INTROGRESI GEN TAHAN BLAS DARI ORYZICA LLANOS-5 PADA POPULASI SILANG BALIK LANJUT
WAY RAREM X ORYZICA LLANOS-5
WENING ENGGARINI
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI DISERTASI DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa disertasi saya yang berjudul
INTROGRESI GEN TAHAN BLAS DARI ORYZICA LLANOS-5 PADA POPULASI SILANG BALIK LANJUT WAY RAREM X ORYZICA LLANOS-5 adalah hasil karya sendiri dengan arahan Komisi Pembimbing dan belum pernah diajukan dalam bantuk apapun untuk memperoleh gelar program sejenis di perguruan tinggi lain mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun yang tidak diterbitkan dari penulis lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir disertasi ini.
Bogor, Agustus 2012
Wening Enggarini
ABSTRACT
WENING ENGGARINI. Blast resistance introgression of Oryzica Llanos-5 on the advanced backcross population Way Rarem x Oryzica Llanos-5. Under supervision of SURJONO HADI SUTJAHJO, TRIKOESOEMANINGTYAS, SRIANI SUJIPRIHATI (Almh.), UTUT WIDYASTUTI, and SUGIONO MOELJOPAWIRO.
Upland rice is an alternative resource for enhancing rice productivity in Indonesia. However upland rice cultivation technology is still not optimum, particularly in controlling blast pathogen (Pyricularia oryzae). The development of cultivar multiline using NIL (Near isogenic lines) population is one of the important efforts to prevent the breaking down of blast resistance in field. The advanced backcross lines were developed using the recurrent parent Way Rarem, which is high yielding but recently was destroyed by blast, and the durable blast resistant Oryzica Llanos-5, as a donor parent. The study was aimed 1) to develop NIL Way Rarem x Oryzica Llanos-5 having a higher blast resistance compared to Way Rarem; 2) to obtain molecular markers which could detect the introgression segment of blast resistance-carrying donor on the advanced backcross lines; and 3) to get information about the blast resistance inheritance of Oryzica Llanos-5. The evaluation of blast resistance demonstrated that the percentage of the blast resistant plant on BC4F1 and BC5F1 were higher than that on BC3F1 and BC4F1,
respectively. The blast test using each 133 and 173 blast races on BC5F1 showed
that there were several BC5F1 plants having a higher blast resistance to each both
blast races compared to Way Rarem. This high blast resistance character might be derived from the introgression segments of Oryzica Llanos-5. The detection of blast resistance introgression from Oryzica Llanos-5 using SSR primers on BC3F1
population demonstrated that three markers (RM411, RM7 and RM251) on chromosome 3 and three markers (RM276, RM549 and RM585) on chromosome 6 were polymorphic markers and could identify the donor introgression based on the appearance of heterozygous allele on PCR amplification. The six SSR markers and the evaluation of blast resistance were used to select the plant on each BC4F1,
BC5F1 and BC6F1 generation which was backcrossed to Way Rarem. Based on the
genome wide scan in the blast resistant BC6F1 plant we found several SSR
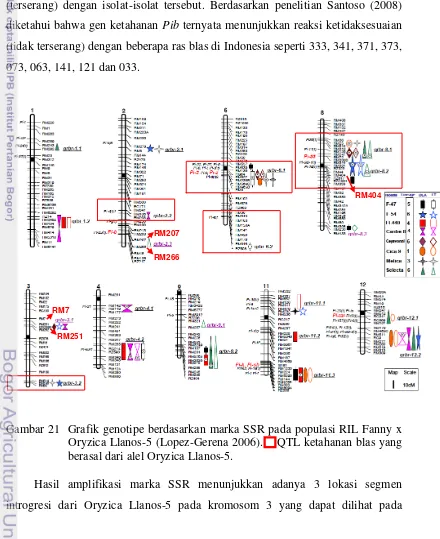
markers which could detect the donor introgression on chromosome 2, 3, 6 and 8. RM207 and RM266 were on chromosome 2, also RM404 was on chromosome 8. This is interesting information because the introgression segment on both chromosomes could not detect in earlier research by Lestari et al. (2011). Both introgression segments were located quite close to blast resistance QTL derived from Oryzica Llanos-5 alleles in RIL population Fanny x Oryzica Llanos-5 that the introgression segment on chromosome 2 was located close to Pib gene and on chromosome 8 was located close to Pi33 gene. In addition there were also introgressions segment of Oryzica Llanos-5 on chromosome 3 that could be detected by RM565 and RM514 which were similar position with blast resistance QTLs on population of cross between Fanny x Oryzica Llanos-5 varieties.
RINGKASAN
WENING ENGGARINI. Introgresi gen tahan blas dari Oryzica Llanos-5 pada populasi silang balik lanjut Way Rarem x Oryzica Llanos-5. Dibimbing oleh:
SURJONO HADI SUTJAHJO, TRIKOESOEMANINGTYAS, SRIANI
SUJIPRIHATI (Almh.), UTUT WIDYASTUTI, dan SUGIONO
MOELJOPAWIRO.
Padi gogo merupakan salah satu alternatif dalam usaha peningkatan produksi beras di Indonesia. Namun saat ini penerapan teknologi budidaya padi gogo belum optimal, terutama dalam pengendalian penyakit blas yang disebabkan oleh cendawan Pyricularia oryzae. Cendawan ini mudah membentuk ras baru dengan tingkat virulensi yang lebih tinggi sehingga dapat dengan cepat mematahkan ketahanan varietas tanaman yang sebelumnya tahan terhadap blas. Salah satu cara yang dapat mencegah patahnya ketahanan varietas terhadap blas adalah dengan pembentukan varietas multilini. Varietas multilini (multilines) merupakan populasi NIL (Near isogenic lines) yang dibentuk melalui beberapa kali silang balik sehingga dihasilkan galur-galur dengan sifat agronomis yang sama tetapi masing-masing memiliki sifat ketahanan yang berbeda yang berasal dari segmen introgresi tetua donornya.
Way Rarem merupakan varietas padi gogo Indica dari Indonesia. Varietas ini memiliki daya hasil yang cukup tinggi dan ketahanan terhadap blas pada awal dilepas pada tahun 1994. Namun ketahanannya terhadap blas terpatahkan setelah beberapa kali musim tanam sehingga perlu ditingkatkan melalui persilangan dengan tetua donor tahan blas, Oryzica Llanos-5. Penelitian ini dilakukan bertujuan untuk: 1) Mendapatkan galur-galur NIL Way Rarem x Oryzica Llanos-5 yang memiliki ketahanan blas lebih tinggi dibandingkan dengan tetua berulangnya, Way Rarem; 2) Memperoleh informasi tentang pola pewarisan sifat ketahanan terhadap patogen blas pada varietas Oryzica Llanos-5; dan 3) Mendapatkan marka molekuler yang mampu mendeteksi introgresi segmen genom dari Oryzica Llanos-5 yang terkait dengan sifat ketahanan terhadap patogen blas pada generasi silang balik lanjut Way Rarem x Oryzica Llanos-5.
Materi tanaman pada awal penelitian adalah populasi BC2F5 Way Rarem x
Oryzica Llanos-5 yang merupakan hasil selfing beberapa kali dari tanaman BC2F2
Way Rarem x Oryzica Llanos-5 yang digunakan pada penelitian pemetaan QTL ketahanan blas oleh Lestari et al. (2011). Setiap kali melakukan silang balik dilakukan seleksi fenotipe dan genotipe terlebih dahulu untuk mendapatkan tanaman-tanaman yang tahan blas dan memiliki segmen introgresi dari Oryzica Llanos-5, yang selanjutnya disilang balik terhadap Way Rarem agar didapatkan benih turunan silang balik selanjutnya hingga generasi BC6.
Seleksi fenotipe dilakukan dengan mengevaluasi ketahanan blas pada populasi turunan silang balik. Inokulum blas yang digunakan untuk evaluasi pada populasi BC3F1 dan BC4F1 terdiri atas campuran 4 ras blas: 033, 123, 133 dan
173, sedangkan pada evaluasi BC5F1 digunakan 4 ras blas tersebut secara terpisah.
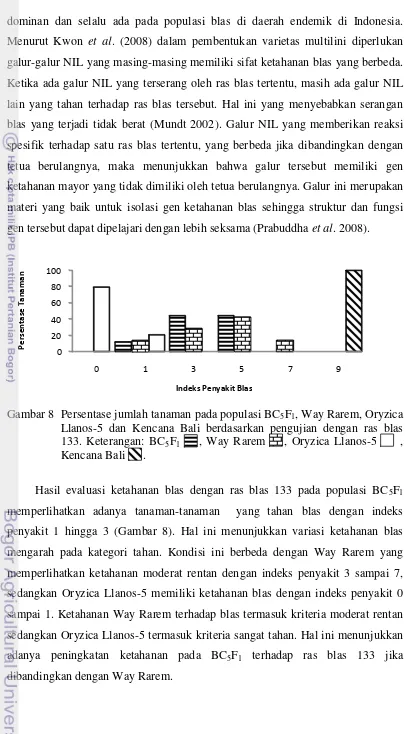
Hasil evaluasi ketahanan blas pada populasi BC3F1 dan BC4F1 memperlihatkan
tahan, sedangkan Oryzica Llanos-5 memiliki indeks penyakit 0 sampai 1 yang merupakan tanaman sangat tahan. Demikian juga dengan hasil evaluasi pada populasi BC5F1 diperoleh tanaman-tanaman yang tahan terhadap masing-masing
ras blas 133 dan 173. Hal ini menunjukkan adanya peningkatan ketahanan blas pada populasi BC3F1, BC4F1 dan BC5F1 jika dibandingkan dengan Way Rarem
sebagai tetua berulangnya. Sumber ketahanan blas tersebut diduga berasal dari segmen introgresi dari Oryzica Llanos-5. Pada pengujian BC5F1 masing-masing
dengan ras blas 033 dan 123 memperlihatkan bahwa Way Rarem dan Oryzica Llanos-5 tahan terhadap kedua ras tersebut dengan indeks penyakit 0 sampai 1 yang merupakan kriteria tanaman sangat tahan. Hal ini terjadi diduga bahwa baik Way Rarem dan Oryzica Llanos-5 memiliki gen ketahanan yang dapat mengatasi ras blas 033 dan 123.
Keberhasilan program pemuliaan tanaman tahan blas tidak lepas dari adanya pengetahuan mengenai mekanisme penurunan sifat ketahanan blas sehingga dapat ditentukan metode pemuliaan yang tepat. Oleh karena itu dilakukan studi pewarisan sifat ketahanan blas pada Oryzica Llanos-5. Pada penelitian ini dilakukan persilangan antara varietas moderat rentan Way Rarem dengan varietas tahan Oryzica Llanos-5 untuk mendapatkan populasi F1, F1 resiprokal, F2, BCP1
dan BCP2. Semua populasi hasil persilangan beserta kedua tetua diinokulasi
dengan ras blas 133 dan 173. Hasil penelitian menunjukkan bahwa ketahanan terhadap ras blas 133 dan 173 merupakan ketahanan horisontal namun terdapat pula peran gen mayor di dalam kendali genetiknya dan tidak ada pengaruh dari tetua betina. Ketahanan terhadap kedua ras tersebut dikendalikan oleh gen mayor yang bersifat dominan dan beberapa gen minor. Aksi gen pengendali ketahanan terhadap ras blas 133 dipengaruhi oleh efek aditif, dominan serta interaksi aditif x dominan dan dominan x dominan; sedangkan ketahanan terhadap ras blas 173 dipengaruhi oleh efek aditif, dominan serta interaksi aditif x aditif dan dominan x dominan. Peran gen mayor pengendali ketahanan terhadap ras blas 133 mengikuti nisbah 13:3 (tahan:rentan), sedangkan ketahanan terhadap ras blas 173 mengikuti nisbah 37:27 (tahan:rentan). Ekspresi gen mayor pada umumnya memiliki nilai heritabilitas yang tinggi dan tidak dipengaruhi oleh perubahan lingkungan. Karakter yang dikendalikan oleh gen mayor sangat baik untuk digunakan sebagai gen donor pada teknik silang balik.
Seleksi genotipe menggunakan marka molekuler penting dilakukan untuk mendeteksi introgresi dari tetua donor pada turunan hasil silang balik. Seleksi genotipe pada pada populasi BC2F5 menggunakan 3 marka SSR (Simple sequence repeat) berdasarkan Lestari et al. (2011) yaitu RM148, RM541 dan RM342B yang masing-masing berada pada kromosom 3, 6 dan 9. Pada BC3F1 ketiga marka
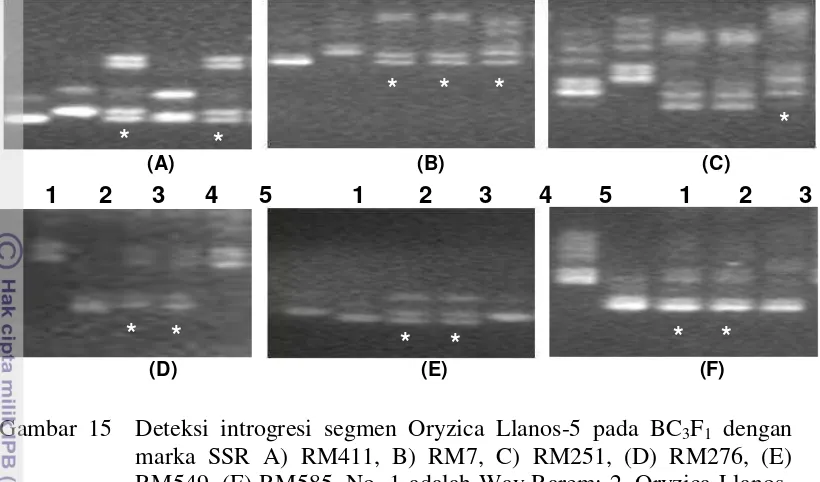
SSR tersebut tidak dapat mendeteksi lagi introgresi dari Oryzica Llanos-5. Hal ini diduga karena jarak antar marka dengan lokus target masih terlalu jauh (lebih dari 10 cM). Oleh karena itu dilakukan eksplorasi primer SSR untuk mendapatkan marka SSR baru pada kromosom 3, 6 dan 9. Dari 120 primer SSR yang digunakan, ditemukan 3 marka (RM411, RM7 dan RM251) pada kromosom 3 dan 3 marka (RM276, RM549 dan RM585) pada kromosom 6 yang dapat mendeteksi introgresi dari Oryzica Llanos-5 pada tanaman BC3F1. Berdasarkan analisis marka
selanjutnya digunakan untuk kegiatan seleksi pada generasi BC4F1 hingga BC6F1.
Jumlah marka SSR yang polimorfik yang didapatkan pada penelitian ini masih terbatas sehingga cukup banyak lokus-lokus yang belum diketahui kondisi genotipenya. Hal ini dapat terjadi karena tetua Way Rarem dan Oryzica Llanos-5 sama-sama merupakan padi Indica yang memiliki banyak kemiripan sifat (Bustamam et al. 2004). Hal yang dapat dilakukan untuk mengatasi permasalahan ini adalah 1) dengan meningkatkan jumlah primer SSR yang digunakan terutama primer-primer SSR pada padi yang baru ditemukan dan 2) menggunakan marka molekuler lain yang memiliki kelimpahan di genom lebih tinggi dibandingkan SSR seperti marka SNP (single nucleotide polymorphism).
Marka molekuler selain digunakan untuk mendeteksi introgresi dari tetua donor, juga bermanfaat untuk menduga seberapa besar proporsi segmen dari tetua berulang sudah kembali kepada turunan hasil silang balik. Pendugaan proporsi segmen dari kedua tetua telah banyak dilakukan dengan menggunakan simulasi teori matematika. Pada penelitian ini dilakukan analisis pendugaan proporsi segmen dari tetua berulang dan tetua donor langsung pada populasi BC2F7 dan
BC4F2 Way Rarem x Oryzica Llanos-5 menggunakan satu set marka SNP. Hasil
penelitian menunjukkan bahwa segmen introgresi dari Oryzica Llanos-5 masih terdapat di hampir seluruh kromosom populasi BC4F2. Terdapat peningkatan
proporsi segmen tetua berulang sebesar 30% dari BC2F7 ke BC4F2, sedangkan
proporsi genom segmen donor menurun sebesar 76% dari BC2F7 ke BC4F2. Hal
ini sesuai dengan pernyataan oleh Sleeper dan Poehlman (2006) bahwa proporsi genom segmen tetua berulang meningkat sebesar 25%, sedangkan segmen donor berkurang sebesar 75% setelah dua kali silang balik. Selain itu, diperoleh marka SNP yang dapat mendeteksi segmen introgresi tetua donor pada populasi BC4F2.
Dalam rangka pembentukan galur-galur NIL (Near isogenic lines) dibutuhkan beberapa kali silang balik sehingga perlu dianalisis keberadaan segmen introgresi dari tetua donor yang diinginkan tetap berada pada galur NIL. Selain untuk mengetahui adanya kemungkinan ditemukan segmen introgresi di lokasi lain yang sebelumnya tidak terdeteksi. Pada penelitian ini digunakan 400 marka SSR polimorfik yang berada pada 12 kromosom padi untuk mendeteksi lokasi segmen introgresi pada tanaman BC6F1 Way Rarem x Oryzica Llanos-5.
Hasil dari penelitian ini didapatkan marka SSR yang mampu mendeteksi segmen introgresi dari Oryzica Llanos-5 yaitu RM207 dan RM266 pada kromosom 2 dan RM404 pada kromosom 8. Hal ini merupakan informasi tambahan dimana pada hasil penelitian sebelumnya berdasarkan Lestari et al. (2011) lokasi segmen introgresi pada kedua kromosom tersebut tidak diketahui. Berdasarkan informasi sebelumnya tentang pemetaan gen ketahanan blas (gen Pi) pada padi, maka diketahui bahwa segmen introgresi dari Oryzica Llanos-5 pada kromosom 2 letaknya dekat dengan gen Pib, sedangkan pada kromosom 8 dekat dengan gen
Pi33. Selain itu terdapat juga segmen introgresi pada kromosom 3 yang dapat dideteksi oleh RM565 dan RM514, segmen introgresi ini terletak cukup jauh dengan yang sebelumnya dapat dideteksi oleh RM7 dan RM411. Sampai saat ini belum ditemukan informasi mengenai adanya gen Pi pada kromosom 3 maka diduga segmen introgresi dari Oryzica Llanos-5 pada kromosom 3 merupakan gen-gen yang berperan dalam ketahanan kuantitatif.
@ Hak Cipta milik IPB, tahun 2012 Hak Cipta dilindungi oleh Undang-Undang
1. Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumber.
a. Pengutipan hanya untuk kepentingan pendidikan, penelitian, penulisan
karya ilmiah, penyusunan laporan, penulisan kritik, atau tinjauan suatu
masalah
b. Pengutipan tersebut tidak merugikan kepentingan yang wajar IPB
2. Dilarang mengumumkan dan memperbanyak sebagian atau seluruh Karya
INTROGRESI GEN TAHAN BLAS DARI ORYZICA LLANOS-5 PADA POPULASI SILANG BALIK LANJUT
WAY RAREM x ORYZICA LLANOS-5
WENING ENGGARINI
Disertasi
Sebagai salah satu syarat untuk memperoleh gelar
Doktor pada
Program Studi Pemuliaan dan Bioteknologi Tanaman
SEKOLAH PASCASARJANA INSTITUT PERTANIAN BOGOR
Penguji pada Ujian Tertup
Penguji pada Ujian Terbuka
: Dr. Ir. Hajrial Aswidinnoor, MSc.
Dr. Ir. Widodo, MSc.
: Prof. Dr. Ir. Bambang Sapta Purwoko, MSc.
Judul Disertasi : Introgresi Gen Tahan Blas dari Oryzica Llanos-5
pada Populasi Silang Balik Lanjut Way Rarem x
Oryzica Llanos-5
Nama : Wening Enggarini
NRP : A263070071
Mayor : Pemuliaan dan Bioteknologi Tanaman
Disetujui,
Komisi Pembimbing
Prof. Dr. Ir. Surjono Hadi Sutjahjo, M.S. Ketua
Dr. Ir. Trikoesoemaningtyas, M.Sc. Prof. Dr. Ir. Sriani Sujiprihati, M.S. Anggota Anggota (Almh.)
Dr. Ir. Utut Widyastuti, M.S Prof. (R). Dr. Ir. Sugiono Moeljopawiro, M.Sc. Anggota Anggota
Diketahui,
Ketua Mayor Dekan Sekolah Pascasarjana Pemuliaan dan Bioteknologi Tanaman
Dr. Ir. Trikoesoemaningtyas, M.Sc. Dr. Ir. Dahrul Syah, M.Sc.Agr
PRAKATA
Syukur alhamdulillah, penulis panjatkan ke hadirat Allah SWT yang telah nemberikan rahmat dan karunia-Nya yang sungguh besar sehingga penelitian dan karya tulis ini dapat diselesaikan. Topik yang dipilih dalam penelitian yang dilaksanakan sejak Agustus 2008 ini adalah mengenai masalah ketahanan padi gogo terhadap patogen blas,
dengan judul “Introgresi Gen Tahan Blas dari Oryzica Llanos-5 pada Populasi
Silang Balik Lanjut Way Rarem x Oryzica Llanos-5”.
Disertasi ini memuat 5 bab percobaan, dimana bab yang berjudul “Characterization of Donor Introgression of Advanced Backcross Population Way Rarem x Oryzica Llanos-5 Detected by SNP Markers” telah diterbitkan di dalam Jurnal Agrobiogen Volume 8 Nomor 1 Tahun 2012. Disertasi ini merupakan bagian dari penelitian yang didanai oleh Proyek Generation Challenge Program Capacity Building sejak tahun 2007 sampai 2009 serta mendapatkan beasiswa pendidikan dari Badan Litbang Pertanian, Kementerian Pertanian pada tahun 2010-2011.
Pada kesempatan ini penulis menyampaikan terima kasih yang sebesar-besarnya kepada Prof. Dr. Ir. Surjono Hadi Sutjahjo, MS., Prof. Dr. Ir. Sriani Sujiprihati, MS. (almh.), Dr. Ir. Trikoesoemaningtyas, MSc., Dr. Ir. Utut Widyastuti, MS., dan Prof. (R). Dr. Ir. Sugiono Moeljopawiro, MSc., selaku komisi pembimbing; Dr. Ir. Trikoesoemaningtyas, MSc. yang juga selaku Ketua Mayor Pemuliaan dan Bioteknologi Tanaman, Dr. Casiana Vera-Cruz dan Dra. Masdiar Bustamam, MSc selaku Peneliti Utama beserta Dr. Kurniawan Rudi Trijatmiko selaku Peneliti dalam Proyek GCP
Capacity Building atas kesempatan, bantuan dan wawasan keilmuan yang diberikan. Ungkapan terima kasih juga disampaikan kepada Dr. Achmad Suryana dan Dr. Haryono (Kepala Badan Litbang Pertanian) beserta Dr. Sutrisno dan Dr. Karden Mulya (Kepala BB Litbang Biogen) yang telah mengijinkan penulis untuk melaksanakan tugas belajar ini. Terima kasih disampaikan kepada Dr. Ir. Hajrial Aswidinnoor, MSc. dan Dr. Ir. Sobir, MS. selaku penguji ujian prelim, Dr. Ir. Widodo, MSc. selaku penguji ujian tertutup, Prof. Dr. Ir. Bambang Sapta Purwoko, MSc. dan Dr. Ir. Suwarno, MS. selaku penguji ujian terbuka atas saran dan kritik yang sangat membangun.
Di samping itu penulis menyampaikan terima kasih kepada Dr. Joko Prasetiyono, Dr. Reflinur dan Tasliah, MSi. yang selalu turut membantu dalam diskusi penelitian; beserta anggota tim penelitian: Bapak Mahrup, Bapak Iman Ridwan, Ibu Ma’sumah, SSi. dan Bapak Fajar Suryawan; Ani Lestari, MSi dan Rinda Fadzila, SSi selaku mahasiswa yang juga turut membantu dalam kegiatan penelitian; teman-teman seperjuangan PBT dan AGR 2007: Dr. Diana Sofia Hanafiah, Dr. Budi Santosa, Dr. Arifah Rahayu dan lainnya; beserta teman-teman PBT dan AGR yang telah banyak memberikan semangat dan dukungan. Terakhir, namun yang paling berarti, ungkapan terima kasih sedalam-dalamnya kepada keluarga tercinta: suamiku, Roberdi, MSi.; anak kami, Innayati Cahaya Althafunnisa; ayahku, Dr. H. Nasuka dan ibuku, Hj. Yuliastuti Nasuka; ibu mertuaku, Sidaria; kakakku, Dody Saptadi dan Siti Halimah; Ir. Hj. Anny Slamet, MM., Ibu Hj. Siti Sundiyah, SE. beserta seluruh keluarga atas segala doa, kasih sayang, bantuan dan semangat yang tak henti-hentinya selalu diberikan.
Akhir kata penulis menaruh harapan semoga karya ilmiah ini bermanfaat.
Bogor, Agustus 2012
RIWAYAT HIDUP
Penulis dilahirkan di Bandung pada tanggal 11 Agustus 1978 sebagai anak
bungsu dari pasangan Bapak Dr. H. Nasuka dan Ibu Hj. Yuliastuti Nasuka.
Pendidikan sarjana di Jurusan Biologi, Fakultas Matematika dan Ilmu
Pengetahuan Alam, Institut Teknologi Bandung diperoleh pada tahun 2001. Pada
tahun 2002 penulis diterima di Program Studi Biologi pada Program Pasca
Sarjana ITB dan tamat pada tahun 2005. Pada tahun 2007 penulis diterima di
Program Studi Pemuliaan dan Bioteknologi Tanaman, Sekolah Pasca Sarjana,
Institut Pertanian Bogor, dimana biaya sekolah dan penelitian diperoleh dari
Proyek Generation Challenge Program Capacity Building pada tahun 2007-2009 dan dilanjutkan beasiswa pendidikan dari Badan Litbang Pertanian, Kementerian
Pertanian pada tahun 2010-2011.
Penulis bekerja sebagai staf peneliti di Balai Besar Penelitian dan
Pengembangan Bioteknologi dan Sumberdaya Genetik Pertanian (BB Litbang
Biogen) sejak tahun 2003 dan ditempatkan di Kelompok Peneliti Biologi
Molekuler.
Selama mengikuti program S3, penulis mendapat kesempatan sebagai PhD scholar selama 4 bulan di International Rice Research Institute (IRRI). Satu karya
ilmiah yang merupakan bagian dari disertasi telah diterbitkan pada jurnal
Agrobiogen (2012).
Penulis menikah dengan Roberdi, SP. MSi. pada tahun 2007 dan dikaruniai
DAFTAR ISI
Beras sebagai Bahan Pangan Utama di Indonesia ... 9
Patogen Cendawan Pyricularia oryzae ... 11
Siklus Infeksi Pyricularia oryzae ... 12
Gejala Penyakit Blas pada Tanaman Padi ... 14
Variabilitas Genetik Cendawan Pyricularia oryzae ... 15
Ketahanan Tanaman Padi terhadap Penyakit Blas ... 17
Strategi Pemuliaan untuk Meningkatkan Ketahanan Tanaman Padi terhadap Penyakit Blas ... 20
VARIASI KETAHANAN TERHADAP BLAS PADA POPULASI SILANG BALIK LANJUT WAY RAREM x ORYZICA LLANOS-5 ... 27
Abstrak ... 27
STUDI PEWARISAN KETAHANAN ORYZICA LLANOS-5 TERHADAP PENYAKIT BLAS YANG DISEBABKAN OLEH Pyricularia oryzae... 41
DETEKSI SEGMEN INTROGRESI KETAHANAN BLAS DARI ORYZICA LLANOS-5 PADA POPULASI SILANG BALIK LANJUT
WAY RAREM x ORYZICA LLANOS-5 DENGAN MARKA SSR ... 63 Abstrak ... 63
Abstract ... 63 Pendahuluan ... 64 Bahan dan Metode ... 65 Hasil dan Pembahasan ... 69 Kesimpulan.. ... … 74 Daftar Pustaka ... 74
KARAKTERISASI INTROGRESI DONOR PADA POPULASI SILANG BALIK LANJUT WAY RAREM x ORYZICA LLANOS-5
MENGGUNAKAN MARKA SNP ... 77 Abstrak ... 77
Abstract ... 77 Pendahuluan ... 78 Bahan dan Metode ... 80 Hasil dan Pembahasan ... 83
Kesimpulan………. 87
Daftar Pustaka ... 88
DETEKSI LOKASI SEGMEN INTROGRESI DARI ORYZICA LLANOS-5 PADA GENOM TANAMAN BC6F1 TAHAN BLAS HASIL
SILANG BALIK LANJUT WAY RAREM x ORYZICA LLANOS-5 ... 91 Abstrak ... 91
Abstract ... 91 Pendahuluan ... 92 Bahan dan Metode ... 94 Hasil dan Pembahasan ... 95
Kesimpulan………...100
Daftar Pustaka ... ….101
PEMBAHASAN UMUM... 103
KESIMPULAN UMUM DAN SARAN ... 111 Kesimpulan Umum ... 111 Saran ... 112
DAFTAR TABEL
Halaman
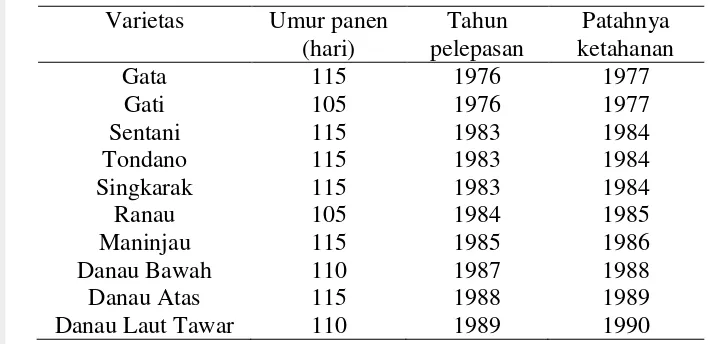
1. Varietas unggul padi gogo yang dilepas dari tahun 1976 sampai
1989 dan perubahan reaksinya terhadap blas pada kondisi lapang ... 21
2. Nisbah fenotipe frekuensi karakter ketahanan tanaman terhadap
patogen dalam populasi F2 ... 47
3. Jumlah tanaman pada setiap populasi persilangan Oryzica Llanos-5 x
Way Rarem berdasarkan persentase luas daunyang terserang ras blas 133 .... 50
4. Jumlah tanaman pada setiap populasi persilangan Oryzica Llanos-5 x
Way Rarem berdasarkan persentase luas daun yang terserang ras blas 173 ... 50
5. Nilai rata-rata persentase luas daun yang terserang ras blas 133 dan 173 serta hasil Uji Beda Nilai Tengah populasi F1 dan F1R persilangan
Oryzica Llanos-5 x Way Rarem... 52
6. Hasil Uji Skala, Uji Skala Gabungan kesesuaian model genetik dan nilai heritabilitas untuk ketahanan populasi persilangan Oryzica Llanos-5 x
Way Rarem terhadap ras blas 133 ... 57
7. Hasil Uji Skala, Uji Skala Gabungan kesesuaian model genetik dan nilai heritabilitas untuk ketahanan populasi persilangan Oryzica Llanos-5 x
Way Rarem terhadap ras blas 173 ... 58
8. Analisis marka tunggal terhadap 6 marka SSR hasil eksplorasi primer
SSR pada kromosom 3,6 dan 9 ... 72
9. Marka SNP yang polimorfik pada kedua tetua ... 83
10.Rata-rata panjang, ukuran dan proporsi segmen genom tetua berulang dan tetua donor pada turunan hasil silang balik ... 86
11.Analisis marka tunggal terhadap 2 marka SNP di kromosom 2 pada
DAFTAR GAMBAR
Halaman
1. Diagram alir penelitian ... 8
2. Konidia cendawan blas hasil pengamatan menggunakan mikroskop pada perbesaran 100x ... 30
3. Skoring serangan blas berdasarkan Sistim Standar Evaluasi dari
IRRI (1996) ... 31
4. Variasi reaksi serangan blas pada individu BC3F1 Way Rarem x
Oryzica Llanos-5 ... 32
5. Persentase jumlah tanaman pada populasi BC3F1, Way Rarem,
Oryzica Llanos-5 dan Kencana Bali berdasarkan skor ketahanan blas ... 33
6. Persentase jumlah tanaman pada populasi BC4F1, Way Rarem,
Oryzica Llanos-5 dan Kencana Bali berdasarkan skor ketahanan blas ... 33
7. Persentase jumlah tanaman pada populasi BC3F1 dan BC4F1, Way Rarem
Oryzica Llanos-5 dan Kencana Bali berdasarkan skor ketahanan blas ... 34
8. Persentase jumlah tanaman pada populasi BC5F1, Way Rarem,
Oryzica Llanos-5 dan Kencana Bali berdasarkan pengujian dengan
ras blas 133 ... 35
9. Persentase jumlah tanaman pada populasi BC5F1, Way Rarem,
Oryzica Llanos-5 dan Kencana Bali berdasarkan pengujian dengan
dengan ras blas 173 ... 36
10.Persentase jumlah tanaman Way Rarem, Oryzica Llanos-5 dan
Kencana Bali berdasarkan pengujian dengan ras blas 033 ... 37
11.Skema posisi relatif nilai tengah F1 terhadap kedua tetuanya berdasarkan
persentase luas daun yang terserang ... 52
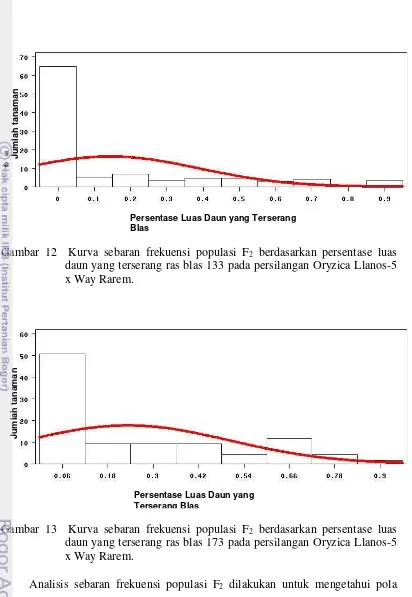
12.Kurva sebaran frekunsi populasi F2 berdasarkan persentase luas daun yang
terserang ras blas 133 pada persilangan Oryzica Llanos-5 x Way Rarem ... 53
13.Kurva sebaran frekuensi populasi F2 berdasarkan persentase luas daun yang
terserang ras blas 173 pada persilangan Oryzica Llanos-5 x Way Rarem ... 53
14.Hasil amplifikasi DNA tanaman dengan RM541 ... 70
15.Deteksi introgresi segmen Oryzica Llanos-5 pada BC3F1 dengan
16.Deteksi introgresi segmen Oryzica Llanos-5 pada BC4F1 dengan
marka SSR ... 73
17.Skema pemuliaan silang balik... 81
18.Marka SNP pada 12 kromosom padi yang digunakan untuk populasi
silang balik BC4F2 dan BC2F7 ... 82
19.Grafik genotipe pada populasi hasil silang balik ... 85
20.Grafik genotipe berdasarkan marka SSR pada tanaman BC6F1 ... 96
21.Grafik genotipe berdasarkan marka SSR pada populasi RIL Fanny x
Oryzica Llanos-5 ... 97
22.Grafik genotipe kromosom 2, 3, 6 dan 8 yang berdasarkan marka SNP
DAFTAR LAMPIRAN
Halaman
PENDAHULUAN
Latar Belakang
Beras merupakan makanan pokok bagi penduduk negara berkembang di
dunia, dimana ketersediaan pangan menjadi masalah utama. Di Indonesia usaha
peningkatan produksi padi nasional terus dilakukan oleh pemerintah seiring
dengan meningkatnya kebutuhan beras setiap tahunnya. Data Badan Pusat
Statistik (BPS) menunjukkan produksi padi pada tahun 2010 mencapai 66.46 juta
ton Gabah Kering Giling (GKG), meningkat sebesar 10% dibandingkan produksi
padi pada tahun 2008 yang mencapai 60.33 juta ton. Kenaikan produksi ini terjadi
karena peningkatan luas panen padi sawah dan padi gogo di pulau Jawa dan luar
Jawa sebesar 926 ribu hektar (1.48%) (Kementan 2011).
Padi gogo merupakan salah satu alternatif dalam usaha peningkatan
produksi beras di Indonesia. Secara umum budidaya padi gogo dilakukan petani
pada: a) lahan terbuka (ladang/tradisional) dan sekitar bantaran sungai, b) sekitar
perbukitan Daerah Aliran Sungai (DAS), dan c) ditumpangsarikan dengan
tanaman perkebunan dan hutan tanaman industri yang umurnya masih muda.
Namun penerapan teknologi budidaya padi gogo belum optimal, terutama dalam
penggunaan varietas unggul dan pengendalian penyakit blas (Toha 2007b).
Penyakit blas disebabkan oleh cendawan Pyricularia oryzae yang mampu merusak tanaman padi hampir pada semua bagian tanaman seperti daun, nodus,
malai dan bulir (Crill et al. 1982). Penularan penyakit blas yang berat terutama saat fase generatif yang terjadi di malai dan bulir dapat menggagalkan panen
(Amir & Nasution 1994). Cendawan P. oryzae memiliki variasi genetik yang tinggi sehingga mudah untuk membentuk ras baru yang mempunyai tingkat
virulensi yang lebih tinggi. Ras baru tersebut dapat dengan cepat mematahkan
ketahanan varietas tanaman yang sebelumnya tahan terhadap patogen blas (Ou
1985). Berdasarkan hal itu maka perlu dilakukan langkah-langkah
penanggulangan penyakit blas antara lain melalui pemakaian pupuk berimbang,
pengaturan waktu tanam, pengendalian secara kimiawi dengan penggunaan
fungisida (Semangun 2006) dan penanaman varietas unggul tahan patogen blas
diketahui sebaiknya disesuaikan dengan populasi ras cendawan blas yang
berkembang di lapangan (Ou 1985). Oleh karena itu, informasi mengenai
keragaman populasi ras di daerah endemik sangat diperlukan.
Sejak tahun 1960 banyak varietas unggul padi gogo yang telah dilepas dan
umumnya memiliki ketahanan terhadap patogen blas. Sebagian besar diantaranya
berubah menjadi peka (ketahanannya terhadap blas patah) setelah ditanam secara
luas selama 2-3 musim tanam (Amir & Nasution 1994). Akibatnya banyak petani
padi gogo yang kembali menerapkan sistem pertanaman tradisional dengan
menggunakan padi lokal. Padi lokal pada suatu pertanaman umumnya beragam
(heterogen) sehingga tidak pernah tertular berat oleh blas namun memiliki hasil
panen yang rendah (Suwarno et al. 2008).
Menurut Mundt (2002) dalam penanggulangan patogen blas terdapat hal
lain yang penting untuk diperhatikan yaitu adanya keragaman genetik berupa
keragaman sifat ketahanan blas pada suatu wilayah pertanaman padi. Hasil
penelitian Suwarno et al. (2008) menunjukkan bahwa terdapat kurang lebih 24 galur yang telah diseleksi memiliki ketahanan blas dan daya hasil tinggi.
Galur-galur tersebut merupakan materi tanaman untuk membuat paket 20 varietas padi
gogo. Paket 20 varietas padi gogo yang memiliki keragaman ketahanan terhadap
blas digunakan untuk pertanaman secara multivarietas atau pergiliran varietas
dalam rangka menanggulangi penyakit blas. Selain itu, hal lain yang dapat
mencegah patahnya ketahanan varietas terhadap blas adalah dengan menggunakan
varietas multilini (Mundt 2002, Abe 2004, Kwon et al. 2008).
Pada tahun 2000 International Rice Research Institute (IRRI) membentuk konsorsium untuk penelitian pemuliaan padi gogo (Upland Rice Research Consortium), dimana Indonesia menjadi salah satu anggotanya. Pembentukan konsorsium bertujuan untuk memfasilitasi pertukaran benih padi gogo di antara
anggotanya dan melakukan identifikasi galur-galur padi gogo hasil persilangan
yang memiliki ketahanan terhadap blas dan daya hasil tinggi yang dapat
diterapkan oleh anggota konsorsium. Selain itu, galur-galur ini memiliki manfaat
lain yaitu: 1) menjadi sumber gen ketahanan untuk digunakan sebagai tetua
persilangan, 2) dievaluasi lebih lanjut dalam kegiatan pemetaan QTL
sebagai tetua donor untuk pengembangan populasi silang balik lanjut (advanced backcross lines) sehingga didapatkan galur-galur NIL (Near isogenic lines) tahan
blas untuk pembentukan varietas multilini (Suwarno et al. 2001a).
Silang balik merupakan teknik persilangan yang dapat diterapkan dalam
pemuliaan tanaman untuk mentransfer satu atau beberapa gen yang diinginkan
dari tetua donor kepada tetua berulang. Melalui pengembangan populasi dengan
beberapa kali silang balik dapat dihasilkan populasi NIL. Galur-galur NIL yang
didapatkan memiliki introgresi segmen genom dari tetua donor yang memberikan
sifat tambahan seperti misalnya ketahanan terhadap stres biotik, yang tidak
terdapat pada tetua berulang (recurrent parent). Oleh karena itu NIL yang dihasilkan memiliki kesamaan genom dengan tetua berulang namun berbeda
hanya pada satu segmen saja yang membawa sifat ketahanan terhadap patogen
tertentu dari tetua donor (Collard et al. 2005).
Penelitian mengenai pengembangan populasi NIL padi untuk
menanggulangi patogen blas telah banyak dilakukan, umumnya pada padi
subspesies Japonica seperti yang dilakukan di Jepang (Abe 2004) dan di Korea (Kwon et al. 2008). Sebaliknya penelitian NIL pada padi subspesies Indica, yang merupakan tipe padi di Indonesia, belum banyak dilakukan. Populasi NIL juga
bermanfaat dalam pengembangan varietas diferensial untuk penentuan ras pada
isolat blas seperti pada padi Japonica (Telebanco-Yanoria et al. 2010) dan padi
Indica (Telebanco-Yanoria et al. 2011).
Populasi NIL dapat digunakan untuk identifikasi gen yang berperan dalam
ketahanan terhadap patogen blas. Cara mengidentifikasi gen tersebut adalah
dengan membandingkan antara ekspresi alel ketahanan pada NIL dengan ekspresi
alel rentan tetua berulang yang memiliki latar belakang genetik yang sama dengan
NIL (McCouch et al. 1994). Melalui hasil identifikasi dapat dipilih gen-gen ketahanan yang cocok untuk program pengembangan suatu varietas padi yang
baru dan varietas padi yang telah ada (Chen et al. 1996).
Secara umum pengembangan populasi NIL melalui pemuliaan konvensional
membutuhkan proses yang rumit dan waktu yang lama. Seiring dengan
perkembangan teknologi maka digunakan bantuan marka molekuler untuk
mengidentifikasi segmen genom tetua donor yang diinginkan dan mengetahui
besarnya proporsi segmen genom tanaman yang telah kembali ke tetua penerima
pada galur-galur NIL (Hospital 2005). Marka molekuler juga membantu
menyeleksi segmen genom tanaman yang diinginkan dan tidak diinginkan dari
tetua donor selama prosedur silang balik dilakukan (Sleper & Poehlman 2006).
Sejumlah marka molekuler telah digunakan dalam pemuliaan tanaman, salah
satunya adalah SSR (Simple sequence repeats). SSR merupakan salah satu marka yang paling banyak digunakan untuk pemetaan genetik, analisis keragaman
genetik, dan studi evolusi. Marka ini muncul sebagai marka yang sangat variatif
(hipervariable), mudah diulang (reproducible) dan mudah dikerjakan karena berbasis teknik PCR (polymerase chain reaction). Hal ini menjadikannya sangat ideal untuk pemetaan genom (genome mapping) (McCouch et al. 1997).
Pada penelitian sebelumnya, Bustamam et al. (2001) menggunakan Oryzica Llanos-5, varietas tahan blas asal Kolombia, sebagai tetua donor untuk
meningkatkan ketahanan blas varietas padi gogo Indonesia, Way Rarem. Oryzica
Llanos-5 (IRGC 117017) termasuk padi gogo jenis indica yang berasal dari Kolombia. Oryzica Llanos-5 (OL5) merupakan varietas komersial populer yang
memiliki ketahanan terhadap blas yang tidak terpatahkan sejak dilepas tahun 1989
di Kolombia (Correa-Victoria & Zeigler 1995). Varietas ini kemungkinan
memiliki beberapa gen ketahanan terhadap blas (R gene) yaitu gen Pi2, Piz, Pi33,
Pita2, Pib, Pik dan Pish. Berdasarkan studi QTL ketahanan blas didapatkan
beberapa QTL ketahanan blas yang berasal dari alel Oryzica Llanos-5 pada
populasi hasil persilangan antara kultivar Fanny x Oryzica Llanos-5
(Lopez-Gerena 2006).
Tetua berulang yang digunakan merupakan padi gogo varietas unggul dari
Indonesia yang sudah dilepas yaitu Way Rarem. Varietas ini termasuk jenis indica
dan memiliki ketahanan terhadap cekaman aluminium (Al) sehingga
direkomendasikan untuk ditanam di daerah tegalan atau kering yang memiliki
tanah podsolik merah-kuning. Way Rarem tahan terhadap blas saat pertama kali
dilepas tahun 1994 (Puslitbangtan 1999) namun ketahanannya mulai terpatahkan
beberapa tahun setelah dilepas sehingga membutuhkan usaha pengembangan lebih
Pada penelitian sebelumnya (Bustaman et al. 2001) telah dilakukan persilangan antara Way Rarem dan tetua donor, Oryzica Llanos-5 yang
menghasilkan populasi F1, BC1F1 dan BC2F1. Selain itu, telah dilakukan juga
seleksi terhadap marka RFLP (Restriction Fragment Length Polymorphism) dan SSR untuk mendapatkan marka molekuler yang polimorfik terhadap kedua tetua,
Way Rarem dan Oryzica Llanos-5. Selanjutnya Lestari et al. (2011) menggunakan marka SSR yang polimorfik untuk membuat pemetaan QTL ketahanan blas pada
generasi BC2F2 Way Rarem x Oryzica Llanos-5.
Tujuan Penelitian
1. Mendapatkan galur-galur NIL Way Rarem x Oryzica Llanos-5 yang memiliki
ketahanan blas lebih tinggi dibandingkan dengan tetua berulangnya, Way
Rarem.
2. Memperoleh informasi tentang pola pewarisan sifat ketahanan terhadap
patogen blas pada varietas Oryzica Llanos-5.
3. Mendapatkan marka molekuler yang mampu mendeteksi introgresi segmen
genom dari Oryzica Llanos-5 yang terkait sifat ketahanan terhadap patogen
blas pada generasi silang balik lanjut Way Rarem x Oryzica Llanos-5.
Hipotesis Penelitian
1. Terdapat galur-galur NIL Way Rarem x Oryzica Llanos-5 yang memiliki
ketahanan terhadap patogen blas lebih tinggi dibandingkan dengan tetua
berulangnya, Way Rarem.
2. Sifat ketahanan Oryzica Llanos-5 terhadap patogen blas merupakan sifat
kuantitatif dengan aksi gen aditif dominan.
3. Terdapat marka molekuler yang dapat mendeteksi segmen introgresi
ketahanan terhadap patogen blas dari Oryzica Llanos-5 pada populasi silang
balik lanjut Way Rarem x Oryzica Llanos-5.
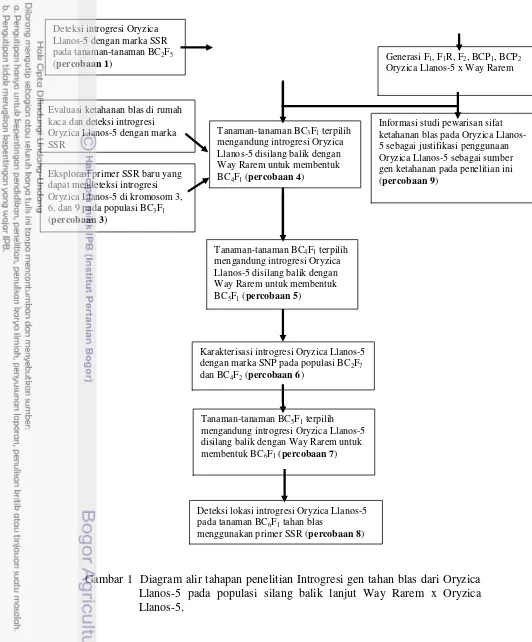
Penelitian dilakukan secara bertahap yang terdiri atas beberapa percobaan,
yaitu:
Percobaan 1:
Melakukan konfirmasi marka molekuler menggunakan 13 marka SSR yang
merupakan hasil penelitian Lestari et al. (2011). Dari 13 marka SSR tersebut, terdapat 3 marka SSR (RM148, RM541 dan RM342B) yang terpaut dengan
QTL ketahanan blas dari alel Oryzica Llanos-5 pada kromosom 3, 6 dan 9.
Konfirmasi ini dilakukan pada populasi BC2F5 Way Rarem x Oryzica Llanos-5
untuk menyeleksi tanaman-tanaman yang diduga memiliki gen ketahanan
terhadap patogen blas.
Percobaan 2:
Melakukan silang balik tanaman-tanaman BC2F5 Way Rarem x Oryzica
Llanos-5 yang terpilih hasil konfirmasi dengan marka SSR dengan tetua
penerima Way Rarem untuk mendapatkan benih BC3F1. Percobaan 3:
Melakukan silang balik tanaman-tanaman BC3F1 yang terpilih tahan blas
melalui pengujian fenotip dan telah dikonfirmasi menggunakan marka SSR
dengan Way Rarem untuk mendapatkan benih BC4F1. Percobaan 4:
Melakukan eksplorasi primer SSR pada kromosom 3, 6, dan 9 yang dapat
mendeteksi segmen introgresi dari Oryzica Llanos-5 pada populasi BC3F1. Dari
hasil eksplorasi akan dipilih marka SSR baru yang polimorfik yaitu yang dapat
menghasilkan larik yang berbeda dari kedua tetua, Way Rarem dan Oryzica
Llanos-5 dan dapat mendeteksi introgresi dari Oryzica Llanos-5 pada populasi
hasil silang balik. Marka SSR baru yang terpilih digunakan untuk konfirmasi
molekuler pada populasi BC4F1, BC5F1 dan BC6F1 yaitu RM411, RM7,
RM251, RM276, RM549 dan RM585.
Percobaan 5:
Melakukan silang balik tanaman-tanaman BC4F1 yang telah dievaluasi
ketahanannya terhadap blas dan dikonfirmasi menggunakan marka SSR
Melakukan karakterisasi terhadap segmen introgresi dari Oryzica Llanos-5
pada populasi BC2F7 dan BC4F2 Way Rarem x Oryzica Llanos-5 menggunakan
marka SNP (Single nucleotide polymorphism). Melalui karakterisasi ini dapat diketahui adanya perbedaa panjang dan proporsi segmen introgresi donor pada
populasi BC2F7 dan BC4F2. Percobaan 7:
Melakukan silang balik tanaman-tanaman BC5F1 yang telah dievaluasi
ketahanannya terhadap blas dan dikonfirmasi menggunakan marka SSR
dengan Way Rarem untuk mendapatkan benih BC6F1. Percobaan 8:
Melakukan identifikasi lokasi introgresi dari Oryzica Llanos-5 pada genom
tanaman BC6F1 tahan blas yang memiliki indeks penyakit 1 menggunakan 400
primer SSR yang tersebar pada 12 kromosom.
Percobaan 9:
Melakukan studi pewarisan sifat ketahanan Oryzica Llanos-5 terhadap patogen
blas dengan cara membuat populasi F1, F1 resiprokal (F1R), F2, BCP1 (hasil
silang balik dengan tetua pertama) dan BCP2 (hasil silang balik dengan tetua
kedua) dari persilangan Oryzica Llanos-5 x Way Rarem. Masing-masing
populasi dievaluasi ketahanannya terhadap ras blas 133 dan 173 kemudian
diambil data ketahanannya berupa persentase luas daun yang terserang blas.
Secara lengkap tahapan penelitian dan jenis percobaan yang dilakukan
ditunjukkan pada Gambar 1.
Tanaman-tanaman BC2F5 terpilih
mengandung introgresi Oryzica Llanos-5 disilang balik dengan Way Rarem untuk membentuk BC3F1 (percobaan 2)
Gambar 1 Diagram alir tahapan penelitian Introgresi gen tahan blas dari Oryzica dengan marka SNP pada populasi BC2F7
dan BC4F2 (percobaan 6)
menggunakan primer SSR (percobaan 8)
Generasi F1, F1R, F2, BCP1, BCP2
Tanaman-tanaman BC3F1 terpilih
mengandung introgresi Oryzica Llanos-5 disilang balik dengan Way Rarem untuk membentuk BC4F1 (percobaan 4)
Tanaman-tanaman BC4F1 terpilih
mengandung introgresi Oryzica Llanos-5 disilang balik dengan Way Rarem untuk membentuk BC5F1 (percobaan 5)
Tanaman-tanaman BC5F1 terpilih
TINJAUAN PUSTAKA
Beras sebagai bahan pangan utama di Indonesia
Padi (Oryza sativa) termasuk dalam genus Oryza, yang didalamnya terdapat lebih dari 25 spesies liar. Indica dan Japonica adalah subspesies Oryza sativa
yang banyak ditanam dan dikonsumsi di dunia. Padi telah dibudidayakan di India
dan Cina Selatan sejak kurang lebih 8000 tahun yang lalu dan saat ini merupakan
salah satu bahan pangan utama di dunia termasuk di Indonesia (IRRI 2002).
Di Indonesia padi merupakan makanan pokok lebih dari 95% rakyat
Indonesia. Penyediaan pangan terutama beras, dalam jumlah yang cukup dan
harga terjangkau tetap menjadi prioritas utama pembangunan nasional. Padi juga
telah menjadi sumber mata pencaharian dengan menyediakan lapangan kerja bagi
sekitar 20 juta rumah tangga petani di pedesaan (Departemen Pertanian 2008).
Produksi padi perlu ditingkatkan untuk memenuhi kebutuhan pangan
penduduk Indonesia yang terus bertambah dari tahun ke tahun. Di sisi lain,
tantangan yang dihadapi dalam pengadaan produksi padi semakin berat. Laju
pertumbuhan penduduk dan tingkat konsumsi beras yang relatif masih tinggi
menuntut peningkatan produksi yang berkesinambungan, sementara sebagian
besar lahan sawah yang subur telah beralih fungsi untuk usaha lainnya seperti
pemukiman penduduk, kawasan industri dan infrastruktur. Perubahan iklim global
juga menjadi ancaman bagi upaya peningkatan produksi pangan. Ancaman
kekeringan dan kebanjiran di sudah semakin sering melanda pertanaman petani
(Departemen Pertanian 2008).
Berdasarkan data dari Badan Pusat Statistik diketahui bahwa pada tahun
2010 produksi padi di Jawa sebesar 36.3 juta ton, mengalami kenaikan sebesar
1.49 juta ton (4.3 %) dibandingkan pada tahun 2009. Produksi padi di luar Jawa
sebesar 30.1 juta ton, mengalami kenaikan sebesar 575 ribu (2 %) dibandingkan
pada tahun 2009. Hal ini terjadi karena adanya peningkatan luas panen padi sawah
sebesar 322 ribu ha (2.7%) dan luas panen padi gogo sebesar 49 ribu ha (4.5%)
pada tahun 2010 (Kementan 2011).
Peningkatan luas panen tersebut terjadi antara lain disebabkan adanya
adalah hamparan lahan yang tidak pernah tergenang atau digenangi air sepanjang
waktu. Berbeda dengan lahan sawah yang tingkat kesuburannya relatif tinggi,
seragam, dan sering tergenang, lahan kering umumnya memiliki tingkat
kesuburan yang rendah dan beragam, kandungan bahan organik rendah dan sulit
dipertahankan sehingga produktivitasnya cepat menurun. Selain itu lahan tersebut
bergantung pada curah hujan sehingga suplai air tidak konsisten (Toha 2007b).
Luas lahan kering di Indonesia sekitar 25.09 juta ha, 35% di antaranya
sesuai untuk usaha tani tanaman semusim seperti padi gogo dan jagung. Luas
lahan kering yang potensial untuk pengembangan padi gogo diperkirakan sekitar 7
juta ha (Sukarman et al. 2008). Lahan kering yang tersedia untuk perluasan areal tanaman pangan di Indonesia sebagian besar berupa tanah masam. Pengembangan
lahan kering yang saat ini merupakan kantong-kantong kemiskinan dapat
dijadikan salah satu alternatif dalam upaya pengadaan pangan di masa depan, dan
diharapkan dapat meningkatkan ketahanan pangan nasional (Toha 2007b).
Produksi padi gogo di Indonesia pada tahun 2010 sebesar 3.45 juta ton atau
meningkat sebesar 9.3 % dibandingkan pada tahun 2008. Hasil produksi padi
gogo lebih banyak di luar Jawa yaitu sebesar 1.9 juta ton dibandingkan produksi
di Jawa sebesar 1.5 juta ton. Dengan demikian padi gogo menyuplai sebesar 5.2 %
dari keseluruhan produksi padi di Indonesia (Kementan 2011).
Dalam usaha peningkatan produksi beras di Indonesia padi gogo merupakan
alternatif setelah padi sawah. Secara umum budidaya padi gogo dilakukan petani
pada: a) lahan terbuka (ladang/tradisional) dan sekitar bantaran sungai, b) sekitar
perbukitan daerah aliran sungai (DAS), dan c) ditumpangsarikan dengan tanaman
perkebunan dan hutan tanaman industri yang umurnya masih muda. Kondisi lahan
kering yang merupakan tempat penanaman padi gogo perlu dikelola agar dapat
memberikan produksi yang baik (Lubis et al. 1994). Pengelolaan Tanaman Terpadu (PTT) merupakan pendekatan dalam usaha meningkatkan produktivitas
dan efisiensi usahatani padi. PTT padi gogo di lahan kering yang perlu diterapkan
yaitu: 1) penambahan bahan organik tanah, 2) pemupukan berimbang berdasarkan
status kesuburan tanah, dan 3) efisiensi pemupukan dengan memberi pupuk secara
larikan dan pada waktu yang tepat (Sembiring & Widiarta 2008).
padi gogo yaitu penyakit blas. Serangan patogen blas pada padi lahan kering lebih
berat dibandingkan dengan padi lahan sawah. Faktor lingkungan berupa
kelembaban yang cukup tinggi terkait erat dengan kerentanan padi gogo terhadap
blas (Amir & Kardin 1991). Salah satu strategi utama dalam pengendalian blas
adalah dengan penggunaan varietas tahan patogen blas (Santoso et al. 2007).
Patogen cendawan Pyricularia oryzae
Pyricularia oryzae Cav. (teleomorf Magnaporthe oryzae) merupakan patogen penyebab penyakit blas pada padi dan 50 spesies tanaman lain dalam
genus Gramineae (Ou 1985). Cendawan blas memiliki beberapa penamaan yang didasarkan pada fase perkembangannya. P.oryzae mengacu pada fase perkembangan aseksual (anamorf) dari cendawan blas yang banyak ditemukan di
lapang. Fase perkembangan seksual (teleomorf) dari cendawan blas dinamakan M. oryzae yang ditemukan secara kultur dalam laboratorium.
Pada mulanya terdapat dua nama spesies yang diberikan kepada anamorf
dari M. grisea yaitu P. grisea dan P. oryzae Cavara. P. grisea berasal dari tanaman inang Digitaria sanguinalis dan P. oryzae Cavara berasal dari tanaman inang Oryza sativa. Pengamatan secara morfologi menunjukkan bahwa tidak ada perbedaan antara P. grisea dengan P. oryzae (Ou 1985). Rossman et al. (1990) menyatakan bahwa P. grisea sebagai nama yang benar untuk fase perkembangan
aseksual patogen blas pada tanaman padi dan tanaman monokotil lainnya. Oleh
karena itu, nama P. grisea (teleomorf M. grisea) umum digunakan oleh peneliti di lapangan sebagai patogen penyebab penyakit blas pada padi.
Isolat Pyricularia terdiri atas empat kelompok utama yang dibedakan berdasarkan tanaman inangnya yaitu: 1) isolat yang berasal dari Digitaria ciliaris
dan Eragrostis sp., 2) isolat yang berasal dari Cenchrus echinatus L., 3) isolat yang berasal dari Cyperus brevifolius dan Cyperus rotundus, dan 4) isolat yang berasal dari Oryza sativa dan tanaman rumput lainnya (Borromeo et al. 1993). Berdasarkan data DNA mitokondria (mtDNA) dengan marka RFLP menunjukkan
bahwa isolat Pyricularia yang berasal dari tanaman inang Digitaria sp. dan
Berdasarkan pengamatan secara morfologi isolat M. grisea identik dengan hasil persilangan antar isolat yang berasal dari tanaman inang padi. Sebagian
isolat dari beberapa jenis rumput dapat melakukan persilangan dengan isolat yang
berasal dari padi, namun isolat yang berasal dari tanaman Digitaria tidak dapat melakukan persilangan dengan isolat yang berasal dari padi. Analisis koefisien
kesamaan dengan marka RFLP juga dilakukan antara kelompok isolat blas dari
tanaman padi dengan isolat blas dari Digitaria. Hasil analisis didapatkan bahwa tingkat kesamaan antara kedua isolat tersebut hanya sekitar 5% (Kato et al. 2000). Menurut Couch dan Kohn (2002) M. grisea yang berasal dari tanaman inang padi dan beberapa jenis rumput merupakan spesies yang berbeda dengan M. grisea
yang berasal dari tanaman inang Digitaria. Perbedaan tersebut terletak pada 3 lokus gen yaitu gen aktin, gen beta-tubulin dan gen kalmodulin. Selain itu
persilangan antar kedua isolat yang dilakukan di laboratorium bersifat infertil.
Dengan demikian hasil dari beberapa penelitian ini mematahkan pendapat
Rossman et al. (1990) serta menunjukkan bahwa M. oryzae adalah isolat yang berasal dari tanaman inang padi dan berbeda spesies dengan M. grisea yang berasal dari tanaman inang Digitaria.
Siklus infeksi Pyricularia oryzae
Tahapan perkembangan penyakit oleh cendawan P. oryzae dimulai dari
proses inokulasi, yaitu proses deposisi atau kontaknya inokulum pada tanaman
inang. Konidiofor menghasilkan konidia yang tersusun secara simpodial. Konidia
ini merupakan sumber inokulum untuk siklus infeksinya. Konidia memiliki spore tip mucilage (ujung yang berlendir) untuk dapat melekat dengan kuat pada permukaan organ tanaman inang. Lendir ini diproduksi selama perkembangan
konidia. Konidia tidak melakukan metabolisme aktif saat melekat pada
permukaan tanaman inang. Konidia selanjutnya berkecambah membentuk germ tube (tabung kecambah). Germ tube akan berdiferensiasi membentuk apresorium. Selama pematangan apresorium akan terbentuk lapisan melanin pada dinding
apresorium, kecuali pada pori apresorium yang menempel pada permukaan daun
padi. Pembentukan apresorium ini tidak memerlukan sinyal yang spesifik dari
Lapisan melanin merupakan lapisan semipermeabel dimana air dapat
berdifusi melalui melanin masuk ke dalam apresorium namun ion-ion dan
molekul kecil tidak dapat masuk atau keluar dari apresorium. Senyawa nutrisi
berupa karbohidrat dan lipid ditransportasikan dari konidium menuju apresorium.
Terjadi pemecahan karbohidrat dan lipid di dalam apresorium yang menghasilkan
peningkatan kosentrasi senyawa organik terutama gliserol. Peningkatan senyawa
gliserol mengakibatkan air masuk ke dalam apresorium melalui melanin sehingga
terbentuk tekanan turgor yang besar di apresorium (Ebbole 2007).
Setelah proses inokulasi, terjadi proses penetrasi yaitu masuknya patogen
blas ke dalam tanaman inang melalui terbentuknya penetration peg (kapak penetrasi) yang disebabkan tekanan turgor yang besar pada apresorium. Lapisan
melanin yang melapisi dinding apresorium tidak terbentuk pada pori apresorium.
Penetration peg terbentuk pada pori apresorium yang berada di dasar apresorium yang menempel pada lapisan kutikula tanaman inang. Penetration peg mampu menembus lapisan kutikula dan dinding sel tanaman inang karena tekanan turgor
yang terus meningkat pada apresorium (Howard & Valent 1996)
Tahapan selanjutnya adalah proses infeksi dimana cendawan blas telah
menetap di dalam sel dan mampu menyerap nutrisi dari inangnya. Pada tahap ini
kapak penetrasi berkembang menjadi hifa infeksi primer yang memiliki
tonjolan-tonjolan pada permukaannya dan bercabang-cabang di dalam lumen sel tanaman
inang. Hifa infeksi primer dapat keluar dari lumen sel tanaman tempat awal
infeksinya untuk mengkolonisasi sel lain yang berada di sebelahnya melalui pori
plasmadesmata lalu membentuk hifa infeksi sekunder. Hifa infeksi sekunder
memiliki lapisan pelindung pada ujungnya. Lapisan pelindung tersebut terdiri atas
membran lamela yang berfungsi untuk mensekresikan protein cendawan ke dalam
sitoplasma sel tanaman inang. Protein ini diduga berperan dalam mengubah
kandungan sel inang menjadi bentuk unit-unit yang dapat diserap dan diasimilasi
oleh cendawan sehingga terjadilah perluasan pertumbuhan cendawan dalam
tanaman inang (Ebbole 2007).
Selama penetration peg berkembang menjadi hifa infeksi primer, kerusakan yang terjadi pada jaringan tanaman belum dapat terlihat secara kasat mata. Sekitar
sel-sel tanaman inang, baru mulai terlihat gejala pada organ tanaman berupa bercak
luka kecil berbentuk oval. Sesaat setelah hifa infeksi sekunder terbentuk maka sel
tanaman tempat awal infeksi akan kehilangan viabilitasnya sehingga terjadi
plasmolisis. Hal ini berbeda dengan sel di sampingnya yang tetap stabil, tidak
kehilangan viabilitasnya karena masih terdapat hifa infeksi sekunder yang
berkembang di dalamnya (Ribot et al. 2008).
Saat cendawan blas telah merusak jaringan yang terkolonisasi sekitar 4
hingga 5 hari muncul bercak coklat lokal. Selanjutnya bercak luka berkembang
menjadi nekrotik dan meluas di organ tanaman inang. Koloni cendawan blas
dalam jaringan tanaman akan mengalami sporulasi untuk menghasilkan konidia
yang berperan dalam infeksi sekunder untuk memperluas serangan patogen blas
ke tanaman lain (Howard & Valent 1996).
Gejala penyakit blas pada tanaman padi
Cendawan P. oryzae dapat menginfeksi tanaman padi hampir pada semua organnya seperti daun (leaf blast), buku (node blast), leher malai (neck blast), malai (panicle blast), bulir (spikelet blast), dan kolar daun (leaf collar) (Semangun 2006). Infeksi cendawan blas pada kolar daun dapat segera menyebar
dan mematikan daun. Buku dan antar buku padi (internode blast) yang terinfeksi cendawan blas saat waktu pembungaan dapat mengakibatkan terbentuknya malai
padi yang berwarna putih atau patahnya batang pada buku yang terinfeksi. Pada
waktu pembungaan, cendawan blas dapat menginfeksi buku leher malai (node neck panicle) yang merupakan serangan paling merusak dan disebut blas leher malai (neck blast). Jika infeksi pada leher malai terjadi pada tahap perkembangan awal maka malai tidak akan terisi bulir sehingga malai masih tetap tegak. Jika
infeksi pada leher malai terjadi pada tahap perkembangan akhir maka malai akan
terisi bulir sebagian namun batang malai akan patah karena tidak mampu
menahan beban (Agrios 2005).
Gejala penyakit blas pada fase vegetatif dapat diamati pada daun berupa
bercak berbentuk belah ketupat dengan ujung runcing. Bagian tengah bercak
berwarna abu-abu atau keputihan dan bagian pinggirnya umumnya berwarna
akhirnya menutupi seluruh permukaan daun yang mengakibatkan kematian daun.
Pada fase generatif, gejala dapat berupa nekrosis berwarna coklat abu-abu pada
leher malai yang terserang, atau dapat juga terjadi pada cabang malai dan kulit
gabah (Amir & Kardin 1991).
Ukuran bercak bervariasi bergantung pada lingkungan dan ketahanan
varietas tanaman. Bercak pada varietas yang rentan dan berada di lingkungan
yang lembab memperlihatkan pinggiran yang berwarna coklat dengan sedikit
warna kuning (’halo area’), sedangkan pada varietas yang tahan bercak hanya
berupa titik coklat sebesar jarum atau lebih dan tidak berkembang lagi. Perbedaan
bentuk, warna, dan ukuran bercak dapat digunakan untuk membedakan ketahanan
suatu varietas (Ou 1985).
Perkembangan patogen blas dipengaruhi oleh faktor lingkungan antara lain
kesuburan tanah dan kelembaban udara. Pada wilayah dengan kelembaban udara
yang tinggi tanaman mengalami masa berembun yang lebih panjang (dew period). Kondisi ini sangat mendukung perkembangan patogen blas. Hujan lebat yang
terjadi dalam waktu singkat tidak membantu perkembangan patogen blas,
sebaliknya hujan rintik-rintik yang lama merupakan kondisi yang menguntungkan
bagi patogen blas untuk berkembang dan menginfeksi tanaman inang (Amir &
Nasution 1994).
Variabilitas genetik cendawan Pyricularia oryzae
Cendawan P. oryzae mempunyai keragaman genetik yang tinggi di alam. Banyak ras P. oryzae yang terdapat di alam dan ras-ras tersebut memiliki sifat patogenitas yang bervariasi. Keragaman genetik pada cendawan dapat terjadi
akibat adanya hibridisasi seksual, rekombinasi aseksual dan mutasi. Hibridisasi
seksual merupakan peristiwa rekombinasi gen sebagai akibat pencampuran secara
acak kromosom dua cendawan induk dan pindah silang (crossing over) selama meiosis. Sebagai akibatnya dihasilkan genotipe keturunannya yang berbeda
dengan genotipe kedua induknya. Diperkirakan banyak ras baru cendawan
patogen yang muncul karena hibridisasi seksual (Semangun 2006). Hibridisasi
melalui metode pengkulturan di laboratorium (Notteghem & Silue 1992).
Zhang dan Zhou (2006) melaporkan bahwa fusi hifa dan rekombinasi
aseksual dapat terjadi pada P. oryzae dan berkontribusi pada peningkatan variabilitas genetik P. oryzae. Beberapa proses rekombinasi aseksual yang terjadi pada cendawan adalah: 1) heterokariosis, 2) rekombinasi paraseksual, dan 3)
heteroploidi. Banyak jenis cendawan yang diketahui memiliki tingkat variasi
genetik yang tinggi sebagai akibat heterokariosis. Sel heterokariotik adalah sel
yang mengandung dua atau lebih inti yang secara genetik berbeda. Hifa cendawan
dapat memiliki sejumlah sel heterokariotik, di samping sel-sel yang homokariotik
(mempunyai inti yang identik secara genetik). Keadaan heterokariotik dapat
terjadi karena: a) terjadinya fusi atau anastomosis hifa-hifa yang menyebabkan
tercampurnya inti yang berbeda ke dalam miselium dan b) terjadinya mutasi pada
hifa homokariotik (Semangun 2006).
Salah satu rekombinasi aseksual yang terjadi pada cendawan adalah
rekombinasi paraseksual. Rekombinasi paraseksual adalah proses dimana
rekombinasi genetik terjadi antara dua hifa yang heterokarion serta tidak
melibatkan segregasi meiosis (Clutterbuck 1996). Pertukaran genetik melalui
rekombinasi paraseksual dapat terjadi pada P. oryzae dan terdeteksi di lapang (Zeigler et al. 1997). Rekombinasi paraseksual berperan dalam meningkatkan variasi genetik P. oryzae (Zhang & Zhou 2006) dan kemampuan P. oryzae untuk
bertahan hidup pada kondisi lingkungan yang tidak optimal (Noguchi et al. 2007) sehingga memperluas spektrum patogenitas P. oryzae terhadap tanaman inangnya (Noguchi et al. 2006).
Peningkatan variasi genetik pada cendawan juga dapat melalui mutasi.
Mutasi adalah perubahan yang bersifat genetik berupa penambahan atau
pengurangan pada satu basa atau lebih pada untaian nukleotida DNA. Pada
kebanyakan mutasi perubahan terjadi pada gen individual, meskipun
kadang-kadang juga terjadi pada kromosom. Pada P.oryzae frekuensi mutasi dapat meningkat karena adanya agen fisik maupun kimiawi seperti adanya elemen
transposon POT2 dan MAGGY dalam genomnya (Nakayashiki et al. 1999).
mematahkan ketahanan suatu varietas. Lebih dari 30 ras P. oryzae yang teridentifikasi di Indonesia menggunakan tujuh varietas diferensial Indonesia
(Mogi et al. 1992). Varietas unggul yang sebelumnya memiliki ketahanan terhadap blas dapat berubah menjadi peka setelah ditanam secara luas selama 2-3
musim tanam (Amir & Nasution 1994).
Ketahanan tanaman padi terhadap patogen blas
Tanaman yang tahan terhadap patogen tertentu disebabkan oleh: 1) tanaman
tidak termasuk dalam kelompok taksonomi tanaman inang bagi patogen sehingga
patogen tidak dapat tumbuh pada tanaman tersebut (nonhost resistance), 2) tanaman memiliki gen-gen ketahanan yang dapat membangun mekanisme
pertahanan tanaman saat patogen masuk ke dalam tanaman sehingga pertumbuhan
patogen terhambat (true resistance), dan 3) tanaman memiliki struktur pelindung atau senyawa kimia sebagai pertahanan tanaman yang dapat menghalangi
keberhasilan infeksi patogen. Tanaman yang merupakan inang suatu patogen dan
bersifat rentan dapat juga terhindar dari serangan patogen akibat faktor
lingkungan atau faktor fisiologi tanaman (disease escape) (Semangun 2006). Ketahanan tanaman terhadap patogen blas berdasarkan cara kerja gen
tanaman inang yang dapat mempengaruhi perkembangan patogen terbagi menjadi
dua macam yaitu ketahanan vertikal dan ketahanan horisontal. Umumnya gen
tanaman yang mengatur ketahanan vertikal maupun horisontal berada di inti sel.
Selain itu terdapat juga beberapa ketahanan terhadap penyakit tanaman yang
dikontrol oleh gen yang berada di sitoplasma. Ketahanan seperti ini disebut
dengan ketahanan sitoplasmik (Ballini et al. 2008).
Ketahanan vertikal terjadi pada kondisi tanaman inang tidak kompatibel
terhadap patogen. Saat patogen akan menginfeksi tanaman maka patogen akan
mengeluarkan senyawa protein tertentu. Protein patogen tersebut mengaktifkan
sistem pertahanan tanaman berupa protein ketahanan (R protein) yang menghambat perkembangan awal inokulum atau menghambat perkembangbiakan
patogen setelah terjadi infeksi (Agrios 2005).
Sistem pertahanan tanaman pada ketahanan vertikal berupa terekspresinya
umumnya merupakan gen mayor (Silue et al. 1992). Gen R umumnya mengkode protein yang memiliki domain N-terminal nucleotide binding (NB) dan C-terminal leucine-rich repeats (LRRs). Sekitar 50 gen R untuk ketahanan penyakit
blas pada tanaman padi telah berhasil ditemukan dan tersebar di 12 kromosom
padi. Lima belas gen R di antaranya telah berhasil dilakukan fine mapping yaitu
IPi, IPi3, Pb1, Pi15, Pi24, Pi25(t), Pi33, Pi34, Pi39, Pi42, Pi5, Pigm(t), Pik, Pi-x(t), Pi-y1(t), dan Pi-y2(t) (Ballini et al. 2008). Terdapat beberapa gen R yang telah berhasil diklon yaitu Pi37 di kromosom 1 (Lin et al. 2007), Pib di kromosom 2 (Wang et al. 1999), Pi9 (Qu et al. 2006), Pi2/Pizt (Zhou et al. 2006) dan Pid2 (Chen et al. 2006) di kromosom 6; Pi36 di kromosom 8 (Liu et al. 2007), Pikh (Sharma et al. 2005) dan Pikm (Ashikawa et al. 2008) di kromosom 8, serta Pita di kromosom 12 (Bryan et al. 2000). Umumnya gen R mengekspresikan ketahanan terhadap ras patogen yang spesifik, namun
berdasarkan pengkajian yang terus dilakukan ternyata ditemukan bahwa beberapa
gen R dapat juga mengekspresikan ketahanan dengan spektrum yang luas, artinya
dengan ketahanan yang dimiliki dapat mengatasi beberapa ras patogen sekaligus.
Sebagai contoh adalah gen Pi40(t) (Jeung et al. 2007).
Ketahanan horisontal pada tanaman umumnya diatur oleh ekspresi dari
banyak gen (poligenik) dan mampu mengatasi beberapa ras patogen sekaligus.
Berbeda dengan ketahanan vertikal, ketahanan horisontal terjadi pada kondisi
tanaman inang kompatibel terhadap patogen dimana patogen dapat masuk ke
dalam tanaman dan menginfeksinya. Dengan ketahanan yang dimilikinya,
tanaman dapat memperlambat perluasan infeksi patogen sehingga memperlambat
penyebaran penyakit dan perkembangan epidemi pada lahan pertanaman tersebut
(Agrios 2005).
Varietas dengan ketahanan horisontal umumnya memiliki gen-gen
pengontrol sifat ketahanan yang merupakan gen-gen minor yang memberikan sifat
ketahanan akumulatif dan stabil. Umumnya gen-gen tersebut dapat diketahui
Sawata-Zenbayashi et al. 2007). Beberapa gen mayor lain yang juga mengatur ketahanan kuantitatif pada tanaman antara lain Pif dan Pb1 (Fujii et al. 2000).
Varietas dengan ketahanan vertikal umumnya menunjukkan ketahanan
penuh yang stabil terhadap ras patogen yang spesifik. Jika terjadi mutasi pada
patogen yang menyebabkan terbentuknya ras baru maka ras baru tersebut dapat
menginfeksi varietas yang sebelumnya tahan. Komposisi ras-ras yang membentuk
populasi patogen di suatu wilayah dapat berubah. Ras-ras patogen yang
sebelumnya mendominasi dapat digantikan oleh ras-ras patogen yang lain.
Perubahan ras dominan ini dapat juga menyebabkan varietas yang sebelumnya
tahan dengan ras tertentu akhirnya dapat terserang oleh ras patogen yang lain
(Suwarno et al. 2008).
Suatu varietas yang memiliki kombinasi dari kedua tipe ketahanan yaitu
vertikal dan horisontal sekaligus memiliki ketahanan blas yang lebih tinggi
dibandingkan varietas yang hanya memiliki salah satu dari tipe ketahanan
tersebut. Berbeda halnya dengan varietas yang memiliki ketahanan horisontal
yang bersifat tidak stabil dan bervariasi dalam reaksi ketahanannya terhadap
patogen. Varietas yang memiliki kombinasi kedua tipe ketahanan lebih fleksibel
pertahanannya dalam menghadapi mutasi yang terjadi pada patogen dan
perubahan populasi patogen (Wang et al. 1993).
Sebagai contoh adalah varietas padi gogo asal Kolombia, Oryzica Llanos-5.
Varietas ini merupakan varietas komersil popular yang memiliki ketahanan
terhadap blas yang tidak terpatahkan (durable) sejak dilepas tahun 1989 di Kolombia. Oryzica Llanos-5 telah dievaluasi ketahanannya terhadap ratusan isolat
blas asal Kolombia. Isolat-isolat blas tersebut telah dikelompokkan dalam 6
lineage berdasarkan analisis marka molekuler. Selain itu, Oryzica Llanos-5 juga telah diinokulasi dengan 202 isolat blas di IRRI. Isolat-isolat tersebut juga telah
dikelompokkan dalam 6 lineage berdasarkan analisis marka molekuler. Hasil kedua penelitian tersebut menunjukkan bahwa Oryzica Llanos-5 tahan terhadap
seluruh isolat blas (Correa-Victoria & Zeigler 1993a, Correa-Victoria & Zeigler
1995). Kemungkinan varietas ini memiliki beberapa gen ketahanan terhadap