DARI EKSTRAK TAPE
PERIS PARIAMAN SITANGGANG
DEPARTEMEN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
ISOLASI DAN KARAKTERISASI ENZIM PROTEOLITIK
DARI EKSTRAK TAPE
PERIS PARIAMAN SITANGGANG
Skripsi
sebagai salah satu syarat untuk memperoleh gelar
Sarjana Sains pada
Departemen Kimia
DEPARTEMEN KIMIA
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
PERIS PARIAMAN SITANGGANG. Isolasi dan Pencirian Enzim Proteolitik dari Ekstrak Tape. Dibimbing oleh IRMA H SUPARTO dan YANTI.
Tape ketan putih merupakan produk fermentasi beras ketan putih oleh bantuan ragi. Bakteri di dalamnya diduga mengandung enzim protease fibrinolitik yang dapat membantu proses penguraian darah beku. Penelitian ini bertujuan mengisolasi bakteri tersebut lalu mencirikan enzim yang dihasilkannya. Ada 9 isolat yang diujikan, dan diketahui isolat TP-4 diketahui memiliki aktivitas protease maksimum, yaitu 0.724 U/ml. Ektrak enzim protease yang dihasilkan dimurnikan melalui beberapa tahapan: (1) presipitasi amonium sulfat 65%, (2) dialisis (cut off 10 kD), dan (3) pemekatan dengan poli(etilena glikol) (PEG). Hasil aktivitas spesifik dialisat adalah 0.881 U/mg dengan tingkat kemurnian 0.701 dibandingkan dengan ekstrak enzim kasar (1.257 U/mg). Analisis SDS-PAGE 10% pada enzim kasar, presipitat, dan dialisat mempunyai 1 buah pita protein (11.60 kD). Uji zimografi menunjukkan bahwa enzim protease ini mempunyai aktivitas kaseinolitik, fibrinolitik, dan gelatinolitik. Ekstrak enzim kasar dihambat secara spesifik oleh etilenadiamina tetraasetat sehingga digolongkan ke dalam kelompok protease logam sedangkan dialisat dihambat secara spesifik oleh N-p-tosil-L-lisinklorometil keton, dan fenilmetilsulfonil fluorida sehingga digolongkan ke dalam protease serin.
ABSTRACT
PERIS PARIAMAN SITANGGANG. Isolation and Characterization of Proteolytic Enzyme from Fermented Glutinous White Rice Extract. Supervised by IRMA H SUPARTO and YANTI.
PRAKATA
Puji syukur penulis panjatkan dalam nama Yesus Kristus atas segala kasih-Nya sehingga penulis dapat menyelesaikan karya ilmiah ini. Penulis melakukan penelitian dari bulan Agustus sampai dengan Desember di Universitas Katolik Atma Jaya, Jakarta.
Penulis mengucapkan terima kasih kepada dr. Irma H Suparto, M.S dan Yanti, Msi selaku pembimbing yang telah memberikan saran, bimbingan, pengarahan serta dorongan moral. Demikian juga kepada Komar Sutriah, MSi selaku pembimbing akademik yang telah memberikan semangat, saran, dan nasehat yang sangat berharga selama ini. Begitu juga kepada Mas Hery yang telah banyak membantu penulis selama kuliah dalam hal administrasi.Penulis juga mengucapkan terima kasih kepada Kak Budi Arifin yang telah banyak membantu penulis dalam hal penulisan skripsi. Ungkapan terima kasih juga penulis sampaikan untuk keluarga tercinta Ayah, Ibu, dan kakak yang telah memberikan bantuan baik material maupun spiritual dalam membantu penulis menyelesaikan karya ilmiah ini. Penulis juga mengucapkan terima kasih kepada Mas Yudi, Mas Nurdin, Pak Bambang, dan Pak Ridwan yang telah banyak membantu penulis selama penelitian, demikian juga rekan-rekan mahasiswa Atma Jaya. Ucapan terima kasih juga ditujukan penulis kepada Andre,Dian, Selvi, Soli,Santi, dan Wiwit selaku rekan-rekan seperjuangan selama penelitian. Terima kasih penulis ucapkan kepada teman-teman angkatan 36, 37 dan 38 atas masukan dan kebersamaannya. Terima kasih juga penulis ucapkan kepada rekan-rekan kosan C-2 atas saran dan masukannya.
Semoga karya ilmiah ini dapat bermanfaat.
Bogor, Maret 2006
Penulis dilahirkan di Sait ni huta, Pulau Samosir Nauli pada tanggal 26 September 1982 dari Ayah P Sitanggang dan ibu S Naibaho merupakan putra bungsu (siampudan) dari delapan bersaudara. Tahun 2001 penulis lulus dari SMU Negeri 1 Pangururan dan pada tahun yang sama penulis masuk IPB melalui jalur Undangan Seleksi Masuk IPB. Penulis diterima di Program Studi Kimia, Departemen Kimia, Fakultas Matematika dan Ilmu pengetahuan Alam.
DAFTAR ISI
Halaman
DAFTAR TABEL ... ....viii
DAFTAR GAMBAR ...ix
DAFTAR LAMPIRAN ... x
PENDAHULUAN TINJAUAN PUSTAKA Tape ... 1
Pangan Fermentasi ... 1
Pencirian Enzim Protease ... 1
BAHAN DAN METODE Alat dan Bahan ... 3
Metode Penelitian... 3
HASIL DAN PEMBAHASAN Penapisan Isolat Sampel Tape Ketan Putih... 6
Pemurnian Enzim... 6
Penentuan Aktivitas Maksimum Enzim kasar ... 7
Pengaruh Suhu Terhadap Enzim ... 8
Pengaruh pH Terhadap Enzim ... 8
Pengaruh Inhibitor ... 9
Pengaruh Pelarut ... 10
Penentuan Bobot Molekul (SDS-PAGE) ... 10
Zimografi ... 11
SIMPULAN DAN SARAN Simpulan ... 12
Saran ... 12
DAFTAR PUSTAKA... 12
Halaman
1 Komposisi gel pemisah dan gel penahan untuk SDS-PAGE dan zimografi ... 5
2 Ringkasan tahap pemurnian enzim dari isolat ekstrak tape ketan putih ... 7
3 Ciri biokimia protease dari isolat bakteri TP-4 ... 11
DAFTAR GAMBAR
Halaman 1 Isolat TP-4 ... 62 Pengaruh konsentrasi amonium sulfat terhadap kadar protein ... 7
3 Kurva penentuan aktivitas maksimum enzim kasar ... 8
4 Kurva pengaruh suhu terhadap aktivitas enzim dan aktivitas sisa enzim ekstrak kasar dan dialisat... 8
5 Kurva pengaruh pH terhadap aktivitas enzim dan aktivitas sisa enzim ekstrak kasar dan dialisat... 8
6 Diagram pengaruh inhibitor terhadap aktivitas sisa enzim ekstrak kasar dan dialisat ... 9
7 Diagram pengaruh pelarut organik terhadap aktivitas sisa enzim eksrak kasar dialisat ... 10
8 Analisis SDS-PAGE 10% pada C, enzim kasar, P, endapan, dan D, dialisat... 10
9 Zimogram kasein 12% pada enzim kasar (C), endapan (P) dan dialisat (D)... 11
10 Zimogram fibrinogen 12% pada enzim kasar (C), endapan (P) dan dialisat (D) ... 11
DAFTAR LAMPIRAN
Halaman
1 Diagram alir penelitian ... 15
2 Prosedur pembuatan pereaksi kimia ... 16
3 Data seleksi 9 isolat bakteri ... 19
4 Contoh perhitungan aktivitas enzim (U/ml) dan aktivitas enzim spesifik... 20
5 Pembuatan kurva standar Bradford ... 21
6 Data pengaruh suhu, pH, waktu, inhibitor, dan pelarut organik terhadap aktivitas protease enzim kasar tape ketan putih ... 22
7 Data pengaruh suhu, pH, waktu, inhibitor, dan pelarut organik terhadap aktivitas protease dari dialisat tape ketan putih ... 23
PENDAHULUAN
Fermentasi merupakan proses berban-tuan mikro-organisme yang salah satunya di-manfaatkan dalam pembuatan dan peningkatan cita rasa makanan. Keju, tempe, tape, minuman anggur, dan tuak merupakan beberapa contoh bahan pangan pangan produk fermentasi. Salah satunya yang diteliti disini ialah tape ketan putih. Tape tersebut merupakan produk fermentasi dari beras ketan putih dengan bantuan ragi. Bakteri yang dihasilkan oleh tape berperan dalam membentukan cita rasa dan aroma yang khas dari tape. Sehubungan dengan hal ini, diduga tape mengandung komponen enzim protease tertentu yang bisa membantu proses penguraian darah beku, yaitu protease fibrinolitik.
Enzim fibrinolitik yang dihasilkan di-harapkan dapat mengatasi masalah trombosis, yaitu suatu keadaan yang menimbulkan penggumpalan darah. Trombosis dapat terjadi pada pembuluh koroner yang disebabkan oleh pecahnya plak aterosklerosis yang labil.
Bakteri diisolasi dari tape ketan putih lalu diseleksi isolat yang dianggap berpotensi baik, untuk dicirikan pada tahap selanjutnya. Isolat yang tertinggi aktivitas proteasenya dipakai untuk produksi enzim protease kasar, yang kemudian dimurnikan dan dicirikan pada aktivitas protease tertinggi. Tahapan pe-murnian yang dilakukan meliputi presipitasi, dialisis, dan pemekatan. Penentuan bobot molekul dan aktivitas enzim spesifik enzim protease dilakukan dengan elektroforesis dan zimografi.
Penelitian ini bertujuan mendapatkan isolat dan mengetahui ciri enzim protease dari isolat ekstrak tape terbaik. Data pencirian enzim tersebut dapat dijadikan informasi bahwa produk pangan fermentasi tradisional Indonesia (khususnya tape) dapat bermanfaat sebagai “nutraceutical” yang kaya enzim untuk peningkatan kesejahteraan masyarakat, baik dalam bidang pertanian, pangan maupun kesehatan. Diagram alir penelitian secara lengkap terdapat pada Lampiran 1.
TINJAUAN PUSTAKA
Tape
Tape merupakan salah satu produk pangan tradisional Indonesia yang dihasilkan melalui proses fermentasi dengan bantuan ragi. Ragi yang berperan dalam pembuatan
tape merupakan populasi campuran spesies genus Aspergillus, Saccharomyces, Candida, Hansenula, dan Acetobacter. Genus−genus tersebut hidup bersama secara sinergetik. Aspergillus dapat menyederhanakan amilum, sedangkan Saccharomyces, Candida dan Hansenula dapat mengubah gula menjadi alkohol dan bermacam-macam zat organik lainnya. Acetobacter berperan mengubah alkohol menjadi asam cuka (Dwidjoseputro 1977).
Pangan Fermentasi
Produk fermentasi meliputi (1) minuman beralkohol seperti bir, sake, brem, dan anggur; (2) produk fermentasi berbahan susu seperti keju, dan yoghurt; (3) produk fermentasi sayuran seperti acar kubis dan acar timun; (4) produk serelia seperti roti, lalu (5) produk kacang kedelai seperti tempe, oncom, tauco, dan kecap; sementara (6) produk beras ketan meliputi tape ketan putih dan tape ketan hitam selain ada juga tape berbahan singkong yang disebut peuyeum.
Mikrob yang bersifat fermentatif dapat mengubah karbohidrat dan turunannya menjadi alkohol, asam, dan karbon dioksida. Mikrob proteolitik dapat memecah protein dan komponen nitrogen lainnya sehingga menghasilkan bau busuk yang tidak diingin-kan. Sementara itu, mikrob lipolitik akan menghidrolisis lemak, fosfolipid, dan turunannya dengan menghasilkan bau tengik. Bila alkohol dan asam yang dihasilkan mikrob cukup tinggi, maka pertumbuhan mikrob proteolitik dan lipolitik dapat dihambat. Jadi pada prinsipnya, fermentasi adalah me-ningkatkan pertumbuhan mikrob pem-bentuk alkohol dan asam, dan menekan per-tumbuhan mikrob proteolitik dan lipolitik (Afrianti 2005).
Fermentasi juga menurunkan kadar senyawa beracun seperti anti-tirosin pada kedelai sehingga kadarnya menurun ketika dijadikan tempe. Fermentasi turut mem-pertinggi nilai gizi, karena mikrob memecah senyawa kompleks menjadi senyawa sederhana. Misalnya, Rhizopus oligosporus dapat meningkatkan vitamin B12 pada tempe. Begitu pula dengan kandungan niasin dan riboflavin (Afrianti 2005).
Pencirian Enzim Protease
2
lain peptidase, yaitu enzim yang menguraikan peptida menjadi asam-asam amino. Contoh lainnya adalah gelatinase, yaitu enzim yang menguraikan gelatin dan renin, yaitu enzim yang menguraikan kasein dari susu.
Ada enzim yang dikeluarkan oleh sel bakteri guna mengambil zat makanan di sekelilingnya; enzim semacam ini disebut eksoenzim atau enzim luar-sel. Sebaliknya, di dalam sel sendiri terdapat juga banyak enzim yang memegang peranan dalam proses pencernaan dan pembongkaran zat makanan, seperti pada peristiwa respirasi atau fermentasi. Enzim-enzim ini telah ada di dalam sel enzim dan disebut endoenzim atau enzim dalam-sel.(Dwidjoseputro 1977).
Enzim yang dihasilkan oleh bakteri juga dapat dibedakan menjadi enzim konstitutif dan induktif. Enzim konstitutif jumlahnya tetap dan tidak bergantung pada keadaan metabolisme organisme tersebut, misalnya enzim yang terlibat dalam lintasan glikolisis. Di sisi lain, enzim induktif terdapat dalam berbagai konsentrasi. Dalam keadaan normal jumlahnya sedikit, tetapi konsentrasinya akan meningkat dengan cepat bila substratnya terdapat dalam medium, terutama jika substrat tersebut merupakan sumber karbon satu-satunya bagi sel Beberapa enzim mempunyai spesifisitas yang tinggi terhadap substrat tertentu dan tidak akan bekerja bahkan terhadap senyawa yang mirip secara struktural (Lehninger 1993). Aktivitas enzim terhadap substrat dipengaruhi oleh beberapa faktor, yaitu pH, suhu, konsentrasi substrat, dan aktivator (koenzim dan kofaktor) dan inhibitor.
Nilai pH
Efek pH pada enzim berkaitan dengan keadaan ionisasi dari sistem yang dikatalisis, termasuk substrat dan enzim itu sendiri. Perubahan pH dapat memengaruhi keadaan ionisasi dari asam-asam amino pada sisi aktif enzim sehingga memengaruhi interaksinya dengan molekul substrat. Nilai pH yang terlalu tinggi atau terlalu rendah akan menyebabkan ketidakstabilan konformasi enzim sehingga struktur enzim rusak. Enzim mempunyai pH optimum yang khas yang akan menyebabkan aktivitas maksimal. Keadaan optimum ini dihubungkan dengan saat gugus pemberi proton atau penerima proton yang aktif pada sisi aktif enzim berada pada kondisi ionisasi yang tepat. Keadaan optimum tidak harus sama dengan pH lingkungannya. Aktivitas enzim dalam sel
sebagian diatur oleh pH media kulturnya (Lehninger 1993).
Suhu
Suhu mempunyai dua pengaruh yang saling bertentangan. Suhu dapat meningkatkan aktivitas enzim, tetapi dapat pula merusak struktur enzim. Suhu optimum merupakan batas di antara keduanya. Peningkatan suhu sebelum tercapainya suhu optimum akan meningkatkan laju reaksi katalitik enzim karena meningkatnya energi kinetik molekul-molekul yang bereaksi. Sebaliknya, jika suhu dinaikkan sesudah suhu optimum kompleks enzim-substrat yang melampaui energi aktivasi terlalu besar, sehingga memecah ikatan sekunder pada konformasi enzim dan sisi aktifnya. Hal ini mengakibatkan enzim terdenaturasi dan kehilangan sifat katalitiknya
Konsentrasi Substrat
Konsentrasi substrat sangat meme-ngaruhi laju reaksi enzimatik. Laju maksimum (vmaks) suatu enzim tercapai ketika
enzim telah jenuh oleh substratnya. Konsentrasi substrat tertentu pada saat enzim mencapai setengah laju maksimum disebut Km atau tetapan Michaelis-Menten. Nilai Km
bersifat khas pada setiap enzim tertentu. Hubungan antara Km dan vmaks dinyatakan
dalam persamaan Michaelis−Menten (Lehninger 1993).
[S] [S]
m maks
0 = +
K v v
Persamaan Michaelis–Menten merupa kan dasar bagi segi kinetika kerja enzim. Apabila nilai Km dan vmaks diketahui, maka
laju reaksi suatu enzim pada setiap konsentrasi substrat dapat dihitung. Nilai Km
dan vmaks yang lebih tepat dapat diperoleh
dengan memetakan data yang ada ke dalam persamaan Linewaver-Burk: maks maks m
v
S
v
K
v
1
]
[
1
1
0+
=
Aktivator (Kofaktor dan Koenzim) dan Inhibitor
Pada beberapa enzim, kofaktor dan koenzim terlibat langsung pada proses katalitik, tetapi ada juga yang berfungsi sebagai pembawa gugus fungsional tertentu. Hampir semua enzim dapat dihambat oleh senyawa kimia tertentu misalnya ion logam, senyawa peng-kelat, senyawa organik, atau bahkan substrat enzim itu sendiri.
Elektroforesis dan Zimogram
Elektroforesis ialah pergerakan molekul bermuatan di dalam medan listrik. Per-pindahan partikel bermuatan ditentukan oleh gugus-gugus yang terionisasi. Jenis dan besar muatan gugus-gugus yang terionisasi bergantung pada permukaan partikel, kekuatan ionik, dan pH medium. Ukuran dan bentuk partikel juga akan memengaruhi mobilitas.
Bahan hayati banyak yang bermuatan listrik, yang besarnya bergantung pada jenis molekul, pH, dan komponen medium pelarutnya. Molekul-molekul ini dalam larutan akan bergerak ke arah elektrode yang polaritasnya berlawanan dengan muatan itu. Prinsip inilah yang digunakan dalam elektroforesis untuk memisahkan molekul-molekul dengan muatan berbeda.
Macam-Macam Elektroforesis
Ada beberapa macam teknik di dalam elektroforesis antara lain elektroforesis per-gerakan batas (moving boundary elek-troforesis) dan elektroforesis zone. Metode yang digunakan pada penelitian ini ialah elektroforesis zone.
Metode elektroforesis pergerakan batas pertama kali dikemukakan oleh ahli biokimia Swedia, Tiselius, pada tahun 1973. Tiselius menggunakan tabung berbentuk huruf U yang ke dalamnya dimasukkan larutan protein yang akan dianalisis. Larutan bufer dimasukkan ke dalam tabung sedemikian rupa sehingga diperoleh batas yang tajam antara kedua larutan tersebut. Elektrode dipasang pada kedua ujung kaki tabung, kemudian arus listrik dialirkan sehingga terjadi pergerakan molekul protein pada kedua batas larutan ke arah elektrode. Pergerakan molekul dapat diikuti dengan mengukur indeks bias larutan. Apabila larutan protein mengandung asam amino dari berbagai titik isoelektrik, maka protein ini akan bergerak dengan laju yang berbeda-beda sehingga dapat dipisahkan satu dengan yang lain. Metode Tiselius ini tidak dapat memisahkan protein dengan sempurna
karena gaya gravitasi memengaruhi batas tajam antara kedua larutan. Demikian pula, terjadinya konveksi mengakibatkan kaburnya batas larutan antara kedua daerah pemisahan.
Metode elektroforesis zone menggunakan penyangga seperti kertas, selulosa asetat, pati, atau poliakrilamida untuk mengatasi kelemahan elektroforesis pergerakan batas. Berdasarkan penyangga yang digunakan tersebut, dikenal beberapa metode elektroforesis zone yaitu elektroforesis kertas, selulosa asetat, pati, gel agar, dan gel poliakrilamida. Elektroforesis zone digunakan untuk pemisahan sempurna suatu campuran protein ataupun senyawa bermuatan lainnya, sehingga sesuai untuk maksud analitik dan preparatif. Semakin tipis zone yang terbentuk maka akan semakin sensitif metodenya atau proses pemisahan semakin sempurna.
BAHAN DAN METODE
Alat dan Bahan
4
Metode Penelitian
Isolasi mikrob
Tape ketan diekstraksi (diblender) secukupnya dengan bufer universal 50 mM. Sebanyak 100 µl ekstrak tape ketan diencerkan dengan 900 µl larutan garam fisiologis (NaCl 0,85%) sebanyak 5 kali dari keadaan awal. Kultur disebar pada media agar yang telah dituang pada cawan petri, kemudian diinkubasi pada suhu 37 oC selama 2 hari dan diamati mikrob yang memiliki zone bening. Koloni mikrob yang diperoleh digores lagi pada media SMA yang baru sampai diperoleh koloni bakteri tunggal. Isolat yang terpilih dimasukkan ke dalam gliserol 15% sebagai stok bakteri selanjutnya.
Produksi protease
Sebanyak 2 ose bakteri ditumbuhkan ke dalam 200 ml campuran 130 ml media LB steril dengan 70 ml susu skim steril, kemudian diinkubasi pada inkubator bergoyang berlaju 150 rpm pada suhu 37 oC selama 2 hari. Kemudian disentrifugasi berlaju 7500 rpm selama 15 menit dengan suhu 4 oC untuk memisahkan sel bakteri dengan enzim kasar. Pengujian aktivitas dilakukan terhadap supernatan yang mengandung enzim kasar.
Analisis aktivitas protease
Aktivitas protease diukur secara kuantitatif dengan modifikasi metode Bergmeyer (1983) menggunakan substrat kasein Hammarsten 2% (b/v). Terdapat tiga perlakuan analisis yang dilakukan, yaitu blangko, standar, dan sampel. Larutan enzim 50 µl yang sudah dipanaskan pada suhu dan waktu inkubasi tertentu (yang menghasilkan aktivitas maksimum) ditambahkan ke dalam tabung Eppendorf yang berisi 250 µl kasein 2%,dan 250 µl bufer fosfat 50 mM dengan pH 7. Pada blangko dan standar, enzim digantikan dengan akuades dan tirosin 50 µl.
Larutan diinkubasi pada suhu 37 oC selama 10 menit (suhu dan inkubasi optimum enzim). Reaksi hidrolisis dihentikan dengan cara penambahan 500 µl TCA 0,1 M. Tabung blangko dan standar ditambahkan 50µl akuades, kemudian larutan diinkubasi kembali pada suhu 37 oC selama 10 menit, kemudian disentrifugasi pada laju 6000 rpm pada suhu 4 oC selama 10 menit.
Supernatan sebanyak 375 µl ditambahkan ke dalam tabung yang berisi 1,25 ml Na2CO3
0,4 M dan 250 µl pereaksi Folin−Ciocalteu, lalu diinkubasi kembali pada suhu 37 oC selama 20 menit. Absorbans larutan diukur pada panjang gelombang 578 nm. Aktivitas enzim protease dihitung berdasarkan persamaan berikut: inkubasi waktu n pengencera faktor blangko standar blangko sampel _ _ ml) / ( = × A A A A U
Analisis kadar protein
Kadar protein ditentukan dengan metode Bradford (1976). Sebanyak 100 µL larutan enzim ditambahkan dengan 1 mL akuades dan 1 mL pereaksi Bradford dalam tabung reaksi. Pada perlakuan blangko, larutan enzim diganti dengan akuades. Kemudian divorteks dan didiamkan selama 20 menit pada suhu ruang (tabung reaksi ditutup dengan foil aluminium). Pengukuran absorbans dilakukan pada panjang gelombang 595 nm. Konsentrasi protein ditentukan dengan persamaan kurva standar (standar protein BSA Fraction dengan kisaran 0-250 µL)
Presipitasi
Enzim dalam filtrat diendapkan dengan amonium sulfat. Pengujian aktivitas enzim hasil pengendapan oleh garam dengann konsentrasi 30−85%(b/v), untuk menentukan konsentrasi garam yang optimal. Penambahan dilakukan sedikit demi sedikit pada suhu 0−4
o
C sambil diaduk hingga larut, lalu dibiarkan 2 jam pada suhu rendah. Endapan enzim yang dipisahkan dengan sentrifugasi dingin pada 7500 rpm selama 15 menit dilarutkan kembali dalam bufer universal 50 mM pH 7.
Dialisis dan pemekatan
Pencirian protease ekstrak kasar
Uji kuantitatif untuk mencirikan enzim protease dilakukan dengan metode spektrofotometri (Bergmeyer 1983), yang meliputi pH dan suhu optimum, pengaruh alkohol (pelarut organik), inhibitor, dan spesifisitas substrat. Sementara uji kualitatif dilakukan dengan SDS-PAGE dan zimografi.
Penentuaan pH dan suhu optimum
Optimalisasi pH dilakukan dengan menggunakan bufer universal dan substrat kasein yang diatur pada pH 4−10. Pengujian aktivitas dilakukan pada suhu optimum (50
o
C). Optimum suhu dilakukan dengan dengan uji aktivitas enzim pada berbagai variasi suhu yaitu; 27, 37, 50, 60, dan 70 oC. Aktivitas tertinggi menunjukkan suhu optimum enzim.
Pengaruh spesifisitas substrat
Uji aktivitas protease pada suhu dan pH optimum dilakukan pada berbagai substrat protein dengan konsentrasi 2% (b/v). Substrat yang digunakan adalah kasein, gelatin, fibrin, fibrinogen dan albumin. Substrat dilarutkan dalam bufer universal 50 mM.
Pengaruh pelarut
Uji aktivitas protease menggunakan variasi jenis pelarut organik, yaitu: kloroform, n-heksana, benzena, dan toluena. Pengujian pengaruh pelarut dilakukan dengan cara mencampur enzim dengan pelarut organik [2% (v/v)], nisbah 1:1 setelah itu, ditempatkan pada suhu 4 oC selama 1 jam. Kemudian larutan divorteks, dan diuji sesuai dengan metode Bergmeyer.
Pengaruh inhibitor
Pengaruh penambahan senyawa inhibitor terhadap aktivitas protease ditentukan dengan cara inkubasi 100 µL enzim dan 100 µL larutan senyawa inhibitor yang diujikan meliputi EDTA 0.01 mM, PMSF 0.01mM, TLCK 0.1 mM dan STI 0.01 mg/ml selama satu jam pada suhu ruang, lalu aktivitas residunya dianalisis secara kuantitatif.
Analisis SDS-PAGE dan zimografi
Kombinasi antara elektroforesis gel poli-akrilamida dengan detergen SDS digunakan untuk memisahkan dan meneliti jumlah dan
ukuran bobot molekul rantai protein dan rantai subunit protein. Sementara itu, elektroforesis dengan teknik zimografi juga merupakan bertujuan mendeteksi aktivitas enzim proteolitik secara langsung.
Tahapan kerja pada analisis SDS-PAGE dan zimografi meliputi: penyiapan gel pemisah dan penahan, penyiapan sampel dan pemuatan, kondisi running, pewarnaan gel, dan pelunturan warna.
Penyiapan gel pemisah dan penahan
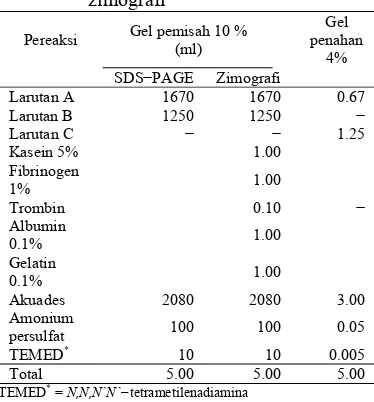
Pembuatan gel pemisah 12% dan gel penahan 4% untukSDS−PAGE dan zimografi dilakukan dengan komposisi yang tertera pada Tabel 1.
Tabel 1 Komposisi gel pemisah dan gel penahan untuk SDS−PAGE dan zimografi
Pereaksi Gel pemisah 10 % (ml)
Gel penahan
4%
SDS−PAGE Zimografi
Larutan A 1670 1670 0.67
Larutan B 1250 1250 −
Larutan C − − 1.25
Kasein 5% 1.00
Fibrinogen
1% 1.00
Trombin 0.10 −
Albumin
0.1% 1.00
Gelatin
0.1% 1.00
Akuades 2080 2080 3.00
Amonium
persulfat 100 100 0.05
TEMED* 10 10 0.005
Total 5.00 5.00 5.00
TEMED*
= N,N,N`N`−tetrametilenadiamina
Penyiapan sampel dan pemuatan
Khusus SDS−PAGE, 20 µL sampel ditambahkan dengan 5 µL bufer sampel yang mengandung 2-merkaptoetanol, lalu dipanas-kan pada suhu 100 oC selama 3−5 menit. Sementara pada zimografi, sampel dilarutkan ke dalam bufer yang tidak mengandung 2-mer-kaptoetanol dan tidak memerlukan pema-nasan. Tiap sampel dimuat ke dalam sumur gel dengan kisaran volume 10−20 µL, sedang-kan volume LMW yang digunasedang-kan sebanyak 5 µL, dengan menggunakan plasmin sebagai standar.
Kondisi running, pewarnaan, dan pelunturan warna
6
Pada SDS-PAGE, setelah elektroforesis langsung diwarnai dengan menggunakan larutan pewarna commassie brilliant blue R−250 selama 15 menit. Pelunturan warna pada gel dilakukan dengan larutan peluntur berulang kali sampai diperoleh pita protein berwarna biru dengan latar gel bening. Sementara pada zimografi, setelah elektroforesis, gel didenaturasi terlebih dahulu dalam larutan Triton X-100 2.5% (v/v) sambil digoyang selama 1 jam. Kemudian gel ditempatkan dalam bufer fosfat 50 mM pH optimum selama 30 menit. Gel diwarnai dengan larutan pewarna commassie brilliant blue R-250) selama 15 menit. Pelunturan warna gel dilakukan dengan peluntur berulang kali sampai diperoleh pita enzim proteolitik putih dengan latar gel biru.
HASIL DAN PEMBAHASAN
Penapisan Isolat Sampel Tape Ketan Putih
Ekstrak tape diencerkan lebih dahulu dengan garam fisiologis sebelum disebar dalam media agar. Pengenceran yang dilakukan berkisar dari 10 sampai 105 kali dengan interval 10. Hasil pengamatan menunjukkan bahwa semakin besar faktor pengencerannya, jumlah isolat bakteri yang memiliki koloni tunggal semakin banyak. Hal ini disebabkan isolat memiliki ruang gerak yang lebih luas pada pengenceran yang lebih tinggi.
Terdapat 9 isolat tape (Lampiran 3) yang diproduksi pada media LB yang aktivitasnya diuji untuk mencari aktivitas enzim maksimum. Jenis isolat dengan aktivitas maksimum adalah TP-4 sebesar 0.724 U/ml yang digunakan untuk pencirian selanjutnya. Contoh perhitungan aktivitas enzim dapat dilihat pada Lampiran 4. Sementara itu, aktivitas spesifik dan kadar protein ekstrak enzim kasar sebesar 1.257 U/mg dan 0.576 mg/ml. Nilai kadar protein tersebut diperoleh melalui persamaan kurva standar BSA (bovine serum albumin). Kurva standar BSA terdapat pada Lampiran 5. Isolat TP-4 dapat dilihat pada Gambar 1.
Pemurnian Enzim
Prinsip presipitasi adalah penggumpalan protein non-enzim dengan penambahan
garam. Ion garam yang ditambahkan memengaruhi kelarutan protein. Pada konsentrasi rendah, ion-ion ini akan mengelilingi molekul protein dan mencegah mereka bersatu sehingga protein melarut. Peristiwa ini disebut salting in. Pada konsentrasi tinggi, terjadi peningkatan muatan
Gambar 1 Isolat TP−4
listrik di sekitar protein, yang akan menarik mantel air dari koloid protein. Peristiwa hidrofobik antarmolekulprotein pada suasana ionik tinggi akan menurunkan kelarutan protein. Peristiwa tersebut dikenal dengan salting out. Molekul air yang berikatan dengan ion-ion garam yang semakin banyak menyebabkan penarikan selubung air yang mengelilingi permukaan protein. Peristiwa ini mengakibatkan protein saling berinteraksi, beragregasi, dan kemudian mengendap (Harris 1989, Scopes 1989). Amonium sulfat merupakan garam yang paling sering digunakan untuk mengendapkan protein karena memiliki daya larut tinggi di dalam air, dan relatif tidak mahal (Scopes 1989).
Tabel 2 Ringkasan tahap pemurnian enzim dari isolat ekstrak tape ketan putih
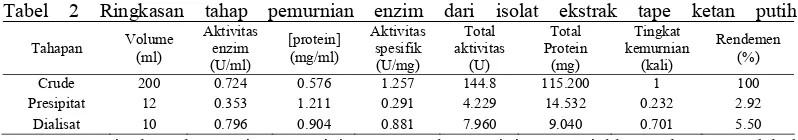
Tahapan Volume (ml) Aktivitas enzim (U/ml) [protein] (mg/ml) Aktivitas spesifik (U/mg) Total aktivitas (U) Total Protein (mg) Tingkat kemurnian (kali) Rendemen (%)
Crude 200 0.724 0.576 1.257 144.8 115.200 1 100
Presipitat 12 0.353 1.211 0.291 4.229 14.532 0.232 2.92
Dialisat 10 0.796 0.904 0.881 7.960 9.040 0.701 5.50
Penurunan tingkat kemurnian presipitat disebabkan oleh kadar protein yang semakin meningkat sementara aktivitas enzim menurun. Hasil pengamatan menunjukkan bahwa penurunan aktivitas enzim disebabkan oleh penambahan amonium sulfat yang menyebabkan kelarutan protein menurun. Selain itu menyebabkan protein saling berinteraksi, beragregasi kemudian mengendap.
Gambar 2 menunjukkan hubungan antara kejenuhan amonum sulfat dan kadar protein supernatan. Kadar protein supernatan terkecil mengindikasikan kadar protein yang terendapkan maksimal. Nilai tersebut dapat dilihat pada titik terndah yang ditunjukkan oleh kurva, yaitu protein untuk mendapatkan titik terendah tersebut.pada konsentrasi 65%. Uji pengendapan dilakukan dengan volume total (enzim dengan amonium sulfat) kecil yakni sebesar 10 ml. Setelah didapatkan kadar protein terkecil supernatan (65%) kemudian dilakukan presipitasi dalam jumlah besar yakni sebesar 200 ml. Setelah presipitasi dilanjutkan dengan dialisis.
Ukuran ini menunjukkan ukuran molekul minimum yang dapat tertahan di dalam membran. Dialisat yang diperoleh memiliki aktivitas enzim sebesar 0.796 U/ml. Nilai ini lebih besar daripada aktivitas enzim kasar dan presipitat. Sebaliknya kadar protein dialisat lebih kecil dibandingkan dengan presipitat tetapi lebih besar dari aktivitas enzim kasar, yakni sebesar 0.904 mg/ml. Di sisi lain aktivitas spesifik meningkat menjadi 0.881 U/mg. Disamping itu tingkat kemurnian dialisat juga meningkat terhadap presipitat, tetapi masih lebih kecil dari enzim kasar.
0.00 0.10 0.20 0.30 0.40 0.50 0.60 0.70
0 20 40 60 80 100
kejenuhan amonium sulfat (%)
K a da r pr o te in ( m g /m l)
Gambar 2 Pengaruh konsentrasi amonium sulfat terhadap kadar protein supernatan.
Berdasarkan pengamatan dapat disimpul-kan bahwa kenaidisimpul-kan aktivitas spesifik dialisat tersebut disebabkan oleh kenaikan aktivitas enzim.hal ini karena kelarutan enzim me-ningkat dengan bantuan bufer yang berdifusi melalui membran sehingga enzim lebih leluasa untuk melakukan aktivitasnya.aktivitas yang dimaksud adalah kemampuan enzim untuk menghidrolisis sumber karbon (substrat) Sebaliknya kadar protein yang berkurang, disebabkan adanya sebagian protein yang keluar dari kantung dialisis. Protein yang dapat bertahan di dalam kantung adalah ukurannya yang lebih besar dari ukuran pori kantung dialisis
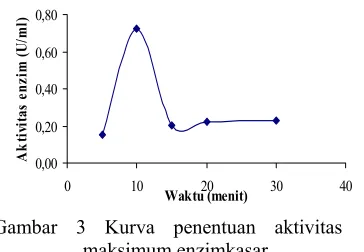
Penentuan Aktivitas Maksimum Enzim Kasar
Dari kesembilan isolat diproduksi sebagai enzim protease kasar dan ditentukan pada variasi suhu, pH, dan waktu optimumnya.. Pada uji aktivitas enzim, setiap sampel, blangko, dan standar ditambahkan buferunuk menjaga stabilitas pH-nya. Selanjutnya setelah diinkubasi, diberi asam trikloro asetat (TCA). Pemberian TCA bertujuan mengendapkan protein (kasein) dan peptida yang belum dipecah oleh enzim sekaligus untuk menginaktifkan enzim. Setelah disentrifugasi, supernatan diambil kemudian ditambah dengan Na2CO3. Penambahan Na2CO3
bertujuan memberikan suasana basa pada supernatan. Suasana basa diperlukan agar pewarna Folin Ciocalteu dapat bereaksi dengan asam amino dan mengubah warna supernatan, sebab pereaksi Folin hanya dapat bekerja pada suasana basa. Pereaksi Folin digunakan karena pereaksi ini lebih mudah Dialisis bertujuan lebih memurnikan
8
didapat, dan warna biru yang dihasilkannya lebih bagus dan bersih. Semakin pekat warna biru yang terbentuk meng-indikasikan bahwa larutan tersebut mengandung banyak asam amino.
Perlakuan sentrifugasi dilakukan pada laju 6000 rpm dengan suhu 4 oC selama 10 menit. Sentifugasi pada suhu rendah bertujuan mencegah kerusakan enzim akibat panas yang ditimbulkan. Dengan ini, sel yang berbobot molekul lebih besar dari enzim akan mengendap karena gaya gravitasi sedangkan enzim tetap berada pada cairan atau supernatannya.
0,00 0,20 0,40 0,60 0,80
0 10 20 30 4
Waktu (menit) A k ti vi ta s e n z im (U /m l) 0
Gambar 3 Kurva penentuan aktivitas maksimum enzimkasar.
Pengaruh Suhu Terhadap Enzim
Suhu merupakan salah satu faktor yang memengaruhi kinerja enzim. Penentuan suhu optimum dilakukan dengan mereaksikan enzim pada pH 7. berbagai suhu: 27, 37, 50, 60, dan 70oC. Gambar 4 menunjukkan bahwa suhu optimum enzim dari ekstrak kasar dan dialisat dicapai pada suhu 37 oC, dengan nilai aktivitas berturut-turut sebesar 0.724 U/ml dan 0.729 U/ml. Tabel lengkap pengaruh suhu, pH, waktu, pelarut, dan inhibitor terhadap enzim kasar dapat terdapat pada Lampiran 6).
Pada suhu di bawah 37 oC, aktivitas enzim meningkat karena terjadi peningkatan energi kinetik yang mempercepat gerak vibrasi, translasi, serta rotasi enzim dan substrat sehingga memperbesar peluang keduanya untuk saling bertumbukan (Suhartono 1989). Pada suhu di atas suhu optimum, aktivitas enzim menurun karena enzim adalah molekul protein yang dapat terdenaturasi pada suhu tinggi. Menurut Palmer (1991), peningkatan suhu hingga batas tertentu
dapat menyebabkan semakin meningkatnya aktivitas katalitik enzim dan semakin bertambahnya kerusakan enzim.
Gambar 4 juga menunjukkan bahwa pada enzim kasar aktivitas residu hilang sebesar 90% pada suhu 50 dan 70 oC. Sementara pada dialisat
hilang sebesar 98% pada suhu 50 oC dan 82% pada suhu 70oC. Berdasarkan rata-rata aktivitas residu, daya tahan enzim kasar cenderung lebih besar dibandingkan dengan dialisat enzim. Hal ini disebabkan aktivitas spesifik dari enzim kasar lebih besar dari dialisatnyameskipun kadar protein dari dialisat enzim lebih besar.
0,00 0,20 0,40 0,60 0,80 1,00 1,20
27 37 50 60 70
Suhu (oC)
A k ti v ita s e n z im (U /m l) 0 20 40 60 80 100 120 A k ti vi at as r e si d u a l
Aktivitas enzim dialisat (U/ml)
Aktivitas enzim crude (U/ml)
Aktivitas residual crude (%)
Aktivitas residual dialisat (%)
Gambar 4 Kurva pengaruh suhu terhadap aktivitas enzim dan aktivitas residual enzim ekstrak kasar dan dialisat.
Enzim yang diisolasi dari Bacillus sp. galur CK 11-4 (Kim et al. 1996) dilaporkan memiliki kisaran suhu optimal sebesar 70oC dan pH 10,5. Enzim fibrinolitik hasil fermentasi Bacillus firmus NA-1 stabil pada suhu 40 oC, tetapi aktivitas residunya hilang pada suhu 50 oC (Seo J dan Lee 2004).
Pengaruh pH Terhadap Enzim
Reaksi enzim dipengaruhi oleh pH. Peningkatan pH sebelum titik optimum menyebabkan terusnya meningkatnya aktivitas enzim,sampai seluruh tapak enzim berikatan dengan substrat membentuk kompleks enzim-substrat.
Pada Gambar 5 diperlihatkan bahwa pada kondisi mendekati pH 7 (pH 4,5, dan 6), aktivitas enzim kasar dan dialisat cenderung meningkat dan mencapai optimum pada pH 7 dengan nilai aktivitas sebesar 0.724 U/ml dan 0.729 U/ml. Pada kondisi pH tersebut tapak aktif enzim sudah seluruhnya berikatan dengan substrat membentuk kompleks enzim-substrat.
0,00 0,20 0,40 0,60 0,80 1,00 1,20
4 5 6 7 8 9 10 12
pH A k ti v ita s e n z im ( U /m l) 0 20 40 60 80 100 120 A k ti v ita s r e si d u a l (% )
Aktiviatas enzim dialisat (U/ml) Aktivitas enzim crude (U/ml) Aktivitas residual crude (%) Aktiviatas residual dialisat (%)
Gambar 5 Kurva pengaruh pH terhadap aktivitas enzim dan aktivitas residual enzim ekstrak kasar dan dialisat.
Aktivitas residu rata-rata pada dialisat cenderung lebih besar dibandingkan dengan aktivitas enzim kasar. Data tersebut me-nunjukkan bahwa masih banyak enzim pada ekstrak kasar yang tidak mampu mendegradasi substrat sehingga kompleks yang terbentuk antara enzim dengan substrat tidak maksimal. Sementara itu, pada dialisat, enzim sudah dimurnikan sehingga pembentukan kompleks antara enzim dan substrat sudah maksimal, sehingga daya tahan terhadap pH di bawah maupun di atas pH optimum cenderung lebih besar. Tabel lengkap pengaruh suhu, pH, waktu, pelarut, dan inhibitor terhadap dialisat enzim terdapat pada Lampiran 7).
Hasil pemurnian dan pencirian enzim protease dari Alkaligenes faecalis (Thangam & Rajkumar 2002) memiliki aktivitas optimal pada pH 9. Enzim protease yang diisolasi dari Bacillus subtilis galur 38 memiliki pH optimum pada 6.5 (Chantawannakul et al. 2002).
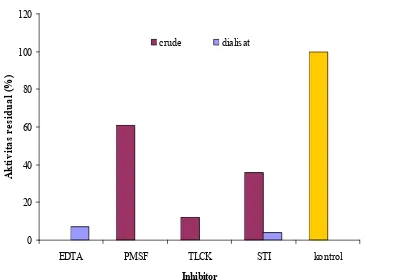
Pengaruh Inhibitor
Ikatan inhibitor (atau aktivator) dengan enzim dapat mengubah kemampuan enzim dalam mengikat substrat sehingga mengubah daya katalisisnya. Hal ini disebabkan karena struktur enzim mengalami perubahan fisik dan kimia sedemikian rupa sehingga aktivitas hayatinya pun berbeda. (Suhartono 1989).
Jenis atau nama protease ditentukan dari jenis inhibitor yang menghambat enzim tersebut. PMSF dan TLCK termasuk jenis protease serin yang dicirikan kehadiran gugus serin pada sisi aktifnya. Protein serin aktifnya pada kondisi pH netral dan basa (pH 7−11) (Rao et al.1998). Gambar 6 menunjukkan bahwa PMSF dan TLCK menghambat aktivitas dialisat, sehingga dapat disimpulkan bahwa dialisat enzim termasuk enzim protease serin.
EDTA merupakan senyawa pengkelat yang dapat menstabilkan enzim. Cara kerjanya ialah dengan mengikat logam penganggu dan mencegah interaksi enzim dengan asam atau golongan –SH (Schwimmer 1981). Pengaruh EDTA (Gambar 6) menghambat aktivitas enzim kasar (0.01 mM), sehingga enzim kasar digolongkan sebagai protease logam. 0 20 40 60 80 100 120
EDTA PMSF TLCK STI kontrol
Inhibitor A k ti v ita s r e si d u a l (% ) crude dialisat
Gambar 6 Diagram pengaruh inhibitor terhadap aktivitas residual enzim ekstrak kasar dan dialisat.
10
aktivitas sisa sebesar 15% pada konsentrasi 1.0 mM, 7% pada konsentrasi 2.5 mM dan 0 % pada konsentrasi 5 mM.
Pada penelitian lain (Kim et al.1996) enzim fibrinolitik yang diisolasi dari Bacillus sp. galur CK 11-4 diturunkan aktivitas residunya sebesar 2.9% dan 58.6% pada konsentrasi 0.1 mM berturut-turut untuk inhibitor EDTA dan PMSF. Lebih lanjut pada konsentrasi 1.0 mM aktivitas tersebut diturunkan 21.4% dan 100 % berturut-turut oleh inhibitor dan EDTA dan PMSF. Berdasarkan data yang diperoleh dapat disimpulkan bahwa konsentrasi inhibitor berbanding lurus dengan daya hambat inhibitornya.
Pengaruh Pelarut
Dalam penggunaan pelarut organik, peubah yang harus diperhatikan adalah konsentrasi pelarut, protein, pH, kekuatan ionik, dan suhu (Harris 1989). Penambahan pelarut organik ke dalam larutan protein akan menurunkan tetapan dielektrik dan menyebabkan medium kurang sesuai bagi permukaan enzim yang polar. Berdasarkan teori kelarutan, hal ini akan menurunkan kelarutan protein. Adanya residu asam amino hidrofobik kadang-kadang akan menyebabkan terjadinya pelipatan molekul baru dan enzim menjadi tidak aktif. Oleh karena itu, penambahan pelarut organik dapat menginduksi proses inaktivasi enzim. Disamping itu, pelarut ini dapat memperbesar kemungkinan terjadinya denaturasi terutama pada suhu yang agak tinggi. Kelemahan utama dalam penggunaan pelarut organik adalah sifatnya yang mudah terbakar dan mahal.
Pada suhu diatas 10 oC, konformasi protein akan berubah yang memungkinkan molekul-molekul pelarut organik mendapatkan jalan masuk ke dalam struktur protein, kemudian merusak interaksi hidrofobik dan akhirnya terjadi denaturasi (Harris 1989; Scopes 1987). Pada Gambar 7 dapat dilihat bahwa rata -rata aktivitas sisa enzim kasar lebih besar dibandingkan dengan aktivitas residual dialisat. Hal ini menunjukkan bahwa penambahan pelarut organik lebih berpengaruh terhadap enzim kasar.
Pengaruh pelarut organik yang berbeda pada enzim kasar dan dialisat disebabkan jenis enzim Enzim kasar termasuk jenis enzim protease logam sedangkan dialisat termasuk enzim protease serin.
0 20 40 60 80 100 120
cloroform n-heksana benzena toluena kontrol
Pelarut organik A k ti v ita s r e si d u a l (% ) crude dialisat
Gambar 7 Diagram pengaruh pelarut organik terhadap aktivitas residual enzim eksrak kasar dan dialisat.
Penentuan Bobot Molekul (SDS−PAGE)
Elektroforesis dengan berbagai jenis gel telah banyak dimanfaatkan dalam teknik preparatif, di antaranya gel pati, agarosa, dan poliakrilamida. Polimer ini disusun oleh akrilamida dan N N ’-metilena bis-akril amida yang berpolimerisasi dengan bantuan suatu katalis atau sistem radikal bebas, seperti amonium persulfat dan N,N,N’,N’-tetrametilena diamina (TEMED). Penggunaan SDS berfungsi untuk memisahkan dan mengetahui jumlah dan ukuran rantai protein atau rantai subunit protein dan membuat keseluruhan protein diselimuti muatan negatif. SDS merupakan detergen lemah yang akan mengikat protein dan memutuskan ikatan antarsubunit protein. Dalam hal ini dipergunakan juga suatu pereaksi tiol seperti β-merkaptoetanol untuk mereduksi semua ikatan disulfida yang ada pada protein.
Metode elektroforesis SDS-PAGE 10% di-gunakan untuk menentukan bobot molekul ekstrak kasar enzim protease dari tape ketan putih. Perlakuan dilakukan pada enzim kasar, presipitat, dan dialisat. Gambar 8 menunjukkan nilai bobot molekul dari enzim kasar, presipitat, dan dialisat dari enzim protease tape ketan putih. Nilai BM yang diperoleh sebesar 11.60 kD. Nilai diperoleh dari protein baku yang telah diketahui bobot molekulnya dan dengan membandingkan nilai Rf yang diperoleh.
segera dianalisis sifat kehomo-genannya. Protein homogen menghasilkan satu pita (Gambar 8), sedangkan sub unit yang ukurannya berbeda menghasilkan lebih dari satu pita. Kurva standar SDS-PAGE dapat dilihat pada Lampiran 8).
Gambar 8 Analisis SDS−PAGE 10% pada C, enzim kasar,P, presipitat, dan D, dialisat. Dari pemurnian dan pencirian enzim protease dari A. faecalis (Thangam & Rajkumar 2002), dilaporkan bahwa bobot molekul yang diperoleh sebesar 67 kDa. Bobot molekul tersebut sekitar 6 kali lebih besar dari bobot molekul enzim protease yang diisolasi dari ekstrak tape.
Zimografi
Prinsip kerja analisis zimografi hampir sama dengan SDS-PAGE, hanya saja pada gel pemisah akrilamida ditambahkan jenis substrat yang akan dianalisis. Penggunan Triton-X 100 pada zimografi berfungsi untuk merenaturasi enzim. Triton-X 100 merupakan detergen nonionik yang dapat meningkatkan aktivitas enzim. Hal ini mungkin disebabkan oleh menjadi terbukanya sisi pengikatan substrat enzim yang memudahkan substrat terikat pada enzim. Pembukaan sisi pengikatan substrat oleh Triton-X 100 ini lebih lemah dibandingkan dengan oleh detergen ionik sehingga aktivitas enzim meningkat.
Tahap zimografi adalah tahap akhir untuk memastikan jenis enzim yang diteliti. Berdasarkan zimogram yang
diperoleh dapat disimpulkan bahwa enzim yang diisolasi dari tape ketan putih mampu menghidolisis substrat kasein, fibrinogen, dan gelatin.
Kemampuan enzim untuk menghidrolisis substrat dapat dilihat dari pita putih dengan latar gel biru. Berdasarkan hasil zimogram, substrat kasein menunjukkan satu buah pita (Gambar 9), berbeda dengan substrat fibrinogen dan gelatin (Gambar 10 dan 11).
LMW C P
Gambar 9 Zimogram kasein 12%
Pita putih yang dihasilkan pada substrat fibrinogen dan gelatin lebih besar dibandingkan dengan substrat kasein. Hal ini mengindikasikan bahwa pada substrat fibrinogen dan gelatin terdapat kompleks enzim dan substrat yang lebih banyak bila dibandingkan dengan kasein. Sementara itu pada albumin menunjukkan hasil yang negatif. Hal ini mengindikasikan bahwa jenis enzim yang dihasilkan oleh tape ketan putih tidak mampu mendegradasi albumin sehingga latar gel yang dihasilkan berwarna biru tanpa pita putih.
Gambar 10 Zimogram fibrinogen 12 %.
Enzim kasar Presipitat Dialisat
Enzim kasar Presipitat Dialisat
—
246.50
—
—
2911.60
—
20
—
12
Gambar 11 Zimogram gelatin 10 %.
Berdasarkan penelitian yang dilakukan dapat diketahui ciri isolat bakteri TP-4 yang diisolasi dari tape ketan putih secara umum (Tabel 3). Ciri biokimia tersebut terdiri dari enzim kasar, presipitat dan dialisat.
Tabel 3 Ciri biokimia protease dari isolat bakteri TP−4
SIMPULAN DAN SARAN
Simpulan
Berdasarkan hasil penelitian, enzim kasar dan dialisat enzim protease tape ketan putih mempunyai aktivitas maksimum pada suhu 37 oC, dan pH 7 dengan waktu inkubasi 10 menit. Enzim kasar merupakan protease logam karena dihambat oleh EDTA sedangkan dialisat enzim tergolong protease serin karena dihambat oleh PMSF dan TLCK. Tingkat
kemurnian dialisat enzim lebih besar dari presipitat tetapi lebih kecil dibandingkan dengan tingkat kemurnian enzim kasar. Enzim protease yang dihasilkan mempunyai aktivitas kaseinolitik, fibrinolitik, dan gelatinolitik dan tergolong protein homogen dengan bobot molekul sebesar 11.60 kD
Saran
Sebaiknya dilakukan identifikasi jenis bakteri yang dihasilkan dan perlu diteliti lebih lanjut korelasi antara kadar alkohol dengan enzim proteolitik yang dihasilkan oleh tape ketan putih.
Enzim kasar Presipitat Dialisat
DAFTAR PUSTAKA
Bergmeyer HU, Grassl MM. 1983. Method of Enzymatic Analysis. Ed ke-2. Weinheim: Verlag Chemie.
Bradford MM. 1976. A Rapid and Sensitive Method for Quantitation of Microgram Quantities of Protein Utilizing The Principle of Protein dyebinding. Anal Biochem 72: 234−254.
Parameter Enzim
kasar Dialisat
pH
optimum 7 7
Suhu
optimum 37 37
Inhibitor Tahan PMSF, STI, dan TLCK Dihambat EDTA Tahan EDTA Dihambat PMSF dan TLCK Pelarut organik Tahan cloroform, n-heksana dan benzena Tahan n−heksana dan toluena
BM 11.6 kD 11.6 kD Spesifisitas substrat Kasein, gelatin dan fibrinogen Kasein, gelatin dan fibrinogen
Jenis enzim Protease logam
Protease serin
Buckle KA. et al. 1987 Ilmu pangan. Alih bahasa oleh Hari purnomo Adiono. Jakarta: UI Pr. Chantawannakul P et al. 2002. Characterization of
proteases of Bacillus subtilis strain 38 isolated from traditionally fermented soybean in Northern Thailand. App Sci 28: 241-245.
Crueger W, Crueger A. 1984. Biotechnology: A Text Book of Industrial Microbiology. Madison: Science Tech.
Dwidjoseputro D.1977. Dasar− dasar Mikrobiologi. Jakarta: Djambatan.
Girindra A. 1993. Biokimia 1. Jakarta: PT Gramedia Pusaka Utama. Hlm. 88−113.
Gumbira−Sa’id E. 1987. Bioindustri: Penerapan Teknologi Fermentasi. Jakarta: Mediatama Sarana Perkasa.
Harris, 1989. Initial Planning in Protein Purification Methods. Di dalam: Haris S, Angal, editor. Protein Purification Methods Practical Approach. IRL Pr. Oxford. Hlm. 15−16.
produce from Bacillus sp strain CK 11−4 screened from chungkook−jang. Environ M icrobial. 62: 2482−2488.
Lehninger AL. 1994. Dasar−Dasar Biokimia. Jilid ke−1. Maggy Thenawidjaja, penerjemah; Jakarta: Erlangga.
Afrianti LH. Keunggulan makanan fermentasi. http:// www.yahoo.com. [11 November 2005].
Palmer T. 1991. Understanding Enzymes. Ellis Horwood Limited, West Sussex, England: Ellis Horwood Limited. Hlm. 40-55. Pelezar, Chan. 1986. Dasar−dasar
Mikrobiologi. (terjemahan Elements of Microbiology oleh Hadi Oetomo RS,Imas T, Tjitrosomo SS, dan Angka SL). Jakarta: Universitas Indonesia Pr. Hlm. 120−122.
Rahman A. 1992. Teknologi Fermentasi. Jakarta: Arcan.
Rao MB et al .1998. Molecular and Biotechnologi Aspect of Microbial Proteases. Microbiol. Mol. Biol. Rev. 63 (3): 597−635.
Scopes RK. 1989. Protein Purification New York: R.R. Donnelley and Sons.
Suhartono MT. 1989. Enzim dan Bioteknologi. Dirjendikti, PAU Bioteknologi, IPB. Bogor.
Seo. Ji−hyum, Sam−Pin− Lee. 2004. Production of Fibrinolytic Enzyme from Soybean Grifts Fermented by Bacillus firmus NA−1. Appl J Med Food 442−449. Thangam, Rajkumar. 2002. Purification and
Characterization of alkalin protease from Alcaligenes faecalis. Appl. Biochem. 35: 144−154.
9
Lampiran 1 Diagram alir penelitian
KARAKTERISASI ENZIM
ANALISA AKTIVITAS PROTEASE
ANALISIS SDS-PAGE DAN ZIMOGRAM
DIALISIS PRESIPITASI
PRODUKSI ENZIM KASAR ISOLAT BAKTERI
10
Lampiran 2 Prosedur pembuatan pereaksi kimia
• Kasein hammarsten 2% (b/v)
Sebanyak 2 gram kasein Hammarsten atau kasein teknis ditimbang, lalu dilarutkan dalam 10 ml buffer pH 7 sambil dipanaskan (50-60) kemudian distirer.
• Tirosin 5 mM
Sebanyak 0.091 ggram tirosin dilarutkan dalam akuades (tambahkan sedikit-demi sedikit sambil distirer) lalu ditera hingga volume total 100 ml.
• TCA 0.1 M
Larutan stok TCA 1 M dibuat dengan cara melarutkan 1.634 gram TCA dalam 100 ml akuades lalu diaduk dengan pengaduk magnetik hingga larut homogen. Larutan kerja TCA 0,1 M dibuat dengan cara mengencerkan 10 ml larutan TCA 1 M dengan akuades hingga volume total 100 ml(gunakan sarung tangan!)
• Na2CO3 0,4 M sebanyak 4.2404 gram Na2CO3 dilarutkan dalam 100 ml akuades, lalu diaduk
dengan pengaduk magnettik hingga larut. • Pereaksi Folin- Ciocalteau
Sebanyak 25 ml pereaksi Folin-Ciocalteu diencerkan dengan akuades hingga volume 50 ml dan diaduk dengan stirrer hingga homogen.
• Pereaksi Bradford
Sebanyak 0,1 g Coomassie Brilliant Blue G-250 dalam 50 ml etanol 95% v/v, dan 100 ml asam fosfat 85% v/v, lalu ditera dengan akuades hingga volume 200 ml. Larutan kerja Bradford dibuat dengan cara mengencerkan 5 ml larutan stok Bradford dengan akuades hingga volume total 50 ml.
• Komposisi media pertumbuhan dan media produksi Media Luria Bertani broth (LB 10%), pH 7
Yeast ekstrak 0.05 gram Tripton 0.1 gram
NaCl 0.05 gram Akuades 100 ml
lanjutan Lampiran 2.
Pereaksi untuk analisis SDS-PAGE dan zimografi
• Larutan A ( 30 % b/v akrilamida; 0,8 % b/v bis- akrilamida)
Sebanyak 14,6 g akrilamida dan 0,4 g bis-akrilamida dilarutkan dalam 50 ml akuades dan diaduk dengan pengauk magnetik hingga larutan homogen.
• Larutan B (bufer gel pemisah, Tris-HCl 2 M pH 8,8)
Sebanyak 75 ml larutan Tris-HCl 2 M pH 8,8 dan 5 ml larutan SDS 10 % (b/v) ditambahkan dengan akuades hingga volume total 100 ml.
• Larutan C ( bufer gel penahan, Tris-HCl 1 M pH 6,8)
Sebanyak 50 ml larutan Tris-HCl 1 M pH 6,8 dan 4 ml larutan SDS 10 % (b/v) ditambahkan dengan akuades hingga volume total 100 ml.
• Ammonium persulfat 10% (b/v)
Sebanyak 0,1 g ammonium persulfat dilarutkan dengan 1ml akuades. • Buffer elektroforesis
Sebanyak 1,803 g Tris, 8,648 g glisin dan 0,6 g SDS dilarutkan dengan 600 ml akuades, lalu ditera hingga pH 8,3 dengan HCl 1 M.
• Buffer sampel
Komposisi buffer sampel untuk SDS-PAGE terdiri atas 0,3 ml Tris-HCl 1 M pH 6,8, 2,5 ml gliserol 50 % (v/v), 1,0 ml SDS 10 % (b/v), 0,25 ml 2-merkaptoetanol, 0,5 ml bromfenol biru 1 % (b/v), dan 0,45 ml akuabides dengan volume total 5,0 ml. Sementara komposisi buffer sampel untuk zimografi terdiri atas 0,5 g SDS, 1 ml gliserol 50% (v/v), 1 ml bromfenol blue, 0,625 ml Tris-HCl 1 M pH 6,8 , dan 2,375 ml akuades denan volume total 5,0 ml.
• Larutan pewarna
Sebanyak 0,5 g coomassie brilliantb blue R-250 dilarutkan dalamcampuran 225 ml metanol, 50 ml asam asetat glacial, dan 225 ml akuades dengan volume total 500 ml.
• Larutan peluntur
Komposisi larutan peluntur terdiri dari 50 ml metanol, 50 ml asam asetat glacial, dan 400 ml akuades dengan volume total 500 ml.
• Triton X-100 2,5 %(v/v)
Sebanyak 2,5 ml larutan triton X-100 dilarutkan denagan akuades hingga volume total 100 ml. • Larutan berbagai bufer
Larutan kerja bufer 50 mM dibuat dengan cara mengencerkan 25 ml larutan stok bufer 0,2 M dengan akuades hingga volume total 100 ml. Bufer yang digunakan dalam penelitian ini adalah bufer-bufer universal (4-12), glisin HCl (pH 8.3), dan bufer Tris-HCl pH 6.8 dan 8.8
lanjutan Lampiran 2.
12
Larutan A: Asam sitrat (6.008 g) + KH2PO4 ( 3.895 g) + H3PO3 (1.769 g ) + asam dietilbarbiturat
(5.266 g ) dilarutkan ke dalam 1 liter akuades Larutan B : 0.2 N NaOH.
100 ml larutan A + X larutan B X ( ml) pH
6.4 3.0 15.5 4.0 27.1 5.0 38.9 6.0 50.6 7.0 63.7 8.0 72.7 9.0 80.8 10 86.0 11 99.6 12
• Penanda LMW
Lampiran 3. Data seleksi 9 isolat bakteri
Tape (TP)
A blanko (nm) Asampel (nm) Astandar (nm) Aktivitas enzim
(U/ml)
1 0.703 0.876 1.384 0.152 2 0.824 2.436 1.686 1.122 3 0.612 0.895 1.307 0.244
4 0.505 0.780 0.733 0.724
14
Lampiran 4. Contoh perhitungan aktivitas enzim (U/ml) dan aktivitas enzim spesifik
Perhitungan aktivitas enzim (U/ml)
Aktivitas protease (U/ml) = Asampel – Ablanko x fp A standar – Ablanko t Keterangan :
Fp = Faktor pengenceran T = waktu hidrolisis
Contoh perhitungan pada suhu 27oC selam 10 menit ( ekstrak protease kasar) Aktiviatas protease (U/ml) = 0.641- 0.619 x 6
0.961- 0.619 10
= 0.039
Perhitungan kadar protein
Persamaan kurva standar
Y = 0.641x + 0.1228 x = [BSA] mg/ml
y = Absorbansi (mg/ml)
contoh perhitungan kadar protein ekstrak protease kasar dengan nilai absorbansi 0.492 Y = 0.641x + 0.1228
0.492 - 0.1228 = 0.641x
x = 0.576 [Protein](mg/ml) = 0.576
Perhitungan aktivitas spesifik enzim (U/ml)
aktivitas spesifik enzim (U/mg) = aktivitas enzim (U/ml) [Protein](mg/ml)
= 0.039
Lampiran 5. Pembuatan kurva standar Bradford
No [BSA] mg/ml Absorbansi (nm)
1 0.025 0.135
2 0.050 0.147
3 0.075 0.172
4 0.100 0.189
5 0.125 0.211
6 0.150 0.220
7 0.175 0.243
8 0.200 0.251
9 0.225 0.262
10 0.250 0.288
11 0.275 0.292
12 0.300 0.313
Kurva Standar BSA
y = 0.641x + 0.1228
R2 = 0.9914
0.00 0.05 0.10 0.15 0.20 0.25 0.30 0.35
0.000 0.025 0.050 0.075 0.100 0.125 0.150 0.175 0.200 0.225 0.250 0.275 0.300 0.325
Konsentrasi BSA (mg/ml)
Ab
so
r
b
a
n
16
Lampiran 6. Data pengaruh suhu, pH, waktu, inhibitor, dan pelarut organik terhadap aktivitas protease enzim kasartape ketan putih
Pengaruh suhu
Suhu (oC)
A blanko (nm) Asampel (nm) Astandar
(nm)
Aktivitas enzim (U/ml)
Aktivitas residual (%)
27 0.619 0.641 0.961 0.039 5
37 0.505 0.780 0.733 0.724 100
50 0.593 0.620 0.820 0.071 10
60 0.571 0.708 0.940 0.223 31
70 0.589 0.673 1.298 0.071 10
Pengaruh pH
pH A blanko (nm) Asampel (nm) Astandar (nm) Aktivitas enzim
(U/ml)
Aktivitas residual (%)
4 0.638 0.691 1.030 0.081 11
5 0.707 0.733 0.895 0.083 11
6 0.699 0.775 2.074 0.033 5
7 0.505 0.780 0.733 0.724 100
8 0.642 0.730 1.104 0.114 16
9 0.720 0.727 0.850 0.032 4
10 0.611 0.669 0.955 0.101 14
12 0.662 0.697 1.011 0.060 8
Pengaruh waktu
waktu A blanko (nm) Asampel (nm) Astandar
(nm)
Aktivitas enzim (U/ml)
Aktivitas residual (%)
5 0.644 0.753 1.069 0.154 21
10 0.505 0.780 0.733 0.724 100
15 0.601 0.939 1.610 0.201 28
20 0.630 0.804 1.093 0.225 31
30 0.637 0.805 1.081 0.227 31
Pengaruh inhibitor
Inhibitor A blanko (nm) Asampel (nm) Astandar
(nm)
Aktivitas enzim (U/ml)
Aktivitas residual (%)
EDTA 0.407 0.378 0.544 0 0
PMSF 0.333 0.419 0.697 0.142 61
TLCK 0.440 0.447 0.590 0.028 12
STI 0.370 0.414 0.689 0.083 36
kontrol 0.631 0.728 0.883 0.231 100
Pengaruh pelarut organic
Pelarut A blanko (nm) Asampel (nm) Astandar
(nm)
Aktivitas enzim (U/ml)
Aktivitas residual (%) Cloroform 0.384 0.501 1.092 0.099 43 n-heksana 0.375 0.406 0.962 0.032 14 Benzena 0.4231 0.447 0.741 0.045 19
Toluena 0.404 0.396 0.748 0 0
Lampiran 7. Data pengaruh suhu, pH, waktu, inhibitor, dan pelarut organik terhadap aktivitas protease dari dialisat tape ketan putih
Pengaruh suhu
Suhu (oC)
A blanko (nm) Asampel (nm) Astandar
(nm)
Aktivitas enzim (U/ml)
Aktivitas residual (%)
27 0.533 0.625 0.921 0.142 18
37 0.251 0.747 0.625 0.796 100
50 0.537 0.546 0.960 0.013 2
60 0.532 0.579 1.225 0.041 5
70 0.448 0.555 0.890 0.145 18
Pengaruh pH
pH A blanko (nm) Asampel (nm) Astandar
(nm)
Aktivitas enzim (U/ml)
Aktivitas residual (%)
4 0.431 0.664 0.873 0.316 40
5 0.509 0.697 0.825 0.357 45
6 0.547 0.684 0.758 0.390 49
7 0.251 0.747 0.625 0.796 100
8 0.532 0.571 0.691 0.147 18
9 0.511 0.572 0.765 0.144 18
10 0.485 0.659 0.878 0.266 33
12 0.357 0.552 0.832 0.246 31
Pengaruh waktu
waktu A blanko (nm) Asampel (nm) Astandar
(nm)
Aktivitas enzim (U/ml)
Aktivitas residual (%)
5 0.517 0.630 1.004 0.139 17
10 0.251 0.747 0.625 0.796 100
15 0.486 0.594 0.971 0.134 17
20 0.627 0.696 0.933 0.135 17
30 0.575 0.587 0.935 0.020 3
Pengaruh inhibitor
Inhibitor A blanko (nm) Asampel (nm) Astandar
(nm)
Aktivitas enzim (U/ml)
Aktivitas residual (%)
EDTA 0.357 0.419 1.002 0.058 7
PMSF 0.418 0.325 0.808 0 0
TLCK 0.353 0.343 0.822 0 0
STI 0.404 0.423 0.807 0.028 4
kontrol 0.251 0.747 0.625 0.796 100
Pengaruh pelarut organic
Pelarut A blanko (nm) Asampel (nm) Astandar
(nm)
Aktivitas enzim (U/ml)
Aktivitas residual (%) Cloroform 0.458 0.463 1.037 0.005 1 n-heksana 0.477 0.552 0.773 0.152 19
Benzena 0.492 0.429 0.755 0 0
18
Lampiran 8. Kurva standar SDS-PAGE 10 %
Marker LMW (pita) Rf (cm) BM (kDA) Log BM
1 0.08 6.5 0.81 2 0.14 14.2 1.15 3 0.29 20 1.30 4 0.41 24 1.38 5 0.59 29 1.46 Enzim kasar,
presipitat, dialisat 0.25 11.60 1.0644
Kurva standar SDS-PAGE 10 %
y = 1.1029x + 0.8869
R
2= 0.7887
0.00
0.50
1.00
1.50
2.00
0.00
0.20
0.40
0.60
0.80
Rf (cm)
log B