Habitat preferences of nest-seeking bumble bees (Hymenoptera: Apidae)
in an agricultural landscape
Birgitta Svensson
a,∗, Jan Lagerlöf
a, Bo G. Svensson
baDepartment of Ecology and Crop Production Science, SLU, P.O. Box 7043, S-750 07 Uppsala, Sweden
bDepartment of Animal Ecology, Evolutionary Biology Centre, Uppsala University, Norbyvägen 18 D, S-752 36 Uppsala, Sweden Received 14 December 1998; received in revised form 27 May 1999; accepted 23 July 1999
Abstract
The aim of this study was to find out where bumble bee queens place their nests in the agricultural landscape. Nest-seeking behaviour was used to indicate nesting site preferences. Four types of agricultural landscape were investigated near Uppsala, Sweden: open, relatively open, relatively wooded and wooded. Twelve 500-metre-long transects, each including several intermingled habitats (e.g., field boundaries, pastures, clearings), were inspected repeatedly over a two-month period, from April to June 1991. In total, 147 observations of bumble bee queens (Bombus spp.) of eight species were recorded. Nest-seeking queens were observed most frequently along forest boundaries and field boundaries, in open uncultivated areas and in the relatively open landscape, whereas they were least frequent in forest and in clearings. Fields, pastures and road boundaries had intermediate frequencies. Differences among species were found in terms of both landscape type and habitat preferences:
B. terrestris, B. lapidarius, B. sylvarum and B. subterranius preferred open terrain, whereas B. lucorum and B. pascuorum
preferred forest boundaries. Withered grass and tussocks were the preferred nest-seeking patches among all species. ©2000 Published by Elsevier Science B.V. All rights reserved.
Keywords: Bumble bees; Bombus; Nest-seeking; Agricultural landscape; Habitat selection; Sweden
1. Introduction
With the modernisation of agriculture, the habitats that many animals and plants rely on for their survival have deteriorated. Meadows and pastures have disap-peared to a great extent, and covered drains have re-placed ditches. In the intensively utilised agricultural landscapes of today, field boundaries and roadsides have come to serve as major refuges for many species
∗Corresponding author. Tel.: +46-18-672526; fax: +
46-18-673430.
E-mail address: [email protected] (B. Svensson).
because they are the only non-cultivated habitats left (see, e.g., Lagerlöf and Wallin, 1993, for review).
Bumble bees are vital from the standpoint of agri-culture and conservation because they pollinate crops, fruit trees and wildflowers. However, for bumble bees to be effective, they must have local access to suitable nesting and hibernation sites, as well as to alternative food sources (i.e., wildflowers) during periods when crop plants are not flowering (Fridén, 1967; Fussell and Corbet, 1992a). A Norwegian study found the home range size of bumble bees to be only 250–350 m from the nest (Dramstad, 1996).
For recording the occurrence and activities of bumble bees, one has to distinguish between castes
(queens, workers and males) in order to gain a proper understanding of their use of resources over time and space (Svensson and Lundberg, 1977). Only a few studies have been made on bumble bee nesting-site preferences and characteristics of the nesting sites (Skovgaard, 1936; Svensson and Lundberg, 1977; Richards, 1978; Fussell and Corbet, 1992b).
Nest-site preferences of bumble bees have been in-vestigated by setting out artificial nests in the field. However, only a very small percentage of such nests are ever used (Richards, 1978; Fussell and Corbet, 1992b). It can, therefore, be advantageous to observe nest-seeking queens in spring, because a larger amount of data can be gathered in this way. This method was mentioned by Skovgaard (1936), and used by Richards (1973) and Svensson and Lundberg (1977).
The present investigation aimed at estimating the abundance of nest-seeking bumble bee queens in dif-ferent types of agricultural landscapes and habitats, with special emphasis on uncultivated areas.
2. Material and methods
2.1. Study area
The study was carried out between 13 April and 12 June 1991, within a 30 km2area near Funbo-Lövsta, about 20 km south-east of Uppsala, Sweden (59◦51′N, 17◦41′E). The region is dominated by plains at an altitude of less than 50 m above sea level. Intensive large-scale agriculture is carried out on clay soil, and mixed broad-leaved and coniferous forest grows on sand and gravely glacial till, esker material and shal-low soils over granite bedrock.
Within this area, the occurrence of nest-seeking bumble bee queens (Bombus spp.) was quantified in 12 smaller study areas (quadrats, 1 km×1 km), repre-senting four different landscape types:
1. Open (O), 0–15% forest; large agricultural fields, surrounded only by fields and roads. Three quadrats were investigated.
2. Relatively open (RO), 15–50% forest; large cultural fields, mostly surrounded by other agri-cultural fields and roads, but also by ditches, bushes, small woods and pastures. Four quadrats were investigated.
3. Relatively wooded (RW), 50–85% forest; small agricultural fields surrounded by forests and roads, but also by several small-habitat types, such as ditches, pastures and woods. Two quadrats were investigated.
4. Wooded landscape (W), 85–100% forest; small agricultural fields, almost always surrounded by forest. Three quadrats were investigated.
The quadrats were randomly chosen from a map covering the main study area. The longest distance between quadrats was 9 km. Within each quadrat, a 500-metre-long transect was laid out. Each transect included several intermingled habitats, and was made as representative as possible for the area. However, arable fields were only occasionally included (as a transport way between two other habitats), because they were presumed not to be possible nesting sites. The transects were not straight, but somewhat curved, in order to cover most habitats of interest within the quadrat. Table 1 gives details concerning the transects.
Eight habitat types were represented in this study: 1. Field (F). Agricultural field with cereals, grass ley
or fallow with a vegetation of weeds.
2. Field boundary (Fb). A grass-dominated strip be-tween two agricultural fields or bebe-tween an agri-cultural field and other open ground, pasture or clearing.
3. Pasture (P). Natural pasture, partly covered with broad-leaved trees and bushes, and grazed by horses, cattle or sheep.
4. Other open ground (Op). Mainly uncultivated open ground with abundant grass, tussocks and stones.
5. Clearing (C). An area with coniferous forest that had been clear cut 3–4 years earlier and then replanted. Tall herbs and bushes dominate the vegetation.
6. Road boundary (Rb). A grass-dominated strip be-tween a trafficable road and an agricultural field, other open ground or clearing.
7. Forest boundary (Fob). The edge between a mixed forest and agricultural field, pasture or clearing. The vegetation mainly consists of grass, herbs and bushes.
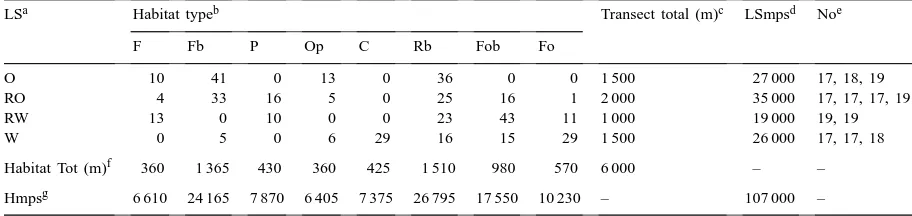
Table 1
Description of the study area near Uppsala, Sweden. The proportional distribution of habitat types (%) in each landscape type (LS) where nest-seeking bumble bees were counted during April–June 1991.(Each of the twelve transects was 500 m long.)
LSa Habitat typeb Transect total (m)c LSmpsd Noe
F Fb P Op C Rb Fob Fo
O 10 41 0 13 0 36 0 0 1 500 27 000 17, 18, 19
RO 4 33 16 5 0 25 16 1 2 000 35 000 17, 17, 17, 19
RW 13 0 10 0 0 23 43 11 1 000 19 000 19, 19
W 0 5 0 6 29 16 15 29 1 500 26 000 17, 17, 18
Habitat Tot (m)f 360 1 365 430 360 425 1 510 980 570 6 000 – –
Hmpsg 6 610 24 165 7 870 6 405 7 375 26 795 17 550 10 230 – 107 000 – aO, open; RO, relatively open; RW, relatively wooded; and W, wooded landscape.
bF, field; Fb, field boundary; P, pasture; Op, other open ground; C, clearing; Rb, road boundary; Fob, forest boundary; and Fo, forest. cTotal length of transects in each LS in the total study area.
dTotal length inspected in each LS during the season (in metres per season). eNumber of inspections in each transect during the season.
fTotal length of transects of each habitat in the total study area.
gTotal length inspected of each habitat during the season (in metres per season).
2.2. Queen bee studies
Bumble bee queens searching for a nesting site char-acteristically fly in a zigzag pattern close to the ground within a small area (termed a ‘patch’), occasionally going down to investigate the ground surface (Lund-berg and Svensson, 1975). The size of each patch characterised was about 1 m×1 m, which was chosen from the behaviour of the queens at each nest-seeking event. To characterise each patch, the occurrences of withered grass, new grass, tussocks, stones and moss within it, and in the area immediately surrounding it was classified as low (<10%), medium (10–50%) or high (>50%).
Bumble bee nests are difficult to find in numbers large enough to allow statistical analysis. Therefore, the number of queens performing nest-seeking be-haviour in an area of a given type was used as an indication of their relative preference for nesting in that type of area compared with other types of areas (cf. Richards, 1973; Svensson and Lundberg, 1977). Queens were counted using the transect method (‘belt method’, Banaszak, 1980), with all observations of nest-seeking bumble bee queens in an area extend-ing 5 m on either side of the transect beextend-ing counted. For each observation, the habitat type was noted and the characteristics of the environment adjoining the patch were described. Once detected, a queen was observed continuously until she stopped performing
nest-seeking behaviour, i.e., flew away from the patch. The observation time varied from 1–10 min. A queen could visit more than one patch, and each visit to a particular patch was counted as one observation. Most queens visited only one patch.
In most cases, each transect was inspected twice a week (occasionally, only once). Temperature and wind speed during the inspections varied within the range of 2–19◦C and 1–4 m/s, respectively. No inspections were made in rainy weather. Walking speed during a transect inspection was 20 m/min (1.2 km/h). The time of day of the inspection was varied randomly between 0900 and 1900 hours, Swedish Summer Time (+one hour GMT).
Bumble bees were identified according to Løken (1973). Bumble bee species differ in the mean date on which they break hibernation (Løken, 1973), so that nest-seeking observations need to be made throughout the spring season in order to be able to record all species. During April 1991, the mean temperature was near normal, whereas the weather was very cold, with a mean temperature 2◦C lower than normal for the months of May and June. This may have delayed the emergence of some species. Under normal conditions, the late-appearing species, B. hortorum, in particular, could be expected to appear more frequently.
queens’ nest-seeking behaviour. This parasite ap-parently disturbs a queen’s orientation capacity. As a result, infected queens search and dig in unusual places and continue nest-seeking until they die in the later part of summer (Lundberg and Svensson, 1975).
2.3. Statistical analysis
To compare the activities of bumble bees in dif-ferent landscape types and habitats, all results were transformed to total number of observations per 100 m during the season(No.of observations100 m ):
No.of observations
mps ×100
where mps (metres per season) for landscape types =length of transect ×number of inspections during the period (Table 1), and mps for habitat =length of each habitat in the total study area × number of inspections during the period (Table 1).
The data were log transformed before the statisti-cal analysis. One-factor ANOVA (the SAS procedure GLM; SAS Institute Inc., 1982) was used to anal-yse the data on landscape type (n=12) and habitat (n=96), respectively. To compare the landscape types and habitats pairwise, a PDIFF command was used.
To analyse the data on bumble bee species, a split-plot design was used (the SAS procedure GLM; SAS Institute Inc., 1982), with the bumble bee species as the dependent variable. The independent variables were landscape type (quadrats as error term) and habitat (n=96).
3. Results and discussion
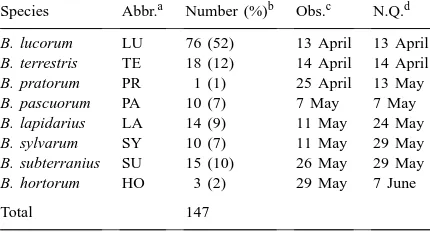
Some 147 observations of nest-seeking bumble bee queens (Bombus spp.) were recorded in this study and the following species were observed: Bombus lucorum L., Bombus terrestris L., Bombus pratorum L., bus pascuorum Scop., Bombus lapidarius L., Bom-bus sylvarum L., BomBom-bus subterraneus L. and BomBom-bus hortorum L. B. lucorum accounted for 76 of the ob-servations (52%), whereas B. pratorum and B. horto-rum were recorded only once and thrice, respectively (Table 2).
Bumble bees (all observations) showed significant preferences for certain landscape types (p<0.05,
Table 2
Bumble bee (Bombus spp.) species observed searching for nesting sites in an agricultural landscape near Uppsala during April–June 1991
Species Abbr.a Number (%)b Obs.c N.Q.d
B. lucorum LU 76 (52) 13 April 13 April
B. terrestris TE 18 (12) 14 April 14 April
B. pratorum PR 1 (1) 25 April 13 May
B. pascuorum PA 10 (7) 7 May 7 May
B. lapidarius LA 14 (9) 11 May 24 May
B. sylvarum SY 10 (7) 11 May 29 May
B. subterranius SU 15 (10) 26 May 29 May
B. hortorum HO 3 (2) 29 May 7 June
Total 147
aAbbreviations for species names.
bNumber of individuals observed during the period; and per-centage of total numbers observed (within parentheses).
cDate of first observation (obs.).
dDate of the first observed nest-seeking queen of each species. n=12). There were more observations of bumble bees in RO than in the other landscape types (Fig. 1). In pairwise comparisons, differences between RO and RW and between RO and W were statistically signif-icant (p<0.05, n=12) and the difference between RO and O was almost so (p=0.051, n=12). The number of species observed was also highest in RO.
One reason for these high values in the relatively open landscape could be that this type of landscape had the greatest habitat diversity (seven out of eight habi-tats were represented, Table 1). Richards (1978) found that Canadian bumble bee species differed in their nesting-site preferences and ascribed these differences to niche specialisation. Other authors have also re-ported differences in habitat (Skovgaard, 1936; Fussell and Corbet, 1992b) and altitudinal zone (Svensson and Lundberg, 1977) preferences among bumble bee species.
When landscape types with the same number of habitats were compared, more nest-seeking queens were found in the open landscape than in the rela-tively wooded one. In addition, the open landscape had a relatively high species diversity (five species), even though there were only four habitats present. Thus, habitat diversity is apparently not the only factor affecting the number of bumble bee species nesting in a given landscape type.
statisti-Fig. 1. Number of nest-seeking bumble bees observed per 100 m transect in different landscape types. (Abbreviations for landscape types and species defined in Tables 1 and 2, respectively).
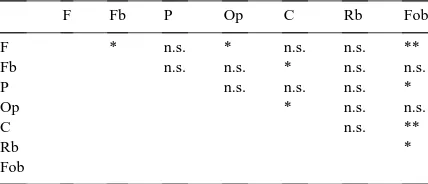
Table 3
Result of pairwise comparisonsabetween numbers of nest-seeking bumble bee queens observed in seven different habitatsb
F Fb P Op C Rb Fob
F * n.s. * n.s. n.s. **
Fb n.s. n.s. * n.s. n.s.
P n.s. n.s. n.s. *
Op * n.s. n.s.
C n.s. **
Rb *
Fob
a*p<0.05; **p<0.01; n.s., no significant difference. bHabitat abbreviations defined in Table 1.
cal differences among the landscape types and the results are consistent with Skovgaard’s (1936) find-ings in agricultural landscapes in Denmark, which showed that nest-selecting queens are influenced by the landscapes’ character and vegetation, and that species-specific preferences exist.
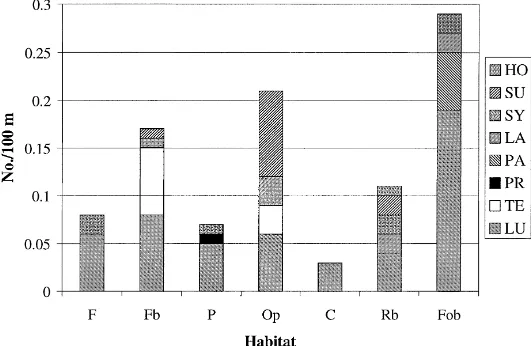
Bumble bees’ preference for the habitats differed significantly (p<0.01, n=96 (8 habitats×12 tran-sects)). Nest-seeking queens were observed most fre-quently in Fob (Fig. 2), and they were also frefre-quently encountered in Op and Fb. The frequency of encoun-ters was somewhat higher in Rb, than in F and P. The frequency of nest-seeking queens was low in C, and none was observed in Fo. Table 3 shows significant differences among habitats in pairwise comparisons.
The highest occurrence of queens in forest bound-aries probably reflects a preference for sites offering some degree of shelter. In their survey in England (where the public participated in reporting observa-tions of bumble bee nest sites), Fussell and Corbet (1992b) found that most nests were sheltered from wind. However, the high score for forest boundaries could also be ascribed to the strong preference of B. lucorum queens for this type of habitat. When this species was excluded from the analysis, the habitat ‘other open ground’ ranked highest in terms of fre-quency.
The habitat with the highest number of species of bumble bees was road boundary (five species were ob-served nest-seeking in this habitat type), although the density of observations (total per 100 m of investigated transect) was lower than in forest boundary and field boundary types. The number of observations in pas-ture varied between transects. Most observations were made in pastures with only a few grazing animals and with an abundance of grass.
Fig. 2. Number of nest-seeking bumble bees observed per 100 m transect in different habitats. (Abbreviations for habitat and species defined in Tables 1 and 2, respectively).
and Corbet, 1992b), showing that uncultivated areas are of greater value as nesting places, compared with cultivated areas.
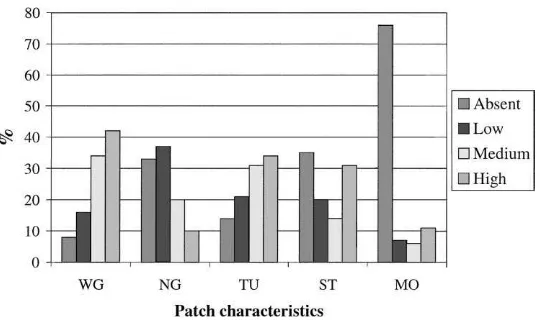
Fig. 3 shows characteristics of the patches where bumble bee queens were searching for nesting places. Withered grass and tussocks are typically found in un-cultivated areas. Almost all bumble bees of all species showed searching behaviour in patches with withered grass (92%), and most of them searched where the oc-currence of withered grass was considerable. In addi-tion, 86% of the queens were observed in patches with tussocks, and 70% of the bumble bees were observed in patches where new grass was uncommon or absent. Of the observations, 31% were made in patches where stones were frequent, and 35% were made in stoneless patches. Only 24% of the queens showed nest-seeking behaviour in patches with moss.
Queens of the latest emerging species were ob-served nest-seeking on patches with new grass more frequently than queens of the earlier emerging species. This difference is probably the result of higher abun-dance of new grass later in the season, when the late-appearing queens are most active.
Skovgaard (1936) concluded that the association be-tween bumble bee nests and stony ground is not as strong as some studies indicate. In the present study, one third of the observed queens were nest-seeking in patches without stones, which accords with Skov-gaard’s (1936) observations. The frequency and
place-ment of underground bumble bee nests are probably dependent on the abundance of abandoned rodents’ nests (Svensson and Lundberg, 1977; Harder, 1986).
Only eight of the 14 species of bumble bees men-tioned by Løken (1973) as occurring in this part of Sweden were recorded in this study. In studies car-ried out in recent years in Norway (Dramstad and Fry, 1995; Saville et al., 1997) and Great Britain (Fussell and Corbet, 1991), only seven and six species, respec-tively, were found in landscapes of similar, or greater, size than in the present investigation. In eastern Fin-land, Ranta and Tiainen (1982) found ten species and Skovgaard (1936) mentioned 13 species in his report from Denmark. Pekkarinen (1984) found that numbers of species in European areas were similar throughout the continent. The lower numbers recorded in recent investigations may reflect a decline in species diver-sity, as has been reported from England (Williams, 1982).
Fig. 3. Characteristics of patches (within all habitats where nest-seeking activities were observed) as a percentage of total number of observations for each set of patch characteristics (n=147). Occurrence of characteristics within patches is classified as absent, low, medium or high (WG, withered grass; NG, new grass; TU, tussocks; ST, stone; and MO, moss).
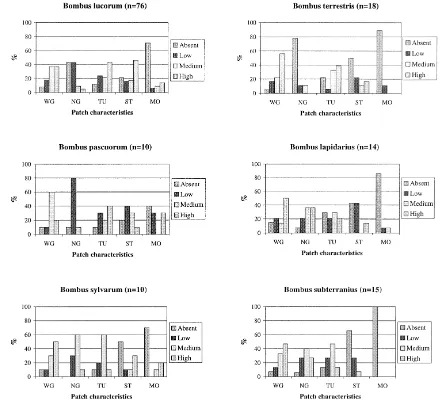
B. lucorum was the earliest emerging and most gen-eralistic species. This conflicts with the assertion by Richards (1978) that early species tend to be special-ists. In the present study, this species was observed in all landscape types (Fig. 1) and habitats, except for-est (Fig. 2). The densities in different habitats were significantly different (p<0.05, n=96). It was most abundant in forest boundaries. During the course of the study period, it became the most frequent species as well. Of this species, 80% were observed in patches with stones and they tended to search where stones were frequent (Fig. 4). The only patch characteristic that was associated with few observations of B. lu-corum, was new grass, probably because the queens were nest-seeking so early in the season. Therefore, it is concluded that although queens of B. lucorum can be found searching for nesting sites in almost all areas in the agricultural landscape, they seem to prefer for-est boundaries in the relatively open landscape with lots of withered grass, tussocks and stones.
The difficulty in discerning between workers of B. lucorum and B. terrestris, because of their similar colour patterns, has led to them being treated as a sin-gle species in some studies (e.g., Fussell and Corbet, 1991; Dramstad and Fry, 1995; Saville et al., 1997). However, morphological differences are more distinct between queens of the two species. Thus, the present
study was able to document differences in their choice of habitat for nesting places.
B. terrestris was observed only in field boundaries and other open ground (Fig. 2), and the frequency with which it was encountered differed significantly among the habitats (p<0.01, n=96). The first queens also appeared very early in the season (Table 2). This species, in particular, tended to be observed in patches where there was little new grass, whereas all species that appeared later in the season ,searched for nests predominantly in patches with new grass, i.e., at least 90% of the observations for each species (Fig. 4). B. terrestris was strongly associated with withered grass and seemed to prefer field boundaries and other open ground in various types of agricultural landscape with much withered grass and tussocks. It was often found searching in patches lacking most of the characteristics included in our analysis.
Fig. 4. Patch characteristics for the six most frequent bumble bee species (within all habitats where nest-seeking activities were observed). The result is shown as the percentage of the total number of observations for each species and for each set of patch characteristics. Occurrence of characteristics within patches is classified as absent, low, medium or high (WG, withered grass; NG, new grass; TU, tussocks; ST, stone; and MO, moss).
In northern Sweden, Svensson and Lundberg (1977) found that this species seemed to prefer open areas with tussocks. It seems probable that bumble bees in southern Sweden prefer locations sheltered from wind and sun, whereas queens in northern Sweden prefer open areas, in order to get more warmth from di-rect sunlight. The present findings agree with those of Skovgaard (1936), who found that B. pascuorum is,
despite its trivial name ‘field bumble bee’, associated with woodland, where it builds its nest in moss.
not appear to be keenly interested in tussocks or stones, despite this species sometimes being called the ‘stone bumble bee’. This agrees with Skovgaard (1936), who found this species to be associated with grassy areas and to have generalistic habitat preference.
Almost all B. sylvarum observed were found in rela-tively open landscape (Fig. 1), and this difference was statistically significant when comparing species dis-tributions among landscape types (p<0.05, n=96). Skovgaard (1936) also observed that B. sylvarum, de-spite its species name, is not found in forest, a fact borne out by the present study.
Queens of B. subterraneus also preferred open type landscape, as found by Reinig (1972). They were found only in open and relatively open landscapes (Fig. 1). This species was observed in other open ground, road boundaries and field boundaries (Fig. 2), and the frequencies with which it was observed differed significantly among the habitats (p<0.01, n=96). B. subterraneus was the only species that was never observed in mossy patches. It was also uncommon in stony patches.
4. Conclusion
In this study, as well as in other recent studies, the number of bumble bee species have been found to be lower than in earlier investigations. Complementary studies are needed to fully evaluate the consequences of landscape heterogeneity on bumble bee fauna, but it seems probable that the species diversity of bum-ble bees is declining. Because of this and the fact that species-specific preferences exist, a protection of existing boundaries and other uncultivated areas are needed to maintain bumble bee populations and diversity.
Acknowledgements
The authors thank Josef Stark for valuable discus-sions on bumble bee behaviour, Jan Olofsson for valu-able advice throughout the work, Erik Petersson and Ulf Olsson for skilful statistical advice, David Tilles for the linguistic revision and Jeremy Flower-Ellis and three anonymous referees for critical comments on earlier drafts of this manuscript. This project was fi-nancially supported by the foundation Stiftelsen Oscar och Lili Lamms minne.
References
Banaszak, J., 1980. Studies on methods of censuring the numbers of bees (Hymenoptera Apoidea). Pol. Ecol. Stud. 6, 355–366. Dramstad, W.E., 1996. Do bumble bees (Hymenoptera: Apidae) really forage close to their nests?. J. Insect Behav. 9, 163–182. Dramstad, W., Fry, G., 1995. Foraging activity of bumble bees (Bombus) in relation to flower resources on arable land. Agric. Ecosyst. Environ. 53, 123–135.
Fridén, F., 1967. Humlorna och jordbruket. Medd. Sveriges Fröodlareförbund 8, 1–16.
Fussell, M., Corbet, S.A., 1991. Forage for bumble bees and honey bees in farmland: a case study. J. Apic. Res. 30, 87–97. Fussell, M., Corbet, S.A., 1992a. Flower usage by bumble-bees: a
basis for forage plant management. J. Appl. Ecol. 29, 451–465. Fussell, M., Corbet, S.A., 1992b. The nesting places of some
British bumble bees. J. Apic. Res. 31, 32–41.
Harder, L.D., 1986. Influences on the density and dispersion of bumble bee nests (Hymenoptera, Apidae). Holarct. Ecol. 9, 99– 103.
Lagerlöf, J., Wallin, H., 1993. The abundance of arthropods along two field margins with different types of vegetation composition: an experimental study. Agric. Ecosyst. Environ. 43, 141–154.
Lundberg, H., Svensson, B.G., 1975. Studies on the behaviour of Bombus Latr. Species (Hym., Apidae) parasitized by
Sphaerularia bombi Dufour (Nematoda) in an alpine area.
Norw. J. Ent. 22, 129–134.
Løken, A., 1973. Studies on Scandinavian bumble bees (Hymenoptera, Apidae). Nor. Entomol. Tidsskr. 20, 1–218. Pekkarinen, A., 1984. Resource partitioning and coexistence in
bumblebees (Hymenoptera, Bombinae). Ann. Entomol. Fenn. 50, 97–107.
Ranta, E., Tiainen, M., 1982. Structure in seven bumble bee communities in eastern Finland in relation to resource availability. Holarct. Ecol. 5, 48–54.
Reinig, W.F., 1972. Ökologische Studien an mittel- und südost-europäischen Hummeln (Bombus Latr., 1802) (Hym., Apidae). Mitt. Münchner Ent. Ges. 60, 1–56.
Richards, K.W., 1973. Biology of Bombus polaris Curtis and B. hyperboreus Schönherr at Lake Hazen, Northwest Territories (Hymenoptera: Bombini). Quaest. entomol. 9, 115–157. Richards, K.W., 1978. Nest site selection by bumble bees
(Hymenoptera: Apidae) in southern Alberta. Can. Entomol. 110, 301–318.
SAS Institute Inc. 1982. SAS User’s Guide: Statistics. SAS Institute, Inc., Cary, NC.
Saville, N.M., Dramstad, W.E., Fry, G.L.A., Corbet, S.A., 1997. Bumblebee movement in a fragmented agricultural landscape. Agric. Ecosyst. Environ. 61, 145–154.
Skovgaard, O.S., 1936. Rødkløverens bestøvning, humlebier og humleboer. K. danske Selsk, (Skr. Nat. Math) Afd. 6, pp. 1–140. Svensson, B.G., Lundberg, H., 1977. Distribution of bumble bee nests in a subalpine/alpine area in relation to altitude and habitat (Hymenoptera, Apidae). Zoon 5, 63–72.