GAMMA OLEH KAPANG Penicillium sp. DAN Trichoderma sp.
YELVI ERIDA
PROGRAM STUDI BIOLOGI
FAKULTAS SAINS DAN TEKNOLOGI
UNIVERSITAS ISLAM NEGERI SYARI HIDAYATULLAH
JAKARTA
GAMMA OLEH KAPANG Penicillium sp. DAN Trichoderma sp
.
SKRIPSI
Sebagai Salah Satu Syarat untuk Memperoleh Gelar Sarjana Sains Pada Program Studi Biologi Fakultas Sains dan Teknologi
Universitas Islam Negeri Syarif Hidayatullah Jakarta
YELVI ERIDA
106095003217
PROGRAM STUDI BIOLOGI
FAKULTAS SAINS DAN TEKNOLOGI
UNIVERSITAS ISLAM NEGERI SYARI HIDAYATULLAH
JAKARTA
yang diciptakan Allah di langit dan di bumi, benar-benar terdapat
tanda-tanda (kekuasaan-Nya) bagi orang-orang yang bertakwa” (QS. Yunus: 6)
Pengetahuan diperlikan karena ia adalah akar, tempat bertaut cabang.
Setiap sayap tak akan mampu terbang melintasi kekuasaan samudra.
Hanya ilmu yang terlimpah lansung dari Allah dapat mengantarkan
seseorang dari haribaan-Nya (Anonim)
Skripsi ini dipersembahkan
SEBAGAI SKRIPSI ATAU KARYA ILMIAH PADA PERGURUAN TINGGI ATAU LEMBAGA MANAPUN.
Jakarta, Desember 2010
Biosolubilisasi adalah teknologi yang memiliki potensi untuk mengubah padatan batubara menjadi bahan bakar cair atau kimia. Tujuan dari penelitian ini adalah untuk mengkarakterisasi enzim ekstraseluler dan mengetahui produk biosolubilisasi batubara hasil iradiasi gamma oleh kapang Penicillium sp. dan
Trichoderma sp. Dosis iradiasi gamma yang digunakan pada batubara adalah 5 kGy dan batubara yang tidak diiradiasi. Parameter yang diukur adalah uji kualitatif enzim ekstraseluler kapang (fenoloksidase, lignin peroksidase, dan Mn-peroksidase), pH medium, nilai absorbansi hasil biosolubilisasi batubara (λ250 nm
and λ450 nm), pengukuran hidrolisis FDA, pengukuran kadar protein ekstraseluler
kapang dengan metode Lowry, analisis senyawa hasil biosolubilisasi batubara dengan GC-MS dan karakteristik enzim ekstraseluler dengan SDS-PAGE. Hasil penelitian menunjukkan bahwa kedua kapang memiliki ketiga jenis enzim dan iradiasi gamma pada batubara (5 kGy) tidak mempengaruhi pH medium, biosolubilisasi, hidrolisis FDA dan kadar protein ekstraseluler kecuali pada karakteristik enzim ekstraseluler kapang Trichoderma sp. Enzim yang terdeteksi pada kapang Penicillium sp. adalah mangan peroksidase (BM=48 KDa) dan lakase (BM=56 KDa) baik pada batubara yang tidak diiradiasi (0 kGy) dan batubara yang diiradiasi (5 kGy), sedangkan pada kapang Trichoderma sp. terdeteksi hanya lakase (BM=56 KDa) pada batubara yang diiradiasi (5 kGy) dan mangan peroksidase (BM=48 KDa) dan lakase (BM=56 KDa) pada batubara yang tidak diiradiasi (0 kGy). Produk solubilisasi menunjukkan kecenderungan setara bensin pada perlakuan batubara yang diiradiasi dan tidak diiradiasi untuk
Trichoderma sp., sedangkan Penicillium sp. hanya pada batubara yang tidak diiradiasi.
Biosolubilization is a technology that has a potential for converting solid coal into liquid fuel or chemical. The objectives of this research were to characterize extracellular enzyme and to know biosolubillization products of gamma irradiation coal by Penicillium sp. and Trichoderma sp. The dosages which has been used for gamma irradiation were 5 kGy and unirradiated coal. The parameters were the qualitative test of extracellular enzyme enzyme (phenoloxidase, peroxidase, and Mn-peroxidase), pH of medium, absorbance of the coal biosulubilization (λ250 nm and λ450 nm), hydrolysis of FDA, protein
concentration by Lowry method, analysis of coal biosolubilization product by GC-MS, and characteristics of the extracellular enzyme by SDS-PAGE. The result showed both of fungi were positive for all enzymes test and the gamma irradiated did not effect on the pH medium, biosolubilization, FDA hydrolysis and extracellular protein concentration but effected on the characteristic of extracellular enzyme from Trichoderma sp. The enzymes detected from
Penicillium sp. were manganese-peroxidase (MW = 48 KDa) and laccase (MW = 56 KDa) in both irradiated coal (5 kGy) and unirradiated coal (0 kGy), whereas on
Trichoderma sp. was detected only laccase (MW = 56 KDa) in irradiated coal (5 kGy), and manganese-peroxidase (MW = 48 KDa) and laccase (MW = 56 KDa) in unirradiated coal (0 kGy). The biosolubilization product showed the similarity with gasoline in irradiated and unirradiated coal for Trichoderma sp., but for
Penicillium sp. only occurred on unirradiated coal .
dengan yang diharapkan, dalam skripsi ini penulis mengambil judul tentang ”Karakterisasi Enzim Ekstraseluler dan Produk Biosolubilisasi Batubara Hasil Iradiasi Gamma oleh Kapang Penicillium sp. dan Trichoderma sp.” sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains pada Program Studi Biologi Universitas Islam Negeri Syarif Hidayatullah Jakarta.
Shalawat dan salam senantiasa tercurahkan kepada junjungan Nabi Muhammad SAW beserta para sahabat, keluarga, dan pengikut-pengikut beliau hingga hari akhir.
Pada kesempatan ini penulis tidak lupa mengucapkan terima kasih yang sebesar-besarnya atas bimbingan dan saran-saran serta pentunjuk kepada :
1. `Dr. Sopiansyah Jaya Putra, M.Sis, selaku Dekan Fakultas Sains dan Teknologi Universitas Islam Negeri Syarif Hidayatullah Jakarta.
2. Dr. Lily Surraya Eka Putri, M.Env.Stud selaku Ketua Program Studi Biologi Fakultas sains dan Teknologi.
3. Irawan Sugoro, M.Si dan Megga Ratnasari Pikoli, M.Si selaku Pembimbing I dan Pembimbing II, yang dengan sabar memberikan petunjuk serta bimbingan, sehingga penulis dapat menyelesaikan penelitian dan penyusunan skripsi ini.
4. Dra. Nani Radiastuti, M.Si dan La Ode Sumarlin, M.Si, selaku Penguji I dan II pada seminar proposal dan seminar hasil yang telah memberikan pengarahan dalam penyusunan skripsi ini.
5. Dr. Lily Surraya Eka Putri, M.Env.Stud dan Dasumiati, M.Si, selaku Penguji I dan II pada sidang Munagosyah yang telah memberikan pengarahan dalam penyusunan skripsi ini.
6. Kedua orang tua tercinta, Mama dan Papa yang selalu hidup di hati penulis sampai kapanpun, yang telah memberikan kasih sayang, do’a, dan pengorbanan yang tak terkira demi kehidupan dan masa depan penulis.
titi, Nte ja, Ni on, Om yos, Om pen, ka pup dan Uncu) yang selalu memberi motivasi, kasih sayang, doa maupun materi pada penulis.
9. Keluarga di Citayam (Nte fifi, Om rid, Om al dan Nte adek) yang selalu memberikan motivasi, doa, perhatian dan kasih sayang kepada penulis. 10.Mitha, Riska, Dede, Astri dan Rian yang menemani penulis dalam
melaksanakan Penelitian di Batan (makasih atas kerjasamanya plend). 11.Biologi Angkatan 2006 khususnya ayang-ayangan (Rinae, Ise, Noe yang
selalu menemeni penulis) dan teman-teman seperjuanganku, Nene, Anggi, Lidi, Mpit, Nungky, Nita, Zian, Hera, Eko, Ipin, Muhe, Deden, Malik, Iqbal, Adeng, dan Geleng) tetap kompak sampai kapanpun.
12.Teman masa kecil penulis ( Ami, Yelin dan Ci) atas doa dan perhatiannya. 13.Pihak-pihak lain yang tidak bisa saya sebutkan satu persatu, tapi saya
ucapkan sekali lagi banyak terima kasih karena dengan bantuan semuanya segala masalah dapat terselesaikan dengan lebih mudah dan mendapatkan hasil yang baik.
Harapan penulis semoga skripsi ini dapat bermanfaat bagi penulis khususnya serta para pembaca umumnya dalam melengkapi ilmu pengetahuan yang berhubungan dengan ”Karakterisasi Enzim Ekstraseluler dan Produk Biosolubilisasi Batubara Hasil Iradiasi Gamma oleh Kapang Penicillium sp.
dan Trichoderma sp.” Akhir kata hanya kepada Allah SWT dipanjatkan do’a
untuk membalas segala budi baik untuk semua pihak yang terkait.
Jakarta, 14 November 2010
Yelvi Erida
KATA PENGANTAR ... i
2.3 Kapang Pengsolubilisasi Batubara... 10
2.3.1 Kapang Trichoderma sp. ... 12
2.3.2 Kapang Penicillium sp. .... ... 14
2.4 Enzim ... 15
2.5 Kromatografi Gas Spektroskopi Massa (GCMS) ... 20
2.6 Elektroforesis ... 21
2.7 Iradiasi Gamma ... 23
BAB III METODOLOGI PENELITIAN 3.1 Tempat dan Waktu Penelitian ... 24
3.3.2.1 Medium Potato Dextrose Agar (PDA) .... 25
3.3.2.2 Medium Minimal Salt (MMS) .... ... 26
3.3.2.3 Medium Potato Dextrose Agar Minimal Salt (PDAM) .... ... 26
3.3.2.4 Medium Minimal Salt + Sukrosa (MMSS) ... 26
3.3.7 Pengujian Biosolubilisasi Batubara ... 28
3.3.7.1 Pengukuran Biosolubilisasi Batubara .... 29
3.3.8 Pengukuran Hidrolisis FDA ... 30
3.3.9 Pengukuran Kadar Protein Ekstraseluler dengan Metode Lowry ... 30
3.3.10 Analisis Hasil Biosolubilisasi Batubara oleh Kapang Penicilium sp. dan Trichoderma sp. Dengan Menggunakan GC-MS ... 31
3.3.11 Karakteristik Enzim Ekstraseluler dengan Elektroforesis ... 31
3.4 Analisis Data ... 33
BAB IV HASIL DAN PEMBAHASAN 4.1 Hasil Analisis Kualitatif Enzim Ekstraseluler Kapang ... 34
4.2 Hasil Pengukuran pH Medium Kapang ... 35
4.3 Hasil Biosolubilisasi Batubara ... 38
4.7 Karakteristik Enzim Ekstraseluler Kapang Penicillium sp.
dan Trichoderma sp. ... 53
BAB V KESIMPULAN DAN SARAN 5.1 Kesimpulan ... 56
5.2 Saran... 56
DAFTAR PUSTAKA... 57
LAMPIRAN... 61
.
Gambar 2. Kapang Trichoderma sp. ... 12 Gambar 3. Kapang Penicillium sp. . ... 15 Gambar 4. Uji kualitatif adanya enzim ekstraseluler kapang Penicillium sp.
dan Trichoderma sp. ... 34 Gambar 5. Nilai pH medium kapang Penicillium sp. dan Trichoderma sp. ... 37 Gambar 6. Nilai absorbansi supernatan pada panjang gelombang 250 nm
hasil biosolubilisasi kapang Penicillium sp. (A) dan
Trichoderma sp. (B) ... 39 Gambar 7.Nilai absorbansi supernatan pada panjang gelombang 450 nm
hasil biosolubilisasi kapang Penicillium sp. (A) dan
Trichoderma sp (B).. ... 41 Gambar 8. Nilai absorbansi pada panjang gelombang 490 nm hasil aktivitas
enzim kapang Penicillium sp. (A) dan Trichoderma sp. (B).. . ... 45 Gambar 9. Nilai kadar protein ekstraseluler kapang Penicillium sp.(A) dan
Trichoderma sp. (B). ... 47 Gambar 10.Karakteristik enzim ekstraseluler kapang Penicillium sp. dan
Trichoderma sp. pada medium yang mengandung batubara yang tidak diiradiasi (0 kGy) dan batubara yang diiradiasi (5 kGy)... 54
pelapuk putih (White Rot Fungi) ... 18
Tabel 2. Komposisi medium perlakuan ... 25
Tabel 3. Data perlakuan kultur kapang ... 29
Tabel 4. Senyawa hasil biosolubilisasi batubara menggunakan GC-MS... 51
viii
Lampiran 2. Bagan kerja ... 62
Lampiran 3. Komposisi medium ... 63
Lampiran 4. Komposisi reagen Lowry ... 63
Lampiran 5. Komposi larutan elektroforesis ... 64
Lampiran 6. Hasil pengujian pH medium kapang ... 65
Lampiran 7. Hasil pengujian biosolubilisasi batubara... 65
Lampiran 8. Hasil pengujian hidrolisis FDA... 65
Lampiran 9. Hasil pengujian kadar protein ekstraseluler kapang... 66
Lampiran 10. Kurva standar protein BSA ... 66
Lampiran 11. Kurva standar marker... 67
Lampiran 12. Uji statistik analisis varians... 68
Lampiran 13. Kromatogram hasil GC-MS kontrol ... 70
Lampiran 14. Kromatogram hasil GC-MS biolubilisasi batubara oleh kapang Penicillium sp. ... 71
Lampiran 15. Kromatogram hasil GC-MS biosolubilisasi batubara oleh kapang Trichoderma sp. ... 72
1.1. Latar Belakang
Kebutuhan bahan bakar minyak mengalami peningkatan dari tahun ke tahun, sedangkan cadangan minyak mentah diperkirakan akan habis dalam kurun waktu 20 tahun. Sumber energi yang akan memegang peranan yang penting adalah batubara. Indonesia memiliki cadangan batubara sekitar 36,3 juta ton dan 49 % dari cadangan tersebut masih dalam bentuk batubara peringkat rendah. Penggunaan batubara terutama untuk kebutuhan energi listrik, tetapi Indonesia tidak mungkin membakar habis batubara dalam bentuk PLTU karena akan mengotori lingkungan dengan dihasilkannya polutan seperti sulfur oksida (SOx), nitrogen oksida (NOx), karbondioksida (CO2) dan logam berat. Salah satu solusi
untuk mengatasi masalah kelangkaan bahan bakar minyak adalah dengan mengolah batubara sehingga menjadi energi bersih seperti melalui proses pencairan batubara (Novia et al., 2009).
Biosolubilisasi batubara adalah suatu upaya untuk mencairkan batubara yang nantinya dapat digunakan sebagai bahan bakar pengganti minyak bumi. Pada umumnya metode yang digunakan dalam proses pencairan batubara adalah dengan metode kimia dan fisika. Kelemahan metode kimia dan fisika yaitu membutuhkan biaya operasional yang cukup tinggi karena prosesnya dilakukan dalam temperatur dan tekanan yang tinggi, memerlukan instalasi yang cukup rumit dan menghasilkan CO2 dua kali lipat dari bahan bakar minyak biasa.
Alternatif lainnya dengan metode biologi yaitu dengan bantuan mikroorganisme. Pencairan batubara dengan metode biologi relatif dapat menekan biaya operasional karena tidak dilakukan dalam tekanan dan temperatur yang tinggi serta lebih ramah lingkungan karena tidak menghasilkan produk sampingan yang berbahaya (Scott et al., 1991). Hasil biosolubilisasi batubara akan menghasilkan senyawa yang setara dengan autput minyak bumi (Cohen et al., 1990).
Mikrooganisme dapat diterapkan dalam proses biosolubilisasi batubara (Laborda et al., 1999). Menurut Cohen dan Gabriele (1982) mikroorganisme menggunakan material batubara sebagai sumber energi untuk pertumbuhan dan kemampuannya dalam mensolubilisasi batubara. Mikroorganisme yang digunakan dalam penelitian ini adalah kapang Penicillium sp. dan Trichoderma sp. Penelitian Sugoro et al. (2009) membuktikan bahwa kapang Penicillium sp. dan
Trichoderma sp. memiliki nilai absorbansi tertinggi, artinya kapang tersebut telah melakukan pendegradasian batubara tertinggi dan mampu tumbuh menggunakan medium batubara. Pembuktian ini diperkuat oleh penelitian Laborda et al. (1999) membuktikan bahwa kapang Penicillium sp. dan Trichoderma sp. terlibat dalam proses biosolubilisasi batubara. Biosolubilisasi batubara oleh kapang Penicillium
sp. dan Trichoderma sp. tumbuh dengan adanya batubara.
dan lakase, masing-masing mempunyai berat molekul 38-47 KDa, 38-50 KDa, dan 53-110 KDa (Fakuosa dan Hofrichter, 1999).
Biosolubilisasi dapat ditingkatkan dengan cara praperlakuan terhadap batubara. Batubara diiradiasi dengan iradiasi gamma. Pemakaian dosis dalam iradiasi gamma mempengaruhi proses biosolubilisasi batubara karena pada iradiasi terjadi proses oksidasi yang dapat menghasilkan senyawa asam. Penelitian ini menggunakan batubara yang tidak diiradiasi (0 kGy) dan batubara yang diiradiasi (5 kGy) (Selvi, 2009). Pemakaian dosis iradiasi gamma bertujuan untuk mengubah senyawa kompleks pada batubara menjadi senyawa sederhana sehingga hasil biosolubilisasi batubara menjadi lebih sempurna. Selain itu, iradiasi gamma dapat meningkatkan site adsorpsi pada batubara sehingga enzim atau sel dapat bekerja maksimum (Sugoro, 2009).
Berdasarkan uraian di atas, tujuan dari penelitian ini adalah untuk mengetahui pengaruh batubara hasil iradiasi gamma terhadap karakteristik enzim ekstraseluler dari kultur kapang Penicillium sp. dan Trichoderma sp. Hasil penelitian ini diharapkan dapat memberikan informasi tentang enzim ekstraseluler yang mampu mensolubilisasi batubara.
1.2. Perumusan Masalah
2. Apakah ada perbedaan karakteristik enzim ekstraseluler kultur kapang
Penicillium sp. dan Trichoderma sp. dalam medium yang mengandung batubara hasil iradiasi gamma?
3. Apakah ada perbedaan produk biosolubilisasi batubara oleh Penicillium
sp. dan Trichoderma sp. dalam medium yang mengandung batubara hasil iradiasi gamma?
1.3. Hipotesis
1. Iradiasi gamma pada batubara berpengaruh terhadap kadar protein ekstraseluler kapang Penicillium sp. dan Trichoderma sp.
2. Karakteristik enzim ekstraseluler kultur kapang Penicillium sp. dan
Trichoderma sp. dalam medium yang mengandung batubara hasil iradiasi gamma berbeda.
3. Produk biosolubilisasi batubara oleh Penicillium sp. dan Trichoderma sp. dalam medium yang mengandung batubara hasil iradiasi gamma berbeda.
1.4. Tujuan Penelitian
1. Mengetahui pengaruh iradiasi gamma pada batubara terhadap kadar protein ekstraseluler kultur kapang Penicillium sp. dan Trichoderma sp. 2. Mengetahui karakteristik enzim ekstraseluler kultur kapang Penicillium
3. Mengetahui perbedaan produk biosolubilisasi batubara oleh Penicillium
sp. dan Trichoderma sp. dalam medium yang mengandung batubara hasil iradiasi gamma.
1.5. Manfaat Penelitian
Karakteristik enzim ekstraseluler kultur kapang Penicillium sp. dan
2.1. Batubara
Batubara adalah mineral organik yang dapat terbakar, terbentuk dari sisa
tumbuhan purba yang mengendap yang selanjutnya berubah bentuk akibat proses
fisika dan kimia yang berlangsung jutaan tahun. Oleh karena itu, batubara
termasuk dalam kategori bahan bakar fosil, proses yang mengubah tumbuhan
menjadi batubara tadi disebut dengan pembatubaraan. Batubara adalah batuan
yang mudah terbakar yang lebih dari 50%-70% berat volumenya merupakan
bahan organik. Bahan organik utamanya yaitu tumbuhan yang dapat berupa kulit
pohon, daun, akar, spora, polen, dan lain-lain. Selanjutnya bahan organik tersebut
mengalami berbagai tingkat pembusukan (dekomposisi) sehingga menyebabkan
perubahan sifat-sifat fisik maupun kimia baik sebelum ataupun sesudah tertutup
oleh endapan lainnya (Speight, 1994).
2.1.1. Penyusun Batubara
Menurut Speight (1994) batubara berasal dari sisa tumbuhan diperkuat
dengan ditemukannya fosil tumbuhan di dalam lapisan batubara. Dalam
penyusunannya batubara diperkaya dengan berbagai macam polimer organik yang
berasal dari antara lain karbohidrat, lignin, dan protein.
Lignin merupakan suatu unsur yang memegang peranan penting dalam
mengubah susunan sisa tumbuhan menjadi batubara. Sementara ini susunan
molekul umum dari lignin belum diketahui dengan pasti, namun susunannya dapat
diketahui dari lignin yang terdapat pada berbagai macam jenis tanaman. Sebagai
contoh lignin yang terdapat pada rumput (Hatakka, 2001).
Karbohidrat terdiri dari disakarida, trisakarida, ataupun polisakarida. Jenis
polisakarida inilah yang umumnya menyusun batubara, karena dalam tumbuhan
jenis inilah yang paling banyak mengandung polisakarida (khususnya selulosa)
yang kemudian terurai dan membentuk batubara (Speight, 1994)
Protein merupakan bahan organik yang mengandung nitrogen yang selalu
hadir sebagai protoplasma dalam sel mahluk hidup. Struktur dari protein pada
umumnya adalah rantai asam amino yang dihubungkan oleh rantai amida. Protein
pada tumbuhan umunya muncul sebagai steroid (Laborda et al.,1999).
2.1.2. Klasifikasi Batubara
Batubara diklasifikasikan berdasarkan tingkat pembatubaraan dan tujuan
pemanfaatannya. Misalnya, batubara bituminus banyak digunakan untuk bahan
bakar pembangkit listrik, pada industri baja atau genteng serta industri semen.
Tipe batubara berdasarkan tingkat pembatubaraan ini dapat dikelompokkan yaitu
A. B.
C. D.
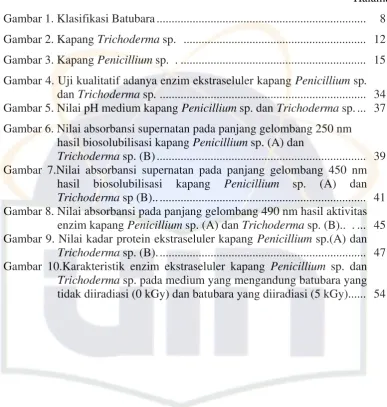
Gambar 1. Klasifikasi batubara (American Coal Foundation, 2007) (A. lignit; B. subbituminus; C. bituminus; dan C. antrasit)
Lignit merupakan batubara muda dengan kualitas terendah. Lignit adalah
batubara yang sangat lunak dan mengandung air 70% dari beratnya. Batubara ini
berwarna hitam, sangat rapuh, nilai kalor rendah dengan kandungan karbon yang
sangat sedikit, kandungan abu dan sulfur yang banyak (Gambar 1 A). Batubara
jenis ini dijual secara eksklusif sebagai bahan bakar untuk pembangkit listrik
tenaga uap (PLTU).
Subbituminus merupakan batubara yang berada di antara batubara lignit
subbituminus mengandung sedikit karbon dan banyak air, oleh karena itu menjadi
sumber panas yang tidak efisien (Gambar 1 B).
Bituminus adalah batubara yang tebal, biasanya berwarna hitam
mengkilat, terkadang coklat tua (Gambar 1 C). Batubara bituminus mengandung
86% karbon dari beratnya dengan kandungan abu dan sulfur yang sedikit.
Umumnya dipakai untuk PLTU, tapi dalam jumlah besar juga dipakai untuk
pemanas dan aplikasi sumber tenaga dalam industri.
Antrasit adalah batubara peringkat teratas, biasanya dipakai untuk bahan
pemanas ruangan di rumah dan perkantoran. Batubara antrasit berbentuk padat,
keras dengan warna hitam mengkilat dan mengandung antara 86% – 98% karbon
dari beratnya, terbakar lambat, dengan nyala api berwarna biru dengan sedikit
sekali asap (Gambar 1 D).
2.2. Biosolubilisasi Batubara
Biosolubilisasi adalah proses pelarutan dalam suatu medium dengan bantuan
mikroorganisme. Biosolubilisasi dapat berupa upaya untuk mencairkan batubara
yang nantinya dapat digunakan sebagai bahan bakar pengganti minyak bumi. Di
samping untuk mencairkan batubara, biosolubilisasi dapat pula digunakan untuk
mengurangi kandungan sulfur atau logam toksik pada batubara (Faison et al.,
1989).
Batubara cair yang dihasilkan dari proses biosolubilisasi adalah berupa
campuran senyawa yang larut dalam air, senyawa-senyawa polar dengan berat
molekul relatif lebih tinggi. Teknologi ini memiliki potensi besar, tetapi masih ada
cocok, produk yang dihasilkan tetap padat. Produk terlarut ini memiliki
kandungan energi tinggi dan memungkinkan digunakan sebagai bahan bakar,
tetapi belum dapat digunakan sebagi bahan bakar sarana transportasi. Selain itu,
kebanyakan mikroorganisme membutuhkan gula dan media pertumbuhan lebih
dari dua minggu. Media murah dan mampu mempercepat pertumbuhan
mikroorganisme untuk aplikasi komersial. Produksi batubara cair dapat dilakukan
dengan memanfaatkan enzim hasil isolasi mikroorganisme (Liu et at., 1989).
2.3. Kapang Pengsolubilisasi Batubara
Kapang (mould/filamentous fungi) merupakan mikroorganisme anggota
Kingdom Fungi yang membentuk hifa (Carlile dan Watkinson 1994). Kapang
bukan merupakan kelompok taksonomi yang resmi, sehingga anggota-anggota
dari kapang tersebar ke dalam filum Glomeromycota, Ascomycota, dan
Basidiomycota (Hibbett et al., 2007).
Sifat-sifat fisiologi kapang dipengaruhi oleh kebutuhan air, suhu
pertumbuhan, kebutuhan oksigen dan pH, makanan, dan komponen penghambat.
Pada umumnya kebanyakan kapang membutuhkan aw minimal untuk
pertumbuhan lebih rendah. Kapang bersifat mesofilik, yaitu tumbuh baik pada
suhu kamar. Suhu optimum pertumbuhan kapang adalah sekitar 25-30°C, tetapi
ada beberapa dapat tumbuh pada suhu 35-37°C atau lebih tinggi, misalnya
Aspergillus. Semua kapang bersifat aerofilik, yaitu membutuhkan oksigen untuk
pertumbuhannya. Kebanyakan kapang dapat tumbuh pada kisaran pH yang luas,
atau pH rendah. Kapang dapat menggunakan berbagai komponen makanan dari
yang sederhana sampai kompleks. Kapang memproduksi enzim hidrolitik,
misalnya amilase, pektinase, proteinase, dan lipase. Kapang mengeluarkan
komponen yang dapat menghambat organisme lainnya. Komponen ini disebut
antibiotik. Pertumbuhan kapang biasanya berjalan lambat bila dibandingkan
pertumbuhan bakteri dan khamir (Fardiaz, 1989).
Habitat kapang sangat beragam, namun pada umumnya kapang dapat
tumbuh pada substrat yang mengandung sumber karbon organik (Carlile dan
Watkinson 1994). Kapang yang tumbuh dan mengkolonisasi bagian-bagian di
dalam ruangan telah banyak diteliti. Kapang tersebut mudah dijumpai pada
bagian-bagian ruangan yang lembab, seperti langit-langit bekas bocor, dinding
yang dirembesi air, atau pada perabotan lembab yang jarang terkena sinar
matahari. Genus kapang yang sering dijumpai tumbuh di dalam ruangan adalah
Cladosporium, Penicillium, Alternaria, dan Aspergillus (Mazur et al., 2006).
Kapang melakukan reproduksi dan penyebaran menggunakan spora. Spora
kapang terdiri dari dua jenis, yaitu spora seksual dan spora aseksual (Carlile dan
Watkinson 1994). Menurut Champe et al. (1981) dan Carlile dan Watkinson
(1994), spora aseksual dihasilkan lebih cepat dan dalam jumlah yang lebih banyak
dibandingkan spora seksual. Spora aseksual memiliki ukuran yang kecil (diameter
1-10 μm) dan ringan, sehingga penyebarannya umumnya secara pasif
menggunakan aliran udara.
Sejumlah strain jamur dan bakteri berfilamen diketahui berinteraksi
medium yang lebih gelap selama proses kultur atau cairan gelap pada permukaan
batubara ketika ditumbuhkan pada permukaan kultur agar. Diketahui bahwa
tedapat beberapa jenis mikroorganisme dari jenis bakteri maupun fungi yang
dapat mengubah batubara padat menjadi produk cair, dengan minimalisasi
hilangnya kandungan energi total awal (Faison et al., 1989).
2.3.1. Kapang Trichoderma sp.
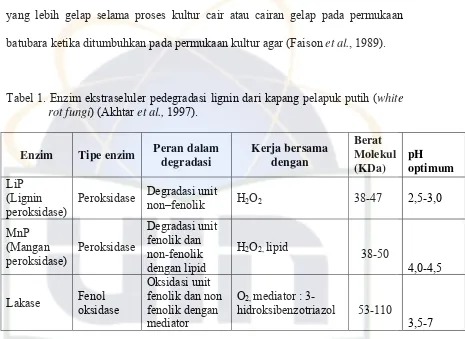
Ciri-ciri spesifik kapang Trichoderma sp. adalah mempunyai konidia,
sterigmata, konidiofora, miselium berseptat (Gambar 2). Koloni kapang
Trichoderma sp. tersebut berumur 7 hari, penampakan koloninya dilihat
menggunakan mikroskop pada perbesaran 400 X (Kuraesin, 2009). Kapang
Trichoderma sp. mempunyai konidiofora bercabang banyak, ujung
percabangannya merupakan sterigma, membentuk konidia bulat atau oval,
berwarna hijau terang, dan berbentuk bola-bola berlendir ( Fardiaz, 1989).
A. Konidia
B. Sterigma
C. Konidiofora
Kapang Trichoderma sp. diklasifikasikan menurut sistem nama binomial
yaitu: Kingdom Fungi; Filum Eumycota; Sub Filum Deuteromycota; Kelas
Hyphomycetes; Ordo Hyphomycetales; Famili Moniliaceae; Genus Trichoderma
dan Spesies Trichoderma sp.Koloni dari kapang Trichoderma sp. berwarna putih,
kuning, hijau muda, dan hijau tua. Susunan sel kapang Trichoderma sp. bersel
banyak berderet membentuk benang halus yang disebut dengan hifa. Hifa pada
jamur ini berbentuk pipih, bersekat, dan bercabang-cabang membentuk anyaman
yang disebut miselium. Miseliumnya dapat tumbuh dengan cepat dan dapat
memproduksi berjuta-juta spora, karena sifatnya inilah Trichoderma sp.dikatakan
memiliki daya kompetitif yang tinggi. Dalam pertumbuhannya, bagian permukaan
akan terlihat putih bersih, dan bermiselium kusam. Setelah dewasa, miselium
memiliki warna hijau kekuningan (Carlile dan Watkinson, 1994).
Trichoderma sp. adalah salah satu jamur tanah yang tersebar luas
(kosmopolitan), yang hampir dapat ditemui di lahan-lahan pertanian dan
perkebunan. Trichoderma sp. bersifat saprofit pada tanah, kayu, dan beberapa
jenis bersifat parasit pada jamur lain. Pada spesies saprofit, kapang tumbuh pada
kisaran suhu optimal 22-30°C. Suhu optimal untuk pertumbuhan kapang ini
adalah 32-35°C dan pH optimal sekitar 4.0. Trichoderma sp. berkembangbiak
secara aseksual dengan membentuk spora di ujung fialida atau cabang dari hifa
(Mazur et al., 2006).
Miselium Trichoderma sp. dapat menghasilkan suatu enzim yang
bermacam-macam, termasuk enzim selulase (pendegradasi selulosa) dan kitinase
tumbuh secara langsung di atas kayu yang terdiri atas selulosa sebagai polimer
dari glukosa. Oleh karena adanya kitinase, Trichoderma sp. dapat bersifat sebagai
parasit bagi jamur yang lainnya. Secara alami seseorang dapat sering menemukan
Trichoderma sp. yang menjadi parasit pada badan buah dan miselia dari jamur
yang lain (Carlile dan Watkinson, 1994).
Penelitian sebelumnya Sugoro et al. (2009) membuktikan bahwa kapang
Trichoderma sp. memiliki nilai absorbansi tertinggi, artinya berarti telah
melakukan pendegradasian batubara tertinggi pada hari ke-2 inkubasi yaitu 1,936.
Kapang Trichoderma sp. mampu tumbuh menggunakan medium batubara dan
memiliki nilai pH medium yang berfluktuasi, artinya telah terjadi proses degradasi
selama proses inkubasi.
2.3.2. Kapang Penicillium sp.
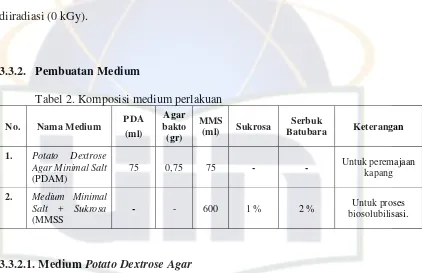
Ciri-ciri spesifik Penicillium sp. (Gambar 3) adalah mempunyai hifa
berseptat, konidia, sterigma , konidiospora (Kuraesin, 2009). Kapang Penicillium
sp. mempunyai hifa berseptat, miselium bercabang, konidiospora septat dan
muncul di atas permukaan, kepala yang membawa spora berbentuk seperti sapu
dengan sterigma muncul dalam berkelompok, dan konidia membentuk rantai
A. Konidia
B. Sterigma
C. Konidiofora
Gambar 3. Kapang Penicillium sp. (Kuraesin, 2009)
Kapang Penicillium sp. diklasifikasikan menurut sistem nama binomial
yaitu: Kingdom Fungi; Filum Ascomycota; Kelas Eurotiomycetes; Ordo
Eurotiales; Famili Trichocomaceae; Genus Penicillium dan Spesies Penicillium
sp. Kapang Penicillium sp. banyak tersebar di alam. Penicillium juga digunakan
dalam industri untuk memproduksi antibiotik, misalnya penisilin yang diproduksi
oleh Penicillium notatum dan Penicillium chrysogenum (Fardiaz, 1989).
Pada hasil penelitian Sugoro et al. (2009) membuktikan bahwa kapang
Penicillium sp. memiliki nilai absorbansi tertinggi, artinya berarti telah melakukan
pendegradasian batubara tertinggi, mampu tumbuh menggunakan medium
batubara dan memiliki nilai pH medium yang berfluktuasi, artinya telah terjadi
proses degradasi selama proses inkubasi.
2.4. Enzim
Enzim adalah biokatalis atau substansi yang dapat mempercepat atau
membantu suatu reaksi kimia tanpa harus ikut terlibat di dalam reaksi itu sendiri.
sederhana sampai organisme multiseluler yang kompleks, termasuk manusia.
Enzim termasuk molekul protein. Reaksi biokimia yang paling sering saat
mengaplikasian enzim secara industri adalah peruraian hidrolitik komponen bahan
pangan yang memiliki berat molekul (BM) tinggi seperti pati, protein, selulosa,
dan sebagainya (Poedjiadi dan Supriyanti, 2006). Setiap jenis enzim memiliki
kisaran pH tertentu yang sangat menentukan enzim beraktivitas secara optimal.
Enzim bersifat spesifik artinya hanya mengkatalisis suatu reaksi yang dirancang
khusus untuk enzim tertentu, misalnya pektinase hannya dapat mendegradasi
pektin, bukan pati atau selulosa. Sekitar 80% dari enzim industrial adalah enzim
hidrolitik, yang digunakan untuk depolimerisasi (pemecahan molekul-molekul
yang kompleks menjadi yang lebih sedarhana) bahan-bahan alami (Hidayat et al.,
2006).
Mikroba merupakan sumber penting dari beberapa jenis enzim. Sebagai
sumber enzim, mikroba memiliki beberapa kelebihan jika dibandingkan dengan
hewan maupun tanaman, yaitu : produksi enzim pada mikroba lebih murah,
kandungan enzim dapat diprediksi dan dikontrol, pasokan bahan baku terjamin,
dengan komposisi konstan dan mudah dikelola (Hidayat et al., 2006). Ada 3
keuntungan yang berkaitan dengan enzim ekstra sel : pertama, tidak memerlukan
proses penghancuran sel saat memanen enzim (proses penghancuran sel tidak
selalu mudah dilakukan dalam skala besar). Kedua, enzim protein yang
disekresikan keluar sel umumnya terbatas jenisnya. Ini berarti enzim ekstrim sel
terhindar dari kontaminasi berbagai jenis protein. Ketiga, secara alami enzim
Ada tiga golongan utama sumber enzim yaitu tanaman, hewan, dan mikroba.
Enzim tanaman pada dasarnya diperoleh dari tanaman. Di antara kelompok ini
yang sudah dikenal luas yaitu papain, bromelin, ficin, dan enzim amilolitik dari
sereal, lipoksigenase dari kedelai dan specialized enzymes dari buah jeruk.
Sebagian besar enzim tanaman tersedia dalam bentuk unpurified powder extracts,
meski demikian ada juga yang tersedia dalam bentuk lain seperti papain dalam
bentuk stabilized dan purified liquid. Enzim dari hewan umumnya di peroleh dari
glandula. Sedangkan enzim mikrobial adalah diperoleh dari mikroba misalnya
yang bersumber dari fungi, bakteria maupun khamir seperti amilase, diastase,
lipase, dan sebagainya. Enzim yang berasal dari fungi menempati urutan teratas,
disusul dari bakteri dan khamir (Hidayat et al., 2006).
Penggolongan enzim berdasarkan tempat bekerjanya adalah endoenzim
dan eksoenzim. Endoenzim disebut juga enzim intraseluler, yaitu enzim yang
bekerjanya di dalam sel. Umumnya merupakan enzim yang digunakan untuk
proses sintesis di dalam sel dan untuk pembentukan energi (ATP) yang berguna
untuk proses kehidupan sel, misalnya dalam proses respirasi. Eksoenzim disebut
juga enzim ekstraseluler, yaitu enzim yang bekerjanya di luar sel. Umumnya
berfungsi untuk “mencernakan” substrat secara hidrolisis untuk dijadikan molekul
yang lebih sederhana dengan BM lebih rendah sehingga dapat masuk melalui
membran sel. Energi yang dibebaskan pada reaksi pemecahan substrat di luar sel
tidak digunakan selama proses kehidupan sel (Lehninger, 1982).
Kapang mendegradasi batubara menggunakan enzim ekstraseluler, hal
dikatalis melalui aktifitas enzim ekstratseluler (Ward, 1990). Enzim ekstraseluler
adalah enzim yang diekskresikan oleh kapang ke luar tubuhnya untuk
mendegradasi substrat. Enzim ekstraseluler tersebut akan menghasilkan medium
yang lebih gelap selama proses kultur cair atau cairan gelap pada permukaan
batubara ketika ditumbuhkan pada permukaan kultur agar (Faison et al., 1989).
Tabel 1. Enzim ekstraseluler pedegradasi lignin dari kapang pelapuk putih (white rot fungi) (Akhtar et al., 1997).
Enzim Tipe enzim Peran dalam degradasi
Peroksidase Degradasi unit
non–fenolik H2O2 38-47 2,5-3,0
Enzim pendegradasi lignin secara umum terdiri dari dua kelompok utama
yaitu lakase dan peroksidase yang terdiri dari lignin peroksidase dan mangan
peroksidase. Ketiga enzim tersebut bertanggung jawab terhadap pemecahan awal
polimer lignin dan menghasilkan produk dengan berat molekul rendah, larut
dalam air dan CO2 (Akhtar et al., 1997). Mangan peroksidase (MnP), lignin
peroksidase (LiP) atau lakase mampu mendegradasi komponen aromatik di
batubara dan mendepolimerisasinya menjadi komponen yang kaya oksigen dan
Lignin peroksidase (LiP) merupakan enzim utama dalam proses degradasi
lignin karena mampu mengoksidasi unit non fenolik lignin. Unit non fenolik
merupakan penyusun sekitar 90 persen struktur lignin. Oksidasi substruktur lignin
yang dikatalis oleh LiP dimulai dengan pemisahan satu elektron cincin aromatik
substrat donor dan menghasilkan radikal aril. LiP memotong ikatan Cα-Cβ
molekul lignin, pemotongan tersebut merupakan jalur utama perombakan lignin
oleh berbagai kapang pelapuk putih (Hammel, 1996).
Mangan peroksidase (MnP) berperan dalam oksidasi unit fenolik, sehingga
LiP dan MnP dapat bekerja secara sinergis. Siklus katalitik MnP dimulai dengan
pengikatan H2O2 atau peroksida organik dengan enzim ferric alami dan
pembentukan kompleks peroksida besi. Pemecahan ikatan oksigen peroksida
membutuhkan Fe okso-porpirin-radikal kompleks dalam pembentukan
MnP-komponen I, kemudian ikatan dioksigen dipecah dan dikeluarkan satu molekul air.
Reaksi berlangsung sampai terbentuk MnP-komponen II, ion Mn2+ bekerja
sebagai donor 1-elektron untuk senyawa antara porfirin dan dioksidasi menjadi
Mn3+. Mn3+ merupakan oksidasi kuat yang dapat mengoksidasi senyawa fenolik
tetapi tidak dapat menyerang unit non fenolik lignin (Perez et al., 2002).
Lakase ditemukan pada kapang, khamir, dan bakteri. Enzim ini tidak
membutuhkan H2O2 tetapi menggunakan molekul oksigen. Lakase mereduksi
oksigen menjadi H2O dalam substrat fenolik melalui reaksi satu elektron
membentuk radikal bebas yang dapat disamakan dengan radikal kation yang
2.5. Kromatografi Gas Spektroskopi Massa (GC-MS)
Kromatografi Gas-Spektroskopi Massa atau sering disebut GCMS (Gas
Chromatography Mass Spectrometry) adalah teknik analisis yang
menggabungkan dua metode analisis yaitu Kromatografi Gas dan Spektroskopi
Massa. Kromatografi gas adalah metode analisis, dimana sampel terpisahkan
secara fisik menjadi bentuk molekul-molekul yang lebih kecil (hasil pemisahan
dapat dilihat berupa kromatogram). Sedangkan spektroskopi massa adalah metode
analisis, dimana sampel yang dianalisis akan diubah menjadi ion-ion gasnya, dan
massa dari ion-ion tersebut dapat diukur berdasarkan hasil deteksi berupa
spektrum massa (Hermanto, 2008).
Pada GC hanya terjadi pemisahan untuk mendapatkan komponen yang
diinginkan, sedangkan bila dilengkapi dengan MS (berfungsi sebagai detektor)
akan dapat mengidentifikasi komponen tersebut, karena bisa membaca spektrum
bobot molekul pada suatu komponen, juga terdapat reference pada software
(Hermanto, 2008).
Pemisahan komponen senyawa dalam GCMS terjadi di dalam kolom
(kapiler) GC dengan melibatkan dua fase, yaitu fase diam dan fase gerak. Fase
diam adalah zat yang ada di dalam kolom, sedangkan fase gerak adalah gas
pembawa (Helium maupun Hidrogen dengan kemurnian tinggi, yaitu ± 99,995%).
Proses pemisahan dapat terjadi karena terdapat perbedaan kecepatan alir dari tiap
molekul di dalam kolom. Perbedaan tersebut dapat disebabkan oleh perbedaan
afinitas antar molekul dengan fase diam yang ada di dalam kolom. Selanjutnya
yang berfungsi sebagai detektor secara instrumentasi, MS adalah detektor bagi GC
(Hermanto, 2008).
2.6. Elektroforesis
Elektroforesis merupakan proses bergeraknya molekul bermuatan pada suatu
medan listrik. Kecepatan molekul yang bergerak pada muatan listrik tergantung
pada muatan, bentuk dan ukuran (Lehninger, 1982). Dengan demikian
elektroforesis dapat digunakan untuk separasi makromolekul (seperti protein dan
asam nukleat). Posisi molekul yang terseparasi pada gel dapat dideteksi dengan
pewarnaan.
Menurut Yuwono (2005), elektroforesis adalah suatu teknik pemisahan
molekul seluler berdasarkan atas ukurannya, dengan menggunakan medan listrik
yang dialirkan pada suatu medium yang mengandung sampel yang akan
dipisahkan. Kecepatan gerak molekul tergantung pada nisbah (rasio) muatan
terhadap massanya, serta tergantung pula pada bentuk molekulnya.
Kegunaan elektroforesis adalah menentukan berat molekul (estimasi).
Penetapan BM secara lebih teliti dapat dilakukan dengan ultrasentrifuge,
meskipun dengan elektroforesis cukup memenuhi syarat, dapat mendeteksi
terjadinya pemalsuan bahan, dapat mendeteksi terjadinya kerusakan bahan seperti
protein dalam pengolahan dan penyimpanan, untuk memisahkan spesies molekul
yang berbeda secara kualitatif, yang selanjutnya masing-masing spesies dapat
Salah satu jenis elektroforesis adalah elektroforesis SDS-PAGE. Sodium
Sodecyl Sulphate Polyacrylamide Gel Electrophoresis (SDS-PAGE) terutama
dilakukan untuk mengetahui apakah suatu protein monometrik ataukah
oligometrik, selain itu untuk menetapkan berat molekul dan jumlah rantai
polipeptida sebagai subunit atau monomer.
Pada mekanisme SDS-PAGE, protein bereaksi dengan SDS yang merupakan
deterjen anionic membentuk kompleks yang bermuatan negatif. Protein akan
terdenaturasi dan terlarut membentuk kompleks berikatan dengan SDS, berbentuk
elips atau batang, dan berukuran sebanding dengan berat molekul protein. Protein
dalam bentuk kompleks yang bermuatan negatif ini dipisahkan berdasarkan
muatan negatif ini dipisahkan berdasarkan muatan dan ukurannya secara
elektroforesis di dalam matriks gel poliakrilamid. Berat molekul protein dapat
diukur dengan menggunakan protein standar yang telah diketahui berat
molekulnya (Ummubalqis, 2000).
SDS-PAGE dilakukan pada pH normal. Pada metoda ini digunakan
anionic deterjent yang bersama dengan beta-merkaptoetanol dan pemanasan
menyebabkan rusaknya struktur tiga dimensi protein menjadi konfigurasi random
coil. Hal ini menyebabkan oleh terpecahnya ikatan disulfida yang selanjutnya
tereduksi menjadi gugus-gugus sulfihidril. SDS akan membentuk kompleks
dengan protein dan komleks ini bermuatan negatif karena gugus-gugus anion dari
SDS. Pada pH 7, SDS 1% dan merkaptoetanol 0,1 M sebagian besar rantai protein
mengikat sekitar 1,4 gr SDS per gram protein, dengan demikian jumlah SDS
hanya berdasarkan ukurannya (BM) di dalam kompleks SDS protein yang lebih
besar mempunyai mobilitas yang lebih kecil dibandingkan dengan kompleks yang
lebih kecil (Hames, 1998).
2.7. Iradiasi Gamma
Iradiasi merupakan pemancaran suatu energi elektromagnetik atau
partikel-partikel dengan kecepatan tinggi. Iradiasi dapat dibedakan menjadi dua macam,
yaitu iradiasi panas dan iradiasi pengion. Iradiasi panas menggunakan frekuensi
rendah atau dengan panjang gelombang tinggi, misalnya infra merah. Iradiasi
pengion menggunakan frekuensi tinggi, misalnya sinar alfa (α), sinar beta (β), dan
sinar gamma (Akhadi, 1997). Aplikasi sumber iradiasi pengion telah meluas
dalam berbagai bidang. Pemanfaatan sumber iradiasi misalnya bidang kedokteran,
bidang industri dan bidang pertanian. Efek kerusakan yang ditimbulkan
bergantung pada jenis dan kualitas iradiasi karena mempunyai daya tembus yang
rendah dan bermuatan positif, bentuk sinar alfa (α) merupakan partikel inti He
yang bergerak cepat. Partikel beta (β) memiliki daya tembus lebih besar
dibandingkan sinar alfa (α) dan bermuatan negatif (Darussalam, 1996).
Sinar gamma merupakan jenis iradiasi yang bisa digunakan dalam berbagai
bidang karena muatan netral, panjang gelombang pendek dan daya tembus paling
tinggi sehingga energi sinar gamma yang dipancarkan sumber terhadap target
dapat menimbulkan perubahan pada komposisinya. Perubahan dapat terjadi secara
acak dan tiba-tiba. Besar kecilnya efek iradiasi gamma tergantung dari energi dan
BAB III
METODOLOGI PENELITIAN
3.1. Tempat dan Waktu Penelitian.
Penelitian ini dilakukan pada bulan April sampai dengan bulan Juli tahun 2010 dan bertempat di Badan Tenaga Nuklir Nasional, Pasar Jum’at Jakarta.
3.2. Alat dan Bahan
Alat yang digunakan dalam penelitian ini adalah timbangan analitik, mikropipet, refrigerator, sentrifuse, pH meter, spektrofotometer Uv-vis, autoklaf, pipet tetes, Gas Chromatograph Mass Spectrometer (GC-MS) Shimadzu dan Mini Protein-Gel Elektroforesis “Atto”.
Bahan yang digunakan dalam penelitian ini adalah batubara subbituminus, kertas whatman No.1, alkohol 70 %, NaCl 0,85 %, isolat kapang Penicillium sp. dan Trichoderma sp. koleksi dari BATAN (Irawan Sugoro), aseton, akuades, medium Potato Dextrose Agar (PDA), Medium Minimal Salt (MMS), agar bakto, sukrosa 1 %, Lowry I ( 2% Na2CO3 dalam NaOH 0,1 N ; 2,7% K.Na tartat; 2%
CuSO4), Lowry II (Folin dan akuades (1:1) ), Larutan Standar BSA, buffer
sampel, Separating Gel (10%) dan Stacking Gel (45%), tanin, methylene blue,
3.3. Cara Kerja
3.3.1. Persiapan Serbuk Batubara
Batubara dihaluskan menggunakan mortar secara aseptik, kemudian disaring menggunakan saringan berukuran 0,2 mm (70 mesh). Serbuk batubara yang telah siap ditimbang sebanyak 5 gr dan dimasukkan ke dalam plastik
polyetilen. Serbuk batubara ditutup rapat dan diiradiasi gamma (5 kGy) dan tidak diiradiasi (0 kGy).
3.3.2. Pembuatan Medium
Tabel 2. Komposisi medium perlakuan
No. Nama Medium PDA
1. Potato Dextrose
Agar Minimal Salt
3.3.2.1. Medium Potato Dextrose Agar
3.3.2.2. Medium Minimal Salt (MMS)
Sebanyak 0,52 gr MgSO4.7H2O; 0,003 gr ZnSO4.7H2O pH 5,5 ; 5 gr
K2HPO4; 0,005 gr FeSO4, dan 1 gr NH4(SO4) dilarutkan dalam 1 liter akuades,
kemudian dilarutkan sampai homogen. Medium MMS tersebut disterilisasi menggunakan autoklaf dengan suhu 121°C selama 15 menit (Silva et al., 2007).
3.2.2.3. Medium Potato Dextrose Agar Minimal Salt (PDAM)
Medium PDAM dibuat dengan mencampurkan medium PDA dan MMS dengan perbandingan 1:1 atau 75 ml : 75 ml. Medium PDA ditambahkan 1 % agar bakto atau 0,75 gr. Medium PDA dan MMS disterilisasi dulu menggunakan autoklaf dengan suhu 121°C dengan waktu 15 menit sebelum dicampurkan. Medium PDAM dihomogenkan dengan cara pengadukan. Sebanyak 5 ml dimasukkan tabung reaksi untuk peremajaan kapang pada tabung reaksi (agar miring).
3.2.2.4. Medium Minimal Salt + Sukrosa (MMSS)
3.3.3. Peremajaan Kultur Kapang
Sebanyak 5 ml medium PDAM di masukkan ke dalam tabung reaksi (untuk agar miring). Kultur stok/isolat kapang diinokulasikan pada medium PDAM tersebut menggunakan ose. Kultur kapang tersebut diinkubasi pada suhu ruang sampai kapang menghasilkan spora, selama 5-7 hari pada suhu ruang.
3.3.4. Kultur Inokulum Spora
Kultur spora yang sudah diremajakan pada medium PDAM, kemudian diambil sporanya menggunakan ose. Kultur spora pada medium ditambahkan 5 ml NaCl steril 0,85 % sambil dilepaskan sporanya menggunakan ose, kemudian dimasukkan ke dalam yellow tube steril.
3.3.5. Uji Enzim Ekstraseluler 3.3.5.1. Fenoloksidase
Pengujian berdasarkan reaksi kimia warna Bavendamm. Kapang ditumbuhkan pada medium PDA yang mengandung tanin 4 mM/L. Lingkaran warna coklat yang terbentuk menunjukkan adanya ekskresi fenoloksidase kapang (Tao et al., 2009).
3.3.5.2. Peroksidase
Metode penambahan warna medium untuk pengujian enzim peroksidase.
methylene blue sebanyak 0,1 gr/L ditambahkan kedalam medium PDA dan
3.3.5.3. Mangan Peroksidase
Sebanyak 1 gr/L Manganese cloride ditambahkan dalam medium PDA dan kapang diinokulasikan lalu diinkubasi selama 2 minggu. Selama masa inkubasi dilakukan pengamatan terhadap titik-titik hitam-coklat yang terbentuk. MnP mengkatalis terjadinya oksidasi MnCl2 menjadi MnO2 yang dihasilkan
warna hitam-coklat (Tao et al., 2009).
3.3.6. Pengukuran pH medium
Medium diukur nilai pHnya dengan menggunakan pH meter. Selanjutnya dibuat grafik perubahannya.
3.3.7. Pengujian Biosolubilisasi Batubara
Tabel 3. Data perlakuan kultur kapang
Kapang Dosis Iradiasi
Serbuk Batubara
3.3.7.1. Pengukuran Biosolubilisasi Batubara
3.3.8. Pengukuran Hidrolisis FDA
Supernatan dimasukkan 1 ml ke dalam tabung reaksi, kemudian ditambahkan 4 ml KH2PO4 buffer (pH 7,6) 60 mM. Reaksi dimulai dengan
menambahkan 40 µg FDA (Fluorescien diacetate) dalam 4 ml aseton, kemudian diinkubasi dalam pengocokan selama 20 menit. Setelah penginkubasian segera ditambahkan aseton sebanyak 4 ml untuk menghentikan reaksi. Suspensi disaring dengan kertas Whatman No. 1, filtrat dimasukkan ke dalam tabung reaksi, ditutup dengan kertas parafilm dan disimpan dalam es batu untuk menguapkan aseton. Nilai OD ditera dengan menggunakan spektrofotometer pada panjang gelombang 490 nm (Breeuwer, 1996).
3.3.9. Pengukuran Kadar Protein Eketraseluler dengan Metode Lowry
Sebanyak 0,5 ml sampel ditambahkan 2,5 ml larutan Lowry I dan diinkubasi pada suhu ruang selama 10 menit. Kemudian ditambahkan 0,25 ml larutan Lowry II, divortek dan diinkubasi pada suhu ruang selam 30 menit. Setelah itu absorbansi dibaca dengan Spektrofotometer UV-Vis pada panjang gelombang 750 nm dan dibandingkan dengan standar Bovine Serum Albumin
3.3.10.Analisis Hasil Biosolubilisasi Batubara oleh Kapang Penicilium sp. dan Trichoderma sp. Dengan Menggunakan GC-MS
Supernatan dan pelarut dicampurkan dengan perbandingan 1:1. Pelarut yang digunakan adalah benzena : heksana : dietil eter dengan perbandingan 3:1:1. Campuran tersebut dimasukkan ke dalam corong Buchner lalu diaduk sampai bercampur kemudian didiamkan beberapa saat sampai terbentuk fase atas dan bawah. Fase atas dipakai untuk identifikasi jenis senyawa produk hasil biosolubilisasi batubara dan menentukan kadarnya dengan menggunakan GC-MS Shimadzu. Kolom yang digunakan adalah Dimethyl polysiloxana dengan kondisi suhu kolom oven 50 0C, suhu injeksi 280 0C, laju alir 1,54 ml/menit, dan fase gerak gas helium. Kontrol yang digunakan adalah medium MMSS yang ditambahkan serbuk batubara yang diiradiasi dan tidak diiradiasi (Silva et al., 2007).
3.3.11.Karakteristik Enzim Ekstraseluler dengan Elektroforesis
Karakterisasi enzim ekstraseluler dilakukan melalui elektroforesis dengan menggunakan SDS-PAGE. Adapun tahapan kerja yang dilakukan antara lain:
1. Preparasi sampel
2. Preparasi gel elektroforesis a. Separatinggel (10%)
Sebanyak 6 ml Acrylamid 30 % dicampurkan dengan separating gel buffer (1,5 Tris-HCl, pH 8,8) sebanyak 4,5 ml kemudian akuabides 7,5 ml, SDS 50 µl, amonium persulfate (APS) 0,08 ml dan TEMED 0,01 ml.
b. Stacking gel (45%)
Sebanyak 0,9 ml Acrylamid 30 % dicampurkan dengan separating gel buffer (1,5 Tris-HCl, pH 8,8) sebanyak 1,5 ml kemudian akuabides 3,6 ml, SDS 25 µl, amonium persulfate (APS) 0,02 ml dan TEMED 0,01 ml.
3. Proses Elektroforesis
Setelah separating gel dibuat, kemudian dimasukkan sedikit demi sedikit ke dalam alat elektroforesis dengan mikropipet, lalu ditambahkan akuades untuk meratakan separating gel tersebut. Setelah separating gel membeku, akuades dibuang dan dimasukkan stacking gel sedikit demi sedikit, lalu dipasang sisir pembentuk kolom dan dibiarkan hingga stacking gel
membeku kemudian sisir diangkat.
Gel diangkat lalu diwarnai dengan staining solution coomassie blue R-250 kurang lebih selama 24 jam.Gel dicuci dengan larutan destaining yang terdiri atas methanol 40 %, asam asetat 7,5 % dan akuades kurang lebih selama 24 jam. Protein yang telah didestaining kemudian discan dan dianalisa dengan Lab.Image untuk menentukan nilai RF (Retensi Faktor) sebagai representasi dari profil protein yang dihasilkan dan berat molekulnya.
3.4. Analisa Data
4.1. Hasil Analisis Kualitatif Enzim Ekstraseluler Kapang
Pengujian enzim ekstraseluler kapang sangatlah penting dilakukan sebelum
proses biosolubilisasi batubara. Pengujian ini bertujuan mengetahui ada atau
tidaknya enzim ekstraseluler kapang yang berperan dalam proses biosolubilisasi
batubara. Uji enzim ini menggunakan substrat atau pereaksi (tanin, methylene blue
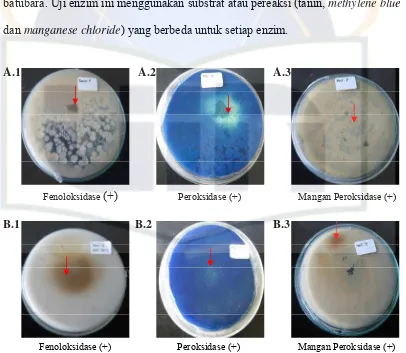
dan manganese chloride) yang berbeda untuk setiap enzim.
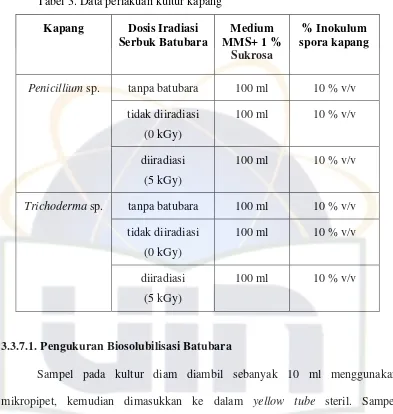
A.1 A.2 A.3
Fenoloksidase (+) Peroksidase (+) Mangan Peroksidase (+)
B.1 B.2 B.3
Fenoloksidase (+) Peroksidase (+) Mangan Peroksidase (+)
Gambar 4. Uji kualitatif adanya enzim ekstraseluler kapang Penicillium sp. (A) dan Trichoderma sp. (B)
Hasil pengujian kualitatif enzim ekstraseluler kapang Penicillium sp. dan
Trichoderma sp. menunjukkan hasil yang positif untuk terdapatnya enzim
fenoloksidase, peroksidase, dan mangan peroksidase (MnP) (Gambar 4).
Enzim-enzim ekstraseluler yang terlibat dalam proses biosolubilisasi batubara adalah
peroksidase, esterase, fenoloksidase atau lakase, MnP dan lignin peroksidase.
Enzim-enzim ini berperan untuk memecah senyawa kompleks menjadi senyawa
yang lebih sederhana, kemudian digunakan sebagai sumber energi untuk
pertumbuhan kapang (Laborda et al., 1999; Fakuosa dan Hofrichter, 1999).
Tao et al. (2009) mengindikasikan perubahan medium untuk mengamati
terjadinya lingkaran coklat oleh fenoloksidase yang dihasilkan oleh kapang
(Gambar 4 A.1 dan B.1). Pada (Gambar 4 A.2 dan B.2) positif terdapatnya enzim
peroksidase karena pudarnya warna medium pada bagian medium yang ditumbuhi
oleh kapang. (Gambar 4 A.3 dan B.3) adanya titik-titik coklat yang terbentuk
sehingga positif terdapatnya enzim MnP. Hal itu terjadi karena MnP mengkatalis
terjadinya oksidasi MnCl2 menjadi MnO2 hingga dihasilkan warna coklat.
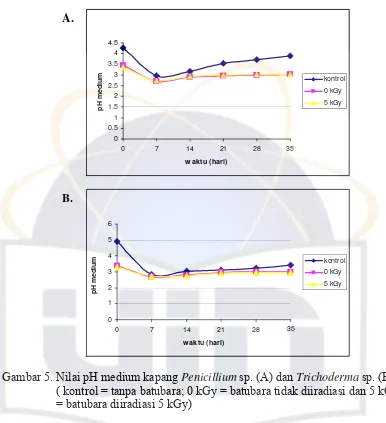
4.2. Hasil Pengukuran pH Medium Kapang
Pola perubahan pH medium, baik kapang Penicillium sp. dan Trichoderma
sp. cenderung sama (Gambar 5). Nilai pH medium setelah 7 hari inkubasi
mengalami penurunan dan setelah itu mengalami kenaikan sampai hari ke-35
inkubasi. Penurunan pH medium yang mengandung batubara yang tidak diiradiasi
(0 kGy) dan diiradiasi (5 kGy) yang terjadi pada hari ke-7 inkubasi tidak terlalu
biosolubilisasi batubara seperti asam karboksil dan asam fulvat yang merupakan
senyawa humat yang terdapat dalam batubara (Cerniglia,1992).
Menurut penelitian Mustikasari (2009), nilai pH cenderung mengalami
kenaikan seiring dengan bertambahnya masa inkubasi. Kondisi peningkatan pH
medium dapat disebabkan oleh dua kemungkinan. Kemungkinan pertama karena
hasil degradasi lignin menghasilkan berbagai macam senyawa fenolik yang
memiliki gugus –OH. Keberadaan gugus tersebut dapat meningkatkan nilai pH
pada masa akhir inkubasi dan kemungkinan kedua adalah terjadinya peningkatan
jumlah sel yang lisis. Menurut Judoamidjojo et al. (1992) sel yang lisis tersebut
terdeaminasi dan menyebabkan peningkatan pH.
Nilai pH medium kapang Penicillium sp. dan Trichoderma sp. selama proses
inkubasi berkisar antara 2,69 - 4,9 pada kontrol (tanpa batubara), pada medium
yang mengandung batubara yang tidak diradiasi (0 kGy) dan yang diradiasi (5
kGy). Nilai pH medium kapang Penicillium sp. berkisar antara 2,71- 4,62
(Gambar 5 A), sedangkan pada medium kapang Trichoderma sp. adalah 2,69- 4,9
(Gambar 5 B). Enzim-enzim ekstraseluler pendegradasi lignin bekerja pada pH
yang optimum. Kondisi pH medium mendukung kerja enzim fenoloksidase atau
lakase yang bekerja optimum pada pH 3,5-7. Lignin peroksidase bekerja optimum
pada pH 2,5-3,0 dan mangan peroksidase bekerja optimum pada pH 4,0-4,5
(Akhtar, 1997). pH medium kultur kapang Penicillium sp. dan Trichoderma sp.
masih pada kisaran pH yang optimum untuk bekerjanya enzim fenoloksidase atau
A.
Gambar 5. Nilai pH medium kapang Penicillium sp. (A) dan Trichoderma sp. (B). ( kontrol = tanpa batubara; 0 kGy = batubara tidak diiradiasi dan 5 kGy = batubara diiradiasi 5 kGy)
Hasil pengujian statistik menunjukkan nilai pH medium kapang Penicillium
sp. menunjukkan perbedaan yang signifikan (probabilitas ≤ 0,05) (Lampiran 12).
Selama proses inkubasi, medium yang mengandung batubara yang diiradiasi (5
kGy) nilai pH cenderung lebih tinggi daripada medium yang mengandung
batubara yang tidak diiradiasi (0 kGy). Nilai pH medium yang mengandung
batubara lebih asam daripada medium yang tidak mengandung batubara (kontrol).
Diduga iradiasi menyebabkan terlepasnya senyawa-senyawa fenolik yang
kapang Penicillium sp., hasil pengujian statistik pada penggunaan kapang
Trichoderma sp. tiap perlakuan menunjukkan tidak adanya perbedaan yang
signifikan pada seluruh perlakuan (probabilitas ≥ 0,05) (Lampiran 12)
Perubahan pH merupakan hal yang menjadi salah satu faktor pengukuran
dalam proses biosolubilisasi batubara. Nilai pH medium kapang Penicillium sp.
dan Trichoderma sp. cenderung asam, yaitu 2,69- 4,9 dan memiliki pola yang
sama setelah 35 hari inkubasi. Kisaran pH tersebut memungkinkan untuk
pertumbuhan kapang Penicillium sp. dan Trichoderma sp. karena kapang tersebut
dapat tumbuh pada kisaran pH yang luas yaitu pH 2 - 8,5 dan pertumbuhan
kapang akan lebih baik pada kondisi asam atau pH rendah (Fardiaz, 1989).
Dengan pertumbuhan kapang yang lebih baik itu dapat mempengaruhi kadar dan
aktivitas enzim yang dihasilkan oleh kapang selama proses biosolubilisasi
batubara. Nilai pH medium sangat mempengaruhi hasil biosolubilisasi batubara
oleh kapang Penicillium sp. dan Trichoderma sp. sehingga hasilnya dapat dilihat
pada biosolubilisasi batubara pada panjang gelombang 250 nm dan 450 nm.
4.3. Hasil Biosolubilisasi Batubara
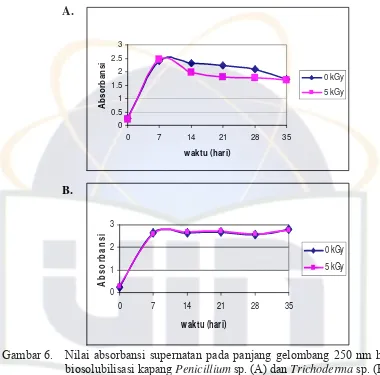
Pengukuran nilai absorbansi supernatan pada panjang gelombang 250 nm
bertujuan untuk mendeteksi adanya gugus fenolik produk hasil biosolubilisasi
batubara oleh kapang Penicillium sp. dan Trichoderma sp. Gugus fenolik
terbentuk oleh proses solubilisasi senyawa lignin yang merupakan komponen
penyusun batubara. Penguraian senyawa lignin ini dibantu oleh enzim lignin
Semakin tinggi nilai absorbansi maka semakin baik pula hasil biosolubilisasi
batubara (Selvi dan Banerjee, 2007).
A.
Gambar 6. Nilai absorbansi supernatan pada panjang gelombang 250 nm hasil biosolubilisasi kapang Penicillium sp. (A) dan Trichoderma sp. (B)
Hasil pengukuran supernatan kultur medium kapang Penicillium sp. dan
Trichoderma sp. pada panjang gelombang 250 nm dan 450 nm memiliki pola
yang sama, baik pada batubara yang tidak diiradiasi (0 kGy) maupun batubara
yang diiradiasi (5 kGy) (Gambar 6 dan 7). Hasil pengujian statistik juga
menunjukkan tidak adanya perbedaan yang signifikan pada seluruh perlakuan
(probabilitas ≥ 0,05) (Lampiran 12). Pada (Gambar 6) terlihat bahwa nilai
diiradiasi (0 kGy) dan batubara yang diiradiasi (5 kGy) mengalami peningkatan
pada hari ke-7 inkubasi dan setelah itu mengalami penurunan pada panjang
gelombang 250 nm. Pada medium kapang Penicillium sp. yang mengandung
batubara yang diiradiasi (5 kGy) tidak mempengaruhi biosolubilisasi batubara.
Tanpa radiasi gamma, nilai absorbansi kapang Penicillium sp. memililki nilai
absorbansi yang tinggi.
Nilai absorbansi supernatan meningkat dan menurun seiring dengan
bertambahnya masa inkubasi. Nilai absorbansi yang meningkat pada hari ke-7
inkubasi disebabkan karena proses biosolubilisasi batubara padat diurai menjadi
batubara terlarut dan selain itu dihasilkan pula gas CO2. Nilai absorbansi yang
menurun pada hari ke-14 dan sampai akhir inkubasi disebabkan karena proses
degradasi atau biosolubilisasi batubara yang sudah melarut kemudian diurai
kembali menjadi komponen-komponen yang lebih sederhana dan dihasilkan pula
gas CO2 (Ward, 1990). Unit fenolik hasil degradasi lignin dioksidasi oleh enzim
lakase yang berperan dalam oksidasi unit fenolik (Perez et al.,2002).
Tiap kapang memiliki waktu yang berbeda dalam hal memiliki nilai
absorbansi tertinggi pada panjang gelombang 250 nm. Kapang Penicillium sp.
nilai absorbansi tertinggi terjadi pada inkubasi hari ke-7 dalam medium yang
mengandung batubara yang tidak diiradiasi (0 kGy) dan diirasiasi (5 kGy).
Kapang Trichoderma sp. nilai absorbansi tertinggi pada inkubasi hari ke-35 dalam
medium yang mengandung batubara yang tidak diiradiasi (0 kGy) dan diirasiasi (5
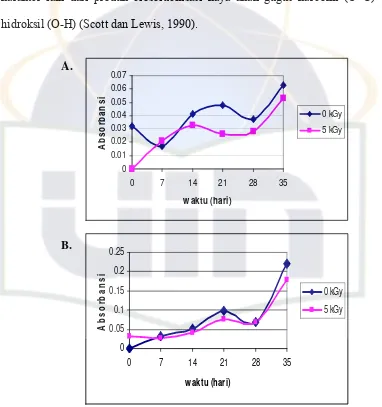
Tingkat biosolubilisasi juga diamati melalui nilai absorbansi pada panjang
gelombang 450 nm. Pengukuran nilai absorbansi pada panjang gelombang
tersebut bertujuan untuk mendeteksi produk hasil biosolubilisasi berupa gugus
karboksil dan hidroksil oleh kapang Penicillium sp. dan Trichoderma sp. karena
karakter lain dari produk biosolubilisasi kaya akan gugus karbonil (C=O) dan
hidroksil (O-H) (Scott dan Lewis, 1990).
A.
Pada (Gambar 7A) terlihat bahwa nilai absorbansi pada masa inkubasi hari
ke-7 dan 28 pada medium yang mengandung batubara yang tidak diiradiasi (0 kGy)
mengalami penurunan. Nilai absorbansi terus meningkat pada masa inkubasi hari
ke-14 dan 21, kemudian nilai absorbansi meningkat lagi pada hari ke-35.
Peningkatan nilai absorbansi tertinggi pada masa inkubasi hari ke-35 yaitu,
sebesar 0,063 pada medium yang mengandung batubara yang tidak diiradiasi (0
kGy). Nilai absorbansi pada medium yang mengandung batubara yang diiradiasi
(5 kGy) pada masa inkubasi dari hari ke-0, 7 dan 14 terus mengalami peningkatan,
kemudian nilai absorbansi menurun lagi pada hari ke-28 dan 35. Peningkatan nilai
absorbansi tertinggi pada masa inkubasi hari ke-35 yaitu, sebesar 0,053 pada dosis
batubara yang diiradiasi (5 kGy). Iradiasi gamma 5 kGy pada batubara tidak
mempengaruhi biosolubilisasi. Hasil pengujian statistik kapang Penicillium sp
menunjukkan tidak adanya perbedaan yang signifikan pada seluruh perlakuan
(probabilitas ≥ 0,05) (Lampiran 12).
(Gambar 7 B) terlihat bahwa nilai absorbansi pada medium yang mengandung
batubara yang tidak diiradiasi (0 kGy) dari ke-0, 7, 14, 21 dan 35 terus mengalami
peningkatan, kemudian nilai absorbansi menurun pada hari ke-28 inkubasi.
Peningkatan absorbansi tertinggi pada masa inkubasi hari ke-35 yaitu, 0,220. Nilai
absorbansi pada dosis batubara yang diiradiasi (5 kGy) menurun pada hari ke-7
dan 28. Kemudian meningkat pada hari ke-14 dan ke-21. Nilai absorbansi
meningkat lagi pada hari ke-35 yaitu, sebesar 0,178. Peningkatan nilai absorbansi
sp. menunjukkan tidak adanya perbedaan yang signifikan pada seluruh perlakuan
(probabilitas ≥ 0,05) (Lampiran 12)
Jika dibandingkan dengan nilai pH medium (Gambar 5) maka dapat dilihat
adanya hubungan yang berbanding terbalik antara pH medium dengan absorbansi
supernatan. Ketika nilai pH medium meningkat maka nilai absorbansi supernatan
mengalami penurunan sedangkan jika nilai pH menurun maka nilai absorbansi
supernatan menigkat. Hal ini sesuai dengan hasil penelitian yang dilakukan oleh
Selvi dan Banerje (2007), yang menghasilkan biosolubilisasi yang tertinggi pada
pH yang rendah.
Kapang Trichoderma sp. memiliki nilai absorbansi lebih tinggi daripada
kapang Penicillium sp. pada medium yang mengandung batubara yang tidak
diiradiasi (0 kGy) selama proses inkubasi. Hal ini sesuai dengan penelitian Sugoro
et al. (2009) kapang Trichoderma sp. memiliki nilai absorbansi tertinggi atau
memiliki kemampuan tertinggi dalam mendegradasi batubara subbituminus
Sumatera Selatan dibandingkan isolat kapang Penicillium sp., Mucor sp.,
Aspergillus sp. dan isolat lainnya (belum teridentifikasi). Menurut Ward (1990)
perbedaan absorbansi menunjukkan adanya perbedaan pada tingkat biosolubilisasi
batubara oleh kapang melalui aktivitas enzim ekstraseluler menjadi produk yang
dapat larut dan mencair, sehingga dalam penelitian ini perlu dilakukan analisis
4.4. Hasil Hidrolisis FDA
Nilai absorbansi diukur bertujuan untuk melihat aktivitas dari enzim
intraseluler dan enzim ektraseluler dalam kegiatannya menghidrolisis FDA
(Fluorescien diacetate). FDA dalam pengujian ini merupakan substrat sebagai
pengganti batubara. Jumlah FDA yang terhidrolisis menunjukkan jumlah enzim
intraseluler dan ekstraseluler. Selain ketiga enzim seperti lakase, lignin
peroksidase dan mangan peroksidase ternyata masih ada enzim ektraseluler lain
seperti lipase, protease dan terutama esterase. Aktivitas dari enzim akan
menghasilkan senyawa yang berpendar berwarna kuning (Lampiran 16.5
)(Breeuwer, 1996).
Hasil pengujian statistik kapang Penicillium sp. menunjukkan tidak adanya
perbedaan yang signifikan pada seluruh perlakuan (probabilitas ≥ 0,05) (Lampiran
12).(Gambar 8 A) terlihat bahwa nilai absorbansi pada kontrol (tanpa batubara),
batubara yang tidak diiradiasi (0 kGy) dan batubara yang diiradiasi (5 kGy)
mengalami peningkatan pada inkubasi hari ke-7. Pada kontrol (tanpa batubara),
nilai absorbansi menurun pada hari ke-14, dan terus meningkat pada hari ke-21,
28 dan 35, sedangkan pada batubara yang tidak diiradiasi (0 kGy) dan batubara
yang diiradiasi (5 kGy) terus meningkat pada hari ke-14 dan nilai absorbansi
menurun pada hari ke-21, 28, dan 35. Nilai absorbansi tertinggi pada batubara
yang tidak diiradiasi (0 kGy) dan batubara yang diiradiasi (5 kGy) yaitu, pada hari
ke-14 yaitu sebesar 0, 85, sedangkan pada kontrol (tanpa batubara) nilai aborbansi
FDA karena dipengaruh oleh pH medium, disaat hidrolisis FDA terus meningkat
maka nilai pH medium juga meningkat (Gambar 5).
A.
Gambar 8. Nilai absorbansi pada panjang gelombang 490 nm hasil hidrolisis FDA kapang Penicillium sp. (A) dan Trichoderma sp. (B). ( kontrol = tanpa batubara; 0 kGy = batubara tidak diiradiasi dan 5 kGy = batubara diiradiasi 5 kGy).
(Gambar 8 B) terlihat bahwa nilai absorbansi batubara yang tidak diiradiasi (0
kGy) dan batubara yang diiradiasi (5 kGy) mengalami peningkatan pada hari ke-7
sampai hari ke-14, 21 dan 28. Nilai absorbansi hidrolisis FDA menurun pada
hari-35. Pada kontrol nilai absorbansi menurun pada hari ke-7 dan meningkat pada hari
batubara yang diiradiasi (5 kGy) yaitu pada masa inkubasi hari ke-28, sedangkan
pada kontrol (tanpa batubara) nilai absorbansi tertinggi pada hari ke-21. Hasil
pengujian statistik kapang Trichoderma sp. menunjukkan perbedaan yang
signifikan (probabilitas ≤ 0,05) (Lampiran 12)
Peningkatan nilai absorbansi hidrolisis FDA tiap masa inkubasi karena jumlah
kapang yang terus meningkat sehingga aktivitas enzim juga tinggi, dan menurut
penelitian Indahwati (2009), hasil degradasi batubara yang kompleks menjadi
senyawa yang terlarut dalam medium menyebabkan peningkatan kekeruhan
medium sehingga nilai absorbansi juga meningkat. Menurut Fengel dan Weneger
(1995), enzim-enzim pendegradasi lignin harus bekerja secara aktif untuk
mendegradasi zat-zat makromolekul batubara, untuk itu perlu dilakukan analisis
kadar protein ekstraseluler kapang. Kadar protein ekstraseluler berhubungan
dengan enzim-enzim yang dihasilkan oleh kapang, sehingga bisa dihubungkan
dengan biosolubilisasi.
4.5. Hasil Kadar Protein Ekstraseluler Kapang
Pengukuran kadar protein ekstraseluler kapang Penicillium sp. dan
Trichoderma sp. bertujuan untuk mengetahui seberapa besar enzim ekstraseluler
diekresikan oleh kapang. Hasil uji menunjukkan bahwa kadar protein ektraseluler
yang tinggi menyebabkan warna supernatan akan lebih gelap setelah proses uji
(Lampiran 16.4), karena menurut Faison et al. (1989), biosolubilisasi batubara
oleh kapang melalui proses ektraseluler akan menghasilkan medium yang lebih
0
Gambar 9. Nilai kadar protein ekstraseluler kapang Penicillium sp.(A) dan Trichoderma sp. (B) (kontrol = tanpa batubara; 0 kGy = batubara tidak diiradiasi dan 5 kGy = batubara diiradiasi 5 kGy).
(Gambar 9 A), pada kapang Penicillium sp. terlihat bahwa kadar protein
ekstraseluler pada kontrol (tanpa batubara) dan medium yang mengandung
batubara yang tidak diiradiasi (0 kGy) menurun pada hari ke-7 inkubasi dan
meningkat pada hari ke-14 inkubasi. Pada kontrol (tanpa batubara) kadar protein
ekstraseluler tertinggi yaitu pada hari ke-35 inkubasi, sedangkan pada medium
yang mengandung batubara yang tidak diiradiasi (0 kGy) tertinggi pada hari ke-21