Characterization of a new antifungal non-specific lipid transfer
protein (nsLTP) from sugar beet leaves
Anne K. Kristensen
1, Janne Brunstedt, Klaus K. Nielsen
2, Peter Roepstorff
3,
Jorn D. Mikkelsen *
Danisco Biotechnology,Langebrogade 1,P.O. Box17,DK-1001 Copenhagen K, Denmark Received 20 August 1999; received in revised form 27 October 1999; accepted 16 December 1999
Abstract
A novel protein (IWF5) comprising 92 amino acids has been purified from the intercellular washing fluid of sugar beet leaves using cation exchange chromatography and reversed phase high performance liquid chromatography. Based on amino acid sequence homology, including the presence of eight cysteines at conserved positions, the protein can be classified as a member of the plant family of non-specific lipid transfer proteins (nsLTPs). The protein is 47% identical to IWF1, an antifungal nsLTP previously isolated from leaves of sugar beet. A potential site for N-linked glycosylation present in IWF5 (Asn-Xxx-Ser/Thr) was found not to be glycosylated. The amino acid sequence data were used to generate a polymerase chain reaction (PCR) clone, employed for the isolation of a corresponding cDNA clone. According to the cDNA clone, IWF5 is expressed as a preprotein with an N-terminal signal sequence of 26 amino acid residues. The protein shows a strong in vitro antifungal activity against
Cercospora beticola (causal agent of leaf spot disease in sugar beet) and inhibits fungal growth at concentrations below 10 mg ml−1. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Antifungal protein; nsLTP; Mass spectrometry;Beta6ulgaris;Cercospora beticola
www.elsevier.com/locate/plantsci
1. Introduction
In the last decade, many plant proteins with antimicrobial activity in vitro have been identified.
These include chitinases [1], b-1,3-glucanases [2],
chitin-binding proteins [3], thionins [4], plant de-fensins [5] and non-specific lipid transfer proteins (nsLTPs) [6]. nsLTPs are basic, 9 – 10 kDa proteins which are able to bind and transfer a variety of very different lipids between membranes in vitro [6]. The plant nsLTPs are synthesized as precur-sors with N-terminal extensions having the se-quence characteristics of a signal peptide [7] and recent studies have demonstrated that some plant nsLTPs are secreted [8] or bound in cell walls [9]. This extracellular localization of plant nsLTPs makes the originally proposed role in membrane biogenesis and turnover unlikely and it has instead been suggested that they are actively participating in the biosynthesis of the cutin layer and surface wax by transporting acyl monomers [10]. In addi-tion, it has been shown that the expression of nsLTPs can be induced by environmental stress factors, such as heat shock or NaCl treatment [11] and several nsLTP genes in barley are upregulated
Abbre6iations: DTT, dithiotreitol; EDTA, ethylenediaminete-traacetic acid; FPLC, fast protein liquid chromatography; HPLC, high performance liquid chromatography; IC50, concentration
re-quired for 50% growth inhibition; IWF, intercellular washing fluid; MW, molecular weight; nsLTP, non specific lipid transfer protein; PCR, polymerase chain reaction; SDS PAGE, sodium dodecyl poly-acrylamide gel electrophoresis; TFA, trifluoroacetic acid.
* Corresponding author. Tel.: +45-32-662200; fax: + 45-32-662167.
E-mail addresses: [email protected] (A.K. Kristensen), g7jdm@danisco. com (J.D. Mikkelsen)
1Present address: M&E Biotech A/S, Kogle Alle 6, DK-2970
Horsholm, Denmark. Tel.: +45-45-162525; fax: +45-45-162500.
2Present address: DLF-Trifolium, Hoejerupvej 31, P.O. Box 19,
DK-4660 Store Heddinge, Denmark.
3Present address: Department of Molecular Biology, Odense
Uni-versity, Campusvej 55, DK-5230 Odense M, Denmark.
in response to infection by various strains of fun-gal pathogens [12]. These results indicate a possi-ble role in the protection against pathogens, a role which has been confirmed by the finding that nsLTPs isolated from seeds of radish and onion
[13,14], and leaves of barley, maize [15],Arabidop
-sis, spinach [16] and sugar beet [17] displayed
antipathogenic activity in vitro.
A major focus of the laboratory is to investigate
the interaction between sugar beet (Beta 6ulgaris
L.) and the pathogen Cercospora beticola (Sacc.),
the cause of leaf spot disease in sugar beet. Previ-ously, a number of chitinases, glucanases and plant defensin-like proteins have been isolated and their biological role examined [18 – 24].
Recently, the isolation of two nearly identical proteins from sugar beet showing antifungal
activ-ity in vitro against the pathogen C. beticola has
been described [17]. The proteins were isolated from the intercellular washing fluid (IWF) of sugar beet leaves and showed sequence homology to
plant nsLTPs. During infection, C. beticola
re-mains in the extracellular environment [25] and secreted proteins with antifungal activity could be advantageously applied against this specific patho-gen. The isolation and characterization of a new protein, designated IWF5, from sugar beet leaves is reported here. The protein consists of 92 amino acids and shows homology to nsLTPs including the presence of eight conserved cysteine residues and displays strong antifungal activity in vitro
against spore cultures of C. beticola.
2. Materials and methods
2.1. Biological materials and induction of resistance
Plants of sugar beet (B.6ulgarisL., cv. Monova,
Danisco Seed) were grown in growth chambers (at 26°C from 07:30 h and 18°C from 18:30 h with a 54% relative air humidity). After 6 weeks, they were sprayed four times with 25 ppm
2,6-dichloroisonicotinic acid (INA; CGA 41369,
kindly supplied by Dr Theo Staub, Plant Protec-tion Division, Novartis, Basel, Switzerland) at a 2-day interval. The INA, suspended in 0.05% Tween-20, was applied by spraying the adaxial leaf surface (INA acts as a chemical inducer of
resis-tance againstC. beticola [17]) Two days after final
treatment, intercellular washing fluid was isolated as described below.
2.2. Isolation and purification of IWF
The IWF from 5 to 700 g sugar beet leaves was isolated as described previously [17]. Briefly, 5 – 700 g sugar beet leaves were submerged in 20 mM acetic acid, pH 4.5, incubated in vacuo in an exicator (5 min, 4 torr) and infiltrated by release of the vacuum. After air-drying of the leaves, the
IWF was isolated by centrifugation at 1500×gfor
15 min. The IWF was subsequently purified by cation exchange chromatography on a CM-Sep-harose column (Pharmacia LKB, Sweden) pre-equilibrated with starting buffer, 20 mM acetic acid, pH 4.5. Bound proteins were eluted by step-wise increasing the salt concentration in the start-ing buffer: 0.1 M, 0.3 M and 0.5 M NaCl. The 0.3 M NaCl eluate displaying antifungal activity was further purified by cation exchange fast protein liquid chromatography (FPLC) on a Mono S HR
5/5 column (Pharmacia LKB, Sweden)
equili-brated in 20 mM acetic acid, pH 4.5, containing 5% betaine (A-buffer). Proteins were eluted with a linear salt gradient from 0 to 0.3 M NaCl in 30 ml
A-buffer with a flow rate of 1 ml min−1.
2.3. Protein analysis methods
The individual protein peaks from the Mono S column were further purified by reversed-phase high performance liquid chromatography (HPLC)
on a Vydac C4 silica column (The Separations
Group, CA) using 0.1% trifluoroacetic acid (TFA) in water for A-buffer and 0.1% TFA in acetoni-trile for B-buffer as described [17]. Discrete protein peaks were collected manually and freeze-dried. Protein concentrations were determined as described [24]. Peptides were separated on a
Nu-cleosil C18-column (The Separations Group, CA)
using 0.1% TFA as A-buffer and 0.1% TFA in 90% acetonitrile as B-buffer. Peptides were eluted by applying a linear gradient from 5 to 60% B-buffer in 30 min, followed by 4 min of washing with 80% B-buffer using a flow rate of 1 ml min−1.
City, CA). The peptide samples were dissolved in 0.1% TFA or 70% formic acid and spotted onto polybrene coated PVDF membranes (Immobilon, Millipore).
Proteins were separated by SDS-PAGE on the Phast system (Pharmacia LKB), using precast high-density gels or using 16% Tricine (Novex, CA) gels on the ‘Small’ system (Hoefer, CA). Protein bands were fixed and stained with silver
according to Manufacturer’s instructions
(Pharmacia).
Digestion of the protein was performed after modification of the Cys-residues. The freeze-dried
protein was dissolved in 200 ml 0.5 M Tris – HCl,
pH 8.5, 6 M guanidine – HCl, 5 mM EDTA and 5
ml of 1.4 mM DTT added. Reaction took place at
37°C for 10 min, after which 1 ml of
4-vinylpyridine was added. After 10 min, the
reac-tion was stopped by the addireac-tion of 5 ml of 1.4
mM DTT. The protein was subsequently desalted
on a Vydac C4-column and freeze-dried.
Digestion with endoproteinase Lys-C was
con-ducted by dissolving the reduced and alkylated
protein in 5ml 8 M urea. After 30 min at 37°C 45
ml of 50 mM NH4HCO3, pH 7.8 and 0.5mg Lys-C
(from Lysobacter enzymogenes, Boehringer
Mannheim) was added. The digestion took place at 37°C for 8 h.
Digestion with endoproteinase Asp-N (from
Pseudomonas fragi mutant, Boehringer
Mannheim) was performed using the same buffer and reaction conditions.
The sequence of IWF5 was analyzed for similar-ity using the BLAST program (version 2.0.5, May 1998) and aligned with similar sequences using the Multialign program (version 5.3.3) [26].
2.4. Mass spectrometry
The purity of each individual fraction was ver-ified by matrix-assisted laser desorption ionization mass spectrometry [28] before further analysis. Mass spectrometry was performed using either a Voyager DE instrument (PerSeptive Biosystems, Framingham, MA) in the linear mode using de-layed extraction [29], or a Bruker reflex TOF (Bruker-Franzen, Bremen, Germany). All samples were analyzed in the positive mode using an accel-eration voltage of 20 kV and external calibration, providing a mass accuracy of 0.1% or better. The
matrix used wasa-cyano 4-hydroxy cinnamic acid
(Sigma, St. Louis, MO) at a concentration of 15
mg ml−1in 70% CH
3CN, 0.1% TFA [30]. Spectra
were accumulated for an average of at least 100 laser shots.
2.5. Antifungal assay
Protein fractions were tested for their in vitro
inhibitory effects on the growth of C. beticola
spore cultures using the microtiter plate bioassay described previously [20]. Briefly, the growth of the submerged spore cultures was followed by measuring the increase in absorbance at 620 nm and by microscopical analysis.
2.6. cDNA cloning
The cDNA sequence of IWF5 was obtained by
3%and 5%RACE essentially as described previously
[17] using the following primers.
3% RACE Primers.
QT:
5%-CCAGTGAGCAGAGTGACGAGGACTCGA
GCTCAAGC(T)17-3%
Q0: 5%-CCAGTGAGCAGAGTGACG-3%
Q1: GAGGACTCGAGCTCAAGC-3%
5% RACE primers.
5%-Anchor:
5%-GGCCACGCGTCGACTAGTACGGGGGGG
GGG-3%
5%-UNI: 5%-GGCCACGCGTCGACTAGTACG
2.7. 3% RACE
The amino acid sequence of the IWF5 protein was used to construct two degenerated oligonucle-otide primers for the isolation of a partial cDNA
clone by 3%RACE. Total RNA was purified from
leaves of sugar beet (cv. Monova) 6 days after
inoculation with C.beticola according to Ref. [27].
Reverse transcription followed by PCR were done with the RT-PCR kit from Perkin Elmer (Applied Biosystems, CA) and according to their protocol.
Briefly, 1 mg of total RNA and 2.5 pmol QT
PCR 40 pmol of the primer Q0 was used as
downstream primer and the upstream primer was
150 pmol of the degenerated primer P1 (5%
- GC(ACGT)TG(CT)(AC)G(ACGT)TG(CT)ATG-AA: position 315 – 331 in the IWF5 cDNA se-quence). In the second nested PCR 50 pmol of the
primer Q1was used as downstream primer and the
upstream primer was 50 pmol of the degenerated
primer P2 (5%
-GG(ACGT)AT(ACT)AA(CT)-CA(CT)AA(GA)TA: position 354 – 370 in the IWF5 cDNA sequence). The PCR conditions were: 1 min at 94°C, 2 min at 42°C, 1 min at 50°C and 5 min at 72°C for 1 cycle followed by 1 min at 94°C and 2 min at 42°C and 3 min at 72°C for 35 cycles followed by 10 min at 72°C. After the second PCR a single DNA product of 390 bp was obtained. The DNA product was cloned into the pT7Blue vector (Novagen, Madison, WI) and se-quenced using a termo sequenase fluorescent cycle sequencing kit (Amersham Pharmacia Biotech, Uppsala, Sweden) and an ALF DNA sequencer (Amersham Pharmacia Biotech, Uppsala, Swe-den).
2.8. 5% RACE
The sequence of the 5%end of IWF5 cDNA was
obtained by 5% RACE using the 5% RACE system
from Gibco BRL (MD, USA) with 3 gene specific primers constructed from the partial cDNA
se-quence obtained by 3%RACE. Briefly, 1 mg of the
same total RNA as used for 3% RACE and 2.5
pmol of a gene specific primer GSP5-1 (5%
-TG-GAATTGGAGATTATGTAAG: position 619 – 643 in the IWF5 cDNA sequence) was incubated at 70°C for 10 min followed by the addition of reverse transcriptase and incubating at 42°C for 30 min, 70°C for 15 min and the addition of RNaseH and incubating further 10 min at 55°C. The cDNA was dC-tailed according to the protocol of Gibco BRL. The tailed cDNA was subjected to two rounds of PCR. In the first PCR 20 pmol of the
5%-Anchor primer was used as upstream primer
and the downstream primer was 20 pmol of the
gene specific primer GSP5-2 (5%
-TCACTTTA-GATGTAAGAAGCACACATG: position 596 – 622 in the IWF5 cDNA sequence). In the second
PCR 50 pmol of the 5%-UNI primer was used as
upstream primer and the downstream primer was
50 pmol of the gene specific primer GSP5-3 (5%
-TAAGCAGAAAGTTCCAGAAAGCATG:
posi-tion 548 – 572 in the IWF5 cDNA sequence). The condition for the first PCR was: 1 min at 94°C and 1 min at 51°C and 2 min at 72°C for 35 cycles followed by 10 min at 72°C. The condition for the second PCR was: 1 min at 94°C and 1 min at 55°C and 2 min at 72°C for 35 cycles followed by 10 min at 72°C. The single 600 bp DNA product was cloned into the pT7Blue vector (Novagen) and sequenced using a Termo Sequenase fluorescent cycle sequencing kit (Amersham) and an ALF DNA sequencer (Pharmacia).
2.9. Southern hybridization
Genomic DNA was isolated from sugar beet leaves according to Ref. [31], digested with appro-priate restriction enzymes and separated on a 0.8% agarose gel. A DNA probe of IWF5 cDNA was
32P-labeled by random priming using the
Ready-To-Go Labeling Kit from Pharmacia. Southern transfer and hybridization was performed
accord-ing to [32] usaccord-ing Hybond N+ membranes
(Amer-sham) following the recommendations of the Manufacturer.
3. Results
3.1. Purification and characterization of IWF5
The IWF isolated from sugar beet leaves was subjected to cation exchange chromatography and bound proteins eluted by stepwise increasing the salt concentration in the starting buffer (0.1, 0.3 and 0.5 M NaCl). The 0.3 M NaCl eluate dis-played strong antifungal activity and was further purified on a Mono S FPLC column using a salt gradient. Five major and a number of smaller protein peaks were eluted (data not shown). One of the major protein peaks, showing antifungal activity, separated into five major peaks and a number of minor peaks when subjected to rpH-PLC (Fig. 1). Mass spectrometric analysis of the individual fractions revealed the presence of two previously identified proteins in fraction 2 (AX2) and 3 (IWF2) (Table 1), while fraction 4 contained a protein with a MW of 9319.07, in the following named IWF5 (Fig. 2). In Fig. 2, both the doubly
charged (MH22+) molecular ion at m/z 4660.98
9319.07 are observed as well as the molecular ion
of the dimer (M2H+) atm/z18634.1. In fraction 5,
a number of smaller proteins were present, none of which could be identified on basis of their molecu-lar weight. Analysis by SDS-PAGE of fraction 4 using a high-density gel, revealed that this fraction contained a protein with an apparent MW of ca. 9 kDa as well as a minor component with a MW of ca. 8.5 kDa (Fig. 3). The fraction was further analyzed by tricine SDS-PAGE performed with and without reducing agent and silver stained (data not shown), and no shift in MW was ob-served when adding the reducing agent. Thus, the protein exists as a monomer. The N-terminal se-quence of fraction 4 was determined after
reduc-tion and carboxymethylareduc-tion and the first 38 Fig. 2. Mass spectrometric analysis of fraction 4 from thehigh performance liquid chromatography (HPLC) purifica-tion shown in Fig. 1. In addipurifica-tion to the molecular ion atm/z
9319.07, the doubly charged molecular ion at m/z 4660.98 and the molecular ion of the dimer at m/z 18634.1 are observed.
Fig. 1. High performance liquid chromatography (HPLC) elution profile of one of the major fractions from the CM-Sepharose purification. IWF5 was present in peak 4. Proteins were eluted with a linear gradient from 5 to 80% of the B-buffer as indicated (dotted line). The flow rate was 1 ml min−1.
Fig. 3. Sodium dodecyl polyacrylamide gel electrophoresis (SDS-PAGE) analysis of the purified sugar beet non specific lipid-transfer protein using high-density gels (Pharmacia). Molecular mass markers were CNBr-derived myoglobin frag-ments.
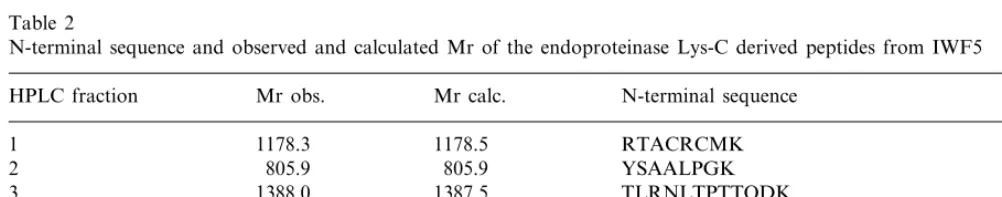
residues were determined (Fig. 6, sequence in italic). The complete sequence was obtained by cleavage of the reduced and alkylated protein with
endoproteinase Lys-C, resulting in six peptides
(Fig. 6, peptides derived from cleavage at Lys-residues (bold) are underlined). All peptides were completely sequenced after mass spectrometric analysis. For all peptides, the determined sequence could be verified by the observed mass (Table 2). Alignment of the cysteines present in the IWF5-derived peptides with those of IWF1 (a nsLTP previously isolated from sugar beet), made it pos-sible to deduce the entire sequence of IWF5 from this single digest (Fig. 6). The deduced sequence of IWF5 was confirmed by mass analysis of overlap-ping peptides generated by a second digestion with
endoproteinase Asp-N (data not shown).
Table 1
Mass spectrometric analysis of the different fractions (Fig. 1) from the rpHPLC purification of the active fraction from the fast protein liquid chromatography (FPLC) cation exchange chromatography of the fraction eluting at 0.3 M NaCl from a crude cation exchange purification stepa
Mr obs.
Fraction Mr calc. Protein
4015.6 N.I.
1 N.I.
5178.8
2 5176.9 AX2b
8937.7
3 8948.3 IWF2c
9319.1
4 9317.7 IWF5
N.I.
5 2–4000 N.I.
aMr obs. and Mr calc. are the observed and calculated
molecular masses of the fractions. N.I. not identified.
The MWs determined by SDS-PAGE and mass spectrometry (9319.1) are in good agreement with that deduced from the amino acid sequence data (9317.7). In summary, IWF5 is a 92 amino acid protein, eight of which are cysteines. A potential
site for N-linked glycosylation (Asn-Xxx-Ser/Thr)
at position 37 – 39 was found not to be glycosy-lated as verified by the good agreement between the measured molecular mass and that deduced from the sequence.
3.2. Characterization of the IWF5 cDNA
The nucleotide sequence of the full-length IWF5
cDNA clone, obtained by a combination of 3%and
5%RACE, and the deduced amino acid sequence is
shown in Fig. 4. The IWF5 cDNA contains 722 nucleotides including a poly(A) tail of 20 nucle-otides. The sequence contains an open reading frame encoding 118 amino acid residues with the
first in frame ATG at position 96 and a stop codon at position 450. In comparison with the amino acid sequence of IWF5, the cDNA seems to encode a precursor protein with an N-terminal signal sequence of 26 amino acid residues followed by a functional domain of 92 amino acid residues, identical to the amino acid sequence of the iso-lated IWF5.
3.3. Antifungal acti6ity
The in vitro antifungal activity of IWF5 was
tested against spore cultures of C. beticola grown
in liquid growth medium in microtiter plates (Fig. 5). The protein showed a strong growth inhibiting
effect against C. beticola and the antifungal
activ-ity increased with increasing protein
concentra-tions. In control cultures, growth started
approximately 30 h after incubation start (Fig. 5, triangles), while the addition of IWF5 caused a
Table 2
N-terminal sequence and observed and calculated Mr of the endoproteinase Lys-C derived peptides from IWF5
HPLC fraction Mr obs. Mr calc. N-terminal sequence
1 1178.3 1178.5 RTACRCMK
2 805.9 805.9 YSAALPGK
1388.0
3 1387.5 TLRNLTPTTQDK
4 1164.9 1165.3 SAASAIPGINHK
CGVSIPGPVGPQAD
2075.3 2075.4
5
3644.3 AINCGLVSQSLAACLGFLENGQPNAACCNGVK
3647.0 6
Fig. 5. Antifungal activity of IWF5 against spore cultures of
C. beticola tested in the microtiter plate assay. The fungal growth was measured as increase in absorbance at 620 nm. Two different concentrations of IWF5 were used in the wells: 16mg ml−1(circles) and 40mg ml−1(squares). Growth of the
control culture is indicated by the triangles.
Microscopical analysis of spore cultures of C.
beticola grown in microtiter plates in the presence
of 16 mg ml−1 IWF5 shows that the hyphae were
shorter and less branched as compared to control culture (data not shown).
3.4. Homology to non-specific lipid transfer proteins
The amino acid sequence of IWF5 shows ho-mology to the family of nsLTPs. In Fig. 6 the sequence of IWF5 is compared to the seven plant nsLTPs to which the highest degree of similarity was found. The sequences were aligned [26] and gaps introduced to optimize the alignment. IWF5 shows 49% identity to an nsLTP isolated from spinach [33] and is 47% identical to IWF1 (Fig. 6). IWF1 has previously been isolated from sugar beet leaves [17] and comprises part of a small gene family in sugar beet. When the amino acid se-quences of nsLTPs from different plant species are compared, only few residues are conserved [34]. Among these are the eight cysteines and 12 posi-tions occupied by hydrophobic or aromatic residues [35]. The amino acid sequence of IWF5 corresponds largely to this sequence homology including the presence of eight cysteines at con-served positions (at positions 4, 14, 28, 29, 49, 51, 75 and 89, numbering as in Fig. 6). However, two of the positions normally occupied by aromatic or hydrophobic residues, have been exchanged with a threonine at position 39 and a glycine at position 81.
concentration dependent delay in growth start
(Fig. 5, circles and squares). At 16 mg ml−1, the
growth was markedly delayed and strongly inhib-ited (Fig. 5, filled circles).
At 40 mg ml−1 no fungal growth could be
detected even at 160 h after assay start (Fig. 5, filled squares). The concentration required for 50%
growth inhibition (IC50) after 100 – 120 h
incuba-tion time was 2 – 4 mg ml−1. The amount of
protein needed for 50% growth inhibition is simi-lar to the level found for the antifungal proteins IWF1 and IWF2 [17].
3.5. Southern hybridization
Genomic DNA from sugar beet was digested
by the restriction enzymes BamHI or XbaI and
subjected to Southern hybridization analysis us-ing IWF5 cDNA as probe. Each restriction en-zyme digest resulted in only one hybridizing band (using stringent conditions), indicating that only one gene (or a small number of genes) for IWF5 is present in the genome of sugar beet.
4. Discussion
The IWF isolated from sugar beet leaves con-tained several proteins with antifungal activity, some of which have already been characterized [17,24]. A novel protein of 9319.07 Da was iden-tified and purified to homogeneity. The protein, designated IWF5, comprises 92 amino acids, eight of which are cysteines. The amino acid se-quence of IWF5 is partially homologous to the sequence of other non-specific lipid transfer proteins, including the presence of eight con-served cysteines and ten aromatic or hydropho-bic residues. The three-dimensional structure has been determined for LTPs isolated from wheat [36], barley [37] and maize seeds [38,39]. The overall structure consists of four helical frag-ments connected by three loops and a C-terminal fragment forming a saxophone-like shape [36]. Hydrophobic interactions and the four disulfide bridges play important roles in maintaining the tertiary structure of the protein [38]. Interest-ingly, the protein has a tunnel-like hydrophobic cavity which could be a potential binding site for lipids [36,38]. IWF5 shows 49% identity to a nsLTP isolated from spinach [34] and is 47% identical to IWF1 (Fig. 6). IWF1 has previously been isolated from sugar beet leaves [17] and comprises part of a small gene family in sugar beet. The hybridization studies using southern blotting resulted in only one band, indicating that only one gene (or a small number of genes) for IWF5 is present in the genome of sugar beet. However, it can not be ruled out that several genes are present as tandem repeats since the hybridizing fragments are relatively large.
IWF5 exists as a monomer both in its reduced and unreduced state, this was also found for the antifungal nsLTPs IWF1 and IWF2 [17]. In
comparison, an antifungal protein isolated from radish was found to exist as a dimer in its unre-duced form [13].
The amino acid sequence data were used to obtain a cDNA clone. The cDNA sequence of IWF5 indicates that the protein is expressed with a signal peptide of 26 amino acid residues. Most of the nsLTPs isolated so far are synthesized as preproteins with putative signal peptides which is in accordance with their extracellular localiza-tion.
Many of the nsLTPs have antimicrobial activ-ity when tested in vitro against a number of
mi-croorganisms. IWF5 demonstrated a strong
inhibitory effect (IC50 2 – 4 mg ml−1) when tested
in a bioassay against C. beticola. In comparison,
an nsLTP from onion seed is extremely active
against a broad range of fungi with IC50 values
from 1 to 6 mg ml−1, an nsLTP from radish
seed is moderately active (IC50 value from 7 to
100 mg ml−1) and an nsLTP from maize and
wheat seed is inactive to most fungi [14]. In comparison with these proteins, IWF5 has a strong antifungal activity against the pathogen C. beticola. The biological role of nsLTPs is still unknown, and although many of the nsLTPs isolated so far are able to transfer lipids in vitro, this activity is not shared by all members of the nsLTP family [14]. Many of the nsLTPs have antimicrobial activities when tested in vitro, thus indicating a possible role in plant defense [14]. However, as earlier suggested [13], the different
functions are not mutually exclusive. The
nsLTPs could play their defensive role after be-ing deposited in the cell wall with the trans-ported cutin precursors [13].
So far, five proteins with in vitro antifungal activity have been isolated from leaves of sugar beet. Two of these proteins belong to the family of plant defensins (AX1 and AX2) [24], one to the chitin-binding peptides (IWF4) [40] and two are members of the nsLTPs (IWF1 and IWF2) [17]. IWF5 is the third nsLTP isolated from sugar beet and demonstrates that a number of
proteins are present belonging to different
families. In combination, these proteins could act to delay the invading fungal hyphae and thus giving the plant time to upregulate other defense mechanisms. The strong antifungal potency of
IWF5 against C. beticola makes it a good
Acknowledgements
We wish to thank Jette Skovsgaard, Susanne Dreboldt and Lene D. Hansen for excellent techni-cal assistance. The work was supported by Novar-tis Seeds (Stanton, USA) and by a grant EF590 from the Danish Academy of Technical Sciences (ATV).
References
[1] D.B. Collinge, K.M. Kragh, J.D. Mikkelsen, K.K. Nielsen, U. Rasmussen, K. Vad, Plant chitinases, Plant J. 3 (1993) 31 – 40.
[2] F. Mauch, B. Mauch-Mani, T. Boller, Antifungal hydro-lases in pea tissue II. Inhibition of fungal growth by combinations of chitinases and b-1,3-glucanases, Plant Physiol. 88 (1988) 936 – 942.
[3] K. De Bolle, K.M.M. David, S.B. Rees, J. Vanderleyden, B.P.A. Cammue, W.F. Broekaert, Cloning and charac-terization of a cDNA encoding an antimicrobial protein from amaranth, Amaranthus caudatus, Plant Mol. Biol. 22 (1993) 1187 – 1190.
[4] H. Bohlmann, S. clausen, S. Behnke, H. Giese, C. Hiller, U. Reimann-Philipp, G. Schrader, V. Barkholt, K. Apel, Leaf-specific thionins of barley — a novel class of cell wall proteins toxic to plant-pathogenic fungi and possi-bly involved in the defense mechanism of plants, EMBO J. 7 (1988) 1559 – 1565.
[5] F.R.G. Terras, K. Eggermont, V. Kovaleva, N.V. Raikhel, R.W. Osborn, A. Kester, S. Rees, S. Torrekens, F. Vanleuven, J. Vanderleyden, B.P.A. Cammue, W.F. Broekaert, Small cysteine-rich antifungal proteins from radish — their role in host defense, Plant Cell 7 (1995) 573 – 588.
[6] J.-C. Kader, Lipid-transfer proteins in plants, Annu. Rev. Plant Physiol. Plant Mol. Biol. 47 (1996) 627 – 654. [7] W.R. Bernhard, S. Thoma, J. Botella, C.R. Somerville, Isolation of a cDNA clone for spinach lipid transfer protein and evidence that the protein is synthesized by the secretory pathway, Plant Physiol. 95 (1991) 164 – 170. [8] P. Sterk, H. Booji, G.A. Schellekens, A. van Kammen, S.C. De Vries, Cell specific expression of the carrot EP2 lipid transfer protein gene, Plant Cell 3 (1991) 907 – 921. [9] S. Thoma, U. Hecht, A. Kippers, J. Botella, S. De Vries, C. Somerville, Tissue-specific expression of a gene encod-ing a cell wall-localized lipid transfer protein fromAra
-bidopsis, Plant Physiol. 105 (1994) 35 – 45.
[10] Hendriks, M., Meijer, E.A., Thoma, S., Kader, J.-C., De Vries, S.C., 1994. The carrot extracellular lipid transfer protein EP2: Quantitative aspects with respect to its putative role in cutin synthesis, In: G. Coruzzi and P. Puigdome`nech (Ed.), Plant Molecular Biology, Springer-Verlag, Berlin, NATO ASI Series H 81, 1994, pp. 85 – 94. [11] S. Torres-Schumann, J.A. Godoy, J.A. Pintor-Toro, A probable lipid transfer protein gene is induced by NaCl in stems of tomato plants, Plant Mol. Biol. 18 (1992) 749 – 757.
[12] F. Garcı´a-Olmedo, A. Molina, A. Segura, M. Moreno, The defensive role of non specific lipid-transfer proteins in plants, Trends Microbiol. 3 (1995) 72 – 74.
[13] F.R.G. Terras, I.J. Goderis, F. Van Leuven, J. Vander-leyden, B.P.A. Cammue, W.F. Broekaert, In vitro anti-fungal activity of a radish (Raphanus sati6us L.) seed protein homologous to non specific lipid transfer proteins, Plant Physiol. 100 (1992) 1055 – 1058.
[14] B.P.A. Cammue, K. Thevissen, M. Hendriks, K. Egger-mont, I.J. Goderis, P. Proost, J. Van Damme, R.W. Osborn, F. Guerbette, J.-C. Kader, W.F. Broekaert, A potent antimicrobial protein from onion seeds showing sequence homology to plant lipid transfer proteins, Plant Physiol. 109 (1995) 445 – 455.
[15] A. Molina, A. Segura, F. Garcı´a-Olmedo, Lipid transfer proteins (nsLTPs) from barley and maize leaves are potent inhibitors of bacterial and fungal plant pathogens, FEBS Lett. 316 (1993) 119 – 122.
[16] A. Segura, M. Moreno, F. Garcı´a-Olmedo, Purification and antipathogenic activity of lipid transfer proteins (LTPs) from leaves of Arabidopsis and spinach, FEBS Lett. 332 (1993) 243 – 246.
[17] K.K. Nielsen, J.E. Nielsen, S.M. Madrid, J.D. Mikkelsen, New antifungal proteins from sugar beet (Beta6ulgarisL.) showing homology to non-specific lipid transfer proteins, Plant Mol. Biol. 31 (1996) 539 – 552. [18] L. Berglund, J. Brunstedt, K.K. Nielsen, Z. Chen, J.D.
Mikkelsen, K.A. Marcker, A proline-rich chitinase from
Beta6ulgaris, Plant Mol. Biol. 27 (1995) 211 – 216. [19] J.D. Mikkelsen, L. Berglund, K.K. Nielsen, H.
Chris-tiansen, K. Bojsen, Structure of endochitinase genes from sugar beets, in: C. Brine, S.A. Sandford, J.P. Zikakis (Eds.), Advances in Chitin and Chitosan, Elsevier Ap-plied Science, New York, 1992, pp. 344 – 353.
[20] K.K. Nielsen, P. Jørgensen, J.D. Mikkelsen, Antifungal activity of sugar beet chitinase against Cercospora beti
-cola: an autoradiographic study on cell wall degradation, Plant Path. 43 (1994) 979 – 986.
[21] K.K. Nielsen, K. Bojsen, P. Roepstorff, J.D. Mikkelsen, A hydroxyproline containing class IV chitinase of sugar beet is glycosylated with xylose, Plant Mol. Biol. 25 (1994) 241 – 257.
[22] K.K. Nielsen, J.D. Mikkelsen, K.M. Kragh, K. Bojsen, An acidic class III chitinase in sugar beet: induction by
Cercospora beticola, characterization and expression in transgenic tobacco plants, Mol. Plant-Microbe Interact. 6 (1993) 495 – 506.
[23] A. Susi, J.D. Mikkelsen, K. Von Weissenberg, K.K. Nielsen, Sugar beet chitinase inhibits the growth of a spruce pathogen, Eur. J. Forest Path. 25 (1995) 61 – 64. [24] K.M. Kragh, J.E. Nielsen, K.K. Nielsen, S. Dreboldt,
J.D. Mikkelsen, Characterization and localization of new antifungal cysteine-rich proteins from Beta 6ulgaris L., Mol. Plant-Microbe Interact. 8 (1995) 424 – 434. [25] M.P. Steinkamp, S.S. Martin, L.L. Hoefert, E.G.
Rup-pel, Ultrastructure of lesions produced by Cercospora beticolain leaves ofBeta6ulgaris, Physiol. Plant Path. 15 (1979) 13 – 26.
[27] D.B. Collinge, D.E. Milligan, J. Maxwell Dow, G. Scofield, M.J. Daniels, Gene expression in Brassica campestris showing a hypersensitive response to the in-compatible pathogenXanthomonas campestris p6.6itians, Plant Mol. Biol. 8 (1987) 405 – 414.
[28] M. Mann, G. Talbo, Developments in matrix-assisted laser desorption/ionization peptide mass spectrometry, Curr. Opin. Biotechnol. 7 (1996) 11 – 19.
[29] M. Vestal, P. Juhasz, S.A. Martin, Delayed extraction matrix-assisted laser desorption time-of-flight mass spec-trometry, Rapid Commun. Mass Spectrom. 9 (1995) 1044 – 1050.
[30] R. Beavis, T. Chaudhary, B.T. Chait,a -cyano-4-hydrox-ycinnamic acid as a matrix for matrix assisted laser desorption mass spectrometry, Org. Mass Spectrom. 27 (1992) 156 – 158.
[31] S.L. Dellaporta, J. Wood, J.B. Hincks, A plant DNA minipreparation: Version II, Plant Mol. Biol. Rep. 1 (1983) 19 – 21.
[32] T. Maniatis, E.F. Fritsch, J. Sambrook, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, New York, 1982.
[33] P. Bouillon, C. Drischel, H. Duranton, J.-C. Kader, The primary structure of spinach-leaf phospholipid-transfer protein, Eur. J. Biochem. 166 (1987) 387 – 391.
[34] P. Coutos-Thevenot, T. Jouenne, O. Maes, F. Guerbette, M. Grosbios, J.P. Le Caer, M. Boulay, A. Deloire, J.-C. Kader, J. Guern, Four 9-kDa proteins excreted by
so-matic embryos of grapevine are isoforms of lipid-transfer proteins, Eur. J. Biochem. 217 (1993) 885 – 889.
[35] W.F. Broekaert, B.P.A. Cammue, M. De Bolle, K. Thevissen, G.W. De Samblanx, R.W. Osborn, Antimi-crobial peptides from plants, Crit. Rev. Plant Sci. 16 (1997) 297 – 323.
[36] E. Gincel, J.-P. Simorre, A. Caille, D. Marion, M. Ptak, F. Vovelle, Three-dimensional structure in solution of a wheat lipid-transfer protein from multidimensional 1
H-NMR data, Eur. J. Biochem. 226 (1994) 413 – 422. [37] M. Lerche, B.B. Kragelund, L.M. Bech, F.M. Poulsen,
Barley lipid-transfer protein complexed with palmitoyl CoA: the structure reveals a hydrophobic binding site that can expand to fit both large and small lipid-like ligands, Structure 5 (1997) 291 – 306.
[38] D.H. Shin, J.Y. Lee, K.Y. Hwang, K.K. Kim, S.W. Suh, High-resolution crystal structure of the non-specific lipid transfer protein from maize seedlings, Structure 3 (1995) 189 – 199.
[39] J. Gomar, M.-C. Petit, P. Sodano, D. Sy, D. Marion, J.-C. Kader, F. Vovelle, M. Ptak, Solution structure and lipid binding of a non specific lipid transfer protein extracted from maize seeds, Prot. Sci. 5 (1996) 565 – 577. [40] K.K. Nielsen, J.E. Nielsen, S.M. Madrid, J.D. Mikkelsen, Characterization of a new antifungal chitin-binding peptide from sugar beet leaves, Plant Physiol. 113 (1997) 83 – 91.

![Fig. 6. Comparison of the amino acid sequences of antifungal sugar beet proteins IWF5, IWF1 [16], an non-specific lipid transferprotein (nsLTP) from spinach [30] and five nsLTPs isolated from rice (or) and almond (pr)](https://thumb-ap.123doks.com/thumbv2/123dok/1034177.928775/7.612.43.255.47.212/comparison-sequences-antifungal-proteins-specic-transferprotein-spinach-isolated.webp)
