HASIL DAN PEMBAHASAN
Isolasi Enterobacter sakazakii dari Susu Formula dan Makanan Bayi
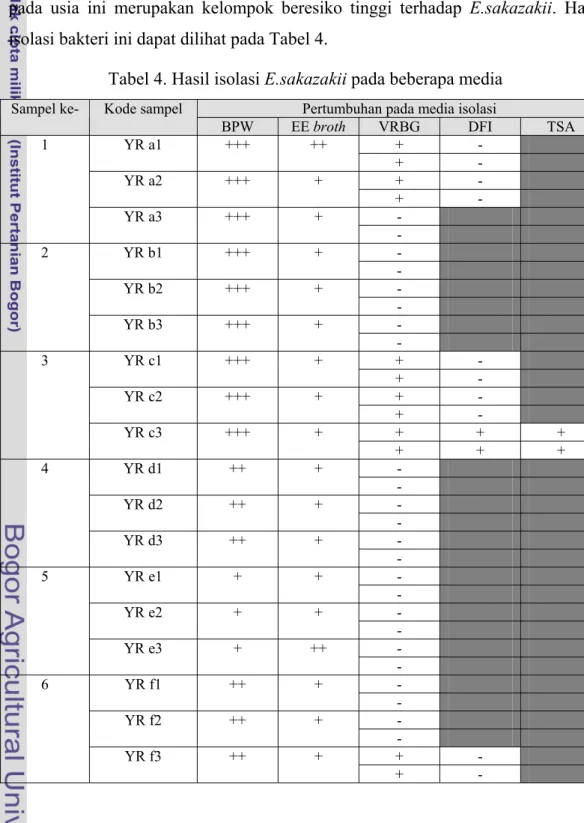
Isolasi E.sakazakii dilakukan dari beberapa sampel susu formula dan makanan bayi yang ditujukan untuk bayi yang berusia di bawah 6 bulan. Penggunaan sampel yang ditujukan untuk bayi di bawah 6 bulan dikarenakan bayi pada usia ini merupakan kelompok beresiko tinggi terhadap E.sakazakii. Hasil isolasi bakteri ini dapat dilihat pada Tabel 4.
Tabel 4. Hasil isolasi E.sakazakii pada beberapa media Pertumbuhan pada media isolasi Sampel ke- Kode sampel
BPW EE broth VRBG DFI TSA + - YR a1 +++ ++ + - + - YR a2 +++ + + - - 1 YR a3 +++ + - - YR b1 +++ + - - YR b2 +++ + - - 2 YR b3 +++ + - + - YR c1 +++ + + - + - YR c2 +++ + + - + + + 3 YR c3 +++ + + + + - YR d1 ++ + - - YR d2 ++ + - - 4 YR d3 ++ + - - YR e1 + + - - YR e2 + + - - 5 YR e3 + ++ - - YR f1 ++ + - - YR f2 ++ + - + - 6 YR f3 ++ + + -
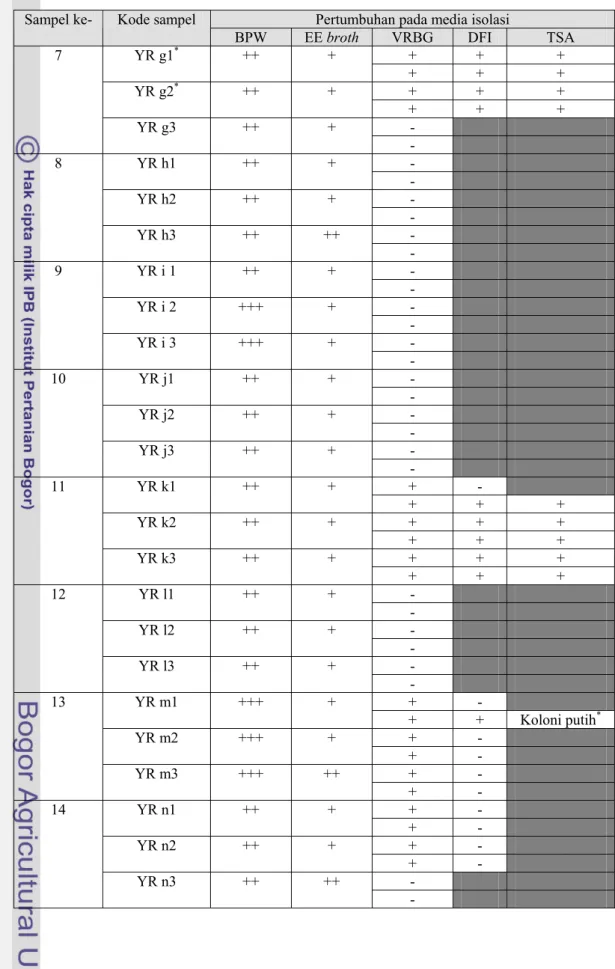
Tabel 4. Hasil isolasi E.sakazakii pada beberapa media (lanjutan)
Pertumbuhan pada media isolasi Sampel ke- Kode sampel
BPW EE broth VRBG DFI TSA + + + YR g1* ++ + + + + + + + YR g2* ++ + + + + - 7 YR g3 ++ + - - YR h1 ++ + - - YR h2 ++ + - - 8 YR h3 ++ ++ - - YR i 1 ++ + - - YR i 2 +++ + - - 9 YR i 3 +++ + - - YR j1 ++ + - - YR j2 ++ + - - 10 YR j3 ++ + - + - YR k1 ++ + + + + + + + YR k2 ++ + + + + + + + 11 YR k3 ++ + + + + - YR l1 ++ + - - YR l2 ++ + - - 12 YR l3 ++ + - + - YR m1 +++ + + + Koloni putih* + - YR m2 +++ + + - + - 13 YR m3 +++ ++ + - + - YR n1 ++ + + - + - YR n2 ++ + + - - 14 YR n3 ++ ++ -
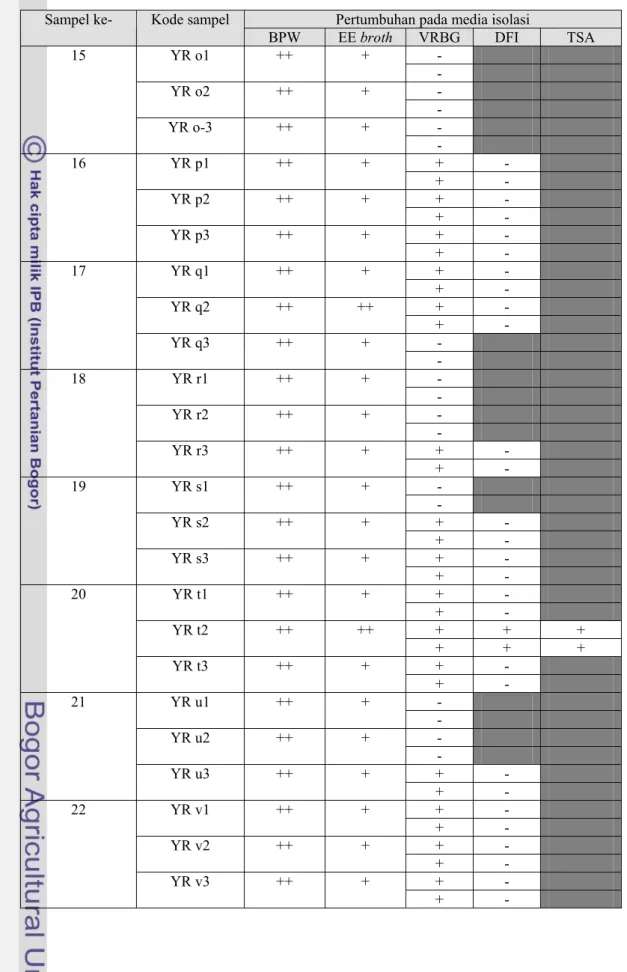
Tabel 4. Hasil isolasi E.sakazakii pada beberapa media (lanjutan)
Pertumbuhan pada media isolasi Sampel ke- Kode sampel
BPW EE broth VRBG DFI TSA - YR o1 ++ + - - YR o2 ++ + - - 15 YR o-3 ++ + - + - YR p1 ++ + + - + - YR p2 ++ + + - + - 16 YR p3 ++ + + - + - YR q1 ++ + + - + - YR q2 ++ ++ + - - 17 YR q3 ++ + - - YR r1 ++ + - - YR r2 ++ + - + - 18 YR r3 ++ + + - - YR s1 ++ + - + - YR s2 ++ + + - + - 19 YR s3 ++ + + - + - YR t1 ++ + + - + + + YR t2 ++ ++ + + + + - 20 YR t3 ++ + + - - YR u1 ++ + - - YR u2 ++ + - + - 21 YR u3 ++ + + - + - YR v1 ++ + + - + - YR v2 ++ + + - + - 22 YR v3 ++ + + -
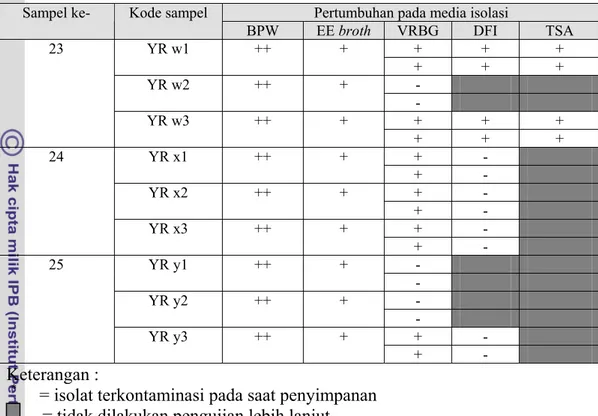
Tabel 4. Hasil isolasi E.sakazakii pada beberapa media (lanjutan)
Pertumbuhan pada media isolasi Sampel ke- Kode sampel
BPW EE broth VRBG DFI TSA + + + YR w1 ++ + + + + - YR w2 ++ + - + + + 23 YR w3 ++ + + + + + - YR x1 ++ + + - + - YR x2 ++ + + - + - 24 YR x3 ++ + + - - YR y1 ++ + - - YR y2 ++ + - + - 25 YR y3 ++ + + - Keterangan :
* = isolat terkontaminasi pada saat penyimpanan
= tidak dilakukan pengujian lebih lanjut
+++ = pada BPW terbentuk gumpalan dan bau menyegat
++ = pada BPW terbentuk gumpalan; pada EE broth terjadi perubahan warna + = pada BPW terbentuk endapan; pada EE broth terjadi kekeruhan; pada
DFI terbentuk koloni tipikal hijau kebiruan; pada TSA terbentuk koloni kuning kecuali pada YR m1
- = tidak terdapat pertumbuhan koloni
Proses isolasi diawali dengan tahap pra pengkayaan dengan media BPW. Tahap ini dilakukan karena jumlah E.sakazakii yang mungkin terdapat pada susu formula atau makanan bayi diperkirakan rendah yaitu berkisar antara 0.22 hingga 1.61/100 g produk (Santos 2006). Pertumbuhan bakteri pada BPW yang sudah diinkubasi selama 24 jam pada 37oC ditunjukkan dengan adanya koagulasi, produksi gas, dan bau.
Tahap pengkayaan lanjutan pada EE broth dilakukan untuk memberikan kondisi yang lebih ideal bagi pertumbuhan E.sakazakii dan Enterobacter lainnya. Penelitian yang dilakukan oleh Oh et al. (2006) menunjukkan bahwa EE broth merupakan media pengkayaan selektif untuk E.sakazakii karena pada media tersebut terkandung dekstrosa untuk mendukung pertumbuhan dari sebagian besar
laktosa-negatif, namun media ini juga mengandung bahan selektif berupa garam empedu dan brillian green untuk menekan pertumbuhan bakteri Gram positif (Iversen & Forsythe 2007). Beberapa isolat yang ditumbuhkan pada EE broth dapat mengubah warna media dari hijau bening menjadi keruh, beberapa isolat mengubah media menjadi kuning, dan beberapa menunjukkan pembentukan lendir.
Isolasi pada VRBG agar menunjukkan pertumbuhan pada sebagian sampel dan sebagian sampel lainnya tidak terlihat adanya pertumbuhan. Tidak adanya pertumbuhan menunjukkan bahwa tidak terdapat Enterobaceriaceae pada sampel tersebut karena pada media VRBG terkandung bahan selekif berupa garam empedu. Pertumbuhan yang terjadi pada VRBG dapat dilihat dari pembentukan koloni yang menggumpal yang berwarna ungu bergradasi ke oranye atau kuning. Koloni yang menggumpal tersebut menandakan terjadinya presipitasi dari garam empedu.
Koloni yang positif pada VRBG selanjutnya digoreskan pada DFI, sementara sampel yang tidak menunjukkan pertumbuhan pada VRBG (negatif) dihentikan pengujiannya. Isolat yang menunjukkan koloni tipikal E.sakazakii pada media DFI (berwarna hijau kebiruan) antara lain YR c3a, YR g1a, YR g1b, YR g 2a, YR g 2b, YR k 1b, YR k2a, YR k2b, YR k3a, YR k3b, YR m1, YR t2a, YR t2b, YR w1, dan YR w3. Isolat YR c3a, YR g1a, YR g1b, YR g2a, YR g2b, YR t2a, dan YR t2b berasal dari manufaktur A. Isolat YR k2a, YR k2b, YR k3a, YR k3b, YR w1, dan YR w3 berasal dari manufaktur B. Sedangkan isolat YR m1 berasal dari produk dari manufaktur C namun diproduksi oleh manufaktur A.
Pada proses isolasi, beberapa koloni yang tumbuh pada VRBG tidak dapat tumbuh pada DFI atau dapat tumbuh namun tidak menunjukkan koloni tipikal sebagai E.sakazakii melainkan membentuk koloni lainnya yang berwarna bening seperti pada isolat YR a1, YR a2, YR c1, YR c2, YR m1, YR m2, YR m3, YR n1, YR n2, YR p1, YR p2, YR p3, YR q1, YR q2, YR q3, YR t3, YR u3, YR v1, YR v2, YR v3, YR x1, YR x2, YR x3, dan YR y3. Hal ini menunjukkan bahwa pada sampel produk-produk tersebut masih terdapat cemaran Enterobacteriaceae.
Koloni tipikal pada DFI yang berwarna hijau kebiruan selanjutnya digoreskan pada TSA, sedangkan koloni yang bening dihentikan pengujiannya.
Pada metode FDA (2002) penggoresan pada TSA dilakukan setelah penggoresan pada VRBG, namun karena tidak semua E.sakazakii berpigmen kuning, seperti yang dilaporkan oleh Farmer (1980) yang menyatakan bahwa beberapa galur
E.sakazakii produksi pigmen sangat dipengaruhi oleh suhu, maka untuk
menghindari kesalahan negatif pada saat isolasi digunakan media DFI yang bersifat kromogenik sebelum TSA. Hasil pengujian menunjukkan bahwa semua isolat yang menunjukkan koloni tipikal sebagai E.sakazakii pada DFI menunjukkan pertumbuhan koloni yang berwarna kuning pada TSA, kecuali isolat YR m1 yang menghasilkan koloni putih pada TSA.
Berdasarkan hasil isolasi E.sakazakii ini dapat dinyatakan bahwa dari 25 kemasan susu formula dan makanan bayi diperoleh 6 sampel yang menghasilkan isolat yang memiliki koloni tipikal sebagai E.sakazakii. Isolat-isolat yang menunjukkan koloni tipikal di media DFI dan TSA kemudian diuji secara biokimia menggunakan perangkat API 20E. Selain itu morfologi isolat juga diamati dengan menggunakan mikroskop. Hasil pengamatan mikroskop dapat dilihat pada Gambar 5.
Gambar 5. Penampakan E.sakazakii secara morfologi di bawah mikroskop dengan pembesaran 1000x. Sel berwarna merah yang menunjukkan bakteri Gram negatif dan berbentuk batang pendek.
Secara mikroskopis dapat dikatakan bahwa isolat sudah homogen yang ditunjukkan dengan sel berbentuk batang dan bersifat Gram negatif. Beberapa isolat mati selama penyimpanan sehingga yang tersisa hanya 8 isolat yaitu YR c3a, YR t2a YR t2b, YR k1b, YR k2a, YR k3a, YR w1, dan YR w3, dimana 8 isolat tersebut berasal dari 4 produk yang berbeda yaitu sampel 3, sampel 11, sampel 20, dan sampel 23.
Karakterisasi Sifat Fenotipik Isolat
Sifat fenotipik isolat yang diamati melalui sifat biokimianya dianalisis dengan program apiweb™ (Lampiran 3). Untuk dapat memunculkan angka persentase kemiripan, analisis tambahan ditentukan berdasarkan sifat E.sakazakii. Uji oksidase (OX) dinilai negatif karena E.sakazakii bersifat oksidase negatif, reaksi terhadap NO2 adalah positif, reaksi terhadap N2 adalah negatif. E.sakazakii
bersifat motil sehingga pengujian motilitas (MOB) adalah positif. Berdasarkan hasil-hasil di atas maka identifikasi biokimiawi dengan perangkat API 20E dapat dilihat pada Tabel 5.
Tabel 5. Hasil analisis uji biokimia E.sakazakii dengan menggunakan program apiweb™
Kode Produk Persentase kemiripan dengan E.sakazakii
YR c3a 18,5% YR t2a 2,6% YR t2b - * YR k1b -** YR k2a 98,4% YR k3a 98,4% YR w 1 98,4% YR w 3 98,4% E.sakazakii ATCC 352/7 98.4%
* = kemiripan lebih sebagai E.amnigenus (90.0%)
** = kemiripan lebih sebagai Pantoea spp. (80.3%)
Berdasarkan Tabel 5, dapat dilihat bahwa 4 isolat menunjukkan kemiripan sebagai E.sakazakii secara biokimia sebesar 98.4%, yakni isolat YR k2a, YR k3a, YR w1, YR w3, yang sama seperti kontrolnya yaitu E.sakazakii ATCC 352/7. Sebanyak 2 isolat meskipun menunjukkan koloni tipikal pada DFI, namun dengan API 20E kemiripannya lebih ke arah Enterobacteriaceae lainnya, yaitu isolat YR c3a yang hanya memiliki 18.5% kemiripan dengan E.sakazakii dan kemiripannya lebih ke arah E.cloacae (81,4%) dan isolat YR t2a yang lebih memiliki kemiripan ke arah Enterobacter amnigenus yaitu sebesar 90.6% sementara kemiripan dengan
E.sakazakii hanya sebesar 2.6%. Sebanyak 2 isolat lainnya tidak memiliki
kemiripan secara biokimia dengan E.sakazakii yaitu isolat YR t2b yang lebih mirip sebagai E.amnigenus sebesar 90.0% serta YR k1b yang lebih mirip sebagai Pantoea spp 3 (80,3%) dibandingkan dengan E.sakazakii. Untuk memastikan
bahwa isolat yang diperoleh merupakan E.sakazakii dilakukan konfirmasi dengan pengujian secara genetik melalui PCR dan sekuensing.
Analisis Keragaman Genetik E.sakazakii Isolasi DNA Genom
DNA dari bakteri dapat dimurnikan dengan metode yang berbeda-beda bergantung pada apakah DNA merupakan kromosomal atau ekstra kromosom. Pada penelitian ini digunakan ekstraksi DNA kromosomal dimana ditambahkan
sodium dodecyl sulphate (SDS) untuk menghancurkan (lisis) dinding sel dari
bakteri. Pada tahap selanjutnya dilakukan penambahan enzim proteinase K untuk mendegradasi protein-protein pengotor yang terdapat pada isolat. Penambahan larutan CTAB/NaCl juga sebagai detergen yang dapat membantu menyempurnakan penghancuran dinding sel dari bakteri.
Residu-residu pengotor seperti protein, oligopeptida, dan sisa-sisa dinding sel selanjutnya diekstrak dengan pelarut-pelarut organik seperti campuran fenol, kloroform, dan isoamil alkohol serta campuran antara kloroform dan isoamil alkohol yang berfungsi membantu denaturasi dan koagulasi protein (Taylor et al, 1993). Sebagian besar protein akan terdenaturasi dan memasuki fase organik atau akan terpresipitasi pada interfase antara fase organik dan fase aqueous. Fase aqueous yang bening dan mengandung DNA dapat dipindahkan ke tabung yang baru. Penambahan isopropanol serta perlakuan dingin dan penambahan garam dan asam dapat mengendapkan DNA pada fase aqueous tersebut sehingga membentuk sedikit endapan atau serabut-serabut yang berwarna putih. Penambahan etanol dapat mencuci DNA atau memisahkan DNA dari oligonukleotida-oligonukleotida kecil, sisa-sisa detergen, dan sisa-sisa pelarut organik yang digunakan untuk menghilangkan protein. Selanjutnya DNA yang diperoleh harus disimpan pada tempat yang bersuhu -20oC untuk menghindari dari kerja enzim nuklease. Pada penelitian ini, untuk masing-masing isolat bakteri dilakukan isolasi sebanyak 2 sampai 3 kali. Hasil isolasi DNA genom dapat diamati dengan elektroforesis gel agarosa. Hasil elektroforesis DNA genom hasil ekstraksi dapat dilihat pada Gambar 6.
Berdasarkan Gambar 6 dapat dilihat bahwa DNA genom E.sakazakii terdapat dalam bentuk DNA sirkular (nicked circle) yang berukuran lebih dari 10000 bp, DNA plasmid berbentuk utuh (supercoiled) yang berukuran 1200 bp, serta DNA plasmid yang berbentuk linier yang berukuran 2000 bp. Selain karena perbedaan ukurannya, laju migrasi DNA pada gel elektroforesis juga ditentukan oleh konformasi molekulnya. DNA dengan bentuk covalently closed circular (CCC) akan bergerak paling cepat, disusul berikutnya konformasi open circular (OC), dan yang terakhir adalah yang berbentuk linier. Oleh karena perbedaan konformasi menyebabkan perbedaan laju migrasi, maka penentuan ukuran suatu fragmen DNA selalu dilakukan pada konformasi linier (Currier & Nester, 1976)
M 1 2 3 4 5 6 7 8 9 10 11 12
Gambar 6. Isolasi DNA genom total E.sakazakii dari kultur murni. Visualisasi DNA dilakukan pada gel agarosa 1.5%. Sampel terdiri atas; Marker 1kb (M), E.sakazakii ATCC 352/7 (1), E8 (2), E6 (3), E12 (4), YR t2a (5), YR t2b (6), YR c3a (7), YR w1 (8), YR w3 (9), YR k 1b (10), YR k2a (11), dan YR k3a (12).
Sebelum diamplifikasi dengan PCR, hasil isolasi DNA genom diamati kemurniannya dengan melihat rasio absorbansi pada panjang gelombang 260 nm dan 280 nm, dimana batas kemurnian DNA adalah 1.8 – 2.0. Pada panjang gelombang 260 nm yang terdeteksi adalah material genetik DNA sedangkan pada panjang gelombang 280 nm yang terdeteksi adalah protein (Sambrook et al.1989). Pengamatan kemurnian DNA genom dapat dilihat pada Lampiran 4.
Data pada Lampiran 4 menunjukkan bahwa DNA hasil isolasi belum murni. Ketidakmurnian DNA genom terlihat dari perbandingan absorbansi antara panjang gelombang 260 nm dengan panjang gelombang 280 nm yang tidak masuk dalam cakupan nilai 1.8 – 2.0. Perbandingan yang kurang dari 1.8 ataupun lebih
2000 bp 1000 bp 10000 bp
dari 2 menunjukkan bahwa preparasi DNA telah terkontaminasi oleh fenol atau protein lainnya (Brown 1992). Beberapa sampel juga menunjukkan hasil absorbansi negatif, sedangkan minimal pembacaan absorbansi adalah 0. Hal ini disebabkan oleh ketidakbersihan kuvet spektrofotometer itu sendiri.
Hasil pengamatan spektrofotometri digunakan untuk mengetahui konsentrasi dari DNA hasil isolasi tersebut. Perhitungan konsentrasi DNA tersebut sangat penting bagi aplikasi pada tahap selanjutnya. Pemanfaatan informasi tersebut diantaranya untuk penentuan jumlah DNA saat pemotongan dengan enzim restriksi maupun penggunaan sebagai DNA cetakan pada saat PCR (Brown 1992).
Amplifikasi Gen 16S-rRNA dan Sekuensing
Amplifikasi gen 16S rRNA dari isolat E.sakazakii pada penelitian ini dilakukan dengan menggunakan pasangan primer 16SUNI-L/ Saka-2b (segmen 1) dan ESA1/16SUNI-R (segmen 2) menghasilkan produk PCR masing-masing berukuran 977 bp dan 408 bp. Pemilihan primer tersebut berdasarkan pada penelitian yang dilakukan oleh Hassan et al. (2007). Primer 16SUNI-L dan 16SUNI-R merupakan primer universal untuk sekuensing 16S rRNA Escherichia
coli yang berturut-turut berlokasi pada basa 8-28 dan basa 1410-1391 (Kuhnnert et al.1996), sementara primer ESA1 dan Saka-2b merupakan primer yang
didisain oleh Hassan et al. (2007) setelah melakukan pensejajaran antara sekuen
E.sakazakii dengan Enterobacteriaceae lainnya dimana berada pada posisi basa
1006 – 1023 dan 1004 -1020 dari gen 16S rRNA E.coli. Alasan utama dari pemilihan primer tersebut yaitu karena Hassan et al. (2007) juga menggunakan isolat lokal E.sakazakii yang diisolasi oleh Estuningsih (2006) sehingga sekuen parsial yang diperoleh pada penelitian ini dapat dibandingkan dengan sekuen parsial E.sakazakii yang juga merupakan isolat lokal.
Visualisasi pada amplifikasi E.sakazakii segmen 1 dapat dilihat pada Gambar 7 dan visualisasi hasil amplifikasi E.sakazakii segmen 2 dapat dilihat pada Gambar 8. Elektroforesis segmen 1 menggunakan marker berupa DNA
ladder berukuran 1 kb, sedangkan elektroforesis segmen 2 menggunakan marker
400 bp
M 1 2 3 4 5 6 7 8 9 10 11
Gambar 7. Hasil amplifikasi E.sakazakii menggunakan pasangan primer 16SUNI-L/Saka-2b. Visualisasi pita DNA dilakukan pada gel agarosa 1.5%. Sampel terdiri atas; Marker 1kb (M), E.sakazakii ATCC 352/7 (1), E4 (2), YR t2a (4), YR t2b (5), YR c3a (6), YR w1 (7), YR w3 (8), YR k1b (9), YR k2a (10), dan YR k3a (11). Lajur (1) dan (2) merupakan kontrol positif E.sakazakii.
Berdasarkan Gambar 7 dapat dikatakan bahwa kedelapan isolat yang diperoleh pada penelitian ini adalah E.sakazakii karena dapat menghasilkan amplikon berukuran 977 bp dengan pasangan primer 16SUNI-L/ Saka-2b (segmen 1) sama seperti kontrol positifnya (E.sakazakii ATCC 352/7 dan E4)
M 1 2 3 4 5 6 7 8 9 10 11 12
Gambar 8. Hasil amplifikasi E.sakazakii menggunakan pasangan primer ESA1/16SUNI-R. Visualisasi pita DNA dilakukan pada gel agarosa 1.5%. Sampel terdiri atas; Marker 100 bp (M), E.sakazakii ATCC 352/7 (1), E4 (2), E8 (3), E12 (4), YR t2a (5), YR t2b (6), YR c3a (7), YR w1 (8), YR w3 (9), YR k1b (10), YR k2a (11), dan YR k3a (12). Lajur (1) – (4) merupakan kontrol positif E.sakazakii.
Hasil amplifikasi dengan pasangan primer ESA-1/16SUNI-R (segmen 2) juga menunjukkan bahwa isolat yang diperoleh pada penelitian ini adalah
E.sakazakii seperti yang tervisualisasi pada Gambar 8. Hal ini karena isolat dapat
menghasilkan amplikon yang sama dengan kontrolnya yaitu E.sakazakii ATCC 352/7, E4, E8, dan E12. Untuk memastikan persen kemiripan genotipik isolat dengan E.sakazakii berdasarkan gen 16S rRNA nya maka dilakukan sekuensing. Urut-urutan basa DNA (hasil sekuensing) dapat dilihat pada Lampiran 5.
Hasil sekuensing dibandingkan dengan beberapa sekuen E.sakazakii yang ada pada GenBank dan dianalisis dengan program BLAST. Perbandingan dilakukan dengan sekuen-sekuen yang paling mirip (highly similar sequence) dengan Enterobacteriaceae. Berdasarkan hasil analisis program BLAST, sekuen parsial gen 16S rRNA isolat-isolat E.sakazakii pada penelitian ini memiliki kemiripan yang variatif dengan berbagai sekuen parsial E.sakazakii pada GenBank.
Sekuen yang dijadikan acuan dalam perbandingan hasil sekuen antara lain
E.sakazakii ATCC BAA-894, complete genome (nomor akses CP000783) dan
sekuen parsial yang berasal dari isolat lokal E.sakazakii hasil amplifikasi oleh Hassan et al. (2007) yaitu gen 16S rRNA galur 6a, 10a, 39a, dan 39d (nomor akses berturut-turut AY624069, AY624071, AY624070, dan AY624073). Sekuen parsial isolat E.sakazakii yang dibandingkan adalah sekuen hasil amplifikasi segmen 1 yang menggunakan pasangan primer 16SUNI-L dan Saka-2b. Hal ini dikarenakan sekuen E.sakazakii hasil isolasi Estuningsih et al. (2006) yang didaftarkan pada GenBank oleh Hassan et al. (2007) merupakan hasil amplifikasi sekuen parsial E.sakazakii segmen 1. Hasil perbandingan yang dilihat dari persen similaritas antara sekuen yang dijadikan acuan tersebut dapat dilihat pada Tabel 6.
Seluruh isolat dapat dianalisis dengan sekuen yang sangat mirip dengan
Enterobacteriaceae (highly similar sequence) kecuali untuk sekuen parsial isolat
YR t2a yang menggunakan perbandingan dengan beberapa sekuen yang mirip (somewhat similar sequence) karena tidak ditemukannya sekuen
Enterobacteriaceae yang sangat mirip dengan isolat YR t2a tersebut dan isolat ini
lebih memiliki kemiripan dengan sekuen berbagai bakteri selain
Enterobacteriaceae. Hasil analisis dengan program BLAST dapat menunjukkan
terdapat pada GenBank. Suatu sekuen dapat dikatakan homolog bila memiliki similaritas lebih dari 70% (Claverie & Notredame, 2007). Semakin besar persentase similaritas maka semakin dekat pula kekerabatan suatu organisme.
Tabel 6. Perbandingan tingkat homologi gen 16S rRNA isolat E.sakazakii hasil isolasi menggunakan pasangan primer 16SUNI-L/Saka-2b dengan beberapa sekuen GenBank yang dianalisis menggunakan program BLAST
Kemiripan dengan sekuen E.sakazakii pada GenBank Kode1
Isolat ATCC BAA-894a Galur 6ab Galur 10ac Galur 39ad Galur 39de
YR t2a 92% - - - - YR t2b 96% 96% 96% 96% 96% YR c3a 96% 95% 95% 95% 96% YR w1 96% 89% 89% 89% 89% YR w3 94% 93% 93% 93% 93% YR k1b 97% 97% 97% 96% 96% YR k2a 96% 95% 95% 95% 95% YR k3a 93% 93% 93% 93% 93%
Keterangan: 1 Dianalisis dengan highly similar sequence terhadap Enterobacteriaceae kecuali
pada isolat YR t2a yang menggunakan somewhat similar sequence pada program BLAST
a = nomor akses CP000783 b = nomor akses AY624069 c = nomor akses AY624071 d = nomor akses AY624070 e = nomor akses AY626073
- = tidak ada kemiripan
Hasil penelitian menunjukkan bahwa isolat YR t2a hanya memiliki kemiripan dengan genom lengkap (complete genome) E. sakazakii ATCC BAA-894 (nomor akses CP000783) sebesar 92% dan tidak menunjukkan kemiripan yang signifikan dengan 4 sekuen parsial E.sakazakii yang diamplifikasi dengan menggunakan primer yang sama oleh Hassan et al. (2007). Tidak adanya kemiripan dilihat dari tidak munculnya sekuen tersebut pada saat dianalisis dengan program BLAST sebagai somewhat similar sequence. Isolat YR t2a memiliki similaritas lebih besar dengan genom lengkap Citrobacter koseri ATCC BAA-895 (nomor akses CP0082221) yaitu sebesar 95%. Meskipun demikian, sesuai dengan pernyataan Claverie & Notredame (2007) yang menyatakan bahwa suatu sekuen dapat dikatakan homolog bila memiliki similaritas lebih dari 70% maka dapat tetap dapat dikatakan bahwa isolat YR t2a merupakan isolat yang homolog dengan E. sakazakii.
Isolat YR t2b memiliki kemiripan sebesar 96% dengan genom lengkap
E.sakazakii ATCC BAA-894 dimana kemiripan dengan sekuen parsial isolat lokal E.sakazakii galur 6a, 10a, 39a, dan 39d sama-sama sebesar 96%. Isolat YR c3a
memiliki kemiripan 96% dengan sekuen genom lengkap E.sakazakii ATCC BAA-894, dimana kemiripan dengan sekuen parsial isolat lokal E.sakazakii adalah 95% pada galur 6a , 10a, dan 39a serta 96% pada galur 39d. Isolat YR w1 memiliki kemiripan 96% dengan sekuen genom lengkap E.sakazakii ATCC BAA-894, dimana kemiripan dengan sekuen parsial isolat lokal E.sakazakii hanya sebesar 89% pada keempat galur pembanding tersebut. Isolat YR w3 memiliki kemiripan sebesar 94% dengan genom lengkap E. sakazakii ATCC BAA-894 sedangkan kemiripan dengan sekuen parsial isolat lokal E.sakazakii sebesar 93% pada keempat galur tersebut. Isolat YR k1b memiliki kemiripan sebesar 97% dengan genom lengkap E. sakazakii ATCC BAA-894 serta sekuen parsial isolat lokal
E.sakazakii galur 6a dan 10a. Untuk galur 39a dan 39d, isolat YR k1b memiliki
kemiripan sebesar 96%. Isolat YR k2a memiliki kemiripan sebesar 96% dengan genom lengkap E. sakazakii ATCC BAA-894, dan kemiripan sebesar 95% dengan keempat sekuen parsial isolat lokal E.sakazakii pembanding. Isolat YR k3a mempunyai kemiripan sebesar 93% baik terhadap sekuen genom lengkap E.
sakazakii ATCC BAA-894 maupun dengan keempat galur sekuen parsial isolat
lokal E.sakazakii.
E.sakazakii ATCC BAA-894 merupakan genom lengkap E.sakazakii
dari basa 1 hingga 4368373. Berdasarkan situs NCBI, isolat tersebut berasal dari susu formula pada bagian perawatan bayi baru lahir di rumah sakit yang terdapat infeksi E.sakazakii berdasarkan keterangan dari CDC (Centers for Disease
Control and Prevention). Namun Asakura et al.(2007) menyatakan bahwa isolat E.sakazakii ATCC BAA-894 berasal dari manusia, namun tidak ada keterangan
lebih lanjut mengenai asal isolat tersebut.
Berdasarkan penelitian ini dapat dikatakan bahwa antara hasil uji biokimia (API 20E) dengan hasil amplifikasi gen 16S rRNA tidak saling mendukung. Hal ini dapat saja terjadi karena gen 16S rRNA merupakan gen yang spesifik terhadap spesies tertentu (species specific) sehingga meskipun isolat
merupakan spesies yang sama namun gen yang mengekspresikan sifat biokimia dari isolat tersebut bisa saja berbeda.
Terdapatnya berbagai ragam isolat menyebabkan adanya usulan pengelompokan kembali E.sakazakii menjadi empat cluster (Iversen et al. 2004), sampai pada pengelompokan E.sakazakii ke dalam satu genus baru yaitu
Cronobacter spp. yang juga memisahkannya menjadi 5 spesies baru yang
dikelompokkan berdasarkan perbedaan dalam reaksi biokimianya (Iversen et al. 2007). Isolat YR t2a, YR t2b, YR c3a, dan YR k1b yang teridentifikasi dengan API 20E bukan sebagai E.sakazakii namun tetap memiliki kemiripan dengan
E.sakazakii secara genotipik dapat dikelompokkan ke dalam E.sakazakii cluster 3
hasil pengelompokkan Iversen et al. (2004). Isolat sisanya yakni YR w1, YR w3, YR k2a, dan YR k3a yang secara biokimia dan genotipik teridentifikasi sebagai
E.sakazakii dapat dikelompokkan dalam E.sakazakii cluster 2 hasil
pengelompokan Iversen et al. (2004). Hal ini dikarenakan meskipun isolat memiliki kemiripan dengan E.sakazakii namun persentasenya tidak terlalu besar (keragamannya 1.6 hingga 1.9%). Untuk menentukan pengelompokan lebih lanjut isolat ke dalam spesies Cronobacter gen.nov perlu dilakukan pengujian secara biokimia lebih lanjut.
Kekerabatan isolat hasil isolasi pada penelitian ini dengan beberapa sekuen parsial E.sakazakii dapat digambarkan dengan menggunakan dendogram. Pohon filogeni yang dibentuk berdasarkan metode neighbour joining dengan menggunakan program MEGA 4 dapat dilihat pada Gambar 9.
Sekuen-sekuen acuan yang dipakai antara lain 4 sekuen parsial yang berasal dari isolat lokal E.sakazakii Estuningsih et al. (2006) yang diamplifikasi oleh Hassan et al. (2007) yakni sekuen parsial E.sakazakii galur 6a gen 16S rRNA (nomor akses AY624069), sekuen parsial E.sakazakii galur 10a gen 16S rRNA (nomor akses AY624071), sekuen parsial E.sakazakii galur 39a gen 16S rRNA (nomor akses AY624070), sekuen parsial E.sakazakii galur 39d gen 16S rRNA (nomor akses AY624073). Selain itu digunakan juga beberapa sekuen parsial
E.sakazakii lainnya dari GenBank sekuen parsial E. sakazakii gen 16S rRNA
galur ATCC 51329 (nomor akses AY752937), sekuen parsial E. sakazakii gen 16S rRNA galur E796 (nomor akses EF059876), sekuen parsial E.sakazakii gen
E.sakazakii ATCC 51329 (AY752937) E.sakazakii galur E627 (EF059856) E.sakazakii galur E739 (EF059867) E.sakazakii galur E761 (EF059870) E.sakazakii galur E768 (EF059871) E.sakazakii galur E775 (EF059873) E.sakazakii galur 39d (AY624073) E.sakazakii galur 39a (AY624070) E.sakazakii galur E736 (EF059866) E.sakazakii galur 6a (AY624069) E.sakazakii galur 10a (AY624071) E.sakazakii (AB004746)
E.sakazakii galur E796 (EF059876) E.sakazakii galur E632 (EF059857) E.sakazakii YR k2a E.sakazakii YR w1 E.sakazakii YR k3a E.sakazakii YR c3a E.sakazakii YR w3 E.sakazakii YR t2b E.sakazakii YR k1b E.sakazakii YR t2a 2
16S rRNA galur E627 (nomor akses EF059856), sekuen parsial E.sakazakii gen 16S rRNA galur E775 (nomor akses EF059873), sekuen parsial E.sakazakii gen 16S rRNA galur E768 (nomor akses EF059871), sekuen parsial E.sakazakii gen 16S rRNA galur E761 (nomor akses EF059870), sekuen parsial E.sakazakii gen 16S rRNA galur E739 (nomor akses EF059867), sekuen parsial E.sakazakii gen 16S rRNA galur E736 (nomor akses EF059866), sekuen parsial E.sakazakii gen 16S rRNA galur E632 (nomor akses EF059857), sekuen parsial E.sakazakii gen 16S rRNA galur E627 (nomor akses EF059856), sekuen parsial E. sakazakii gen 16S rRNA galur v266 (nomor akses EF088350), dan sekuen parsial Enterobacter
sakazakii gen 16S rRNA (nomor akses AB004746).
Gambar 9. Dendogram Neighbour-Joining berdasarkan sekuen parsial 16S rRNA pada isolat E.sakazakii hasil isolasi dengan beberapa sekuen parsial
Berdasarkan dendogram dapat dilihat bahwa E.sakazakii hasil isolasi pada penelitian ini berada pada sub cluster yang berbeda dengan isolat lokal
E.sakazakii (galur 6a, 10a, 39a, dan 39d) Estuningsih et al. (2006) yang
diamplifikasi oleh Hassan et al. (2007). Hal ini menunjukkan bahwa isolat yang diisolasi pada penelitian ini meskipun sama-sama merupakan isolat lokal
E.sakazakii namun tetap memiliki perbedaan dalam penempatan cluster nya.
Isolat YR k2a dan YR w1 berada pada cluster yang sama dengan isolat-isolat E.sakazakii dari GenBank (termasuk isolat-isolat lokal Estuningsih et al. (2006)), yang selanjutnya disebut sebagai cluster 1. Isolat YR k3a, YR c3a, YR w3, dan YR t2b berada pada cluster yang berbeda dengan cluster 1, namun masih berkerabat dekat, selanjutnya disebut sebagai cluster 2, isolat YR k1b dapat dikelompokkan menjadi suatu cluster tersendiri (cluster 3). Pada dendogram dapat dilihat bahwa isolat YR t2a merupakan outgroup dari isolat-isolat
E.sakazakii hasil isolasi pada penelitian ini. Hal ini sejalan dengan hasil analisis
program BLAST yang menunjukkan bahwa isolat YR t2a lebih memiliki kemiripan dengan C.koseri (similaritas 95%) daripada dengan E.sakazakii (92%).
Isolat YR t2a dan YR t2b merupakan isolat yang berasal dari kemasan yang sama, hal ini menunjukkan bahwa terdapat dua filogeni pada satu kemasan susu formula. Hal serupa juga diemukan oleh Lehner et al. (2004) yang menemukan perbedaan filogenik setelah membandingkan sekuen gen 16S rRNA pada 13 galur E.sakazakii. Adanya perbedaan filogeni tersebut diduga berasal dari sumber kontaminasi bakteri yang berbeda pada tahap manufaktur.
Pengaruh Suhu Rekonstitusi terhadap E.sakazakii
Persiapan Kultur. Isolat yang digunakan pada pengujian ini adalah
sebanyak 16 isolat yaitu E.sakazakii ATCC 352/7; 7 isolat dari Estuningsih (2006) yaitu E1, E2, E3, E4, E5, E6, dan E7; serta 8 isolat hasil isolasi pada penelitian ini yaitu YR c3a, YR t2a, YR t2b, YR k1b, YR k2a, YR k3a, YR w 1, dan YR w3. Tahap persiapan dilakukan dengan metode pengeringan beku (freeze
dry) diharapkan untuk mendapatkan isolat dalam bentuk kering sehingga dapat
dicampurkan ke dalam susu bubuk sebagai simulator. Pemilihan metode kering beku juga dilakukan dengan harapan mendapat jumlah E.sakazakii yang cukup
tinggi sebagai jumlah awal sebelum rekonstitusi. Kondisi yang ekstrim diciptakan unuk melihat seberapa besar pengaruh suhu rekonstitusi tersebut terhadap
E.sakazakii. Jumlah E.sakazakii dalam susu bubuk sebelum rekonstitusi dapat
dilihat pada Tabel 7.
Untuk melihat besar penurunan log CFU/ml antara susu terinokulasi sebelum rekonstitusi dan sesudah direkonsitusi, maka pada kondisi sebelum rekonstitusi, yang mana data tersedia dalam bentuk log CFU/g, maka angka sebelum rekonstitusi merupakan konversi dari log CFU/g ke log CFU/ml yang diperoleh dari perhitungan secara matematis. Karena jumlah susu terinokulasi yang direkonstitusi adalah 10g dan air yang merekonstitusi membuat volume susu terekonstitusi adalah 100 ml maka jumlah awal inokulum yang terdapat pada susu sebelum direkonstitusi adalah 1 log di bawah jumlah awalnya yang diperoleh dalam log CFU/g. Pengamatan plating susu terinokulasi selengkapnya dapat dilihat pada Lampiran 6.
Tabel 7. Jumlah E.sakazakii dalam susu bubuk pasca inokulasi sebelum rekonstitusi
Kode isolat Jumlah E.sakazakii (log CFU/g) ATCC 352/7 5,72 E1 5,75 E2 6,86 E3 6,71 E4 5,93 E5 6,85 E6 6,30 E7 6,35 YR t 2a 8,13 YR t 2b 7,02 YR c 3a 7,05 YR k 1b 7,72 YR k 2a 6,76 YR k 3a 8,22 YR w1 6,55 YR w3 6,47
0 1 2 3 4 5 6 7 8 AT CC 35 2/7 E1 E2 E3 E4 E5 E6 E7 YR t2a YR t2b YR c3a YR k1b YR k2 a YR k3a YR w1 YR w3 kode isolat lo g CF U/ m l
sebelum rekonstitusi (terhitung) setelah rekonstitusi (plating)
Simulasi Rekonstitusi dalam Penyiapan Susu Formula. Susu bubuk
yang telah dicampur E.sakazakii dalam jumlah tertentu (Tabel 7) direkonstitusi di berbagai suhu yang berbeda untuk melihat seberapa besar pengaruh rekonstitusi terhadap susu yang mengandung E.sakazakii. Rekonstitusi dilakukan pada suhu 100oC, 70oC, 40oC, dan 4oC. Pada rekonstitusi 100oC plating dilakukan pada tingkat pengenceran 100 – 102 karena diduga jumlah E.sakazakii yang bertahan akan sangat rendah (< 2.5 x 102). Hasil penelitian menunjukkan bahwa seluruh isolat tak terdeteksi bila direkonstitusi dengan suhu 100o C kecuali isolat E2 dan YR w3 yang masing-masing masih menyisakan bakteri sebesar 1.15 x 101
CFU/ml dan 1 CFU/ml (< 2.5 x 102). Namun jumlah tersebut tidak bisa dianggap akurat mengingat limit deteksi pada metode plating adalah 2,5 x 102 dan jumlah tersebut jauh di bawah dosis infeksi yang dapat ditimbulkan oleh E.sakazakii. Data pengamatan rekonstitusi suhu 100oC dapat dilihat pada Lampiran 7. Hal ini menunjukkan bahwa E.sakazakii mudah terbunuh dengan suhu rekonstitusi 100oC.
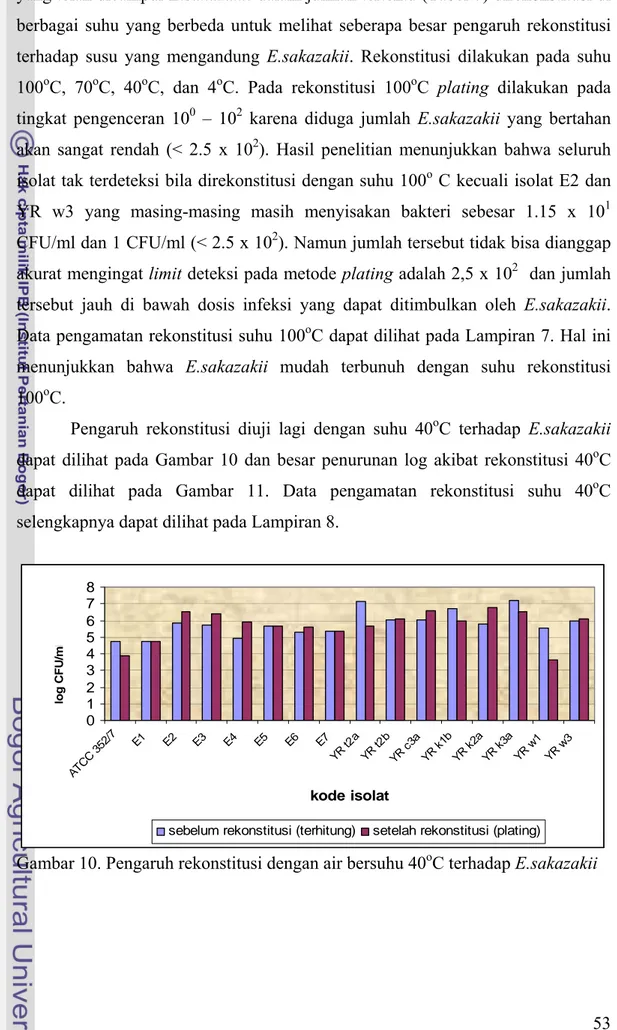
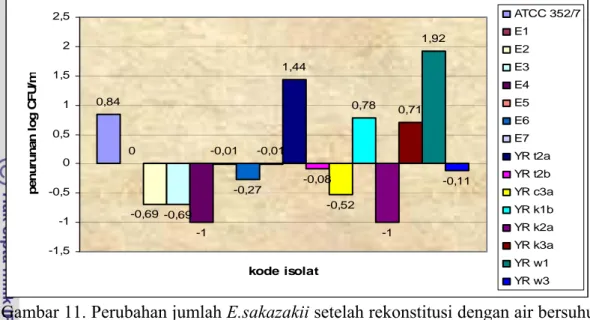
Pengaruh rekonstitusi diuji lagi dengan suhu 40oC terhadap E.sakazakii dapat dilihat pada Gambar 10 dan besar penurunan log akibat rekonstitusi 40oC dapat dilihat pada Gambar 11. Data pengamatan rekonstitusi suhu 40oC selengkapnya dapat dilihat pada Lampiran 8.
0,84 0 -0,69 -1 -0,01 -0,27 -0,01 1,44 -0,08 -0,52 0,78 -1 1,92 -0,11 -0,69 0,71 -1,5 -1 -0,5 0 0,5 1 1,5 2 2,5 kode isolat penur unan l o g C FU /m ATCC 352/7 E1 E2 E3 E4 E5 E6 E7 YR t2a YR t2b YR c3a YR k1b YR k2a YR k3a YR w1 YR w3
Gambar 11. Perubahan jumlah E.sakazakii setelah rekonstitusi dengan air bersuhu 40oC
Perubahan jumlah E.sakazakii pada susu mengandung E.sakazakii yang direkonstitusi pada suhu 40oC berbeda-beda antara isolat satu dengan isolat lainnya. Sebagian besar isolat justru mengalami kenaikan jumlah E.sakazakii setelah direkonstitusi pada suhu 40oC seperti pada isolat E2, E3, E4, E5, E6, E7, YR t2b, YR c3a, YR k2a, dan YR w3 meskipun kenaikannya tidak terlalu besar yaitu maksimum 1 siklus log. Hal ini diduga karena suhu 40oC termasuk dalam suhu optimum pertumbuhan E.sakazakii dimana menurut Iversen et al. (2004) bahwa suhu optimum pertumbuhan E.sakazakii berada pada kisaran 37oC hingga 43oC, sehingga dengan rekonstitusi pada suhu 40oC tidak memberikan pengaruh berarti terhadap kematian E.sakazakii. Faktor lain yang berkontribusi adalah keterbatasan dalam metode plating, selain itu juga karena jumlah awal sel bakteri sebelum direkonstitusi berasal dari perhitungan, bukan dari hasil plating yang sesungguhnya maka perbandingan keduanya tidak terlalu akurat. Beberapa isolat mengalami penurunan jumlah bakteri, namun penurunannya tidak terlalu besar. Isolat ATCC mengalami penurunan sebesar 0.84 siklus log dan menyisakan sejumlah 3.88 log CFU/ml bakteri, YR t2a mengalami penurunan sebesar 1.44 siklus log yang merupakan penurunan terbesar kedua akibat rekonstitusi dengan 40oC namun masih menyisakan sejumlah 5.69 log CFU/ml bakteri, YR k1b mengalami penurunan sebesar 0,70 log CFU/ml dan menyisakan sejumlah 5,94 log CFU/ml bakteri, YR k3a yang mengalami penurunan sebesar 0.71 siklus log
dan masih menyisakan sejumlah 6.51 log CFU/ml bakteri, terakhir adalah YR w1 mengalami penurunan terbesar akibat rekonstitusi 40oC yaitu sebesar 1.92 siklus log, namun masih menyisakan sejumlah bakteri sebesar 3.63 log CFU/ml. Jadi, suhu rekonstitusi 40oC belum memadai untuk membunuh E.sakazakii bila jumlah awal bakteri dalam susu bubuk lebih dari 5.5 log CFU/ml. Bahkan menurut Iversen et al. (2004) suhu 37-43oC termasuk suhu optimum dari E.sakazakii untuk hidup dalam susu formula.
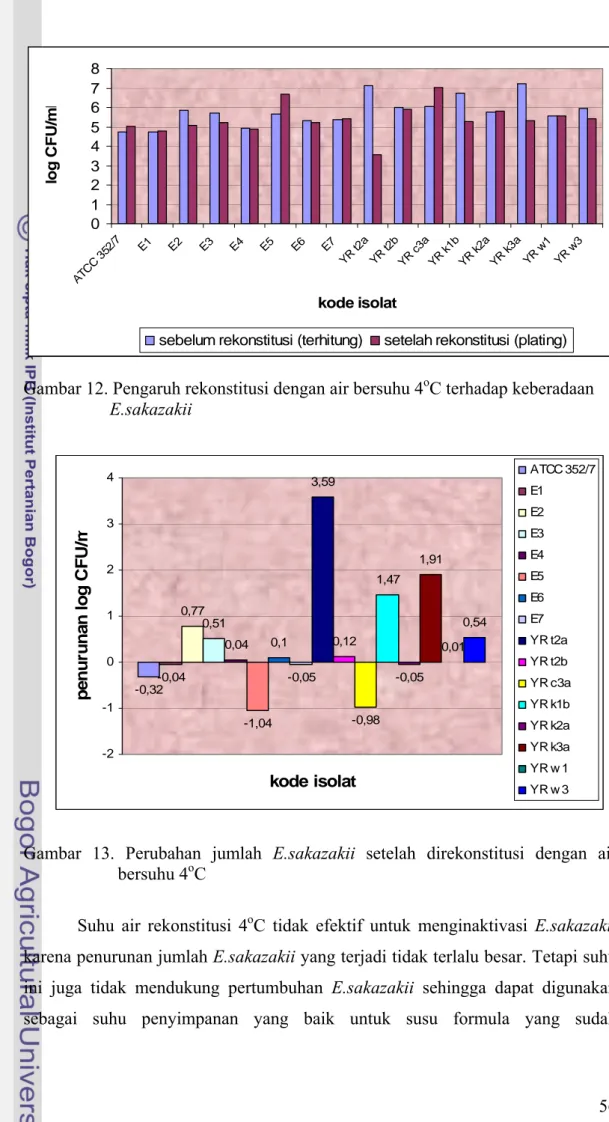
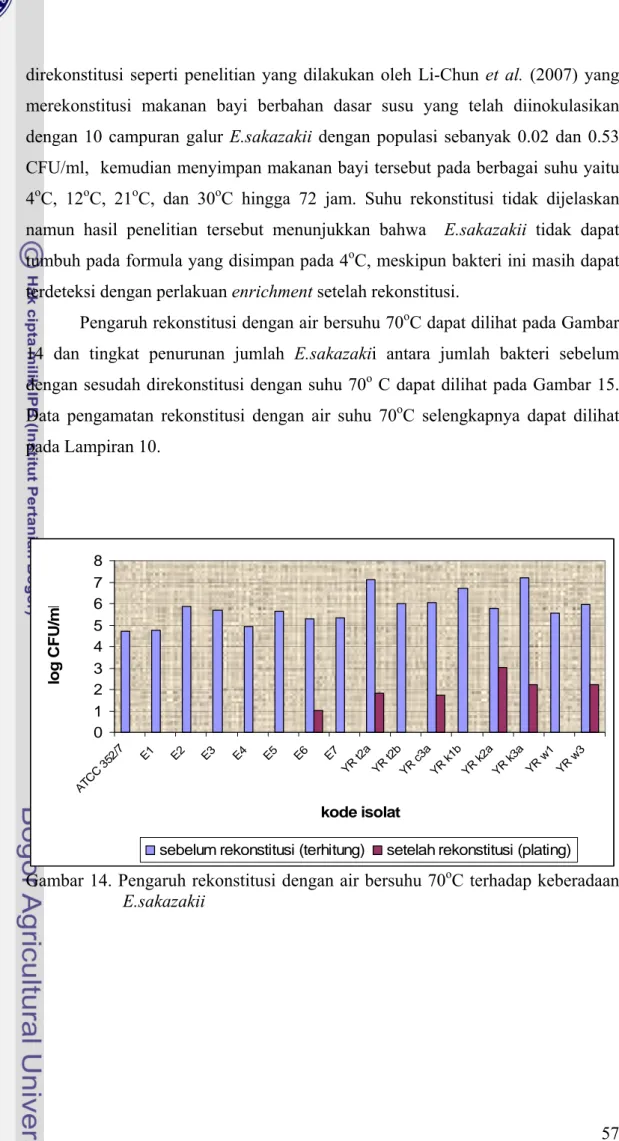
Perubahan jumlah E.sakazakii pada susu yang direkonstitusi dengan air bersuhu 4oC cukup bervariasi (Gambar 12 dan Gambar 13). Penurunan tertinggi
terjadi pada isolat YR t2a yaitu sebesar 3.59 siklus log. Namun dengan jumlah awal yang terdapat pada susu bubuk cukup tinggi yaitu sebesar 7.13 log CFU/ml maka jumlahnya setelah mengalami penurunan masih di atas dosis infeksinya yaitu sebesar 3.54 log CFU/ml. Penurunan terbesar kedua terjadi pada isolat YR k3a yang mengalami penurunan sebesar 1.91 siklus log dan masih menyisakan 5.31 log CFU/ml bakteri. Isolat YR k1b mengalami penurunan sebesar 1.47 log CFU/ml setelah direkonstitusi dengan air 4oC dan masih menyisakan bakteri sebanyak 5.25 log CFU/ml. Beberapa isolat mengalami kenaikan jumlah bakteri meski tidak terlalu besar yaitu isolat ATCC, E1, E4, E5, E7, YR c3a, dan YR k2a, dimana kenaikannya berturut-turut sebesar 0.32 log CFU/ml, 0.04 log CFU/ml, 1.04 log CFU/ml, 0.05 log CFU/ml, 0.98 log CFU/ml, dan 0.05 log CFU/ml. Untuk isolat lainnya penurunan jumlah bakteri akibat rekonstitusi dengan 4oC besarnya tidak lebih dari 1 siklus log. Isolat E2 mengalami penurunan sebesar 0.77 siklus log, E3 mengalami penurunan sebesar 0.51 siklus log, E4 mengalami penurunan sebesar 0.04 siklus log, E6 mengalami penurunan sebesar 0.10 siklus log, YR t2b mengalami penurunan sebesar 0.12 siklus log, YR w1 mengalami penurunan sebesar 0.01 siuklus log, dan YR w3 mengalami penurunan sebesar 0.54 siklus log. Penurunan jumlah bakteri tersebut tidak memadai bila jumlah bakteri yang terdapat pada susu besarnya lebih dari 5.5 log CFU/ml sehingga dapat dikatakan bahwa suhu 40oC dan 4oC tidak memadai dalam mengurangi
jumlah E.sakazakii. Data pengamatan rekonstitusi E.sakazakii dengan suhu 4oC dapat dilihat pada Lampiran 9.
0 1 2 3 4 5 6 7 8 ATC C 35 2/7 E1 E2 E3 E4 E5 E6 E7 YR t2a YR t2 b YR c 3a YR k 1b YR k2 a YR k3a YR w 1 YR w 3 kode isolat lo g C F U /m l
sebelum rekonstitusi (terhitung) setelah rekonstitusi (plating)
-0,32 -0,04 0,77 0,51 0,04 -1,04 0,1 -0,05 3,59 0,12 -0,98 1,47 -0,05 1,91 0,01 0,54 -2 -1 0 1 2 3 4 kode isolat pe nur u na n l o g C F U /m ATCC 352/7 E1 E2 E3 E4 E5 E6 E7 YR t2a YR t2b YR c3a YR k1b YR k2a YR k3a YR w 1 YR w 3
Gambar 12. Pengaruh rekonstitusi dengan air bersuhu 4oC terhadap keberadaan
E.sakazakii
Gambar 13. Perubahan jumlah E.sakazakii setelah direkonstitusi dengan air bersuhu 4oC
Suhu air rekonstitusi 4oC tidak efektif untuk menginaktivasi E.sakazakii karena penurunan jumlah E.sakazakii yang terjadi tidak terlalu besar. Tetapi suhu ini juga tidak mendukung pertumbuhan E.sakazakii sehingga dapat digunakan sebagai suhu penyimpanan yang baik untuk susu formula yang sudah
0 1 2 3 4 5 6 7 8 ATC C 3 52/7 E1 E2 E3 E4 E5 E6 E7 YR t2 a YR t2 b YR c3a YR k 1b YR k2a YR k3a YR w 1 YR w 3 kode isolat log C F U/ m l
sebelum rekonstitusi (terhitung) setelah rekonstitusi (plating)
direkonstitusi seperti penelitian yang dilakukan oleh Li-Chun et al. (2007) yang merekonstitusi makanan bayi berbahan dasar susu yang telah diinokulasikan dengan 10 campuran galur E.sakazakii dengan populasi sebanyak 0.02 dan 0.53 CFU/ml, kemudian menyimpan makanan bayi tersebut pada berbagai suhu yaitu 4oC, 12oC, 21oC, dan 30oC hingga 72 jam. Suhu rekonstitusi tidak dijelaskan namun hasil penelitian tersebut menunjukkan bahwa E.sakazakii tidak dapat tumbuh pada formula yang disimpan pada 4oC, meskipun bakteri ini masih dapat terdeteksi dengan perlakuan enrichment setelah rekonstitusi.
Pengaruh rekonstitusi dengan air bersuhu 70oC dapat dilihat pada Gambar
14 dan tingkat penurunan jumlah E.sakazakii antara jumlah bakteri sebelum dengan sesudah direkonstitusi dengan suhu 70o C dapat dilihat pada Gambar 15. Data pengamatan rekonstitusi dengan air suhu 70oC selengkapnya dapat dilihat pada Lampiran 10.
Gambar 14. Pengaruh rekonstitusi dengan air bersuhu 70oC terhadap keberadaan
5,86 5,65 4,27 5,35 6,02 4,3 6,72 2,74 5,01 5,55 4,72 4,75 5,71 4,93 5,3 3,77 0 1 2 3 4 5 6 7 8 kode isolat p e n u runa n l o g C F U /m ATCC 352/7 E1 E2 E3 E4 E5 E6 E7 YR t2a YR t2b YR c3a YR k1b YR k2a YR k3a YR w1 YR w3
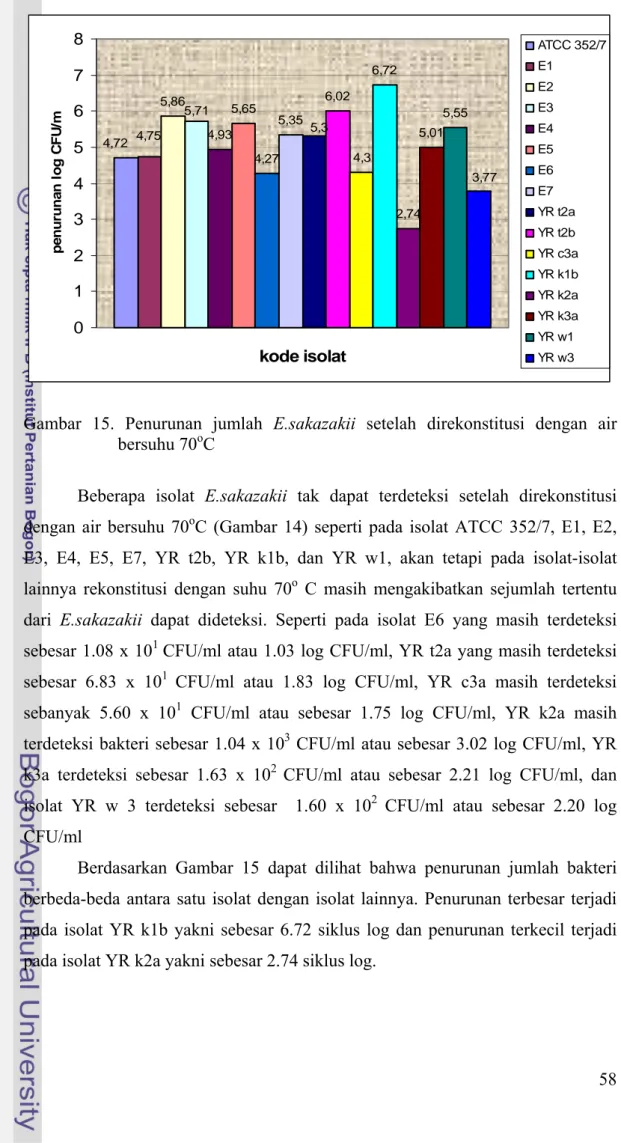
Gambar 15. Penurunan jumlah E.sakazakii setelah direkonstitusi dengan air bersuhu 70oC
Beberapa isolat E.sakazakii tak dapat terdeteksi setelah direkonstitusi dengan air bersuhu 70oC (Gambar 14) seperti pada isolat ATCC 352/7, E1, E2, E3, E4, E5, E7, YR t2b, YR k1b, dan YR w1, akan tetapi pada isolat-isolat lainnya rekonstitusi dengan suhu 70o C masih mengakibatkan sejumlah tertentu dari E.sakazakii dapat dideteksi. Seperti pada isolat E6 yang masih terdeteksi sebesar 1.08 x 101 CFU/ml atau 1.03 log CFU/ml, YR t2a yang masih terdeteksi sebesar 6.83 x 101 CFU/ml atau 1.83 log CFU/ml, YR c3a masih terdeteksi
sebanyak 5.60 x 101 CFU/ml atau sebesar 1.75 log CFU/ml, YR k2a masih terdeteksi bakteri sebesar 1.04 x 103 CFU/ml atau sebesar 3.02 log CFU/ml, YR k3a terdeteksi sebesar 1.63 x 102 CFU/ml atau sebesar 2.21 log CFU/ml, dan isolat YR w 3 terdeteksi sebesar 1.60 x 102 CFU/ml atau sebesar 2.20 log CFU/ml
Berdasarkan Gambar 15 dapat dilihat bahwa penurunan jumlah bakteri berbeda-beda antara satu isolat dengan isolat lainnya. Penurunan terbesar terjadi pada isolat YR k1b yakni sebesar 6.72 siklus log dan penurunan terkecil terjadi pada isolat YR k2a yakni sebesar 2.74 siklus log.
Meskipun beberapa isolat E.sakazakii masih dapat bertahan pada rekonstitusi 70oC namun jumlahnya masih dibawah dosis infeksi yang diduga oleh Iversen dan Forsythe (2003) yaitu 1000 sel sebagai konsentrasi awal bakteri ini dapat menimbulkan infeksi, kecuali pada isolat YR k2a yang telah mencapai dosis infeksinya yaitu terdapat sel bakteri sebanyak 1.04 x 103 CFU/ml.
Secara umum dapat dilihat bahwa suhu 70oC cukup efektif dalam menurunkan jumlah E.sakazakii sehingga dapat digunakan sebagai suhu rekonstitusi susu formula dan makanan bayi untuk mengurangi resiko terhadap
E.sakazakii. Sesuai penelitian yang dilakukan oleh Nazarowec-White dan Farber
tahun 1997 bahwa suhu di atas 70oC dapat menurunkan jumlah E.sakazakii hingga jumlah yang terdeteksi kurang dari 2 log CFU/ml. Suhu 70oC juga merupakan suhu rekonstitusi susu formula yang dianjurkan oleh ICMSF (2002). Mengingat terdapat satu galur pada penelitian ini yang masih mencapai dosis infeksinya setelah direkonstitusi dengan air bersuhu 70oC maka perlu diperhatikan dalam penanganan susu formula dan makanan bayi pada skala rumah tangga dan rumah sakit untuk menghindari rekontaminasi silang pada saat penyimpanan susu formula yang sudah terbuka agar jumlah E.sakazakii tidak melebihi 5 log CFU/g sehingga cukup aman bila direkonstitusi dengan air bersuhu 70oC.
Suhu 100oC tidak direkomendasikan sebagai suhu rekonstitusi meskipun dapat menyebabkan E.sakazakii pada sebagian besar isolat tidak dapat dideteksi lagi, namun karena mempertimbangkan rusaknya komponen gizi pada susu formula seperti penelitian yang dilakukan oleh Edelson dan Buchanan (2004).
Isolat YR t2a yang pada saat dianalisis secara genetik lebih memiliki kemiripan sebagai C.koseri ternyata pada saat direkonstitusi terlihat memiliki kecenderungan untuk tereduksi lebih besar dibanding isolat-isolat lokal lainnya seperti pada rekonstitusi dengan air bersuhu 4oC dan 40oC. Pada rekonstitusi dengan air bersuhu 70oC, isolat YR t2a juga mengalami penurunan yang cukup besar, hanya saja karena jumlah awal yang terdapat pada susu cukup tinggi (7,13 log CFU/ml) maka isolat ini masih dapat terdeteksi pasca rekonstitusi dengan air bersuhu 70oC.
Isolat YR k1b yang pada saat analisis genetik berada pada cluster yang berbeda dengan isolat-isolat E.sakazakii lainnya juga memiliki kecenderungan
untuk tereduksi lebih besar dibanding isolat-isolat lainnya, baik pada suhu 4o C, 40oC, dan 70oC. Bahkan pada suhu 70oC isolat ini mengalami reduksi paling besar dibandingkan isolat-isolat lainnya. Berdasarkan hasil penelitian dapat dikatakan bahwa isolat yang memiliki kemiripan lebih besar sebagai E.sakazakii juga memiliki ketahanan lebih besar terhadap suhu air rekonsitusi 70oC.
Uji Hang Time. Hasil pengujian hang time terhadap susu yang
direkonstitusi pada 70oC dapat dilihat pada Lampiran 11. Berdasarkan hasil dapat dilihat bahwa beberapa isolat yang pada saat direkonstitusi dengan air bersuhu 70oC tak dapat dideteksi, bila dibiarkan selama 2 jam akan meningkat jumlahnya. Meskipun demikian isolat E.sakazakii ATCC 352/7, E1, E2, E7, YR t2b, dan YR w1 tidak meningkat jumlahnya sampai terdeteksi pada 2 jam setelah susu direkonstitusi pada isolat tersebut.
Beberapa isolat yang pada saat rekonstitusi dengan 70oC tak terdeteksi, dan tetap tidak menunjukkan pertambahan hingga 4 jam dan 8 jam hang time adalah isolat E1, E2, dan E7. Untuk isolat YR t2b, penambahan jumlah bakteri baru terjadi setelah hang time 8 jam menjadi 1.20 log CFU/ml. Jumlah ini masih di bawah konsentrasi awal bakteri ini dapat menyebabkan infeksi yaitu sebesar 1000 sel berdasarkan dugaan Iversen & Forsythe (2003). Untuk isolat E.sakazakii ATCC 352/7 setelah hang time selama 4 jam mengalami kenaikan jumlah bakteri sebesar 1.05 siklus log dan setelah 8 jam hang time jumlah bakteri yang tumbuh adalah sebesar 1.70 log CFU/ml atau meningkat 0.65 siklus log dari jumlah sebelumnya. Pada isolat YR w1, meskipun pada 2 jam hang time masih tidak dapat terdeteksi, namun setelah 4 jam terjadi pertumbuhan bakteri sebesar 1.26 siklus log dan setelah 8 jam hang time menjadi 2.43 log CFU/ml.
Beberapa isolat E.sakazakii yang bertahan setelah direkonstitusi dengan air bersuhu 70oC meningkat jumlahnya selama hang time 2, 4, dan 8 jam (Gambar 16), sedangkan besar kenaikan jumlah E.sakazakii selama hang time dapat dilihat pada Gambar 17.
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 5
0 jam 2 jam 4 jam 8 jam hang time lo g CF U/ m E6 YR t2a YR c3a YR k2a YR k3a YR w3
Gambar 16. Pertumbuhan E.sakazakii selama hang time pasca rekonstitusi dengan air bersuhu 70oC
Pada Gambar 16 dapat dilihat bahwa setelah hang time selama 8 jam pada suhu ruang (28oC) isolat E.sakazakii berada pada kisaran 4 hingga 4.72 log CFU/ml. Jumlah ini mencapai konsentrasi E.sakazakii dapat menimbulkan infeksi yaitu melewati 1000 sel. Pada 2 jam hang time, isolat YR k2a dan YR w3 telah mencapai dosis infeksi bakteri ini yaitu 3.31 dan 3.55 log CFU/ml. Untuk isolat YR k2a hal ini dikarenakan jumlah sel yang bertahan pada saat direkonstitusi 70oC berada di atas 3 log CFU/ml, sedangkan pada YR w3 meskipun jumlah sel pada saat rekonstitusi di bawah 3 log CFU/ml, setelah 2 jam
hang time isolat ini mempunyai kemampuan pertumbuhan yang cepat hingga
melebihi 3 log CFU/ml. Pada 4 jam hang time, semua isolat mencapai dosis infeksi minimal E.sakazakii karena semua isolat minimal telah mencapai 3 log CFU/ml, seperti pada isolat E6, sementara isolat lainnya yakni YR t2a, YR c3a, YR k2a, YR k3a, dan YR w3 telah mencapai 4 hingga 4.5 log CFU/ml. Hal ini menunjukkan meskipun susu direkonstitusi dengan suhu 70o C, bila dalam susu bubuk sudah terdapat jumlah E.sakazakii sebesar 105 hingga 107 sel (dilihat dari jumlah awal sebelum rekonstitusi) maka setelah 4 jam hang time maka susu dinyatakan tidak aman untuk dikonsumsi. Namun jumlah E.sakazakii yang terdapat pada susu formula sangat kecil yaitu kurang dari 1 sel per gram, meskipun demikian bakteri ini dapat bertahan dalam susu bubuk tersebut selama 12 bulan (Caubilla-Barron et al. 2004).
0,41 0,58 1,03 0,29 0,22 1,36 1,51 1,69 1,54 1,29 1,73 1 1,11 0,33 0,3 0,12 0,36 0,06 0 0,2 0,4 0,6 0,8 1 1,2 1,4 1,6 1,8 2
E6 YR t2a YR c3a YR k2a YR k3a YR w 3 kode isolat k e na ik a n l og C FU /m
2 jam 4 jam 8 jam
Gambar 17. Pertumbuhan E.sakazakii selama hang time pasca rekonstitusi denga air bersuhu 70oC
Berdasarkan Gambar 17 dapat dilihat bahwa bakteri meningkat sampai 4 jam hang time dengan kisaran rata-rata sebesar 1.5 siklus log, kecuali pada isolat YR w3 yang tingkat pertumbuhan paling besar terjadi pada 2 jam hang time. Setelah mencapai puncak pertumbuhannya, maka laju pertumbuhan bakteri ini akan menurun kembali setelah 8 jam hang time. Hal ini menunjukkan beberapa sel bakteri mengalami sub letal pada saat direkonstitusi pada suhu 70oC maka pada saat 2 jam hang time beberapa sel mengalami fase lag dan mencapai puncak fase logaritmik pada saat 4 jam hang time. Kemudian setelah 8 jam hang time sel mengalami fase kematian. Pada isolat YR w3 laju pertumbuhannya lebih cepat dari pada isolat E6, YR t2a, YR c3a, YR k2a, dan YR k3a. Hal ini dapat dilihat dari puncak pertumbuhannya terjadi pada 2 jam hang time, kemudian pada 4 jam setelah rekonstitusi terjadi penurunan laju pertumbuhan hingga pada saat 8 jam
hang time pertumbuhan yang terjadi sangat kecil. Ini menunjukkan bahwa pada
saat 4 jam hang time isolat YR w3 mengalami awal fase kematian.
Beberapa isolat yang tidak menunjukkan pertumbuhan setelah direkonstitusi dengan air bersuhu 70oC ternyata menunjukkan peningkatan jumlah setelah hang time. Isolat E3, E4, E5, dan YR k1b yang pada saat direkonstitusi dengan 70o C tidak terdeteksi, namun setelah 2 jam hang time menunjukkan
-1,00 0,00 1,00 2,00 3,00 4,00 5,00
2 jam 4 jam 8 jam
hang time log C FU /m l E3 E4 E5 YR k1b
pertumbuhan bakteri pada jumlah tertentu, namun jumlahnya di bawah 1 log CFU/ml. Bahkan pada isolat YR k1b jumlah bakteri yang tumbuh pada 2 jam hingga 4 jam pertama hanya 0.5 CFU/ml. Pertumbuhan E.sakazakii untuk isolat yang tak terdeteksi pada saat direkonsitusi dengan air bersuhu 70oC pada 2 jam, 4 jam, 8 jam hang time dapat dilihat pada Gambar 18. Terjadinya pertumbuhan kembali bakteri ini setelah tidak adanya pertumbuhan pada saat direkonstitusi 70oC diduga karena pada saat direkonstitusi beberapa bakteri mengalami luka sub letal dan waktu 2 jam mencukupi untuk bakteri ini melakukan penyembuhan dan tumbuh kembali, meskipun dalam jumlah yang sangat kecil.
Gambar 18. Pertumbuhan E.sakazakii selama hang time yang tidak terdeteksi pasca rekonstitusi dengan air bersuhu 70o C
Besarnya kenaikan jumlah sel yang terjadi setelah 2 jam, 4 jam, dan 8 jam untuk isolat E3, E4, dan E5 dapat dilihat pada Gambar 19. Pada gambar dapat dilihat bahwa hang time selama 2 jam dapat menaikkan jumlah sel sebesar 0.18 log CFU/ml untuk E3, 0.3 log CFU/ml untuk E4, dan 0.94 log CFU/ml untuk isolat E5.
0,18 0,30 0,94 1,71 1,66 1,55 0,58 0,57 1,33 0,00 0,20 0,40 0,60 0,80 1,00 1,20 1,40 1,60 1,80 E3 E4 E5 kode isolat k e na ik a n l og C FU /m l
2 jam 4 jam 8 jam
Gambar 19. Kenaikan jumlah E.sakazakii selama hang time pasca rekonstitusi dengan air bersuhu 70oC
Berdasarkan Gambar 19 dapat dilihat bahwa peningkatan jumlah bakteri paling besar terjadi 4 jam hang time, namun jumlah sel yang tumbuh masih di bawah dosis infeksi yang dapat ditimbulkan. Setelah 8 jam hang time peningkatan jumlah sel tetap terjadi namun lebih rendah dibandingkan pada saat 4 jam hang
time. Pada 8 jam hang time, koloni E.sakazakii yang tumbuh adalah 2.47 log
CFU/ml untuk E3, 2.53 CFU/ml untuk E4, 3.82 CFU/ml untuk E5, dan 1.12 log CFU/ml untuk YR k1b.
Berdasarkan hasil uji hang time secara umum, meskipun susu formula direkonstitusi dengan air bersuhu 70oC, namun waktu jeda antara rekonstitusi dengan konsumsi sebaiknya tidak lebih dari 4 jam, karena waktu 4 jam setelah rekonstitusi masih memungkinkan E.sakazakii pada galur-galur tertentu mencapai dosis infeksinya. Hal ini sejalan dengan rekomedasi ICMSF (2002) yang menyatakan bahwa waktu jeda (hang time) pada susu formula tidak boleh lebih dari 4 jam setelah rekonstitusi.