HUBUNGAN FILOGENETIK Hylarana mocquardii (ANURA : RANIDAE)
DI SULAWESI BERDASARKAN PENGUKURAN MORFOLOGI
DAN MOLEKULER GEN 12S rRNA DAN 16S rRNA
SISTER SIANTURI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN
SUMBER INFORMASI SERTA PELIMPAHAN HAK CIPTA
Dengan ini saya menyatakan bahwa tesis berjudul Hubungan Filogenetik
Hylarana mocquardii (Anura : Ranidae) di Sulawesi Berdasarkan Pengukuran Morfologi dan Molekuler Gen 12S rRNA dan 16S rRNA adalah benar karya saya dengan arahan dari komisi pembimbing dan belum diajukan dalam bentuk apa pun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini.
Dengan ini saya melimpahkan hak cipta dari karya tulis saya kepada Institut Pertanian Bogor.
Bogor, Agustus 2015
Sister Sianturi
RINGKASAN
SISTER SIANTURI. Hubungan Filogenetik Hylarana mocquardii (Anura : Ranidae) di Sulawesi Berdasarkan Pengukuran Morfologi dan Molekuler Gen 12S rRNA dan 16S rRNA. Dibimbing oleh ACHMAD FARAJALLAH dan MIRZA DIKARI KUSRINI.
Hylarana mocquardii merupakan spesies yang awalnya merupakan anggota spesies H.chalconota yang masih termasuk kompleks spesies karena terdapat banyak variasi dalam spesies ini. H.mocquardii pada akhirnya ditetapkan sebagai spesies yang hanya terdapat di Pulau Sulawesi. Pulau Sulawesi merupakan salah satu pulau di Indonesia yang memiliki sejarah geologi yang kompleks. Sejarah geologi yang kompleks dengan kombinasi reorganisasi habitat, aktivitas pegunungan yang aktif, dan adanya fluktuasi permukaan laut menjadi faktor pendukung terbentuknya fragmentasi habitat, migrasi, kolonisasi spesies, hingga terjadinya spesiasi.
Konstruksi filogeni dihasilkan dengan membandingkan sekuen
H.mocquardii dari kawasan Sulawesi yang mewakili semua bagian Sulawesi, yaitu Sulawesi Utara, Barat, Tengah, Tenggara, dan Selatan. Topologi filogeni juga terdiri atas sekuen H.chalconota, H.megalonesa, H.erythraea, H.signata,
H.grandocula, H.similis, Sanguirana luzonensis, Limnonectes cf.kuhlii, dan Occydozyga baluensis yang diunduh dari genebank. Konstruksi filogeni dengan gen 12S rRNA dan 16S rRNA menghasilkan pola pengelompokan spesies
H.mocquardii yang monofiletik dan membentuk empat kelompok, yaitu kelompok spesies di kawasan Sulawesi Utara, kelompok spesies di kawasan Sulawesi Tengah dan Sulawesi Barat, kelompok spesies di kawasan Sulawesi Tenggara, dan kelompok spesies di kawasan Sulawesi Selatan. Berdasarkan gen 12S rRNA diperoleh pengelompokan spesies H.mocquardii dengan dua subclade yaitu kelompok yang berasal dari kawasan Sulawesi Tengah dan Barat dan kelompok yang berasal dari kawasan Sulawesi Utara, Sulawesi Tenggara, dan Sulawesi Selatan. Berdasarkan gen 16S rRNA diperoleh pengelompokan spesies
H.mocquardii juga menghasilkan dua subclade yaitu dengan kelompok spesies yang berasal dari kawasan Sulawesi Selatan memiliki pengelompokan sendiri.
Hasil analisis molekuler ini kemudian dibandingkan dengan hasil analisis pengukuran morfologi. Dalam analisis morfometri ini dipisahkan antara analisis individu betina dengan jantan. Penelitian ini melakukan perbandingan antara hasil analisis morfometri dengan analisis molekuler. Pada hasil analisis morfometri pada individu betina setelah dilakukan uji lanjut tidak terdapat perbedaan dan pengelompokan. Pada hasil analisis morfometri individu jantan juga pada awalnya tidak terlihat adanya pengelompokan, tetapi setelah dilakukan uji lanjut Duncan terlihat adanya pengelompokan yang juga mendukung pada hasil analisis molekuler gen 16S rRNA yang menunjukkan bahwa kelompok spesies
H.mocquardii di Sulawesi Selatan memiliki pengelompokan tersendiri. Berdasarkan penelitian ini juga diketahui bahwa proses invasi spesies
H.mocquardii ke Pulau Sulawesi terjadi seiring dengan proses terbentuknya Pulau Sulawesi dan nenek moyang spesies ini berasal dari daratan Asia.
SUMMARY
SISTER SIANTURI. Phylogenetic Relationships of Hylarana mocquardii in Sulawesi based on Morphometrics and Molecular 12S rRNA with 16S rRNA gene. Supervised by ACHMAD FARAJALLAH and MIRZA DIKARI KUSRINI.
Hylarana mocquardii was originally member of species Hylarana chalconota complex because there are many variation in this species. The H. mocquardii was then defined as species only found on Sulawesi island. Sulawesi is an island in Indonesia with a complex geological history. Complex geological history with a combination of reorganization habitat, the activity of the active mountains, and the fluctuation of sea level are factors that support the formation of habitat fragmentation, migraton,colonization of that species, and resulted in the occurence of speciation.
A phylogeny construction was made by comparing the sequence of
H.mocquardii from all parts of Sulawesi: North Sulawesi, West Sulawesi, Central Sulawesi, East Sulawesi, and South Sulawesi and added sequence of H. chalconota, H. megalonesa, H. erythraea, H. signata, H. grandocula, H. similis,
Sanguirana luzonensis, Limnonectes cf. kuhlii, and Occydozyga baluensis from genebank. The phylogeny construction of the 12S rRNA and 16S rRNA produce monophyletic grouping patterns of H.mocquardii and consist of four groups: North Sulawesi region, Central Sulawes and West Sulawesi region, Southeast Sulawesi region, and South Sulawesi region. Based on12S rRNA gene this group was divided in two subclade, the first group consist of species from West Sulawesi and Central Sulawesi region and the second group consist of species from North Sulawesi, Southeast Sulawesi, and South Sulawesi region. Based on 16S rRNA gene the grouping was divided in two subclade that separated species from South Sulawesi region from other region.
Molecular analysis were then compared with morphometrics analysis of females and males frog. Morphometric analysis results were unable to show significant difference grouping of H.mocquardi because it produce only one grouping. However, final morphometric analysis of males using Duncan Test showed that H.mocquardii from South Sulawesi differed from other region. It conclude that morphometric analysis of male frog showed the same pattern and supported molecular analysis of 16S rRNA gene. This research was able to analysis the spread of H.mocquardii to Sulawesi island. This process occured during the formation of Sulawesi island and predicted occured during Miocene. Based on this I concluded that ancestor of this species originated from Mainland Asia.
© Hak Cipta Milik IPB, Tahun 2015
Hak Cipta Dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan atau menyebutkan sumbernya. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan IPB
Tesis
sebagai salah satu syarat untuk memperoleh gelar Magister Sains
pada
Program Studi Biosains Hewan
HUBUNGAN FILOGENETIK Hylarana mocquardii (ANURA : RANIDAE)
DI SULAWESI BERDASARKAN PENGUKURAN MORFOLOGI DAN
MOLEKULER GEN 12S rRNA dan 16S rRNA
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
BOGOR 2015
Puji dan Syukur penulis panjatkan kepada Tuhan Yang Maha Esa atas segala berkat dan karuniaNya sehingga penulisan tesis ini dapat diselesaikan. Tesis ini disusun dalam rangka memenuhi persyaratan memperoleh gelar Magister Sains pada Program Studi Biosains Hewan (BSH) Sekolah Pascasarjana Institut Pertanian Bogor.
Pada kesempatan ini penulis mengucapkan terima kasih kepada:
1. Dr. Ir. Achmad Farajallah, M.Si. dan Dr. Ir. Mirza Dikari Kusrini, M.Si. selaku komisi pembimbing yang telah memberikan arahan dan masukan dalam penyusunan karya ini
2. Kepada Dr.Ir.R. R. Dyah Perwitasari, M.Sc.selaku ketua program studi Biosains Hewan.
3. Museum Zoologicum Bogorienses (MZB) yang telah memberikan akses terh adap penggunaan spesimen yang digunakan dalam penelitian.
4. Dr.Amir Hamidy M.Sc selaku kepala Laboratorium Herpetologi Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong, Dr. rer. nat. Evy Arida, M.Sc. dan seluruh staff Laboratorium Herpetologi LIPI Cibinong
5. Dirjen Pendidikan Tinggi (DIKTI) Indonesia yang telah memberikan dana melalui beasiswa Pendidikan Pasca Sarjana Dalam Negeri (BPPDN) 2013 6. Rekan-rekan BSH 2013 yang telah memberikan bantuan, dukungan dan
motivasi selama perkuliahan dan penelitian berlangsung
7. Bapak Adi Surahman dan Ibu Tini Wahyuni yang banyak membantu dalam penyediaan alat dan bahan laboratorium
8. Kedua orang tua, saudara-saudara, dan teman-teman yang telah memberi dukungan dan doa kepada penulis
Bogor, Agustus 2015
DAFTAR ISI
Waktu dan Tempat Penelitian 4 Sampel Penelitian 5Analisis Pengukuran Morfologi 10 Analisis Molekuler 14
Kongruensi Analisis Morfometri dengan Analisis Molekuler Gen 16S rRNA 15
DAFTAR TABEL
DAFTAR GAMBAR
1 Spesimen H.mocquardii tampak dorsal dan tampak ventral 1 2 Karakter penting untuk identifikasi H.mocquardii 2 3 Kondisi Geologi dan Tektonik Pulau Sulawesi pada
masing-masing pembagian kawasan
3 4 Titik pengukuran morfologi pada individu H.mocquardii 7 5 Karakter Nuptial pad pada H.mocquardii individu jantan 7 6 Hasil analisis biplot pengelompokan spesies H.mocquardii
betina
12 7 Hasil analisis biplot pengelompokan spesies H.mocquardii
jantan
13
8 Proses sejarah tektonik Pulau Sulawesi 16
9 Pembagian kawasan daratan yang terdiri dari kawasan daratan Sunda, Wallacea, dan kawasan daratan Sahul
17 10 Konstruksi pohon filogeni dengan metode Neighbour-joining 18 11 Pola filogeografi H.mocquardii di Sulawesi berdasarkan gen
12S rRNA dan 16S rRNA
19
DAFTAR LAMPIRAN
1 Analisis kontribusi masing-masing variabel terhadap komponen pada spesies H.mocquardii betina
26 2 Analisis kontribusi masing-masing variabel terhadap komponen
pada spesies H.mocquardii betina
27 3 Data spesimen H.mocquardii yang digunakan untuk analisis
pengukuran morfologi
28 1 Data Spesimen H.mocquardii yang digunakan dalam penelitian
untuk analisis molekuler
6 2 Daftar Spesies Hylarana dan akses Genebank yang digunakan
dalam analisis molekuler dan konstruksi filogeni
9 3 Hasil Pengukuran morfologi spesies H.mocquardii betina (n=79) 10 4 Hasil Pengukuran morfologi spesies H.mocquardii jantan (n=189) 11 5 Analisis komponen utama pengukuran morfologi spesies
H.mocquardii betina
12 6 Analisis komponen utama pengukuran morfologi spesies
H.mocquardii jantan 9 Jumlah perbedaan nukleotida (bawah diagonal) dan jarak genetik
(atas diagonal) berdasarkan gen 12S rRNA
20 10 Jumlah perbedaan nukleotida (bawah diagonal) dan jarak genetik
(atas diagonal) berdasarkan gen 16S rRNA
1 PENDAHULUAN
Latar Belakang
Penelitian terkait sistematika pada famili Ranidae masih perlu dilakukan karena taksonomi pada taksa ini masih diperdebatkan. Pada anggota famili Ranidae masih banyak spesies yang memiliki hubungan filogenetik yang belum dapat dipecahkan. Penyebabnya adalah karena penyebaran spesies yang sangat luas dan revisi sistematika terkait dengan sinonim antar lokasi geografis yang masih terjadi pada anggota spesies famili Ranidae (Dubois 1992).
Hylarana mocquardii (Werner, 1901) merupakan salah satu anggota famili Ranidae yang awalnya memiliki nama spesies Hylarana chalconota
(Schlegel, 1837). Spesies H. chalconota memiliki penyebaran yang luas di Indonesia, yaitu Jawa, Bali, Sumatera, Kalimantan, dan Sulawesi (Inger 1966). H. chalconota yang berasal dari Sulawesi ditempatkan sebagai spesies yang sah setelah ditemukan ciri morfologi pada bagian dorsal yang lebih mencolok dibandingkan dengan H. chalconota yang berasal dari kawasan di luar Sulawesi (Iskandar & Colijin 2000). Hal ini diperkuat dengan pernyataan Inger & Voris (2001) yang menyatakan bahwa fauna Sulawesi, khususnya katak keturunan
Sundaland, menunjukkan hampir tidak ada kesamaan dengan fauna lain pada tingkat spesies. Hasil penelitian yang dilakukan oleh Inger et al. (2009) dengan menggunakan gen 16S rRNA dan beberapa karakter morfologi sebagai marker menemukan bahwa spesies H. chalconota dipecah menjadi beberapa kelompok. Kelompok H. chalconota yang hidup di Kalimantan terbagi menjadi dua jenis, yaitu H. megalonesa dan H. raniceps, kelompok yang hidup di Sumatera dibagi menjadi tiga jenis, yaitu H. chalconota, H. parvaccola, dan H. rufipes, kelompok yang hidup di Jawa tetap sebagai jenis H. chalconota, dan kelompok yang hidup di Sulawesi ditetapkan sebagai H. mocquardii. Inger et al. (2009) kemudian menghapus H. mocquardii sebagai sinonim dari H. chalconota.
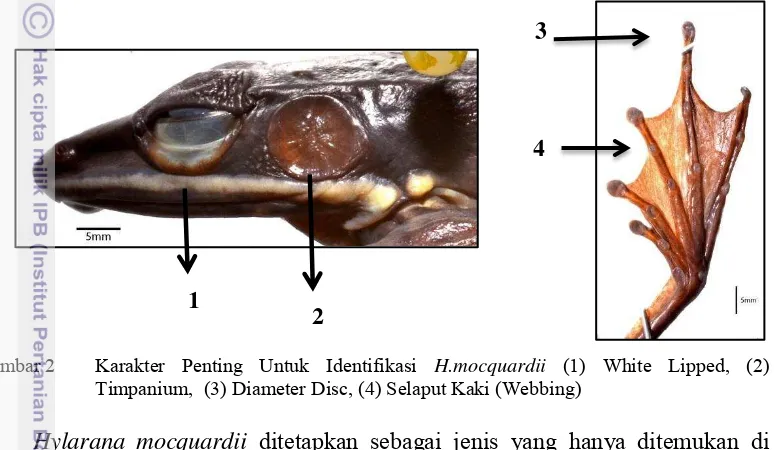
Spesies H. mocquardii pada jantan memiliki kisaran ukuran 15.87 mm - 55.73 mm sedangkan pada betina 35.74 mm - 83.87 mm, memiliki permukaan dorsal yang berwarna kuning hingga coklat, sedangkan permukaan ventral memiliki warna putih dengan bintik hitam, memiliki kaki panjang dan ramping (Gambar 1) (Iskandar 1998). Karakter penting dalam identifikasi spesies ini adalah memiliki bibir putih (white lipped) yang berada di sepanjang bawah mata hingga timpanium, memiliki timpanium berwarna coklat tua, dan memiliki selaput kaki (webbing) penuh (Gambar 2) (Iskandar 1998).
Hylarana mocquardii ditetapkan sebagai jenis yang hanya ditemukan di Pulau Sulawesi (Frost 2009). Inger et al. (2009) membuktikan bahwa
H.mocquardii merupakan spesies kriptik dalam grup H.chalconota yang ada di Sulawesi.
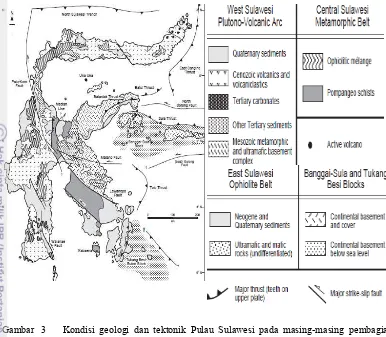
Pulau Sulawesi merupakan salah satu pulau di Indonesia yang memiliki sejarah geologi yang kompleks (Stelbrink et al. 2011). Sejarah geologi dan topologi yang kompleks (Gambar 3) dengan kombinasi reorganisasi habitat terestrial, aktivitas pegunungan yang aktif, dan adanya fluktuasi permukaan laut menjadi faktor pendukung terbentuknya fragmentasi habitat, migrasi, kolonisasi spesies pada habitat baru, hingga terbentuknya spesiasi allopatrik (Esselstyn et al. 2009). Pulau Sulawesi terbentuk dari beberapa proses tabrakan beberapa lempeng yang berbeda (Hall 1996). Keadaan ini merupakan barier yang potensial untuk menghasilkan pola distribusi hewan yang memiliki pola sesuai dengan fragmentasi dan diversifikasi habitat (Evans et al. 2003). Selain itu adanya topografi yang kompleks dan keragaman habitat iklim mikro memberi pengaruh dalam diversifikasi, spesiasi, dan variasi spesies (Bridle et al. 2001). Studi molekuler pada biota Sulawesi menemukan bukti bahwa sejarah vikarian dan seleksi ekologi adalah sebagai faktor utama yang mendorong terjadinya diversifikasi (Evans et al. 2001).
Gambar 2 Karakter Penting Untuk Identifikasi H.mocquardii (1) White Lipped, (2) Timpanium, (3) Diameter Disc, (4) Selaput Kaki (Webbing)
2
1
2
3
Gambar 3 Kondisi geologi dan tektonik Pulau Sulawesi pada masing-masing pembagian kawasan (Moss & Wilson 1998)
Morfometri merupakan salah satu cara untuk mengetahui keanekaragaman dari suatu spesies dengan menguji karakter morfologi. Data morfometri dapat digunakan untuk menjelaskan perbedaan dan persamaan antar populasi dan mendeskripsikan kekerabatan populasi secara morfologi (Hill & Wiens 2000). Sebagai aplikasi lanjut, hasil analisis secara morfometri dapat memberikan gambaran umum tentang tingkat variabilitas dari suatu taksa (Chernoff 1982). Setiap karakter yang diamati umumnya merupakan akibat adanya interaksi gen-gen yang ekspresinya dipengaruhi oleh lingkungan (Munshi & Dutta 1996). Keterbatasan dari penggunaan data morfometri adalah nilai konsistensi rendah dalam menunjukkan hubungan filogenetik pada tingkat variasi spesies kriptik dan subspesies sehingga perlu dikomparasi dengan data molekuler (Wiens 2004). Studi dengan menggunakan kedua pendekatan ini dapat memberi informasi yang lebih luas dan sangat membantu untuk informasi sistematika dan rekonstruksi filogeni (Hillis 1987).
gen yang digunakan sebagai marker adalah gen 12S rRNA. Hedges and Maxson (1993) yang melakukan penelitian filogeni pada cecilia membuktikan bahwa gen 12S rRNA dan 16S rRNA dalam genom mitondria merupakan gen yang sangat potensial dalam aspek filogeni amfibi.
Perumusan Masalah
Spesies H. mocquardii pada awalnya memiliki nama H. chalconota, merupakan kompleks spesies. Inger et al. (2009) menemukan bahwa H. mocquardii memiliki perbedaan dengan spesies lain dalam chalconota grup baik secara morfologi maupun secara molekuler. H. mocquardii yang ada di Sulawesi akhirnya ditetapkan sebagai spesies yang terpisah. Penegakan sebagai spesies yang terpisah dari chalconota group pada H. mocquardii di kawasan Sulawesi dipengaruhi oleh kondisi geologis, habitat, dan batasan-batasan ekologis. Penelitian ini ingin membuktikan bagaimana kontribusi dari batasan tersebut terhadap kekerabatan spesies H. mocquardii di Sulawesi dan bagaimana pola pengelompokan spesies berdasarkan sekuens gen 12S rRNA dan 16S rRNA dalam genom mitokondria. Selain itu, penelitian ini juga menggunakan pengukuran morfologi untuk memperkuat hasil pengelompokan tersebut.
Tujuan Penelitian
Penelitian ini bertujuan untuk:
1. Untuk menganalisis kekerabatan intraspesies H. mocquardii
menggunakan gen 12S rRNA dan 16S rRNA genom mitokondria
2. Untuk menganalisis kekerabatan interspesies H. mocquardii
berdasarkan pengukuran morfologi
Manfaat Penelitian
Penelitian ini diharapkan bermanfaat sebagai acuan untuk manajemen konservasi spesies endemik di Sulawesi khususnya spesies H. mocquardii
2
METODE PENELITIAN
Waktu dan Tempat Penelitian
Penelitian dilaksanakan pada bulan Agustus 2014 – Maret 2015. Pengukuran morfologi dan pengambilan sampel jaringan untuk analisis molekuler dilakukan di Laboratorium Herpetologi, Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong, sedangkan untuk proses analisis molekuler meliputi proses
Sampel Penelitian
Sampel katak H. mocquardii berasal dari kawasan Sulawesi yang diperoleh dari hasil preservasi yang disimpan di Laboratorium Herpetologi LIPI Cibinong. Analisis morfometri menggunakan 79 individu spesimen betina dan 189 individu spesimen jantan pada semua sampel H. mocquardii yang ada di Museum Zoologicum Bogoriense (MZB) (Lampiran 3). Analisis molekuler menggunakan 12 spesimen yang diambil dari organ hati (liver) yang berasal dari masing-masing kawasan Sulawesi yang dapat mewakili masing-masing bagian pulau yaitu Sulawesi Utara, Sulawesi Barat, Sulawesi Tengah, Sulawesi Tenggara, dan Sulawesi Selatan (Tabel 1).
Prosedur Penelitian
Pengukuran Morfologi
Pengukuran menggunakan Calliper digital 0.1 mm. Titik-titik pengukuran sesuai dengan Matsui (1984) dan Matsui (1994) ( Gambar 4). Selain itu, pada katak jantan dilakukan juga pengukuran pada karakter tambahan, yaitu Nuptial Pad (NP), diameter disc pada kaki depan jari ketiga (DFTD3) dan jari keempat (DFTD4), dan diameter disc kaki belakang pada jari ketiga (DFTB3) dan jari keempat (DFTB4) (Gambar 5).
Analisis Pengukuran Morfologi
Tabel 1 Data Spesimen H. mocquardii yang digunakan dalam penelitian untuk MZB13989 BSI 1272 Desa Balapu, Kec.Kulawi,
Kab.Donggala, Sulawesi Tengah
KR082127 12S rRNA
MZB11450 AK203 Lantai Hutan Salodik, Kab .Banggai,Sulawesi Tengah MZB12400 JAM 5942 Desa Polewali, Massawa R
oad River 1, Sulawesi MZB17486 ATH 2082 Pegunungan Mekongga, D
esa Tinukari, Kec.Wawo, Kolaka,Sulawesi Tenggara
KR082130 KR153319
12S rRNA 16S rRNA MZB17484 ATH 1760 Pegunungan Mekongga,
Desa Tinukari, Kec.Wawo,
MZB23288 KBAA12 Kaki Gunung Boliyuhuto, Marga Satwa Nantu, MZB23289 KBAA14 Kaki Gunung Boliyuhuto,
Marga Satwa Nantu,
Gambar 4 Titik Pengukuran Morfologi pada Individu H. mocquardiii (Matsui 1984) (1): Snout Vent Length (SVL), (2) Head Length (HL), (3) Snout-Nostril Length (SNL), (4) Nostril-Eyelid Length (NEL), (5) Snout-Length (SL), (6) Eye Length (EL), (7) Tympanium-Eye Length (TEL), (8) Tympanium Diameter (TD), (9) Head Width (HW), (10) Internarial Distance (IND), (11)Intercanthal Distance (ICD), (12) Interorbital Distance (IOD), (13)Upper Eyelid Width (UEW), (14) Upper Eyelid Margin Distance (UEMD), (18) Forelimb Length (FLL), (19) Lower Arm Length (LAL), (20) Third Finger Length (TFL), (21) First Finger Length (FFL), (22) Outer Palmar Tubercle Length (OPTL), (23) Inner Palmar Tubercle Length (IPTL), (24) Hand Length (HAL), (25) ForearmWidth (FAW), (26) Hindlimb Length (HLL), (27) Tibia Length (TL), (28) Foot Length (FL), (29) Fourth Toe Length (FTL), (30) Outer Metatarsal Tubercle Length (OMTL), (31) Inner Metatarsal Tubercle Length (IMTL)
Gambar 5. Karakter Nuptial pad pada H.mocquardii individu jantan
Analisis Molekuler
Ekstraksi DNA
Isolasi DNA total dilakukan dengan mengikuti metode Farajallah (2002). Potongan otot katak sekitar 2 mm dalam alkohol 70 % dicuci dengan 500 µl buffer TE sebanyak 2 kali. Sampel otot yang telah dicuci kemudian dipotong kecil-kecil menggunakan gunting dalam bufer 1x STE. Sel-sel otot dilisis dengan menambahkan SDS sampai 10 % dan enzim Proteinase-K sebanyak 0.3 mg/ml. Campuran tersebut kemudian diinkubasi pada suhu 55oC selama 1 jam sambil dikocok perlahan.
Material DNA dipisahkan dari material organik lainnya dengan metode ekstraksi fenol, yaitu dengan menambahkan larutan NaCl 5 M sebanyak 1/10 volume, fenol sebanyak 1x volume, dan Kloroform-isoamil alkohol (CIAA:24:1) sebanyak 1x volume. Campuran tersebut kemudian diinkubasi pada suhu ruang sambil dikocok pelan selama 1 jam. Bahan organik yang masuk ke fase fenol dipisahkan dari fase air dengan sentrifugasi pada kecepatan 5000 rpm selama 5 menit. Fase air kemudian dipindahkan ke dalam tabung baru. Molekul DNA dimurnikan dengan teknik pengendapan alkohol yaitu dengan menambahkan alkohol absolut sebanyak 2x volume dan NaCl 5 M sebanyak 1/10 x volume. Campuran diinkubasi pada suhu 4o C selama 24 jam. Molekul DNA diendapkan dengan sentrifugasi pada kecepatan 5000 rpm selama 5 menit. Endapan DNA yang diperoleh kemudian dicuci dengan alkohol 70 %. Molekul DNA diendapkan kembali kemudian disuspensikan dalam bufer TE 80 % dan disimpan dalam
freezer untuk digunakan lebih lanjut.
Amplifikasi (Perbanyakan) Ruas DNA
Ruas DNA diamplifikasi dengan metode Polymerase Chain Reaction (PCR) menggunakan mesin Thermo Cycler TaKaRa MP4. Primer yang digunakan adalah AF559 (5’ACTGGGATTAGATACCCCACTAT3’) yang menempel pada bagian tengah gen 16S rRNA dan AF560 (5’ATGTTTTTGGTAAACAGGCG-3’) yang menempel pada akhir gen 12S rRNA. Total pereaksi PCR adalah 25µl yang terdiri atas 0.625 mg/ml Gotag Green Master Mix, 0.0026 mg/ml Primer Forward, 0.0028 mg/ml Primer Reverse, 10.5 µl Nuclease Water, dan 0.8 ƞg/ml Template DNA.
Amplifikasi dilakukan dengan kondisi predenaturasi 94o C selama 4 menit, dilanjutkan dengan 30 siklus yang terdiri dari denaturasi 94o C selama 1 menit,
annealing (penempelan) 55o C selama 1 menit 30 detik, pemanjangan 72o C selama 1 menit, dan pemanjangan akhir 72o C selama 7 menit.
Visualisasi Perbanyakan Ruas DNA
Perunutan (Sequencing) DNA Produk PCR
Perunutan DNA dilakukan dengan menggunakan jasa pelayanan sekuensing. Perunutan tersebut dilakukan dengan menggunakan pasangan primer yang digunakan pada saat PCR.
Analisis Data
Urutan nukleotida yang diperoleh dari proses perunutan diedit secara manual dibantu dengan program Bioedit. Runutan nukleotida kemudian disejajarkan bersama spesies lain yang masih dalam satu famili yang diunduh dari
genebank (www.ncbi.nlm.nih.gov) (Tabel 2). Analisis jumlah perbedaan nukleotida, jarak genetik dilakukan dengan model Kimura 2 Parameter (K2P) dan rekonstruksi filogeni dilakukan dengan menggunakan program MEGA version 4 (Tamura et al. 2007).
Tabel 2. Daftar spesies Hylarana dan akses GeneBank yang digunakan dalam analisis molekuler dan konstruksi filogeni
No Spesies No. Akses Sumber Tahun
3 HASIL
Analisis Pengukuran Morfologi
Pengukuran morfologi pada spesies H. mocquardii dilakukan berdasarkan teknik pengukuran sesuai dengan Matsui 1984. Untuk analisis pengukuran, dipisahkan analisis individu jantan dengan individu betina setelah terlebih dahulu dilakukan uji t-test yang menunjukkan adanya perbedaan siginifikan (t268 = 1.69; P < 0.05) antara jantan dan betina. Spesies H. mocquardii betina memiliki nilai
Snout Vent Length (SVL) kisaran 35.74 mm hingga 83.87 mm dengan nilai rata-rata sebesar 65.50 mm (Tabel 3).
Tabel 3 Hasil Pengukuran Morfologi spesies H. mocquardii betina (n = 79) Variabel* Rata-rata Minimum Maksimum Std.deviasi
SVL 65.50 35.74 83.87 11.12
Tabel 4. Hasil Pengukuran Morfologi spesies H. mocquardii jantan (n = 189)
Keterangan : * Singkatan variabel mengacu pada keterangan Gambar 4
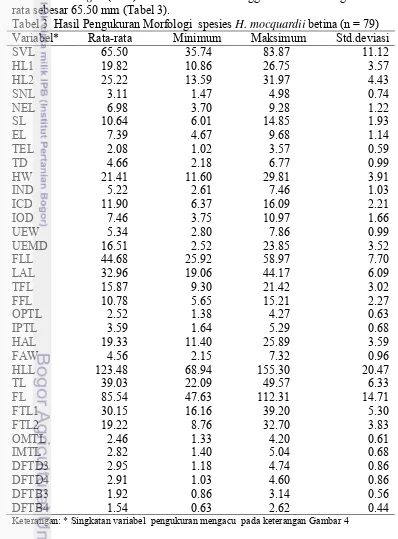
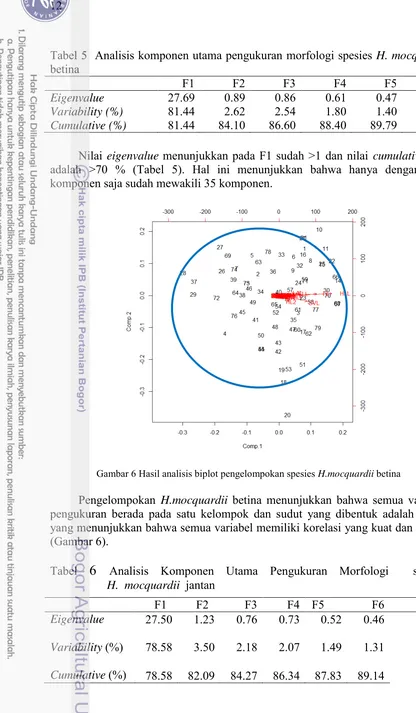
Tabel 5 Analisis komponen utama pengukuran morfologi spesies H. mocquardii
betina
F1 F2 F3 F4 F5 F6 F7
Eigenvalue 27.69 0.89 0.86 0.61 0.47 0.44 0.37
Variability (%) 81.44 2.62 2.54 1.80 1.40 1.30 0.09
Cumulative (%) 81.44 84.10 86.60 88.40 89.79 91.09 92.18
Nilai eigenvalue menunjukkan pada F1 sudah >1 dan nilai cumulative nya adalah >70 % (Tabel 5). Hal ini menunjukkan bahwa hanya dengan satu komponen saja sudah mewakili 35 komponen.
Gambar 6 Hasil analisis biplot pengelompokan spesies H.mocquardii betina
Pengelompokan H.mocquardii betina menunjukkan bahwa semua variabel pengukuran berada pada satu kelompok dan sudut yang dibentuk adalah < 90o yang menunjukkan bahwa semua variabel memiliki korelasi yang kuat dan positif (Gambar 6).
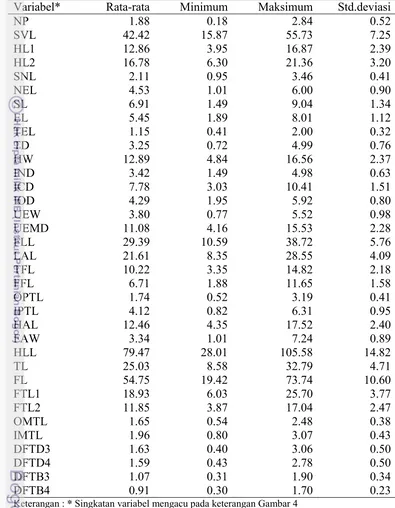
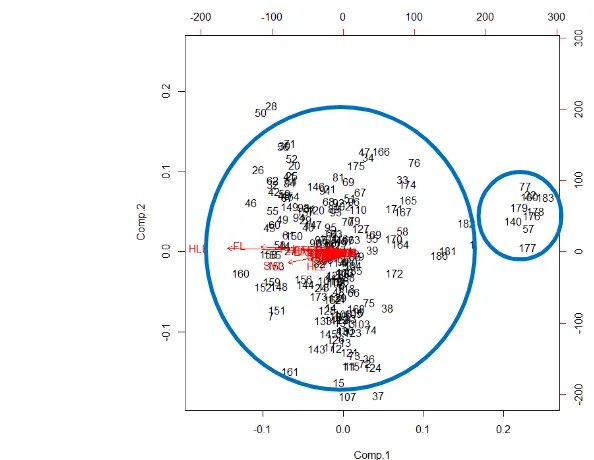
Tabel
6
Analisis Komponen Utama Pengukuran Morfologi spesiesH. mocquardii jantan
F1 F2 F3 F4 F5 F6
Eigenvalue 27.50 1.23 0.76 0.73 0.52 0.46
Variability (%) 78.58 3.50 2.18 2.07 1.49 1.31
Nilai eigenvalue >1 dan nilai cumulative juga >70 % menunjukkan bahwa satu komponen saja sudah dapat mewakili 35 komponen yang dianalisis dan tidak ada pengelompokan dalam variabel pengukuran yang dianalisis (Tabel 6)
Pengelompokan spesies H.mocquardii jantan menunjukkan bahwa semua variabel pengukuran berkumpul cenderung memiliki dua kelompok dan sudut yang dibentuk adalah < 90o yang menunjukkan bahwa semua variabel memiliki korelasi yang kuat dan positif (Gambar 7) .
Gambar 7 Hasil Analisis Biplot pengelompokan spesies H.mocquardii jantan
Variabel pengukuran yang memiliki nilai kontribusi paling besar pada betina adalah variabel HLL dengan nilai 3.51 (Lampiran 1). Uji Lanjut Duncan menunjukkan bahwa tidak ada pengelompokan berdasarkan variabel HLL sehingga tidak ada perbedaan karakteristik yang signifikan pada spesies
H.mocquardii betina di Sulawesi (Tabel 7).
Tabel 7 Hasil analisis Uji Lanjut Duncan spesies H.mocquardii betina pada variabel HLL
Wil N
Subset for alpha = 0.05 1
3.00 2 101.20
5.00 8 112.18
1.00 7 116.69
4.00 32 126.13
2.00 30 126.73
Sig. 0.05
Tabel 8 Hasil analisis Uji Lanjut Duncan spesies H. mocquardii jantan pada
Karakteristik dan Komposisi Sekuens H. mocquardii Berdasarkan Gen 12S rRNA dan 16S rRNA
Analisis gen 12S rRNA yang dianalisis terdiri dari 12 sekuens dengan pensejajaran sebesar 596 bp, yaitu 492 bp conserve sites, 103 variable sites, dan 80 parsimony informative dengan komposisi rata-rata nukleotida adalah T (U) = 24.1 %; C =28.0 %; A=30.3 %; dan G=17.6 % sedangkan analisis gen 16S rRNA terdiri dari 9 sekuens dengan pensejajaran nukleotida sebesar 665 bp, yaitu 584 conserve sites, 79 variable sites, dan 43 parsimony informative dengan komposisi rata-rata adalah T (U) = 24.3 %; C = 23.4 %; A = 37.4 %; dan G = 15.0 %.
Analisis Filogenetik Berdasarkan Gen 12S rRNA
Konstruksi filogenetik berdasarkan gen 12S rRNA membentuk dua kelompok, yaitu kelompok [Sulawesi Barat dan Sulawesi Tengah] dan [Sulawesi Utara, Sulawesi Tenggara, Sulawesi Selatan]. Untuk lebih lanjut lagi, kelompok tersebut terbagi menjadi empat kelompok, yaitu kelompok [Sulawesi Utara], [Sulawesi Barat dan Sulawesi Tengah], [Sulawesi Tenggara], dan [Sulawesi Selatan] (Gambar 10a). Jarak genetik (genetic distance) paling dekat berasal dari Sulawesi Tengah dan Sulawesi Barat sedangkan jarak genetik yang paling jauh yaitu sebesar 0.111 ditemukan pada sekuens yang berasal dari Sulawesi Selatan dan Sulawesi Tengah (Tabel 9). Spesies H. chalconota dan H. megalonesa merupakan sister clade dari kelompok spesies H. mocquardii yang didukung kuat dengan nilai bootstrap = 100. Hal ini menunjukkan kekerabatan yang sangat dekat antara H. mocquardii dengan kelompok H. chalconota yang merupakan salah satu jenis spesies kompleks.
Analisis Filogenetik Berdasarkan Gen 16S rRNA
Konstruksi filogenetik berdasarkan gen 16S rRNA menghasilkan dua kelompok yaitu kelompok [Sulawesi Utara, Sulawesi Barat, Sulawesi Tengah, Sulawesi Tenggara] dan [Sulawesi Selatan]. Untuk lebih lanjut, kelompok tersebut terbagi atas empat kelompok, yaitu kelompok [Sulawesi Utara], [Sulawesi Tengah dan Sulawesi Barat], [Sulawesi Tenggara], dan [Sulawesi Selatan] (Gambar 10b). Jarak genetik yang paling dekat ditemukan pada sekuens
berasal dari Sulawesi Selatan (Tabel 10). H. chalconota dan H. megalonesa
merupakan sister clade dari kelompok spesies H.mocquardii yang didukung dengan nilai bootstrap = 100.
4 PEMBAHASAN
Kongruensi Analisis Morfometri dengan Analisis Molekuler Gen 16S rRNA
Analisis molekuler spesies H. mocquardii di Sulawesi membentuk pengelompokan sesuai dengan pola pembentukan Pulau Sulawesi (Gambar 11). Hasil rekonstruksi filogeni secara konsisten menunjukkan bahwa spesies H. mocquardii di Sulawesi memiliki posisi monofiletik dibandingkan dengan anggota famili Ranidae. Berdasarkan gen 16S rRNA spesies H. mocquardii
membentuk dua kelompok, yaitu kelompok [Sulawesi Utara, Sulawesi Tenggara, Sulawesi Barat, Sulawesi Tengah] dan kelompok [Sulawesi Selatan]. Untuk lebih lanjut lagi, kelompok [Sulawesi Utara], [Sulawesi Barat dan Tengah], [Sulawesi Tenggara], dan [Sulawesi Selatan] membentuk empat kelompok yang terpisah.
Hasil analisis molekuler 16S rRNA menunjukkan hasil yang kongruen dengan hasil pengukuran morfologi pada individu jantan yang menempatkan Sulawesi Selatan sebagai kelompok yang terpisah. Kesesuaian hasil ini membuktikan bahwa kedua metode mampu menunjukkan adanya variasi atau diferensiasi spesies (Wien 2000). Menurut Schuh (2000) bahwa adanya variasi ekologis dan adanya barier geografis dapat memunculkan karakter yang berbeda pada individu dalam satu spesies dan akan meningkatkan diferensiasi genetik. Hal ini menunjukkan bahwa adanya tingkat efisiensi dan konsistensi yang lebih tinggi pada gen 16S rRNA dibanding gen 12S rRNA. Vences et al. (2005) membuktikan bahwa gen 16S rRNA lebih efektif sebagai marker pada amfibi (Xia et al. 2011). Selain itu, gen 16S rRNA memiliki laju evolusi (evolution rate) yang lebih rendah. Penggunaan gen 16S rRNA juga ditinjau dari segi ketersediaan database di
genebank.
Rekonstruksi Filogeni dan Evolusi Pulau Sulawesi
Hylarana mocquardii memiliki distribusi yang luas dan populasi yang besar di Sulawesi. Spesies H. mocquardii membentuk kelompok sendiri satu clade
pada posisi basal pohon filogeni (Inger et al. 2009). Adanya pengaruh faktor ekologi atau geologi dapat memberi batasan pada diversifikasi geografis yang menyebabkan adanya pola biogeografis pada taksa terkait (Evans et al. 2002). Adanya pola distribusi spesies H. mocquardii yang ada di Pulau Sulawesi merupakan hasil sejarah geografi yang kompleks yang terstruktur. Biogeografi dapat membatasi jangkauan kekuatan evolusi yang relevan. Sejarah geologi yang kompleks dengan kombinasi reorganisasi habitat terestrial dengan adanya tabrakan antara dataran Sunda dan Sahul, aktivitas pegunungan yang aktif, dan fluktuasi permukaan laut yang berulang menjadi faktor pendukung terbentuknya fragmentasi habitat, migrasi, kolonisasi spesis pada habitat baru, hingga terbentuknya spesiasi allopatrik (Esselstyn et al. 2009). Pulau ini terbentuk dari beberapa proses tabrakan beberapa lempeng yang berbeda berbeda dan memiliki barier pada empat lengan berbeda pada pulau (Hall 1996). Keadaan ini merupakan
pola sesuai dengan fragmentasi dan diversifikasi habitat (Evans et al. 2003). Selain itu, adanya topografi yang kompleks dan keragaman iklim mikro dan habitat memberi pengaruh dalam diversifikasi, spesiasi, dan variasi spesies (Bridle
et al. 2001). Adanya barrier dalam penyebaran dapat menghasilkan pola variasi pada taksa terkait yang berkerabat (Avise 2000). Kondisi ini menyebabkan Pulau Sulawesi memiliki tingkat variasi yang beragam pada tiap taksa hingga pada tingkat spesies.
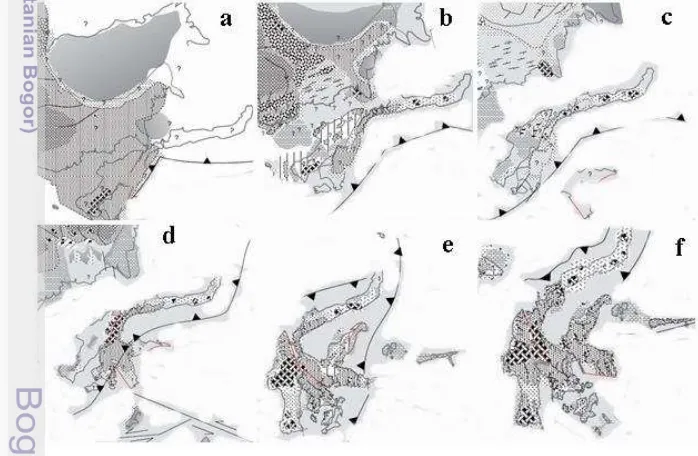
Pola pengelompokan menghasilkan satu kelompok spesies Sulawesi Barat dan Tengah (Gambar 10). Adanya satu pengelompokan antara spesies yang berada di kawasan Sulawesi Barat dan Tengah disebabkan karena kurangnya konkordansi geografis spesies yang berada di zona kontak Sulawesi Barat dan Sulawesi Tengah. Perpindahan spesies pada kedua kawasan ini salah satu faktor pendukungnya adalah adanya kesamaan tipe vegetasi, tipe tanah, dan fluktuasi iklim antara Sulawesi Tengah dan Sulawesi Barat (Whitten et al. 2002). Proses pengelompokan spesies di Sulawesi Tengah dan Sulawesi Barat kemungkinan juga disebabkan karena proses stratifikasi geologi yang menyebabkan sulawesi bagian tengah pernah menyatu (Gambar 8). Villeneuve et al. (2000) menyatakan bahwa pada masa Miocene adanya pergerakan lempeng menuju ke barat menyebabkan adanya benturan besar yang menjadikan bagian tengah Sulawesi bertekuk yang menyebabkan percampuran jenis batuan yang berasal dari lingkungan pengendapan yang berbeda.
Gambar 8 Proses Sejarah Tektonik Pulau Sulawesi (a) Eocene, (b) Middle Eocene, (c) Early Oligocene, (d) EarlyMiocene, (e) LateMiocene, (f) Early Pliocene (Moss & Wilson 1998)
Analisis jarak genetik gen 16S rRNA menunjukkan bahwa spesies Sulawesi Selatan memiliki jarak genetik yang paling jauh. Hal ini dihubungkan dengan proses evolusi tektonik pulau Sulawesi yang menyebabkan adanya perbedaan stratigrafi khususnya pada kawasan Sulawesi Selatan berupa perbedaan sedimen dan vulkanik (Suyono & Kusnama 2010). Aktivitas vukanik pada masa Paleocene
pada saat penurunan permukaan laut 100 m menunjukkan terjadinya pelebaran permukaan antara kawasan Kalimantan Selatan dan Sulawesi Selatan yang memungkinkan terjadinya kontak antara kedua kawasan tersebut yang kemudian terpisah kembali oleh Selat Makassar pada saat naiknya permukaan laut (Katili 1978). Proses penurunan suhu global juga terjadi di kawasan Sulawesi Selatan pada masa pertengahan Eocene (Gower et al. 2012). Hal ini memberi dampak terhadap kondisi ekologis dari spesies H. mocquardii yang rentan terhadap perubahan habitat dan kemungkinan menjadi faktor yang menjadi penyebab spesies di kawasan ini memiliki pengelompokan sendiri dibandingkan dengan spesies di kawasan yang lainnya.
Analisis Proses Invasi Spesies H.mocquardii berdasarkan analisis Filogeni
Berdasarkan hasil ini diasumsikan bahwa nenek moyang dari spesies ini berasal dari daratan Sunda (Gambar 9). Proses masuknya (invasi) spesies ini menuju Pulau Sulawesi yang termasuk kawasan Wallacea diduga memiliki hubungan dengan proses terbentuknya Pulau Sulawesi.
Gambar 9 Pembagian kawasan daratan yang terdiri dari kawasan Daratan Sunda, Wallacea, dan kawasan Daratan Sahul (Bacon et al. 2013).
(a) (b)
19 Gambar 11 Pola Filogeografi H. mocquardii di Sulawesi berdasarkan gen 12S rRNA dan 16S rRNA
Tabel 9 Jumlah perbedaan nukleotida (bawah diagonal) dan jarak genetik (atas diagonal) berdasarkan gen 12S rRNA
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21
21 Tabel 10 Jumlah perbedaan nukleotida (bawah diagonal) dan jarak genetik (atas diagonal) berdasarkan gen 16S rRNA
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18
5
SIMPULAN DAN SARAN
Simpulan
a. Analisis pengukuran morfologi menghasilkan pengelompokan spesies
H.mocquardii betina menjadi satu kelompok, sedangkan pada jantan menjadi dua kelompok, yaitu kelompok dari [Sulawesi Utara, Sulawesi Barat, Sulawesi Tengah, Sulawesi Tenggara] dan [Sulawesi Selatan]. b. Analisis molekuler gen 12S rRNA menghasilkan 2 pengelompokan yaitu
kelompok [Sulawesi Barat dan Sulawesi Tengah] dan [Sulawesi Utara, Sulawesi Tenggara, Sulawesi Selatan]. Gen 16S menghasilkan 2 pengelompokan yaitu [Sulawesi Utara, Sulawesi Barat, Sulawesi Tengah, Sulawesi Tenggara] dan [Sulawesi Selatan]. Untuk lebih lanjut, kedua gen menghasilkan pengelompokan menjadi 4 kelompok, yaitu kelompok [Sulawesi Utara], [Sulawesi Barat dan Sulawesi Tengah], [Sulawesi Tenggara], dan Sulawesi Selatan
c. Hasil pengukuran morfologi individu jantan memiliki hasil pengelompokan yang kongruen dengan hasil molekuler gen 16S rRNA d. Proses invasi spesies H.mocquardii di Sulawesi kemungkinan terjadi pada
saat proses pembentukan Pulau Sulawesi dan nenek moyang spesies ini berasal dari daratan Sunda
Saran
Analisis selanjutnya perlu dilakukan analisis nuclear DNA tingkat populasi pada masing-masing kawasan untuk melihat aliran gen (gene flow)
DAFTAR PUSTAKA
Avise JC. 2000. Phylogeography: The History and Formation of Species. Cambridge (GB). Harvard University Press.
Bacon CD, Michonneau F, Henderson AJ, McKenna MJ, Milroy AM, & Simmons MP. 2013. Geographic and taxonomic disparities in species
diversity: dispersal and diversification rates across Wallace’s line. Evolution. 67-7:2058-2071.
Bridle JR, Butlin RK, Schulter D. 2009. Speciation and Patterns of Diversity. Cambridge (GB). Cambridge University Press.
Campbell P, Putnam AS, Bonney C, Bilgin R, Morales JC, Kunz TH, Ruedas LA. 2007. Contrasting patterns of genetic differentiation between endemic and widespread species of fruit bats (Chiroptera : Pteropodiae) in Sulawesi, Indonesia. Molecular Phylogenetics and Evolution. 44:474-482.
Chernoff B. 1982. Character Variation among population and the analysis of biogeography. Amer.Zool. 22 : 425-439.
Ciani A, Stanton R, Scheffrahn W, Sampurno W. 1989. Evidence of gene flow between Sulawesi Macaques. Am J Primatol. 17:257-270.
Dubois A. 1992. Notes sur la classiffication des Ranidae (Amphibiens Anoures).
Bulletin Mensuel de la Societe Linneenne de Lyon 61:305-352.
Esselstyn J A, Timm RM, and Brown RM. 2009. Do geological or climatic processes drive speciation in dynamic archipelagos? The tempo and mode of diversification in Southeast Asian shrews. Evolution. 63:2595-2610. Evans B J, Supriatna J, Andayani N, Setiadi MI, Cantella DC. 2003. Monkeys
and toads define areas of endemism on Sulawesi. Evolution; international journal of organic evolution. 57 (6) :1436–1443.
Evans BJ, J Supriatna, and D J Melnick. 2001. Hybridization and population genetics of two macaque species in Sulawesi, Indonesia. Evolution
55:1685-1702.
Evans B J, Brow RM, McGuire JA, A Diesmos, and D C Cannatella. 2002.
Crossing Wallace's line : molecular phylogeography of Limnonectes (Ranidae) in Borneo, the Phillipines. and Sulawesi. Kansas City (US). Farajallah A. 2002. Karakterisasi Genom Mitokondria Labi-Labi Dogania
subplana (Trionychidae, Testudines, Reptilia) [Disertasi]. Program Pascasarjana, Institut Pertanian Bogor. Bogor (ID).
Frost D. 1985. Amphibian Species of the World: A Taxonomic and Geographic Reference. USA (US) : Allen Press Inc.
Gower D J, Johnson KG, Richardson JE, Rosen BR, Rulber L, & Williams ST. 2012. Biotic Evolution and Environmental Change in Southeast Asia. New York (US): Cambridge University Press.
Hall R. 1996. Reconstructing Cenocoiz SE Asia and Tectonic Evolution of Southeast Asia. London (UK): Geological Society Special Publication. Hay JM, Ruvinsky I, Hedges SB, & Maxson LR. 1995. Phylogenetic relationships
of amphibian families inferred from DNA sequences of mitochondrial 12S and 16S Ribosomal RNA genes. Mol.Biol.Evol 12(5):928-937.
Hillis DM. 1987. Molecular Versus morphological approaches to systematics.
Ann. Rev. Ecol. Syst. 18:23-42.
Hillis DM & J.J Wiens. 2000. Molecules Versus Morphology in Systematics : Phylogenetic Analysis of Morphological Data. Philadelphia (US ): Smithshonian Institution Press.
Hedges SB, LR Maxson. 1993. Caecilian phylogeny and biogeography inferred from mitocondrial DNA sequences of the 12S and 16S rRNA genes (Amphibia : Gymnophiona). Herpetol. Monogr. 7:64-76.
Inger RF. 1966. The systematics and zoogeography of the Amphibia of Borneo.
Fieldiana: Zoology. 52:1-402.
Inger RF, Stuart BL & Iskandar DT. 2009. Systematics of a widespread Southeast Asian frog, Rana chalconota (Amphibian: Anura: Ranidae). Zoological Journal of the Linnean Society. 155:123-147.
Inger RF & Voris HK. 2001. The biogeographical relations of the frogs and snakes of Sundaland. Journal of Biogeography. 28:863-891.
Iskandar DT. 1998. Amfibi Jawa dan Bali. Indonesia (ID) : Puslitbang Biologi – LIPI.
Iskandar DT & Colijin E. 2000. Preliminary checklist of Southeast Asia and New Guinean herpetfauna Amphibians. Truebia. 31:1-133.
Kampen PN. 1923. The Amphibia of the Indo-Australian Archipelago. Leiden Holland (NL): E.J. Brill Ltd.
Katili. 1978. Past and present geotectonics in the Indonesian Island areas.
Tectonophysics. 45:289-322.
Kimura M. 1968. Evolutionary rate at the molecular level. Nature. 217:624-626. Matsui M. 1984. Morphometric Variation Analyses and Revision of the Japanese
Toads (Genus Bufo, Bufonidae). Tokyo (JP) : Kyoto University Press. McGuire JA. 2002. Bayesian Phylogenetics and Biogeography of the
Dracolineatus grup. p. 212 in Program Book and Abstracts of the Joint Meeting of Ichthologist and Herpetologist. 3-8 July, Kansas.
Moss SJ & Wilson, EJ. 1998. Biogeography and Geological Evolution of SE Asia. Netherlands (NL) : Backbuys Publisher.
Munshi JS & HM Dutta. 1996. Fish Morphology: Horizon of New Research. New York (US): Science Publishers, Inc.
Schuh RT. 2000. Biological Systematic: Principles and Aplications. London (UK): Cornel Universty Press.
Stelbrink B, Albrecht C, Hall R, & Rintelen TV. 2011. The biogeography of Sulawesi revisited : is there evidence for a vicariant origin of taxa on
Wallace’s“anomalous island”?. Evolution. 66 (7):1-20.
Suyana & Kusnama. 2010. Stratigraphy and tectonics of the Sengkang Basin, South Sulawesi. Jurnal Geologi Indonesia. 5 (1):1-11.
Tamura K, Dudley J, Nei M, & Kumar S. 2007. Mega4 : Molecular evolutionary genetics analysis (MEGA) software version 4.0. Molecular Biology and Evolution. 24:1596-1599.
Tegelstrom H. 1986. Mitochondrial DNA in natural population : an improved routine for screening of genetic variation based on sensitive silver staining. Electrophoresis. 7(5):226-229.
Tjandra L. 2011. Analisis Filogenetik Bufo melanostictus, Schneider 1799 dan
Vences M, Kosuch J, Lotters S, Widmer A, Jungfer K, Kohler J, & Veith M. 1999. Phylogeny and classification of poison frog (Amphibia: Dendrobatidae) based on mitochondrial 16S and 12S ribosomal RNA gene sequences.
Molecular Phylogenetics and Evolution 15(1):34-40.
Vences M, Thomas M, Meijden A, Chiari Y, & Vieites DR. 2005. Comparative performance of the 16S rRNA gene in DNA barcoding of amphibians.
Frontiers in Zoology 2:5
Villeneuve M, Gunawan W, Cornee JJ, & Vidal O. 2000. Geology of the Central Sulawesi belt (Eastern Indonesia): constraints for geodynamic models.
Int J Earth Sci .91:524-537.
Whitten T, Henderson GS, & Mustafa M. 2002. The Ecology of Sulawesi. Hongkong (TW): Periplus Editions.
Wien J J. 2000. Phylogenetic, Analysis of Morphological Data. USA (US): Smithsonian Institution Press.
Wiens J J. 2004. The Role of Morphological data ini phylogeny reconstruction.
Syst. Biol. 53(4):653-661.
Xia Y, Gu HF, Peng R, Chen Q, Zheng YC, Murphy RW, & Zeng XM. 2011. COI is better than 16S rRNA for DNA barcoding Asiatic salamanders (Amphibia: Caudata: Hynobiidae). Molecular Ecology Resourches
12:48-56.
LAMPIRAN
Lampiran 1. Analisis kontribusi masing-masing variabel terhadap komponen pada spesies H. mocquardii betina
Lampiran 2. Analisis kontribusi masing-masing variabel terhadap komponen pada spesies H. mocquardii jantan
Lampiran 3 Data Spesimen H.mocquardii yang digunakan untuk analisis morfometri
Provinsi Lokasi No.MZB No.Field Sex Kolektor Tanggal Koleksi
Sulawesi Utara Desa Tudi, Kecamatan
Anggrek, Kabupaten Gorontalo
13953 0642 F Jimmy McGuire et al. 18 Oktober 2004
Sulawesi Utara Desa Bubade, Kecamatan
Kuandang, Kabupaten
Sulawesi Utara Camp. Tulido, Sungai
Hialioda
17716 GT 123 F Dadang R.S. 24 Agustus 2009
Sulawesi Utara Desa Tudi, Kecamatan
Anggrek, Kabupaten Gorontalo
13954 BSIFS 0746 F Jimmy McGuire et al. 18 Oktober 2004
Sulawesi Utara Pos PHPA, Sungai
Matayangan. Kecamatan Dumoga, Kabupaten Bolaang Mangondow. TNBN Warta Bone
3622 3622 F Ibnu Maryanto 16 Oktober 1995
Sulawesi Utara Bogani Nani Wartabone. TN
Kala Bolang Mangandow, Toraut
8048 JAM 3678 F Jimmy McGuire et al. 07 September
2001
Sulawesi Utara Desa Lombongo. Kecamatan
Suwawa
13952 BSIFS 0624 M Jimmy McGuire et al. 18 Oktober 2004
Sulawesi Utara Desa Bubade, Kecamatan
Kuandang
13947 BSIFS 0546 M Jimmy McGuire et al. 18 Oktober 2004
Sulawesi Utara Desa Bubade, Kecamatan
Kuandang
29
Sulawesi Utara Desa Bubade, Kecamatan
Kuandang
13950 BSIFS 0549 M Jimmy McGuire et al. 18 Oktober 2004
Sulawesi Utara Camp. Tulido, Sungai
Hialioda
17714 GT 123 M Dadang R.S. 24 Agustus 2009
Sulawesi Utara Camp. Tulido, Sungai
Hialioda
17723 GT 72 M Dadang R.S. 18 Agustus 2009
Sulawesi Utara Camp. Tulido, Sungai
Hialioda
17715 GT 123 M Dadang R.S. 24 Agustus 2009
Sulawesi Utara Kaki Gunung Boliyuhuto,
Margasatwa Nantu
23288 KBAA 12 M Mirza D. Kusrini, Aria
Nusantara, Luna Raftika
18 Juni 2013
Sulawesi Utara Camp. Tulido, Sungai
Hialioda
17725 GT 62 M Dadang R.S. 18 Agustus 2009
Sulawesi Utara Camp 2, Sungai Poatnudaa,
Suaka margasatwa Nantu
Sulawesi Utara Bogani Nani Wartabone, TN
Kala Bolaang Mangandow, Toraut
8047 JAM 3675 M Jimmy McGuire et al. 07 September
2001
Sulawesi Utara Bogani Nani Wartabone, TN
Kala Bolaang Mangandow,
8049 JAM 3679 M Jimmy McGuire et al. 07 September
Toraut
Sulawesi Utara Desa Manemboku Possi,
Kabupaten Bolang Mangondow
14511 JAM 2036 M Jimmy McGuire et al. 14 Oktober 2004
Sulawesi Utara Desa Manemboku Possi,
Kabupaten Bolang Mangondow
14512 BSIFS 2044 M Jimmy McGuire et al. 14 Oktober 2004
Sulawesi Utara Desa Manemboku Possi,
Kabupaten Bolang
Sulawesi Tengah Desa Mataue 13967 BSIFS 1455 F Rave Brown, Chris
Hayden, Umi , Iqbal, Abdul, Ferdy
04 November 2004
Sulawesi Tengah Desa Mataue 13961 BSIFS 1381 F Rave Brown, Chris
Hayden, Umi , Iqbal, Abdul, Ferdy
03 November 2004
Sulawesi Tengah Desa Mataue 13962 BSIFS 1382 F Rave Brown, Chris
Hayden, Umi , Iqbal, Abdul, Ferdy
03 November 2004
Sulawesi Tengah Desa Mataue 13958 BSIFS 1359 F Rave Brown, Chris
Hayden, Umi , Iqbal, Abdul, Ferdy
03 November 2004
Sulawesi Tengah Desa Kali Bulu, Kecamatan
Bulasan
13955 BSIFS 1217 F David, Freddy, Umi,
Ferdi
29 Oktober 2004
Sulawesi Tengah Desa Porigipun, Kecamatan
Perigi, Kabupaten Mountong
31
Sulawesi Tengah Desa Mataue 13973 BSIFS 1547 F Umi, Freddy, David,
Iqbal
06 November 2004
Sulawesi Tengah Desa Tomado 13970 BSIFS 2142 F Rave Brown,
Iqbal,Chris Hayden, Umi, Abdul, Ferdy
13 November 2004
Sulawesi Tengah Desa Tomado 13969 BSIFS 2113 F Rave Brown,
Iqbal,Chris Hayden, Umi, Abdul, Ferdy
12 November 2004
Sulawesi Tengah Desa Karya Agung,
Kecamatan Moutong, Kabupaten Parimo
11381 AR 40008 F Awal R. & Mulyadi 2004
Sulawesi Tengah Desa Karya Agung,
Kecamatan Moutong, Kabupaten Parimo
11384 AR 40013 F Awal R. & Mulyadi 2004
Sulawesi Tengah Desa Karya Agung,
Kecamatan Moutong, Kabupaten Parimo
11382 AR 40011 F Awal R. & Mulyadi 2004
Sulawesi Tengah Desa Karya Agung,
Kecamatan Moutong, Kabupaten Parimo
11383 AR 40012 F Awal R. & Mulyadi 2004
Sulawesi Tengah Desa Tomado 14010 BSIFS 2115 F Freddy Chain, Ferdi,
Umi Laela
12 Noember 2004
Sulawesi Tengah Desa Tomado 14004 BSIFS 1982 F Freddy Chain, Ari,
Umi Laela
10 November 2004
Sulawesi Tengah Desa Tomado 14019 JAM 4979 F Freddy Chain, Ferdy,
Umi Laela
11 November 2004
Sulawesi Tengah Desa Tomado 14018 BSIFS 4978 F Freddy Chain, Ferdy,
Umi Laela
11 November 2004
Sulawesi Tengah Desa Tomado 14013 BSIFS 2139 F Freddy Chain, Ferdy,
Umi Laela
Sulawesi Tengah Desa Mataue, Kecamatan Kulawi, Kabupaten Donggala
14001 BSIFS 1446 F Rave Brown 03 November
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
13996 BSIFS 1365 F Rave Brown 03 November
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
13993 BSIFS 1358 F Rave Brown 03 November
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
13994 BSIFS 1352 F Rave Brown 03 November
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Mataue, Kecamatan
33
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Dadahitan, Kecamatan
Baolan. Kabupaten Toli-toli
13982 RMB 4643 M Rave Brown, Chris
Hayden, Charles Linken
27 Oktober 2004
Sulawesi Tengah Desa Dadahitan, Kecamatan
Baolan. Kabupaten Toli-toli
13983 RMB 4644 M Rave Brown, Chris
Hayden, Charles Linken
27 Oktober 2004
Sulawesi Tengah Desa Dadahitan, Kecamatan
Baolan. Kabupaten Toli-toli
13984 RMB 4754 M Rave Brown, Chris
Hayden, Charles Linken
27 Oktober 2004
Sulawesi Tengah Desa Karya Agung,
Kecamatan Moutong, Kabupaten Parimo
11386 AR 40015 M Awal R. & Mulyadi 2004
Sulawesi Tengah Desa Karya Agung,
Kecamatan Moutong, Kabupaten Parimo
11387 AR 40024 M Awal R. & Mulyadi 2004
Sulawesi Tengah Desa Karya Agung,
Kecamatan Moutong, Kabupaten Parimo
11388 AR 40048 M Awal R. & Mulyadi 2004
Sulawesi Tengah Desa Karya Agung,
Kecamatan Moutong, Kabupaten Parimo
11389 AR40051 M Awal R. & Mulyadi 2004
Sulawesi Tengah Desa Tomado 14011 BSIFS 2116 M Freddy Chain, Ferdi,
Umi Laela
12 November 2004
Umi Laela 2004
Sulawesi Tengah Desa Tomado 14009 BSIFS 2141 M Freddy Chain, Ferdi,
Umi Laela
13 November 2004
Sulawesi Tengah Desa Tomado 14003 BSIFS 1980 M Freddy Chain, Ari,
Umi Laela
10 November 2004
Sulawesi Tengah Desa Tomado 14005 BSIFS 1984 M Freddy Chain, Ari,
Umi Laela
10 November 2004
Sulawesi Tengah Desa Tomado 14006 BSIFS 1988 M Freddy Chain, Ari,
Umi Laela
10 November 2004
Sulawesi Tengah Desa Tomado 14007 BSIFS 1989 M Freddy Chain, Ari,
Umi Laela
10 November 2004
Sulawesi Tengah Desa Tomado 14020 JAM 05096 M Freddy Chain, Ferdy,
Umi Laela
11 November 2004
Sulawesi Tengah Desa Tomado 14021 JAM 05097 M Freddy Chain, Ferdy,
Umi Laela
11 November 2004
Sulawesi Tengah Desa Tomado 14022 JAM 05098 M Freddy Chain, Ferdy,
Umi Laela
11 November 2004
Sulawesi Tengah Desa Tomado 14015 BSIFS 2153 M Freddy Chain, Ferdy,
Umi Laela
11 November 2004
Sulawesi Tengah Desa Tomado 14016 BSIFS 2156 M Freddy Chain, Ferdy,
Umi Laela
11 November 2004
Sulawesi Tengah Desa Tomado 14017 BSIFS 4912 M Freddy Chain, Ferdy,
Umi Laela
11 November 2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
14000 BSIFS 1387 M Rave Brown 03 November
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
14002 BSIFS 1447 M Rave Brown 03 November
2004
35 Kulawi, Kabupaten
Donggala
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
13998 BSIFS 1383 M Rave Brown 03 November
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
13999 BSIFS 1386 M Rave Brown 03 November
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
13991 BSIFS 1343 M Rave Brown 03 November
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
13992 BSIFS 1357 M Rave Brown 03 November
2004
Sulawesi Tengah Desa Mataue, Kecamatan
Kulawi, Kabupaten Donggala
13995 BSIFS 1363 M Rave Brown 03 November
2004
Sulawesi Tengah Desa Balapapu, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Balapapu, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Balapapu, Kecamatan
Kulawi, Kabupaten
Sulawesi Tengah Desa Salodik, Kecamatan
Luwuk, Kabupaten Banggai
8057 JAM 3730 M Jimmy McGuire et al. 22 September
2001
Luwuk, Kabupaten Banggai 2001
Sulawesi Tengah Desa Salodik, Kecamatan
Luwuk, Kabupaten Banggai
8059 JAM 3734 M Jimmy McGuire et al. 22 September
2001
Sulawesi Tengah Desa Salodik, Kecamatan
Luwuk, Kabupaten Banggai
8050 JAM 3715 M Jimmy McGuire et al. 22 September
2001
Sulawesi Tengah Desa Salodik, Kecamatan
Luwuk, Kabupaten Banggai
8051 JAM 3718 M Jimmy McGuire et al. 22 September
2001
Sulawesi Tengah Desa Salodik, Kecamatan
Luwuk, Kabupaten Banggai
8052 JAM 3719 M Jimmy McGuire et al. 22 September
2001
Sulawesi Tengah Desa Salodik, Kecamatan
Luwuk, Kabupaten Banggai
8053 JAM 3720 M Jimmy McGuire et al. 22 September
2001
Sulawesi Tengah Desa Boya, Kecamatan
Kulawi
13900 BSIFS 1753 M Freddy Chain 12 November
2004
Sulawesi Tengah Desa Labanu, Kecamatn
Dando, Kabupaten Tali-tali
11344 HR 40105 M Awal R. & Mumpuni 2004
Sulawesi Tengah Desa Labanu, Kecamatn
Dando, Kabupaten Tali-tali
11345 HR 40110 M Awal R. & Mumpuni 2004
Sulawesi Tengah Desa Labanu, Kecamatn
Dando, Kabupaten Tali-tali
11346 KR 04 M Awal R. & Mumpuni 2004
Sulawesi Tengah Desa Labanu, Kecamatn
Dando, Kabupaten Tali-tali
11347 KR 06 M Awal R. & Mumpuni 2004
Sulawesi Tengah Sei Kramat, Gunung Dako
(300 m), Lakaton
Kinapasang, Kabupaen Toli-toli
2814 2814 M Boeadi 22-28 Februari
1985
Sulawesi Tengah Sei Kramat, Gunung Dako
(300 m), Lakaton
Kinapasang, Kabupaen Toli-toli
2814 2814 M Boeadi 22-28 Februari
1985
37
Kabupaten Banggai 2005
Sulawesi Tengah Tepi Sungai Salodik,
Kabupaten Banggai
11466 AK 137 M Andre Koch 17 Agustus 2005
Sulawesi Tengah Morawali 8413 UI 1289 M UI Jakarta Juli 2001
Sulawesi Barat Desa Polewali, Massawa
Road River I
12422 JAM 05936 F Jimmy McGuire et al. 30 Oktober 2005
Sulawesi Barat Desa Kabinan, Kabupaten
Majene
12405 JAM 06286 F Jimmy McGuire et al. 07 November
2005
Sulawesi Barat Desa Tomnonga, Kecamatan
Pulowai, Kabupaten
Sulawesi Barat Desa Tomnonga, Kecamatan
Pulowai, Kabupaten
Sulawesi Barat Polewali Massawa Road
(River I)
12400 JAM 05942 M Jimmy McGuire et al. 30 Oktober 2005
Sulawesi Barat Polewali Massawa Road
(River I)
12402 JAM 05934 M Jimmy McGuire et al. 30 Oktober 2005
Sulawesi Barat Desa Tasiu, Tibo Road 12403 JAM 06559 M Jimmy McGuire et al. 09 November
2005
Sulawesi Barat Desa Kabinan, Kabupaten
Majene
12404 JAM 06284 M Jimmy McGuire et al. 09 November
2005
Sulawesi Barat Desa Tasiu, Tibo Road 12406 JAM 06554 M Jimmy McGuire et al. 09 November
2005
Sulawesi Barat Polewali Massawa Road
(River I)
12407 JAM 05938 M Jimmy McGuire et al. 30 Oktober 2005
Sulawesi Barat Desa Tasiu, Tibo Road 12409 JAM 06568 M Jimmy McGuire et al. 09 November
2005
Sulawesi Barat Desa Tasiu, Tibo Road 12410 JAM 06564 M Jimmy McGuire et al. 09 November
Sulawesi Barat Polewali Massawa Road (River I)
12412 JAM 05940 M Jimmy McGuire et al. 30 Oktober 2005
Sulawesi Barat Desa Kabinan, Kabupaten
Majene
12414 JAM 06278 M Jimmy McGuire et al. 01 November
2005
Sulawesi Barat Desa Kabinan, Kabupaten
Majene
12416 JAM 06280 M Jimmy McGuire et al. 07 November
2005
Sulawesi Barat Desa Makuang, Kecamatan
Massawa
12418 JAM 05996 M Jimmy McGuire et al. 07 November
2005
Sulawesi Barat Desa Kabinan, Kabupaten
Majene
12419 JAM 06276 M Jimmy McGuire et al. 07 November
2005
Sulawesi Barat Desa Kabinan, Kabupaten
Majene
12421 JAM 06288 M Jimmy McGuire et al. 07 November
2005
Sulawesi Barat Desa Tasiu, Tibo Road 12423 JAM 06569 M Jimmy McGuire et al. 01 November
2005
Sulawesi Barat Desa Tasiu, Tibo Road 12426 JAM 06561 M Jimmy McGuire et al. 09 November
2005
Sulawesi Barat Desa Polewali, Massawa
39
Sulawesi Tenggara Gunung Putih, Desa
Lampeapi, Pulau Wawani
10833 W59 F Mumpuni 27 April 2004
Sulawesi Tenggara Stream of Nat.Res of
Lambusango, Kecamatan Kapontori, Kabupaten Buton
9444 Km 227 F Mumpuni 05 Juni 2003
Sulawesi Tenggara Stream under 1st bridge on
road North of Talingko
Deve’s sites
7703 GG 079A F Graeme Gillespie 24 Juli 2001
Sulawesi Tenggara Stream under 1st bridge on
road North of Talingko
Sulawesi Tenggara Desa Ewo-ewo, Kecamayan
Pomalaa, Kabupaten Kolaka
21961 PM 054 F Hellen K. &
Syarifudin
23 Mei 2013
Sulawesi Tenggara Air Terjun Kambu, Kota
Kendari
9333 JAM 4441 F Jimmy McGuire et al. 18 Agustus 2005
Sulawesi Tenggara Desa Oko-oko, Kecamatan
Pomalaa, Kabupaten Kolaka
21956 PM 043 F Hellen K. &
Syarifudin
21 Mei 2013
Sulawesi Tenggara Desa Toldo, Kecamatan
Tirauta, Kabupaten Kolaka
Sulawesi Tenggara Pegunungan Mekongga,
Sulawesi Tenggara Desa Larumerui, Kecamatan
41
Sulawesi Tenggara Kendari 9926 T 62 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9927 T 49 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9928 T 30 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9929 T 12 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9930 T 53 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9911 T 33 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9912 T 2 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9913 T 114 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9914 T 56 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9915 T 123 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9916 T 37 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9917 T 25 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9918 T 126 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9919 T 102 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9920 T 40 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9897 T 54 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9898 T 61 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9899 T 14 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9900 T 90 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9901 T 55 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9902 T 63 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9903 T 29 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9904 T 66 M Djoko T. Iskandar 1995
Sulawesi Tenggara Kendari 9905 T 94 M Djoko T. Iskandar 1995
Sulawesi Tenggara Tanjung Peropa, Kabupaten
Kendari
8342 RJL 266 M WCS 04 Agustus 2000
Sulawesi Tenggara Desa Laumerui, Kecamatan
Rauta, Kabupaten Kolaka
23345 B 54 M Ucok 04 Oktober 2013
Sulawesi Tenggara Desa Ewo-ewo, Kecamatan
Pomalaa, Kabupaten Kolaka
21959 PM 052 M Hellen K. &
Syarifudin
Sulawesi Tenggara Desa Ewo-ewo, Kecamatan Pomalaa, Kabupaten Kolaka
21960 PM 053 M Hellen K. &
Syarifudin
23 Mei 2013
Sulawesi Tenggara Desa Oko-oko, Kecamatan
Pomalaa, Kabupaten Kolaka
21957 PM 044 M Hellen K. &
Syarifudin
21 Mei 2013
Sulawesi Tenggara Desa Oko-oko, Kecamatan
Pomalaa, Kabupaten Kolaka
21958 PM 045 M Hellen K. &
Syarifudin
21 Mei 2013
Sulawesi Tenggara Desa Wonakoea, Kecamatan
Pomalaa, Kabupate Kolaka
21953 PM 009 M Hellen K. &
Syarifudin
17 Mei 2013
Sulawesi Tenggara Desa Wonakoea, Kecamatan
Pomalaa, Kabupate Kolaka
21954 PM 010 M Hellen K. &
Syarifudin
17 Mei 2013
Sulawesi Tenggara Desa Wonakoea, Kecamatan
43
Sulawesi Tenggara TN Rawa Aopa, Desa
Tatangge, Kabupaten Kolaka
17570 RAWH 76 M Irine & Arman Rauf 28 November
2011
Sulawesi Tenggara Kendari 6356 UI 78 M Ben Evens 07 Agustus 2000
Sulawesi Tenggara Sungai Enano, Buton 7707 GG KB6601
013
M Graeme Gillespie 31 Agustus 2001
Sulawesi Selatan Desa Bonto Maranu (Loka) 12399 JAM 05502 F Jimmy McGuire et al. 21 Oktober 2005
Sulawesi Selatan Desa Bonto Maranu (Loka) 12401 JAM 05504 F Jimmy McGuire et al. 21 Oktober 2005
Sulawesi Selatan Kelurahan Lewaja, Kabuaten
Enrekang