KAJIAN
PERTUMBITHAN AKAR RAMBUT
Cinchona sp
D A N
INOKULASI
BEBERAPA
I
GALUR
Agrobacterium rhizogenes
PROGRAM
PASCASARlANA
INSTITUT PERTAMAN
BOGOR
Reflini. Study of haixy ruat growth of t'imhonu sp by inoculation af several Agrobaeterium rhizogenes strains. Under the guidance of Dr. Hj,
Anna
PrkganiRoswiem, klS and Dr. Nucita Toruan-Mathius,
MS.
APU.Problem encountered in bcukation of Cinchona species by A, rhizogenes is iow percentage of transformed root initiation and slow growth of hairy root. Strains
of A, rhizugenes and the growth medium
has
a sign&cant effect to the formation of!miry roots. The aim of
tkis
research was to evaluate the potmtia1 of several A. rhizogenes strains far initiation of hairy roots of C. mrccirubrud
C. ledgeriano, and to obtain the opthum composition of the medium for hairy roat culture of Cinchona species. Axenic stem and leaf explants of six to eight-week-old fromC, succirubra and C. ledgeriunca
~~
were inoculated with A. rhizogenes strains15834, 8196, l2-20001, 07-20001, A4,
R
MAFFA, TTSm 509, TISTR 510 rtnd LBA 9453. Inoculatd explants were cugured in solid MS medium. Subculture of thehavy root was performed by transfer of root pieces into fresh liquid basal medium MS,
B5,
White and HeIler. Hairy roots from the best of b a d medium were subcu1tured an the xime medium with addition of 50 mglf and 100 mg/IL-Triptaphane; three (MSJV) and five (M55V) times concentratien of MS vitamins. Thc integration of T-DNA in hairy roat was c o r n e d by using, spesific primer for
TL
and TR-DN A of A. rhizogenesin
Polymerase Chin Reaction (PCR) analysis. The results showed that only A. rhizogeraes LBA 9457 were effective to the ~ e c t i o n of explants from both Cinchona species. The h a q roots showed the fastest growth hMS
medium, while growth irz. others medium was poor. Hairy roots ofC,
ledgerianahas vigor and growth better
than
hairy r m s of C. su~cl'rubra. MSJV is the best medium for the improvement of hairy rout grawth and vigor. Transformed roots ofC. succi'rubra and C. ledgeriana uscd in this experiment were canfirmed t b t fialry
roots examined contained b t b TI, and TR-DNA region of Ri plasmid by PCR
amplificcttion d y s i s of DNA. The results obtained
in
this study indicate b t theSURAT PERNYATAAN
Dengan ini saya m e n y a ~ a n bahwa tesis yanl; berjudul :
Kajian Pertumbuhan Akar Rambut Cinchofla sp dari Inokulasi
Beberapa Gal ur AgrolZacteriu r#iizogeptes
addah benw mmup&an h a i l kmya say8 sendiri dm Mum panah dipublikasikan,
Semua s w b e r data dm informasi ymg digunakan telafi dinyat- secara jelas dm
dapat. diperikszt kebenxannya.
KAJIIAN
PERTUMBIJHAN
AKAR
RAMBUT
Cinchona
sp
DARI INOKULASI
BEBERAPA
GALUR
Agrobacterium
rhizogenes
. I-
Tesis
sebagai salah satu syarat untuk rnemperoleh gelar
Magiszer Sains p d a P r o p m Studi Bioteknologi
PROGRAM
PASCASARJANA
'
INSTTTW PERTANTAN BOGOR
Judul Tesis : Kajian prkurnbuhan rambut Cinchona sp dari inokulasi beberapa g l u r Aphucteriunr rhizogenes
Nama : Iieflini
MRP : 99650
Program Studi : Bioteknologi
Dr. Hj. Anna Priangani Roswiem, MS Ketua
Penufis dilahirkan di Payakurnbuh pada tmgga! 1 Februari 1976 sebstgai an& bungsu dari pasangan Yulizar (Alrn) dm Maridas. Penulis menyelesaikm Pendidikan Dasar tahun 1988, Sekolah hnjutan Pertma tahun 1991 dm Sekolah Lanjutan Atas tahun 1994. Pendidikm sarjana ditempuh di Jurusm Pendidikan Biologi, Fakultas Pendidikan Maternatika dan Ilmu Pengetahan Alam IKIP Padmg, lulus pa& t d u n
1999. P d a tahun<, 1989, penulis rnelanjutkan studi Strata dua (Sz) Program Seudi
Biotehologi, Program Pascasarjma IPB dm mmendapat gelar Magister Sains pada bulm
Puji d m syukur ke hadirat All& SWT atas limpahan &mat dsut h i d a y m y % sehingga tesis yang berjudul Kajian Pertumbuhsln Akar Rambut Cinchona sp dari Inokulasi Beberapst Grr lur Agrobact~rium rhizogmes, dapat diselesaikm.
Terirna kasitz diucapkan kepada Dr, Hj. Anna Priangani Roswiem,
MS
dm Dr. Nurita Toman-Mathius, MS. APU sel&u pembimbing, atas birnbingan, arabn dm dorongan moriel rnelalui diskusi ymg sangat berguna d a l m menmbab sertamemperluas w a w m untuk, menyelesaikm penelitian serta penuiisan tesis ini. Penulis mengucapkm terima kasih kepada kepda Unit Penelitian Biutehoiogi X)erkebunm, Bogar beserta staf yang telah rnernberikan izin serta penggunam failitas
peneiitim, 'Keberhasilan dalam xnenyelesaikan tesis ini tidak terlepas dari bmtuan
dma penelitim dari Dr. Nuritst Torum-Mathius, MS. APU, mtuk itu p w l i s
mengucapkan terima kasih yang sebanywk-banyaknya. Terima kasih juga diucapkan kepacla Katua Program Studi Biotehologi beserta staf, Direktw Program Pascasarjana TPB kserta staf ymg telah mernberikan ilmu dm p l a y a a n selamst penulis menempuh pendidikm.
Terim kasih juga pendis samgaikm kepada Bap& Tolhas Hutabarat, BSc- Dipl.Kim yang tel* memberikan bimbingan dm arahan ymg berhmga, Bapalc Suhartono, Dra. Nurhaimi-Hwis MSi, Mb& Nani dm selunrh s t d Laboratorium
Biarnalekul dip Imunologi atas bantuan dm kerjasama y m g sangat berawti, serta ternan-ternan Program Studi Biotehlogi angkatan 99.
Terirnst kasih ymg sebesar-besmya penulis ucapkan kepada Abak (alm), bu dm k&&-kakalrku y m g selma ini selalu memberikan perhatian, durongm rnoriel, kasih sayang dm da'a. Semuga Allah SWT melimpfiari batasan atas k e b a i h yang telah diberikm.
AXcbimya pexlulis bedlarap sernoga tdisan ini dapat bemanfaat dm
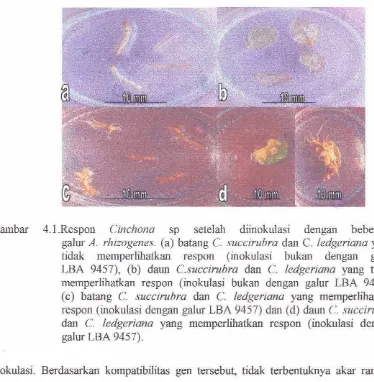
2. Respon Cinchona sp setelah diinokulasi dengm beberapa
gdur A, rhizogcnes ,
.
,. .
,.
, ,. . .
.
.
.
. . .
.
. .
. .
, ,. .
,.
+ ..
383, Respan awal C. succirubra dm C, ledgerianu setelah 3 rninggu
diinokulasi dengm A. rhizogenes galur LBA 9457
. . .
4 14. Respu? C. Succirubra dm C. ledgeriana pada ekplan &m dm
batmg sek1al-i diinalculzisi selama 7 rninggu ,
.
,. .
,.
,.
.
,.
,. . .
,. . . ..
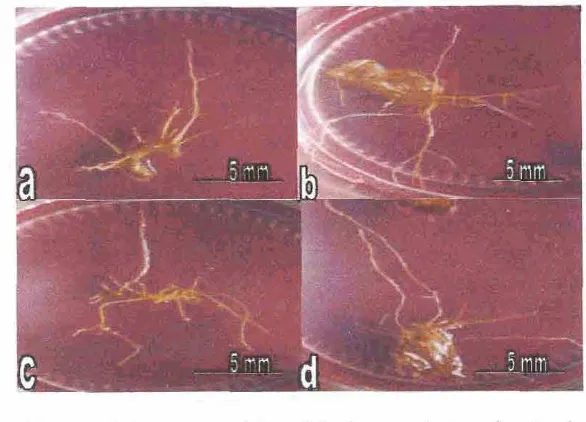
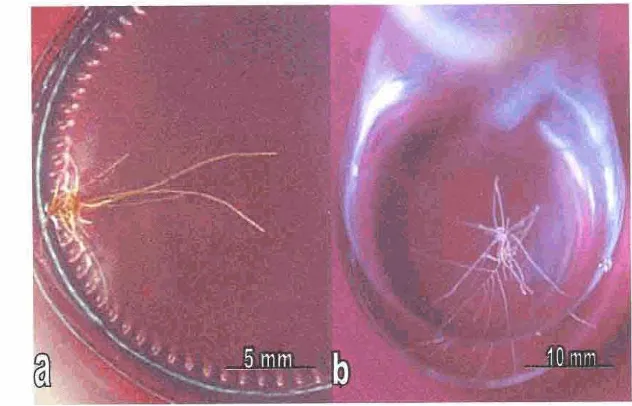
415. A k a rambut Qnmm kina..
. . .
,.
,. .
,. . .
,. ..
436 . Kultur aka ramhut C. Iedgeuiants umur 8 minggu
. .
,. . .
,.
, ,. .
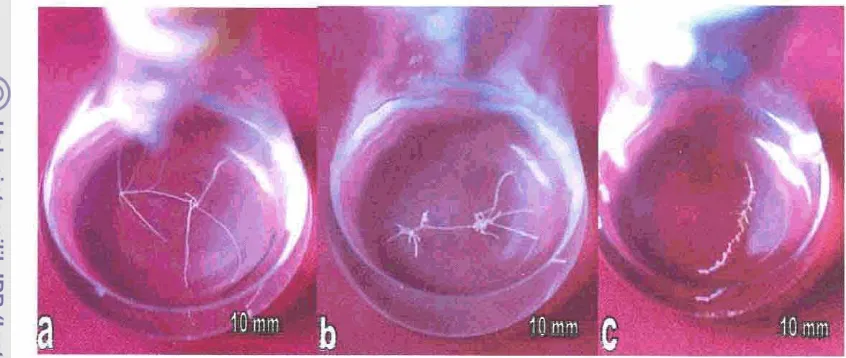
, 467. K u l ~ akar m b u t C. ledgevianu pada medium MS modifikasi.. 47
8, K u l m rarribut C ledgerianu pa& medium MS modifikasi 47
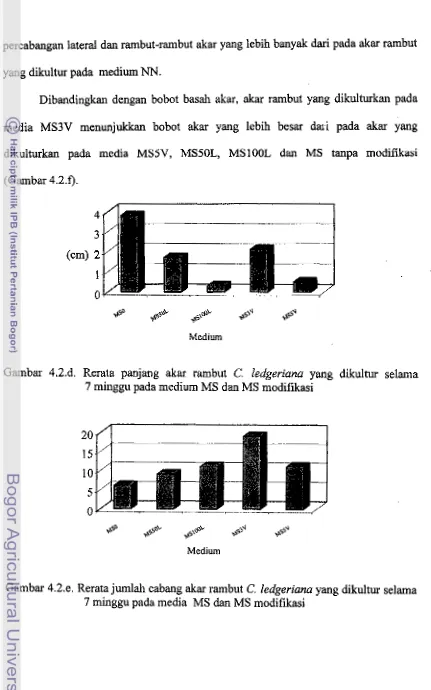
9, Rerata panjmg aka rambut C. Ledgerianu ymg dikultur s e l m
7 minggu pada medium MS dm MS rnadifikasi
. . .
,. . .
. .
.
.
.
,. . .
4910. Rerata jurnlah cabmg &ar rmbut C. ledgerianu yang dikultur
selama 7 Ginggu pada medium MS dm MS modifikasi
. . .
. .
.
.
. .
.
..
491 I . Rerata babat basah akar rambut C. ledgerianu yang dikultur
selama 7 minggu pada medium MS dm MS modifikasi
. . .
,.
,.
5012. Hasif fraksinasi DNA setelah PCR..
. . . .
,. .
,. .
,. . .
, ,.
, ,.
.
.
,No Teks Halaman
1 . Kompasisi beberapa medium dasar untuk pertumt7uhstn &ar rambut
tanaman kina
...
61Sebagim besar senyawa kimia yang digmakm pada indutri abat-abatan,
,*
makanm dm rninyak wmgi
Ire&
dari tumbuRm. Senyawa-senyawa tersebutbiasanya dixlmaicm rnetabolit sekunder atau produk sekunder
.
Senyawa-senyawametabulit s e k d e r ini merupakan h a i l metabulisme Xmjutan druri kxbohidrat, lernak
dan protein. Dalam tumbuhan, metabulit sekunder dapat: dijumpai dalm bentuk
alkaloid, terpenoid, steroid, mtrztsianin dm polifenul,
Tanamm kina (Cinchona sp) mengandung senyawa rnetabolit sekunder dalm
bentuk alkaloid kuinalin yang antara lain terdiri, dari kuinin, kuinidin, sinkonin dm
sinkonidin (Stiiba dm Chung 1981). Kuinin dm kuinidin GI& l m a digunakm
sebak;ai abat-abatan. Kuinin digunakan sebagai obat anti malaria, sedangkan kuinidin
selain digunakm sebagai obat anti malaria juga dapat dig&an sebagai obat unt&
menormalkan denyut jmtung yang tidak teratw (cardiac atyhtzic) (Verstrijden
1975). Pada industri minuman r i n ~ a n , kuinin biasanya digunakm set>@ pmberi
cita rasa (flavoring #gent) kaxena rasanya pakit (Anderson et al. 1986).
kuinolin yang meningbt. Tetapi tmmm kina rnemerlukan w&tu paling sedikit
tujuh tahun untuk &pat dipmen d m produksi alkaloid tertinggi banr dapat dipperoleh
setelah t m m a n berumur 20 tahun. Kendala tersebut rnenyebabkm obat
anti
mdwiasangat mahat dipasam, Untuk mengatttasi hd tersebut- tel& merangsang
obat anti malaria sintetik mengakibatkan kebutuhan alkaloid kuinulin kembali
mengalami peningkatan.
Pmspek pas& kina duniit pada kberapa eahun terakhir ini rnenunjukkm
kecendemgan y m g menggembir&m Permintam alkaloid kuinolin di psar d d a
diperkirdcan &an tcrus meningkat karma pengguamyrt yang semakin meluas
di krbagai bidang industri m t m 1&n industri famzstsi, industri minuman, incfustri
kasrndik dm industri biapestisida. Setiap t&un terjadi peningkatan permin-
alkaloid kuinolin sebsar 20 persen. Pada tahun 2001 kebuhrhan alldoid kuinulin
rnencapai 175 ton &an diperkirakan pada tahm 2005 mendatang a i m rneningkat
rnenjadi 275-300 ton dengm nilai sekitar US $9.9OQ.000 (SIL 2000).
Usaha-rr~aha y m g telah dil&ukm untuk memproduksi dkdoid kuinoIin
antara lain dengan memarifaatkan teknik kdtw jaringan. Menmt Fowler (1983)
salah satu keuntungm teknik kultur jztringan dibmdingkm dengm cara kunvensiond
addah kernampurn dalam menghasilkan senyawa kirnia d a l m wak-.tu ymg relatif
singkat dm kemampuan untuk rnernproduksi senyawa yang sulrar diperoleh seem
alami.
Staba dm Ckung (1 98 1) telah herhasit memproduksi kuinolin pada kultur
daun C, ledgerima sebsar 0,45% dari bobot kcringnya. Sedangkan Mdder-Krieger
er al. (1982) juga berhasil mendapatkan kuinin sebesar 95 pg/g bobat kering dmi
kdus C. pubmeens. Kuinin dan tiinidin juga berfrztsil diperuleh d a i kdtur aka
C. ledgerianu sebesar 0,010 mglg bobut kering dm 0,001 mg/g bubot kering oleb
Anderson el ui. (1982). Scragg (1986) berhasil memprolch kuinidin sebessu 0,053%
Sedangkan Hay (1986) mendapatkan peningkatan lima kali lipat kuinin dm kuinidin
pada kultur akai C ledgerianu dengan pemberian 500 mgll prekmur L-triptofm.
Produksi alkaloid kin# melalui kulhxr jaringan m a i h menunjukkan p r o d h i
y m g rendah. Hal ini diduga karma pertumtruhm sel yang lambat dm d~prlukannya
sejumlah hamon pertumbuhm dengan kunsentrasi yang tepat. modes (1986)
menerangkan bahwa banyak persenyawam tidak ekonamis dipruduksi meidui kultur
suspensi dm tingkat heterugenitasnya jugst tinggi. Kultur cenderung tidak: stabil dari
masa ke masa, Adslnya kcndala-kendala ymg dihadapi tersebut telah rnembuka suatu
pengembangan t e h i k baru mtuk menghasil kstn senyawa rnehbolit sekunder yang
lebih tinggi, yakni dengan memanfaatkm transfumasi Agrobacterium utuk:
merighasilkan &a rambut.
Agrobacteriokm merupakan bakteri tanah gram-negatif yang temasuk pada
kelompok Rhizobiaceae, mempunysti kernampurn untuk mentransfer sebagian bahan
genetiknya (DNA) pada sel tanaman melaiui gelukaan (Nilson dm Qlsson 1997).
DNA ymg ditransfex: disebut dengan T-DNA, yang rnerniliki gen-gen penyandi
enzirn untuk produksi opine (suatu kelompok konjugasi assun amino dm kabbidrat)
(Machstdo 1995). T-DNA rnenrpakan potongan beberapa rcttus kilo basa drtri plasmid
ymg dikenttl dengan Ri plasmid (root inducing plasmid') atau Ti plasmid (tumor
inducing plasmid), fii plasmid terdagat pada A, rhizogenes wdmgkan Ti plasmid
terdapat pada A. tumefascieras, T-DNA aka terintepsi pada kromosom tmamm dm
&an mengekspksikan gen-gen untuk mensintesis senyawa win. Selain gea unt&
rnensintesis senyawa apin, T-DNA juga mengandung onkagen, yaitu gen-gen ymg
terekspresi maka akan terjadi pertwnbuhan cepat dari sel. Ekspresi onkogen pada
plasmid
Ri
rnencirikan pembentukan aka adventif secara besar-besaran padaternpat ymg diinfeksi dm dikenal dengan 'hairy root' (akw rambut)
(Nilson dm OTsson 1 997).
Pemmfagtan kultur aka rambut untuk merighasilkan senyawa mekbolit
sekundcr telah banyak dil&ukm, antara lain lobelin dmi Lobeliu injlanra L.
(Yonemitsu et
QI.
199O), katarantin d m ajmalin dari Cutharc~nthus roseus (Vazquez-Flota ef 01. 18941, saponin dari Solmum aculeatr'ssurn (Ikenaga et al. 19951,
glukotropaeolin dari Tropaeolum majus (Wielmek dan Urbmek 19991, hiosimin
d m
skapalamin dari Atropa beliardonna (Kmada et al, 1986 ; Aoki ee6 al, 1997)
d m
Dutwra innoxia (Boitel-Conti et a!. 2000).
Kultur akar m b u t pada tanaman kina untuk mernproduksi kuinulin juga tel&
dilalrukan oieh Hamifi et al. (1 989) pada C. ledgerianu dengm A. rhizogenes gdur
LBA 9402 dm'R1000, dm Cieerling et al. (1999) pada C, oficinalis dengan galur
I,BA 9402. Narnun produksi kuinolinnya rnasih rendah dm diperoleh dalm periode
waktu lebih dari satu tahun, Hal ini discbabkan istju pcrturnbuhan akar rambut yang
ssuzgat lambat. Namun sistem produksi ini jauh lebih rnenguntmgkan dibandingkan
dengan vanamannya yang baru dapat dipmen setelah berumur lebih d a i 7 tahun.
Adanya kendda dalam kultur &ar rambut tanman kina disebabkan b l u m
ditemukan komposisi medium yank tepat untuk perturnbuh. akax rambut. Menurut
Gixi dm N a a s y (2000) medium tumbuh mcmberikm pengaruh yang nyata terhadap
p e ~ u m b u h m akar rambut yang optimal, kondisi kultur haws diatw padst
tingkat optimum.
Bercfasarkan hasil yang telab dlperoleh d m dalm upaya rri~ndstpatkan
alkaloid kuinoin yang lebih tinggi dilakukw penelitian terhadap kemampuan
A. rhizogenes untuk mengirrfeksi dm rnenghasilkan respon pada tanaman kina,
Penelitian ini rnerupakan pahap awal dari serangkaian penelitian yang d:lrencim&an
untuk menghasilkan teknulogi kultur &ar rmbut C, succirubra dm C. ledgerianu
yang mernproduksi alkaloid kuinolin. T&;tpm penelitian yang dildcuk-nuz
rnencangkup pengujian trmsformasi genetik behrapa galur A. rhizogenes terhadap
C succirubru dan C. ledgerima d m dilmjutkm dengm uji konfrrmasi T-DNA ymg
terintegrasi di "ddarn akar rambut C. succirubra dm C. Jedgeraiana dengan
rnenggunakan primer spesifik dari TL dm TR-DNA A. rhizogenes rnelalui tehik
Poly~ne~ase C h i n Reaction (PCR).
1,2 Tujuan Penelitian
Penelitian ihi krtujuan untuk (1) menyeleksi beberapa galur A. rhizogenes yang
dapat menginokulasi C', succiru bra dm C. ledgerianu, (2) mendapatlcan medium
yang optimum untuk gertumbuhan &a rambut: C, succir:rubra dm C. ledgeriana, dan
(3) mendapatkan akar rambut tanaman kina y m g telah terintegrasi T-DNA.
1.3 Hipatesis
Hipatesis ymg digunalcm d d m penelitim ini addah :
I . A. rhizogenes &pat rnenginduksi pernbentukan akar rambut pada sel jaringan
2, Komposisi medium kuItur d q a r rnempengaruhi pertumbuhan akar rambut
tanaman kin# C. succiuubra dm C. ledgeriana.
3. T-DNA dari A, rhizugenes dapat te~integasi pada &a rambut &~aman kina
C succr'rubra dan C ledgerr'una,
l,4 Kegunaan PeneXitian
Penelitfan ini diharapkan dapat, menglxasilkan teknik trmsformasi pada
tanaman kina dengan rnenggunakan gdur A. rhizogenes yang kumpatibel terhdap
genarn C. succr'rubru dan C. ledgerianu dan m e n d a p a t h medium terbaik ymg &pat
menghasilkm pertumbuhan akw rambut ymg optimal. Teknik y m g d i h a s i l h
2.1 Tinjauan Umum Tanaman Kina
Tanaman kina (Cinchona sp) menrpakan tanaman tropis yang b e r a d dari
benua Amerika Selatan d m termasuk suku Rubisceae (Croraquist 1981 ; Aerts el ai.
1991). Ada berbagai spesies tanaman kina, di mtaranya C, lanc$ulila,
C.
triameIC, cordifdid, C, succirubva, C, oflcinalis, C. micralathu, C. calopdera, C. josephiana
dan C, culisuya (Kerbosch 193 1). Di Indonesia, tanaman kina dibudidayakan
di drierah pegunungan pada ketinggian 800-1625 m di atas permukaan laut atau
kusmg dari 1950 rn di atas permukaan Iaut. Ada dua spesies kina y m g
dibudidaydcan, yaitu C. ledger iana dm C. succkubra,
Pohor- C ledgeriano kurmg kuku!i~, tetapi pertumbuhamya ceprtt, mmpunyai
percabangan ymg banyak dm tidak beratwan. Dam C, ledgeriana tebal, berwarna
~ j a u muda, berbentuk jorong, Xmset, dengm alur ymg kecil pacia tulang-tulmg dam
& p e m h atas dam. Bunga kuning pucat, bakd bbu& berbentuk telur sku tomb&
dengm panjang 8
-
15 mm. Kulit teba1 dengan warna cukeXat (Astika 19751,C, Szdccjrubra rnerniliki puhon ymg besar dm batang luws. Perhunbuhannya
cepat namun percabmgm sedikit, cabang-=bang muda berbulu. D a m y a besar,
tipis, b e n v m a hijau muda, berbentuk jorong & (40
-
50) X (30-
40) ern denganlapisan bulu sepanjang tulmg daun pada permukaan bawah, Bmga b e w m a me&
muda dm perrnukaan atas rnahkota berwma putib. B&al buah rnemanjang
2.2 Alkaloid Tanaman Kina
Tanman kina telah lama dikenal sebagai penghasil metabulit sekunder, yrtitu
alkaloid kuinolin. Kuinolin banyak ditemukan di ddam kulie batang tanaman kina,
sedangkan pada bagian lain seperti kayu, buah dm daun hmya dikmukan d d m
kadar yang relatif sedikit (Musalam er al. 1980). Kurang lebih 35 macam alkaloid
kuinolin telah ditemukan pada tanaman kina, n m u n hmya terdapEtt empat macam
kuinolin u m a yaitu kuinin, kuinidin, sinkonin dm sinkunidin (Urdang 19451,
Kuinin dan kuinidin mempaksm turunan dari 6-metoksi-kuinolin, dm
mempunyai m u s empiris yang sama, yaitu C ~ ~ H 2 4 N 2 0 2 dengan bemt rnolekul 232
(Verstridjen 1975). Keduanya menrpakan stereoisomer yang dibedakm d a m
karakteristik kikia dm fisikanya, sehingga mempunyai sifat yang berbeda. Perbedm
bentuk ruang kedua molekul ini berasal d x i orientasi yang berlainan dari
atom C pusat ymg tidak rnempwyai simefri dm dapat terikat ppada atom-af0In
dengan dua macam susunan, ywg satu merupakan baymgan cemin yang I&
(Verstrijden 19751,
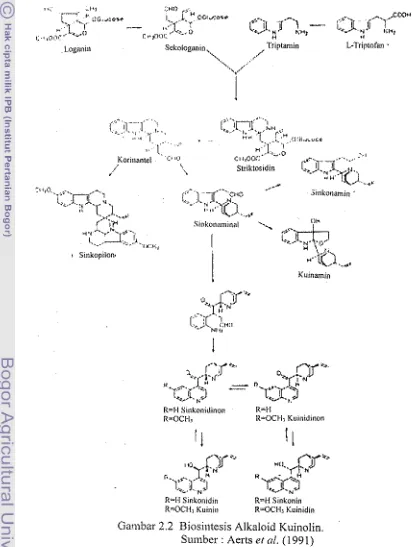
Biasilltesis alkaloid kuinolin dapat dilihat pada gmbar 2.2. P=mbntukan
alkaloid kuinolin diawali dengan perubahan triptofan membentuk triptamin dengm
banturn enzim triptofan dekarbksilase. Selanj utnya terj adi kondensasi ezimatik dari
triptamin dm sekolagmin untuk membcntuk striktosidin, terw rnembenluk sejunlah
turunan dkdoid berikutnya termasuk alkaloid kuinolin (Aerts et al. 199 3).
Setiap jenis Cinchona mempunyai kandungm alkaloid kuinolin yang berbeda.
C. Eedgeriana diketahui memiliki kadar kuinin ymg tinggi (4
-
13 %), sedmgkankadar sinkanidin C, succirtib~.~ lebih tinggi yaitu 3,2 - 5,1 a/o dari pada C Itdgeriunn
R=l.l Sinkonidinon l<=H
R=OCH, tl-OCl,lr K u i ~ ~ i d i n o n
R-1.l Sinkonidin 8-M Sinkonin
[image:101.611.72.484.190.737.2]R=QCI~I Kitirliu R=OCHj Kuinidin
Gambar 2.2 Biosintesis Alkaloid KuinaIin.
2,3 Karakteri~tik Agrubacterilrm rltizoge~les,
Genus Agrobacferium merupakan bakteri gram-llegatif, temasuk kelomgok
Rhizobiweae dm bersifaf aerobik. Ada dua spesies Agrobacterium y ~ , g diketahui
sebagai patugen pada tanaman, yaitu A. tumefusciens
cfan
A. rhizogenes. Keduaspesies tersebut dapat menginfeksi tanaman dikotil dm sebagim kecil monokotil
rneldui pelukaan. hfeksi yang disebabkm oleh A. fumefusciem dapat menyebabkan
terbentuknya tumor pacia tempat infeksi yang disebut dengan crown gall. S e h g k a n
infeksi oleh A. rhizogenes dapat membentuk sernacam akw adventif secara besar-
besaran yang l z i m disebut dengan hairy root (aka rambut). Menurut KIee es ab.
(1987) tumor dm aka r m b u t yang terbentuk dapat tern tumbuh secma alrsenik
wdaupun Agmbacterr'um-nya telah mati, Pertumbufxm tersebut dapat hrianj ut pada
medium tmpa rat pengatw tumbuh,
hduksi tumor dm akar rmbut terjadi karena sebagim DNA atau yang
disebut dengan T-DNA dari Agrobacferiwm ditransfer ke ddam sel tanaman. T-DNA
tersebut memiliki gen-gen untuk mensintesis auksin dm sitokinin, sehingga apabila
terekspresi &an rnengakibatkan terjadiny~l over produksi hormun tersebut di dalm
sel tanaman (Klee et al. 19871,
T-DNA merupakan bagian dari megaplasmid yang ada dalam Ag~.obacierium
. J,
yaitu Ti-plasmid (tumor inducing) pada A. tumefasciens atau =-plasmid (root
inducing) pa& A. rhizuge~zes (Winans 1992). Ti-plasmid h a t : d i b e d a h
berdasarkan tipe opine yang dihasilkan, yaitu nopalin dm oktopin. Sedangkan
Ri-plasmid dibedakan rnenjadi Ri-plasmid kelompok manopin dm kelompok agropin
Pada A, rhizogenes, Ri-plasmid mernpunyai satu atau
dua
macam T-DNA,yaitu Iefr T-DNA (TL-DNA) dm right T-DNA (TR-DNA). Berbeda dengan
TL-DNA, TR-DNA mernpunyai persamactn dengm T-DNA A. tunaefasciem.
TR-DNA mengmdung gen-gen iaaM dm iuuN yang berfungsi unhxk. biosintesis
auksin, dm juga mernbawa gen-gen untuk menyandi sintesis senyawa apin (Giri dm
Narstsu 2000). Sedangkan TL-DNA mengandung gen-gen rul. Slightom et al. (1986)
rnengidentifikctsi 18 open reading frames (OWs) pada TL-DNA pRi A4. Pengujian
yang dil&ukan pada daun dm ba-g lcalanchoe menunjukkan bahwst dari 18 lokus
yang krpatensi pada TL-DNA, hanya empat Iakus ymg tamp& mempengaruhi
morfologi &ar rambut ymg djhmsiih, y h i lokus A-D (ralA-a), Lukus-Iokus
tersebut berupa ORF 10 (rol A), ORF 1 1 (uolB), ORF 12 (ro2 C) dm
O W
I5(rul D). Produk gen rol ini drtpat: mengakibatkan terjadiiya pembentukan akar
rambut. White et rrl. (1985) nzelakukaxl pengujian terhadap plasmid agi- pin pRi A4
yang mernbawa d m dapat melztransfer dua T-DNA (TL dm TR) pada sel tmamm,
TL-DNA pRi A4 rnenunjukkan prsamaan dengan T-DNA gdur 8196 yang
menyandi sembiian gen tranship.
2,4 Mekanisme Infeki Agrobakterium rhizugmes terhadap Sei Twnaman
Secara umum proses infeksi A. rhizogenes mirip dibmdingkan dengan
A. tumefasciens, Ada dua daerah yang terlibat dalam proses transfer DNA yaitu
T-DNA dm daerah vir ((virulence). T-DNA rnerupakm DNA yang ditransfer
ke dalam genom sel tanman, sedmgkan &aerah vir merupaXcan daerah penyandi y m g
%-plasmid mempunyai satu atau bebentpa T-DNA yang berbeda, dibatasi oleh
25 pasany basa susunan berulang (Nilsson dm O l s m 1997).
lnteraksi awal .$grobacterium dengan sel tanamm didrrhului dewm
pengadan Agrobacieriurn terhadap sel yang Iuka. Interaksi ini tmjadi secara kimiawi
dimma sel tanaman yang luka akan mengeluarkan suatu metabalit berupa senyawa
guia, asam amino atau senyawa fen01 (Winms 1992). Dengan admya mebbolit ini,
Agrobacrerium akan bergcrak &if (kemotaksis) menuju sel &arnan. Selanjuhlya
ferjadi kuntak: antara Agro bacterium dengm sel tanaman. Untuk memperkuat kuntak
sel tersebut Agrobacferium mengeluarlcrtn metabulit
P-
I ,2-glukan. Glukm diperlukanuntuk pengikatan sei tanaman bagi Agrobacterium. Behrapa gen &am krumosum
Agrobacierr'um y m g diketahui rnerupakm penyandi enzim yang berperan ddm
sintesis berbsgai senyawa glukan, yaitu chv A (merighasilkan protein yang berfungsi
untuk mentransportasikan
P-
1,2-glukan) dm chv B (mensintesisP-
1,2-gluk:m], Genlain yang ikut bcrperan adalah cel, yaitu beyeranan dalam sintesis senyawa seldom
fibril (Douglas tsef ul. 1985).
Mekanisme selanjutnya addah induksi ftdktor vir ymg &an mengatur
pernotongan dm pernindahan T-DNA ke ddam sel tanaman. Faktor vir terindksi
akibat adanya scnyawa fen01 yang dikeluarkan tanman yang l&a. Beberapa senyawa
fen01 yang dapat rnengindulcsi fkktor vir 'rdaalah asetosiringuri, hidroksiasetosiringon,
koniferil alkohul, koniferin (feniprupunuid g1uHcosida) dm etil ftrulat (Winans 19921,
dapat rnenginduksi faktor vir, apabila asetosiringon tidak diproduksi
(Cangelasi et al, 19891,
Ada enam fakbr virulensi yang diketahui berfungsi dalam proses p e m i n d a h
T-DNA, yaitu vir A, vir B, vir C , vir
D,
vir E dm ~ i r G. Senyawa fenulik yangdikeluukan tanaman yang luka &an menginduksi vir A unh& memprodulcsi suatu
, .1.
protein Vir A. Protein Vir A mengalami autofosforilasi dm &an menginduksi
fosfurilasi protein Vir
G,
Selanjutnya potein Vir G &an mengaktifkan gen vir Iainnya(PawelI er a1.. 1989). Praduk berupa protein Vir Dl dm Vir
Dz
&an memotongT-DNA pa& daerah flanking (daerab y m g susunan DNAnya bedang). Daerah
tersebut sebagai pembatas kiri (lefr border) dan pembatas b a n (~ight border) dari
T-DNA. Karnpanen pembaias kanan rnutlak diperlukm ddam transfer T-DNA
(Klee et al. 1987). Adanya pernotongan oleh gen 17ir D akan rnenyebabkm
terbentuknya rnalekul DNA u h s tunggal (ss-DNA) atau disebut dengan T-strand,
Stmktur T-sirund yang ditransfer dari sel Agrahucferium ke sel tanma*. menrpakan
komplek DNA-protein. Sdanjutnya protein Vir D2 &an mengwahkan patongan
T-DNA tersebut rnenuju nukleus sel ttlnamm. Protein Vir D2 juga diperlukan mtuk
integai T-DNA pada genum tanaman (Mysure e6 a!, 1998). Protein pi-uduk vir I3
akan merneaiasi transfer T-DNA ymg diduga membentuk pri y m g &an dilewati
oteh T-DNA dari sel Agrobacrerium ice sel tanaman. Kemudian proses ligasi dan
sintesis T-DNA utas komplemennya dilakukm oleh topoisomerase dm ligae
tanman. Selma proses transfer, T-DNA dilindwlgi aleh pratein pruduk vir C dm
vir E, Protein Vir C juga dapat berfungsi untuk meningkatkan aktifitas protein Vir
D
berfungsi rneiindunginya dari kerusakan akibat nuklease sel tanama~ Pengikatsn
protein Vir Ez pada T-strand yanp sangat kuat akan menjadikan sWtur T-komplek
stabil dm terjaya selama proses transfer. T-komplek ini diperLr&an panjmgnya
3600 nm dm lebar 3 nm, mzngmdung sekitar. 600 rnolekul Vir E2 dm m u rnalekul
Vir D2 dengm perkiraan berat rnolekdnya 50,000
kD
(Sheng dm Citavshj 1996).2.5 EIrspresi T-DNA Pada Genom Tanaman
T-DNA yang ditrmsfcr ke dalztm sel tanman bisa lebih
dari
satu maiekulT-DNA, sehingga di dalam krornosarn tanaman &an terdapat satu atau beberapa utas
T-DNA. Situs integrasi T-DNA pada kromosom tanaman bersifrtt ac& (Armitage
1 987). T-DNA &an terintegrasi dengan stabiI pada komasom tamm (Peterman
1997 ; Giri d m Narasu 2000). Gen-gen pada T-DNA &an diekspresikan aleh
Canaman untuk keperluan Agmbucterium.
T-DNA mengandung gen-gen yang beperan untuk biasintesis senyawa opin
(opine cufubolism), yaitu merupakan turunan guia dm a r m amino. Opin ti&
dimetabolisme o1eh tanaman tctapi dieksre-esikan ke lingkungan, dm dimetabaiisasi
aleh Agrobacterr'um, Upin dirnwfaatkan aiek Agrobscterium sebrtgai s u b karbon
dm nitrogen, Dengsm demikian t u j w Agrobacferium menginfeksi sel tanaman
adalah wtuk mensintesis senyawa apin (Nillsan dm Olsson 1997).
Di samping gcn-gen untuk rnensintesis apin, T-DNA juga rlrengandung
onkagen, yaitu gen-gen yang rnenyandi pembentukm hornon pertumbuhm auksin
dm sitakinin. Integrasi T-DNA di &lam krumosom tanaman mengakibatkan
homan. Hal tersebut rnenyebablcan perturnbuhan scl yang tidak terkuntroi. Ekspresi
dari gen inilah yang rnengakibatkan terbentuknya a h rambut.
Pada TR-DNA, gen yang b.:rfungsi untuk pemkntukan auksin addah iaaM
(gen 1, ims 1, avx 1, shi) dm iaaH fgen 2, tms 2, avx 2, shr). iaaM rnenyandi
pembentulcan triptafm 2-monoaksigenase yang &an mengkatdisis perubahan
L-tripto fm menj adi senyztwa indole-3 -asetamida. Sedmgkan iaaH rnenyandi enzim
amino hidrolase yang merigubah indale-3-asetamida menjp-di asam indole-3-asetat
(IAA) (Thomashaw et al. 1986). Gen i p f (gan 4, tmr, mi) mempstaan penyandi
enzim iso-penteniltrmsferase yang &an mengkatdisis sintesis sitokinin (sitoknin
iso-penteniladenosine 5'-monofosfat) dari dimetildil-pirofosf dan S'AMP
(Akiyoshi et al. 1 984).
Pada TL-DNA terdapat onkogen yang disebut dengm gen rol (root locus).
Gen yo1 rnenyandi pembentukstn protein, termstsuk protein yang terlibat d d m
regulasi rnetabdisrne hormon tanaman, White et al. (1985;) rnelalcukm pengujian
genet% pada Ri piasmid dari A. rhizogenes dm ditemukan adanya empat gen rol
yaitu rul A, r01 B, rot C dm rub D. Hwil pengujian rnenunjukh bahwa protein
prod& dari hrbagk gen ral beperan dalam meningkatkan sensitivitas sel tanaman
terhadap adanya auksin. Selain itu produk gen rol juga diduga mendorang
tanaman inang untuk mensintesis auksin sehingga mengakibatkan terjadinya
pembentukm &are
Vilaine er al. (1 987) menjelaskan bahwa ixduksi stkar y m g disebabkm oleh
rol A mernberikan suatu perubahan fenotip dm rnenmbah efek pelukaan pada
pembentukan akar rambut. Selain ilu juga terdapat rol C dm rul
D
yang fungshyadidupa juga bzrhubungm dengan pertumbuhan
akar rambut.
Menurut Nillson dmOlsson (1 997) ekspresi gen rol secam bersama akan mernberikan efek yang Iebih kuat
dari p d a hmya satu %en.
2,6 Kultur Akar Rambut Dalam Praduksi MetaboXit Sekunder
Kultur In vitro ttlah banyak digunakan untuk tujuan produksi metablit
sekunder. Tehik kultur yang sering digunakan diantaranya addah kultur kdus,
kultur suspensi sel dm kultur organ. Teknik: hi kmrientasi pada tmri totiptensi sel.
Totipotensi sel tersebut disebabkm semua sel Aiciup suatu tamman mmniliki DNA
yang sama. Hal ini berarti bsthwa sel mengandung semua informasi genetik
yang sama.
Pada kultur organ, akar merupakan surnber metabulit sekunder y m g penting,
temtma untuk jenis-jenis persenyawam yang berasasiasi dengan &ar, Kultur aka
yang diisolasi pada umumnya merniliki sifsat-sifat morfologi ywng sama seperti &ar
yang terdapat pada tanaman. Pada ujung &ar juga terdapat quiescent c e m r dm pula
penyebaran, pembuluh yang spesifik dari spesies masih &pertahadcan (Gunawan
1992). Alcar yang sudah diisoiasi bistsanya diturnbuhkm dalam media cair dengm
pengocokan perlahan-lahm. Untuk rnendapatkan kultur &a dengan pertumbuhan
ymg optimal d m dalam w&tu yang lama dapat dilalcukan dengan memberikm
nutrient dm hormon yang tepat. White (1934) ~ U I I I M Gumwan (1992) berhasil
memperlihatk? perturnbuhm akar yang tidak terbatas dalam kultur &a tomat
hrhmil melakukan kultw akar beberapa spesies tertentu dari Nicotiana seperti
N. langsdorfii dan N fabucurn. Pemanfaatan kultur akar untuk produksi metablit
sekunder teIah banyak dilakuksn, misaXnya produhi ginseng dari aka Panax gimeng
dm produksi shikonin dari
akar
Lifhospermurn. Namun karena pertmbuhanakar
yang relatif lambat, kultur &ar menjadi tidak digunakan Iagi &lam pmduksi
rnetabolit sekunder.
Kdtur akar krkembmg 1agi sejak ditemukan A. rhizogenes yang h a t :
menginduksi pernbentukan akar pada tanaman yang terinfeksi. A h yang terkntuk
dapat tumbuh dengan cepat. dan juga tidak memerlukm human pertumbulnan
eksogen. Akar tersebut sering disebut dengan "Hairy rooff'
(dm
rambut) karenaumumnya mernperlihatkm morfologi ymg bertPeda dengm &ar tamman induknya.
&=-dm
tersebut biasanya rnernbentulc cabang-cabang lateral dm rmbut-mbut&ar yang lebih banyak.
A k a rambut terbentuk kslfena integasi T-DNA pada genorn sel tanman.
T-DNA merupakan bagian dari DNA piasmid A. rhr'zogenes y m g ditrmsfer ke dalm
sel tanaman yang terinfeksi. T-DNA mengandung gen-gen untulc mensintesis harmon
pertumbuhn, Ekspresi dari gen-gen tersebut daiam sel tanman mcnyebabkm
terbentuknya
akar
adventif pada tempat infeksi.Kultur aka rambut telah banyak dikembangkm pada berbagai jenis tanaman,
terutama untuk produksi metabolit sekunder. Sejumlah penelitim kultur A m rambut
untuk produksi metabolit sekunder antara lain produksi alkaloid trogane dari kuttur
belladonna dm Calystegia sepium (lung dm Tepfer 1987) dm produksi sapunin dari
Panax gimeng (Yoshi kawa dm h y a 1 987).
Keunggulan kultur a k a rambut dibandingkm dengan kultur lainnya adalah
perturnbuhm sel yang cepat dm tidak rnernerlukm z6lt pengatw tumbuh, sintetik.
Selain itu sifat gendiknya tetap stabil dengan pmduksi metabofit sekundw yang
tinggi. Momciiovic et al. (1997) memmukm
TR
d m TL-DNA dari A. rhizogenestetap stabil pada &ar rambut Geritima yang telah b e m u r lebih dari satu tahun.
Sedangkan Ladhi d m Charlwoad (1996) bwhwil mendapatkrtn &umulasi total
mtrakuinan dm alizarin dalam aksr rambut Rtkbiu peregrina L masing-masing dua
kdi dan tiga kali lebib tinggi dari pada &ar biasa yang b e m w 1 tahun. Tm&a
(1999) juga melaporkan b&wa Vinca minor yang ditrmsfomasi dengan gdw
MAFFF03-0 1724 merighasilkan vinkmin dm kali lebih tinggi dari tmmm ymg
ti&& ditransfomasi. Sedangksm Ssluerwein d m Shirnamora (I 999) juga mendapatkan
dkaloid yang , s m a sebmyak 2% dmi &ar Hyoscyapnus albui yang ditrmsform%si
dengan galur yang s m a , dm dikuitw pada medium Garnborg dengan 3% sukrosa.
Keuntungan lain dari akm mmmb hhasil transfarmasi gen dari A, rhizogenes
adalah dapat dilakukan pnyisipiur gen-gen ymg bertujuan untuk nreningkatkm
produksi metabof it sekunder. Boehm et a/. (2000) berhwtsil mendapatkan peningkatan
produksi shikonin sebsar 22% dmi kontlrol pada kultw &m rambut Lithospermum
eryfhrorhizon dengan menyisipkan gen uhi A. Akar rambut juga dapat beregenemi
mmjadi tunas d m tanaman lengkap, Kecepatan pertumbuhm dm frekwensi
Selain itu macam-macam fenotip yang timbul dari regenerasi &ar rambut merupakm
HmiU et al, (1989) rnelakukan kultur akw rmbut p#da tanaman kina
in vilro dengan A. rhizogenw ggalur LBA 9402 dan R1000. Hail y m g diproleh
rnenunjukkan bakwa akar rambut lebih banyak terinduksi oleh LBA 9402, namun
perhrmbuhan aka sangat lambat. Beberapa aka hasil inokulasi LBA 9.:i)2 tersebut
d i k u l t w h di ddm medium
BS
dengan 3 % (w/v) sukrosa. Dari kultur tersebuthrhasil diperoleh kinin, sinkonin dan kuinidin sebagai kompanen utama sekik
50pg/g babot basah setelah dikuittar 45 h a i , H a i l teabut masik rendah
dibandingkan dengan tanaman secwi alami. Selmjutnya Geerling el al. (1999)
melstkukan kultur akar rambut padst C. oficinalis dengan menyisipkan dua gen kunci
dalam biasintesis alkaIoid kuinolin ymg krasal, d a i Cutharaptrhus ~.oseus, yaia
triptofan dekarboksilase (TDC) dm striktusidin sintase (STR). Untuk rnendapatkm
akar rambut ymg merigandung gen fdc dan srp, inokulasl dClakuZcan pada dam
tanaman hmi1 perkecambdm benih s e w a rnksenik dengm menggunakan
A. rhizogenes galw 9402 ymg teIafr rnembawa kedua gen tersebut. Dengan
penyisipan gen tersebut diperoleh peningkatan kuinin dm kuinidin masing-masing
sampai SOOpg/g dm 100pg/g b b t Irering, n m u n satu tahun setelah andisis, &a
2.7. Folktor-Faktor yang Mempengaruhi Kultur Aksr Rambut.
Pengguwn kultur &a r m b u t hasii transabrmasi genetik A. rltizogenes unt&
rnemperoleh senyawa metabatit sekunder telah banyak dilakukan pada berbagai jenis
tanaman, Pertumbukan akar rambut yang cepctt dan kemmpuam sel untuk mensinksis
senyawa-senyawer kmia &pat dijadikan sebagai swnbr untuk memgroduksi
rnetabolit sekunder secarst terus-menems. Untuk merzdapah pertumbuhm kultur
dengm produlcsi rnetabolit sekunder y m g tinggi kundisi kulm hams diatur pada
tin@& optimal.
KuItur &a m b u t mengikuti pala pertwnbuhan tertentu, a h tetapi prodhi
metabulit sekunder rnungkin tidalc berhubungan dengan pertumbuhm Inrltur. Menurut
Giri dm Narasu (2000) biosintesis metablit sekunder di ddam kultur aka rmbut
dikontrol secara genetik, teapi juga dipengaruhi oleh nutrisi dm faktoi lingkungan,
Komposisi medium kultur bepengaruh pada pertumbuhan dan produksi metablit
sekunder.
Di ddam kultur jaringan, ada tiga jenls medium kultur yang d i g t d a n yaitu
medium padat, semi padat dm a i r . Medium tersebut: tersusun dari gam-garam
mineral atau nutrient marganik yang terdiri atas unswr-unsur m&o dan &o,
sumber karbon yang biasanya berupa gula atau sukrosa, vitamin, asm mino, zat
pengatur tumbuh dm zat-zlat tslmbahan orgmik Xainnya, seperti air kelapa, ekstrak
ragi, ekstrak kentang dan sebagainya.
Sulxosa rnerupakan sumber k w b a paling baik dm dihidrolisis menjadi
glukosa dm fruktosa oleh sel tanaman selama asimiki. Di ddam aka rambut tingkat
produksi metabolit sekunder. Emawati (1 990) melaporkan - bahwa peningkatan
konsentrasi sukrosa sampai 6% meningkatkan produksi senyawa anti jamur pa&
kultur akar rambut P. tinchioriurn Ait. Sebdiknya Granicher el al. (1992)
rnernperoleh praduksi valepatriates dalam kultur akar rambut V. oflcirulis
L
palingtinggi pada medium setengah Gambarg B5 dengm 2% sukrosa dibandingkm deqan
3% dm 5% sukrosa.
Nutrient anorganik m&o rneliputi unsur-unsur K, N, Mg, P, Ca, S, Na dm
C1, sedangkan nutrient mikra meliputi unsur-unsur I, 3,
Mn,
Zn, Mo, Crr, Co, Al, Nidm Fe. Untuk pertumbuhm akar rambut, diperlukan susunan, nutrient dl, dalam
medium yang h a s dicukupi s w a a kualitatif maupun kuantitatif. Hal i;ti sebagian
besar tergmtung kepda keadaan fisiofogi jaringan tersebut. Di samping susunm
nutrient, tekanan osmosis dm kekuatm ion ymg ditimbdkan di dalam sel juga
b e r p e n g d pada perturnbuhztn akar rambut. Menunat Sikuli dan Derneyer (1997)
perbedam kandungan ion-ion di dalam medium
B5
yang digunakan sebagai mediumkultur stlrar rambut Datura srrumoniuwz ternyata b e r p e n g d t a h d a p produlcsi
hiosiamin dm tropin. Produksi hiosimin diperaleh lebih tinggi pada medium
B5
dengan kandungan SO:- dm K' dorninan. Sedangkan produksi tropin lebih tinggi
jika kandungan
ca2'
dan NU3' dominan. Ernawati (1 990) rnemperoleh media terbdkuntuk merangsang pertumbuhan aka rambut
P.
tiachtorium Aif adalah mediaMS
yang rnengandung 2mM NH4N03 15g/1. Dernikian jugst b b t kering dm antosianin
ymg dihasi I kan, Di samping iru penghi langm fosfor clari media meningkakan
Sebagai zat tambahan beberapa vitamin yang sering digunakan &lam kultur
jaringan adalah tiamin (vitamin B1), asam nikotinat (niasin) dm piridoksin (Bs).
Vitamin-vitamin tersebut irerhgsi sebagai kaenzim dalam mehbolisme karbokidrat
d m prutein, Menurut Lehninger (1975) tiamin menrpakm koenzim dalam proses
transfer kelornpuk aldehida, sedangkan niasin rnerupakm kaenzim dalam proses
transfer elektron atau sttorn hidragen, dm piridoksin mempakm koenzim dalam
proses transfer kelornpak amino.
Kecudi ,susunm medium buatm, pemilihn pengunam medium padat atau
a i r juga rnexzentukan berhasil ridaknya suatu kultur tumbukan, Pada umumnya
pemilihan medium padat atau cair diiakukan sesuai dengan kebutuhan dm tujuan
penelitim serta fmilitas yang tersedia. Pada kultw &ar rambut m m y a
menggunakm medium cair karem laju perkmbuhm kuItur lebih cepat. Menurut
Gunawan (1992) pertimbuhan ymg lebih: cepat pada medium cair kernungkinm
disebabkan aleh p e m u k a n eksplm ymng kontak langsung dengan media lebih luas
dm penyerapan nutrient Iebih efektif,
Faktur lain yang berpengaruh pada perEumbuhan kulhx akw rambut addah
lingkungm tumbuh, dimtumyil suhu, kelembaban u h ,
pH
dm cafraya,Lingkungan tluxlbuh yang optimum untuk pertumbuhm kultur bervariasi ctntara
spesies. Toivonen el a/. (1992) memperoleh pertumbuhan d a r rambut Cutharanrhus
roseus paling cepat apabila dikdturkan pada suhu 32°C dib&dingkan pada suhu
Selain itu Ikenaga et a!. (1995) melaporkan bahwa prtumbuhan dm praduksi
dikulturkan d a l h kondisi terany. Steroidal saponin tidak diperoleh pada akar rmbut
yang dikulturkan dalam kandisi ge1ap.
2,s Zat Pengatur Tumbuh
Zat pengatur tumbuh merupakan persenyawam organik selain dari nutrien
yang dalam jumlafi sedikit (ImM) dapat merangsang, menghrunbat abu rnengub*
pola prtumbukm d m perkernbangan tanaman (Gunawm 1992). Zat pengatur
tumbuh terdapat secara alami di dstlam t m a n (endogen), Selain zat pengatur
t u b & alami juga ada zat pengatur tumbuh sintetik (eksogen) yang telah dijual
secara komersil.
Pada saat ini d i k e d enam icelompok zat pengatur twnbuh yaitu auksin,
giberelin, sitokinin, asam absis f k (ABA), etilen dan retardan. Seny awa-senyawa
palimin @utresin, spermidin, spermin), pulifenotilc, dm alkohol bermtai pmjang
(Triakontanof) sering diguiongkan ice dalam zat pengatur tumbuh (Wattimena 1992).
Di dalm kultur jaringan dua galangan zat pngatur tumbufi yang sangat penting
adalah auksin d m sitokinin. Zat p:ngatur tumbuh ini mempengaruhi perturnbufian
dan morfogenesis sel, jmingan dm organ tanaman,
Auksin digunakm secara has ddam kultur jaringan tamman untulc
merangsang pertumbuhan kdus, suspensi sel dan pertumbuh ak:w atau organ
lainnya. Tindakm auksin yang paling karkteristik adalah rneningkatkan p m b e l k
sel dm mendorung ~"rpanjangan sel (Wilkins 1992).
Pengaruh auksin terhadap perturnbuhan jaringm tanaman diduga kaxena
menyebabkan susunan matrik dinding sel rnerenggang dan ion K' mas& ke dalam sel
sehingga rnengurangi potcnsial air dalam sel
.
Keadaan tersebut menj ibabkan airmasuk ke dalm sel
dan
sel menjadi rnernbesar. Pengaruh lain dari auksin terhadaptrtnman juga diduga h e m dapat m e m g e n g d i metabulime RNA yang b e m i
metahlisme protein. Hal tersebut kemungkinm meldui traylskripsi molekul RldA
(Abidin
X
990 ; Gunawan 1992 ; George dan Sherringtan 1984).Auksin alamiah pada tanaman adalafx a m indol asetat (TAA). TAA disintesis
dwi triptofan dalam primordia daun, dstun muda dm biji yang sedang berkecmbah.
Transhkasi di ddam tanaman terjadi rnelalui flaern. Sedmgkan auksin sintetik, selain
IAA juga ada asam indol butirat dm herbisida yslng bersifat auksin yaitu asam
2.4-diklorofenoksi asetat (2.4-D), asam 2.4.5 -trikloro asetat (2.4.5-T), asam ndhlin
asetat (NAA), Dikamba, Pikloram dll.
Pada &ar rmbue h i 1 transfomasi genetik bakteri A. rhizogems terdapctt gen
untuk menyandi sintesis auksin endogen. Gen yang berasai dari plasmid baktei
tersebut addah iaaM dan iau1-I. Ekspresi dari gen-gen tersebut &an mensintesis
auksin alamiah, yaitu IAA.
Di samping auksin, sitokinin juga penting dalam pengatman p m b e l h sel
dm morfugenesis. Sitokinirr adztiah kelampok senyawa orgmik ymg menyebabh
pembelahm sel yang dikenal dengan proscs sitokinesis. Pemklahan. xl,;tosis tid&
&an terjadi tanpa sitakinin (Wattirnena 1992).
Sitokinin y m g pextama &Xi ditemukm adalah kinetin (6-furfuril mino purin)
yang diisolasi dari DNA ikan Herring (Gunstwan X992), Sitokinin yang terdapat
adenin dm mtai samping add&* isopenti1 yang berasal
dari
asam mevdonat.Biusintesis zeatin terntamst diujung &ar dm ddm biji yang sedang berkecambafi.
Translokasi zeatin terutama meldui xilem (WatEimena 1992). Sitalcinin sintetik terdiri
dari -in, bemil adenin purin (BAP), 2-iP, Kinetin dll.
Sama seerti auksin, gen untuk rnenyandikn sirntesis situkirrirr juga terdapat:
di ddam sel akar rambut. Gen tersebut adalah gen ipr yang juga berasal dmi
&ansfomasi b&teri A. rhizogenes. Oleh karena a h y a gen-gen tersebut, zat
pengatur hunbuh eksogen jarmg digunakm untuk menginduksi akar rambut.
Biasanya di dalam kultur akar rambut, penambahan zat penngatur tumbuh ehagen
bertujuan untuk regenemi aka rambut menjadi m a s atau untuk; pembcntukan kalus.
Sedangkan kultur &a rambut mtuk rnemproduksi rnetabolit sekunder tidak selalu
membutuhkan zat pengatur tumbuh eksogen. Bimanya penmbthn zat pengatur
tumbuh eksogen diperlukm untuk meningkatkan laju pertmbuhan aka
tr
produicsimetabulit sekunder y m g dihasilkm, &an tetapi tergantung pada spesies tanaman d m
jenis zat pengatur tumbuh.
Pengaruh xat pngatur hunbuh eksogen terfradap pertum5uhan dm
pembentukan metabulit sekunder dalam akar r~~llbue sangat beragam antara spesies.
Pada aka rambut Solmum ucatleatissinaum, penmbahan NAA dan EAA 100 pglI
pada medium kultwr dapat rneningkatkan praduksi steroidal saponin walaupun tidak
ternyata naenghambat perturnbuhan &ar d m produlcsi steroidal sapanin (Ikenaga
antara 0,Ul sampai
1,U
mg/l pada medium kultur temyata iidak berpengaruh padaperturnbuhan akar rambut ledgeriana (Hamill el ul. 1989).
2.8 Polinrerase Chain Reucffon PCR)
Pa& akar rambut terjadi penyisi@inte@sasi DNA
dari
A. rhizage~les padagenom sel tanaman. Penyisipm ini akan merub& susunsur nuIclwtidEt DNA seI
tanaman dan rndrubah ekspresi gen dari sel tanaman untuk mernbentuk akar rambut.
Integrasi DNA tersebut dapat dideteksi secara PCR. Menurut Dale (1994) dm
Geerlings et al+ (1999) metoda ymg lebih cepat dm &curat yang banyak dila3cukrtn
saat ini untuk menguji keberhasilan irztegrasi DNA bakteri pada sel tanaman adalah
dengan teIrnik PCR,
PCR merupakan
proses sintesis enzimatis untlrlc mengmplikasi unrtmnukleotida DNA secara in vifro ( h d g r a f dan Wolfes 1993). Amplifikasi DNA
dengan PCR dapat meningkatkm jumlah urutan DNA secaa cepat dm bisa lebih d a i
jutam kopi. Aplikasi PCR antara lain untulc rnendeteksi adanya DNA spesifik,
adisis DNA finger printing dengm rnetode U P D (Random Amplified Polirnorpkic
DNA) dm untuk mendeteksi terjadinya rnutasi (Thomas 1996).
P d a prinsipnya ada tiga proses ymg terjadi ddam reaksi PCR. Proses
pertma adalah denaturasi, mempaketn proses pemisahm rank ganda DNA rnenjadi
utas tunggal. Proses ini memerlukan suhu y m g tinggi, yaitu b l w y a antara
94'C
-
97T. Proses ymg kedua a&l& annealing, merrrpakm proses penempelanprimer sebagai pernula untuk sintesis DNA. Pada proses annealing rnemerlukan suhu
oleh psnjang, banyak G dan C dalam primer dm konsentrasi garam lamtan bufer.
Proses y m g terakhir adalah extention, rnentpakm proses perpanjangm primer. Suhu
extention tergantung pada pmjmg dm konsentrasi dari susunan DNA target.
Urnumnya perganjangm primer tedabi pada suhu 72°C karena pada suhu ini enzirn
Taq palimeruse bekerja optimal untuk. sintesis DNA (Xnnis dm Gel f a d 1990).