BAB VI
JENIS DAN KARAKTERISTIK JAMUR PENYEBAB
PELAPUKAN PADA BANGUNAN RUMAH
Identifikasi Jamur Pelapuk Kayu
Jamur pelapuk yang banyak menyerang bangunan rumah diisolasi dan diberi kode DE, SC dan PB. Jamur DE diisolasi dari komponen kayu plafon bangunan yang terlindung dari sinar matahari, tapi sesekali terkena rembesan air dari atap yang bocor bila sedang turun hujan. Jamur SC diambil dari komponen kayu yang terkena hujan dan sinar matahari langsung. Adapun jamur PB diisolasi dari komponen tiang pintu kamar mandi yang tidak terkena hujan dan sinar matahari langsung, tapi sering terkena percikan air pemakaian di kamar mandi. Ketiga jamur pelapuk kayu berhasil ditumbuhkan pada media PDA dan media baglog yang mengandung serbuk kayu.
Tubuh buah jamur DE berwarna abu-abu dengan pinggiran berwarna putih. Pada permukaan basiodikarp yang menghadap ke bawah terdiri dari banyak pori yang merupakan ujung dari tabung-tabung himenium tempat keluar spora. Tubuh buah jamur PB selain warna abu-abu dan putih ada juga bagian berwarna hitamnya. Pada permukaan putih dan abu-abu terdiri dari banyak pori. Jamur SC berbentuk seperti kipas. Permukaan atas berbulu, pinggirannya melengkung ke bawah. Di bagian bawah tudung terdiri dari banyak lamela yang menggulung memanjang radial seperti insang.
Pada media PDA, miselia ketiga jamur uji pada mulanya berwarna putih (Gambar 12). Setelah sekitar 3 minggu pada kultur SC, selain hamparan miselium putih terdapat bagian yang kuning dan spot coklat. Pada kultur DE, diantara miselium yang putih terdapat bagian penebalan miselium berwarna kuning tidak merata. Adapun kultur PB warnanya merata putih dan tebal.
Berdasarkan ciri-ciri tipe miselium yang diuraikan Stamets (2000), miselium jamur DE dan PB termasuk tipe linier, yaitu terdiri dari garis-garis memanjang radial yang homogen, kecuali pada miselium PB ada sebagian
garis-garis yang menebal. Adapun miselium SC termasuk pseudo-rizomorfik, yaitu seperti jalinan benang-benang terpilin halus.
Pada miselium yang sudah menua terjadi perubahan warna. Miselium DE menjadi tipe appressed berwarna coklat dan terdapat agregat-agregat cairan berwarna coklat kemerahan yang merupakan metabolit yang dihasilkan jamur tersebut. Miselium jamur PB setelah 12 minggu menjadi bertipe appressed tapi putih dengan sedikit bercak-bercak coklat dan hitam; sedangkan miselium SC menjadi tipe cottony berwarna putih dengan banyak spot-spot berwarna coklat kemerahan. Hal ini sesuai dengan yang disampaikan Stamets (2000) bahwa sebagian besar jamur saprofit pada mulanya memiliki miselium warna putih. Warna miselium dapat berubah ketika miselium tersebut dewasa. Seperti
Gambar 12 Isolat jamur DE, SC dan PB pada media PDA (potato dextrose agar).
Ganoderma lucidum¸miselium muda berwarna putih, setelah dewasa menjadi kuning.
Hasil pengukuran kecepatan tumbuh miselium jamur DE, PB dan SC pada media PDA dengan pH 5,6 dan suhu 30 oC menunjukkan bahwa pertumbuhan miselium jamur PB adalah yang paling cepat, sedangkan miselium jamur SC adalah yang paling lambat. Nilai rata-rata kecepatan tumbuh diameter miselium jamur DE, PB dan SC berurutan adalah 1,27 cm/hari, 1,45 cm/hari dan 1,05 cm/hari (Lampiran 15).
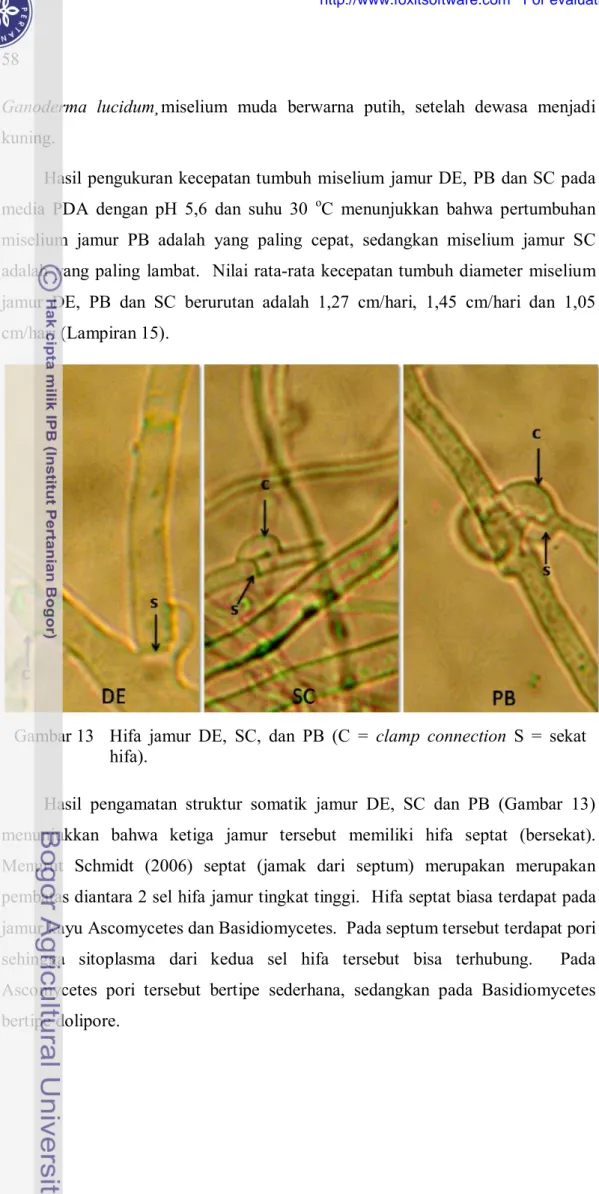
Hasil pengamatan struktur somatik jamur DE, SC dan PB (Gambar 13) menunjukkan bahwa ketiga jamur tersebut memiliki hifa septat (bersekat). Menurut Schmidt (2006) septat (jamak dari septum) merupakan merupakan pembatas diantara 2 sel hifa jamur tingkat tinggi. Hifa septat biasa terdapat pada jamur kayu Ascomycetes dan Basidiomycetes. Pada septum tersebut terdapat pori sehingga sitoplasma dari kedua sel hifa tersebut bisa terhubung. Pada Ascomycetes pori tersebut bertipe sederhana, sedangkan pada Basidiomycetes bertipe dolipore.
Gambar 13 Hifa jamur DE, SC, dan PB (C = clamp connection S = sekat hifa).
Clamp connection (sambungan apit) juga ditemukan pada miselia jamur DE, SC dan PB. Menurut Watanabe (2002) bahwa keberadaan hifa septat dan clamp connection merupakan ciri penting Basidiomycetes. Hifa jamur SC relatif lebih besar dibandingkan dengan hifa jamur DE dan PB. Diameter hifa jamur SC adalah 1,3-2,0 µm. Adapun hifa jamur DE dan PB rata-rata berdiameter antara 0,9-1,7 µm dan 1,0-1,6 µm. Menurut Schmidt (2006) diameter hifa bervariasi antar jenis jamur. Pada Phellinus pini ada hifa mikro berdiameter 0,1-0,4 µm. Sedangkan pada Serpula laacrimans hifa vegetaifnya berdiameter 2-7 µm, tapi juga memiliki hifa pembuluh berdiameter 60 µm.
Selama proses inkubasi, pertumbuhan ketiga jamur DE, SC dan PB tampak dari luar baglog dengan semakin luasnya miselium putih meliputi media serbuk kayu (Gambar 14). Miselium ketiga jamur tersebut sudah meliputi permukaan media baglog (± 200 gram) dalam 2 minggu inkubasi. Setelah 4 minggu inkubasi, miselium jamur SC mengalami penebalan dan membentuk noda-noda berwarna Gambar 14 Pertumbuhan miselia (m) jamur PB, SC dan DE pada media
coklat di antara putihnya warna miselium. Setelah 12 minggu inkubasi noda-noda tersebut menjadi hitam.
Miselium jamur DE dan PB tumbuh putih merata, tapi setelah 8 minggu inkubasi warna miselium jamur DE menjadi kekuningan, sedangkan miselium jamur PB tetap putih hingga 12 minggu inkubasi. Pertumbuhan miselium kedua jamur tampak lebih tebal pada media serbuk kayu sengon. Ini menunjukkan media serbuk sengon lebih disukai kedua jamur tersebut daripada media serbuk pinus yang dikenal memiliki kandungan resin dan ekstraktif pinosylvin yang bersifat racun. Selain itu miselium jamur PB relatif lebih tebal dibandingkan miselim jamur DE
Ketiga spesimen jamur uji dapat ditumbuhkan pada media baglog serbuk kayu pinus dan sengon 200 g. Dalam waktu satu bulan tubuh buahnya sudah tumbuh, kecuali tubuh buah jamur SC pada media serbuk sengon belum tumbuh hingga minggu ke-12. Tubuh buah jamur PB tumbuh pada seluruh media baglog sengon dan pinus. Tubuh buah jamur DE tumbuh pada 90% media serbuk sengon dan 63% media serbuk pinus. Adapun tubuh buah jamur SC tumbuh pada semua media serbuk pinus.
Pembentukan tubuh buah selain dipengaruhi oleh faktor lingkungan, juga oleh faktor nutrisi pada substrat media. Sebagaimana dinyatakan oleh Chang dan Miles (2004) bahwa pertumbuhan tubuh buah Schizophyllum commune dapat terpacu dengan penghilangan atau kehabisan nutrisi. Pertumbuhan tubuh buah juga dapat diinduksi dengan memotong miselium vegetatif, sehingga tubuh buah tumbuh pada potongan tersebut. Selain itu terkadang tubuh buah jamur juga terbentuk pada tepi petri dish.
Bentuk tubuh buah jamur SC yang tumbuh pada baglog serupa dengan yang ditemukan di lapangan yaitu seperti kulit berbentuk menyerupai kipas (Gambar 15), tapi ukurannya 1,5 kali lebih besar dari yang ditemukan di lapangan. Lebar tubuh buah SC pada baglog adalah 1-3 cm. Bagian atasnya berwarna abu-abu dan berbulu. Bagian bawahnya memiliki lamela seperti insang dengan belahan longitudinal yang menggulung ke dalam. Pada tubuh buah tidak terlihat adanya
stipe (tangkai) khusus, kecuali pengecilan tubuh buah langsung membentuk seperti tangkai; dagingnya relatif tipis lunak (yang muda) hingga keras (yang kering).
Jejak spora jamur SC yang dihasilkan berwarna putih kekuningan. Spora merupakan struktur reproduktif yang penting bagi jamur pelapuk kayu karena peranannya dalam reproduksi. Spora dari tubuh buah jamur dapat tersebar ke berbagai tempat dengan bantuan angin, serangga, hujan, dsb. Ketika menempel pada kayu, spora mengeluarkan enzim yang mendegradasi kayu dan mengeluarkan hifa yang mengambil nutrisi dari dinding sel dan isi dari lumen (Harris 2001).
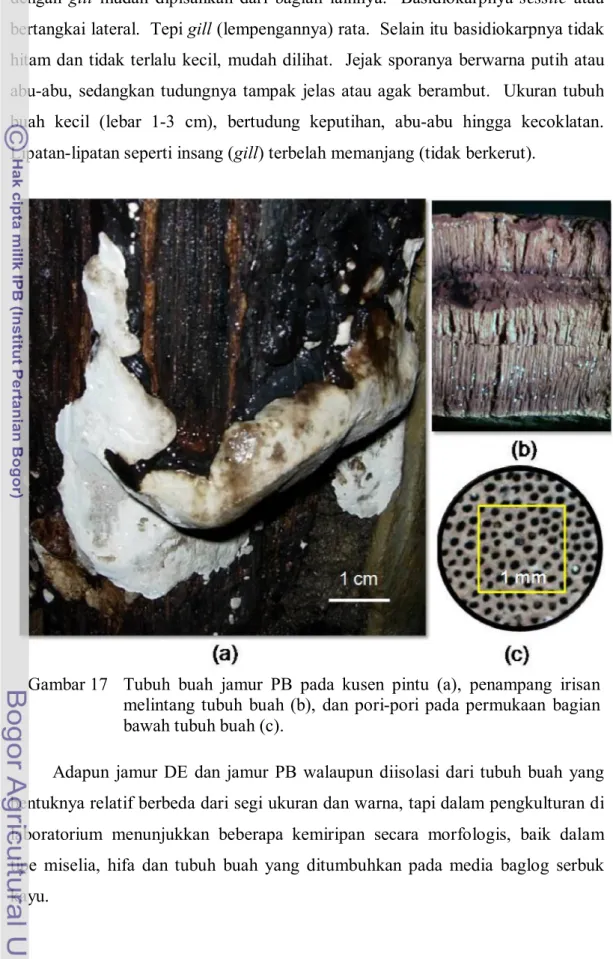
Basidiokarp (tubuh buah) jamur DE (Gambar 16) dan jamur PB (Gambar 17) yang diisolasi tidak bertangkai dan keras. Permukaan tubuh buah jamur DE berwarna abu-abu, coklat, hitam dan putih (di lapangan), sedangkan jamur PB berwarna hitam dan putih. Tubuh buah jamur PB yang tumbuh pada media baglog warnanya coklat tua hingga hitam, lebih gelap dibanding tubuh buah DE yang coklat muda. Bagian konteks tubuh buah kedua jamur tersebut berwarna coklat. Menurut Flood et al. (2000), konteks Ganoderma berbeda-beda warnanya, Gambar 15 Tubuh buah jamur SC pada kayu sengon (a), lamela pada
mulai dari putih hingga coklat tua. Warna konteks ini dapat berubah dalam lingkungan yang berbeda.
Himenofor jamur PB dan jamur DE berbentuk tabung dan tersusun dalam beberapa lapisan. Konteks berwarna coklat dengan tekstur seperti gabus. Ujung-ujung himenofor membentuk lubang-lubang pori dengan jarak cukup rapat pada permukaan bawah tubuh buah kedua jamur. Pori pada tubuh buah jamur DE berdiameter rata-rata 73,98 µm, sedangkan pada jamur PB diameter porinya adalah 91,93 µm. Berdasarkan kerapatan porinya jamur DE lebih tinggi, yaitu 55 pori/mm2, adapun jamur PB adalah 49 pori/mm2. Pori-pori tersebut tempat keluarnya spora-spora yang siap menjadi individu jamur baru. Bentuk spora yang dihasilkan jamur DE dan jamur PB relatif sama, bulat lonjong; permukaannya tidak rata; warnanya coklat muda hingga coklat tua. Pada jamur SC spora yang dihasilkan juga berbentuk bulat lonjong, tapi warnanya putih.
Berdasarkan karakteristik anatomi dan morfologis jamur SC tersebut banyak kesamaan dengan jamur Schizophyllum commune. Sebagaimana ciri-ciri morfologis yang diuraikan oleh Emberger (2006) bahwa jamur S. commune termasuk dalam kelompok yang memiliki gill (bentuk seperti insang). Bagian Gambar 16 Tubuh buah jamur DE (a) pada rangka plafon dan pori-pori pada
bawah basidiokarp terdiri dari lempengan radial atau gill. Fungi ini berdaging dengan gill mudah dipisahkan dari bagian lainnya. Basidiokarpnya sessile atau bertangkai lateral. Tepi gill (lempengannya) rata. Selain itu basidiokarpnya tidak hitam dan tidak terlalu kecil, mudah dilihat. Jejak sporanya berwarna putih atau abu-abu, sedangkan tudungnya tampak jelas atau agak berambut. Ukuran tubuh buah kecil (lebar 1-3 cm), bertudung keputihan, abu-abu hingga kecoklatan. Lipatan-lipatan seperti insang (gill) terbelah memanjang (tidak berkerut).
Adapun jamur DE dan jamur PB walaupun diisolasi dari tubuh buah yang bentuknya relatif berbeda dari segi ukuran dan warna, tapi dalam pengkulturan di laboratorium menunjukkan beberapa kemiripan secara morfologis, baik dalam tipe miselia, hifa dan tubuh buah yang ditumbuhkan pada media baglog serbuk kayu.
Gambar 17 Tubuh buah jamur PB pada kusen pintu (a), penampang irisan melintang tubuh buah (b), dan pori-pori pada permukaan bagian bawah tubuh buah (c).
Berdasarkan kunci identifikasi ciri morfologis dari Emberger (2006) untuk jamur DE dan jamur PB sama-sama termasuk pada kategori bentuk jamur poroid (berpori). Pori-pori tersebut merupakan ujung dari tabung-tabung yang dinding dalamnya menjadi tempat pembentukan spora. Ukuran pori-pori tersebut cukup kecil, yaitu 5-7 pori/mm garis, bentuknya bundar atau membentuk sudut. Basidiokarpnya keras seperti kayu dan ketika dipotong melintang menunjukkan lebih dari satu lapisan (lebih dari satu tahun pertumbuhan). Selain itu, bagian dalamnya (konteks) berwarna coklat kekuningan atau coklat gelap. Basidiokarpnya datar dengan permukaan atas tertutup kerak, sedangkan permukaan bawahnya berpori dan berwarna putih yang bila tergores menjadi coklat tua. Basidiokarpnya sering ditemukan pada berbagai jenis kayu daun lebar yang hidup ataupun telah mati. Ciri-ciri tersebut menunjukan jenis jamur G. applanatum.
Dalam proses identifikasi molekuler jamur pelapuk, amplifikasi PCR (Polymerase Chain Reaction) berhasil dilakukan sehingga pita-pita DNA (Deoxyribonucleic Acid) dari ketiga jamur yang diuji (jamur DE, jamur SC, dan jamur PB) dapat dilihat melalui agarose gel electrophoersis (Gambar 18). Pita-pita DNA dari ketiga spesimen jamur tersebut terpisah berdasarkan ukurannya yang berada diantara 800-900 bp. PCR sequence dan electropherogram ketiga jamur yang diuji dapat dilihat pada Lampiran 8 sampai 14. Hasil perbandingan ITS sequence ketiga jamur dengan DDBJ database (Zhang et al. 2000), terbukti bahwa jamur SC menunjukkan 99% identitas Schizophyllum commune (nomor akses Genbank: EF155505.1). Adapun jamur DE dan jamur PB sama-sama menunjukkan 93% identitas Ganoderma lipsiense (nomor akses Genbank:EF060006.1). Menurut The BayScience Foundation (2009), G. lipsiense adalah nama lain dari G. applanatum, termasuk family Ganodermataceae, ordo Polyporales, class Basidiomycetes, phylum Basidiomycota. Menurut Niemela dan Miettinen (2008), G. applanatum merupakan pelapuk serius pada pohon-pohon di taman dan hutan serta sering ditemukan pada tunggak pohon-pohon bekas tebangan.
S. commune termasuk family Schizophyllaceae, ordo Agaricales, class Homobasidiomycetes, phylum basidiomycota (UniProt Consortium 2010). S. commune merupakan penyebab penyakit pada pohon meranti merah (Shorea smithiana) (Erwin et al. 2008). Menurut Kuo (2003) juga S. commune tumbuh pada kayu dan kadang kadang bersifat parasit pada pohon. S. commune tumbuh secara soliter tapi lebih sering berkelompok atau kluster pada log, ataupun kayu gergajian. Dalam keadaan kering bisa bertahan hingga datang kelembaban untuk tumbuh lagi. Menurut Adejoye et al. (2007), S. commune merupakan salah satu jamur yang dapat di makan, sumber protein, vitamin, lemak dan mineral di Nigeria. Namun pertumbuhannya secara alami hanya pada musim hujan, sehingga perlu dibudidayakan.
Uji Oksidasi Jamur Pelapuk Kayu
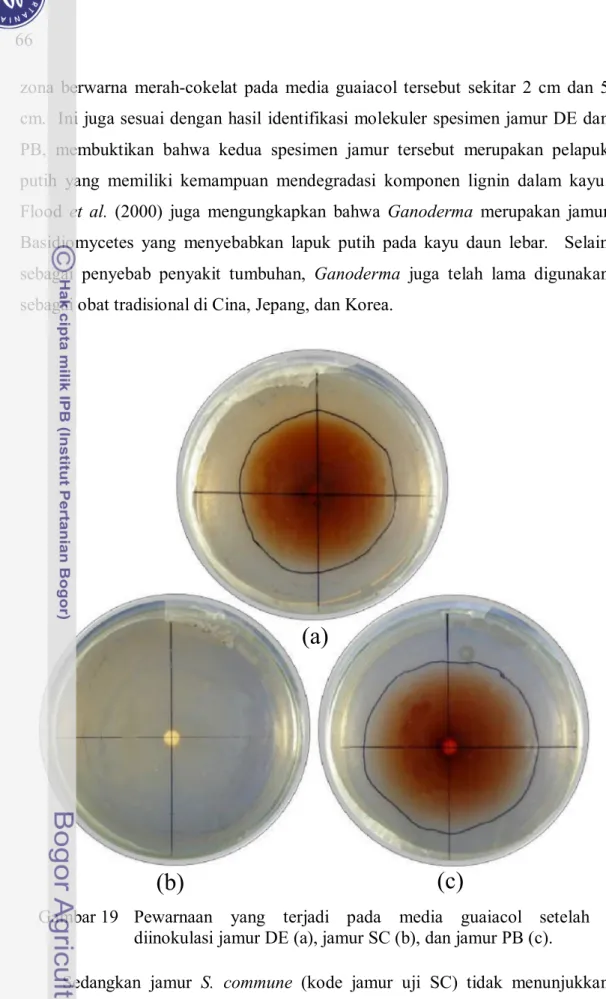
Hasil uji oksidasi jamur pelapuk menggunakan media guaiacol memperlihatkan reaksi enzimatik yang cukup cepat. Media guaiacol yang diinokulasi jamur G. applanatum, kode jamur PB dan DE sama-sama mengalami pewarnaan menjadi coklat (Gambar 19). Setelah 2 dan 8 hari inkubasi, diameter Gambar 18 Hasil gel electrophoresis produk PCR dari rDNA jamur PB, jamur
zona berwarna merah-cokelat pada media guaiacol tersebut sekitar 2 cm dan 5 cm. Ini juga sesuai dengan hasil identifikasi molekuler spesimen jamur DE dan PB, membuktikan bahwa kedua spesimen jamur tersebut merupakan pelapuk putih yang memiliki kemampuan mendegradasi komponen lignin dalam kayu. Flood et al. (2000) juga mengungkapkan bahwa Ganoderma merupakan jamur Basidiomycetes yang menyebabkan lapuk putih pada kayu daun lebar. Selain sebagai penyebab penyakit tumbuhan, Ganoderma juga telah lama digunakan sebagai obat tradisional di Cina, Jepang, dan Korea.
Sedangkan jamur S. commune (kode jamur uji SC) tidak menunjukkan pewarnaan atau bereaksi negatif hingga hari ke 8 inkubasi. Sementara itu miseliumnya tampak tumbuh tipis dengan diameter 5 cm. Diduga dalam
(a)
(b)
(c)
Gambar 19 Pewarnaan yang terjadi pada media guaiacol setelah diinokulasi jamur DE (a), jamur SC (b), dan jamur PB (c).
pengujian guaicol ini S. commune hanya sedikit mengeluarkan laccase. Walaupun tidak menunjukkan perubahan warna, jamur S. commune merupakan jamur pelapuk putih, sebagaimana dinyatakan oleh Ghosh et al. (2005), Hirai et al (2008), dan Tsujiyama & Minami (2005).
Pertumbuhan Jamur Pelapuk pada Berbagai Suhu Inkubasi
Selain makanan dan oksigen, lingkungan yang hangat dan lembab dibutuhkan jamur untuk pertumbuhannya (Harris 2001). Hasil eksperimen menunjukkan bahwa jamur Schizophyllum commune dan Ganoderma applanatum memiliki suhu optimum yang berbeda dalam pertumbuhannya. Suhu optimum pertumbuhan kedua jamur uji dalam eksperimen ini lebih tinggi dari 25 oC (Gambar 20).
Pertumbuhan miselia jamur S. commune dan G. applanatum pada media PDA memberikan respon cukup baik. Suhu optimum pertumbuhan jamur G. applanatum pada media PDA adalah antara 35 oC dan 40 oC, atau tepatnya adalah pada suhu 37 oC. Pada suhu tersebut pertumbuhan diameter miseliumnya sebesar 1,99 cm/hari, termasuk pertumbuhan yang cepat, sedangkan pada suhu 25 oC Gambar 20 Pertumbuhan diameter miselia jamur pelapuk S.commune dan G.
pertumbuhannya agak cepat yaitu 1,24 cm/hari. Data pertumbuhan miselium pada berbagai suhu disajikan pada Lampiran 15. Sebagai perbandingan adalah pertumbuhan jamur G. boninense yang lebih lambat yaitu 0,9 cm/hari pada suhu optimumnya 30 oC (Abadi 1987), sedangkan pertumbuhan G. australe sebesar 0,6 cm/hari (Yeh & Chen 1990).
Jamur S. commune tumbuh optimum antara suhu 25 oC dan 30 oC, atau tepatnya pada suhu 29 oC dengan kecepatan tumbuh diameter miselium 1,07 cm/hari, sedangkan pada suhu 25 oC adalah 0,96 cm/hari. Hal itu berarti pertumbuhan jamur S. commune pada suhu ruang dan suhu optimumnya termasuk klasifikasi agak cepat. Sementara itu berdasarkan percobaan Adejoye et al. (2007) suhu optimum pertumbuhan S. commune pada media PDA yang diperkaya dengan pepton adalah 25 oC.
Pertumbuhan jamur pelapuk kayu pada umumnya lebih cepat dalam keadaan hangat daripada dalam keadaan dingin. Suhu optimum untuk pertumbuhan bervariasi antar jenis jamur, tapi pada umumnya jamur tumbuh baik pada suhu 12-40 oC. Adapun suhu optimum untuk jamur pelapuk yang digunakan dalam pengujian laboratorium berkisar antara 28–36 oC (Nicholas & Crawford 2003). Penelitian yang dilakukan oleh Harris (2001) menunjukkan bahwa suhu optimum pertumbuhan jamur adalah 20–25 oC. Di atas 30 oC aktivitas pertumbuhan jamur menurun bahkan di bawah 0 oC dan di atas 40 oC pada umumnya jamur pelapuk mati.
Pertumbuhan jamur S. commune pada berbagai suhu lebih lambat dibandingkan dengan jamur G. applanatum. Berdasarkan kelas pertumbuhannya pada suhu ruang (25 oC) jamur G.applanatum tergolong cepat, sedangkan jamur S. commune tergolong agak cepat. Pada suhu 45 oC pertumbuhan jamur S. commune terhenti, sedangkan pertumbuhan jamur G. applanatum baru terhenti pada suhu 50
o
C. Dengan demikian untuk tujuan pengendalian serangan kedua jamur pelapuk ini pada kayu diperlukan suhu sekitar 60 oC.
Pertumbuhan Jamur Pelapuk pada Berbagai pH Media
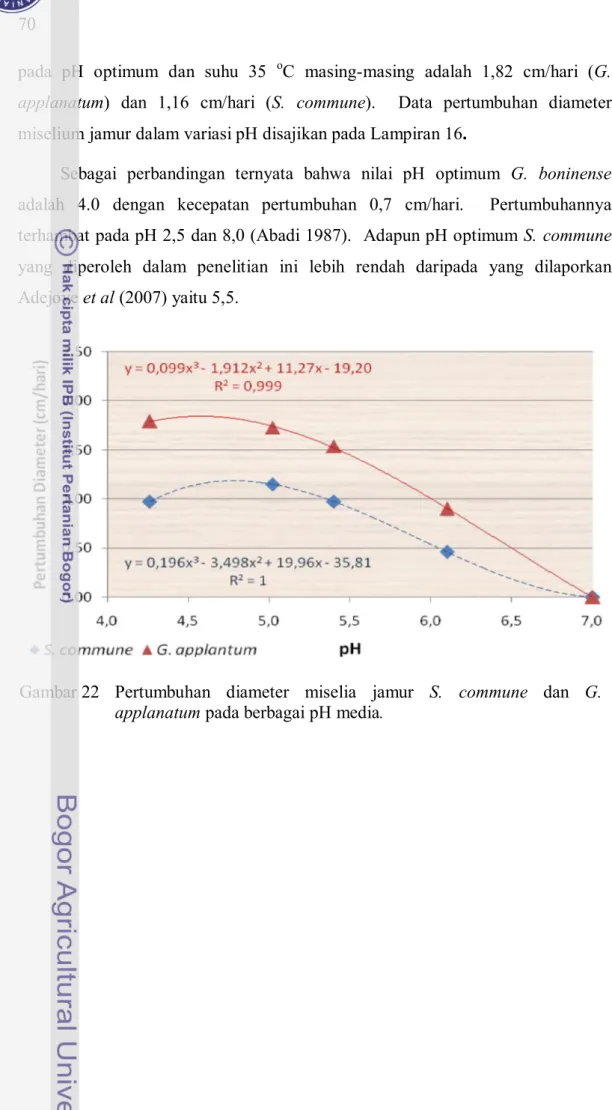
Hasil pengujian menunjukkan bahwa kecepatan tumbuh miselia jamur S. commune dan G. applanatum bervariasi dalam pengaruh pH yang berbeda. Pada media ber-pH 7, jamur G. applanatum tidak menunjukkan pertumbuhan (perluasan miselium), sedangkan jamur S. commune memperlihatkan respon pertumbuhan setelah lebih dari 10 hari inkubasi (Gambar 21). Hal ini menunjukkan kemampuan adaptasi jamur S. commune pada pH yang lebih tinggi dibanding jamur G. applanatum.
Pada Gambar 22 diperlihatkan bahwa jamur S. commune dan G. applanatum membutuhkan kondisi asam untuk tumbuh dengan baik. Bowyer et al. (2003) juga mengungkapkan bahwa jamur lebih suka kondisi asam yaitu pada pH 4-6. Pada pH 5,4 atau lebih tinggi, kecepatan pertumbuhan jamur S. commune dan G. applanatum cenderung menurun. Selain itu, jamur S. commune juga cenderung menurun kecepatannya pada pH kurang dari 4,3.
Nilai pH optimum untuk kedua jamur uji hampir sama, yaitu antara pH 4,3 dan 5,0. Hasil perhitungan regresi, pH optimum G. applanatum adalah 4,6, sedangkan untuk S. commune adalah 4,8. Kecepatan pertumbuhan diameternya Gambar 21 Pertumbuhan miselia jamur pelapuk S. commune pada media
PDA (pH 7) hingga 10 hari inkubasi (a) dan setelah 34 hari inkubasi (b).
pada pH optimum dan suhu 35 oC masing-masing adalah 1,82 cm/hari (G. applanatum) dan 1,16 cm/hari (S. commune). Data pertumbuhan diameter miselium jamur dalam variasi pH disajikan pada Lampiran 16.
Sebagai perbandingan ternyata bahwa nilai pH optimum G. boninense adalah 4.0 dengan kecepatan pertumbuhan 0,7 cm/hari. Pertumbuhannya terhambat pada pH 2,5 dan 8,0 (Abadi 1987). Adapun pH optimum S. commune yang diperoleh dalam penelitian ini lebih rendah daripada yang dilaporkan Adejoye et al (2007) yaitu 5,5.
Gambar 22 Pertumbuhan diameter miselia jamur S. commune dan G. applanatum pada berbagai pH media.
BAB VII
DAMPAK SERANGAN JAMUR PELAPUK TERHADAP
SIFAT-SIFAT KAYU
Pengaruh Serangan Jamur Pelapuk terhadap Struktur Anatomi Kayu
Kayu memiliki struktur kompleks dan dapat menyimpan air yang sangat penting bagi kelangsungan hidup organisme perusak kayu. Selain itu, rongga-rongganya dapat memfasilitasi aktivitas dan pergerakan organisme perusak serta hifa-hifa jamur dalam kayu. Dengan demikian kayu bukan hanya menjadi bahan makanan organisme perusak, tapi juga sebagai media yang mendukung dan melindungi organisme tersebut dalam melangsungkan kehidupannya.
Hasil pengamatan mikroskopis pada preparat slide mikrotom kayu sengon dan pinus yang digunakan dalam uji pelapukan menujukkan keberadaan hifa-hifa jamur dalam sel jari-jari, sel pembuluh kayu sengon dan dalam saluran interseluler kayu pinus (Gambar 23 dan 24). Artinya sel jari-jari, sel pembuluh dan saluran interseluler tersebut menjadi awal akses utama serangan jamur pelapuk ke dalam kayu daun lebar maupun kayu daun jarum. Dengan demikian proporsi, ukuran, Gambar 23 Hifa (h) jamur pelapuk G. applanatum dalam saluran interseluler
(i) dan sel jari-jari (j) kayu pinus serta dalam sel pembuluh (p) kayu sengon (perbesaran 450 x).
distribusi dan orientasi sel-sel dan saluran interseluler tersebut akan menentukan tingkat kolonisasi jamur dalam kayu.
Jamur G. applantum dan S. commune mampu menembus ke bagian dalam jaringan kayu dengan kecepatan invasi yang berbeda. Jamur G. applanatum sendiri bergerak relatif lebih cepat dalam mengkolonisasi jaringan kayu dibandingkan dengan jamur S. commune. Hal ini terkait dengan sifat pertumbuhan jamur yang berlainan antar jenis yang berbeda. Kemampuan hidup berbagai jenis jamur pelapuk dalam keterbatasan air dan nutrisi dalam kayu juga dapat berlainan.
Mekanisme serangan jamur pelapuk pada kayu diawali ketika spora dari sumber infeksi menempel pada kayu dan berkecambah membentuk hifa-hifa jamur yang kemudian bercabang-cabang membentuk miselium yang melekat pada permukaan kayu. Hifa-hifa jamur kemudian masuk melalui sel jari-jari yang merupakan jalan paling mudah bagi hifa jamur untuk mendegradasi kayu. Selain itu, jari-jari merupakan sel-sel parenkim yang mengandung zat-zat makanan bagi Gambar 24 Hifa (h) jamur pelapuk S. commune dalam saluran interseluler
(i) dan sel jari-jari (j) kayu pinus serta dalam sel pembuluh (p) kayu sengon (perbesaran 450 x).
jamur, sebagaimana diungkapkan Bowyer et al. (2003) bahwa gula dan pati terdapat dalam lumen sel dan parenkim jari-jari kayu. Zat-zat tersebut merupakan nutrisi yang mudah dikonsumsi oleh jamur dalam jumlah terbatas. Sel jari-jari juga merupakan jalan yang mudah dilewati oleh hifa-hifa jamur karena secara alami sel jari-jari juga berfungsi sebagai sarana transportasi. Sel jari-jari yang tersusun seperti susunan batu bata pada bangunan, berfungsi dalam sintesis, penyimpanan, transport lateral bahan biokimia dan air (Rowell 2005). Selain itu sel parenkim jari-jari berdinding tipis sehingga relatif mudah terdegradasi dan ditembus oleh hifa jamur. Dalam kayu daun lebar, sel-sel jari-jari yang menjadi sarana pergerakan jamur ini, memiliki ukuran yang lebih bervariasi dan lebih besar daripada dalam kayu daun jarum (Butterfield 2006), bahkan proporsi sel jari-jari dalam kayu daun lebar pada umumnya 30% lebih banyak daripada dalam kayu daun jarum (Ridout 2004). Dengan demikian akses hifa-hifa jamur pada arah radial dalam kayu daun lebar secara fisik relatif lebih besar dibandingkan dengan dalam kayu daun jarum. Selain itu, sel jari jari yang berorientasi pada arah radial menjadikan bidang tangensial kayu lebih terbuka dibandingkan dengan bidang radial sebagai jalan invasi hifa-hifa jamur ke dalam kayu.
Pada arah longitudinal, jalan termudah bagi jamur dalam menginvasi ke dalam kayu daun lebar adalah melalui sel pembuluh, sedangkan pada kayu daun jarum melalui saluran interseluler. Kemudahan itu terjadi karena pergerakan air dalam kayu daun lebar terjadi melalui sel-sel pembuluh yang ujungnya bersambungan satu sama lain. Pada dinding ujung sel-sel pembuluh ini terdapat bidang perforasi sehingga bisa dilewati air (Ridout 2004). Selain itu, sel pembuluh memiliki ukuran cukup besar untuk dimasuki hifa yaitu antara 50-200 µm bahkan bisa lebih dari 300 µm (Rowell 2005), sementara diameter hifa pada umumnya antara 1-30 µm (Walker & White 2005). Sel pembuluh dan saluran interseluler yang berorientasi pada arah longitudinal menjadikan bidang lintang kayu pada umumnya lebih banyak terbuka bagi akses spora dan hifa jamur sehingga bisa menempel dan masuk ke dalam jaringan kayu. Bidang lintang kayu juga lebih tinggi permeabilitasnya dibandingkan dengan bidang radial dan tangensial, sehingga air dan kelembaban dapat cepat masuk ke dalam kayu pada arah longitudinal secara kapiler dan menjadikan bagian dalam kayu kondusif bagi
pertumbuhan dan pergerakan hifa-hifa jamur. Di dalam sel pembuluh, hifa-hifa jamur tidak mendapatkan cukup bahan yang bisa dikonsumsi karena saluran pembuluh memiliki dinding sel sekunder tebal dan mengalami lignifikasi serta tidak mengandung sitoplasma (Zwieniecki & Holbrook 2000), akan tetapi hifa-hifa jamur juga dapat bergerak ke sel-sel lainnya melewati noktah, sehingga terjalin akses pergerakan hifa dalam kayu baik pada arah longitudinal maupun arah lateral.
Noktah merupakan celah di antara dinding sekunder sel kayu. Umumnya noktah berpasangan di antara dua sel yang berhubungan (Gambar 25). Ada dua tipe noktah, yaitu noktah halaman (bordered pits) dan noktah sederhana (simple pits). Noktah sederhana menghubungkan antar sel-sel parenkim seperti antar sel jari-jari, sedangkan noktah halaman pada umumnya terdapat pada dinding sel-sel trakeid kayu daun jarum dan sel-sel serabut kayu daun lebar. Noktah halaman yang menghubungkan antar sel-sel trakeid memiliki membran. Membran tersebut terdiri atas pasangan dinding primer dan lamela tengah. Bagian luarnya disebut margo yang terdegradasi secara alami dan memungkinkan air melewatinya. Adapun bagian tengahnya tetap utuh membentuk torus (Butterfield 2006). Selain Gambar 25 Noktah sederhana (a) menghubungkan dua sel parenkim; noktah
halaman (b) menghubungkan rongga antar dua sel trakeid; dan noktah setengah halaman (c) menghubungkan sel trakeid dengan sel parenkim (Bowyer et al. 2003).
kedua tipe noktah tersebut ada juga noktah setengah halaman (half-bordered pits) yang menghubungkan sel trakeid dengan sel jari jari atau sel parenkim aksial (Fujita & Harada 2001). Pada dinding saluran pembuluh juga terdapat banyak noktah setengah halaman yang menghubungkannya dengan parenkim aksial dan parenkim jari-jari di sekitarnya (Bowyer et al. 2003). Kondisi struktur kayu seperti itu mempermudah bagi hifa-hifa jamur untuk menginvasi dan masuk ke bagian dalam kayu.
Untuk mendapatkan sumber nutrisi yang lebih banyak terutama selulosa dalam dinding sel kayu, jamur pelapuk terhalang oleh lamela tengah dan dinding primer yang kaya lignin. Jalan yang relatif mudah bagi jamur untuk dapat masuk ke dalam sel kayu pada umumnya melewati noktah terutama pada awal serangan dengan terlebih dahulu mendegradasinya secara enzimatik dan secara fisik oleh hifa yang sangat halus (Ø 1 µm). Secara alami, noktah pada dinding sel dapat dilewati air dalam proses fisiologi pohon terutama pada bagian margo yang lebih permeabel. Bahkan dalam kayu daun jarum, pergerakan air melalui sel-sel trakeid yang memiliki banyak noktah halaman (bordered pit) (Butterfield 2006). Dengan terdegradasinya noktah pada dinding sel serabut dan trakeid maka air lebih mudah masuk ke dalam sel kayu secara kapiler. Oleh karena itu kayu-kayu yang terserang jamur pelapuk cenderung lebih tinggi kadar airnya daripada kayu yang utuh.
Setelah berhasil masuk ke dalam rongga, hifa tumbuh dan menempel pada dinding sekunder. Maka terjadilah degradasi dinding sel kayu mulai dari dinding lumen sel (dari dalam rongga sel). Dinding sekunder sel kayu yang kaya selulosa terdegradasi terutama yang disekitar hifa sehingga terbentuk alur-alur erosi dan menyebabkan penipisan dinding sel kayu dari dalam (lumen) ke luar dinding sel (Schwarze et al. 2000 ). Degradasi dinding sel kayu ini terjadi secara enzimatik yang spesifik bagi setiap jenis jamur sehingga laju degradasi yang terjadi juga dapat berlainan antar jenis jamur.
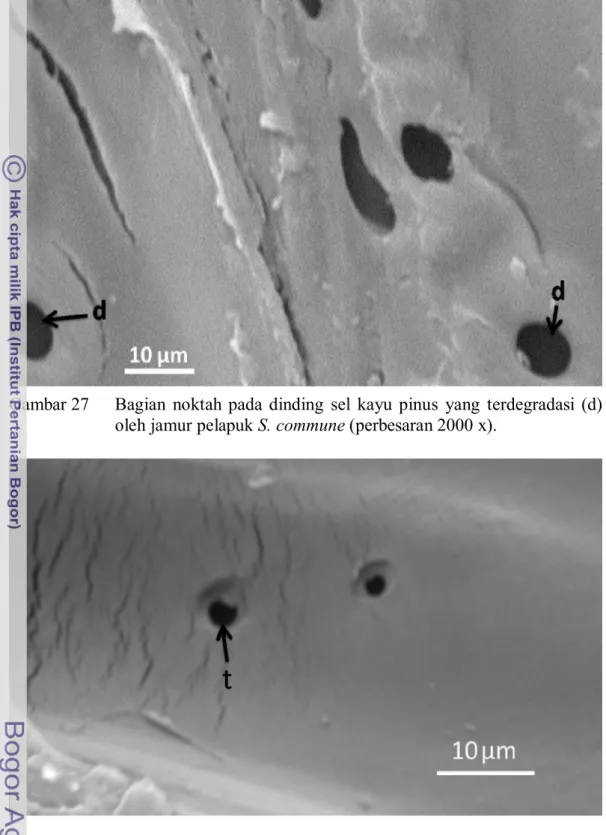
Scanning electron micrograph (5kV) menunjukkan bahwa torus dalam noktah halaman sel kayu Pinus insularis telah terdegradasi jamur G. applanatum dalam 4 minggu inkubasi, sedangkan jumlah tori terdegradasi semakin banyak dalam inkubasi 12 minggu (Gambar 26). Adapun noktah-noktah yang terdegradasi jamur S. commune setelah pengumpanan 4 minggu pada dinding sel kayu pinus relatif sedikit. Baru kemudian setelah 12 minggu inkubasi terjadi banyak noktah yang terdegradasi (Gambar 27). Ini membuktikan bahwa jamur S. commune lebih lambat dalam mendegradasi kayu pinus dibandingkan dengan jamur G. applanatum.
Pada kayu sengon, kerusakan noktah juga terjadi oleh jamur G. applanatum (Gambar 28). Sementara itu studi yang dilakukan oleh Green dan Clausen (1999) selama 3 minggu inkubasi ditemukan bahwa jamur pelapuk putih menghidrolisis semua membran noktah halaman, sedangkan jamur pelapuk coklat hanya merusak torus, melemahkan dan merobek membran noktah. Kedua jenis jamur sama-sama memiliki kapasitas menghidrolisis pektin, merusak membran noktah dan meningkatkan permeabilitas kayu selama kolonisasi sehingga lebih mudah menyerap air.
Gambar 26 Bagian noktah pada dinding sel kayu pinus yang terdegradasi (d) oleh jamur pelapuk G. applanatum (perbesaran 2000 x).
Gambar 28 Bagian noktah pada dinding sel kayu sengon (t) yang terdegradasi jamur pelapuk G. applanatum (perbesaran 2000 x).
Gambar 27 Bagian noktah pada dinding sel kayu pinus yang terdegradasi (d) oleh jamur pelapuk S. commune (perbesaran 2000 x).
Hasil pengamatan dengan mikroskop stereo pada kayu pinus dan sengon setelah uji pelapukan terhadap jamur G. applanatum dan S. commune menunjukkan terbentuknya lubang dan rongga-rongga yang dapat dilihat dengan mikroskop stereo (Gambar 29 dan 30). Rongga-rongga yang tampak pada bidang radial terutama disebabkan degradasi sel jari-jari dan sel-sel kayu di sekitarnya yang semakin lama semakin luas. Pada kayu pinus, lubang-lubang besar juga terjadi karena sel-sel trakeid di sekitar saluran interseluler terdegradasi sehingga memperbesar saluran tersebut. Adapun pada kayu sengon, lubang-lubang besar terjadi karena degradasi sel-sel parenkim aksial dan sel-sel serabut di sekitar saluran pembuluh. Banyaknya rongga-rongga dan lubang yang terbentuk dapat berpengaruh pada kekuatan kayu.
Masa inkubasi semakin lama, proses dedgradasi semakin keras sehingga terjadi kerusakan struktur sel. Pada kayu yang diinkubasi selama 8 dan 12 minggu lebih banyak terbentuk lubang dibandingkan dengan yang diinkubasi selama 4 minggu. Kayu uji menjadi lebih berongga-rongga setelah inkubasi 8 minggu terutama yang diserang jamur G. applanatum. Pola degradasi kayu Gambar 29 Rongga-rongga (r) yang terbentuk oleh jamur pelapuk G.
sengon dan pinus oleh jamur G. applanatum dan S. commune tersebut merupakan tipe pelapukan simultan.
Miselium yang tumbuh dalam lumen sel dapat membentuk lubang-lubang dan mendegradasi dinding sel secara progresif (Bowyer 2003). Levin dan Castro (1998) juga menemukan pola degradasi yang serupa pada kayu populus yang diserang oleh jamur pelapuk putih Trametes trogii, yaitu kayu menjadi berongga-rongga serta porous seperti spong, sedangkan pada kayu salix meninggalkan sel-sel pembuluh sehingga kayu menjadi stringy (terdiri atas banyak benang). Adapun Luna et al. (2004) mengungkapkan bahwa serangan jamur G. lucidum menyebabkan kombinasi delignifikasi selektif dan pelapukan simultan. Ciri utama delignifikasi selektif adalah terjadinya pemisahan antar sel-sel kayu, sedangkan pelapukan simultan ditandai dengan erosi dan penipisan dinding sel kayu serta terbentuknya rongga-rongga pada kayu. Dengan demikian jamur penyebab pelapukan simultan dapat menurunkan masa dan kekuatan kayu lebih besar dibandingkan dengan jamur penyebab delignifikasi selektif. Schwarze et al. (2000) telah membuktikan bahwa kayu beech yang lapuk simultan oleh jamur
Gambar 30 Rongga-rongga (r) yang terbentuk oleh jamur pelapuk S. commune pada kayu sengon (a) dan pinus (b).
Fomes fomentarius mengalami penurunan nilai keteguhan pukul lebih besar daripada yang mengalami delignifikasi selektif oleh G. pfeifferi.
Pada pelapukan lebih lanjut banyak dinding sel yang menipis bahkan habis terdegradasi sehingga terjadi peningkatan volume rongga dalam kayu dan kekuatannya menurun drastis. Bila kayu tersebut mengering, akan mudah mengalami collapse dan perubahan bentuk.
Pengaruh Serangan Jamur Pelapuk terhadap Sifat Kimia Kayu
Selulosa, hemiselulosa dan lignin adalah komponen kayu yang menjadi sumber nutrisi jamur pelapuk dengan terlebih dahulu diuraikan menjadi molekul-molekul sederhana dengan bantuan metabolit yang dihasilkan pada ujung dan sisi hifa (Bowyer et al. 2003). Hasil uji biodeteriorasi media serbuk kayu pinus dan sengon oleh jamur G. applanatum dan S. commune menunjukkan terjadinya gejala lapuk putih. Hal ini terbukti dengan perubahan warna media serbuk kayu dalam baglog dari coklat menjadi lebih pucat (coklat muda). Perubahan warna tersebut sudah tampak dalam masa inkubasi 2 minggu (Gambar 31) dan merupakan indikasi terjadinya degradasi lignin dalam kayu pinus dan sengon oleh jamur G. applanatum dan S. commune.
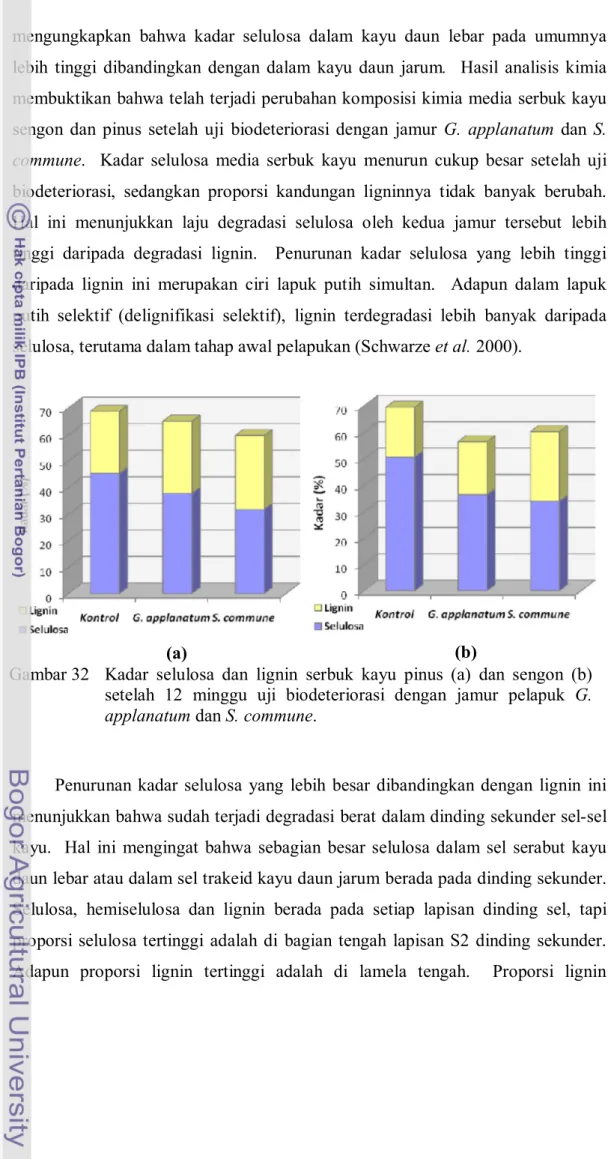
Penelitian ini menujukkan bahwa proporsi kandungan selulosa dalam kayu sengon lebih tinggi daripada dalam kayu pinus (Gambar 32). Walker (2006) juga
Gambar 31 Perubahan warna media serbuk kayu setelah diinokulasi dengan jamur pelapuk G. applanatum (GS) dan S. commune (SS) selama dua minggu (S=kontrol).
mengungkapkan bahwa kadar selulosa dalam kayu daun lebar pada umumnya lebih tinggi dibandingkan dengan dalam kayu daun jarum. Hasil analisis kimia membuktikan bahwa telah terjadi perubahan komposisi kimia media serbuk kayu sengon dan pinus setelah uji biodeteriorasi dengan jamur G. applanatum dan S. commune. Kadar selulosa media serbuk kayu menurun cukup besar setelah uji biodeteriorasi, sedangkan proporsi kandungan ligninnya tidak banyak berubah. Hal ini menunjukkan laju degradasi selulosa oleh kedua jamur tersebut lebih tinggi daripada degradasi lignin. Penurunan kadar selulosa yang lebih tinggi daripada lignin ini merupakan ciri lapuk putih simultan. Adapun dalam lapuk putih selektif (delignifikasi selektif), lignin terdegradasi lebih banyak daripada selulosa, terutama dalam tahap awal pelapukan (Schwarze et al. 2000).
Penurunan kadar selulosa yang lebih besar dibandingkan dengan lignin ini menunjukkan bahwa sudah terjadi degradasi berat dalam dinding sekunder sel-sel kayu. Hal ini mengingat bahwa sebagian besar selulosa dalam sel serabut kayu daun lebar atau dalam sel trakeid kayu daun jarum berada pada dinding sekunder. Selulosa, hemiselulosa dan lignin berada pada setiap lapisan dinding sel, tapi proporsi selulosa tertinggi adalah di bagian tengah lapisan S2 dinding sekunder. Adapun proporsi lignin tertinggi adalah di lamela tengah. Proporsi lignin
(a) (b)
Gambar 32 Kadar selulosa dan lignin serbuk kayu pinus (a) dan sengon (b) setelah 12 minggu uji biodeteriorasi dengan jamur pelapuk G. applanatum dan S. commune.
dibandingkan dengan komponen kimia lainnya semakin berkurang pada lapisan S1, S2, dan S3 (Bowyer et al. 2003).
Pada uji biodeteriorasi ternyata jenis jamur berpengaruh nyata terhadap komposisi selulosa dan lignin dalam kayu (Lampiran 17). Total kadar selulosa dan lignin kayu sengon dalam uji dengan jamur G. applanatum relatif lebih rendah dibandingkan dengan yang diumpankan terhadap jamur S. commune, sedangkan pada kayu pinus total kadar selulosa dan ligninnya lebih rendah pada media serbuk kayu yang diumpankan terhadap S. commune. Hal ini menunjukkan bahwa degradasi kayu sengon oleh jamur G. applanatum lebih besar daripada oleh jamur S. commune. Hasil penelitian Dill dan Kraeplin (1986) juga membuktikan bahwa G. applanatum dapat menimbulkan delignifikasi ekstensif dan penguraian sel-sel serabut dalam kayu.
Kayu yang terserang jamur pelapuk pada umumnya lebih lunak bila ditekan dengan kuku atau benda keras dan bila dicungkil dengan benda tajam serabut-serabutnya mudah putus. Adapun kayu utuh bila dicungkil benda tajam serabutnya tidah mudah putus. Menurut Senese (2010) selulosa merupakan komponen utama pada dinding sel kayu yang memberikan kekuatan pada kayu. Selulosa lebih berperan dalam kekuatan tarik aksial, sedangkan hemiselulosa dan lignin lebih menentukan elastisitas dan kekuatan tekan kayu (Ridout 2004). Oleh karena itu perubahan dalam komponen utama kayu ini akan merubah kemampuan struktural kayu. Degradasi selulosa dan lignin kayu akibat serangan jamur pelapuk pada struktur bangunan sangat berbahaya. Sebagai contoh, dalam struktur balok lentur yang biasa dipakai pada bagian dasar kuda-kuda atap, apabila terjadi pelapukan, maka kondisi balok menjadi kritis, karena bagian bawah balok lentur mengalami tegangan tarik, sedangkan pada bagian atasnya mengalami tegangan tekan sejajar serat.
Penurunan kadar selulosa oleh kedua jenis jamur pada kayu sengon lebih besar dibandingkan dengan yang terjadi pada kayu pinus. Kandungan dan komposisi lignin berperan penting dalam ketahanan kayu dari pelapukan. Pada Gambar 32 tampak bahwa proporsi lignin dalam kayu pinus lebih tinggi daripada dalam kayu sengon. Ridout (2004) juga melaporkan bahwa jamur pelapuk putih
lebih banyak menyerang jenis kayu daun lebar, karena banyak pelapuk putih kesulitan mendegradasi lignin dalam kayu daun jarum yang jenis dan kuantitasnya berbeda dengan kayu daun lebar.
Karakteristik lignin pinus yang merupakan kayu daun jarum berbeda dengan lignin pada sengon yang merupakan kayu daun lebar. Penyusun utama lignin dalam kayu daun jarum adalah unit fenilpropan dengan satu metoksil (guaiasil, G), sedangkan penyusun utama lignin dalam kayu daun lebar adalah unit fenilpropan dengan dua metoksil (siringil, S) dan guaiasil (Martinez et al. 2005). Kandungan lignin siringil berkaitan dengan kerentanan kayu dari serangan jamur pelapuk. Kayu yang tinggi kadar lignin siringilnya yaitu kayu red maple, lebih cepat terdegradasi jamur Trametes versicolor daripada kayu boxelder yang rendah kadar liginin siringilnya. Padahal kedua jenis kayu tersebut memiliki komposisi kimia dan sifat anatomi yang agak sama (John et al. 1994).
Selain lignin, perlindungan alami yang dimiliki kayu dari serangan organisme perusak adalah zat ekstraktif. Sebagaimana diungkapkan oleh Rowell et al. (2005) bahwa ekstraktif dalam kayu daun lebar maupun kayu daun jarum dapat menyebabkan kayu awet. Zat ekstraktif terdiri atas lemak, asam lemak, fenol, terpena, steroid, asam resin, rosin, lilin, dan senyawa organic lainnya. Menurut Harris (2001) jenis dan kadar ekstraktif menentukan daya tahan kayu dari pelapukan. Methanol merupakan ekstraktif kayu yang dapat menahan serangan jamur dan bisa digunakan sebagai bahan pengawet. Ekstraktif penahan pelapukan lainnya adalah tanin terlarut, courmarin, alkaloid, terpenoid, dan steroid, semuanya merupakan produk metabolit sekunder. Selain itu Harju et al. (2003) melaporkan bahwa kayu Scot pine (Pinus sylvestris L) yang tahan dari serangan jamur Coniophora puteana memiliki kandungan ekstraktif fenolik lebih tinggi dibandingkan dengan yang tidak tahan. Begitupula Venalainen et al. (2002) melaporkan bahwa keawetan kayu Scots pine dari jamur pelapuk terutama dipengaruhi oleh konsentrasi pinosylvin dan monometil eter yang merupakan senyawa fenolik yang tergolong stilbena.
Pengaruh Serangan Jamur Pelapuk terhadap Berat Kayu
Jamur G. applanatum dan S. commune merupakan jamur pelapuk putih yang mampu mendegradasi dinding sel kayu yang terdiri atas selulosa, hemiselulosa dan lignin menjadi gula sederhana secara enzimatik yang kemudian diserapnya. Dengan demikian, semakin banyak masa kayu yang terurai mengakibatkan penurunan berat kayu yang semakin tinggi dan menujukkan kemampuan mendegradasi jamur yang semakin tinggi.
Jamur G. applanatum dapat secara cepat menyelimuti ketiga jenis kayu uji (kamper, pinus dan sengon), sedangkan S. commune hanya menutup sebagian permukaan kayu uji (Gambar 33). Penurunan berat terjadi pada ketiga contoh uji kayu oleh jamur G. applanatum dan S. commune. Penurunan berat kayu uji tersebut semakin tinggi dengan semakin lamanya masa inkubasi, sebagaimana dapat dilihat pada Gambar 34.
Dalam konstruksi kayu di lapangan kehilangan masa kayu tersebut bisa lebih lambat daripada yang terjadi dalam eksperimen ini, karena di alam banyak mikroorganisme lain yang dapat menjadi penghambat pertumbuhannnya, atau karena kondisi lingkungannya yang tidak selamanya mendukung pertumbuhannya, terutama adalah ketersediaan air pada kayunya.
Gambar 33 Kolonisasi jamur G. applanatum (a) dan S. commune (b) pada contoh uji kayu.
Pada Gambar 35 juga terlihat bahwa kehilangan berat kayu sengon, kamper dan pinus yang disebabkan oleh jamur G. applanatum lebih besar dibandingkan dengan yang diakibatkan oleh jamur S. commune. Hal ini menunjukkan bahwa kemampuan jamur G. applanatum dalam mendegradasi kayu lebih tinggi dibandingkan dengan jamur S. commune. Bahkan pada kayu sengon, G. applanatum menyebabkan kehilangan berat kayu hampir dua kali yang diakibatkan S. commune, yaitu rata-rata masing-masing 12,6% dan 6,7% selama Gambar 34 Penurunan berat kering kayu kamper (a), pinus (b), dan sengon
(c) oleh jamur pelapuk G. applanatum dan S. commune.
(a)
(b)
12 minggu inkubasi. Dengan demikian kehadiran jamur G. applanatum dalam konstruksi harus lebih diwaspadai dan segera dikendalikan serangannya.
Nsolomo (2000) dalam penelitiannya juga membuktikan bahwa G. australe dan S. commune merupakan jamur pelapuk putih yang mengakibatkan penurunan berat kayu Ocotea sebesar 16% dan 2% selama empat bulan inkubasi. Degradasi kayu oleh G. australe ternyata nilainya lebih besar dibandingkan dengan degradasi oleh S. commune.
Di antara ketiga jenis kayu terbukti secara statistik bahwa kayu sengon paling tinggi penurunan beratnya, disusul kemudian oleh kayu kamper dan yang terkecil penurunan beratnya adalah kayu pinus (Lampiran 18). Hal ini menunjukkan bahwa kayu sengon paling tidak tahan dari serangan jamur pelapuk yang digunakan dalam penelitian ini. Sengon yang dikenal jenis kayu cepat tumbuh (fast growing species) sebagian besar terdiri atas kayu gubal dan tidak memiliki ekstraktif bersifat racun yang dapat melindunginya dari serangan jamur pelapuk.
Lapuk berat (lebih dari 10% penurunan berat kayu) oleh G. applanatum hanya terjadi pada kayu sengon, yang sudah terjadi dalam masa inkubasi 8 minggu. Kayu kamper dan pinus mengalami lapuk sedang disebabkan oleh jamur G. applanatum pada masa inkubasi berurutan 8 dan 12 minggu. Dalam penelitian ini jamur S. commune tidak menimbulkan lapuk berat walaupun pada kayu seperti sengon yang termasuk paling tidak awet.
Variasi ketahanan kayu dari serangan jamur pelapuk ini terutama dipengaruhi oleh kandungan zat ekstraktifnya sebagaimana diungkapkan oleh Butterfield (2006) bahwa akumulasi zat ekstraktif jenis polifenol dalam kayu teras menyebabkan penurunan kadar air dan dapat memperlambat pelapukan oleh jamur. Celimene et al. (1999) juga melaporkan bahwa pinosilvin yang diekstrak dari kayu Pinus banksiana dan Pinus resinosa, dapat menahan pertumbuhan jamur pelapuk putih. Ekstrak kayu kamper (Dryobalanops aromatica) banyak mengandung terpena dan asam lemak. Terpena dan polifenol merupakan senyawa antimikroba yang sering terdapat dalam kayu. Beberapa triterpen yang teridentifikasi dengan HPLC (High Performance Liquid Chromatographhy)
dalam kayu kamper adalah kapurone, dryobalanone, dipterocarpol, dan terpinihidrat, dengan proporsi yang bervariasi antar daerah pertumbuhan dan umur pohon (Ali et al. 1990).
Pengaruh Serangan Jamur Pelapuk terhadap Berat Jenis Kayu
Berat jenis (BJ) merupakan indikator kepadatan masa kayu. Kayu dengan BJ tinggi pada umumnya memiliki dinding sel yang tebal. Kayu yang telah diumpankan pada jamur pelapuk dan mengalami penurunan berat, berat jenisnya tampak tidak banyak berbeda dengan kayu kontrol (Gambar 35). Hasil analisis statistik juga menunjukkan bahwa jamur G. applanatum maupun S. commune tidak berpengaruh nyata pada berat jenis (BJ) kayu (Lampiran 19).
Perubahan BJ kayu yang tidak nyata ini dapat disebabkan oleh peristiwa collapse sel-sel kayu sehingga terjadi penurunan volume kayu yang tidak normal. Dinding sel kayu yang menipis dan menurun kekuatannya dapat mengalami collapse, sehingga dinding sel melipat ke dalam. Sebagaimana dilaporkan oleh Gambar 35 Berat jenis kayu kamper, pinus, dan sengon setelah pengumpanan
Erwin et al. (2008) bahwa dinding sel kayu dapat mengalami penipisan akibat serangan jamur pelapuk putih khususnya S. commune.
Perbedaan BJ antar jenis kayu pada umumnya merupakan sifat bawaan masing-masing jenis kayu. BJ kayu kamper yang lebih tinggi daripada BJ kayu sengon mengindikasikan bahwa dinding sel kayu kamper lebih tebal daripada dinding sel kayu sengon. Hal ini juga menentukan ketahanan relatif kayu kamper yang lebih tinggi daripada kayu sengon ketika diserang jamur pelapuk.
Pengaruh Serangan Jamur Pelapuk terhadap Modulus Lentur dan Modulus Patah Kayu
Hasil pengolahan data sifat mekanis modulus lentur (MOE) dan modulus patah (MOR) kayu dalam uji pelapukan terhadap jamur G. applanatum dan S. commune menunjukkan bahwa nilai MOE dan MOR kayu menurun setelah pengumpanan pada jamur pelapuk G. applanatum dan S. commune. Penurunan sifat mekanis kayu terjadi karena hifa-hifa jamur yang masuk ke dalam kayu dengan enzim yang dihasilkannya telah merusak sel-sel kayu sehingga terbentuk rongga-rongga dalam kayu. Dengan demikian terjadi pelemahan kekokohan struktur kayu seiring dengan semakin luasnya serangan jamur di dalam kayu.
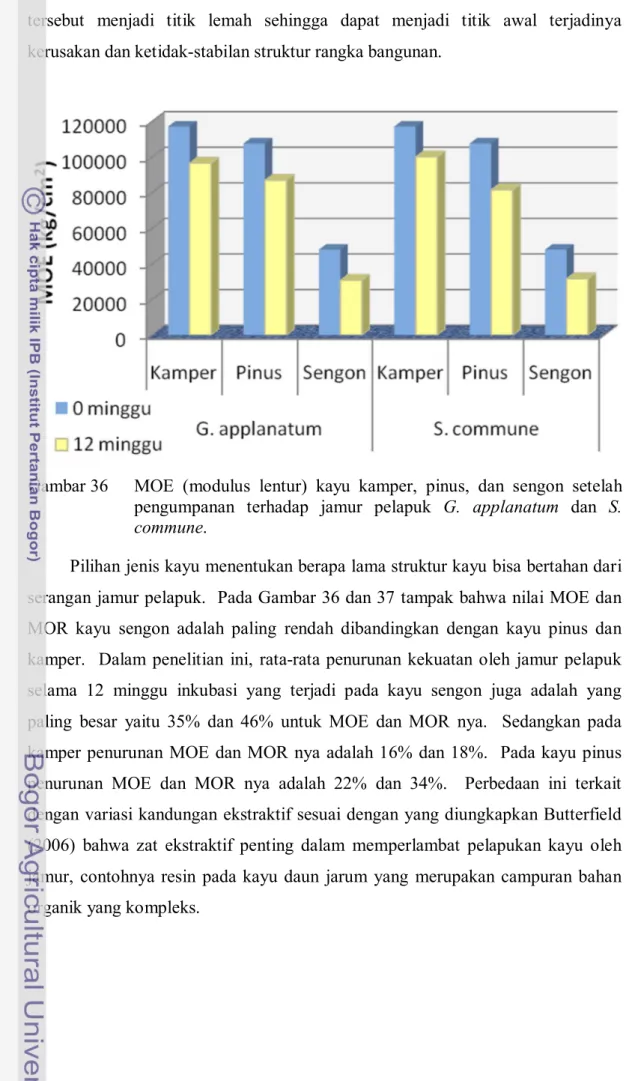
Secara statistik terbukti adanya perbedaan yang nyata pada nilai mekanis kayu yang diumpankan terhadap jamur dengan nilai kontrolnya (Lampiran 20). Pada umumnya nilai rata-rata penurunan sifat mekanis kayu oleh jamur G. applanatum cenderung lebih besar daripada oleh S. commune (Gambar 36 dan 37). Selama 12 minggu inkubasi penurunan MOE kayu kamper oleh G. applanatum dan S. commune adalah 17% dan 14%; penurunan MOR-nya berturut-turut adalah 22% dan 14%. Hal ini menujukkan bahwa daya rusak jamur G. applanatum pada kayu kamper lebih tinggi dibandingkan dengan jamur S. commune. Nilai penurunan sifat mekanis oleh kedua pelapuk putih ini cukup besar.
Dalam struktur rangka bangunan, penurunan kekuatan kayu ini bisa kritis karena walaupun tidak seluruh bagian batang lapuk terserang jamur, tapi bagian
tersebut menjadi titik lemah sehingga dapat menjadi titik awal terjadinya kerusakan dan ketidak-stabilan struktur rangka bangunan.
Pilihan jenis kayu menentukan berapa lama struktur kayu bisa bertahan dari serangan jamur pelapuk. Pada Gambar 36 dan 37 tampak bahwa nilai MOE dan MOR kayu sengon adalah paling rendah dibandingkan dengan kayu pinus dan kamper. Dalam penelitian ini, rata-rata penurunan kekuatan oleh jamur pelapuk selama 12 minggu inkubasi yang terjadi pada kayu sengon juga adalah yang paling besar yaitu 35% dan 46% untuk MOE dan MOR nya. Sedangkan pada kamper penurunan MOE dan MOR nya adalah 16% dan 18%. Pada kayu pinus penurunan MOE dan MOR nya adalah 22% dan 34%. Perbedaan ini terkait dengan variasi kandungan ekstraktif sesuai dengan yang diungkapkan Butterfield (2006) bahwa zat ekstraktif penting dalam memperlambat pelapukan kayu oleh jamur, contohnya resin pada kayu daun jarum yang merupakan campuran bahan organik yang kompleks.
Gambar 36 MOE (modulus lentur) kayu kamper, pinus, dan sengon setelah pengumpanan terhadap jamur pelapuk G. applanatum dan S. commune.
Uji Lapang Pelapukan Kayu
Hasil uji lapang pelapukan kayu tidak menyentuh tanah di berbagai daerah pada umumnya menunjukkan penurunan kekuatan kayu. Dalam hal ini nilai modulus lentur (MOE) dan modulus patah (MOR) kayu yang dipaparkan terhadap lingkungan selama 12 minggu lebih rendah dibandingkan dengan kayu kontrol baik sengon maupun kamper. Hal ini menunjukkan telah terjadinya degradasi kayu selama uji lapangan tersebut. Proses pelapukan pada umumnya terjadi secara dinamis oleh berbagai organisme. Sebagaimana diungkapkan oleh Nicholas dan Crawford (2003) bahwa bakteri, Actinomycetes dan Ascomycetes sering mendahului serangan pada kayu sebelum serangan jamur pelapuk Basidiomycetes yang lebih agresif dalam mendegradasi kayu.
Semakin tinggi indeks pelapukan (IP) daerah, maka nilai MOE dan MOR kayu yang diumpankan di daerah tersebut cenderung semakin rendah. Sebagai contoh di Bogor yang IP nya tinggi (157) nilai MOE dan MOR kayunya rata-rata Gambar 37 MOR (modulus patah) kayu kamper, pinus, dan sengon setelah
pengumpanan terhadap jamur pelapuk G. applanatum dan S. commune.
83.018,1 kg/cm2 dan 1.139,0 kg/cm2 untuk kamper, sedangkan untuk kayu sengon 28.235,5 kg/cm2 dan 325,6 kg/cm2. Adapun di Bekasi yang IP-nya 65, nilai MOE dan MOR kayu kamper setelah uji lapang pelapukan adalah 103.919,5 kg/cm2 dan 1.321,9 kg/cm2, sedangkan untuk kayu sengonnya 47.561,0 kg/cm2 dan 550,4 kg/cm2. Secara statistik terdapat korelasi yang kuat antara indeks pelapukan (IP) daerah dan degradasi yang terjadi pada kayu yang diumpankan di daerah tersebut yang dinyatakan dengan nilai MOE dan MOR-nya.
Hasil analisis ragam menunjukkan bahwa indeks pelapukan berpengaruh sangat nyata terhadap modulus lentur dan modulus patah kayu kamper dan sengon. Hasil uji korelasi dan analisis ragam indeks pelapukan (IP) dengan sifat mekanis kayu tersaji dalam Lampiran 21. Korelasi IP dengan sifat mekanis kayu tersebut ternyata lebih kuat dibandingkan dengan korelasi antara masing-masing faktor iklim dengan sifat mekanis kayu. Gambar 38 dan 39 mencerminkan hubungan indeks pelapukan dengan nilai MOE dan MOR kedua jenis kayu.
Pada indeks pelapukan di bawah 35 yang menunjukkan kelas bahaya pelapukan sedang, nilai MOE dan MOR kayu setelah uji pelapukan tampak tidak berbeda dengan kontrolnya. Hal ini menunjukkan bahwa dalam masa uji pelapukan tidak menyentuh tanah selama 12 minggu di daerah kelas bahaya pelapukan sedang, sifat mekanis kayu belum menurun. Dengan demikian untuk mendeteksi degradasi kayu berdasarkan sifat mekanis di daerah kelas bahaya sedang perlu waktu yang lebih lama dari 12 minggu.
Faktor suhu tidak menunjukkan korelasi yang baik dengan sifat mekanis kayu sebagai indikator degradasi kayu, mungkin karena perbedaan suhu antar kota ataupun kabupaten di Jawa relatif tidak besar, yang terendah 20 oC (Lembang) yang tertinggi 28 oC (Jakarta Utara). Secara umum jamur memiliki rentang toleransi suhu yang cukup lebar untuk hidupnya, sebagaimana dijelaskan Nicholas dan Crawford (2003), bahwa jamur tumbuh pada suhu 12-40 oC. Menurut Rao (2005) jamur pelapuk juga pada umumnya tumbuh dalam kondisi yang cukup lembab dan suhu yang relatif tinggi, yaitu pada kelembaban 95% dan suhu 20-40
o
C. Tapi menurut Bowyer et al. (2003) pertumbuhan jamur agak lambat pada suhu di atas 35 oC dan pada umumnya terhenti pada suhu lebih dari 38 oC. Di
daerah iklim sedang yang memiliki empat musim, suhu di bawah 12oC atau di atas 40oC bisa sering terjadi. Sehingga wajar serangan jamur pelapuk pada komponen bangunan di Indonesia relatif lebih tinggi dibandingkan dengan di negara-negara lain yang beriklim sedang.
Kelembaban udara relatif rendah korelasinya dibandingkan dengan curah hujan dengan nilai mekanis kayu setelah uji pelapukan. Bahkan secara statistik diantara faktor-faktor cuaca yang paling kuat korelasinya dengan degradasi kayu uji adalah curah hujan. Kelembaban udara di Lembang lebih tinggi dibandingkan dengan di Bogor, sedangkan degradasi kayunya lebih tinggi yang diumpankan di Bogor. Kelembaban di Lembang adalah 84,7%, di Bogor 83,9%. Dibandingkan dengan nilai mekanis kayu yang diuji pelapukan di Bogor (tersebut di atas), kayu yang diuji pelapukan di Lembang lebih tinggi MOE dan MOR nya, yaitu 92.984,9 kg/cm2 dan 1.203,1 kg/cm2 untuk kamper, sedangkan pada kayu sengon 34.240,9 kg/cm2 dan 397,6 kg/cm2. Di Bogor volume curah hujan dan jumlah hari hujan bulanannya, yaitu 346 mm dan 13 hari hujan per bulan. Sedangkan di Lembang 141 mm dan 12 hari hujan per bulan. Hal ini sesuai dengan penjelasan Leicester et al. (2003) bahwa laju pelapukan yang terjadi dipengaruhi oleh jenis kayu,
Gambar 38 Pengaruh indeks pelapukan daerah terhadap modulus lentur (MOE) kayu dalam uji lapang pelapukan.
indeks iklim (berdasarkan suhu dan curah hujan) serta waktu tenggang awal pelapukan.
Nilai mekanis kayu dalam uji pelapukan ini lebih kuat berkorelasi dengan jumlah hari hujan yang lebih dari 0.25 mm per bulan dibandingkan dengan curah hujan rata-rata bulanan. Kondisi tersebut juga tampak pada grafik hubungan faktor-faktor cuaca dengan degradasi kayu yang dinyatakan dengan variasi nilai MOE dan MOR kayu (Lampiran 22). Sebagai contoh, dua daerah yang memiliki volume curah hujan bulanan yang sama bisa berbeda jumlah hari hujannya. Bila volume curah hujan hariannya tinggi, maka jumlah hari hujannya akan lebih sedikit dibandingkan dengan yang volume curah hujan hariannya relatif rendah. Dalam hal ini kontinyuitas pembasahan dan ketersediaan air bagi pertumbuhan jamur lebih terjamin pada frekwensi hujan yang tinggi.
Gambar 39 Pengaruh indeks pelapukan daerah terhadap modulus patah (MOR) kayu dalam uji lapang pelapukan.
Dalam uji pelapukan tanpa menyentuh tanah ini terjadi serangan jamur pewarna terlihat dengan munculnya pewarnaan pada kayu yang diumpankan (Gambar 40). Secara visual serangan jamur pelapuk belum dapat dilihat dalam masa pengumpanan 12 minggu ini. Walau demikian tumbuhnya jamur pewarna pada kayu dapat menjadi petunjuk ke arah pelapukan kayu, karena proses pelapukan pada umumnya terjadi secara dinamis oleh sejumlah mikroorganisme. Sebagaimana yang diungkapkan oleh Nicholas dan Crawford (2003) bahwa kolonisasi kayu oleh mikroorganisme terjadi secara dinamis yaitu oleh bakteri, jamur pewarna dan pelapuk lunak, kemudian Basidiomycetes. Proses suksesi kolonisasi ini dalam kayu yang tidak diawetkan bisa terjadi dalam 70 hari-an.
Proses biodeteriorasi dalam uji lapangan ini banyak dipengaruhi oleh faktor cuaca. Pembasahan kayu oleh air hujan menjadikan kadar air kayu naik dan dapat ditumbuhi jamur. Selain itu, radiasi ultraviolet dari sinar matahari dapat menyebabkan degradasi lignin pada permukaan kayu walaupun sangat kecil dan dalam waktu yang lama. Sebagaimana dijelaskan oleh Kutz (2005) bahwa ultraviolet dari cahaya matahari dapat mengoksidasi komponen lignin pada kayu. Kerusakan yang tampak pada kayu akibat fotodegradasi adalah perubahan warna dan retak-retak. Padahal lignin merupakan komponen kimia pelindung dalam kayu.
Demikian pula halnya dengan pemanasan sinar matahari dan pembasahan air hujan yang berulang kali dalam waktu lama dapat menyebabkan perubahan dimensi dan retakan kayu. Pembasahan air hujan yang berulang kali juga dapat menyebabkan pencucian ekstraktif kayu secara bertahap. Sehingga ekstraktif dalam kayu kamper yang menjadikannya awet, lama kelamaan mengalami pencucian dan berkurang sifat proteksinya dari serangan organisme perusak.
Gambar 40 Pewarnaan pada kayu kamper (K) dan sengon (S) dalam uji lapang pelapukan.
Di Pulau Jawa banyak daerah yang memiliki curah hujan rata-rata bulanan yang tinggi, sehingga memiliki potensi pelapukan kayu yang tinggi. Pembasahan menyebabkan kayu lebih disukai jamur dan sangat mendukung untuk pertumbuhannya. Kayu pada kondisi pemakaian atau pada keadaan kering udara berkadar air sekitar 16%, sehingga jamur pelapuk akan sulit tumbuh. Hal ini sebagaimana dijelaskan Nicholas dan Crawford (2003) bahwa untuk mendukung terjadinya pelapukan kayu dibutuhkan kadar air minimal pada titik jenuh serat (28–30)%. Air sangat dibutuhkan untuk pertumbuhan jamur pelapuk, yaitu sebagai pelarut dalam proses metabolisme, pengangkutan metabolit, enzim dan organel, sebagai pengisi penting kerangka dan sebagai kekuatan pendorong pertumbuhan.
Dalam uji pelapukan lapangan ini juga terbukti bahwa dibandingkan dengan kayu kamper, kayu sengon menunjukkan penurunan sifat mekanis yang lebih besar, terutama nilai modulus patahnya (MOR) karena keawetan kayu sengon lebih rendah daripada kayu kamper. Sehingga kayu sengon lebih cepat terdegradasi oleh organisme perusak dibandingkan dengan kayu kamper. Kayu kamper dan sengon digunakan dalam pengujian ini mewakili kayu awet dan kayu tidak awet yang banyak digunakan masyarakat dalam konstruksi rumah. Menurut Martawijaya et al. (1989) kayu kamper termasuk kelas awet II, sedangkan kayu sengon termasuk kelas awet IV. Walupun termasuk katagori awet, kayu kamper juga terdegradasi yang ditandai dengan nilai mekanis (MOE dan MOR) lebih rendah dari kontrolnya.