POSISI FILOGENI
Orlitia borneensis
DAN
Hieremys annandalei
(TESTUDINES) BERDASARKAN GEN 16S rRNA
D. KURNIAWATI
G04400049
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
D. KURNIAWATI. Posisi Filogeni Orlitia borneensis dan Hieremys annandalei
(TESTUDINES) Berdasarkan Gen 16S rRNA. Dibimbing oleh ACHMAD FARAJALLAH dan RIKA RAFFIUDIN.
Urutan DNA mitokondria (mtDNA) banyak digunakan sebagai penanda molekuler dalam menentukan hubungan filogeni suatu organisme karena struktur organisasinya yang stabil. Seiring dengan kemajuan bioteknologi, pengelompokkan kura-kura Bataguridal masih menjadi perdebatan. Hal ini disebabkan oleh perbedaan karakter yang diamati. Beberapa hasil penelitian mengelompokkan kura ini ke dalam famili Emydidae tetapi ada yang menempatkan kura-kura ini ke dalam famili tersendiri yaitu famili Bataguridae.Pada penelitian ini digunakan urutan nukleotida gen 16S rRNA O. borneensis dan H. annandalei (Testudines) yang disejajarkan dengan urutan nukleotida beberapa spesies Testudines lainnya yang terdapat di Genbank. Hasil rekonstruksi pohon filogeni berdasarkan urutan nukleotida 16S rRNA menunjukkan bahwa O. borneensis dan H. annandalei berkerabat dengan Chinemys reevesi (famili Bataguridae). Hasil amplifikasi gen 16S rRNA O. borneensis dan H. annandalei menunjukkan komposisi nukleotida didominasi oleh nukleotida adenin dan timin dengan nilai perbandingan antara A/T dan G/C = 60.1%:39.9% untuk O. borneensis dan 63%:37% untuk H. annandalei. Rasio transisi terhadap transversi (ts/tv) gen 16S rRNA berkisar antara 0.67 sampai dengan 6.75. Nilai jarak genetik terkecil terjadi antara Trachemys scripta dan Chrysemys picta dan antara Dogania subplana dan
Tracemys scripta yaitu sekitar 0.005 sedangkan nilai jarak genetik terbesar terjadi antara
Pelomedusa subrufa dan Emydurasubglobusa yaitu sekitar 0.869.
ABSTRACT
D. KURNIAWATI. Phylogenetic Position of Orlitia borneensis and Hieremys
annandalei (TESTUDINES) Based on 16S rRNA Gene. Supervised by ACHMAD
FARAJALLAH and RIKA RAFFIUDIN.
Mitochondrial gene has been commonly used as molecular marker in phylogenetic analysis. Due to its stable organization structure, mtDNA is often used to determine phylogenetic relationship of organisms. Along with the increasing improvements in biotechnology methods, classification of Bataguridal turtles was still in debate among scientists. This is mainly due to the differences in the observed characteristics. Several studies placed Bataguridal turles into Emydidae family, but others placed Bataguridal turtles into a separated family: Bataguridae. This research used sequence data of 16S rRNA of Orlitia borneensis and Hieremys annandalei aligned with those of several Testudines species deposited in Genbank. DNA alignment conducted using Clustal W 1.8 software. Nucleotide composition, transition and transvertion substitution type (ts/tv), determination of genetic distance and phylogenetic trees reconstruction were performed using MEGA 3.0 software. The result of phylogenic trees reconstruction based on 16S rRNA showed that Orlitia borneensis and Hieremys annandalei were closely related to Chinemys reevesi
(Bataguridae). Amplification of 16S rRNA gene of Orlitia borneensis and Hieremys annandalei
showed that the nucleotide composition was dominated by adenine and timin nucleotides with A/T:G/C value of 60.1%:39.9% for Orlitia borneensis and 63%:37% for Hieremys annandalei.
POSISI FILOGENI
Orlitia borneensis
DAN
Hieremys annandalei
(TESTUDINES) BERDASARKAN GEN 16S rRNA
D. KURNIAWATI
Skripsi
sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains pada Fakultas Matematika dan IPA
Institut Pertanian Bogor
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
Judul Skripsi : Posisi Filogeni
Orlitia borneensis
dan
Hieremys annandalei
(Testudines) Berdasarkan Gen 16S rRNA
Nama
: D. Kurniawati
NRP
: G04400049
Menyetujui:
Pembimbing I
Pembimbing II
Dr. Ir. Achmad Farajallah, M.Si. Dr. Ir. Rika Raffiudin, M.Si.
NIP.131878947 NIP.131999583
Mengetahui:
Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
Prof.Dr. Ir. Yonny Koesmaryono, M.S
NIP 131473999
PRAKATA
Segala puji hanya untuk Allah SWT atas segala nikmat, cahaya dan karunia-Nya kepada penulis sehingga karya ilmiah dengan topik Posisi Filogeni Orlitia borneensis dan Hieremys annandalei (TESTUDINES) Berdasarkan gen 16S rRNA ini dapat selesai dibuat sebagai salah satu syarat memperoleh gelar Sarjana Sains di Departemen Biologi Fakultas Matematika dan IPA, Institut Pertanian Bogor.
Ucapan terima ingin penulis sampaikan kepada Bapak Dr. Ir. Achmad Farajallah dan Ibu Dr. Ir. Rika Raffiudin, M.Si. selaku pembimbing karya ilmiah atas ilmu dan bimbingan yang telah diberikan kepada Bapak Dr. Ir.Iman Rusmana, M.Si. selaku Wakil Komisi Pendidikan atas saran dan perbaikannya pada karya ilmiah ini.
Penghargaan setingginya ingin penulis sampaikan kepada keluarga tercinta, Bapak, dan ade atas dukungan dan doanya. Terima kasih yang tak terhingga kepada Mamah Ayumi dan Keluarga, Mbak Novi dan keluarga atas bantuannya serta seluruh sahabat dan teman yang membuat hidup ini lebih berwarna:Eni, Uci, Alfi, Mira, Mb Gadi, Ritma, Mamy Pity dan Tria untuk keindahan persahabatan, bantuan, dan semangat yang selalu diberikan. Untuk teman seperjuangan Nina Ranty terima kasih telah jadi editor dan teman setia. Buat Suamiku tercinta, terima kasih atas kesabaran dan pengorbanannya selama ini, maaf sering meninggalkan kewajiban. Semoga Allah SWT membalas semuanya dengan kebaikan yang melimpah. Untuk Ummi tercinta karya ini kupersembahkan untukmu, semoga Allah menempatkan dirimu di tempat yang terbaik.
Penulis menyadari bahwa tulisan ini masih jauh dari sempurna. Semoga karya ilmiah ini dapat bermanfaat.
Bogor, April 2007
DAFTAR TABEL
Halaman
1 Spesies Testudines yang diperoleh dari Genbank...... 5
2 Komposisi nukleotida gen 16S rRNA pada beberapa spesies Testudines... 5
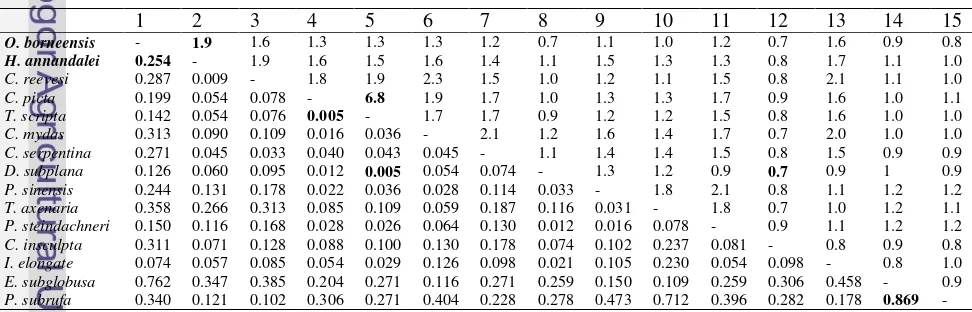
3 Jarak genetik (dibawah diagonal) dan rasio transisi terhadap transversi (ts/tv) (diatas diagonal) O. borneensis dan H. annandalei dengan 13 spesies Testudines berdasarkan gen 16S rRNA... 9
DAFTAR GAMBAR Halaman 1 O. borneensis (a) tampak plastron (b) tampak karapas... 2
2 H. annandalei (a) tampak plastron (b) tampak karapas... 2
3 C. amboinensis (a) tampak plastron (b) tampak karapas... 2
4 S. crassicolis (a) tampak plastron (b) tampak karapas... 2
5 C. dentata (a) tampak plastron (b) tampak karapas... 3
6 Posisi primer AF05 dan AF08 pada mtDNA Dogania subplana (nomor akses AF366350)... 3 7 Hasil amplifikasi O. borneensis dan H. annandalei menggunakan primer AF05 dan AF08... 4 8 Hasil perunutan DNA O. borneensis gen 16S rRNA. Angka di dalam tanda”[ ]” menunjukkan posisi nukleotida terakhir dalam baris... 5 9 Hasil perunutan DNA H. annandalei gen 16S rRNA. Angka di dalam tanda”[ ]” menunjukkan posisi nukleotida terakhir dalam baris... 6 10 Urutan nukleotida hasil penjajarangen 16S rRNA dari 15 individu ordo Testudines. Keterangan : Tanda ”–” menunjukkan gap dan tanda ”.” menunjukkan nukleotida yang sama dengan nukleotida pada baris pertama. Angka di dalam tanda”[ ]” menunjukkan posisi nukleotida terakhir dalam suatu baris... 9 11 Hasil rekontruksi pohon filogeni 15 spesies Testudines berdasarkan urutan nukleotida 16S rRNA menggunakan metode (a) ME (b) NJ (c) MP... 10
DAFTAR LAMPIRAN Halaman 1 Klasifikasi Ordo Testudines (Gaffney 1975)... 15
2 Taksonomi O. borneensis... 16
3 Taksonomi H. annandalei... 17
DAFTAR ISI
Halaman
DAFTAR TABEL...
vii
DAFTAR GAMBAR... vii
DAFTAR LAMPIRAN... vii
PENDAHULUAN Latar Belakang... 1
Tujuan... 2
Waktu dan Tempat... 2
BAHAN DAN METODE Sampel Kura-kura... 2
Ekstraksi dan Isolasi DNA... ... 3
Amplifikasi (Perbanyakan) Fragmen DNA... 3
Visualisasi Fragmen DNA... 3
Pengurutan (Sequencing) DNA Produk PCR... 4
Analisis Urutan Nukleotida dan Analisis Filogeni... 4
HASIL Hasil Amplifikasi dan Visualisasi Fragmen DNA... 4
Hasil Perunutan DNA Produk PCR... 4
Hasil Penjajaran (Alignment) DNA... 4
Hasil Analisis Filogeni... 9
PEMBAHASAN... 11
SIMPULAN DAN SARAN... 12
DAFTAR PUSTAKA... 12
DAFTAR SINGKATAN
TE : Tris HCl 20 mM, EDTA 2 mM
1xTE : Tris- HCl 20 mM, EDTA 100 mM
SE : Sarkosil 1 %, 5 mM EDTA
SDS : Sodium dodesil sulfat
CIAA : Klorofom: isoamil alkohol = 24:1
PCR : Polymerase Chain Reaction
dNTP : Campuran dari dATP, dGTP, dCTP dan dTTP dengan perbandingan yang sama
PAGE : Polyacrilamide Gel Electroforesis
1xTBE : Tris 0.5 M, asam borat 0.65 M, EDTA 0.02 M
12S rRNA : 12S ribosomal RNA
16S rRNA : 16S ribosomal RNA
tRNA : Transfer RNA
A : Adenin
G : Guanin
T : Tirosin
C : Timine
ME : Minimum Evolution
NJ : Neighbor Joining
MP : Maximum Parsimony
ts : Transisi
tv : Transversi
RIWAYAT HIDUP
Penulis dilahirkan di Bogor pada tanggal 01 Desember 1982 dari pasangan Darus Salam dan Maemunah (Alm). Penulis merupakan anak pertama dari lima bersaudara.
Lulus dari MAN I BOGOR pada tahun 2000 penulis melanjutkan studinya di Institut Pertanian Bogor, Fakultas Matematika dan Ilmu Pengetahuan Alam, Departemen Biologi pada tahun 2000 lewat jalur Undangan Seleksi Masuk IPB (USMI).
PENDAHULUAN
Latar Belakang
Kura-kura diklasifikasikan sebagai klas Reptilia subklas Anapsida dan ordo Testudines (Ernst dan Barbour 1989). Berdasarkan klasifikasi oleh Gaffney (1975) ordo Testudines atau kura-kura modern terbagi menjadi 2 subordo yaitu subordo Cryptodira dan subordo Pleurodira. Subordo Cryptodira terbagi atas 3 superfamili yaitu superfamili Testudinoidea, Chelonioidea dan Trionychoidea, sedangkan subordo Pleurodira terbagi atas dua famili yaitu Pelomedusidae dan Chelidae. Bagan klasifikasi Testudines selengkapnya dapat dilihat pada Lampiran 1.
Ordo Testudines atau kura-kura modern mempunyai ciri-ciri tengkorak masih primitif, kranium utuh tanpa lubang temporal dan tubuhnya berada dalam cangkang. Cangkang terdiri atas dua bagian yang saling berhubungan yaitu karapas (sisi bagian atas) dan plastron (sisi bagian bawah). Umumnya cangkang terbuat dari bahan tanduk yang keras tetapi pada beberapa taksa ada yang bersifat lunak yaitu pada famili Trionychidae. Subordo Cryptodira memiliki ciri-ciri mempunyai leher yang berukuran pendek dan bergerak secara vertikal sehingga kepala dan leher dapat ditarik masuk ke dalam cangkang (kecuali anggota famili Cheloniidae (penyu laut)). Sedangkan anggota Pleurodira mempunyai leher dengan ukuran yang panjang, bergerak secara lateral sehingga kepala dan leher tidak dapat ditekuk dan ditarik masuk kedalam cangkang (Lim dan Das 1999).
Famili Bataguridae merupakan anggota dari subordo Cryptodira, termasuk ke dalam
Old World Pond Turtles, memiliki keragaman fenotipe yang tinggi, hidup di daerah semi akuatik, semi terestrial dan tropik. Famili ini tersebar terutama di daerah Asia Tenggara tetapi juga di daerah Eropa Barat, Afrika Utara dan Amerika Latin (Iverson 1992). Karakter umum famili Bataguridae adalah tubuhnya terdiri atas cangkang yang keras, karapas mempunyai permukaan yang licin dan terdiri atas 5 vertebral, plastron keras, kaki depan tidak berbentuk seperti stik golf (not club-shaped), selaput diantara jari-jarinya ada atau kadang tidak ada, kaki mempunyai 4 atau 5 cakar, kepala dan leher dapat ditekuk dan ditarik masuk ke dalam cangkangnya (Lim dan Das 1999).
Orlitia borneensis dan Hieremys
annandalei merupakan anggota famili Bataguridae. Daerah penyebarannya di Asia seperti Thailand, Malaysia dan Indonesia terutama di daerah Sumatera (Ernst dan Barbour 1989, Lim dan Das 1999).
Orlitia borneensis (Malayan Giant Turtles) merupakan kura-kura air tawar terbesar di pulau Borneo dan Malaysia. Ciri-ciri O. borneensis adalah panjang karapas hingga mencapai 800 mm, ukuran kepala besar dan cangkang berwarna gelap. Habitatnya terutama di sungai yang besar dan daerah-daerah air tawar yang mempunyai ukuran yang luas dan termasuk ke dalam omnivora (Lim dan Das 1999). Status menurut IUCN tahun 2000 termasuk fauna yang dilindungi. Taksonomi O. borneensis dapat dilihat pada Lampiran 2.
Hieremys annandalei (Yellow-headed Temple Turtles) memiliki ciri-ciri karapas berwarna gelap kadang dengan garis punggung berwarna orange. Panjang karapas dapat mencapai 506 mm, habitat terutama di sungai, kubangan dan rawa dan termasuk ke dalam herbivora (Lim dan Das 1999). Status menurut IUCN tahun 2000 termasuk ke dalam fauna yang dilindungi. Taksonomi H. annandalei dapat dilihat pada Lampiran 3.
Seiring dengan kemajuan bioteknologi, pengelompokan kura-kura Bataguridal (Bataguridae, subfamili Batagurinae, famili Testudinidae) masih menjadi perdebatan. Hal ini disebabkan karena perbedaan karakter yang diamati. McDowell (1964) berdasarkan karakter tengkorak meletakkan kura-kura Bataguridal kedalam empat kelompok (complex) yaitu Hardella complex (Hardella, Morenia dan Geoclemys), Orlitia complex
(Orlitia dan Siebenrockiella), Batagur complex
Emydidae menempatkan dua spesies Emydidae yang diteliti sebagai outgroup dari spesies-spesies Bataguridae. Gaffney dan Meylan (1988) berdasarkan data osteologi meletakkan kura-kura Bataguridal ke dalam status famili Bataguridae dengan mengklasifikasikannya menjadi dua subfamili yaitu Batagurinae (termasuk Batagur, Hardella, dan Orlitia complex ( klasifikasi McDowell, 1964)) dan Geomydinae (Geomyda complex (klasifikasi McDowell, 1964)). Ernst dan Barbour (1989) berdasarkan morfologi tengkorak mengklasifikasikan kura-kura Bataguridal ke dalam famili Emydidae. Iverson (1992) berdasarkan penyebaran kura-kura di dunia mengklasifikasikan kura-kura Bataguridal ke dalam famili Emydidae.
Genom mitokondria sangat populer digunakan sebagai penanda molekuler dalam analisis filogeni, mempelajari evolusi, struktur populasi dan aliran genetik, hibridisasi, biogeografi dan berbagai fenomena mikro dan makro evolusi hewan. Genom mitokondria diwariskan secara maternal, memiliki laju evolusi yang cepat dan tanpa intron. Struktur organisasi mtDNA bersifat stabil, sehingga mtDNA hewan memiliki jumlah gen dan ukuran yang sama, yakni terdiri atas 13 gen yang menyandikan protein (terlibat dalam respirasi sel), 2 gen menyandikan rRNA (12S dan 16S), 22 gen menyandikan tRNA dan beberapa daerah lain yang tidak menyandikan protein D-loop (Control Region) dan daerah
intergenic (Moritz et al. 1987) (Lampiran 4). Meski demikian, gen-gen dalam mtDNA memiliki laju evolusi yang berbeda-beda, evolusi tRNA dan rRNA mitokondria relatif lambat jika dibandingkan dengan gen lainnya. Karena itu tRNA dan rRNA mitokondria banyak digunakan untuk mempelajari hubungan filogeni pada tingkat interspesies (Randi 2000).
Zardoya dan Meyer (1998) telah menggunakan genom mitokondria untuk analisis filogeni diantara kelompok vertebrata. Honda et al (2002) juga menggunakan runutan mtDNA untuk menentukan hubungan filogeni Asian Box Turtles.
Tujuan
Penelitian ini bertujuan untuk menentukan posisi filogeni O. borneensis dan
H. annandalei (Testudines) berdasarkan gen 16S rRNA.
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Januari sampai dengan Juni 2006 bertempat di Laboratorium Zoologi Departemen Biologi Fakultas MIPA, Institut Pertanian Bogor, Bogor.
BAHAN DAN METODE Sampel Kura-kura
Sampel yang digunakan pada penelitian ini adalah 4 spesimen ujung ekor dan 1 spesimen karapas O. borneensis (Gambar 1), 1 spesimen ujung ekor H. annandalei
(Gambar 2), 2 spesimen ujung ekor Cuora amboinensis (Gambar 3), 1 spesimen ujung ekor Siebenrockiella crassicolis (Gambar 4) dan 1 spesimen ujung ekor Cyclemys dentata
(Gambar 5). Semua sampel yang digunakan merupakan koleksi Achmad Farajallah (Lab. Zoologi Dept. Biologi FMIPA IPB, Bogor ) yang diambil dari Kalimantan Tengah.
(a) (b) Gambar 1 O. borneensis (a) tampak plastron (b) tampak
karapas.
(a) (b)
Gambar 2 H. annandalei (a) tampak plastron (b) tampak karapas.
(a) (b)
(a) (b)
Gambar 4 S. crassicolis (a) tampak plastron (b) tampak karapas.
(a) (b)
Gambar 5 C. dentata (a) tampak plastron (b) tampak karapas.
Ekstraksi dan Isolasi DNA
Isolasi DNA total dilakukan mengikuti metode Farajallah (2002). Sampel dalam alkohol 70% dicuci dengan 500 µl bufer TE sebanyak 2 kali. Sampel otot yang telah dicuci kemudian dihancurkan menggunakan gunting dalam bufer 1X STE 300 µl dan SE 3%. Penghancuran protein dilakukan dengan menambahkan Proteinase K 0.3 mg/mL dan SDS 10% yang kemudian diinkubasi pada suhu 55 0C sambil dikocok pelan selama 2,5 jam.
Material DNA dipisahkan dari material organik lainnya dengan metode ekstraksi fenol, yaitu dengan menambahkan larutan fenol 1X volume, NaCl 5 M 1/10 X volume,
dan CIAA 1X volume. Campuran di atas kemudian dikocok pelan selama 2 jam pada suhu ruang. Bahan organik yang masuk ke fase fenol dipisahkan dari fase air menggunakan sentrifus pada kecepatan 5000 rpm selama 2 menit. Fase air yang terbentuk pada lapisan atas dipindahakan ke tabung baru kemudian DNA dimurnikan dengan teknik pengendapan alkohol yaitu dengan cara menambahkan NaCl 5M 1/10 X volume dan
alkohol 2X volume. Campuran diinkubasi pada suhu 4 0C selama satu malam kemudian molekul-molekul DNA diendapkan dengan
sentrifus 7000 rpm selama 5 menit. Endapan DNA yang diperoleh dicuci dengan alkohol 70%. Setelah alkohol pencuci dibuang dan dievaporasi dalam ruang vakum, molekul-molekul DNA disuspensikan dalam bufer TE 80% dan disimpan dalam freezer untuk digunakan lebih lanjut.
Amplifikasi (Perbanyakan) Fragman DNA
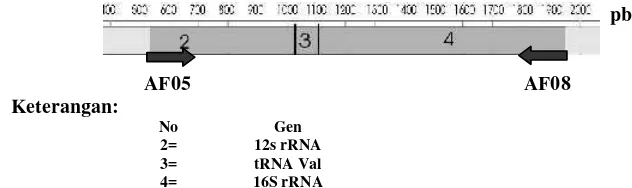
Perbanyakan fragmen DNA dilakukan secara in vitro dengan metode PCR menggunakan mesin Thermo Cyler TaKaRa MP4. Primer yang digunakan yaitu AF05 (5`- ACT GGG ATT AGA TAC CCC ACT AT-3`) dan AF08 (5`- ATG TTT TTG GTA AAC AGG CG-3`).
Primer AF05 menempel pada bagian akhir gen 16S rRNA dan AF08 menempel pada bagian tengah gen 16S rRNA. Kedua primer tersebut mengapit ruas DNA sebesar 1474 bp atau setara dengan posisi nukleotida 505 sampai 1978 genom mitokondria Dogania subplana (Farajallah 2002) (Gambar 6). Komposisi 25 µl reaksi PCR adalah DNA cetakan 10-100 ng, Taq Polymerase (Promega) 0.75 Unit beserta bufernya, Mg Cl2
5 nM, dNTP 600 pM dan masing-masing primer 25 nM.
Perbanyakan DNA dilakukan pada kondisi suhu denaturasi awal 94 0C selama 5 menit. Selanjutnya dilakukan 30 siklus perbanyakan pada suhu denaturasi 94 0C selama 1 menit, suhu penempelan (annealing)
50 0C selama 2 menit dan suhu pemanjangan (elongasi) 70 0C selama 2 menit dan diakhiri dengan pemanjangan akhir pada suhu 70 0C selama 10 menit.
Visualisasi Fragmen DNA
Produk PCR dimigrasikan menggunakan PAGE 5% dalam buffer 1XTBE. Elektroforesis dijalankan pada kondisi 165 mV selama 75 menit. Selanjutnya gel diwarnai dengan pewarnaan sensitif perak (Tegelstrom 1986).
pb
AF05 AF08 Keterangan:
No Gen
Gambar 6 Posisi primer AF05 dan AF08 pada mtDNA Dogania subplana (nomor akses AF366350).
Perunutan (Sequencing) DNA produk PCR
Perunutan DNA merupakan tahap akhir untuk memperoleh data urutan nukleotida dari fragmen DNA hasil amplifikasi. Perunutan DNA dilakukan menggunakan jasa Biologi Molekuler CHAROEN POPKHAND Indonesia menggunakan metode Big Dye Determination dalam mesin ABI PRISM. Produk PCR yang berupa pita tunggal (single band) dijadikan sampel dalam reaksi perunutan nukleotida. Perunutan nukleotida dilakukan menggunakan primer AF08.
Analisis Urutan Nukleotida dan Analisis Filogeni
Urutan nukleotida diedit secara manual menggunakan pogram Genetyx win versi 4.0.1 (Genetyx Software Jepang) berdasarkan pada elektropherogram. Urutan nukleotida dari semua sampel kemudian disejajarkan bersama urutan nukleotida beberapa spesies Testudines yang terdapat di Genbank dengan nomor akses terdapat pada Tabel 1. Proses penjajaran dilakukan menggunakan program Clustal W 1.8 kemudian diedit secara manual. Komposisi nukleotida, tipe subsitusi transisi dan transversi (ts/tv) dihitung menggunakan program MEGA 3.0 (Kumar et al. 2001). Penghitungan jarak genetik dan konstruksi pohon filogeni dilakukan menggunakan program MEGA 3.0 (Kumar et al. 2001) dengan metode ME, MP dan NJ dengan
bootstrap 1000X. Analisis bootstrap 1000X dilakukan untuk menguji kestabilan posisi filogeni dari klad tertentu pada pohon filogeni yang dihasilkan.
HASIL
Hasil Amplifikasi dan Visualisasi Fragmen DNA
Hasil amplifikasi O. borneensis dan H. annandalei menggunakan primer AF05 dan AF08 menunjukkan fragmen DNA berukuran sekitar 1500 bp (Gambar 7). Hal
ini menunjukkan panjang DNA yang dihasilkan berada di antara gen 12S dan 16S rRNA. Amplifikasi C. amboinensis, S.
crassicolis dan C. dentata tidak
menghasilkan pita pada gel elektoforesis sehingga dari ketiga spesies ini tidak dilakukan perunutan nukleotida pada lembaga Biologi Molekuler CHAROEN POPKHAND.
Hasil Perunutan (Sequencing) DNA produk PCR
Perunutan DNA dilakukan dari arah primer AF08. Setelah diedit menghasilkan panjang DNA sebesar 681 nt untuk O. borneensis (Gambar 8) dan 673 nt untuk H. annandalei (Gambar 9).
Hasil Penjajaran (Alignment) DNA
Setelah disejajarkan, panjang DNA yang diperbandingkan sebesar 695 nt untuk
O. borneensis dan 696 nt untuk H. Annandalei. Terdiri dari 317 basa yang sama (conserved) dan 359 basa yang berbeda (variabel). Dari 359 basa yang berbeda terdapat 211 basa parsimony dan 143 basa tunggal (singleton). Untuk selanjutnya, basa yang digunakan untuk analisis merupakan basa yang bersifat parsimony (Gambar 10).
Komposisi nukleotida H. annandalei
didominasi oleh nukleotida T sebesar 42.0%, kemudian berturut-turut diikuti oleh nukleotida G (21.7%), A (21.4%) dan C (14.9%). Nilai perbandingan antara A/T dan G/C yaitu 63.4% :36.6%. Komposisi 1 2 M
1500 bp 1000 bp 400 bp
100 bp
Gambar 7 Hasil amplifikasi O. borneensis dan H.
annandalei menggunakan primer AF05 dan
nukleotida O. borneensis didominasi oleh nukleotida T sebesar 43.4%, kemudian berturut-turut diikuti oleh nukleotida G (23.3%), A (19.9%) dan C (13.4%). Nilai perbandingan antara A/T dan G/C yaitu
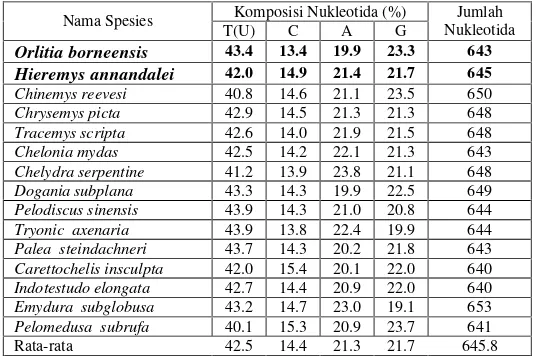
63.3% :36.7%. Dari Tabel 2 dapat dilihat bahwa sebagian besar spesies memiliki pola komposisi nukleotida yang sama yaitu T>G>A>C.
Tabel 1 Spesies Testudines yang diperoleh dari Genbank
Tabel 2 Komposisi nukleotida gen 16S rRNA pada beberapa spesies Testudines
CAATGAAACCGCTCACTTACCCACAAAAAGCAAAAATTAATCCTTATACCTTTTCATCAT [ 60] GATTTAGCCAGCACACCCAAGCAAAGACCCCTAAAGTCTGAAACCCCGAAACCAAATGAG [120] CTACTTAAAGGCAGCTACTGCACACACCCAAACCCATCTCTTGGCAAAAGAGTGGCGAGA [180] CCTCTAAGTAGAGCTGAAAAGCCTAACGAATTTCGTAATA?CT?CTTGCTCAACAAAAGA [240] ATATAAGTTCAACCTTAAACCTTCTAAAAACAACTTAAAGTGAAAAGAAAAGTTTAAGAT [300] TCATTCAATTGAGGTACAGCCCAATTGAAAAAGGATACAACCTATAACGGAGGACAAAAC [360] ACCAAAAACACAACCACCGTAGGCTTTAAAGCACCCACCACCAAAGAAAGCGTCAAAGCT [420] CCAAAATACTAAATAATAACACAAATTTTTTCCCCAAAACAACATTGAGCCATTCTACTT [480] AAATATAAGAACTAATGCTGAAATGAGTAACAAGAAGATAAAACTTCTCTAACGCGCTAG [540] CTTAAATCATAATGGACAAACCACTGATTATTAACAACTAACACTATGGTACAACAACAC [600] TTAAATCACCCTATTAAATACAACTCTTAATGGCAACACAGGAGCGCATGAAAGAAAGAG [660] TAAAATTTTAAAAAAACTACC [681]
Spesies Famili Subordo No. Akses
Chinemys reevesi Emydidae Cryptodira AY676201
Chrysemys picta Emydidae Cryptodira AF069423
Trachemys scripta Emydidae Cryptodira L28077
Chelonia mydas Cheloniidae Cryptodira AB012104
Chelydra serpentine Chelydridae Cryptodira DQ283320
Dogania subplana Trionychidae Cryptodira AF366350
Pelodiscus sinensis Trionychidae Cryptodira AY687385
Tryonyx axenaria Trionychidae Cryptodira AY583695
Palea steindachneri Trionychidae Cryptodira AY743418
Carettochelis insculpta Cheloniidae Cryptodira Puspa (2005)
Indotestudo elongata Testudinidae Cryptodira DQ656607
Emydura subglobusa Chelidae Pleurodira unpub. data
Pelomedusa subrufa Pelomedusidae Pleurodira NC001947
Komposisi Nukleotida (%) Nama Spesies
T(U) C A G
Jumlah Nukleotida
Orlitia borneensis 43.4 13.4 19.9 23.3 643
Hieremys annandalei 42.0 14.9 21.4 21.7 645
Chinemys reevesi 40.8 14.6 21.1 23.5 650
Chrysemys picta 42.9 14.5 21.3 21.3 648
Tracemys scripta 42.6 14.0 21.9 21.5 648
Chelonia mydas 42.5 14.2 22.1 21.3 643
Chelydra serpentine 41.2 13.9 23.8 21.1 648
Dogania subplana 43.3 14.3 19.9 22.5 649
Pelodiscus sinensis 43.9 14.3 21.0 20.8 644
Tryonic axenaria 43.9 13.8 22.4 19.9 644
Palea steindachneri 43.7 14.3 20.2 21.8 643
Carettochelis insculpta 42.0 15.4 20.1 22.0 640
Indotestudo elongata 42.7 14.4 20.9 22.0 640
Emydura subglobusa 43.2 14.7 23.0 19.1 653
Pelomedusa subrufa 40.1 15.3 20.9 23.7 641
Rata-rata 42.5 14.4 21.3 21.7 645.8
Gambar 8 Hasil perunutan DNA O. borneensis gen 16S rRNA. Angka di dalam tanda”[ ]” menunjukkan posisi nukleotida terakhir dalam baris.
CTTAGCTTTAAAAAAGCTCAAGAATTACCCTTTATTCTCTTTCGCATCATGATTTTGTCA [ 60] GCTCATTCGAACAAAGAGCCCTAAAGTCCGAACCCCCGAAACCAAGTGAGCTACTTAGAG [120] GCAGCCACACCAAGGCTAAATCCGTCTCTGTGGCAAAAGAGTGGAAAGACCCCTAAGTAG [180] AGGTGAAAATCCTAACGAACTTGGTGATAGCCTGGTTGTTCAATAAAAGAATATAAGTTC [240] AACCTTAAACCTTCTAAAAACAGCCTAAAGTATGAAAGAAAAGTTTAAGATTCATTCAAT [300] TGAGGTACAGCCCGATTGAAAAAGGACACAACCTAATAATGGAGGATAAAACATTAAAGC [360] ACAACAACCGTAGACCTTAAAGCAGCCATCACCAAAGAAAGCGTCAAAGCTCACCTTAAT [420] AATTAAATAATAACATAAAAATTTTACCCCAAAAAACATTGAACCACTCTACCCAAATAG [480] AAGAACTAATGGTAAAATGAGTAACAAGAAGACAAAACTTCTCTAACGCGCCAGCTTAAG [540] TCATTATAGACAAACTACTGACTATTAACAATTAACACTATGAAACAAAAATACTTAAAA [600] CACCCTATAAACCTAACTGTTAACGGAACACAGGAGCGCACACAAGAAAGAGTAAAATTG [660] TAAAAGAACTAGC [673]
Gambar 9 Hasil perunutan DNA H. annandalei gen 16S rRNA. Angka di dalam tanda”[ ]” menunjukkan posisi nukleotida terakhir dalam baris.
O. borneensis TTTCTTT-TGTGCGCTCCTGTGTTCCGTTAACAGTTAGGTT-TAATAGGGTGATTTAAG- [ 60]
H. annandalei ...G-...-..-...T...- [ 60]
C. eevesi ...-A...GG...C-..-...G...AA [ 60] C. picta ...-AC...GG...TA..--..-..T....T...- [ 60]
T. scripta ...-A...CGG...TA..--..-..T....T...- [ 60]
C. mydas ...G-.A...GG...T....-..-..T...ATG..T..- [ 60]
C. erpentine ...-A.A...AT...GG...T...A-..-..T..C.TG...- [ 60]
D. subplana ...G-.T.A...GG...TATG.-..-..T.T..TG..T..- [ 60]
P. sinensis ...ATCC.A...GG...TATG.-..-..T.T.ATG..T.A- [ 60] T. axenaria ...AT.T.A...GG...T.TG.-..-..TAT.ATG..T.A- [ 60] P. steindachneri...GT.T.A...G...GG...TATG.-..-..T.T.TTG..T..- [ 60]
C. insculpta ...AG-.A...GG.G...A..T....-..-..T.-.T.G....T- [ 60] I. elongata ...A.A...GG...TA...-..-....A..T...- [ 60]
E. subglobusa ...G-..C..AT...GG...TT..C-..-....A.TTG...- [ 60]
P. subrufa ...GG-.CA..A...GG...G...GA...G.T...T.T.TTG..T..- [ 60]
O. borneensis TGTTGTTGTTTC--ATA-GTGT-TAGTTGTTAATAATCAGTGGTTTGTCTATT-ATGATT [120] H. annandalei .A..T...--...-....-..A...G...A...A-....C. [120] C. reevesi ...-T-...GAG..--G...G...A...G..-T...C. [120] C. picta ...AT...-..A.-.G...A..C...-... [120]
T. scripta ...AT...-T.A.-..A...A....A...-... [120]
C. mydas .A...G..-TT...ATAA.--...A...-... [120]
C. serpentine ...G..-TT...A-AA.-...A...G..-G... [120]
D. subplana ..C...-TT..---G..--G...A....A...G..-C... [120]
P. sinensis .A...C-TT...--GA---G...A...G..-C... [120]
T. axenaria .T..A..A.AC-TT...--GA---G...A...G..-C... [120]
P. steindachneri.AG.A...-TT..---.-.G.C...A...G..-C... [120]
C. insculpta .C...TG..-TT...---A---G..C...A.G.GT...G..AC....C [120] I. elongata ....A...G.A-TT...---A.A.GAC...A...G..-... [120]
E. subglobusa .T...TG...CT...A..-.-...A..C...GG-C... [120]
P. subrufa ....T...--.GGT...---A.-.--.C...A...CT...GAA-C.A.CG [120] O. borneensis TAAGCTAGCGCG-TTAGAGAAGTTTTAT-CTTCTTGTTACTCATTTTACCATTAGTTCTT [180] H. annandalei ...G...-...G.-...G... [180]
C. reevesi ...-...-...G...C. [180] C. picta ...-CA...-.C...G... [180]
T. scripta ....-.G...-.A...-.-...G....G... [180]
C. mydas ..G...G....A-...G.-...T...A... [180]
C. serpentine ...AT..A-.A...-..-...G...A... [180]
D. subplana ...TA-GG...G....AT.T...G...A... [180]
P. sinensis ...G...TA-.-.A...GAAG..TGT...G....G... [180]
T. axenaria ...G...TA-.-.A...GAAG.ATAT...G....G... [180]
P. steindachneri...G...TA---.T...GAAG..T.T...G...A.T... [180]
C. insculpta .GG...T....AA...G..G.A-GT-...G... [180]
I. elongata ...-...G.C...G... [180]
E. subglobusa ...AT..--...TAA-...A...G...A... [180]
P. subrufa ...T...--.T...AG..----...A...G....G... [180]
O. borneensis CTATTTAGGTAGAATGGCTCAATG--TTGTTTTGGGG-AAAAAA-TTTG-TGTTATTATT [240] H. annandalei ...G...G...T...--..T...-....--T....T...TA... [240]
C. picta ....GGG...G...G..--..A...-....-.T....G...TA...A.... [240]
T. scripta ....AGG...G.A--..A...-....-GT....G...TA...G.... [240]
C. mydas ...G.A...A...A--...-....-.-....A..G-A...A.... [240]
C. serpentine ....AGGA...A...--..A...-...A-.G....T.G.--...A.... [240]
D. subplana ...T...G..TT..T...GA...-GT.G.-G.A.--...A.... [240]
P. sinensis ...TA...G..--...GA..A-GT...-T.A.--...A.... [240]
T. axenaria ...TA...G..--...GA..A-GT...-T.A.--...A.... [240]
P. steindachneri....A.T...G..--...GA...-GT.G..-.G.--....GA.... [240]
C. insculpta ...GTTA...A..C..G.TT--...GA...TGC.GTGG.A----....A.... [240]
I. elongata ...G...A...G..T--...-....-.-....A...AA....G... [240]
E. subglobusa ....G.G...A...G.AG-AG....A...AG.T..-.T...--.A...A.... [240]
P. subrufa ...GG..A...A...TG--...G..AATGGGC..AGG.T---..GA.... [240]
O. borneensis TA-GTATTTT----GGA---GCTTTGACGCTTTCTTT-GGTGGTGGGTGCTTTAAAG [300] H. annandalei ..-A.TA..A-AGGT..---...-....A...C...G. [300] C. reevesi .--.--A.GG-GTG..GGGGGGG...-....A...C...G. [300] C. picta ..TA..A.GG--T-.A---...T.A...C... [300]
T. scripta ..CA..A.AG--T-..---...-AA...C...G. [300] C. mydas .--..TG..A-GT--..---...C...-.AC...C... [300]
C. serpentine ..--.TA..A-AT--.G---...-A...C...G. [300] D. subplana .--..-G.GG-AT-T..---...-...C... [300]
P. sinensis .--..-A.AG-GT-...---...-...C...GG. [300] T. axenaria .--..-A.GG-AT-...---...-...C...G. [300] P. steindachneri.--...A.GG--T-A.G---...T...-A...A...C...G. [300] C. insculpta .--..T....--T--..---...-.T...C...G. [300] I. elongata .--..TG..A-GT--..A---...T...-....A...C... [300]
E. subglobusa ..-T.-A.GG--T-TT.---...A...A...TAA...C...G. [300] P. subrufa .---.-G-GATATAT..---...A...A....A-A...A...C...G. [300] O. borneensis CCTACGGTGGT-TGTGTTTTTGGT-GTTTTGTCCTCCGTT-A-TAGGTTGTATCCTTTTT [360] H. annandalei T...T..-....C...-AA.-...A...A..-.T...G... [360]
C. reevesi ...T..-...A....-AA.-A...G.T... [360]
C. picta ...T--.T.A....-...-...TA..--T...G... [360]
T. scripta ...A.TAA.A.A....-...-...A..--T...G... [360]
C. mydas ...AA.-AA...G-AA.-...A..--T...G... [360]
C. serpentine ...A..A-G.A...-.A.-...T.A..A..--T...T... [360]
D. subplana ..C..TT..AGT.--T.G.AGT..-..C..T....T.A..--T...G... [360]
P. sinensis ...TA...TAAAT.G..G.A.-A....A....T....--T...G... [360]
T. axenaria ...TAA.AT.AAT.G..GA..-...A....T....--T...G... [360]
P. steindachneri...T..A.----A....GT..A..C..A....T.A..--T...T..G... [360]
C. insculpta ...T...T.GGT..GA..-.T.-.C....C...T.A..--G...T... [360]
I. elongata ...AA.--A...-...-...A...A..--T...G... [360]
E. subglobusa ...T.GTA.TAT...A.A-AA--A....T...TA.G--T.T...T...A. [360] P. subrufa ...TA.TTA---A...GCA.-AG..G..T....A..--G.G...T...C..C. [360] O. borneensis CAATTGGGCTGTACCTCAATTGAATGAATCTTAAAC-TTTTCTTTT---TA--CTTTAGG [420] H. annandalei ....C...-...CA--..--... [420]
C. reevesi ...C...A...-...---..--....G.. [420]
C. picta ...C...AT...-...GG-..AG... [420]
T. scripta ...C.G...AT...-...GGG..AA... [420]
C. mydas ...C..A...G...AT...T-..C....-AAG---TA...T. [420] C. serpentine ...C...AT...T-..C..G..AAA-A.TA....G.. [420]
D. subplana ...T...CT...T.C..C....T-..G..A.G.----.ACTA....- [420] P. sinensis ...T...CT...AGC..C....A-...----.ACT...G.- [420] T. axenaria ...T...C...CT...AGC..C....AA...GG----ACT...G.- [420] P. steindachneri...AA...T...GC..C...-...G.G----ACT....AA [420] C. insculpta ...A...CT...T.AT...T-..C..AG..GGC.TAAGG.ATTT [420] I. elongata ...C...G.AT...T-C...---TA....GT. [420] E. subglobusa T.G...C.T.C.A...ATC...GT-C.C...-.GGGG.GAA....GTT [420]
O. borneensis TTGTTTT-TAGAAGGTTTAAGGTTGAACTTA-TATTCTTTTGT-TGAGCAACCAGCTATT [480] H. annandalei C...-...-...A.-...C [480] C. reevesi G...-...-A...A.-...C [480] C. picta ...-...AA...-C....A...A.-...C [480] T. scripta ...-...A...-C....A...A.-...C [480] C. mydas ...-...GGT...A.-...A.-...C [480] C. serpentine ...-...G..A...-A...A.-...C [480] D. subplana ...-.G.G-T...G...G.C..AC.C...A..A.-...C [480] P. sinensis ...-....TT...G...G....-..C...A.-...C [480] T. axenaria ...-...T...G...G....-..C...A.-...C [480] P. steindachneri...-.G...T...G...G....-..C...A.-...C [480] C. insculpta G..CC..GAT.G.TA...G-...G...A.-..G...C [480] I. elongata A...G...-...A.-...C [480] E. subglobusa G...C.GT...GG.A...A...T-...CC-...C [480] P. subrufa A...GGGG.G.GC.A...A...AT-...A.-...C [480] O. borneensis ACCAAGTTCGTT-AGGCTTTTCACCTCTACTTAGAGGTCTTTCCACTCTTTTGCCACAGA [540]
H. annandalei ...-...A...G... [540]
C. reevesi ...-...T... [540]
C. picta ...A.-...T...T...C... [540]
T. scripta ...A.-...T...T... [540]
C. mydas ...G....-...T... [540]
C. serpentine ...-...T...TT...C... [540]
D. subplana ..T.G..CG...T...A...C....GT... [540]
P. sinensis ..T.G...-...A...AT... [540]
T. axenaria ..T.G...-...A...AT.T... [540]
P. steindachneri..T.G...-...G...AT...GT... [540]
C. insculpta ...G.-...G...T... [540]
I. elongata ...G...A.-...T...T... [540]
E. subglobusa ..T...-...T...T.A... [540]
P. subrufa ..T.GA...-T...T...AG...C... [540]
O. borneensis GATGGGTTT-GGGTGTG---TGAAGT-AGCTGCCTTTAAGTAGCTCATTTGGTTTCG [600] H. annandalei ..C..A..--TA.CC.---..GT..-G...C...C... [600]
C. reevesi ..C..A..G-TA.CC.---...T.C-G...C... [600]
C. picta ..C..A..--TA.CC..---...TA.TG...C... [600]
T. scripta ..C..A..--TA.CC..---...TA.TG...C... [600]
C. mydas ..C..A..--TA.CC..G---...TA.TG...C... [600]
C. serpentine ..C..A..A-TA.CC.TA----G...TA.TG..C...C... [600]
D. subplana ....-AGGG-TA.CT.TACG--GG..T..TT...C..A...G.. [600]
P. sinensis ....-A..G-TA.CT.TGG----ATT.A.TG...G...C..A... [600]
T. axenaria ....-A..A-TA.CT.TGG----ATT.-.TG...G...C..A... [600]
P. steindachneri....-AC.A-CA.CT.TAGTGTGG.TG..CG...C..A... [600]
C. insculpta ..C..T..AATA.CCCTG----A...T..AG...T..C...C... [600]
I. elongata ..C..A..--TT.CC.---.GT.A-G...C... [600]
E. subglobusa ..C..T.AAGAA.CTCT---..TTA.A...T.A...C..A... [600]
P. subrufa ..C..A..G-TAACT.T---..CTAA-..T...A...C... [600]
O. borneensis GGGTTTCAGACTTTAGGGCTCTTTGCTTG---GGTGTGCTGGCTAAATCATGATGCAAAA [660] H. annandalei ...G...G...T.C.---AA..A....A.A...G... [660]
C. reevesi ...G...T--A..A... [660]
C. picta ...G..T...TT...A---AT...T...G.. [660]
T. scripta ...G..T...TT...A---A...T...G. [660] C. mydas ..AG...TT.G...---A...A.T...G. [660] C. serpentine ...G...TT...---A..A.A...A...G. [660] D. subplana ....A.T.A...TT...AGGTAT.A.T...G. [660] P. sinensis ...T.A....C..TT...AGTTAT..--...A....G... [660]
T. axenaria ..A...T.A....A..TT...AGTTAT..--...A... [660]
P. steindachneri....CCT...A..TT...AGGT.T.A--...A....G... [660]
C. insculpta ..A.AA..A...TT.G...G--A-...A...A... [660]
I. elongata ...C...T...---..G...T... [660]
E. subglobusa ..A.C...T...TC...A--A..A....A...T... [660]
O. borneensis GGTATAAGGATTAATTTTTGCTTTTTGTGGGTAAG-- [697] H. annandalei ..A....A.G...C...TAAA.CT.AG- [697] C. reevesi ...C...A...C.T..-- [697] C. picta ...G...C...A.T.C.T..-- [697] T. scripta ...G...C...T.C.T..-- [697] C. mydas ...G...C...C.T..-- [697] C. serpentine ...G..G...C...A...A...C.T..G- [697] D. subplana ....C...AG...C...A...T.T.C.TGA-- [697] P. sinensis ....C....T....CC...T...C.T..-- [697] T. axenaria ....C....G...C...T.T.C.T.A-- [697] P. steindachneri....C....G...C....-C....T.T.C.TTA-- [697] C. insculpta ....C....T...G.C...A..GT.TCA.TT.G- [697] I. elongata ...G....GC...AG..C.T.--- [697] E. subglobusa ....C....T...C...T.T.C.T..G- [697] P. subrufa ....C....G..T..C...T...C.TG.-- [697]
Gambar 10 Urutan nukleotida hasil penjajarangen 16S rRNA dari 15 individu ordo Testudines. Keterangan : Tanda ”–” menunjukkan gap dan tanda ”.” menunjukkan nukleotida yang sama dengan nukleotida pada baris pertama. Angka di dalam tanda”[ ]” menunjukkan posisi nukleotida terakhir dalam baris.
Hasil Analisis Filogeni
Jarak genetik dari gen 16S rRNA berkisar antara 0.005 sampai 0.869. Nilai jarak genetik terkecil ditemukan antara
Trachemys scripta dan Chrysemys picta dan antara D. subplana dan T. scripta sedangkan nilai jarak genetik terbesar ditemukan antara
Pelomedusa subrufa dan Emydura
subglobusa. Rata-rata rasio transisi terhadap transversi (ts/tv) adalah 1.308 dengan selang antara 0.67 sampai 6.75. Perbandingan nilai ts/tv terbesar ditemukan antara C. picta dan
T. scripta sedangkan nilai ts/tv terkecil ditemukan antara D. subplana dan
Carettochelis insculpta (Tabel 3).
Analisis filogeni hasil penjajaran urutan nukleotida Gen 16S rRNA O. borneensis
dan H. annandaei dilakukan menggunakan metode Kimura-2-parameter yaitu metode yang menggunakan parameter transisi dan
transversi untuk menghitung persentase besarnya perbedaan jarak genetik antar sampel (Nei dan Kumar 2000). Berdasarkan metodeME, MPdan NJ dengan bootstrap
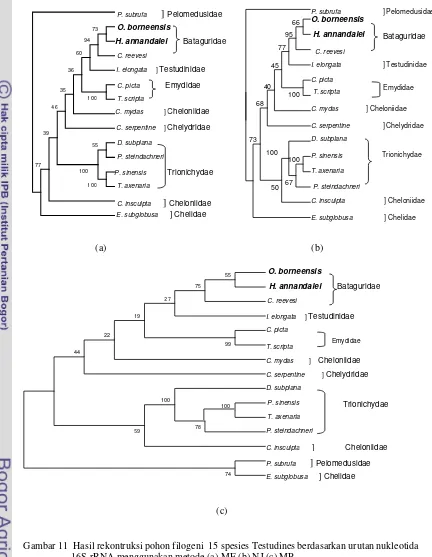
1000X dihasilkan pohon filogeni dengan topologi yang sama, menunjukkan O. borneensis dan H. annandalei berada satu cabang dengan Chinemys reevesi (famili Bataguridae) sedangkan Chrysemys picta
dan T. scripta (famili Emydidae) membentuk cabang tersendiri memisahkan diri dari famili Bataguridae. Spesies
Indotestudo elongata (famili Testudinidae) berada di dalam percabangan antara famili Bataguridae dan famili Emydidae. Spesies
P. subrufa (famili Pelomedusidae) dan spesies E. subglobusa (famili Chelidae) membentuk outgroup karena kedua spesies ini merupakan anggota subordo Pleurodira (Gambar 11).
Tabel 3 Jarak genetik (di bawah diagonal) dan rasio transisi terhadap transversi (ts/tv) (di atas diagonal) O. borneensis dan H. annandalei dengan 13 spesies Testudines berdasarkan gen 16S rRNA
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
O. borneensis - 1.9 1.6 1.3 1.3 1.3 1.2 0.7 1.1 1.0 1.2 0.7 1.6 0.9 0.8
H. annandalei 0.254 - 1.9 1.6 1.5 1.6 1.4 1.1 1.5 1.3 1.3 0.8 1.7 1.1 1.0
C. reevesi 0.287 0.009 - 1.8 1.9 2.3 1.5 1.0 1.2 1.1 1.5 0.8 2.1 1.1 1.0
C. picta 0.199 0.054 0.078 - 6.8 1.9 1.7 1.0 1.3 1.3 1.7 0.9 1.6 1.0 1.1
T. scripta 0.142 0.054 0.076 0.005 - 1.7 1.7 0.9 1.2 1.2 1.5 0.8 1.6 1.0 1.0
C. mydas 0.313 0.090 0.109 0.016 0.036 - 2.1 1.2 1.6 1.4 1.7 0.7 2.0 1.0 1.0
C. serpentina 0.271 0.045 0.033 0.040 0.043 0.045 - 1.1 1.4 1.4 1.5 0.8 1.5 0.9 0.9
D. subplana 0.126 0.060 0.095 0.012 0.005 0.054 0.074 - 1.3 1.2 0.9 0.7 0.9 1 0.9
P. sinensis 0.244 0.131 0.178 0.022 0.036 0.028 0.114 0.033 - 1.8 2.1 0.8 1.1 1.2 1.2
T. axenaria 0.358 0.266 0.313 0.085 0.109 0.059 0.187 0.116 0.031 - 1.8 0.7 1.0 1.2 1.1
P. steindachneri 0.150 0.116 0.168 0.028 0.026 0.064 0.130 0.012 0.016 0.078 - 0.9 1.1 1.2 1.2
C. insculpta 0.311 0.071 0.128 0.088 0.100 0.130 0.178 0.074 0.102 0.237 0.081 - 0.8 0.9 0.8
I. elongate 0.074 0.057 0.085 0.054 0.029 0.126 0.098 0.021 0.105 0.230 0.054 0.098 - 0.8 1.0
E. subglobusa 0.762 0.347 0.385 0.204 0.271 0.116 0.271 0.259 0.150 0.109 0.259 0.306 0.458 - 0.9
(a) (b)
(c)
Gambar 11 Hasil rekontruksi pohon filogeni 15 spesies Testudines berdasarkan urutan nukleotida 16S rRNA menggunakan metode (a) ME (b) NJ (c) MP.
H. annandalei Bataguridae
C. reevesi
I. elongata ] Testudinidae
C. picta
T. scripta Emydidae
C. mydas ] Cheloniidae
C. serpentine ] Chelydridae
D. subplana
Trionichydae
P. sinensis
T. axenaria
P. steindachneri
C. insculpta ] Cheloniidae
P. subrufa ] Pelomedusidae
E. subglobusa ] Chelidae 100 78 100 99 55 75 59 27 19 22 44 74 O. borneensis P. subrufa ] Pelomedusidae
O. borneensis
H. annandalei Bataguridae
C. reevesi
I. elongata ] Testudinidae
C. picta Emydidae
T. scripta
C. mydas ] Cheloniidae
C. serpentine ] Chelydridae
D. subplana
P. steindachneri
P. sinensis Trionichydae
T. axenaria
C. insculpta ] Cheloniidae
E. subglobusa ] Chelidae 100 55 100 100 73 94 60 36 35 46 39 77
P. subrufa ] Pelomedusidae
O. borneensis
Bataguridae H. annandalei
C. reevesi
I. elongata ] Testudinidae
C. picta
Emydidae T. scripta
C. mydas ] Cheloniidae
C. serpentine ] Chelydridae
D. subplana
Trionichydae P. sinensis T. axenaria P. steindachneri
C. insculpta ] Cheloniidae
PEMBAHASAN
Amplifikasi O. borneensis dan H. annandalei menggunakan primer AF05 dan AF08 menghasilkan fragmen DNA berukuran sekitar 1500 bp (Gambar 7). Hal ini menunjukkan bahwa hasil amplifikasi tersebut berasal dari mtDNA yang menjadi target amplifikasi karena sesuai dengan panjang fragmen yang diharapkan saat mendesain primer berdasarkan urutan nukleotida mtDNA D. subplana famili Trionychidae (Farajallah 2002).
Rasio transisi terhadap transversi (ts/tv) 16S rRNA berkisar antara 0.67 sampai dengan 6.75. Rasio ts/tv yang mendekati 1 berkisar 23.8% sedangkan rasio ts/tv yang melebihi 1 berkisar 76.2%. Hal ini menunjukkan bahwa terjadi substitusi transisi dan subsitusi transversi tetapi substitusi transversi lebih jarang terjadi dibandingkan dengan substitusi transisi. Transisi merupakan penggantian satu basa purin oleh satu basa purin yang lain (A/G), atau penggantian satu basa pirimidin oleh basa pirimidin yang lain (C/T), sedangkan transversi merupakan penggantian satu basa purin oleh pirimidin atau penggantian basa pirimidin oleh purin (Nei 1987). Randi (2000) menyatakan bahwa untuk ruas-ruas gen yang tidak menyandikan protein rasio ts/tv cenderung mendekati angka 1.
Perkiraan jarak genetik untuk analisis ini diukur menggunakan Kimura-2-Parameter, yaitu metode yang menggunakan parameter transisi dan transversi untuk menghitung persentase besar perbedaan jarak antar sampel (Nei and Kumar 2000). Jarak genetik terkecil terjadi antara T. scripta dan C. picta dan antara D. subplana
dan T. scripta yaitu sekitar 0.005 sedangkan nilai jarak genetik terbesar terjadi antara P. subrufa dan E. subglobusa yaitu sekitar 0.869.
Hasil rekontruksi pohon filogeni berdasarkan urutan nukleotida 16S rRNA menggunakan metode ME, MP, dan NJ menunjukkan bahwa O. borneensis dan H. annandalei berada pada satu cabang dengan
C. reevesi (famili Bataguridae) sedangkan spesies C. picta dan T. scripta (famili Emydidae) membentuk cabang tersendiri memisahkan diri dari famili Bataguridae. Semua spesies Bataguridae yang dianalisis dalam penelitian ini mengelompok menjadi satu dengan nilai bootstrap 94% pada pohon filogeni ME, 95% pada pohon filogeni NJ dan 75% pada pohon filogeni MP. Hasil
pengklasifikasian ini sesuai dengan pengelompokkan yang dilakukan oleh McDowell (1964) berdasarkan karakter tengkorak, memisahkan kura-kura Bataguridal menjadi famili tersendiri yaitu famili Bataguridae. Berdasarkan pengelompokkan McDowell maka O. borneensis termasuk ke dalam kelompok Orlitia complex dan H. annandalei termasuk ke dalam kelompok Batagur complex. Farajallah (2002) melaporkan bahwa semua spesies Bataguridae mengelompok menjadi satu klad dalam pohon filogeni, menyisihkan Emydidae ke cabang yang lain.
Hirayama (1984) berdasarkan data morfologi fosil membagi famili Emydidae menjadi dua subfamili yaitu Emydinae dan Batagurinae akan tetapi Hirayama berpendapat bahwa pengelompokkan Emydinae dan Batagurinae dalam pandangan kladistik bukan sebagai kelompok alami, karena kedua subfamili ini mempunyai daerah penyebaran yang sangat berbeda. Kura-kura Emydinae baik secara fosil maupun yang masih hidup saat ini mempunyai daerah penyebaran di Eropa dan Amerika sedangkan semua anggota Batagurinae daerah penyebarannya di Asia kecuali genus neotropika, yaitu Rhinoclemmys. Hirayama (1984) juga mengusulkan adanya dua jalur evolusi di antara Batagurinae. Jalur pertama adalah kura-kura yang memiliki kehidupan lebih ke daratan yang diwakili oleh Geomyda dan Testudinidae. Jalur kedua melalui kura-kura yang habitatnya lebih ke air yang diwakili oleh Batagur dan Hardella.
Gaffney dan Meylan (1988) berdasarkan data osteologi juga meletakkan kura-kura Bataguridal ke dalam status famili Bataguridae dengan mengklasifikasikannya menjadi dua subfamili yaitu Batagurinae dan Geomydinae. Jika mengikuti klasifikasi Gaffney dan Meylan maka O. borneensis
dan H. annandalei termasuk ke dalam subfamili Batagurinae.
Pada pohon filogeni terlihat spesies I. elongata (famili Testudinidae) berada di luar percabangan famili Bataguridae dan menempatkan famili Emydidae sebagai pangkal dari famili Testudinidae. Iverson et al. (2001) berdasarkan variasi gen Cytokrom b mtDNA genus Indotestudo yang berasal dari Asia Selatan (I. elongata), India (I. travancorica) dan Indonesia Timur (I.
forstenii) menempatkan C. scripta
sama seperti pohon filogeni yang dihasilkan pada penelitian ini yang menempatkan T. scripta dan C. picta (Emydidae) sebagai pangkal dari I. elongata walaupun pada hasil ini tidak didapat nilai bootstap yang tinggi (kurang dari 50%).
Pada pohon filogeni terlihat spesies C. insculpta dan C. mydas dari famili Cheloniidae tidak berada dalam satu cabang. Hingga saat ini posisi filogeni C. Insculpta
(Carretochelydae) dalam ordo Testudines masih menjadi perdebatan. Meylan (1987) berdasarakan anatomi cangkang menunjukkan bahwa famili Carettochelydae berkerabat dekat dengan Famili Trionychidae. Hirayama (1994) berdasarkan 76 karakter osteologi dari 26 genera Chelonioids menempatkan genus Caretta (Carettochelydae) ke dalam famili Cheloniidae. Puspa (2005) berdasarkan urutan nukleotida gen 12S rRNA meletakkan C. insculpta membentuk politomi dengan kelompok Cryptodira, Pelomedusa dan Chelydae sedangkan berdasarkan urutan nukleotida gen 16S rRNA dan gabungannya meletakkan C. insculpta di antara famili Cheloniidae dan Trionychidae.
Pada pohon filogeni juga terlihat spesies P. subrufa (famili Pelomedusidae) dan spesies E. sublobusa (famili Chelidae) membentuk outgroup karena kedua spesies ini merupakan anggota dari Pleurodira.
KESIMPULAN
Analisis filogeni berdasarkan sebagian gen 16S rRNA menggunakan metode ME, MP dan NJ menghasilkan pohon filogeni dengan topologi yang sama; menempatkan
O. borneensis dan H. annandalei dalam satu klad dengan famili Bataguridae dan spesies dari famili Emydidae membentuk percabangan tersendiri sehingga terpisah dari famili Bataguridae. Hal ini sesuai dengan pengklasifikasian yang dilakukan oleh MC Dowell (1964) dan Gaffney dan Meylan (1988).
SARAN
Gen 16S rRNA dapat digunakan untuk analisis filogeni pada tingkat famili tetapi hal ini perlu didukung oleh data urutan nukleotida gen 16S rRNA yang lebih lengkap sehingga menghasilkan topologi pohon filogeni yang lebih stabil.
DAFTAR PUSTAKA
Bickham JW, Carr JL. 1983. Taxonomy and phylogeny of the higher categories of Cryptodiran turtles based on a cladistic analysis of chromosomal data. Copera
1983: 918-932.
Ernst CH, Barbour. 1989. Turtles of the World. Washington DC: Smithsonian Institution Pr.
Farajallah A. 2002. Karakterisasi genom mitokondria labi-labi, Dogania subplana (Trionychidae, Testudines, Reptilia) [disertasi]. Bogor. Program Pascasarjana, Institut Pertanian Bogor. Gaffney ES. 1975. A phylogeny and classification of the higher categories of turtles. Bull Am Mus Nat Hist 155: 387-436
Gaffney ES, Meylan PA. 1988. A phylogeny of turtles. In the phylogeny and classification of the Tetrapods vol 1 ed by MJ Benton. Oxford Universuty Pr. New York. 157-219.
Hirayama R. 1984. Cladistic analysis of batagurinae turtles (Batagurinae: Emydidae, Testudinoidea): a preliminary result. Stud Geol Salmanticcensia vol Especial 1.Studia Paleocheloniolog:141-157.
Hirayama R. 1994. Phylogenetic systematics of Chelonioid Sea Turtles. The Island Arc 3: 270-284.
Honda M, Yasukawa Y, Hirayama R, Ota H.
2002. Phylogenetic Relationships of the Asian Box Turtles of the Genus Cuora sensu Lato (Reptilia: Bataguridae) inferred from Mitochondrial DNA Sequences. Zool Sci 19:1305-131
Iverson JB. 1992. A Revised Checklist with Distribution Maps of the Turtles of the World. Indiana: Earlham College. Iverson JB, Spinks PQ, Mc Cord WP, Das I.
Testudinidae). Hamadryad 26(2): 272-275.
Kumar S, Tamura K, Nei M. 2001.
Molecular Evolution Genetics
Analysis (MEGA) Version 2.0. New York: Oxford Univ Pr.
Lim LB, Das I. 1999. Turtles of Borneo and
Peninsular Malasyia. Kinabalu:
Natural History Pub.
McDowell SB. 1964. Partition of the genus Clemys and related problems in the taxonomy of the aquatic Testudinidae.
Proc. Zool. Soc.London 143: 239-279. Moritz C, Dowling TE, Brown WM. 1987.
Evolution of animal mithochondrial DNA: relevance for population biology and systematics. Ann Rev Ecol Syst 18:269-291
Nei M, Kumar S. 2000. Molecular Evolution and Phylogenetics. New York: Oxford Univ Pr.
Puspa Cs. 2005. Posisi Filogeni Kura-kura
Carettochelis insculpta (Reptilia:
Testudines) [skripsi]. Bogor: Institut Pertanian Bogor.
Randi E. 2000. Mitochondrial DNA. Di dalam: Baker AJ,ed. Molecular
Methods in Ecology. Oxford:
Blackwell Sci Ltd.
Sites JW, Bickman JW, Pytel BA, Grenbaum IF, Bates BA. 1984. Biochemical characters and recontruction of the turtles phylogenies: relationships among Batagurid genera. Syst Zool 33: 137-158.
Tegelstrom H. 1986. Mithochondrial DNA in natural population: An improved routine for sreening of genetic variation based on sensitives silver staining. Electrophoresis 7:226-229. Zardoya R, Meyer A. 1998. Complete
Lampiran 1 Klasifikasi Ordo Testudines (Gaffney 1975)
Klas Reptilia
Subklas Anapsida
Ordo Testudines
Subordo Cryptodira
Superfamili Testudinoidea
Famili Chelydridae
Famili Emydidae
Subfamili Emydinae
Subfamili Batagurinae
Famili Testudinidae
Superfamili Chelonioidea
Famili Cheloniidae
Famili Dermochelyidae
Superfamili Trionychoidea
Famili Dermatemydidae
Famili Kinosternidae
Famili Carettochelyidae
Famili Trionychidae
Subfamili Cyclanorbinae
Subfamili Trionychinae
Subordo Pleurodira
Famili Pelomedusidae
Lampiran 2 Taksonomi Orlitia borneensis
Klas : Reptilia
Subklas : Anapsida
Ordo : Testudines (Chelonia)
Superfamili : Testudinoidea
Famili : Bataguridae (Geomyda)
Genus : Orlitia
Spesies : Orlitia borneensis
Nama Umum : Malayan Giant Turtles, Malayan Giant Terrapin (Inggris)
Bajuku/Biuku, Tuntung, Kura-kura Gading (Indonesia)
Lampiran 3 Taksonomi Hieremys annandalei
Klas : Reptilia
Subklas : Anapsida
Ordo : Testudines (Chelonia)
Superfamili : Testudinoidea
Famili : Bataguridae (Geomyda)
Genus : Hieremys
Spesies : Hieremys annandalei
Nama Umum : Yellow headed Turtles (Inggris)
This document was created with Win2PDF available at http://www.daneprairie.com.
POSISI FILOGENI
Orlitia borneensis
DAN
Hieremys annandalei
(TESTUDINES) BERDASARKAN GEN 16S rRNA
D. KURNIAWATI
G04400049
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ABSTRAK
D. KURNIAWATI. Posisi Filogeni Orlitia borneensis dan Hieremys annandalei
(TESTUDINES) Berdasarkan Gen 16S rRNA. Dibimbing oleh ACHMAD FARAJALLAH dan RIKA RAFFIUDIN.
Urutan DNA mitokondria (mtDNA) banyak digunakan sebagai penanda molekuler dalam menentukan hubungan filogeni suatu organisme karena struktur organisasinya yang stabil. Seiring dengan kemajuan bioteknologi, pengelompokkan kura-kura Bataguridal masih menjadi perdebatan. Hal ini disebabkan oleh perbedaan karakter yang diamati. Beberapa hasil penelitian mengelompokkan kura ini ke dalam famili Emydidae tetapi ada yang menempatkan kura-kura ini ke dalam famili tersendiri yaitu famili Bataguridae. Pada penelitian ini digunakan urutan nukleotida gen 16S rRNA O. borneensisdanH. annandalei (Testudines) yang disejajarkan dengan urutan nukleotida beberapa spesies Testudines lainnya yang terdapat di Genbank. Hasil rekonstruksi pohon filogeni berdasarkan urutan nukleotida 16S rRNA menunjukkan bahwa O. borneensisdan H. annandaleiberkerabat dengan Chinemys reevesi(famili Bataguridae). Hasil amplifikasi gen 16S rRNAO. borneensisdanH. annandaleimenunjukkan komposisi nukleotida didominasi oleh nukleotida adenin dan timin dengan nilai perbandingan antara A/T dan G/C = 60.1%:39.9% untuk O. borneensis dan 63%:37% untuk H. annandalei. Rasio transisi terhadap transversi (ts/tv) gen 16S rRNA berkisar antara 0.67 sampai dengan 6.75. Nilai jarak genetik terkecil terjadi antara Trachemys scripta danChrysemys pictadan antaraDogania subplanadan
Tracemys scripta yaitu sekitar 0.005 sedangkan nilai jarak genetik terbesar terjadi antara
Pelomedusa subrufa dan Emydura subglobusayaitu sekitar 0.869.
ABSTRACT
D. KURNIAWATI. Phylogenetic Position of Orlitia borneensis and Hieremys
annandalei (TESTUDINES) Based on 16S rRNA Gene. Supervised by ACHMAD
FARAJALLAH and RIKA RAFFIUDIN.
Mitochondrial gene has been commonly used as molecular marker in phylogenetic analysis. Due to its stable organization structure, mtDNA is often used to determine phylogenetic relationship of organisms. Along with the increasing improvements in biotechnology methods, classification of Bataguridal turtles was still in debate among scientists. This is mainly due to the differences in the observed characteristics. Several studies placed Bataguridal turles into Emydidae family, but others placed Bataguridal turtles into a separated family: Bataguridae. This research used sequence data of 16S rRNA of Orlitia borneensis and Hieremys annandalei aligned with those of several Testudines species deposited in Genbank. DNA alignment conducted using Clustal W 1.8 software. Nucleotide composition, transition and transvertion substitution type (ts/tv), determination of genetic distance and phylogenetic trees reconstruction were performed using MEGA 3.0 software. The result of phylogenic trees reconstruction based on 16S rRNA showed thatOrlitia borneensisandHieremys annandaleiwere closely related toChinemys reevesi
(Bataguridae). Amplification of 16S rRNA gene of Orlitia borneensis and Hieremys annandalei
showed that the nucleotide composition was dominated by adenine and timin nucleotides with A/T:G/C value of 60.1%:39.9% for Orlitia borneensis and 63%:37% for Hieremys annandalei.
PENDAHULUAN
Latar Belakang
Kura-kura diklasifikasikan sebagai klas Reptilia subklas Anapsida dan ordo Testudines (Ernst dan Barbour 1989). Berdasarkan klasifikasi oleh Gaffney (1975) ordo Testudines atau kura-kura modern terbagi menjadi 2 subordo yaitu subordo Cryptodira dan subordo Pleurodira. Subordo Cryptodira terbagi atas 3 superfamili yaitu superfamili Testudinoidea, Chelonioidea dan Trionychoidea, sedangkan subordo Pleurodira terbagi atas dua famili yaitu Pelomedusidae dan Chelidae. Bagan klasifikasi Testudines selengkapnya dapat dilihat pada Lampiran 1.
Ordo Testudines atau kura-kura modern mempunyai ciri-ciri tengkorak masih primitif, kranium utuh tanpa lubang temporal dan tubuhnya berada dalam cangkang. Cangkang terdiri atas dua bagian yang saling berhubungan yaitu karapas (sisi bagian atas) dan plastron (sisi bagian bawah). Umumnya cangkang terbuat dari bahan tanduk yang keras tetapi pada beberapa taksa ada yang bersifat lunak yaitu pada famili Trionychidae. Subordo Cryptodira memiliki ciri-ciri mempunyai leher yang berukuran pendek dan bergerak secara vertikal sehingga kepala dan leher dapat ditarik masuk ke dalam cangkang (kecuali anggota famili Cheloniidae (penyu laut)). Sedangkan anggota Pleurodira mempunyai leher dengan ukuran yang panjang, bergerak secara lateral sehingga kepala dan leher tidak dapat ditekuk dan ditarik masuk kedalam cangkang (Lim dan Das 1999).
Famili Bataguridae merupakan anggota dari subordo Cryptodira, termasuk ke dalam
Old World Pond Turtles, memiliki keragaman fenotipe yang tinggi, hidup di daerah semi akuatik, semi terestrial dan tropik. Famili ini tersebar terutama di daerah Asia Tenggara tetapi juga di daerah Eropa Barat, Afrika Utara dan Amerika Latin (Iverson 1992). Karakter umum famili Bataguridae adalah tubuhnya terdiri atas cangkang yang keras, karapas mempunyai permukaan yang licin dan terdiri atas 5 vertebral, plastron keras, kaki depan tidak berbentuk seperti stik golf (not club-shaped), selaput diantara jari-jarinya ada atau kadang tidak ada, kaki mempunyai 4 atau 5 cakar, kepala dan leher dapat ditekuk dan ditarik masuk ke dalam cangkangnya (Lim dan Das 1999).
Orlitia borneensis dan Hieremys
annandalei merupakan anggota famili Bataguridae. Daerah penyebarannya di Asia seperti Thailand, Malaysia dan Indonesia terutama di daerah Sumatera (Ernst dan Barbour 1989, Lim dan Das 1999).
Orlitia borneensis (Malayan Giant Turtles) merupakan kura-kura air tawar terbesar di pulau Borneo dan Malaysia. Ciri-ciri O. borneensis adalah panjang karapas hingga mencapai 800 mm, ukuran kepala besar dan cangkang berwarna gelap. Habitatnya terutama di sungai yang besar dan daerah-daerah air tawar yang mempunyai ukuran yang luas dan termasuk ke dalam omnivora (Lim dan Das 1999). Status menurut IUCN tahun 2000 termasuk fauna yang dilindungi. Taksonomi O. borneensis dapat dilihat pada Lampiran 2.
Hieremys annandalei (Yellow-headed Temple Turtles) memiliki ciri-ciri karapas berwarna gelap kadang dengan garis punggung berwarna orange. Panjang karapas dapat mencapai 506 mm, habitat terutama di sungai, kubangan dan rawa dan termasuk ke dalam herbivora (Lim dan Das 1999). Status menurut IUCN tahun 2000 termasuk ke dalam fauna yang dilindungi. Taksonomi H. annandalei dapat dilihat pada Lampiran 3.
Seiring dengan kemajuan bioteknologi, pengelompokan kura-kura Bataguridal (Bataguridae, subfamili Batagurinae, famili Testudinidae) masih menjadi perdebatan. Hal ini disebabkan karena perbedaan karakter yang diamati. McDowell (1964) berdasarkan karakter tengkorak meletakkan kura-kura Bataguridal kedalam empat kelompok (complex) yaitu Hardella complex (Hardella, Morenia dan Geoclemys), Orlitia complex
(Orlitia dan Siebenrockiella), Batagur complex
Emydidae menempatkan dua spesies Emydidae yang diteliti sebagai outgroup dari spesies-spesies Bataguridae. Gaffney dan Meylan (1988) berdasarkan data osteologi meletakkan kura-kura Bataguridal ke dalam status famili Bataguridae dengan mengklasifikasikannya menjadi dua subfamili yaitu Batagurinae (termasuk Batagur, Hardella, dan Orlitia complex ( klasifikasi McDowell, 1964)) dan Geomydinae (Geomyda complex (klasifikasi McDowell, 1964)). Ernst dan Barbour (1989) berdasarkan morfologi tengkorak mengklasifikasikan kura-kura Bataguridal ke dalam famili Emydidae. Iverson (1992) berdasarkan penyebaran kura-kura di dunia mengklasifikasikan kura-kura Bataguridal ke dalam famili Emydidae.
Genom mitokondria sangat populer digunakan sebagai penanda molekuler dalam analisis filogeni, mempelajari evolusi, struktur populasi dan aliran genetik, hibridisasi, biogeografi dan berbagai fenomena mikro dan makro evolusi hewan. Genom mitokondria diwariskan secara maternal, memiliki laju evolusi yang cepat dan tanpa intron. Struktur organisasi mtDNA bersifat stabil, sehingga mtDNA hewan memiliki jumlah gen dan ukuran yang sama, yakni terdiri atas 13 gen yang menyandikan protein (terlibat dalam respirasi sel), 2 gen menyandikan rRNA (12S dan 16S), 22 gen menyandikan tRNA dan beberapa daerah lain yang tidak menyandikan protein D-loop (Control Region) dan daerah
intergenic (Moritz et al. 1987) (Lampiran 4). Meski demikian, gen-gen dalam mtDNA memiliki laju evolusi yang berbeda-beda, evolusi tRNA dan rRNA mitokondria relatif lambat jika dibandingkan dengan gen lainnya. Karena itu tRNA dan rRNA mitokondria banyak digunakan untuk mempelajari hubungan filogeni pada tingkat interspesies (Randi 2000).
Zardoya dan Meyer (1998) telah menggunakan genom mitokondria untuk analisis filogeni diantara kelompok vertebrata. Honda et al (2002) juga menggunakan runutan mtDNA untuk menentukan hubungan filogeni Asian Box Turtles.
Tujuan
Penelitian ini bertujuan untuk menentukan posisi filogeni O. borneensis dan
H. annandalei (Testudines) berdasarkan gen 16S rRNA.
Waktu dan Tempat
Penelitian ini dilaksanakan pada bulan Januari sampai dengan Juni 2006 bertempat di Laboratorium Zoologi Departemen Biologi Fakultas MIPA, Institut Pertanian Bogor, Bogor.
BAHAN DAN METODE Sampel Kura-kura
Sampel yang digunakan pada penelitian ini adalah 4 spesimen ujung ekor dan 1 spesimen karapas O. borneensis (Gambar 1), 1 spesimen ujung ekor H. annandalei
(Gambar 2), 2 spesimen ujung ekor Cuora amboinensis (Gambar 3), 1 spesimen ujung ekor Siebenrockiella crassicolis (Gambar 4) dan 1 spesimen ujung ekor Cyclemys dentata
(Gambar 5). Semua sampel yang digunakan merupakan koleksi Achmad Farajallah (Lab. Zoologi Dept. Biologi FMIPA IPB, Bogor ) yang diambil dari Kalimantan Tengah.
[image:32.595.325.504.380.447.2]
(a) (b) Gambar 1 O. borneensis (a) tampak plastron (b) tampak
karapas.
[image:32.595.321.506.467.674.2](a) (b)
Gambar 2 H. annandalei (a) tampak plastron (b) tampak karapas.
(a) (b)
Emydidae menempatkan dua spesies Emydidae yang diteliti sebagai outgroup dari spesies-spesies Bataguridae. Gaffney dan Meylan (1988) berdasarkan data osteologi meletakkan kura-kura Bataguridal ke dalam status famili Bataguridae dengan mengklasifikasikannya menjadi dua subfamili yaitu Batagurinae (termasuk Batagur, Hardella, dan Orlitia complex ( klasifikasi McDowell, 1964)) dan Geomydinae (Geomyda complex (klasifikasi McDowell, 1964)). Ernst dan Barbour (1989) berdasarkan morfologi tengkorak mengklasifikasikan kura-kura Bataguridal ke dalam famili Emydidae. Iverson (1992) berdasarkan penyebaran kura-kura di dunia mengklasifikasikan kura-kura Bataguridal ke dalam famili Emydidae.
Genom mitokondria sangat populer digunakan sebagai penanda molekuler dalam analisis filogeni, mempelajari evolusi, struktur po


![Gambar 9 Hasil perunutan DNA H. annandalei gen 16S rRNA. Angka di dalam tanda”[ ]” menunjukkan posisi nukleotida terakhir dalam baris](https://thumb-ap.123doks.com/thumbv2/123dok/497512.331267/15.595.89.507.290.760/gambar-hasil-perunutan-annandalei-menunjukkan-posisi-nukleotida-terakhir.webp)