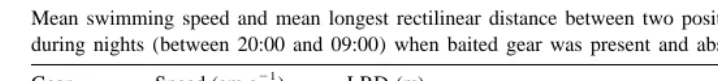L
Journal of Experimental Marine Biology and Ecology 247 (2000) 195–208
www.elsevier.nl / locate / jembe
Food-search strategy in ling (Molva molva L.): crepuscular
activity and use of space
a ,* b b
¨ Svein Løkkeborg , Katrine Skajaa , Anders Ferno
a
Institute of Marine Research, Fish Capture Division, P.O. Box 1870, N-5817 Bergen, Norway
b
Department of Fisheries and Marine Biology, University of Bergen, N-5020 Bergen, Norway Received 8 January 1999; received in revised form 14 December 1999; accepted 18 December 1999
Abstract
The activity rhythm and use of space in the food-searching behaviour of ling (Molva molva L.) were studied by means of a stationary positioning system. The system consisted of a fixed array of three hydrophones, which monitored the position of each individual being tracked once every three minutes. Five ling were tagged in situ by allowing them to ingest a transmitter wrapped in bait and were tracked continuously for 6 to 11 days. Ling showed a crepuscular activity rhythm with higher levels of swimming activity at dawn and dusk. The fish occupied a home range throughout the study, and during inactive periods (about 65% of the time) they remained within a small core area. Most of the time spent outside the core area was during the period of high activity at dawn. Food-search behaviour was studied by setting mackerel-baited fishing gear in the experimental area. When baits were present, ling moved more slowly and within a more limited area, indicating area-restricted searching in the presence of a food odour. Ling responded to and located baits in both their active and inactive periods during the day, but were not observed to react at night. Comparing these findings with those from similar tracking experiments on cod (Gadus morhua L.) suggested that ling are less active at night and show a more restricted use of space by occupying a core area. This may reflect different feeding strategies between these gadoid species with ling
taking a higher proportion of mobile prey and being more dependent on visual stimuli. 2000
Elsevier Science B.V. All rights reserved.
Keywords: Activity rhythm; Foraging behaviour; Ling; Telemetry; Use of space
*Corresponding author. Tel.: 147-55236826; fax: 147-55236830.
E-mail address: [email protected] (S. Løkkeborg)
1. Introduction
The success of an animal in obtaining food depends on its efficiency in locating resources under various environmental conditions (Bell, 1990). The probability of locating, encountering and capturing prey may vary through time and space, and thus lead to disproportionate use of time and space. Several studies have demonstrated diel
¨ ¨ rhythms in feeding activity in fish (e.g. Thorpe, 1978; Fraser et al., 1993; Alanara and
¨ ¨
Brannas, 1997), and such rhythmicities have often been explained by behavioural adaptations to temporal variations in the probability of detecting and capturing prey
¨
(Løkkeborg, 1998; Løkkeborg and Ferno, 1999). Diel variations in the utilization of space have been observed in several field studies on fishes (Hawkins et al., 1974;
˚
Hawkins et al., 1980; Bradbury et al., 1995; Engas et al., 1996). Disproportionate use of space may be a reflection of temporal variations in activity or an adaptation to spatial variations in food availability. To understand the feeding behaviour of animals, therefore, it may be essential to study spatio–temporal variations in activity patterns.
The sensory modalities involved in food search are an important aspect of feeding behaviour. Vision and chemoreception are vital senses in food search, but the senses by which prey are most effectively located and captured may vary with time and space, as well as depending on prey type (Atema, 1980; Løkkeborg, 1998). Their relative importance is affected by environmental conditions such as light level, turbidity, current speed and turbulence, and by prey characteristics and behaviour. Chemoreception and rheotaxis are probably the most important factors in locating food at some distance, due to the limited visibility in water (Atema, 1980).
In this field experiment we studied several aspects of the foraging behaviour of ling (Molva molva L.), such as activity rhythm, use of space and the role of the senses. Ling is a commercially important species (Bjordal and Løkkeborg, 1996). It is solitary and benthic or semi-benthic and inhabits rocky and sandy substrates at depths of 60 to 1000
˚
m in the east and north-west Atlantic (Pethon, 1989; Sunnana, 1992). Adult ling are regarded as top predators (Bergstad, 1991), and their behaviour can therefore be related mainly to feeding. Ling have a piscivorous diet supplemented by cephalopods, crustaceans and echinoderms (Rae and Shalton, 1982; Svetovidov, 1986). Since they are predators in relatively deep water where light is limited, it is reasonable to assume that they use both chemoreception and vision for locating food. To the best of our knowledge the activity pattern and food search behaviour of ling have not been studied previously. Behavioural observations in the field using underwater TV cameras have contributed to our understanding of the feeding behaviour of animals (Wilson and Smith, 1984;
¨
Ferno et al., 1986; Zimmer-Faust et al., 1995). However, this method is not capable of demonstrating the patterns of movement of marine animals. In this study we used a stationary positioning system with a fixed hydrophone array to study the movement patterns of ling under natural conditions and in response to sources of food odours. Similar tracking experiments have been carried out to study the feeding behaviour of cod
¨
2. Materials and methods
2.1. Experimental area and procedure
The study was carried out in April 1995 in a branch fjord in western Norway (2.5 km long and approximately 800 m wide, Fig. 1) with depths ranging from 80 to 100 m. The muddy bottom is relatively flat, but with some stones. On each side of the fjord is a steep rock wall, and where the fjord joins the main fjord the depth falls steeply to about 400 m. The current velocity and direction were recorded every 10 min by two current meters (SD6000, Sensordata A / S, Bergen, Norway) suspended one metre above the
21
seabed in the experimental area. The current velocity was usually below 2 cm s .
A stationary telemetry system (VRAP, Vemco Ltd., Halifax, Canada) was used to track the fish, which were tagged with acoustic transmitters. The positioning system consisted of a fixed array of three hydrophone buoys anchored to the seabed in a triangular configuration with radio transmission to a base station installed on board a research vessel. The distances between the buoys were between 381 and 592 m. The hydrophone buoys received acoustic pulses from acoustic transmitters. Using the time delays of pulse arrivals at the hydrophones, the base station calculated the position of the transmitters relative to the three buoys. A number of received pulse-times were aligned and an average position was calculated. The system was capable of tracking up to ten fish tagged with transmitters that operated at different frequencies. However, the system monitored only one frequency at a time, and the elapsed time between successive positional fixes thus varied with the number of animals being tracked. In this study each fish was tracked approximately every three minutes except during the first days, when fewer individuals were tracked. Under optimal acoustic conditions the transmitter can be localized to an accuracy of about one metre within the triangle of buoys. When the transmitter was outside the triangle the accuracy decreased.
The cylindrical transmitters had their electronics and batteries sealed in epoxy, and produced one pulse per second. Two types of transmitters were used; one, 65 mm long and 16 mm in diameter (weighing 10 g in water), that gave horizontal (two-dimensional) positional fixes (x,y coordinates) and one, 80 mm long and 16 mm in diameter (weighing 12 g in water), that contained a depth sensor and gave three-dimensional positional fixes (x,y,z coordinates).
S
.
Løkkeborg
et
al
.
/
J.
Exp
.
Mar
.
Biol
.
Ecol
.
247
(2000
)
195
–
208
Table 1
Summary of tracking information on individual ling in a fjord in western Norway. Dark bars indicate periods with baits present in the experimental ova
and ingesting the transmitter were videotaped. A more detailed description of the ˚
positioning system and tagging procedure has been provided by (Engas et al., 1996). During the first four days of the experiment five ling were tagged, one of them with a depth transmitter. Their movements and basic behaviour were tracked continuously for 2–6 days. Responses to food odour were then studied by setting a mackerel-baited longline (415 hooks, 1.3 m hook spacing) inside the triangle of hydrophone buoys each night between 18:45 and 20:15 for five days and retrieved the following morning between 08:00 and 09:00. In addition, on two of the days, five mackerel-baited pots (about 100 g bait in each pot) in a chain and a single pot were set (Table 1). A transmitter was attached to the baited gear to determine their positions and provide a fixed reference point. In the period with no gear in the area, a transmitter was placed on the seabed as a reference point in order to indicate the accuracy of the positional fixes.
2.2. Data analysis
area was defined for each individual as the fish were seldom inactive for longer than 10 min at locations outside this area (only on three occasions for longer than 3 h). The positional fixes of the core area were trimmed to compensate for drift of the hydrophone buoys and inaccuracy of the fixes (the 20 most extreme values in each direction were removed), before the size of the area was calculated. This was defined as the area of the rectangle given by the minimum and maximum X- and Y-values.
Using the tracking data of the first part of the experiment, when basic behaviour was studied, diel rhythms in activity were studied by dividing the 24-h cycle into 3-h periods. A Friedman ANOVA was run to compare differences in swimming speed and LRD between times of day. This nonparametric test adjusts for potential individual differences and allows for deviation from normality. It does not, however, take into account possible interaction between time and individual. The test requires that all time periods during a day contain data and therefore only 24-h cycles fulfilling this requirement were included in the analysis. The data were log-transformed before the analysis to stabilize the variance between groups. Diel rhythms in ‘staying within the core area’ were analysed by Friedman ANOVA. An ANOVA was run to test whether the direction of current influenced direction of movement.
A Mann–Whitney U test was performed to compare differences in activity (mean speed per hour and LRD) between nights (from 20:00 to 09:00) with and without baits in the experimental area. As baits were set only early in the night, only data recorded at night were compared as the release rate of feeding attractants is highest shortly after setting (Løkkeborg, 1990).
3. Results
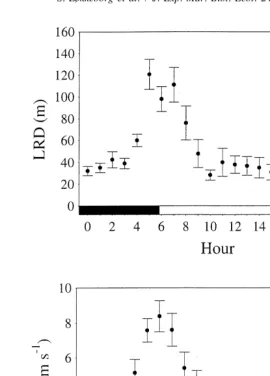
All ling stayed inside or close to the buoy triangle, except for one fish that swam for a few hours beyond the range of the hydrophones. The ling were most active at dawn and dusk, with peak activity between 04:00 and 08:00, and a less clear activity peak from 20:00 to 22:00 (Fig. 2). There were significant differences during the 24-h cycle in mean
swimming speed and LRD (P,0.001). The ling tagged with a depth transmitter
remained near the bottom at depths ranging from 90 to 100 m during both movement and periods of inactivity.
Usually at dawn, but sometimes at dusk, the fish swam around a large area, but with few exceptions they returned to the same area where they spent the day and night. Most of the time when the fish were inactive was spent within this core area, which averaged
62379 m (Table 2). The fish spent 65% (SD54.9) of their time on average within the
core area. The real size of the core area was smaller than the estimate due to the drifting of the buoys, as was indicated by the observation that the static transmitter serving as a
reference point had a core area of 36368 m (when calculated as for the fish). Ling had
thus presumably a very small core area.
There were significant differences in ‘time spent within the core area’ (percentage of
Fig. 2. Longest rectilinear distance between two positions (LRD) and swimming speed per hour (mean6S.E.,
n55) of ling throughout the 24-h cycle. Dark and light sections indicate night and day defined by the times of sunrise and sunset in the middle of the study period.
time being spent outside the core area in the morning (from 05:00 to 08:00, Fig. 3). This coincided with the high-activity period at dawn.
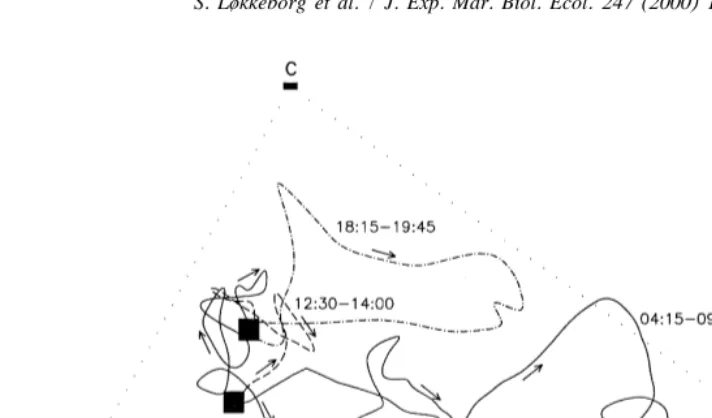
The movement of one ling during a 24-h period is shown in Fig. 4. Its most wide-ranging movements were made at dawn, but it also moved from 12:30 to 14:00 and from 18:15 to 19:45. In the active period at dawn, the fish often moved to a certain area where it swam around for a while before returning to the core area. On three occasions a ling entered the core area of another ling when the holder was present.
Table 2
a
Size of and time spent within the core area
Ling no. Core area Length of sides Time spent
2
(m ) of core area (m) within core area (%)
1 8419 703120 69
2 3835 58367 72
3 2328 53344 62
4 3287 49367 61
5 7526 79395 62
Average 5079 62379 65
a
The core area is the rectangle (x3y) within which the fish stayed when it was inactive.
preferred direction varied between individuals, and there was no correlation between
current direction and movement direction (P.0.05). As the seabed in the area was
relatively flat and uniform, it is unlikely that the ling moved in relation to the topography.
Three of the five tracked ling located baits; two of them made three localisations and one made two. These eight localisations occurred throughout the day, except between 00:30 and 06:30. Ling showed significantly lower swimming speed and smaller LRD during nights (from 20:00 to 09:00) with baits present in the area than during nights with
no baits in the area (P,0.001; Table 3).
Fig. 4. Movement of one ling throughout an entire day (from midnight to midnight). Arrows indicate direction of movement. Times and alternating symbols indicate successive periods of activity and squares indicate the position of the fish during inactive periods. A, B and C indicate the positions of the hydrophone buoys.
4. Discussion
The study was conducted in a natural environment for ling. Potential effects of the tagging procedure were minimised by allowing the ling to voluntarily ingest the transmitter. Ingestion of a transmitter does not seem to decrease feeding motivation in fish (Lucas and Johnstone, 1990; Priede et al., 1990; Løkkeborg, 1998), and the tagging method should thus have ensured that the tracked individuals behaved in the same way as non-tagged fish in the area under study. The species and size of the tagged fish were readily identified, but sex and maturity status are not known. The study might have been conducted in or close to the spawning period for ling in the north-east Atlantic (reported
´ to be in different periods between March and June for different regions; Magnusson et al., 1997), and the largest tagged ling had reached a size at which they might be
´
expected to be mature (60–75 cm; Magnusson et al., 1997). However, several factors indicate that their behaviour was not influenced by spawning. Examination of ling caught in the area showed that fish of similar size to the smallest tagged fish had not yet reached maturity, and small and large tagged fish showed similar behaviour. Further-more, the fish that were tagged remained in shallower water than spawning ling
´
(150–300 m; Magnusson et al., 1997).
activity pattern has been found in several fish species, especially in predators exploiting the transitional nature of the crepuscular periods (Hobson, 1965; Major, 1977; Helfman, 1981; Hobson et al., 1981; Potts, 1990). For ling, the risk of predation should be low throughout the 24-h cycle as they are regarded as top predators (Bergstad, 1991), and the observed activity rhythm probably reflected the advantage of feeding at dawn and dusk. As ling are piscivorous (Bergstad, 1991), the transitional nature of the crepuscular periods may increase the probability of encountering, surprising and capturing prey (see Helfman, 1993). There may be an optimal light level for successfully attacking prey, and a balance between visual detection of the prey and invisibility of the predator may explain the decreased activity at noon. Activity patterns in fishes may also be strongly determined by the activity patterns of their prey (Helfman, 1993). Furthermore, the demand for food and energy might have been fulfilled through the feeding activity at dawn and dusk, leading to low activity and energy-saving behaviour during the rest of
¨ the day. The activity patterns of many fish species vary throughout the year (Muller,
¨
1969; Eriksson, 1978; Ferno et al., 1986; Løkkeborg et al., 1989; Clark and Green, 1990), and studies during other seasons are needed in order to establish whether ling show seasonal changes in activity rhythm.
Ling were found to stay within the area covered by the positioning system, indicating
2
that they occupied a home range much smaller than 1 km (the range of the system under optimal acoustic conditions). They spent most of the time in a core area within this area. A home range is the area within which an animal regularly moves in pursuit of its daily activities, and the core area is the part of the home range that is used more intensively than the rest (Burt, 1943). Home ranges have earlier been described in fish (Reese, 1975; Hawkins et al., 1980; Fish and Savitz, 1983; Clark and Green, 1990; Morrissey and Gruber, 1993; Bradbury et al., 1995). Occupying a home range may offer advantages in terms of knowing where to find shelter and sites with a high probability of encountering prey (Wootton, 1992), and may enable fish to maintain a stable social structure (Sale, 1971).
The fish spent about 65% of their time within the small core area. Less time was spent inside the core area during the period of high activity at dawn. Disproportionate utilization of space within the boundaries of a home range has also been shown for cunner (Tautogolabrus adspersus Walbaum) (Bradbury et al., 1995) and whiting
˚
(Merlangius merlangus L.) (Engas et al., 1996). The home ranges of individual ling overlapped, but the core areas did not and were fairly evenly distributed throughout the area. Staying at one place may partly be associated with feeding, as fish hiding behind stones in a core area may increase the chances of taking prey by surprise during the day when the predator is visible. The movements of the gadoid pollack (Pollachius
pollachius L.) have been shown to cover a restricted part of a small reef (Sarno et al.,
certain location in passive periods for a fish that maintains a home range should be negligible.
There was no correlation between the movement of the fish and the direction of the current in the area. This does not mean that chemically mediated food search does not exist in ling. Ling may normally display a food search pattern independent of the current direction and respond by initiating a rheotactic search when they encounter a food odour source. Such food search behaviour has been observed in cod (Løkkeborg, 1998;
¨
Løkkeborg and Ferno, 1999). Ling were seen to localize baits from a distance, and the fact that baited fishing gear is the most important method of harvesting ling (Bjordal and Løkkeborg, 1996) is a strong indication that chemical stimuli play a role in food search. Although behavioural distortions that do not relate to the natural conditions may arise when the fish comes into contact with the baited gear, the behaviour of fish responding to bait odour is regarded to be similar to the behaviour of a foraging fish as the bait is regarded by the fish as a food item (Løkkeborg, 1994; Bjordal and Løkkeborg, 1996). Localizations of baited gear occurred throughout the day, but no localizations were observed between 00:30 and 06:00 in spite of the fact that the baited gear was set early at night and the concentration of feeding attractants thus should be expected to be high at night (Løkkeborg, 1990). Although the total number of localizations was limited, this indicates that the fish did not react to chemical attractants at night. The explanation for this may be that ling depend on vision in locating or capturing their prey. During the day ling were observed locating baited gear during both active and inactive periods, suggesting that they reacted to food odour during the day whenever the concentration of attractants was high enough to elicit a response, regardless of their state of activity.
Ling swam more slowly and moved within a smaller area when baits were present. Upon detecting attractants the fish can expect to find food in the vicinity and should therefore search within a smaller area to increase the probability of encountering the odour source. In addition to reduced swimming speed within the odour plume of the baits, these findings may also be explained by fish staying in the vicinity of the baits upon encountering the gear. By covering a larger area when no baits were present, the fish will increase the likelihood of encountering odour plumes and thereby food. Area-restricted search behaviour has been observed in species that exploited patchily
¨ distributed food sources (Thomas, 1974; Bell, 1990; Løkkeborg and Ferno, 1999).
To link the spatio–temporal activity functionally with use of the senses in ling, it is of interest to compare the feeding behaviour of ling with that of another gadoid fish. Ling displayed a crepuscular activity cycle, a rhythm often shown by piscivorous species that hunt by vision (Hobson, 1965; Major, 1977; Hobson et al., 1981; Potts, 1990; Helfman, 1993). Ling also responded to chemical stimulation, but the lack of such responses at night supports the idea that this species depends on visual stimuli to capture prey. Studies of cod in a similar habitat to that studied here suggested that they depend less on visual stimuli than do ling. This is indicated by greater activity in periods of reduced light and by the occurrence of olfactory mediated responses during the day and at night
¨
Table 3
Mean swimming speed and mean longest rectilinear distance between two positions (LRD) per hour of ling
a
during nights (between 20:00 and 09:00) when baited gear was present and absent 21
Gear Speed (cm s ) LRD (m)
Absent 6.1 (0.3) 68.8 (4.0)
Present 4.8 (0.3) 58.3 (3.8)
a
Standard errors are given in brackets.
interspecies difference in prey choice, with cod taking a high proportion of stationary prey (Klemetsen, 1982) that may become depleted when feeding is restricted to a small area. In species feeding on stationary prey, chemical stimuli should be expected to be more important. Ling may fulfil their need for food by attacking larger and more mobile prey (Bergstad, 1991) passing through their home range or core area. Likewise, in their behaviour study on the gadiods saithe (Pollachius virens L.) and pollack, Sarno et al. (1994) reported species specific movement pattern and use of habitat, and the observed differences were explained by diversification in feeding strategies. The suggested differences between ling and cod in the relative role of visual and chemical stimuli, prey choice and feeding strategy are supported by morphological differences, with ling having a more elongated body shape and cod a lower mouth terminality compatible with the fact that it is adapted to feed near the bottom (Mattson, 1990).
Acknowledgements
We thank Dr. I. Heuch for help and advice with the statistics. [AU]
References
¨ ¨ ¨ ¨
Alanara, A., Brannas, E., 1997. Diurnal and nocturnal feeding activity in Arctic char (Salvelinus alpinus) and rainbow trout (Oncorhynchus mykiss). Can. J. Fish. Aquat. Sci. 54, 2894–2900.
Atema, J., 1980. Chemical senses, chemical signals and feeding behaviour in fishes. In: Bardach, J.E., Magnuson, J.J., May, R.C., Reinhart, J.M. (Eds.), ICLARM Conf. Proc., Manila, Fish Behaviour and Its Use in the Capture and Culture of Fishes, pp. 57–101.
Bell, W.J., 1990. Searching Behaviour, the Behavioural Ecology of Finding Resources, Chapman and Hall, London.
Bergstad, O.A., 1991. Distribution and trophic ecology of some gadoid fish of the Norwegian Deep. Sarsia 75, 269–313.
˚
Bjordal, A., Løkkeborg, S., 1996. Longlining, Fishing News Books, Oxford.
Bradbury, C., Green, J.M., Bruce-Lockhart, M., 1995. Home ranges of female runner, Tautogolabrus
adspersus (Labridae), as determined by ultrasonic telemetry. Canadian J. Zool. 73, 1268–1279.
Burt, W.H., 1943. Territoriality and home range concepts as applied to mammals. J. Mammal. 24, 346–352. Clark, D.S., Green, J.M., 1990. Activity and movement patterns of juvenile Atlantic cod, Gadus morhua, in
Cote, D., Scruton, D.A., Niezgoda, G.H., McKinley, R.S., Rowsell, D.F., Lindstrom, R.T., Ollerhead, L.M.N., Whitt, C.J., 1998. A coded acoustic telemetry system for high precision monitoring of fish location and movement: application to the study of nearshore nursery habitat of juvenile Atlantic cod (Gadus morhua). Mar. Technol. Soc. J. 32, 54–62.
˚
Engas, A., Løkkeborg, S., Øvredal, J.T., 1996. Natural and fishing-gear-induced behaviour of tagged fish, studied by means of a stationary positioning system. In: Baras, E., Philippart, J.C. (Eds.), Proceedings of the First Conference and Workshop on Fish Telemetry in Europe, Underwater Telemetry, University of Liege, Liege, Belgium, pp. 203–211.
Eriksson, L.O., 1978. Nocturnalism versus diurnalism- dualism within fish individuals. In: Thorpe, J.E. (Ed.), Rhythmic Activity in Fishes, Academic Press, London, pp. 69–89.
¨
Ferno, A., Solemdal, P., Tilseth, S., 1986. Field studies on the behaviour of whiting (Gadus merlangus L.) towards baited hooks. FiskDir. Skr. Ser. HavUnders. 18, 83–95.
Fish, P.A., Savitz, J., 1983. Variations in home ranges of largemouth bass, yellow perch, bluegills, and pumpkinseeds in an Illinois lake. Trans. Am. Fish. Soc. 112, 147–153.
Fraser, N.H.C., Metcalfe, N.B., Thorpe, J.E., 1993. Temperature-dependent switch between diurnal and nocturnal foraging in salmon. Proc. R. Soc. London Ser. B 252, 135–139.
Hawkins, A.D., MacLennan, D.N., Urquhart, G.G., Robb, C., 1974. Tracking cod Gadus morhua L. in a Scottish sea loch. J. Fish Biol. 6, 225–236.
Hawkins, A.D., Urquhart, G.G., Smith, G.W., 1980. Ultrasonic tracking of juvenile cod by means of a large spaced hydrophone array. In: Amlaner, Jr. C.J., MacDonald, D.W. (Eds.), A Handbook of Biotelemetry and Radio Tracking, Pergamon Press, Oxford, pp. 461–470.
Helfman, G.S., 1981. Twilight activities and temporal structure in a freshwater fish community. Can. J. Fish. Aquat. Sci. 38, 1405–1420.
Helfman, G.S., 1993. Fish behaviour by day, night and twilight. In: Pitcher, T.J. (Ed.), Behaviour of Teleost Fishes, Chapman and Hall, London, pp. 479–512.
Hobson, E.S., 1965. Diurnal-nocturnal activity of some inshore fishes in the Gulf of California. Copeia 1965, 291–302.
Hobson, E.S., McFarland, W.N., Chess, J.R., 1981. Crepuscular and nocturnal activities of Californian nearshore fishes, with consideration of their scotopic visual pigments and the photic environment. Fish. Bull. 79, 1–30.
Klemetsen, A., 1982. Food and feeding habits of cod from the Balsfjord, northern Norway during a one-year period. J. Cons. Int. Explor. Mer 40, 101–111.
Løkkeborg, S., 1990. Rate of release of potential feeding attractants from natural and artificial bait. Fish. Res. 8, 253–261.
¨
Løkkeborg, S., 1994. Fish behaviour and longlining. In: Ferno, A., Olsen, S. (Eds.), Marine Fish Behaviour in Capture and Abundance Estimation, Fishing News Books, Oxford, pp. 9–27.
Løkkeborg, S., 1998. Feeding behaviour of cod, Gadus morhua: activity rhythm and chemically mediated food search. Anim. Behav. 56, 371–378.
˚ ¨
Løkkeborg, S., Bjordal, A., Ferno, A., 1989. Responses of cod (Gadus morhua) and haddock (Melanogrammus
aeglefinus) to baited hooks in the natural environment. Can. J. Fish. Aquat. Sci. 46, 1478–1483.
¨
Løkkeborg, S., Ferno, A., 1999. Diel activity pattern and food search behaviour in cod, Gadus morhua. Env. Biol. Fish. 54, 345–353.
Lucas, M.C., Johnstone, A.D.F., 1990. Observations on the retention of intragastric transmitters, and their effects on food consumption, in cod, Gadus morhua L. J. Fish Biol. 37, 647–649.
´ ´
Magnusson, J.V., Bergstad, O.A., Hareide, N.R., Magnusson, J., Reinert, J., 1997. Ling, blue ling and tusk of the northeast Atlantic. In: TemaNord, Vol. 535, Nordic Council of Ministers, Copenhagen, p. 57. Major, P.F., 1977. Predator-prey interactions in schooling fishes during periods of twilight: a study of the
silverside Pranesus insularum in Hawaii. Fish. Bull. 75, 415–426.
Mattson, S., 1990. Food and feeding habits of fish species over a soft sublittoral bottom in the northeast Atlantic. 1. Cod (Gadus morhua L.) (Gadidae). Sarsia 75, 247–260.
Morrissey, J.F., Gruber, S.H., 1993. Home range of juvenile lemon sharks, Negaprion brevirostris. In: Copeia, pp. 425–434.
¨
Pethon, P., 1989. Aschehougs Store Fiskebok, 2nd edition, H. Aschehoug and Co., A / S, Oslo.
Potts, G.W., 1986. Predator-prey interactions between pollack and sand-eels. Prog. Underwater Sci. 11, 69–79. Potts, G.W., 1990. Crepuscular behaviour of marine fishes. In: Herring, P.J., Campbell, A.K., Whitfield, M., Maddock, L. (Eds.), Light and Life in the Sea, Cambridge University Press, Cambridge, pp. 221–227. Priede, I.G., Smith, Jr. K.L., Armstrong, J.D., 1990. Foraging behaviour of abyssal grenadier fish: inferences
from acoustic tagging and tracking in the North Pacific Ocean. Deep-Sea Res. 37, 81–101.
Rae, B.B., Shalton, R.G.J., 1982. Notes on the food on nine species of elasmobranch (Part I) and nine species of demersal teleost (Part II) fishes from Scottish waters. ICES C.M. 1982 / G:56, 5 pp.
Reese, E.S., 1975. A comparative field study of the social behavior and related ecology of reef fishes of the family Chaetodontidae. Z. Tierpsychol. 37, 37–61.
Sale, P.F., 1971. Extremely limited home range in a coral reef fish, Dascyllus aruanus (Pisces: Pomacentridae). In: Copeia, pp. 324–327.
Sarno, B., Glass, C.W., Smith, G.W., 1994. Differences in diet and behaviour of sympatric saithe and pollack in a Scottish sea loch. J. Fish Biol. 45 (Supplement A), 1–11.
˚
Sunnana, K., 1992. Torskefisker. In: Jonsson, B., Semb-Johansson, A. (Eds.), Norges Dyr, Vol. Fiskene 2, J.W. Cappelens Forlag as, Oslo, pp. 85–122.
Svetovidov, A.N., 1986. Gadidae. In: Whitehead, P.J.P., Bauchot, M.L., Hureau, J.C., Nielsen, J., Tortonese, E. (Eds.), Fishes of the Northeastern Atlantic and the Mediterranean, Vol II, Unesco, Paris, pp. 680–710. Thomas, G., 1974. The influence of encountering a food object on the subsequent searching behaviour in
Gasterosteus aculeatus L. Anim. Behav. 22, 941–952.
Thorpe, J.E., 1978. Rhythmic Activity of Fishes, Academic Press, London.
Wilson, R.R., Smith, K.L., 1984. Effect of near-bottom currents on detection of bait by the abyssal grenadier fishes Coryphaenoides spp., recorded in situ with a video camera on a free vehicle. Mar. Biol. 84, 83–91. Wootton, R.J., 1992. Fish Ecology, Chapman and Hall, New York.