnya. Peningkatan pertumbuhan tersebut mengindikasikan adanya peranan glutamat dalam memberi suplemen terhadap peptidoglikan sehingga dinding sel tahan terhadap etanol 20 g/l.
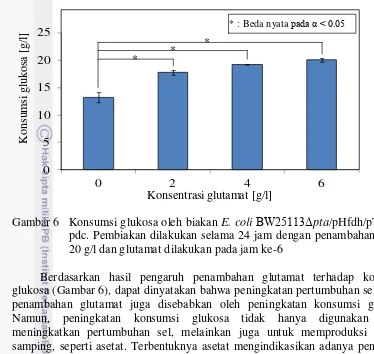
Untuk mengklarifikasi adanya pengaruh penambahan glutamat terhadap peningkatan pertumbuhan sel, maka dilakukan analisis glukosa sebagai sumber karbon yang dikonsumsi E. coli (Gambar 6). Semakin meningkatnya konsentrasi glutamat, semakin banyak glukosa yang dikonsumsi. Penambahan glutamat 2 g/l mengakibatkan konsumsi glukosa meningkat drastis sebesar 4.5 g/l dari perlakuan tanpa penambahan glutamat.
Gambar 5 Pengaruh penambahan glutamat terhadap bobot sel kering E. coli
BW25113Δpta/pHfdh/pTadhB-pdc. Pembiakan dilakukan selama 24 jam dengan penambahan etanol 20 g/l dan glutamat dilakukan pada jam ke-6
* : Beda nyata pada α < 0.05
*
0
Gambar 6 Konsumsi glukosa oleh biakan E. coliBW25113Δpta /pHfdh/pTadhB-pdc. Pembiakan dilakukan selama 24 jam dengan penambahan etanol 20 g/l dan glutamat dilakukan pada jam ke-6
Berdasarkan hasil pengaruh penambahan glutamat terhadap konsumsi glukosa (Gambar 6), dapat dinyatakan bahwa peningkatan pertumbuhan sel akibat penambahan glutamat juga disebabkan oleh peningkatan konsumsi glukosa. Namun, peningkatan konsumsi glukosa tidak hanya digunakan untuk meningkatkan pertumbuhan sel, melainkan juga untuk memproduksi produk samping, seperti asetat. Terbentuknya asetat mengindikasikan adanya penurunan pertumbuhan sel (Luli dan Strohl 1990). Dengan demikian, penting untuk mengetahui produksi asetat dalam metabolisme E. coli.
Asetat adalah produk samping yang dihasilkan dari jalur metabolisme E. coli. Asetat terbentuk karena adanya perubahan aliran karbon. Aliran karbon yang seharusnya menuju siklus TCA dari asetil CoA berubah menuju asetat melalui jalur PTA-ACK (phosphotransacetylase-acetate kinase). Pengalihan aliran ini terjadi karena tingginya NADH pada asetil CoA. Tingginya NADH bisa menjadi penghambat bagi enzim sitrat sintase untuk aktifitas siklus TCA sehingga mengakibatkan perubahan aliran karbon menuju asetat (Vemuri et al. 2006). Perubahan aliran karbon akibat pembentukkan asetat dapat mengakibatkan menurunnya pertumbuhan sel dan produk protein rekombinan lainnya (Underwood et al. 2002). Selain itu, pembentukkan asetat juga bisa melalui jalur POX (pyruvate oksidase) yang mengkonversi piruvat secara langsung menjadi asetat (Ojima et al. 2012).
Terbentuknya asetat mengindikasikan terjadinya metabolisme berlebih pada metabolisme E. coli. Terjadinya fenomena tersebut disebabkan oleh kondisi aerobik dan kandungan glukosa yang tinggi (Vemuri et al. 2006). Pada penelitian sebelumnya, penggunaan galur E. coli MC1060 dengan media pembiakan sebanyak 27 ml dalam flask 300 ml yang mengandung glukosa 5 g/l pada laju agitasi 300 rpm mampu menghasilkan asetat sebanyak 0.92 g/l (Luli dan Strohl 1990). Terbentuknya asetat tersebut membuktikan tingginya konsumsi glukosa dan laju oksigenasi. Pada kondisi aerob atau level oksigenasi tinggi, sel menggunakan NADH untuk aktifitas respirasi. Aktifitas tersebut mengoksidasi
* : Beda nyata pada α < 0.05
*
0 glikolisis dan aktivitas siklus TCA. Meningkatnya konsumsi glukosa akan menghasilkan NADH yang tinggi pula. Sementara itu, peningkatan aktifitas siklus TCA juga akan menghasilkan NADH. Dengan demikian, akan terjadi akumulasi NADH. Tingginya ketersediaan NADH akan menjadi hambatan alosterik bagi enzim sitrat sintase untuk aktifitas siklus TCA. Hal ini mengakibatkan sedikitnya aliran karbon menuju siklus TCA sehingga terjadi pembentukkan asetat (Eiteman dan Altman 2006; Vemuri et al. 2006). Dengan demikian, kondisi aerobik dan kandungan glukosa yang tinggi telah berpengaruh terhadap pembentukkan asetat.
Untuk mengklarifikasi pengaruh peningkatan konsumsi glukosa terhadap pertumbuhan sel, maka dilakukan analisis asetat. Gambar 7 menunjukkan perbedaan produksi asetat oleh sel E. coli pada perlakuan dengan dan tanpa penambahan glutamat. Kurva asetat hasil analisis dengan HPLC bisa dilihat pada Lampiran 8. Asetat yang terbentuk pada perlakuan dengan penambahan glutamat 2 g/l menurun hampir dua kali lipat dari perlakuan tanpa penambahan glutamat. Rendahnya produksi asetat pada perlakuan dengan penambahan glutamat mengkonfirmasi bahwa peningkatan konsumsi glukosa untuk meningkatkan pertumbuhan sel.
Gambar 7 Produksi asetat sel E. coli BW25113Δpta/pHfdh/pTadhB-pdc dalam media pembiakan selama 24 jam
Asetat Asetat
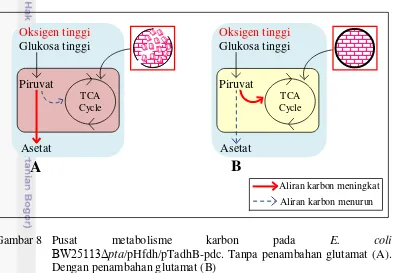
penambahan glutamat mengakibatkan terjadinya perubahan pada pusat metabolisme karbon E. coli. Perbaikan dinding sel tersebut mengakibatkan pertumbuhan sel meningkat. Hal ini mengindikasikan pemanfaatan sumber kabon dari glukosa untuk pertumbuhan sel. Peningkatan pertumbuhan sel akibat penambahan glutamat mengindikasikan banyaknya aliran karbon menuju siklus TCA sehingga asetat yang terbentuk rendah. Sebaliknya, tanpa penambahan glutamat mengakibatkan rendahnya pertumbuhan sel. Hal ini mengindikasikan sedikitnya pemanfaatan karbon untuk pertumbuhan sel. Dengan demikian, aliran karbon menuju asetat lebih banyak daripada menuju siklus TCA sehingga asetat yang terbentuk tinggi dan pertumbuhan sel rendah.
Gambar 8 Pusat metabolisme karbon pada E. coli
BW25113Δpta/pHfdh/pTadhB-pdc. Tanpa penambahan glutamat (A). Dengan penambahan glutamat (B)
Berdasarkan perbedaan hasil antara adanya pengaruh penambahan dan tanpa penambahan glutamat, mengindikasikan adanya perubahan pusat metabolisme karbon pada E. coli BW25113Δpta/pHfdh/pTadhB-pdc. Penurunan pertumbuhan akibat penambahan etanol 20 g/l telah mengkonfirmasi adanya kerusakan dinding sel akibat tekanan osmosis. Penambahan glutamat telah terbukti mampu mengatasi tekanan osmosis karena glutamat berperan sebagai penambah suplemen untuk menguatkan dinding sel. Hal tersebut telah dikonfirmasi dengan peningkatan pertumbuhan sel yang drastis pada penambahan glutamat 2 g/l (Gambar 5). Terjadinya peningkatan pertumbuhan sel akibat penambahan glutamat juga disebabkan oleh banyaknya glukosa yang dikonsumsi (Gambar 6). Adanya pengaruh konsumsi glukosa sebagai sumber karbon untuk meningkatkan pertumbuhan sel mengindikasikan banyaknya aliran karbon menuju siklus TCA. Hal tersebut dibuktikan dengan tingginya pertumbuhan sel dan rendahnya asetat yang terbentuk. Sebaliknya, rendahnya pertumbuhan sel akibat rendahnya konsumsi glukosa, mengindikasikan aliran karbon lebih banyak menuju asetat daripada menuju siklus TCA. Hal tersebut telah dikonfirmasi dengan tingginya asetat yang terbentuk dan rendahnya pertumbuhan sel (Gambar 5 dan Gambar 7).
Selain dipengaruhi oleh konsumsi glukosa yang tinggi, terbentuknya asetat juga disebabkan oleh oksigen yang tinggi (kondisi aerobik). Glukosa dan oksigen yang tinggi secara bersamaan berpengaruh terhadap meningkatnya ketersediaan NADH. Tingginya NADH akan menjadi penghambat bagi enzim sitrat sintase sehingga mengakibatkan perubahan aliran karbon menuju asetat. Dengan demikian, adanya penambahan glutamat telah mampu menekan pembentukkan asetat dan meningkatkan pertumbuhan sel.
SIMPULAN DAN SARAN
Simpulan
Terjadinya krisis energi mampu diatasi dengan adanya bahan bakar alternatif seperti etanol. Produksi etanol menggunakan E. coli mampu mencegah penggunaan pati dan gula tebu sebagai bahan baku etanol. Namun, produksi etanol tinggi mampu mengakibatkan tekanan osmosis sehingga mampu menghambat pertumbuhan sel E. coli. Adanya penambahan glutamat sebagai pelidung tekanan osmosis mampu meningkatkan ketahanan dinding sel sehingga pertumbuhan sel meningkat. Penambahan glutamat mengakibatkan perubahan padapusat metabolisme karbon E. coli sehingga meningkatkan konsumsi glukosa dan aliran karbon menuju siklus TCA. Meningkatnya aliran karbon menuju siklus TCA mengakibatkan tingginya pertumbuhan sel dan rendahnya asetat yang terbentuk. Sebaliknya, tanpa penambahan glutamat mengakibatkan rendahnya konsumsi glukosa dan sedikitnya aliran karbon menuju siklus TCA sehingga asetat yang terbentuk tinggi dan pertumbuhan sel rendah.
Saran
Perlu dilakukan penelitian lebih lanjut untuk mengetahui pengaruh penambahan glutamat terhadap produk-produk pada siklus TCA dan metabolisme lainnya pada E. coli. Selain itu, juga perlu diteliti pengaruh glutamat terhadap gen gltA yang berperan dalam siklus TCA.
DAFTAR PUSTAKA
Alterthum F dan LO Ingram. 1989. Efficient ethanol production from glucose, lactose, and xylose by recombinant Escherichia coli. Appl Environ Microbiol. 55: 1943–1948.
Bothast RJ dan MMA Schlicher. 2005. Biotechnological processes for conversion of corn into ethanol. Appl Microbiol Biotechnol. 67: 19–25.
Deanda K, M Zhang, C Eddy, dan S Picataggio. 1996. Development of an arabinose-fermenting Zymomonas mobilis strain by metabolic pathway engineering. Appl Environ Microbiol. 62: 4465–4470.
Doublet P, JV Heijenroot, dan DM Lecreulx. 1992. Identification of the Escherichia coli murI gene, which is required for the biosynthesis of D-glutamic acid, a specific component of bacterial peptidoglycan. J Bacteriol. 174: 5772–5779.
Eiteman MA dan E Altman. 2006. Overcoming acetate in Escherichia coli recombinant protein fermentations. Trends Biotechnol. 24: 530–536.
El-Mansi EMT dan WH Holms. 1989. Control of carbon flux to acetate excretion during growth of Escherichia coli in batch and continuous cultures. J Gen Microbiol. 135: 2875–2883.
Ghareib M, KA Youssef, dan AA Khalil. 1988. Ethanol tolerance of Saccharomyces cerevisiae and its relationship to lipid content and composition. Folia Microbiol. 33: 447–452.
Hahn-Hagerdal B, M Galbe, MF Gorwa-Grauslund, G Liden, dan G Zacchi. 2006. Bio-ethanol - the fuel of tomorrow from the residues of today. Trends Biotechnol. 24: 549–556.
Ho HT, PJ Falk, KM Ervin, BS Krishnan, LF Discotto, TJ Dougherty, dan MJ Pucci. 1994. UDP-N-acetylmutamyl-L-alanine function as an activator in the regulation of the Escherichia coli glutamate racemase activity. Biochem. 34: 2464–2470.
Huffer S, ME Clark, JC Ning, HW Blanch, dan DS Clark. 2011. Role of alcohols in growth, lipid composition, and membrane fluidity of yeast, bacteria, and archaea. Appl Environ Microbiol. 77: 6400–6408.
Ingram LO dan NS Vreeland. 1980. Differential effects of ethanol and hexanol on the Escherichia coli cell envelope. J Bacteriol. 144: 481–488.
Ingram LO. 1986. Microbial tolerance to alcohols: role of the cell membrane. Trends Biotechnol. 4: 40–44.
Ingram LO, T Conway, DP Clark, GW Sewell, dan JF Preston. 1987. Genetic engineering of ethanol production in Escherichia coli. Appl Environ Microbiol. 53: 2420–2425.
Kim KH, YJ Bong, JK Park, KJ Shin, KY Hwang, dan EE Kim. 2007. Structural basis for glutamate racemase inhibition. J Mol Biol. 372: 434–443.
Luli GW dan WR Strohl. 1990. Comparison of growth, acetate production, and acetate inhibition of Escherichia coli strains in batch and fed-batch fermentations. Appl Environ Microbiol. 56: 1004–1011.
Lundqvist T, SL Fisher, G Kern, RHA Folmer, Y Xue, DT Newton, TA Keating, RA Alm, dan BLM de Jonge. 2007. Exploitation of structural and regulatory diversity in glutamate racemace. Nature. 447: 817–822.
Lustig KD, K Kroll, E Sun, R Ramos, H Elmendorf, dan MW Kirschner. 1996. A Xenopus nodal-related gene that acts in synergy with noggin to induce complete secondary axis and notochord formation. Development. 122: 3257–3282.
Panja S, S Saha, dan T Basu. 2006. Role of membrane potential on artificial transformation of E. coli with plasmid DNA. J Biotechnol. 127: 14–20. Skotnicki ML, KJ Lee, DE Tribe, dan PL Rogers. 1981. Comparison of ethanol
production by different Zymomonas mobilis. Appl Environ Microbiol. 41: 889–893.
Strom AR. 1998. Osmoregulation in the model organism Escherichia coli: genes governing the synthesis of glycine betaine and trhalose and their use in metabolic engineering of stress tolerance. J Biosci. 23: 437–445.
Stimson H. 2011. The essential chromatography and spectroscopy catalog 2011 2012 edition. Canada: Agilent Technologies Inc.
Suryadarma P, Y Ojima, K Tsuchida, dan M Taya. 2012. Design of Escherichia coli cell culture for regulating alanine production under aerobic conditions. 2012. J Chem Eng Jpn. 45: 604–608.
Underwood SA, ML Buszko, KT Shanmugam, dan LO Ingram. 2002. Flux through citrate synthase limits the growth of ethanologenic Escherichia coli KO11 during xylose fermentation. Appl Environ Microbiol. 68: 1071–1081. Underwood SA, S Zhou, TB Causey, LP Yomano, KT Shanmugam, dan LO
Ingram. 2002. Genetic change to optimize carbon partitioning between ethanol and biosynthesis in ethanologenic Escherichia coli. Appl Environ Microbiol. 68: 6263–6272.
Vemuri GN, E Altman, DP Sangurdekar, AB Khodursky, dan MA Eiteman. 2006. Overflow metabolism in Escherichia coli during steady-state growth: Transcriptional regulation and effect of the redox ratio. Appl Environ Microbiol. 72: 3653–3661.
Lampiran 1 Prosedur transformasi metode kejut panas(Panja et al. 2006) Pembuatan sel kompeten
Galur E. coliBW25113Δpta dibiakan terlebih dahulu dalam 2 ml media LB yang mengandung 15 ppm kanamisin kemudian diinkubasi pada suhu 37 °C dengan kecepatan 200–250 rpm selama semalam. Sebanyak 0.5 ml biakan sel E. coli dibiakan hingga OD600 mencapai 0.4–0.5. Kemudian 1.5 ml biakan sel E. coli diinkubasi dalam dalam es selama 10 menit. Selanjutnya disentrifugasi dengan kecepatan 3000 rpm pada suhu 4 °C selama 10 menit. Supernatan yang dihasilkan kemudian dibuang sedangkan pelet diresuspensi dengan 495 µ l CaCl2. Selanjutnya diinkubasi dalam es selama 10 menit kemudian disentrifugasi dengan kecepatan 3000 rpm pada suhu 4 °C selama 10 menit. Pelet yang telah dipisahkan dari supernatan kemudian diresuspensi dengan 125 µ l CaCl2 lalu ditambahkan DMSO dan diinkubasi dalam es selama 10 menit. Sel kompeten yang dihasilkan kemudian digunakan untuk transformasi. Setiap transformasi membutuhkan 100 µ l sel kompeten.
Transformasi metode kejut panas
Sebanyak 10 µ l plasmid pHfdh ditambahkan ke dalam 100 µ l sel kompeten. Kemudian diinkubasi dalam es selama 30 menit lalu dipanaskan pada suhu 42 °C selama 45 detik. Kemudian diinkubasi dalam es selama 5 menit. Setelah diinkubasi, sel kompeten tersebut ditambahkan dengan 100 µ l media 2xYT lalu diinkubasi pada suhu 37 °C dengan kecepatan 250 rpm selama 1 jam.
Sel E. coli yang telah disisipi plasmid pHfdh kemudian disebar ke dalam media LB agar/padat yang mengandung 34 ppm kloramfenikol dan diinkubasi pada suhu 37 °C selama semalam. Selanjutnya akan dilakukan analisis plasmid dengan menggunakan GeneJETTM miniprep kit. Untuk menyisipkan plasmid pTadhB-pdc, prosedurnya sama dengan menyisipkan pHfdh tetapi menggunakan media seleksi (LB) yang mengandung 50 ppm ampisilin.
Lampiran 2 Prosedur pembuatan persediaan biakan (Lustig et al. 1996)
Sebanyak 200 µ l gliserol 50% dan 200 µ l sel E. coli hasil prapembiakan dimasukkan ke dalam eppendorf. Kemudian diresuspensi agar homogen. Selanjutnya simpan dalam lemari pendingin.
Lampiran 3 Prosedur prapembiakan (Ojima et al. 2012)
Sebanyak 50 ml LB dimasukkan ke dalam erlenmeyer kemudian ditambahkan ampisilin, kloramfenikol, kanamisin, dan galur E. coli
BW25113Δpta/pHfdh/ pTadhB-pdc masing-masing 50 µ l. Selanjutnya, diinkubasi di dalam kultur bergoyang dengan kecepatan 120 rpm pada suhu 37 °C selama 12 jam.
media LB cair. Namun, jika nilai OD660 di bawah 1 maka proses inkubasi dilanjukan kembali hinggan nilainya 1–1.5.
Lampiran 4 Prosedur pembiakan (Ojima et al. 2012)
Sebanyak 2.5 ml galur E. coli yang telah mencapai nilai OD660 1–1.5 dimasukkan ke dalam baffled conical flask yang mengandung 40 ml media LB, l0 ml glukosa 40 g/l, 50 µ l IPTG 0.5 mM, 50 µ l ampisilin 50 mg/l, 50 µ l kloramfenicol 34 mg/l, 50 µ lkanamisin15 mg/l, 1 ml format 4 g/l, dan 1 g CaCO3 20 g/l. Nilai pH media diatur hingga 7 dengan penambahan NaOH 1 M. Selanjutnya, diinkubasi di dalam kultur bergoyang dengan kecepatan 250 rpm pada suhu 37 °C selama 24 jam.
Pada saat jam ke-6, dilakukan penambahan 1.266 ml etanol 20 g/l, dan glutamat dengan berbagai konsentrasi yaitu 2, 4, dan 6 g/l. Kemudian proses inkubasi dilanjutkan hingga jam ke-24. Setiap perlakuan dilakukan perulangan sebanyak 3 kali.
Format, etanol, dan glutamat yang digunakan harus disterilisasi terlebih dahulu dengan menggunakan syringe filter 0.2 µ m. Sedangkan untuk CaCO3 disterilisasi dengan dioven pada suhu 180 °C selama 2 jam.
Lampiran 5 Prosedur analisis bobot sel kering (Ojima et al. 2012)
Sebanyak 4.5 ml HCl 1 M dimasukkan ke dalam kuvet kemudian dicampur dengan 0.5 ml sampel dan diresuspensi. Kemudian diukur nilai OD pada absorbansi (λ) 660 nm dengan spektrofotometer Hach DR 2500.
Bobot sel kering (g/l) = 0.36 x OD660
Lampiran 6 Prosedur analisis glukosa (F-Kit 716251) (Suryadarma et al. 2012) Sebelum dianalisis, sampel disentrifugasi terlebih dahulu dengan kecepatan 10 000 rpm pada suhu 4 °C selama 2 menit. Setelah disentrifugasi, sampel disaring dengan syringe filter 0,2 µm. Kemudian sampel diencerkan berseri yaitu 1:10, 1:50, 1:100, 1:200, dan 1:400.
Pertama ukur nilai absorbansi (A) blangko. Sebanyak 0.5 ml larutan 1 dicampur dengan 1.05 ml aqua bidestilata kemudian diresuspensi. Selanjutmya tunggu sekitar 3 menit lalu diukur nilai A1 blangko. Kemudian tambahkan 0.01 ml suspensi 2 dan resuspensi hingga homogen. Kemudian tunggu sekitar 10–15 menit lalu ukur nilai A2 blangko.
ΔA (g/l) = (A2 - A1)sampel - (A2 - A1)blangko
c = x ΔA (g/l) V = volume akhir (ml)
v = volume sampel (ml) MW = Bobot molekul (g/mol) d = light path (cm)
ε = extinction coefficient NADPH pada: 340 nm = 6.3 (l x mmol-1 x cm-1) Hg 365 nm = 3.5 (l x mmol-1 x cm-1) Hg 334 nm = 6.18 (l x mmol-1 x cm-1)
Lampiran 7 Prosedur analisis asetat (Stimson 2011)
Pengukuran asetat dilakukan dengan menggunakan HPLC dengan kolom ZORBAX SB-Aq 883975-914. Fase bergerak yang digunakan adalah 99% NaH2PO4 20 mM, pH2, 1% ACN. Suhu, laju alir, dan panjang gelombang yang digunakan, berturut-turut yakni 35 ºC, 1 ml/menit, dan 210 nm. Volume sampel yang diinject sebanyak 20 µ l.
Lampiran 8 Hasil analisis asetat dengan HPLC
Keterangan : (A) Sampel dengan perlakuan penambahan glutamat. (B) Sampel dengan perlakuan tanpa penambahan glutamat
V x MW ε x d x v x 1000
A
B
RIWAYAT HIDUP
Penulis dilahirkan di Padang pada tanggal 21 Agustus 1991 dari ayah Yusman dan ibu Marnis. Penulis merupakan anak ketiga dari tiga bersaudara. Tahun 2010 penulis lulus dari SMA Negeri 2 Jember dan pada tahun yang sama penulis lulus seleksi masuk Institut Pertanian Bogor (IPB) melalui jalur Seleksi Nasional Masuk Perguruan Tinggi Negeri (SNMPTN) dan diterima di Departemen Teknologi Industri Pertanian, Fakultas Teknologi Pertanian.
Selama menjadi mahasiswa, penulis aktif di berbagai organisasi. Pada tahun 2010–2011 penulis menjabat sebagai ketua Kominfo CIA (Club Ilmiah Asrama). Kemudian di tingkat dua (2011–2012) menjadi anggota FORCES (Forum Scientific of Students) IPB dan anggota departemen syiar LDF (Lembaga Dakwah Fakultas) Forum Bina Islami (FBI) Fateta. Selanjutnya pada tahun 2012–2013, penulis mendapatkan amanah sebagai ketua LDF (Lembaga Dakwah Fakultas) Forum Bina Islami (FBI) Fateta.
Semasa mengikuti perkuliahan, penulis pernah memperoleh berbagai prestasi, di antaranya PKM Penelitian 2012 berhasil didanai DIKTI (Direktorat Jenderal Pendidikan Tinggi), juara 3 LKTI Nasional BEM FMIPA ITS 2012, dan juara 3 LKTA JMMI ITS 2012.