UNIVERSITAS INDONESIA
KOMPETISI DAN TUMPANG-TINDIH RELUNG ANTARA
SIAMANG (
Symphalangus syndactylus
) DAN MAMALIA
ARBOREAL LAINNYA DI TAMAN NASIONAL BUKIT
BARISAN SELATAN
SKRIPSI
MARSYA CHRISTYANTI 1006675991
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM DEPARTEMEN BIOLOGI
UNIVERSITAS INDONESIA
KOMPETISI DAN TUMPANG-TINDIH RELUNG ANTARA
SIAMANG (
Symphalangus syndactylus
) DAN MAMALIA
ARBOREAL LAINNYA DI TAMAN NASIONAL BUKIT
BARISAN SELATAN
SKRIPSI
Diajukan sebagai salah satu syarat untuk memperoleh gelar Sarjana Sains
MARSYA CHRISTYANTI 1006675991
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM DEPARTEMEN BIOLOGI
iv
Puji syukur penulis panjatkan kepada Allah Tritunggal karena hanya oleh
kasih karunia dan penyertaan-Nya, penulis dapat menyelesaikan skripsi ini.
Skripsi ini dapat dituntaskan karena dukungan dan bantuan berbagai pihak. Oleh
karena itu, penulis mengucapkan terima kasih kepada:
1. Dr. Noviar Andayani, M.Sc. sebagai pembimbing I dan Dr. Nurul Laksmi
Winarni, M.Sc. sebagai pembimbing II yang telah membimbing penulis dari
awal pencetusan ide penelitian sampai akhir masa penulisan. Penulis
berterima kasih untuk seluruh masukan, gagasan, kritik, dukungan, dan
terutama inspirasi yang telah diberikan.
2. Drs. Wisnu Wardhana, M.Si. sebagai penguji I dan Dr.rer.nat. Yasman, M.Sc.
sebagai penguji II yang telah memberikan banyak masukan dan kritik dalam
rancangan penelitian dan penulisan hasil penelitian.
3. Dr. Anom Bowolaksono, M.Si. sebagai Penasihat Akademik atas bimbingan
dan dukungan selama masa perkuliahan.
4. Dr. Andi Salamah sebagai Ketua Prodi S1 Departemen Biologi FMIPA UI dan
Dr.rer.nat. Yasman, M.Sc. sebagai Ketua Departemen Biologi FMIPA UI.
5. Seluruh staf pengajar dan karyawan Departemen Biologi FMIPA UI atas
bantuan selama masa perkuliahan.
6. Dr. Susan Lappan untuk ilmu, kritik, dan saran dalam pembuatan rancangan
penelitian.
7. Wildlife Conservation Society-Indonesia Program yang telah memberikan
kesempatan untuk melakukan penelitian di Stasiun Penelitian Way Canguk.
Penulis berterima kasih kepada Pak Opo yang telah memberikan perizinan dan
saran untuk penelitian, Mas Seti sebagai asisten lapangan, dan Mas Gawik,
Mas Jayus, Mas Lazi, dan Mas Rahman yang telah membantu selama
pengambilan data di lapangan.
v
setia berdoa dan memberikan dukungan finansial.
10.Almarhum Ayah yang semasa hidupnya merupakan seorang petualang dan
pecinta alam. Jiwa petualang dan kisah hidupnya yang hanya penulis dengar
sebagai cerita selalu menjadi motivasi bagi penulis untuk tidak berhenti
mengeksplorasi alam.
11.Partner ekologi hewan, yaitu Sheherazade, Ardiantiono, Shafia Zahra, Pramita
Indrarini, dan Achmad Ridha Junaid, yang telah menjadi sahabat diskusi yang
menyenangkan; Dyna, Abinubli, Ricky, Ninda, Elisabeth, Ayu, Deka, Cindy
Kus, dan seluruh anggota keluarga B10GENESIS lainnya untuk persahabatan
dan kekeluargaan selama masa perkuliahan.
12.Teman-teman, junior, dan senior Biologi serta ketiga BSO (KSHL Comata,
OMPT Canopy, dan SIGMA-B UI), terutama Nuruliawati, Niken Ekatiwi,
Lucia, Kak Sasha, Kak Sisil, Kak Eman, Kak Wendy, untuk tumpahan ilmu,
diskusi, motivasi, dan pengalaman lapangan.
13.Teman-teman persekutuan dalam Kristus, yaitu Octy, Gina, Aviana, Livia,
Septi, Zendy, Inez, dan seluruh anggota Persekutuan Alumni SMAN 8 untuk
doa dan semangat.
Walaupun naskah ini tidak sempurna, penulis berharap hasil penelitian ini dapat
berkontribusi bagi perkembangan ilmu pengetahuan, terutama untuk konservasi
biodiversitas hutan hujan tropis Indonesia.
Depok, 30 Juni 2014
vii
Nama : Marsya Christyanti
Program Studi : S1 Biologi
Judul : Kompetisi dan Tumpang-tindih Relung antara Siamang (Symphalangus syndactylus) dan Mamalia Arboreal Lainnya di Taman Nasional Bukit Barisan Selatan
Siamang hidup berdampingan dengan berbagai spesies mamalia arboreal yang berpotensi sebagai kompetitor di Taman Nasional Bukit Barisan Selatan (TNBBS). Tujuan penelitian ini adalah untuk mengetahui apakah terjadi kompetisi antara siamang dan mamalia arboreal lainnya serta mengetahui tumpang-tindih relung berdasarkan penggunaan habitat dan pemilihan pakan di antara komunitas mamalia arboreal di Stasiun Penelitian Way Canguk, TNBBS. Pengambilan data dilakukan pada bulan Februari hingga April 2014 dengan dua metode, yaitu metode focal instantaneous sampling untuk pengamatan perilaku siamang dan metode transek garis untuk survei mamalia arboreal. Hasil penelitian menunjukkan bahwa siamang berkompetisi dengan simpai, bajing kelapa, dan jelarang hitam. Tumpang-tindih relung terbesar terjadi antara siamang dengan jelarang hitam berdasarkan pemilihan pakan (Ro = 0,418) dan penggunaan habitat
(Uji Wilcoxon, p-value > 0,05). Dari 57 interaksi interspesifik antara siamang dan mamalia arboreal lainnya, terdapat 61,40% interaksi netral, 19,30% agresi, dan 19,30% dominansi. Kesimpulan penelitian ini adalah kompetisi interferensi dan eksploitatif terjadi antara siamang dan ketiga spesies mamalia arboreal serta terdapat tumpang-tindih relung antara siamang dan ketiga spesies mamalia arboreal.
Kata Kunci : kompetisi interspesifik, mamalia arboreal, relung, siamang.
viii
Name : Marsya Christyanti
Study Program : S1 Biologi
Judul : Competition and Niche Overlap between Siamang (Symphalangus syndactylus) and Other Arboreal Mammals in Bukit Barisan Selatan National Park
Siamang coexists with various arboreal mammal species which are potential competitors to siamang in Bukit Barisan Selatan National Park (BBSNP). The aims of this study are to determine whether interspecific competition occurs between siamang and other arboreal mammals and to determine niche overlap in terms of habitat use and food selection among mammals community in Way Canguk Research Station, BBSNP. Data collection was conducted on February until April 2014 using two methods: focal instantaneous sampling to measure siamang behavior and line transect method to survey coexisting mammals. The result of this research suggests that siamang competes with banded langur, plaintain squirrel, and black giant squirrel. Niche overlap is the highest between siamang and black giant squirrel based on food preference (Ro = 0,418) and habitat use (Uji Wilcoxon, p-value > 0,05). Among 57 interspecific interactions between siamang and other arboreal mammals, 61,40% are netral interactions, 19,30% are agressions, and 19,30% are dominance interactions. This research concludes that interference and exploitative competition occur between siamang and three other arboreal mammals and there is niche overlap among them.
Keywords : interspecific competition, arboreal mammals, niche, siamang.
ix
HALAMAN JUDUL ... i
HALAMAN PERNYATAAN ORISINALITAS ... ii
LEMBAR PENGESAHAN ... iii
KATA PENGANTAR ... iv
LEMBAR PERSETUJUAN PUBLIKASI KARYA ILMIAH ... vi
ABSTRAK ... vii
ABSTRACT ... viii
DAFTAR ISI ... ix
DAFTAR GAMBAR ... xi
DAFTAR TABEL ... xii
DAFTAR LAMPIRAN ... xiii
1. PENDAHULUAN... . 1
2. TINJAUAN PUSTAKA... 5
2.1. Kompetisi Interspesifik dan Relung ... 5
2.2. Siamang ... 7
2.2.1. Klasifikasi siamang ... 7
2.2.2. Morfologi siamang... 7
2.2.3. Habitat dan distribusi ... 9
2.2.4. Perilaku ... 10
2.2.5. Struktur sosial ... 11
2.2.6. Pakan ... 12
2.2.7. Peran ekologis ... 13
2.3. Mamalia Arboreal ... 14
2.3.1. Suku Tupaiidae ... 16
2.3.2. Suku Cercopithecidae ... 17
2.3.3. Suku Ursidae ... 17
2.3.4. Suku Mustelidae ... 17
2.3.5. Suku Sciuridae ... 18
2.4. Stasiun Penelitian Way Canguk, Taman Nasional Bukit Barisan Selatan ... 18
3. METODOLOGI PENELITIAN... 22
3.1. Waktu dan Lokasi Penelitian ... 22
3.2. Alat, Bahan, dan Subjek Penelitian ... 22
3.3. Cara Kerja ... 23
3.3.1. Pengambilan data ... 23
3.3.2. Analisis data ... 27
4. HASIL DAN PEMBAHASAN... 31
4.1. Tumpang-tindih Relung Berdasarkan Pakan ... 33
x
4.4. Partisi Relung antara Siamang dan Kompetitornya ... 50
4.4.1. Partisi Relung antara Siamang dan Simpai ... 51
4.4.2. Partisi Relung antara Siamang dan Bajing Kelapa ... 52
4.4.3. Partisi Relung antara Siamang dan Jelarang Hitam ... 53
5. KESIMPULAN DAN SARAN... 54
5.1. Kesimpulan ... 54
5.2. Saran ... 54
DAFTAR ACUAN... . 56
xi
Gambar 2.2.2. Siamang betina (kiri) dan siamang jantan (kanan) ... 8
Gambar 2.2.3. Peta distribusi Hylobatidae di Asia Tenggara ... 10
Gambar 2.2.6. Komposisi pakan siamang di TNBBS ... 12
Gambar 2.4(1) Lokasi Stasiun Penelitian Way Canguk, TNBBS ... 19
Gambar 2.4(2) Area penelitian tenggara (atas) dan area penelitian barat laut (bawah) di Way Canguk ... 21
Gambar 3.3.1. Wilayah jelajah beberapa kelompok siamang yang telah terhabituasi di area penelitian selatan ... 26
Gambar 4. Tiga spesies mamalia arboreal dengan frekuensi pertemuan tertinggi di area survei ... 32
Gambar 4.1(1) Diagram komposisi pakan tiga kelompok siamang subjek penelitian ... 33
Gambar 4.1(2) Diagram perbandingan komposisi pakan siamang, simpai, bajing kelapa, dan jelarang hitam ... 34
Gambar 4.2(1) Proyeksi variabel-variabel penggunaan habitat pada PC1 (sumbu-x) dan PC2 (sumbu-y) ... 39
Gambar 4.2(2) Ordinasipenggunaan habitat oleh empat spesies mamalia arboreal terhadap PC1 (sumbu-x) dan PC2 (sumbu-y) ... 40
Gambar 4.2(3) Boxplot ketiga variabel yang berkorelasi tinggi dengan PC1 dan PC2 ... 42
Gambar 4.3(1) Proyeksi variabel-variabel penggunaan habitat pada sumbu PC1 dan PC2 ... 46
xii
Tabel 2.3. Daftar famili dan spesies mamalia di Way Canguk beserta pola aktivitas dan arborealitas ... 14 Tabel 3.2. Komposisi kelompok siamang subjek penelitian ... 23 Tabel 3.3.2(1) Variabel penggunaan habitat yang dianalisis dengan PCA
untuk mengetahui tumpang-tindih relung di antara mamalia arboreal ... 28 Tabel 3.3.2(2) Variabel kondisi habitat yang dianalisis dengan PCA
untuk mengetahui perbedaan kondisi habitat yang
berhubungan dengan interaksi interspesifik ... 29 Tabel 4.1(1) Tabel ringkasan catatan spesies pakan dan tumpang-tindih
relung berdasarkan pemilihan pakan... 35 Tabel 4.1(2) Daftar spesies tumbuhan pakan siamang yang juga dimakan
oleh simpai, bajing kelapa, dan jelarang hitam ... 36 Tabel 4.2(1) Ringkasan variabel-variabel penggunaan habitat ... 37 Tabel 4.2(2) Hasil uji Kruskal-Wallis variabel penggunaan habitat
dengan faktor keempat spesies mamalia arboreal (siamang, simpai, bajing kelabu, dan jelarang hitam)... 41 Tabel 4.3(1) Frekuensi interaksi interspesifik antara siamang dengan
mamalia arboreal lainnya ... 44 Tabel 4.3(2) Hasil uji Kruskal-Wallis terhadap kedua PC dan variabel
yang berkorelasi tinggi dengan PC1 dan PC2 ... 47 Tabel 4.3(3) Perbandingan ukuran tubuh siamang, simpai, bajing
kelapa, dan jelarang hitam ... 48 Tabel 4.3(4) Perbandingan laju penanganan buah antara siamang dan
xiii
Lampiran 1. Daftar sepuluh mamalia arboreal yang terdapat di ruang
jelajah siamang ... 62 Lampiran 2. Daftar kategori dan spesies pakan siamang, simpai, bajing
kelapa, dan jelarang hitam ... 63 Lampiran 3. Perhitungan indeks tumpang-tindih Horn ... 66 Lampiran 4. Analisis variabel penggunaan habitat oleh siamang dan tiga
spesies mamalia arboreal menggunakan metode PCA ... 72 Lampiran 5. Analisis penggunaan habitat dengan uji Kruskal-Wallis dan
uji Wilcoxon ... 78 Lampiran 6. Analisis dengan metode PCA pada variabel kondisi habitat
yang memungkinkan terjadinya interaksi interspesifik ... 82 Lampiran 7. Analisis kondisi habitat yang memfasilitasi interaksi
1 Universitas Indonesia BAB 1
PENDAHULUAN
Kompetisi interspesifik terjadi jika dua spesies yang berbeda
menggunakan sumber daya yang sama dan hanya tersedia dalam jumlah terbatas.
Menurut MacArthur (1972: 21), mekanisme terjadinya kompetisi tidak terlihat
secara langsung, tetapi dapat disimpulkan dari bukti-bukti yang dapat ditemukan
di alam. Pada prinsipnya, dua spesies disebut berkompetisi jika peningkatan
populasi spesies yang satu berakibat pada penurunan populasi spesies yang lain
(Connell 1961: 715, 722; Krohne 2001: 223).
Subjek penelitian kompetisi interspesifik pada umumnya adalah
spesies-spesies berkerabat dekat yang hidup simpatrik, seperti yang dilaporkan antara lain
pada komunitas kadal di Amerika Utara, Afrika bagian selatan, dan Australia
Barat (Pianka 1973), burung dari famili Bucerotidae di Sumatra (Hadiprakarsa
2000), owa dengan siamang di Sumatra (Elder 2009), serta dua spesies marsupial
dari genus Didelphis di Amerika Selatan (Cáceres & Machado 2013). Hal
tersebut dilatarbelakangi oleh pandangan bahwa kompetisi terkuat terjadi di antara
spesies-spesies yang berkerabat dekat dengan kebutuhan sumber daya yang serupa
(Tokeshi 1999: 155).
Organisme tidak hanya hidup berdampingan dengan spesies yang
berkerabat dekat, melainkan juga dengan spesies-spesies lain dari taksa yang lebih
luas. Kompetisi interspesifik juga dapat terjadi pada spesies-spesies berkerabat
jauh yang memanfaatkan sumber daya serupa, baik pakan maupun ruang.
Fenomena itu pernah dilaporkan oleh Estrada & Coates-Estrada (1985: 34--35)
yang menyatakan bahwa monyet howling (Alouatta palliata) dan serangga
herbivora mengalami kompetisi interspesifik secara asimetris untuk
memperebutkan sumber daya pakan berupa daun di hutan hujan tropis Los
Tuxtlas, Meksiko. Marshall dkk. (2009: 173--175, 181) juga pernah melaporkan
bahwa kompetitor paling berpengaruh terhadap owa kalimantan (Hylobates
(Callosciurus prevostii) yang mengonsumsi buah muda yang menjadi pakan
primata tersebut.
Kompetisi interspesifik telah lama diduga berpengaruh terhadap
pembentukan struktur komunitas satwa. Walaupun hal tersebut masih sering
diperdebatkan hingga saat ini (Schoener 1983: 240; Krohne 2001: 279; Rohde
2013: 371), Beaudrot dkk. (2012: 182) secara tegas mengemukakan bahwa
interaksi interspesifik di dalam dan di antara kelompok-kelompok taksa yang
berbeda memiliki peran penting terhadap distribusi spesies dan struktur
komunitas. Dalam penelitiannya, Beaudrot dkk. (2012: 182) dan Beaudrot dkk.
(2013: 1061) menemukan bahwa struktur komunitas vertebrata di Kalimatan
dipengaruhi oleh competitive exclusion di antara spesies-spesies yang berkerabat
jauh. Oleh karena itu, penelitian mengenai kompetisi interspesifik di dalam suatu
komunitas penting dilakukan untuk memahami dinamika komunitas, terutama
untuk mengetahui bagaimana berbagai spesies berbagi sumber daya yang terbatas.
Siamang merupakan primata arboreal sejati yang menghuni hutan hujan
tropis Semenanjung Malaysia dan Sumatra. Sebagai primata arboreal sejati,
hampir seluruh aktivitas harian siamang dilakukan di lapisan tajuk tengah dan atas
(Gittins & Raemaekers 1980: 80--84; Markhamah 2007: 76). Siamang di
Semenanjung Malaysia dilaporkan lebih bersifat foliovora dibandingkan dengan
kerabatnya di Sumatra yang lebih banyak mengonsumsi buah (Gittins &
Raemaekers 1980: 90; Nurcahyo 1999: 24; O‟Brien dkk. 2003: 120). Komposisi
pakan siamang di Semenanjung Malaysia terdiri atas 43% daun, 22% buah
Ficus spp., 15% materi hewani, 14% buah pohon lain, dan 6% bunga (Gittins &
Raemaekers 1980: 90), sedangkan di Sumatra terdiri atas 52,07% buah, 42,62%
daun, dan 5,3% bunga (Nurcahyo 1999: 24).
Salah satu habitat siamang di Sumatra adalah Taman Nasional Bukit
Barisan Selatan (TNBBS). Kepadatan siamang di dalam kawasan konservasi
tersebut cukup tinggi, yaitu 2,23 kelompok/km2. O‟Brien dkk. (2004: 278--279) menyatakan bahwa TNBBS memiliki populasi siamang yang tinggi dibandingkan
lokasi lainnya di Semenanjung Malaysia dan Sumatra. Berdasarkan hasil
penelitian yang dilakukan di Stasiun Penelitian Way Canguk yang berada di
(Iqbal dkk. 2001: 13). Dari 26 famili mamalia yang terdapat di TNBBS,
setidaknya terdapat lima famili yang memiliki kemiripan cara hidup dengan
siamang, yaitu bersifat arboreal, frugivora, dan diurnal. Kelima famili mamalia
tersebut adalah Tupaiidae, Cercopithecidae, Ursidae, Mustelidae, dan Sciuridae
(Payne 1980: 266--267; Payne & Francis 1985; Iqbal dkk. 2001: 14).
Kompetisi terjadi ketika ketersediaan sumber daya yang dibutuhkan suatu
spesies berkurang akibat aktivitas spesies lain, baik secara eksploitatif maupun
interferensi. Oleh karena itu, fauna yang bersifat arboreal, baik parsial maupun
sejati, berpotensi mengalami kompetisi dengan siamang (Payne 1980: 261).
Tubuh siamang yang besar dan populasi siamang yang tinggi di TNBBS menjadi
dasar untuk menduga bahwa siamang merupakan kompetitor yang dominan.
Penelitian mengenai kompetisi interspesifik di antara tiga spesies primata paling
melimpah di TNBBS telah dilakukan oleh Elder (tidak dipublikasikan). Namun,
interaksi kompetitif antara siamang dan satwa selain kelompok primata belum
banyak diketahui. Penelitian ini dilakukan untuk mengkaji potensi kompetisi
antara siamang dengan spesies-spesies mamalia arboreal di Stasiun Penelitian
Way Canguk.
Penelitian noneksperimental di alam untuk mengetahui terjadinya
kompetisi pada umumnya menggunakan pendekatan indeks tumpang-tindih
relung walaupun indeks tersebut tidak secara tegas mengindikasikan kompetisi
(Estrada & Coates-Estrada 1985: 35; Marshall dkk. 2009:181; Cáceres &
Machado 2013: 13). Tiga parameter umum untuk mendefinisikan relung adalah
penggunaan habitat, pemilihan pakan, dan pola aktivitas harian (Holt 1987: 111).
Melalui penelitian ini, tumpang-tindih relung di antara mamalia arboreal akan
dikaji dari segi penggunaan habitat dan pemilihan pakan, kemudian dianalisis
kaitannya dengan kompetisi. Pola aktivitas harian tidak menjadi bahasan karena
spesies-spesies mamalia subjek penelitian merupakan satwa diurnal.
Tujuan penelitian ini adalah mengetahui apakah terjadi kompetisi antara
siamang dan mamalia arboreal lainnya serta mengetahui tumpang-tindih relung
berdasarkan penggunaan habitat dan pemilihan pakan di antara komunitas
mamalia arboreal di Stasiun Penelitian Way Canguk, TNBBS. Berdasarkan
antara siamang dan mamalia arboreal lainnya serta terdapat tumpang-tindih relung
di antara komunitas mamalia arboreal dari segi penggunaan habitat dan pemilihan
pakan.
Penelitian tentang kompetisi interspesifik dalam cakupan taksa yang luas
penting dilakukan untuk memahami peran kompetisi dalam pembentukan suatu
komunitas. Pengetahuan tentang dinamika komunitas penting dalam manajemen
kawasan konservasi, misalnya dalam menentukan area prioritas konservasi, luas
kawasan, dan lokasi pelepasliaran satwa yang dikembalikan ke hutan tanpa
mengganggu komunitas yang ada. Jika kompetisi merupakan faktor penting
dalam komunitas, pengambilan keputusan dalam manajemen konservasi
sebaiknya tidak hanya didasarkan pada faktor abiotik, melainkan juga
mempertimbangkan interaksi interspesifik.
5
BAB 2
TINJAUAN PUSTAKA
2.1. Kompetisi Interspesifik dan Pembagian Relung
Kompetisi merupakan bentuk interaksi yang menarik untuk dikaji karena
memberikan dampak negatif kepada pihak-pihak yang terlibat. Kompetisi terbagi
menjadi dua tipe, yaitu kompetisi intraspesifik dan kompetisi interspesifik.
Kompetisi intraspesifik terjadi di antara individu dari spesies yang sama,
sedangkan kompetisi interspesifik terjadi pada spesies yang berbeda. Kedua
bentuk kompetisi tersebut terjadi oleh penyebab yang sama, yaitu perebutan
sumber daya (Caughley & Sinclair 1994: 110 & 131). Bagian yang akan dikaji
pada penelitian ini adalah kompetisi interspesifik.
Kompetisi interspesifik adalah interaksi di antara individu dari spesies
yang berbeda akibat penggunaan sumber daya yang sama dan terbatas. Kompetisi
dapat juga terjadi saat sumber daya yang tersedia tidak terbatas jika suatu spesies
melukai spesies lain dalam proses pencarian sumber daya (Birch 1957: 6).
Menurut MacArthur (1972: 21), dua spesies disebut berkompetisi jika
peningkatan populasi suatu spesies berdampak pada penurunan populasi spesies
yang lain. Contoh proses yang memungkinkan antara lain suatu spesies
mengurangi persediaan makanan spesies lain, kedua spesies saling berkelahi, atau
keberadaan suatu spesies meningkatkan predator spesies lainnya. Bukti kompetisi
biasanya tidak teramati secara langsung di alam (MacArthur 1972: 21).
Definisi kompetisi memberikan empat implikasi penting. Pertama,
kompetisi berpengaruh terhadap kesintasan individu-individu yang terlibat.
Pengaruh tersebut dapat terjadi dalam bentuk penurunan kemampuan bertahan
hidup, pertumbuhan, atau reproduksi. Kedua, penggunaan sumber daya yang
sama tidak secara langsung mengindikasikan kompetisi jika tidak diketahui bahwa
sumber daya tersebut terbatas. Ketiga, dua spesies tidak berkompetisi jika tidak
memiliki kemampuan untuk memengaruhi ketersediaan sumber daya untuk
kompetisi eksploitatif atau kompetisi interferensi (Caughley & Sinclair 1994:
131--132). Dalam kompetisi eksploitatif, individu dari suatu spesies mengambil
alih sumber daya yang digunakan oleh spesies lain, sedangkan kompetisi
interferensi terjadi secara langsung melalui perkelahian atau produksi racun
(Schoener 1983: 257).
Kompetisi sudah menjadi topik bahasan ahli ekologi sejak setidaknya
delapan dekade yang lalu. Penelitian eksperimental klasik yang menjadi dasar
teori kompetisi dilakukan oleh Connell (1961). Dalam penelitian tersebut,
Connell menemukan bahwa kompetisi interspesifik di antara dua spesies teritip,
yaitu Balanus dan Chthamalus,merupakan faktor terpenting yang memengaruhi
populasi Chthamalus dibandingkan faktor lainnya seperti predator dan kondisi
fisik. Keberadaan Balanus meningkatkan mortalitas dan menurunkan
kemampuan reproduksi Chthamalus (Connell 1961: 715, 722).
Kompetisi interspesifik memiliki kaitan yang erat dengan relung (Wiens
1989: 147). Konsep relung mulai berkembang sejak tahun 1917 dan telah banyak
terdapat definisi relung. Namun, definisi yang paling sering digunakan adalah
definisi yang diajukan oleh Hutchinson yang mendefinisikan relung secara
matematis sebagai “n-dimensional hypervolume” (Hutchinson 1957: 416).
Berdasarkan perumusan oleh Hutchinson tersebut, relung dapat diartikan sebagai
seluruh kisaran kondisi biotik dan abiotik yang memungkinkan suatu spesies
mempertahankan ukuran populasi yang stabil (Wiens & Graham 2005: 519).
Relung dapat dibagi menjadi dua tipe, yaitu relung fundamental dan relung nyata.
Relung fundamental adalah kondisi abiotik yang memungkinkan suatu spesies
bertahan hidup, sedangkan relung nyata adalah bagian dari relung fundamental
yang ditempati suatu spesies jika dibatasi oleh interaksi dengan organisme lain
(Hutchinson 1957: 416--418).
Kompetisi sering kali dikaitkan dengan tumpang-tindih relung antara dua
spesies. Tumpang-tindih yang besar dapat mengindikasikan kompetisi
interspesifik. Namun, terdapat perdebatan di antara ahli ekologi mengenai
hubungan tumpang-tindih relung dengan kompetisi interspesifik. Tumpang-tindih
relung yang besar dapat juga terjadi akibat toleransi interspesifik, sedangkan
kemudian berusaha menempati relung yang berbeda untuk menghindari kompetisi
(Pianka 1974: 2141; Schoener 1983: 270). Walaupun demikian, indeks
tumpang-tindih relung sering digunakan sebagai pendekatan untuk memperkirakan
kompetisi (Estrada & Coates-Estrada 1985: 35; Marshall dkk. 2009:181; Cáceres
& Machado 2013: 13).
2.2. Siamang
Siamang merupakan salah satu spesies primata dari famili Hylobatidae,
yaitu salah satu famili dari kelompok primata yang bersifat diurnal, arboreal, dan
teritorial (Nowak 1999: 169, 171). Di antara kera lainnya, famili Hylobatidae
merupakan kelompok kera dengan ukuran tubuh terkecil sehingga disebut juga
kera kecil (lesser ape). Selain ukuran tubuh, perbedaan antara Hylobatidae
dengan kera lainnya--orangutan, gorila, dan simpanse--adalah gaya hidup yang
sepenuhnya arboreal, memiliki bantalan duduk (ischial callosities), dan tidak
membuat sarang untuk tidur (Bartlett 1999: 44).
2.2.1. Klasifikasi Siamang
Siamang dikelompokkan ke dalam genus Hylobates bersama owa bertubuh
kecil lainnya oleh beberapa ahli (Miller 1933: 159; Napier & Napier 1985: 161),
tetapi Brandon-Jones dkk. (2004: 153) mengelompokkannya ke dalam genus
tersendiri, yaitu Symphalangus. Genus Symphalangus hanya terdiri atas satu
spesies, yaitu S. syndactylus, dengan dua subspesies. Kedua subspesies siamang
dibedakan berdasarkan distribusinya. Symphalangus syndactylus syndactylus
Raffles(1821) terdistribusi di Sumatra, Indonesia, sedangkan S. syndactylus
continentis Thomas (1908) terdistribusi di Semenanjung Malaysia.
2.2.2. Morfologi Siamang
Siamang mudah dibedakan dengan spesies Hylobatidae lain karena
siamang merupakan spesies yang memiliki ukuran tubuh terbesar. Rentang
tangan siamang dapat mencapai 1,5 m, panjang tubuh termasuk kepala berkisar
antara 750--900 mm, dan berat tubuh sekitar 8--13 kg (Nowak 1999: 168). Tubuh
siamang ditutupi oleh rambut berwarna hitam, tetapi bagian kantung udara di
bawah dagu tidak tertutupi rambut (Napier & Napier 1985: 161).
Siamang tidak menunjukkan adanya dimorfisme seksual. Warna rambut,
ukuran tubuh, dan panjang gigi taring siamang jantan dan betina sama (Bartlett
1999: 45). Baik siamang jantan maupun betina juga memiliki kantung udara yang
berfungsi untuk vokalisasi. Walaupun tidak terdapat dimorfisme seksual, individu
jantan dan betina dapat dibedakan (Gambar 2.2.2.). Individu jantan memiliki
rambut skrotal, sedangkan individu betina tidak (Prasetyaningrum 2001: 30).
Gambar 2.2.2. Siamang betina (kiri) dan siamang jantan (kanan).
Keterangan:
Jenis kelamin siamang dapat dibedakan berdasarkan ada atau tidaknya rambut skrotal. Individu jantan memiliki rambut skrotal (a) di antara kedua kakinya.
[Sumber: Dokumentasi pribadi]
(a)
Seperti spesies dari famili Hylobatidae lainnya, siamang memiliki
ekstremitas anterior yang lebih panjang daripada ekstremitas posterior sebagai
adaptasi untuk pergerakan brakiasi. Di antara spesies-spesies Hylobatidae
lainnya, siamang memiliki intermembral index terbesar, yaitu 145--152 (Miller
1933: 159). Ciri khas siamang yang tidak dimiliki oleh spesies Hylobatidae
lainnya adalah selaput antarjari yang menghubungkan jari kaki kedua dan ketiga
(LBN-LIPI 1982: 85; Nowak 1999: 168).
2.2.3. Habitat dan Distribusi
Siamang terdapat di Sumatra dan Semenanjung Malaysia (LBN-LIPI
1982: 85; Nowak 1999: 168). Siamang pada umumnya dapat ditemukan di
dataran rendah sampai daerah pegunungan setinggi 2000 m (LBN-LIPI 1982: 85).
Menurut Wilson & Wilson (1976: 17), siamang di Sumatra terdapat di rangkaian
Pegunungan Bukit Barisan yang terletak memanjang di bagian barat Sumatra
(Gambar 2.2.3.) dan lebih menyukai hutan dataran rendah dan perbukitan.
Siamang simpatrik dengan dua kerabat dekatnya, yaitu Hylobates agilis
dan H. lar, di rangkaian pegunungan bagian barat Sumatra, tetapi siamang tidak
ditemukan di dataran rendah bagian timur Sumatra. Siamang simpatrik dengan H.
agilis dari bagian selatan Sumatra sampai Simpangkiri, Aceh, dan Sungai
Wampu, Sumatra Utara, sedangkan daerah simpatrik siamang dengan H. lar
meliputi bagian utara dari Simpangkiri dan Sungai Wampu (Wilson & Wilson
1976: 17). Hasil studi O‟Brien (2004: 276--278) menunjukkan bahwa kepadatan
siamang berkorelasi negatif dengan kepadatan Hylobatidae yang simpatrik
dengannya. Kepadatan siamang tertinggi terdapat pada hutan dataran rendah
(< 300 mdpl), terendah di hutan ketinggian sedang, kemudian meningkat kembali
di hutan pegunungan. Sementara itu, kepadatan H. agilis tertinggi terdapat di
hutan ketinggian sedang. Kepadatan siamangmenunjukkan peningkatan dari
lintang utara ke selatan, yaitu dari Semenanjung Malaysia ke Sumatra, sementara
Gambar 2.2.3. Peta distribusi Hylobatidae di Asia Tenggara.
Keterangan:
Siamang terdistribusi di bagian barat Sumatra dan Semenanjung Malaysia (warna hijau).
[Sumber: www.gibbons.de]
2.2.4. Perilaku
Siamang merupakan primata diurnal, yaitu melakukan sebagian besar
aktivitas pada siang hari. Aktivitas harian siamang dimulai antara pukul
06.00--07.00. Secara umum, aktivitas harian siamang dapat dibagi ke dalam lima
kategori, yaitu istirahat, makan, mencari makan, bergerak, dan perilaku sosial.
Aktivitas utama siamang adalah istirahat (56,80%) dan makan (25,96%).
Siamang paling aktif mencari makan pada pagi hari (07.00--08.00) dan sore hari
(14.00--16.00). Aktivitas bergerak terbagi ke dalam empat tipe, yaitu brakiasi,
memanjat, berjalan, dan melompat. Sebagian besar pergerakan siamang (81,64%)
dilakukan dengan cara brakiasi. Aktivitas sosial terdiri atas menggaruk individu
jarang terjadi di antara individu dewasa dengan jenis kelamin yang berbeda.
Namun, individu dewasa terkadang menunjukkan perilaku agresi terhadap
individu pradewasa yang memiliki jenis kelamin sama (Chivers & Raemaekers
1980: 229; Nurcahyo 1999: 37).
2.2.5. Struktur sosial
Siamang hidup dalam kelompok keluarga (selanjutnya hanya disebut
kelompok) yang bersifat teritorial. Satu kelompok siamang pada umumnya terdiri
atas empat individu yang beranggotakan pasangan jantan dan betina dewasa serta
beberapa anak yang belum mandiri (Chivers 1974: 274; Gittins & Raemaekers
1980: 68). Siamang dan juga semua spesies dari famili Hylobatidae memiliki
ukuran kelompok yang kecil untuk mengurangi kompetisi antarindividu saat
mencari makanan. Semakin besar ukuran kelompok, kompetisi antarindividu juga
semakin besar. Sifat teritorial siamang berkaitan dengan distribusi sumber pakan
yang bervariasi dari waktu ke waktu. Walaupun mempertahankan suatu teritori
membutuhkan energi yang besar, siamang mendapatkan keuntungan berupa
jaminan persediaan makanan sepanjang tahun (Gittins & Raemaekers 1980:
101--103).
Berbeda dengan primata dan bahkan sebagian besar mamalia lain, induk
jantan siamang terlibat dalam pengasuhan bayi. Menurut Gittins & Raemaekers
(1980: 70), selama satu tahun sejak kelahiran, bayi siamang diasuh oleh induk
betina, kemudian pada tahun kedua, bayi diasuh oleh induk jantan. Perilaku
tersebut berbeda dengan kerabat simpatriknya, yaitu owa ungko (Hylobates agilis)
yang hanya menunjukkan pengasuhan bayi oleh induk betina.
Siamang pada umumnya bersifat monogami. Namun, beberapa kelompok
poliandri ditemukan pada populasi siamang di Way Canguk. Menurut Lappan
(2008: 1314--1315), keberadaan individu jantan sekunder tidak berhubungan
dengan peningkatan total asuhan yang diterima bayi serta ikatan antara jantan dan
induk betina. Poliandri pada siamang diduga merupakan mekanisme untuk
mengurangi interval antarkelahiran pada siamang betina, dengan kata lain
2.2.6. Pakan
Pakan siamang dapat dibagi menjadi tiga kategori, yaitu bagian
reproduktif tumbuhan (bunga dan buah), bagian vegetatif tumbuhan (daun), serta
materi hewani (Gittins & Raemaekers 1980: 88). Siamang sering dideskripsikan
sebagai foliovora (Gittins & Raemaekers 1980: 100--101; Napier & Napier 1985:
161; Supriatna & Wahyono 2000: 294), tetapi siamang di Sumatra diketahui lebih
bersifat frugivora (Nurcahyo 1999: 37). Komposisi pakan siamang di TNBBS
(Gambar 2.2.6.) didominasi oleh buah dengan persentase sebesar 52,07%
(Nurcahyo 1999: 24).
Gambar 2.2.6. Komposisi pakan siamang di TNBBS.
[Sumber: Nurcahyo 1999:24]
Siamang menunjukkan preferensi terhadap buah Ficus spp. Siamang
cenderung memakan lebih banyak buah Ficus spp. pada pagi hari saat memulai
aktivitas. Hal tersebut diduga karena buah Ficus spp. menyediakan nutrisi yang
melimpah dan siap dicerna untuk memenuhi kebutuhan nutrisi setelah berpuasa
sepanjang malam (Gittins & Raemaekers 1980: 90--93). Persentase buah Ficus
Buah Daun Bunga
Tumbuhan lain 0.45% 0% 0%
Liana 1.28% 11.88% 0%
Pohon lain 18.91% 19.75% 5.30%
Ficus spp. 31.43% 10.99% 0%
0.00% 10.00% 20.00% 30.00% 40.00% 50.00% 60.00%
Tumbuhan lain
Liana
Pohon lain
Ficus spp.Ficus spp.
spp. dalam komposisi pakan siamang adalah 31,43% (Nurcahyo 1999: 24).
Spesies Ficus yang dikonsumsi siamang antara lain F. albipila, F. altissima, F.
caulocarpa, F. depressa, F. drupacea, F. elastica, F. stupenda, F. sundaica, F.
variegata, dan F. virens (Nurcahyo 1999: 24, 27; Rusmanto 2001: 18).
Menurut Nurcahyo (1999: 27) spesies tumbuhan selain Ficus spp. yang
diketahui merupakan pakan siamang adalah Actinodaphne nigrescens, Aglaia
odoratissima, Alseodaphne nigrescens, Anamirta cocculus, Antocephalus sp.,
Atuna racemosa, Bauhinia sp., Beilschmiedia lucida, Bridelia monoica, Cananga
odorata, Cryptocarya ferrea, Daemonorops sp., Diospyros truncata,
Dracontomelon dao, Durio sp., Dysoxylum caulostachyum, Eugenia jamboloides,
Garcinia sp., Horsfieldia bracteosa, Leuconotis eugenifolia, Michelia champaca,
Polyalthia spp., Popowia pisocarpa, Pterospermum sp., Shorea javanica,
Siphonodon celastrineus, Stelechocarpus burahol, dan Xerospermum
noronhianum. Sementara itu, Rusmanto (2001:18--20) mendata 43 spesies
tumbuhan yang dikonsumsi buahnya oleh siamang. Spesies-spesies tersebut
termasuk dalam famili Alangiaceae, Annonaceae, Arecaceae, Boraginaceae,
Clusiaceae, Combretaceae, Ebenaceae, Euphorbiaceae, Lauraceae, Meliaceae,
Menispermaceae, Moraceae, Myristicaceae, Myrtaceae, Olacaceae, Rhamnaceae,
Rutaceae, Sapotaceae, Sapindaceae, dan Vitaceae.
Puncak aktivitas makan siamang bersifat bimodal, yaitu antara pukul
07.00--08.00 dan 14.00--16.00. Aktivitas makan di pagi hari diduga berfungsi
untuk memenuhi kebutuhan energi untuk beraktivitas di siang hari. Sementara
itu, makanan yang dimakan pada sore hari berfungsi sebagai cadangan energi
untuk malam hari (Nurcahyo 1999: 26, 37).
2.2.7. Peran Ekologis
Siamang merupakan pemencar biji yang efektif. Siamang, seperti
Hylobatidae lainnya, tidak mengunyah atau menghancurkan biji dari buah yang
dimakan. Biji ditelan, kemudian dibuang secara utuh dalam bentuk feses
beberapa ratus meter dari pohon pakan (Gittins & Raemaekers 1980: 88).
endozoochory (90,7%), sedangkan sebagian kecil akan hancur selama proses
pencernaan. Jarak pemencaran biji oleh siamang berkisar antara 0 hingga 632 m
dari pohon pakan (Rusmanto 2001: 17--25).
2.3. Mamalia Arboreal
Mamalia arboreal adalah mamalia yang mencari makan di tajuk hutan,
baik pada bagian bawah, tengah, maupun atas (Beaudrot dkk. 2013: 1056).
Mamalia arboreal meliputi berbagai famili, antara lain Pteropodidae, Tupaiidae,
Lorisidae, Cercopithecidae, Hylobatidae, Sciuridae, Muridae, Ursidae, dan
Viverridae (Payne 1980: 266--267). Mamalia arboreal yang bersifat nokturnal
terhindar dari kompetisi secara langsung dengan siamang akibat pembagian
relung secara temporal. Walaupun demikian, mamalia nokturnal dapat
menyebabkan kompetisi karena aktivitas makan mamalia nokturnal dapat
mengurangi ketersediaan sumber daya bagi mamalia arboreal diurnal (Estrada &
Coates-Estrada 1985: 35). Tabel 2.3. berisi daftar famili dan spesies seluruh
mamalia yang terdapat di Way Canguk. Dari tabel tersebut, terlihat
spesies-spesies mamalia arboreal dan diurnal yang menjadi subjek penelitian. Berikut
adalah paparan singkat famili mamalia arboreal diurnal selain Hylobatidae yang
terdapat di Way Canguk, TNBBS.
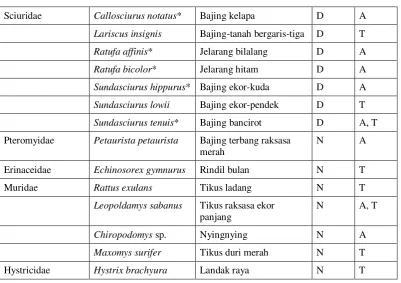
Tabel 2.3. Daftar famili dan spesies mamalia di Way Canguk beserta pola aktivitas dan arborealitas.
Famili Spesies Nama Indonesia N/D A/T
Manidae Manis javanica Trenggiling leusing N T
Tupaiidae Tupaia tana Tupai tanah D T
Tupaia minor* Tupai kecil D A, T
Tupaia glis* Tupai akar D A, T
Cynocephalidae Cynocephalus
variegatus
Kubung malaya N A
Pteropodidae Cynopterus brachyotis Codot krawar N A
Pteropus vampyrus Kalong besar N A
Hipposideridae Hipposideros diadema Barong besar N A
Tabel 2.3. (lanjutan)
Tarsiidae Tarsius bancanus Krabuku ingkat N A
Lorisidae Nycticebus coucang Kukang bukang N A
Cercopithecidae Macaca fascicularis* Monyet kra D A
Macaca nemestrina* Monyet beruk D A, T
Presbytis melalophos* Simpai/cecah D A
Trachypithecus
cristatus*
Lutung kelabu D A
Hylobatidae Hylobates agilis* Owa ungko D A
Symphalangus
Viverridae Arctitis binturong Binturong N A, T
Arctogalidia trivirgata Musang akar N A
Rhinocerotidae Dicerorhinus
Tabel 2.3. (lanjutan)
Sciuridae Callosciurus notatus* Bajing kelapa D A
Lariscus insignis Bajing-tanah bergaris-tiga D T
Ratufa affinis* Jelarang bilalang D A
Ratufa bicolor* Jelarang hitam D A
Sundasciurus hippurus* Bajing ekor-kuda D A
Sundasciurus lowii Bajing ekor-pendek D T
Sundasciurus tenuis* Bajing bancirot D A, T
Pteromyidae Petaurista petaurista Bajing terbang raksasa
merah
N A
Erinaceidae Echinosorex gymnurus Rindil bulan N T
Muridae Rattus exulans Tikus ladang N T
Leopoldamys sabanus Tikus raksasa ekor panjang
N A, T
Chiropodomys sp. Nyingnying N A
Maxomys surifer Tikus duri merah N T
Hystricidae Hystrix brachyura Landak raya N T
Keterangan: N = nokturnal D = diurnal A = arboreal T = terestrial
* = spesies yang menjadi subjek penelitian
[Sumber: Payne 1980: 266—267; Payne & Francis 1985; Iqbal dkk. 2001: 14]
2.3.1. Famili Tupaiidae
Mamalia yang tergolong Tupaiidae memiliki morfologi seperti bajing,
tetapi dengan moncong yang lebih panjang. Tupaiidae memiliki tungkai depan
dan belakang dengan ukuran yang hampir sama (Lawlor 1979: 73). Famili ini
merupakan famili endemik di kawasan Indomalaya. Sebagian besar spesies yang
termasuk famili Tupaiidae bersifat arboreal dan diurnal. Tupaiidae tergolong
omnivora karena memakan buah dan juga invertebrata (Lawlor 1979: 73; Corbet
& Hill 1992: 46). Tiga spesies Tupaiidae terdapat di Way Canguk, yaitu Tupaia
tana, T. minor, dan T. glis. Dari ketiga spesies tupaiidae tersebut, hanya T. tana
2.3.2. Famili Cercopithecidae
Cercopithecidae terdapat di kawasan Indomalaya (enam spesies) dan
Afrika tropis (sepuluh spesies). Semua spesies merupakan primata diurnal dan
arboreal. Secara umum, Cercopithecidae merupakan omnivora. Buah-buahan
merupakan komponen utama pakan dengan tambahan materi hewani dan
dedaunan (Corbet & Hill 1992: 165). Empat spesies Cercopithecidae yang
terdapat di Way Canguk adalah Macaca fascicularis, M. nemestrina, Presbytis
melalophos, dan Trachypithecus cristatus.
2.3.3. Famili Ursidae
Hanya terdapat satu spesies Ursidae di Way Canguk, yaitu beruang madu
(Helarctos malayanus). Mamalia ini aktif pada malam dan siang hari serta
merupakan omnivora yang memakan sarang lebah, rayap, hewan kecil, dan buah.
Beruang tersebut bersifat terestrial dan arboreal (Payne & Francis 1985:
275--276).
2.3.4. Famili Mustelidae
Mustelidae memiliki tubuh yang memanjang, berkaki pendek, dan bersifat
terestrial (Corbet & Hill 1992: 194). Famili ini beranggotakan berang-berang,
musang, dan luwak. Dari lima spesies yang terdapat di Way Canguk, hanya
Martes flavigula yang bersifat diurnal dan arboreal. Martes flavigula pada
dasarnya diurnal, tetapi terkadang beraktivitas di malam hari. Spesies tersebut
merupakan generalis yang memakan berbagai vertebrata dan invertebrata kecil,
sarang lebah, serta nektar (Payne & Francis 1985: 276--277).
2.3.5. Famili Sciuridae
Famili Sciuridae terdiri atas tiga kelompok, yaitu bajing terbang, bajing
tungkai-tungkainya yang berfungsi untuk gliding. Pada umumnya, bajing terbang bersifat
nokturnal. Berbeda dengan bajing terbang, bajing pohon bersifat diurnal dan
tidak memiliki membran gliding. Sebagai adaptasi terhadapcara hidup arboreal,
bajing terbang dan bajing pohon memiliki kepala berbentuk bulat dengan mata
yang besar, cakar melengkung dan tajam, ekor panjang, dan pergelangan kaki
dengan kemampuan rotasi yang sangat baik. Bajing tanah bersifat terestrial
sehingga memiliki adaptasi yang berbeda dengan bajing terbang dan bajing
pohon, yaitu kepala yang relatif datar, cakar lurus, ekor pendek, dan pergelangan
kaki yang tidak terspesialisasi. Bajing tanah pada umumnya bersifat diurnal.
Sciuridae memakan daun, biji, buah, kulit kayu, kacang, resin, jamur, dan nektar.
Beberapa spesies juga memakan serangga (Lawlor 1979: 160). Dari delapan
spesies Sciuridae di Way Canguk, terdapat lima spesies yang merupakan
kompetitor potensial bagi siamang, yaitu Callosciurus notatus, Ratufa affinis,
R. bicolor, Sundasciurus hippurus, dan S. tenuis (Iqbal dkk. 2001: 14).
2.4. Stasiun Penelitian Way Canguk, Taman Nasional Bukit Barisan Selatan
Taman Nasional Bukit Barisan Selatan (TNBBS), diresmikan pada tahun
1981, merupakan salah satu taman nasional yang terletak di Pulau Sumatra.
Taman nasional tersebut meliputi kawasan seluas 3.568 km2 dan terletak pada dua provinsi, yaitu Lampung dan Bengkulu. Kawasan taman nasional berbentuk
memanjang dengan batas kawasan sepanjang lebih dari 700 km. Hal tersebut
menyebabkan perambahan untuk penebangan dan perkebunan lebih mudah
terjadi. Letak geografis TNBBS adalah 4o31'--5o57' LS dan 103o34'--104o43' BT. TNBBS terfragmentasi menjadi tiga blok besar hutan akibat pembangunan dua
jalan lintas di sebelah utara dan selatan taman nasional. TNBSS merupakan
habitat penting bagi mamalia besar Sumatra seperti harimau, badak, dan gajah
Gambar 2.4(1) Lokasi Stasiun Penelitian Way Canguk, TNBBS.
Keterangan:
Stasiun Penelitian Way Canguk terdiri atas dua area penelitian--area barat laut dan tenggara--yang mencakup wilayah seluas 900 ha dengan tipe ekosistem hutan hujan tropis dataran rendah.
[Sumber: WCS-IP 2001: 7]
Stasiun Penelitian Way Canguk terletak di bagian selatan TNBBS,
meliputi area seluas 900 ha, termasuk 165 ha yang terbakar akibat El Niño
-Southern Oscillation (ENSO) pada tahun 1997 (Gambar 2.4(1)). Letak geografis
stasiun penelitian ini adalah 5o39'325'' LS dan 104o24'21'' BT dengan ketinggian 0--100 mdpl. Stasiun Penelitian Way Canguk diresmikan pada bulan Maret 1997.
Tujuan pembangunan stasiun penelitian adalah sebagai tempat penelitian lapangan
jangka panjang dan pelatihan lapangan (WCS-IP 2001: 7).
Area penelitian di Way Canguk terbagi menjadi dua area yang dipisahkan
oleh Sungai Way Canguk, yaitu area di bagian barat laut dengan luas 200 ha dan
area di bagian tenggara dengan luas 600 ha (Gambar 2.4(2)). Pada area
penelitian, terdapat jalur-jalur melintang dan membujur dengan jarak antarjalur
200 m. Sebagian besar area merupakan hutan primer yang masih baik. Dalam
area penelitian terdapat pula blok-blok hutan yang mengalami gangguan akibat
Berdasarkan karakteristik vegetasi, hutan di area penelitian diklasifikasikan
menjadi empat tipe habitat: hutan yang masih utuh dengan tajuk pohon tertutup
dan banyak pohon berukuran besar; hutan yang masih utuh dengan tajuk pohon
tertutup dan banyak pohon berukuran kecil; hutan yang terganggu dengan tajuk
pohon terbuka dan beberapa pohon berukuran kecil; serta hutan terganggu dengan
tajuk pohon terbuka dan beberapa pohon berukuran besar (Hadiprakarsa &
O‟Brien 2000 lihat WCS-IP 2001: 7).
Menurut Iqbal dkk. (2001: 13--16), Way Canguk merupakan habitat bagi
56 spesies mamalia yang tergolong ke dalam 26 famili. Primata yang umum
dijumpai adalah owa ungko (H. agilis), siamang (S. syndactylus), cecah (Presbytis
melalophos), dan beruk (Macaca nemestrina), sedangkan primata yang jarang
dijumpai adalah lutung kelabu (Trachypithecus cristatus) dan monyet ekor
panjang (Macaca fascicularis). Beberapa spesies ungulata yang ada di Way
Canguk adalah kijang muncak (Muntiacus muntjak), rusa sambar (Cervus
unicolor), babi (Sus sp.), pelanduk kecil (Tragulus javanicus), dan pelanduk napu
(T. napu). Terdapat pula mamalia besar seperti gajah asia (Elephas maximus),
tapir tenuk (Tapirus indicus), dan badak sumatra (Dicerorhinus sumatrensis).
Keanekaragaman avifauna di Way Canguk meliputi kurang lebih 207
spesies burung dari 41 famili. Spesies burung yang umum adalah dari famili
Timaliidae (18 spesies), Pycnonotidae (17 spesies), Cuculidae (15 spesies),
Picidae (12 spesies), dan Nectarinidae (12 spesies). Selain itu, terdapat delapan
spesies burung rangkong (famili Bucerotidae) di Way Canguk (Prasetyaningrum
& Hadiprakarsa 2001: 19).
Way Canguk diketahui memiliki lebih dari 330 spesies pohon. Hutan di
Way Canguk merupakan mosaik antara hutan hutan primer, hutan sekunder, dan
hutan yang rusak. Hutan primer ditandai oleh spesies-spesies Dipterocarpaceae
seperti Dipterocarpus spp., Shorea spp., dan Anisoptera costata serta memiliki
tajuk yang tinggi dan rapat. Hutan sekunder pada umumnya dicirikan oleh
Bombax valetonii, Tetrameles nudiflora, dan Octomeles sumatrana yang tidak
dapat tumbuh maksimal di bawah naungan. Way Canguk merupakan habitat bagi
setidaknya delapan spesies Ficus spp. (Prasetyaningrum & Hadiprakarsa 2001:
Gambar 2.4(2) Area penelitian tenggara (atas) dan area penelitian barat laut (bawah) di Way Canguk.
Keterangan:
Perpotongan jalur-jalur melintang dan membujur membentuk petak-petak berukuran 200 x 200 m. Sistem jalur tersebut memungkinkan pencatatan koordinat secara manual berdasarkan sistem Kartesius.
22 Universitas Indonesia BAB 3
METODOLOGI PENELITIAN
3.1. Waktu dan Lokasi Penelitian
Penelitian dilakukan selama tiga bulan pada Februari--April 2014 di
Stasiun Penelitian Way Canguk, TNBBS, Lampung. Kawasan terbagi menjadi
dua area, yaitu area penelitian barat laut dan tenggara (lebih sering disebut area
selatan dan utara), tetapi pengambilan data hanya dilakukan di area selatan. Pada
area tersebut, terdapat beberapa kelompok siamang yang telah terhabituasi dengan
manusia.
3.2. Alat, Bahan, dan Subjek Penelitian
Alat yang digunakan dalam penelitian adalah kamera [Canon PowerShot
SX40 HS], teropong binokular [Nikon], rangefinder [Nikon], dan jam tangan
digital [Eiger LS-88]. Bahan yang digunakan adalah pensil, lembar pengamatan,
pita penanda, dan kantung plastik zip lock. Subjek penelitian adalah tiga
kelompok siamang yang terdapat di area penelitian selatan dan mamalia arboreal
lain yang ditemukan di dalam ruang jelajah kelompok siamang. Kelompok
siamang yang dipilih merupakan kelompok yang seluruh ruang jelajahnya hutan
primer untuk menghindari bias akibat perbedaan struktur habitat. Komposisi
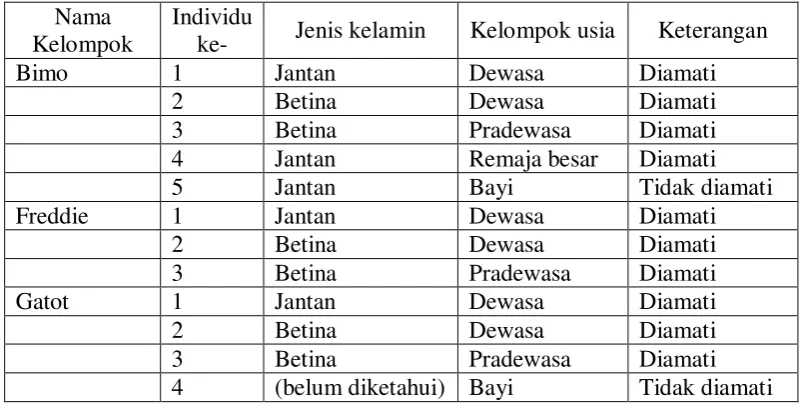
kelompok siamang yang dipilih sebagai subjek penelitian dapat dilihat pada Tabel
3.2. Individu yang masih bayi tidak diamati karena masih bergantung pada
Tabel 3.2. Komposisi kelompok siamang subjek penelitian.
Nama Kelompok
Individu
ke- Jenis kelamin Kelompok usia Keterangan
Bimo 1 Jantan Dewasa Diamati
2 Betina Dewasa Diamati
3 Betina Pradewasa Diamati
4 Jantan Remaja besar Diamati
5 Jantan Bayi Tidak diamati
Freddie 1 Jantan Dewasa Diamati
2 Betina Dewasa Diamati
3 Betina Pradewasa Diamati
Gatot 1 Jantan Dewasa Diamati
2 Betina Dewasa Diamati
3 Betina Pradewasa Diamati
4 (belum diketahui) Bayi Tidak diamati
Keterangan:
Individu yang diamati adalah individu yang sudah tidak bergantung pada induk. Individu ke-4 pada kelompok Gatot belum diketahui jenis kelaminnya.
3.3. Cara Kerja
3.3.1. Pengambilan data
Pengambilan data pada penelitian ini terbagi menjadi dua tahap, yaitu
pengamatan perilaku siamang dan survei mamalia arboreal. Data yang akan
diperoleh dari pengamatan perilaku siamang adalah komposisi pakan siamang,
penggunaan habitat, dan interaksi dengan mamalia arboreal lain. Data yang akan
diperoleh dari survei mamalia arboreal adalah frekuensi pertemuan setiap spesies
mamalia arboreal yang berada di ruang jelajah siamang, komposisi pakan, dan
penggunaan habitat.
1. Pengamatan perilaku siamang
Metode focal instantaneous sampling digunakan untuk pengamatan
perilaku siamang. Focal sampling merupakan metode yang dilakukan dengan
mengamati dan mencatat perilaku satu individu saja pada waktu yang ditentukan.
setiap interval waktu tertentu (Martin & Bateson 2007: 48--61). Selain itu,
metode ad libitum digunakan untuk mencatat interaksi interspesifik antara
siamang dan mamalia arboreal lainnya.
Subjek penelitian adalah tiga kelompok siamang yang seluruh ruang
jelajahnya hutan primer (Tabel 3.2.). Ketiga kelompok tersebut telah terhabituasi
terhadap manusia karena telah menjadi subjek penelitian perilaku sejak tahun
1998 (Nurcahyo 1999). Setiap kelompok siamang diikuti selama empat hari
berturut-turut. Satu hari sebelum memulai pengamatan perilaku, pengamat
mencari kelompok siamang dan mengikutinya hingga siamang kembali ke pohon
tidur. Hal tersebut dilakukan agar pencatatan perilaku keesokan harinya dapat
dimulai saat siamang baru memulai aktivitas, yaitu sekitar pukul 06.00.
Pengamatan dilakukan sampai siamang kembali ke pohon tidur dan berhenti
beraktivitas, yaitu sekitar pukul 17.00 (Chivers 1974: 23--24). Pengulangan
pengamatan perilaku dilakukan satu kali dengan urutan kelompok yang sama.
Waktu pengamatan dibagi menjadi interval-interval dengan periode satu
jam. Setiap satu jam, individu focal diganti secara sistematis. Hal tersebut
dilakukan untuk menghindari bias yang diakibatkan oleh faktor lingkungan,
seperti hari dengan curah hujan tinggi dan hari cerah. Dalam satu jam interval
pengamatan, perilaku individu focal dicatat setiap interval waktu lima menit.
Perilaku yang dicatat diklasifikasikan ke dalam lima kategori yang diadaptasi dari
Gittins & Raemaekers (1980: 96--97) dan O‟Brien & Kinnaird (1997: 326), yaitu
istirahat, makan, bergerak, vokalisasi, dan sosial.
Dua komponen utama dalam pengamatan perilaku siamang yang berfungsi
untuk menjawab pertanyaan penelitian adalah informasi mengenai penggunaan
habitat dan pemilihan pakan. Data penggunaan habitat dan pemilihan pakan akan
dicatat bersamaan dengan pengamatan perilaku. Variabel yang dicatat untuk
menjelaskan penggunaan habitat adalah tipe substrat, diameter substrat,
ketinggian siamang dari tanah, ketinggian pohon, jarak dari pusat tajuk, lebar
tajuk, fenofase pohon, skor fenofase, strata, dan kategori tajuk. Variabel
pemilihan pakan yang dicatat adalah kategori makanan dan spesies yang dimakan.
Jika memungkinkan, waktu yang dibutuhkan untuk menangani makanan juga
2. Survei mamalia arboreal
Survei mamalia arboreal dilakukan dengan metode transek garis. Metode
tersebut dilakukan dengan menyusuri jalur berbentuk garis lurus dan mencatat
jarak tegak lurus dari hewan yang ditemui ke garis transek (Rudran dkk. 1996:
89). Data yang dicatat adalah waktu pertemuan, spesies mamalia, jumlah
individu, jarak tegak lurus, koordinat, perilaku, dan penggunaan habitat. Jika
individu yang ditemukan sedang makan, kategori makanan dan spesies yang
dimakan dicatat. Waktu penanganan makanan dicatat jika memungkinkan.
Variabel penggunaan habitat dan pemilihan pakan sama seperti pada pengamatan
siamang.
Survei transek garis dilakukan di antara pukul 06.00--12.00 karena
sebagian besar mamalia arboreal lebih aktif pada pagi hari daripada sore hari.
Area survei mencakup wilayah jelajah ketiga kelompok siamang subjek
penelitian. Survei hanya dilakukan pada transek hutan primer. Pembatasan
wilayah pengamatan pada wilayah jelajah primata subjek penelitian juga telah
dilakukan oleh Estrada & Coates-Estrada (1985: 29). Jalur transek dapat dilihat
pada Gambar 3.3.1. Wilayah survei mencakup tiga jalur transek melintang
Gambar 3.3.1. Wilayah jelajah beberapa kelompok siamang yang telah terhabituasi di area penelitian selatan.
[Sumber: telah diolah kembali dari Prasetyaningrum 2001: 34 dan WCS-IP 2001: 6] Legenda
Daerah bekas kebakaran tahun 1997
Ruang jelajah siamang subjek penelitian
Ruang jelajah kelompok siamang yang tidak diamati
3.3.2. Analisis data
Analisis data dilakukan menggunakan dua metode utama, yaitu analisis
tumpang-tindih relung dan analisis komponen utama (PCA, Principal Component
Analysis). Analisis tumpang-tindih relung dilakukan dengan menghitung besar
indeks tumpang-tindih Horn. Rumus untuk menghitung indeks tersebut adalah:
𝑅
𝑜=
𝑝𝑖𝑗+𝑝𝑖𝑘 log 𝑝𝑖𝑗+𝑝𝑖𝑘 − 𝑝𝑖𝑗log 𝑝𝑖𝑗− 𝑝𝑖𝑘log𝑝𝑖𝑘
2 log 2
Keterangan:
Ro = Indeks tumpang-tindih Horn untuk spesies j dan k
pij = Proporsi sumber daya i dari sumber daya total yang digunakan spesies j
(spesies mamalia arboreal selain siamang)
pik = Proporsi sumber daya i dari sumber daya total yang digunakan spesies k
(siamang)
Indeks tersebut dipilih karena bias yang dihasilkan dari perhitungan tidak besar
dibandingkan indeks-indeks lainnya (Krebs 1989: 386--390). Perhitungan indeks
tumpang-tindih Horn dilakukan dengan perangkat lunak Microsoft Office Excel
2007.
Principal Component Analysis (PCA) adalah metode statistik yang
menggunakan transformasi ortogonal untuk mengonversikan kumpulan variabel
yang saling berkorelasi menjadi variabel baru yang disebut komponen utama (PC,
Principal Component). Variabel baru yang dihasilkan bersifat independen satu
sama lain. Kelebihan PCA adalah walaupun jumlah variabel berkurang, PC yang
dihasilkan tetap memiliki informasi mengenai hubungan antarvariabel. Reduksi
jumlah variabel menjadi PC memungkinkan analisis aspek ekologis difokuskan
pada beberapa variabel yang terpenting saja (Janžekovič & Novak 2012: 127).
Pada penelitian ini, PCA dilakukan untuk dua tujuan, yaitu (1) mengetahui
tumpang-tindih penggunaan habitat antara siamang dan mamalia arboreal lainnya
serta (2) mengetahui kondisi habitat yang berhubungan dengan interaksi
interspesifik. Analisis dilakukan menggunakan perangkat lunak R i386 3.0.2 dan
package vegan (Oksanen 2013: 8—11).
Untuk tujuan pertama, analisis dilakukan pada 12 variabel penggunaan
habitat yang terangkum dalam Tabel 3.3.2(1). Analisis dengan PCA
berdasarkan koefisien korelasinya dengan PC. Variabel dengan korelasi tinggi
(≥ 0,5) akan digunakan untuk menganalisis tumpang-tindih penggunaan habitat. Setelah itu, data diordinasi pada scatter plot yang terdiri atas dua PC sebagai
sumbu-x dan sumbu-y. Pengelompokan titik data berdasarkan spesies mamalia
arboreal dilakukan dengan membuat elips dengan interval kepercayaan 95%.
Tumpang-tindih penggunaan habitat dapat diketahui secara visual berdasarkan
tumpang-tindih elips. Analisis tambahan, yaitu uji perbedaan rerata, dilakukan
pada PC dan variabel-variabel yang berkorelasi tinggi (≥ 0,5) dengan PC jika
terdapat tumpang-tindih elips. Tujuan uji tersebut adalah untuk mengetahui
apakah terdapat perbedaan signifikan penggunaan habitat berdasarkan variabel
tertentu. Jika distribusi data normal, uji yang digunakan adalah uji Anova dan uji
t. Jika distribusi data tidak normal, uji Kruskal-Wallis dan uji Wilcoxon
digunakan.
Tabel 3.3.2(1) Variabel penggunaan habitat yang dianalisis dengan PCA untuk mengetahui tumpang-tindih relung di antara mamalia arboreal.
Kode Variabel Keterangan Tipe Variabel
Ds Diameter substrat Kontinu
Hdt Ketinggian dari tanah Kontinu
Hp Ketinggian pohon Kontinu
Ratio_H Rasio Hdt:Hp Kontinu
Rsp Jarak dari pusat tajuk Kontinu
Rt Jari-jari tajuk Kontinu
Ratio_R Rasio Rsp:Rt Kontinu
Substrat Tipe substrat Kategori
Fenofase Fenofase pohon Kategori
Skor Skor fenofase Kategori
Strata Strata hutan Kategori
Untuk tujuan kedua, analisis dilakukan pada 19 variabel kondisi habitat
yang terangkum dalam Tabel 3.3.2(2). Tahap-tahap analisis hampir sama dengan
tahap analisis penggunaan habitat. Namun, untuk tujuan kedua, pengelompokan
titik data dilakukan berdasarkan tiga kategori interaksi interspesifik, yaitu:
1. netral (toleransi), jika siamang dan spesies lain dapat berada pada satu
pohon tanpa terjadi agresi,
2. dominansi, jika spesies lain menjauhi siamang tanpa terjadi perilaku
agresif dari siamang, dan
3. agresi, jika terjadi perilaku agresif antarspesies, termasuk mengusir dan
berkelahi.
Hasil PCA kemudian digambarkan dalam bentuk scatter plot yang di-overlay
dengan elips untuk menggambarkan pengelompokan interaksi berdasarkan
kondisi terjadinya. Jika terdapat perbedaan kondisi habitat yang menyebabkan
ketiga kategori interaksi, elips yang terbentuk tidak tumpang-tindih. Demikian
pula sebaliknya.
Tabel 3.3.2(2) Variabel kondisi habitat yang dianalisis dengan PCA untuk mengetahui perbedaan kondisi habitat yang berhubungan dengan interaksi interspesifik.
Kode Variabel Keterangan Tipe Variabel
D Jarak antara siamang dengan kompetitor Kontinu
Ds1 Diameter substrat siamang Kontinu
Ds2 Diamater substrat kompetitor Kontinu
Hdt1 Ketinggian siamang dari tanah Kontinu
Hdt2 Ketinggian kompetitor dari tanah Kontinu
Hp Ketinggian pohon Kontinu
Selisih_H Selisih ketinggian siamang dan kompetitor Kontinu
Rsp1 Jarak siamang dari pusat tajuk Kontinu
Rsp2 Jarak kompetitor dari pusat tajuk Kontinu
Rt Jari-jari tajuk Kontinu
Selisih_R Selisih jarak siamang dan kompetitor dari pusat tajuk
Kontinu
Subs1 Tipe substrat siamang Kategori
Tabel 3.3.2(2) (lanjutan)
Fenofase Fenofase pohon Kategori
Skor Skor fenofase Kategori
Strata1 Strata siamang Kategori
Strata2 Strata kompetitor Kategori
Tajuk1 Kategori tajuk siamang Kategori
31 Universitas Indonesia BAB 4
HASIL DAN PEMBAHASAN
Pengamatan perilaku siamang telah dilakukan pada bulan Februari hingga
April 2014 dengan total 24 hari pengamatan dan meliputi 2.504 titik data yang
setara dengan 208,67 jam observasi. Selama periode pengamatan, terjadi dua
perubahan komposisi kelompok siamang. Pertama, individu pradewasa betina
dari kelompok Bimo memisahkan diri dari kelompoknya dan tidak diketahui
keberadaannya sampai akhir studi. Kedua, individu jantan dewasa pada kelompok
Freddie tercatat melakukan kopulasi dengan betina dewasa dan juga betina
pradewasa. Oleh karena itu, betina pradewasa tersebut selanjutnya dianggap
sebagai betina dewasa.
Survei mamalia arboreal telah dilakukan pada 23 hari pengamatan selama
periode penelitian. Hasil survei menunjukkan bahwa siamang hidup
berdampingan dengan sepuluh spesies mamalia arboreal (Lampiran 1). Namun,
hanya spesies yang melimpah yang dianalisis interaksinya dengan siamang.
Spesies yang hanya berjumlah sedikit tidak memberikan dampak kompetitif yang
besar. Mamalia arboreal yang akan dianalisis ditentukan berdasarkan frekuensi
pertemuan. Hewan dengan kelimpahan yang tinggi memiliki probabilitas
pertemuan yang besar sehingga frekuensi pertemuan merupakan indeks yang
dianggap representatif dan sederhana. Berdasarkan hasil survei (Lampiran 1),
mamalia arboreal dengan frekuensi pertemuan tertinggi (Gambar 4) adalah
Presbytis melalophos (51,16%), Callosciurus notatus (20,16%), dan Ratufa
bicolor (10,08%).
Dari kedua metode pengambilan data tersebut--pengamatan perilaku
siamang dan survei mamalia arboreal--telah diperoleh data yang dibutuhkan untuk
mencapai tujuan penelitian. Pemaparan hasil dan pembahasan dibagi menjadi
empat bagian, yaitu tumpang-tindih relung berdasarkan pakan, tumpang-tindih
relung berdasarkan penggunaan habitat, kompetisi interspesifik, dan partisi relung
Gambar 4. Tiga spesies mamalia arboreal dengan frekuensi pertemuan tertinggi di area survei.
Keterangan:
(a) = simpai (Presbytis melalophos) (b) = bajing kelapa (Callosciurus notatus) (c) = jelarang hitam (Ratufa bicolor)
[Sumber: Dokumentasi pribadi]
(a) (b)
(c) 20 cm
4.1. Tumpang-tindih Relung Berdasarkan Pakan
Komposisi pakan siamang terdiri atas 72,52% buah, 23,57% daun, 3,77%
bunga, dan 0,14% materi hewani (Gambar 4.1(1)). Hasil tersebut menunjukkan
bahwa siamang memiliki preferensi yang tinggi terhadap buah-buahan. Selama
dua bulan pengamatan, siamang tercatat memakan 23 spesies buah, 22 spesies
daun, dan lima spesies bunga. Frekuensi dan keanekaragaman spesies pakan
siamang dapat dilihat pada Lampiran 2. Berdasarkan penelitian Nurcahyo (1999:
24) yang dilakukan di lokasi yang sama, komposisi pakan siamang terdiri atas
52,07% buah, 42,62% daun, dan 5,3% bunga. Proporsi buah yang lebih besar
terjadi karena saat penelitian ini dilakukan, terdapat beberapa pohon pakan
siamang yang berbuah lebat seperti Beilschmiedia dyctioneura, Sandoricum
koetjape, Ficus altissima, dan spesies-spesies dari famili Annonaceae. Siamang
memakan lebih banyak daun jika ketersediaan buah sedikit (Elder 2009: 153).
Gambar 4.1(1) Diagram komposisi pakan tiga kelompok siamang subjek penelitian.
0% 10% 20% 30% 40% 50% 60% 70% 80% 90% 100%
Bimo Freddie Gatot Total
Invertebrata
Bunga
Daun
Gambar 4.1(2) Diagram perbandingan komposisi pakan siamang, simpai, bajing kelapa, dan jelarang hitam.
Keterangan:
Angka menunjukkan jumlah data suatu spesies mamalia tercatat memakan makanan dari kategori tertentu.
Komposisi pakan tiga spesies mamalia arboreal yang hidup di ruang
jelajah siamang dapat dilihat pada Gambar 4.1(2). Simpai diketahui memakan
73,91% buah dan 26,09% daun. Hasil tersebut berbeda dengan hasil yang
dilaporkan oleh Curtin (1980: 138) yang menyatakan bahwa simpai di Malaysia
memakan 48% buah dan 35% daun. Selama periode pengambilan data, bajing
kelapa hanya tercatat enam kali memakan buah dan satu kali memakan kayu
pohon. Berdasarkan hasil penelitian Payne (1980: 273) di hutan hujan tropis
Malaysia, komposisi pakan terbesar bajing kelapa adalah biji-bijian (> 40%),
dilanjutkan dengan kayu dan getah (30%), daun (25%), serta bunga dan materi
hewani. Jelarang hitam tercatat 12 kali memakan buah (92,31%) dan hanya sekali
memakan bunga (7,68%). Sifat frugivora jelarang hitam juga dilaporkan oleh
Payne (1980: 273--274) yang menyatakan bahwa hewan pengerat tersebut
memakan 81% buah. Perbedaan antara hasil pengamatan dengan literatur
kemungkinan besar disebabkan oleh jumlah sampel yang sedikit (< 30 per
Siamang Simpai Bajing Kelapa Jelarang Hitam
Kayu 0 0 1 0
Invertebrata 1 0 0 0
Bunga 27 0 0 1
Daun 168 6 0 0
Buah 520 17 6 12
spesies). Walaupun hasil pengamatan tidak meliputi seluruh variasi pakan, hasil
tersebut dianggap representatif untuk komponen pakan dominan ketiga spesies
mamalia arboreal.
Perhitungan indeks Horn dilakukan untuk mengetahui tumpang-tindih
relung antara siamang dan ketiga mamalia arboreal yang paling melimpah, yaitu
simpai, bajing kelapa, dan jelarang hitam. Kategori pakan materi hewani dan
kayu tidak diikutsertakan dalam perhitungan. Hasil perhitungan (Tabel 4.1(1) dan
Lampiran 3) menunjukkan bahwa tumpang-tindih relung terbesar terdapat di
antara siamang dan jelarang hitam (Ro = 0,418), sedangkan tumpang-tindih
terkecil terdapat di antara siamang dan simpai (Ro = 0,310). Spesies-spesies
tumbuhan pakan siamang yang juga merupakan sumber pakan bagi simpai, bajing
kelapa, dan jelarang hitam dirangkum pada Tabel 4.1(2). Berdasarkan tabel
tersebut, jumlah spesies pakan yang sama antara siamang dan simpai lebih banyak
daripada siamang dan jelarang hitam, tetapi indeks Horn menunjukkan
sebaliknya. Hal itu terjadi karena simpai memakan daun beberapa spesies
tumbuhan yang tidak dimakan oleh siamang.
Tabel 4.1(1) Tabel ringkasan catatan spesies pakan dan tumpang-tindih relung berdasarkan pemilihan pakan.
Spesies Frekuensi catatan
Jumlah spesies pakan
Jumlah spesies pakan yang sama dengan
siamang
Indeks Horn
Siamang 715 50 - -
Simpai 23 13 6 0,310
Bajing kelapa 6 4 3 0,334