HUBUNGAN MAMALIA DENGAN JENIS VEGETASI DI TAMAN NASIONAL
GUNUNG CIREMAI
AGUS PRIYONO KARTONO1), GUNAWAN2), IBNU MARYANTO3), SUHARJONO3) 1) Divisi Keanekaragaman Hayati dan Ekosistem Pusat Penelitian Lingkungan Hidup IPB Bogor,
email: apkartono@yahoo.com
2) Alumni Departemen Konservasi Sumberdaya Hutan & Ekowisata Fakultas Kehutanan IPB 3) Puslitbang Biologi-LIPI, Cibinong Bogor, email: ibnu_mar@yahoo.com
ABSTRACT
Relationship of mammals and vegetation species in Gunung Ciremai National Park. The study of fod resources and the distribution of mammalia was conducted in Palutungan, Linggarjati, Seda, Cibeureum, Sayana, Bandorasa, Pajambon and Sukamukti resort at Gunung Ciremai from May to July 2007. The mammals was observed in this study were Sus scrofa, Muntiacus muntjak, Paradoxurus hermaphroditus, Nycticebus javanicus, Trachypithecus auratus, Macaca fascicularis, and Presbytis aygula. The relationship between number of those species and vegetation density are presented. Further, the existence forrelation of the carnivore of Panthera pardus melas and Prionailurus bengalensis have positive correlation with Paradoxurus hermaphroditus. The observation also indicated that there are no significantly related correlation population distribution of Muntiacus muntjak vs Sus scrofa, Paradoxurus hermaphroditus and Presbytis aygula; Paradoxurus hermaphroditus vs Trachypithecus auratus; and Nycticebus javanicus vs Presbytis aygula.
Key words: Mammals, vegetation, Ciremai PENDAHULUAN
Populasi mamalia herbivora dipenga-ruhi oleh kombinasi antara ketersediaan pakan dengan tekanan predator (Richards & Coley 2007). Kebutuhan pakan berbagai jenis mamalia herbivora berbeda-beda menurut jenis, umur, serta ketersediaan pakan baik jenis tumbuhan pakan maupun produktivitas pakan. Namun demikian, herbivora tidak memakan tumbuhan pakan secara acak tetapi cenderung memilih jenis tumbuhan tertentu yang disukainya (Colebrook et al. 1990, Tolkamp et al. 1998) yang terutama dipengaruhi oleh dua faktor, yakni: (a) kandungan nutrisi dan/atau zat beracun pada bagian tumbuhan tertentu (van Wiesen 1996) dan (b) ketersediaan relatif pakan secara spasial dan temporal (Belovsky & Schmitz 1994). Faktor lain yang mempengaruhi pemilihan jenis pakan oleh jenis-jenis mamalia herbivora adalah ukuran tubuh dan morfologi pencernaan yang pada umumnya dapat dinyatakan sebagai adaptasi evolusioner terhadap faktor-faktor dasar tersebut (Milton 1979, Laska et al. 2003).
Ketersediaan relatif sumber pakan herbivora dipengaruhi oleh produktivitas serta distribusi spasial tumbuhan. Di wila-yah tropis, peningkatan ketinggian tempat mengakibatkan terjadinya penurunan keanekaragaman spesies tumbuhan,
menurunnya tinggi maksimum pohon serta mengecilnya ukuran daun sehingga produk-tivitas biomassa menurun (Ohsawa 1995). Produktivitas hijauan pakan yang tinggi terjadi pada tapak-tapak rumpang akibat pohon rebah karena intensitas cahaya yang sampai ke permukaan tanah lebih tinggi dibanding dengan areal hutan rapat (Richards & Coley 2007).
Kepadatan herbivora dan predator pada areal berumpang ini lebih tinggi diban-ding dengan areal hutan yang relatif rapat. Jika produktivitas hijauan pakan rendah maka sumberdaya pakan menjadi pembatas herbivora, yang pada akhirnya berpengaruh pada populasi predator. Sebaliknya, jika produktivitas hijauan pakan tinggi maka populasi herbivora akan meningkat sampai pada tingkat yang dapat mendukung popu-lasi predator secara substansial sehingga populasi herbivora akan dibatasi oleh predator (Richards & Coley 2007).
Hubungan keanekaragaman spesies mamalia herbivora dengan keanekaragam-an jenis vegetasi di kawaskeanekaragam-an TNGC perlu diketahui sehingga upaya konservasi jenis-jenis mamalia tersebut dapat mencapai sasaran secara baik dan benar. Tujuan penelitian ini adalah mengidentifikasi hu-bungan keanekaragaman mamalia herbi-vora dengan kerapatan jenis-jenis vegetasi serta mengidentifikasi jenis-jenis vegetasi
dominan penentu sebaran populasi mama-lia herbivora.
BAHAN DAN CARA KERJA
Penelitian ini dilakukan di kawasan Seksi Pengelolaan Taman Nasional (SPTN) Wilayah Kuningan Taman Nasional Gunung Ciremai (TNGC), Kabupaten Kuningan Provinsi Jawa Barat. Pengumpulan data lapangan dilakukan pada 16 Mei sampai 29 Juli 2007. Peralatan yang digunakan terdiri atas: teropong binokuler, kompas brunton, altimeter, GPS receiver, kamera digital, pita meter, tali rafia serta buku panduan lapang-an mamalia. Pengumpullapang-an data mamalia dilakukan dengan menggunakan unit con-toh berbentuk transek jalur (strip transect) dengan panjang transek 1.750 m dan lebar 100 m. Jumlah unit contoh pengamatan mamalia sebanyak 12 transek jalur yang diletakkan pada delapan lokasi, yakni tiga jalur di Palutungan, Linggarjati dan Seda masing-masing dua jalur, serta di Cibeu-reum, Sayana, Bandorasa, Pajambon dan Sukamukti masing-masing sebanyakn satu jalur pengamatan. Pengumpulan data vege-tasi dilakukan melalui penarikan unit contoh berbentuk jalur berpetak berukuran panjang 100 m dengan lebar 20 m. Lokasi pengumpulan data vegetasi ditempatkan sesuai dengan letak unit contoh transek jalur pengamatan mamalia. Posisi areal pengamatan terletak pada 06o50'11.6"– 06o56'16.3" LS dan 108o23'48.3"– 108o28'33.0" BT. Analisis data populasi mamalia dilakukan dengan menggunakan metode King, sedangkan analisis hubungan mamalia dengan jenis-jenis vegetasi dilaku-kan dengan menggunadilaku-kan pendekatan regresi linear berganda. Penentuan jenis-jenis vegetasi yang mempengaruhi kebera-daan mamalia herbivora dilakukan dengan menggunakan metode regresi dalam pola
stepwise. Perangkat lunak yang digunakan
untuk analisis data adalah SPSS for Windows
ver 13.
HASIL Kondisi Vegetasi
Kawasan TNGC memiliki tipe penutup-an lahpenutup-an ypenutup-ang terdiri atas: areal kebun,
padang alang-alang, hutan tanaman pinus, dan hutan alam. Berdasarkan formasi hutan, kawasan TNGC merupakan hutan hujan tropis yang terbagi atas tiga formasi, yakni: hutan dataran rendah, hutan pegu-nungan dan hutan sub alpin. Total jenis vegetasi yang ditemukan dalam plot peng-amatan sebanyak 189 jenis, namun yang dapat diidentifikasi sebanyak 166 jenis dari 59 famili, terdiri atas 120 jenis vegetasi pohon dan 46 jenis vegetasi non-pohon (rumput, semak, perdu dan herba). Berda-sarkan jumlah anggota spesies pada satu famili maka terdapat lima famili tumbuhan yang mendominasi, yakni Euphorbiaceae 15 jenis (9,04%), Lauraceae 14 jenis (8,43%), Moraceae 14 jenis (8,43%), Myrtaceae 8 jenis (4,82%), dan Rubiaceae 8 jenis (4,82%). Jenis-jenis vegetasi yang teriden-tifikasi disajikan pada Lampiran 1.
Sebaran jumlah jenis vegetasi pohon berdasarkan formasi hutan adalah di kawasan hutan tanaman pinus (700–850 mdpl) sebanyak 14 jenis, hutan dataran rendah (500–1000 mdpl) sebanyak 69 jenis, hutan pegunungan (1000–2400 m dpl) sebanyak 86 jenis dan hutan sub alpin (2400–2700 mdpl) sebanyak 38 jenis. Total jumlah jenis vegetasi sumber pakan mamalia herbivora yang ditemukan di kawasan TNGC sebanyak 45 jenis, terdiri atas 39 jenis ditemukan dalam plot peng-amatan dan 6 jenis ditemukan di luar plot pengamatan. Namun demikian, jumlah jenis yang ditemukan dalam plot pengamatan yang dapat diidentifikiasi hanya 38 jenis dari 19 famili. Jenis vegetasi sumber pakan dari didominasi oleh famili Moraceae, Arecaceae, Euphorbiaceae dan Myrtaceae. Famili Moraceae, Euphorbiaceae dan Myrtaceae ditemukan hampir pada semua formasi hutan, kecuali di kawasan hutan tanaman pinus. Sebaran jenis-jenis vegetasi potensial sebagai pakan tersebut adalah sebagai berikut: di kawasan hutan tanaman pinus sebanyak 5 jenis, hutan dataran rendah 32 jenis, hutan pegunungan 30 jenis dan hutan sub alpin sebanyak 15 jenis.
Total kepadatan jenis vegetasi poten-sial sebagai sumber pakan bagi mamalia herbivora di TNGC tertinggi terdapat di kawasan hutan sub alpin, yakni sebanyak 112.750,00 ind./ha sedangkan terendah
adalah di kawasan hutan tanaman pinus yakni sebanyak 3.200,00 individu/ha. Kerapatan jenis-jenis vegetasi potensial
sebagai sumber pakan mamalia herbivora berdasarkan habitus disajikan pada Tabel 1. Tabel 1. Kerapatan jenis-jenis vegetasi sumber pakan mamalia herbivora di kawasan TNGC
Habitus Kepadatan vegetasi sumber pakan mamalia herbivora (individu/ha)
HtPn HtDR HtPg HtSA Tumbuhan bawah 0 24.166,66 48.000,00 0 Semai 0 66.666,66 26.000,00 80.000,00 Pancang 3.200,00 15.200,00 10.560,00 27.200,00 Tiang 0 1.600,00 2.240,00 2.550,00 Pohon 0 1.366,65 1.580,00 3.000,00 Jumlah 3.200,00 108.999,97 88.380,00 112.750,00
Keterangan: HtPn=hutan tanaman pinus, HtDR=hutan dataran rendah, HtPg=hutan pegunungan, HtSA=hutan sub
alpin
Jenis-jenis vegetasi tumbuhan bawah yang mendominasi kawasan hutan tanaman pinus adalah Eupatorium odoratum (INP 28,68%), sedangkan pada tingkat semai didominasi oleh jenis Caliandra caliandra (INP 90,47%). Pada tingkat pancang didominasi oleh Dillenia excelsa (INP 86,36%), tingkat tiang dan pohon didomi-nasi oleh Pinus merkusii dengan INP berturut-turut 268,32% dan 275,85%. Di kawasan hutan dataran rendah, jenis-jenis vegetasi yang dominan adalah sebagai berikut: tumbuhan bawah didominasi jenis jukut galunggung (INP 32,71%), tingkat semai didominasi jenis Ficus montana (INP 38,89%), tingkat pancang didominasi jenis
C. caliandra (INP 19,04%), tingkat tiang
adalah jenis pulus Ficus sp. (INP 22,09%) dan tingkat pohon didominasi jenis F.
variegata (INP 31,18%). Jenis-jenis vegetasi
yang dominan di kawasan hutan pegu-nungan adalah Strobilanthes crispus (INP 49,00%) untuk tumbuhan bawah,
Cleroden-drum serrature (INP 35,49%) untuk tingkat
semai, Lithocarpus ewyckii (INP 31,35%) untuk tingkat pancang dan Castanopsis
javanica (INP 34,87%) untuk tingkat pohon.
Di kawasan hutan sub alpin, jenis tumbuhan bawah yang dominan adalah S. crispus (INP 55,64%), vegetasi tingkat semai didominasi jenis Flemingia strobilifera (INP 34,79%), tingkat pancang didominasi jenis
Actino-daphne macroptera (INP 29,52%), tingkat
tiang didominai jenis Psychotria viridiflora (INP 38,78%) dan tingkat pohon didominasi jenis L. ewyckii (INP 41,79%).
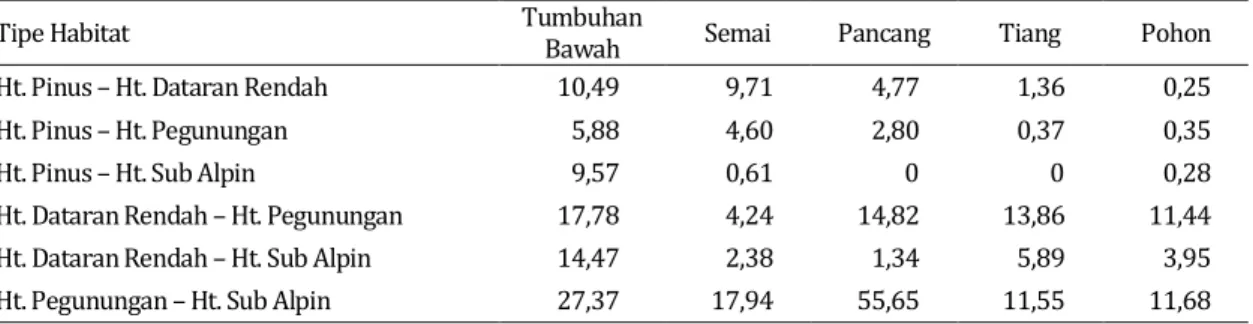
Tingkat kesamaan komunitas berbagai formasi hutan berdasarkan indeks Sorensen untuk berbagai tingkat vegetasi di kawasan TNGC sangar rendah. Maksimum indeks kesamaan komunitas terjadi pada vegetasi tingkat pertumbuhan pancang antara formasi hutan pegunungan dengan hutan sub-alpin. Indeks kesamaan komunitas berbagai formasi hutan di kawasan TNGC disajikan pada Tabel 2.
Tabel 2. Indeks kesamaan komunitas vegetasi pada berbagai formasi hutan di kawasan TNGC
Tipe Habitat Tumbuhan Bawah Semai Pancang Tiang Pohon
Ht. Pinus – Ht. Dataran Rendah 10,49 9,71 4,77 1,36 0,25
Ht. Pinus – Ht. Pegunungan 5,88 4,60 2,80 0,37 0,35
Ht. Pinus – Ht. Sub Alpin 9,57 0,61 0 0 0,28
Ht. Dataran Rendah – Ht. Pegunungan 17,78 4,24 14,82 13,86 11,44
Ht. Dataran Rendah – Ht. Sub Alpin 14,47 2,38 1,34 5,89 3,95
Mamalia Herbivora
Kekayaan Jenis
Mamalia herbivora yang ditemukan di kawasan TNGC sebanyak tujuh jenis dari lima famili, yakni: babi hutan Sus scrofa, kijang muncak Muntiacus muntjak, musang
luwak Paradoxurus hermaphroditus, kukang jawa Nycticebus javanicus, lutung budeng
Trachypithecus auratus, monyet ekor
panjang Macaca fascicularis dan surili
Presbytis aygula. Kepadatan jenis-jenis
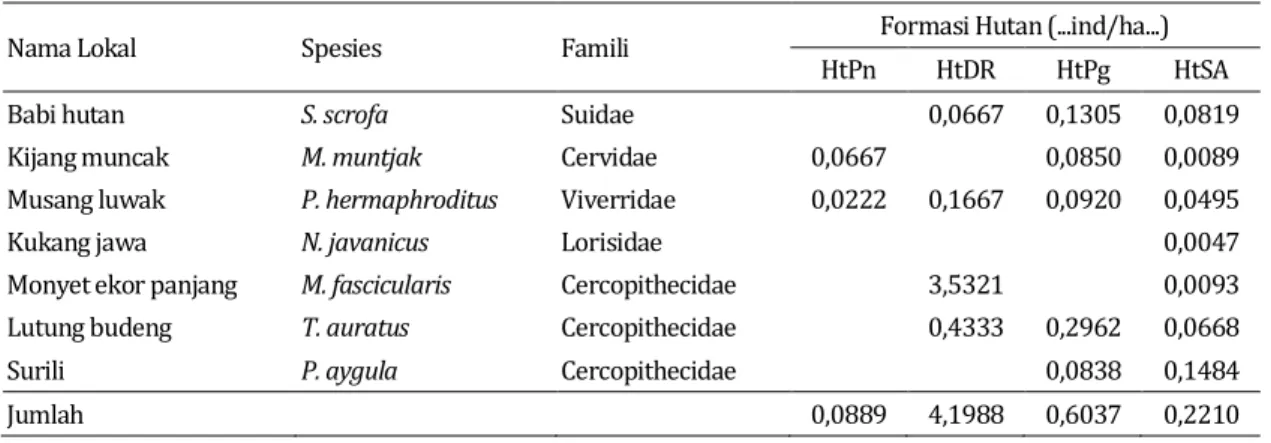
mamalia herbivora disajikan pada Tabel 3. Tabel 3. Rata-rata kepadatan jenis-jenis mamalia herbivora berdasarkan formasi hutan di
kawasan TNGC
Nama Lokal Spesies Famili Formasi Hutan (...ind/ha...)
HtPn HtDR HtPg HtSA
Babi hutan S. scrofa Suidae 0,0667 0,1305 0,0819
Kijang muncak M. muntjak Cervidae 0,0667 0,0850 0,0089
Musang luwak P. hermaphroditus Viverridae 0,0222 0,1667 0,0920 0,0495
Kukang jawa N. javanicus Lorisidae 0,0047
Monyet ekor panjang M. fascicularis Cercopithecidae 3,5321 0,0093
Lutung budeng T. auratus Cercopithecidae 0,4333 0,2962 0,0668
Surili P. aygula Cercopithecidae 0,0838 0,1484
Jumlah 0,0889 4,1988 0,6037 0,2210
Keterangan: HtPn=hutan tanaman pinus, HtDR=hutan dataran rendah, HtPg=hutan pegunungan, HtSA=hutan sub
alpin
Jenis Vegetasi Penentu Sebaran Populasi
Babi hutan (S. scrofa) merupakan jenis mamalia dari famili Suidae yang ditemukan di TNGC. Jenis mamalia ini termasuk omnivora karena pakannya meliputi buah-buahan yang jatuh dan biji-bijian, akar-akaran, terna dan bahan tumbuhan lainnya, cacing tanah, serta binatang kecil lainnya (Payne et al. 2000). Makanan babi hutan selalu berubah-ubah menurut musim sepanjang tahun, namun yang paling banyak dikonsumsi adalah buah-buahan dan biji-bijian. Di South Carolina, babi hutan banyak memakan berbagai jenis rumput pada musim semi, sedangkan pada musim panas banyak mengkonsumsi akar-akaran, tumbuh-tumbuhan, akar umbi dan daun-daunan. Di daerah beriklim lebih hangat, babi hutan memakan apa saja yang menjadi makanannya berdasarkan ketersediaan pakan tersebut di habitatnya (Wood & Roark 1980).
Sebaran babi hutan di kawasan TNGC meliputi kawasan hutan dataran rendah, hutan pegunungan dan hutan sub alpin. Keberadaan babi hutan di kawasan ini ditentukan oleh 11 jenis vegetasi, yakni kiplik Ficus sp., surian Palaquium
impressinervium, kiputri Podocarpus neriifolius, huru kopi Saprosma arborea,
mareme Glochidion arborescens,
cangcaratan Psychotria sp., huru madam
Litsea sanguinolenta, pakis Pityrogramma calomelanos, kimeong Timonius sp., kiseer Antidesma tetandrum, dan pasang dadap Lithocarpus sundaicus (F=8,561; p=0,00; r2=0,299). Hubungan keberadaan babi hutan (Y) dengan kepadatan (individu/ha) jenis-jenis vegetasi disajikan pada Tabel 4.
Salah satu jenis mamalia herbivora dari famili Cervidae yang ditemukan di TNGC
adalah kijang muncak. Jenis ini
membutuhkan makanan terutama berupa buah dan semak, serta hanya memakan secara selektif bagian-bagian tumbuhan. Kijang muncak menempati habitat pada satu atau beberapa tipe vegetasi sepanjang tahun, namun lebih banyak menempati habitat daerah bagian atas (Meijaard & Sheil 2008). Perumputan menempati proporsi yang lebih tinggi dalam pemenuhan kebutuhan pakan bagi kijang muncak dibanding dengan pakan buah (Barrette 1977).
Sebaran jenis kijang muncak di kawasan TNGC meliputi tiga formasi hutan, yakni hutan tanaman pinus, hutan
pegunungan dan hutan sub alpin. Jenis ini tidak ditemukan pada formasi hutan dataran rendah. Jenis-jenis vegetasi penting yang menentukan sebaran kijang muncak di TNGC adalah: mangga pari Mangifera indica, huru langsep Aglaia squamulosa, rukem
Falcourtia rukam, huru kawoyang Prunus arborea, kitiwu Miliosma pinnata, calodas Ficus microcarpa, huru mungkal
Actinodaphne procera, dan kerteu Alangium rotundifolium (F=7,19; p=0,00; r2=0,19). Hubungan kijang muncak dengan jenis-jenis vegetasi disajikan pada Tabel 4.
Musang luwak merupakan jenis mamalia omnivora yang ditemukan pada seluruh formasi hutan di kawasan TNGC. Jenis ini merupakan satwaliar nokturnal arboreal dan terestrial, tetapi lebih aktif di permukaan tanah. Jenis pakan bagi musang luwak meliputi buah-buahan, dedaunan, arthropoda, cacing tanah dan moluska (Payne et al. 2000). Musang luwak dapat hidup pada habitat hutan tinggi, hutan sekunder, perkebunan dan kebun masyarakat. Sebaran jenis musang luwak di TNGC dipengaruhi oleh sebaran jenis nangsi
Villebrunea rubescens dan panggang Trevesia sundaica (F=5,025; p=0,007; r2=0,038). Hubungan antara keberadaan musang luwak dengan jenis vegetasi disajikan pada Tabel 4.
Kukang merupakan primata yang hidup di hutan tropis Indonesia, menyukai hutan primer dan sekunder, semak belukar dan rumpun-rumpun bambu. Di Indonesia terdapat dua spesies kukang, yakni: N.
coucang yang tersebar di Semenanjung
Malaya, Sumatera, Kalimantan serta kepulauan sekitarnya; dan N. javanicus yang penyebarannya hanya di Jawa. Kukang jawa bersifat nokturnal dan arboreal pada sebagian besar pepohonan berukuran kecil sampai sedang. Jenis makanan kukang jawa meliputi binatang-binatang kecil, sebagian besar serangga, dan buah-buahan yang berdaging (Payne et al. 2000). Sebaran populasi kukang jawa di kawasan TNGC dipengaruhi oleh jenis-jenis vegetasi kiputih
Litsea cuceba, kilaki Zizyphus horsfieldii,
huru jambu Pisonia umbellifera, kitambaga
Syzygium antisepticum, saninten C. javanica
dan nangsi V. rubescens (F=9,686; p=0,000;
r2=0,306)(Tabel 4).
Lutung budeng atau lutung jawa terdiri atas dua sub spesies, yakni lutung budeng
T.a. auratus yang berada di kawasan hutan
Jawa Timur dan lutung Jawa Barat T.a.
mauritus. Lutung budeng termasuk dalam
kelompok pemakan daun (folivorous). Sumber pakan bagi lutung budeng adalah jenis-jenis pohon, perdu, epifit/parasit pada pohon dan paku-pakuan. Pergerakan harian kelompok lutung antara 92–132 m/hari dengan luas wilayah jelajah antara 12,4– 18,5 ha. Jenis-jenis vegetasi yang menentukan sebaran populasi lutung budeng di kawasan TNGC adalah kiputri P.
neriifolius, huru kopi S. arborea, kiplik Ficus sp., mareme G. arborescens dan surian Palaquiurn impressinervium (F=5,404; p=0,000; r2= 0,098) (Tabel 4).
Monyet ekor panjang merupakan salah satu jenis mamalia pemakan buah (frugivorous) yang sangat selektif. Primata ini memakan buah dan daun-daun muda dari genus Ficus, Dillenia, Diospyros,
Koordersiodendron, Dracontomelon, Bambusa sp. dan beranekaragam jenis
lainnya (Kurland 1973). Komposisi bagian tumbuhan yang dimakan oleh monyet ekor panjang terdiri atas: daun sebanyak 49,93%, buah 38,54%, bunga 6,60% dan lain-lain sebanyak 5,94% (Sugiharto 1992). Selain sebagai frugivorous, jenis primata ini juga mempunyai alternatif sumber pakan lain, yakni: serangga, rumput, jamur, moluska, krustase, akar, umbi dan telur burung (Lindburg 1980). Ficus spp. merupakan jenis vegetasi yang menjadi sumber pakan utama bagi monyet ekor panjang karena dapat menghasilkan dedaunan muda sepanjang tahun dan berbuah 2-3 kali setiap tahun (Chivers 1980).
Di kawasan TNGC, jenis-jenis vegetasi yang menentukan keberadaan monyet ekor panjang adalah: binuang Tetrameles
nudiflora, kigambir Uncaria gambir, salam Syzygium polyanthum, huru kadu Elaeagnus latifolius, kopi Canthium confertum, carui Pterospermum javanicum, huru kadoya Dysoxylum alliaceum, sawo manggung Casimiroa edulis, kisampang Evodia latifolia,
simpur Dillenia excelsa, benda Artocarpus
elasticus, kibonteng Platea latifolia, kacu Artocarpus sp., gintung, afrika, beringin Ficus benjamina dan ipis kulit Decaspermum
fruticosum (F=12,293; p=0,000; r2=0,47) (Tabel 4).
Surili merupakan primata pemakan daun (folivorous). Komposisi pakan surili meliputi daun 65%, buah 14%, bunga 7% dan biji 1% (Rowe 1996). Di kawasan hutan konsesi Unocal Geothermal Indonesia Gunung Salak Sukabumi, jumlah jenis vegetasi yang dimanfaatkan oleh surili sebagai sumber pakan sebanyak 36 jenis dari 19 famili. Pakan utama surili adalah jenis-jenis vegetasi dari famili Moraceae 26,47% dan Fagaceae 11,76%. Jenis-jenis
vegetasi lain adalah dari famili Euphorbiaceae 8,82%, Araliaceae 5,88%, Myrtaceae 5,88%, Rubiaceae 5,88% dan famili lainnya 35,29% (Siahaan 2002). Di kawasan TNGC, sebaran populasi surili dipengaruhi oleh jenis-jenis vegetasi kiputri
P. neriifolius, huru kopi S. arborea, mareme G. arborescens, surian P. impressinervium,
kiplik Ficus sp., cangcaratan Psychotria sp., huru madam L. sanguinolenta, pasang L.
ewyckii dan pasang dadap L. sundaicus
(F=8,494; p=0,000; r2=0,239) (Tabel 4). Tabel 4. Hubungan keberadaan mamalia (Y) dengan kepadatan (individu/ha) jenis-jenis
vegetasi
Babi hutan (Sus scrofa)
Y = 0,074 + 0,003.Kiplik + 0,001.Surian + 0,006.Kiputri + 0,003.Huru_kopi + 0,003.Mareme + 0,006.Cangcaratan + 0,001.Huru_madam + 0,001.Pakis + 0,005.Kimeong + 0,003.Kiseer + 0,002.Pasang_dadap
Kijang muncak (Muntiacus muntjak)
Y = 0,028 + 0,007.Mangga_pari + 0,014.Huru_langsep + 0,003.Rukem + 0,005.Huru_kawoyang + 0,007.Kitiwu + 0,005.Calodas + 0,006.Huru_mungkal + 0,004.Kerteu
Musang luwak (Paradoxurus hermaphroditus)
Y = 0,065 + 0,001.Nangsi + 0,003.Panggang
Kukang (Nycticebus javanicus)
Y = 0,002 + 0,001.Kiputih + 0,001.Kilaki + 0,005.Huru_jambu + 0,001.Kitambaga + 0,0000539.Saninten + 0,0000669.Nangsi
Lutung budeng (Trachypithecus auratus)
Y = 0,199 + 0,016.Kiputri + 0,008.Huru_kopi + 0,007.Kiplik + 0,009.Mareme + 0,004.Surian
Monyet ekor panjang (Macaca fascicularis)
Y = 0,288 + 0,132.Binuang + 0,395.Kigambir + 0,296.Salam + 0,395.Huru_kadu + 0,296.Kopi + 0,084.Carui + 0,036.Huru_kadoya + 0,065.Sawo_manggung + 0,065.Kisampang + 0,092.Simpur + 0,087.Benda + 0,079.Kibonteng + 0,078.Kacu + 0,133.Gintung + 0,564.Afrika + 0,244.Beringin + 0,034.Ipis_kulit
Surili (Presbytis aygula)
Y = 0,066 + 0,008.Kiputri + 0,004.Huru_kopi + 0,005.Mareme + 0,002.Surian + 0,004.Kiplik + 0,007.Cangcaratan + 0,001.Huru_madam + 0,001.Pasang + 0,003.Pasang_dadap Berdasarkan hubungan jenis-jenis
vegetasi dengan sebaran mamalia herbivora maka terdapat 44 jenis vegetasi dari 23 famili yang menentukan sebaran mamalia di kawasan TNGC. Famili vegetasi yang paling menentukan sebaran mamalia adalah Moraceae, Rubiaceae dan Lauraceae.
Hubungan Antar Mamalia Herbivora
Jenis-jenis satwaliar mamalia herbivora yang ditemukan di kawasan hutan TNGC dapat dikelompokkan ke dalam dua kategori, yakni mamalia herbivora terestrial dan arboreal. Jenis satwaliar yang termasuk dalam herbivora terestrial adalah babi hutan, kijang muncak dan musang luwak; sedangkan herbivora arboreal
adalah kukang jawa, lutung budeng, monyet ekor panjang dan surili.
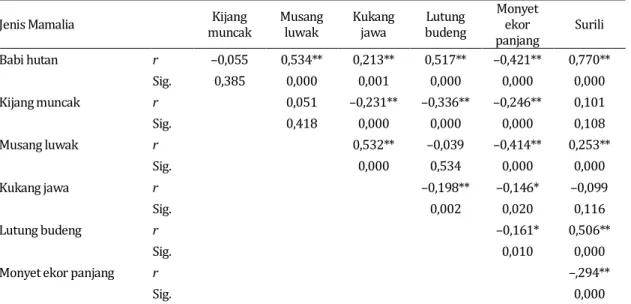
Berdasarkan korelasi Pearson (r) maka sebaran populasi babi hutan memiliki hubungan positif yang sangat nyata dengan musang luwak, kukang jawa, lutung budeng dan surili; tetapi berhubungan negatif sangat nyata dengan monyet ekor panjang. Populasi kijang muncak berhubungan negatif sangat nyata dengan kukang jawa, lutung budeng dan monyet ekor panjang; sedangkan musang luwak berhubungan berhubungan positif sangat nyata dengan kukang jawa dan surili tetapi berhubungan negatif sangat nyata dengan monyet ekor panjang. Jenis mamalia herbivora terestrial kukang jawa berhubungan negatif sangat nyata dengan lutung budeng dan monyet ekor panjang. Lutung budeng memiliki
hubungan negatif nyata dengan monyet ekor panjang tetapi berhubungan positif sangat nyata dengan surili; sedangkan monyet ekor panjang berhubungan negatif
sangat nyata dengan surili. Koefisien korelasi dan tingkat signifikansi antar mamalia herbivora disajikan pada Tabel 5. Tabel 5. Koefisien korelasi antar jenis mamalia herbivora di kawasan TNGC
Jenis Mamalia muncak Kijang Musang luwak Kukang jawa budeng Lutung Monyet ekor
panjang Surili Babi hutan r –0,055 0,534** 0,213** 0,517** –0,421** 0,770** Sig. 0,385 0,000 0,001 0,000 0,000 0,000 Kijang muncak r 0,051 –0,231** –0,336** –0,246** 0,101 Sig. 0,418 0,000 0,000 0,000 0,108 Musang luwak r 0,532** –0,039 –0,414** 0,253** Sig. 0,000 0,534 0,000 0,000 Kukang jawa r –0,198** –0,146* –0,099 Sig. 0,002 0,020 0,116 Lutung budeng r –0,161* 0,506** Sig. 0,010 0,000
Monyet ekor panjang r –,294**
Sig. 0,000
Keterangan: **) = korelasi sangat nyata pada α=0,01; *) = korelasi nyata pada α=0,05
Karnivora
Selain jenis-jenis mamalia herbivora juga ditemukan dua jenis mamalia karnivora, yakni macan tutul jawa Panthera
pardus melas kucing hutan atau kucing
kuwuk Prionailurus bengalensis. Macan tutul jawa merupakan satwa karnivora dari suku Felidae. Pakan macan tutul jawa meliputi binatang melata, ketam, serangga, kelelawar, penyu laut, landak, trenggiling, burung merak, ayam hutan, teledu, musang, owa abu-abu, monyet ekor panjang dan anjing (Hoogerwerf 1970, Grzimek 1975, Lekagul & McNeely 1977, Dit. PPA 1982). Meskipun jenis sumber pakan macan tutul beragam, tetapi sebaran macan tutul di kawasan TNGC hanya berhubungan positif nyata dengan sebaran musang luwak.
Kucing hutan merupakan satwa nokturnal dan arboreal yang dapat menempati habitat berupa areal hutan, perkebunan dan kebun masyarakat. Jenis pakan bagi kucing hutan meliputi mamalia kecil dan serangga besar (Payne et al. 2000). Di kawasan TNGC sebaran kucing hutan hanya berhubungan positif dengan musang luwak.
PEMBAHASAN
Sebaran populasi sebagian jenis mamalia herbivora berhubungan sangat nyata dengan sebaran jenis-jenis vegetasi sumber pakan. Berdasarkan koefisien kesamaan komunitas Jaccard maka hampir semua tipe habitat yang diamati dapat dikategorikan memiliki komunitas yang berbeda antar satu tipe habitat dengan tipe habitat lainnya. Koefisien kesamaan komunitas tertinggi terjadi antara tipe habitat hutan pegunungan dengan hutan sub alpin, yakni untuk komunitas tumbuhan bawah sebesar 27,37%, untuk tingkat pertumbuhan semai sebesar 17,94%, tingkat pertumbuhan pancang sebesar 55,65%, tingkat pertumbuhan tiang sebesar 11,55% dan tingkat pertumbuhan pohon sebesar 11,68%.
Kesamaan komunitas tumbuhan yang rendah antar tipe habitat ternyata memberikan pengaruh terhadap perbedaan jenis-jenis mamalia herbivora yang menempati habitat bersangkutan. Mamalia herbivora di TNGC sebagian besar ditemukan di tipe habitat hutan sub alpin, yakni sebanyak 7 jenis. Koefisien kesamaan komunitas Jaccard untuk mamalia herbivora
tertinggi terjadi antara tipe habitat hutan pegunungan dengan hutan sub alpin, yakni sebesar 70,55%.
Salah satu jenis mamalia di kawasan TNGC yang sebarannya hampir tidak bergantung pada sebaran vegetasi dan tipe ekosistem adalah P. hermaphroditus. Hal ini ditunjukan oleh ditemukannya musang luwak pada seluruh tipe ekosistem yang diamati. Meskipun P. hermaphroditus membutuhkan pakan sebagian besar berupa buah dan daun, namun jenis ini hanya berhubungan dengan V. rubescens dan T. sundaica yang menghasilkan buah keras. Selain itu, berdasarkan tipe habitat maka P. hermaphroditus merupakan salah satu jenis mamalia yang dapat hidup pada habitat hutan tinggi, hutan sekunder, perkebunan maupun kebun masyarakat.
Hutan tanaman pinus merupakan tipe ekosistem yang miskin keanekaragaman jenis-jenis vegetasi. Selain mamalia P.
hermaphroditus, di kawasan ini hanya
ditemukan jenis S. scrofa dan M. muntjak. Jenis M. muntjak dapat menempati habitat beberapa tipe vegetasi sepanjang tahun dengan pakan utama berupa buah dan semak, namun lebih banyak menempati habitat daerah bagian atas (Meijaard & Sheil 2008). Jenis-jenis pohon penghasil buah yang menentukan sebaran M. muntjak di TNGC adalah A. squamulosa, F. rukam, P.
arborea, M. pinnata, F. microcarpa, A. procera, dan A. rotundifolium. Selain itu, di
kawasan hutan tanaman juga ditemukan jenis-jenis sumber pakan M. muntjak antara lain Calliandra callothyrsus, Eupatorium
odoratum, Melastoma sylvaticum dan Desmodium heterophyllum.
Keberadaan M. muntjak berhubungan erat dengan N. javanicus, T. auratus dan M.
fascicularis. Hal ini mengindikasikan bahwa M. muntjak memanfaatkan buah-buahan
dari pohon tinggi seperti F. microcarpa yang terjatuh maupun sisa makanan jenis-jenis primata. Selain M. muntjak, keberadaan S.
scrofa juga hubungan sangat erat dengan
keberadaan primata. Namun demikian, jenis pohon penghasil buah yang dimakan oleh S.
scrofa berbeda dengan M. muntjak.
Jenis-jenis pohon yang menentukan keberadaan
S. scrofa adalah Antidesma tetandrum, Ficus sp., G. arborescens, L. sundaicus, L.
sanguinolenta, P. impressinervium, P. calomelanos, P.s neriifolius, Psychotria sp., S. arborea dan Timonius sp. Kedua mamalia
herbivora terestrial ini diduga memanfaatkan sisa makanan primata sebagai salah satu sumber pakannya.
Di habitat hutan dataran rendah hanya ditemukan jenis-jenis S. scrofa, P.
hermaphroditus, M. fascicularis dan T. auratus. Kondisi hutan dataran rendah yang
cenderung terganggu berpengaruh terhadap jenis-jenis mamalia yang menempatinya, yakni pada umumnya mamalia yang menyukai daerah terbuka seperti Macaca fascicularis dan jenis yang memiliki kemampuan adaptasi tinggi terhadap perubahan lingkungan. Berdasarkan kepadatan populasi maka habitat hutan dataran rendah merupakan habitat yang memiliki total kepadatan populasi tertinggi dibanding dengan tipe habitat lainnya, yakni kepadatan M.
fascicularis 3,5321 individu/ha, T. auratus
0,4333 individu/ha, Sus scrofa 0,0667 individu/ha dan P. hermaphroditus 0,1667 individu/ha.
Jenis-jenis mamalia yang ditemukan di habitat hutan pegunungan P. aygula, T.
auratus, S. scrofa, M. muntjak dan P. hermaphroditus. Meskipun memiliki jumlah
jenis yang lebih tinggi dibanding dengan tipe hutan dataran rendah, namun total kepadatan populasi lebih rendah dibanding dengan habitat hutan dataran rendah. Total kepadatan populasi di habitat hutan pegunungan hanya 0,5956 individu/ha, yang terdiri atas P. aygula 0,0838 individu/ha, T. auratus 0,2962 individu/ha,
S. scrofa 0,1305 individu/ha, M. muntjak
0,0850 individu/ha dan P. hermaphroditus 0,0920 individu/ha. Jenis-jenis tumbuhan yang mendominasi pada habitat ini merupakan jenis yang merupakan sumber pakan bagi jenis-jenis mamalia pemakan daun dan buah.
Semua jenis mamalia herbivora yang ditemukan pada penelitian ini dijumpai di habitat hutan sub alpin. Namun demikian, total kepadatan populasi seluruh mamalia herbivora di habitat ini lebih rendah dibanding dengan tipe habitat hutan pegunungan maupun hutan dataran rendah. Kekayaan jenis mamalia herbivora di
habitat hutan sub alpin ini berbanding terbalik dengan kekayaan jenis tumbuhan. Kekayaan jenis tumbuhan di TNGC berangsur-angsur menurun dari tipe habitat hutan dataran rendah, habitat hutan pegunungan hingga habitat hutan sub alpin. Penurunan kekayaan jenis tumbuhan ini sesua dengan hasil penelitian Kitayama (1992) yang menyatakan bahwa kekayaan jenis tumbuhan tertinggi pada kawasan dengan ketinggian sekitar 1400 m dpl dan akan menurun secara perlahan pada ketinggian tempat 2600–2800 mdpl.
KESIMPULAN
Keberadaan musang luwak P. Hermaphroditus tidak dipengaruhi oleh sebaran vegetasi dan tipe ekosistem, sebaliknya kesamaan komunitas tumbuhan yang rendah antar tipe habitat ternyata memberikan pengaruhi terhadap perbedaan jenis-jenis mamalia.
DAFTAR PUSTAKA
Barrette C. 1977. Some aspects of the behaviour of muntjacs in Wilpattu National Park. Mammalia 41:1–34. Belovsky GE and OJ Schmitz. 1994. Plant
defense and optimal foraging by mammalian herbivores. Journal of
Mammalogy 75:816–832.
Chivers DJ. 1980. The siamang in Malaya: A field study of primate in trophical rain forest. Contribution on
Primatology 4:1–135.
Colebrook WF, JL Black, DB Purger, WJ Collins and RC Rossiter. 1990. Factors affecting diet selection by sheep V. Observed and predicted preference ranking for six cultivars of subterranean clover. Australian
Journal of Agricultural Research
41:957–967.
[Dit PPA] Direktorat Perlindungan dan Pelestarian Alam. 1978. Mamalia di Indonesia. Direktorat Perlindungan dan Pelestarian Alam Direktorat Jenderal Kehutanan. Bogor.
Grzimek B. 1975. Animal Life Encyclopedia Vol. 12, Mammal III. London: Van Nostrand Reinhold Company.
Hoogerwerf A. 1970. Ujung Kulon, The Land of The Last Javan Rhinoceros. Leiden: EJ Brill.
Kitayama K. 1992. An altitudinal transect study of the vegetation on Mount Kinabalu, Borneo. Vegetatio 102:149–171.
Kurland J. 1973. A natural historis of kera macaques (Macaca fascicularis) at Kutai Reserve Kalimantan Timur. Indonesia. Primates 14:245–263 Laska M, JML Baltazar and ER Luna. 2003.
Food preferences and nutrinet composition in captive pacas Agouti
paca (Rodentia, Dasyproctidae). Mammalian Biology 68:31–41.
Lekagul B and JA McNeely. 1977. Mammals of Thailand. Bangkok: Kurusapha Ladprao Press.
Lindburg DG. 1980. The Macaques: Studies in ecology, behavior and evolution (Ed). Van Nostrand Reinhold Company. New York. 384p.
Meijaard E and D Sheil. 2008. The persistence and conservation of Borneo’s mammals in lowland rain forests managed for timber: observations, overviews and opportunities. Ecological Restoration 23: 21–34.
Milton K. 1979. Factors influencing leaf choice by howler monkeys: A test of some hypotheses of food selection by generalist herbivores. The American
Naturalist 114:362–378.
Ohsawa M. 1995. Latitudinal comparison of altitudinal changes in forest structure, leaf-type, and species richness in humid monsoon Asia.
Vegetatio 121:3–10.
Payne J, CM Francis, K Phillipps dan SN Kartikasari. 2000. Panduan Lapangan Mamalia di Kalimantan, Sabah, Sarawak & Brunei Darussalam. Bogor: Wildlife Conservation Society Indonesia Program.
Siahaan AD. 2002. Pendugaan parameter demografi populasi surili (Presbytis
aygula Linnaeus 1758) di kawasan
Unocal Geothermal Indonesia, Gunung Salak [Skripsi]. Jurusan Konservasi Sumberdaya Hutan Fakultas Kehutanan IPB. Bogor .
Sugiharto G. 1992. Studi perilaku makan monyet ekor panjang (Macaca
fascicularis) di Pulau Tinjil, Jawa
Barat [Skripsi]. Jurusan Konservasi Sumberdaya Hutan Fakultas Kehutanan IPB. Bogor .
Richards LA and PD Coley. 2007. Seasonal and habitat differences affect the impact of food and predation on herbivores: a comparison between gaps and understory of a tropical forest. Oikos 116: 31–40.
Tolkamp BJ, RJ Dewhurst, NC Friggens, I Kyriazakis, RF Veerkamp and JD
Oldham. 1998. Diet choice by dairy cows 1: Selection of feed protein content during the first half of lactation. Journal of Dairy Science 81:2657–2669.
van Wiesen SE. 1996. Do large herbivores select a diet that maximizes short-term energy rate? Forest Ecological
Management 88:149–156.
Wood GW and DN Roark. 1980. Food habits of feral hogs in Coastal South Carolina. Journal of Wildlife Management 44:506–511.
Lampiran 1. Jenis-jenis tumbuhan di kawasan TNGC
No. Nama Lokal Spesies Famili Formasi
HPn HDR HPg HSA
1 Kileho Saurauia sp. Actinidiaceae √ √ √
2 Kerteu, kereteup Alangium rotundifolium Alangiaceae √
3 Daho Dracontomelon dao (Blanco) Merr Anacardiaceae √
4 Renghas Gluta renghas Anacardiaceae √
5 Limus Mangifera foetida Lour. Anacardiaceae √
6 Mangga pari Mangifera indica L. Anacardiaceae √
7 Lame Alstonia angustiloba Miq. Apocynaceae √
8 Pulai, kame, lame Alstonia scholaris (L) R.Br. Apocynaceae √
9 Jelutung Dyera lowii Apocynaceae √
10 Angrit Kopsia sp Apocynaceae √
11 Kijago Macropanax dispermum Araliaceae √ √
12 Panggang Trevesia sundaica Miq. Araliaceae √ √
13 Huru dapung Combretum sp. Combretaceae √
14 Huru lancang Connarus sp. Cornaceae √
15 Binuang Tetrameles nudiflora Datiscaceae √ √
16 Simpur Dillenia excelsa Dilleniaceae √ √ √ √
17 Huru kadu Elaeagnus latifolius Elaegnaceae √
18 Wuni Antidesma bunius (L.) Spreng. Euphorbiaceae √ √
19 Pereng Antidesma montanum Bl Euphorbiaceae √ √ √
20 Huru koneng, kiseer Antidesma tetandrum Bl. Euphorbiaceae √ √ √ √
21 Kiendog Aporosa sphaeridophora Merr. Euphorbiaceae √
22 Cerem Breynia cernua (Poir.) M.A. Euphorbiaceae √
23 Huru duren Drypetes longifolia Euphorbiaceae √
24 Kisereh Exoecaria virgata Euphorbiaceae √
25 Mareme Glochidion arborescens Euphorbiaceae √
26 Kipare Glochidion rubrum Euphorbiaceae √
27 Mara Macaranga rhicinoides Euphorbiaceae √ √ √
28 Kurai Mallotus paniculatus Euphorbiaceae √ √
29 Kareumbi Omalanthus populneus Euphorbiaceae √ √ √
30 Tarisi Albizia lebbeck Fabaceae √
31 Kaliandra Caliandra caliandra Fabaceae √ √ √
32 Kihiur, huru saninten Castanopsis accuminatissima (Bl.) DC. Fagaceae √ √ √ 33 Saninten, kingkilaban Castanopsis javanica (Bl.) DC. Fagaceae √ √ √ √
34 Pasang Lithocarpus ewyckii Fagaceae √ √ √
35 Pasang dadap/sapu Lithocarpus sundaicus Fagaceae √ √ √
36 Picung Pangium edule Flacouraceae √
37 Rukem Falcourtia rukam Flacourtiaceae √
38 Kibonteng Platea latifolia Bl. Icacinacae √ √
39 Hambirung, kimeri Engelhardia servata Juglandaceae √ √ √
40 Huru piit Actinodaphne macroptera Lauraceae √ √
41 Huru mungkal Actinodaphne procera Nees Lauraceae √
42 Kiteja Cinnamomum burmanii Ness ex Bl. Lauraceae √ √
43 Kiamis Cinnamomum iners Reinw. ex Bl. Lauraceae √
44 Petag hurang, kihurang Cryptocarya densiflora Bl. Lauraceae √
45 Huru kembang Cryptocarya ferrea Bl. Lauraceae √
46 Huru kawoyang Endiandra rubescens (Bl) Miquel Lauraceae √
47 Kiputih Litsea cuceba (Lauv) Pers. Lauraceae √ √
48 Huru kiteja Litsea lanceolata Lauraceae √
49 Huru madam Litsea sanguinolenta Lauraceae √ √ √
50 Huru menceng Machilus rimosa Lauraceae √
51 Kitales Notaphoebe umbelliflora Lauraceae √ √
Lampiran 1. Lanjutan
No. Nama Lokal Spesies Famili Formasi
HPn HDR HPg HSA
53 Huru nangka Persea rimosa (Bl.) Kosterm. Lauraceae √ √
54 Bungur Lagerstroemia speciosa Lythraceae √
55 Kait besi Memecylon excelsum Bl. Melastomataceae √
56 Huru cantigi Aglaia odorata Meliaceae √ √
57 Huru langsep Aglaia squamulosa Meliaceae √ √
58 Kadoya, huru kadoya Dysoxylum alliaceum Meliaceae √ √ √
59 Andewi, sentul Sandoricum koetjape Merr. Meliaceae √ √
60 Lantoro Leucaena glauca Bth. Mimosaceae √
61 Petai-petaian Parkia javanica Merr. Mimosaceae √
62 Benda Artocarpus elasticus Reinw. ex Bl. Moraceae √ √
63 Kacu Artocarpus sp. Moraceae √ √ √
64 Hampelas Ficus ampelas Moraceae √ √
65 Beringin Ficus benjamina L Moraceae √ √
66 Pangsor Ficus callosa Willd. Moraceae √
67 Beunying Ficus fistulosa Moraceae √ √ √
68 Hamberang Ficus grossularoides Burm. f. Moraceae √ √
69 Bisoro Ficus hispida Moraceae √ √
70 Calodas Ficus microcarpa Moraceae √ √
71 Manis mata Ficus montana Burm. f. Moraceae √
72 Walen Ficus ribes Moraceae √ √
73 Kiplik Ficus sp. Moraceae √ √
74 Pulus Ficus sp. Moraceae √ √
75 Kondang Ficus variegata Moraceae √ √
76 Ipis kulit Decaspermum fruticosum Myrtaceae √ √
77 Kibereum Eugenia spicata Lamk. Myrtaceae √
78 Ceremeh Eugenia uniflora Myrtaceae √
79 Kitambaga Syzygium antisepticum (Bl.) Myrtaceae √ √
80 Petag Syzygium densiflorum Myrtaceae √ √ √
81 Putat manggung Syzygium lineatum Myrtaceae √
82 Salam Syzygium polyanthum Walp Myrtaceae √
83 Petag kopo, huru salam Syzygium pycnanthum Myrtaceae √ √
84 Huru jambu Pisonia umbellifera Seem. Nyctaginaceae √
85 Hahapan Flemingia strobilifera R.Br. Papilionaceae √
86 Pinus Pinus merkusii Pinaceae √
87 Kibima, jamuju Podocarpus imbricatus Bl. Podocarpaceae √ √
88 Kiputri Podocarpus neriifolius D. Don Podocarpaceae √
89 Sawuhem Xanthophyllum excelsum Miq. Polygalaceae √
90 Afrika Maesopsis eminii Rhamnaceae √ √
91 Kilaki Zizyphus horsfieldii Rhamnaceae √ √
92 Kawoyang, kitamiang Prunus arborea (Bl.) Kalkm. Rosaceae √ √
93 Kopi Canthium confertum Rubiaceae √ √ √
94 Cangcaratan Psychotria sp. Rubiaceae √ √
95 Huru tangkil Psychotria viridiflora Rubiaceae √ √
96 Huru kopi Saprosma arborea Rubiaceae √
97 Kimeong Timonius sp. Rubiaceae √ √
98 Kigambir Uncaria gambir Roxb. Rubiaceae √
99 Gempol Nauclea excelsa Rubiaceae √
100 Huru meuhmal Acronychia sp. Rutaceae √ √
101 Kisampang Evodia latifolia Rutaceae √ √
102 Kitiwu Miliosma pinnata Sabiaceae √
103 Lingsir Guioa pleuroptesis Sapindaceae √ √
Lampiran 1. Lanjutan
No. Nama Lokal Spesies Famili Formasi
HPn HDR HPg HSA
105 Rerak Mischocarpus pentapetankul sundaicus Sapindaceae √
106 Sawo manggung Casimiroa edulis Sapotaceae √ √
107 Pungpurutan Mimusops elengi Sapotaceae √ √ √
108 Surian Palaquiurn impressinervium Sapotaceae √
109 Canar Smilax leucophylla Smilacaceae √ √ √
110 Bintinu Melochia umbullata Sterculiaceae √ √
111 Carui Pterospermum javanicum Sterculiaceae √ √
112 Hantap Sterculia coccinea Jack Sterculiaceae √
113 Jirag Symplocos fasciculata Zoll. Symplocaceae √ √
114 Huru sawo Symplocos odoratissima Symplocaceae √ √
115 Huru kibeunter Debregeasia sp. Urticaceae √
116 Kibeunter Macutia diversifolia Urticaceae √ √ √
117 Camun Pipturus incanus (BI.) Wedd Urticaceae √
118 Leksa Poikilospermum suaveolens Urticaceae √
119 Gurunggutu Clerodendrum serrature (L.) Spr. Verbenaceae √ √
120 Nangsi, ruyung Villebrunea rubescens Verbenaceae √ √
121 Antret UK UK √ 122 Baboyoran UK UK √ 123 Beti UK UK √ 124 Bosot UK UK √ 125 Canggogo UK UK √ 126 Depak UK UK √ 127 Gintung UK UK √ √ 128 Huru kapundung UK UK √ √ 129 Kutupuk UK UK √ 130 Singadapa UK UK √
131 Nilam Hemigraphis confinis (Nees) T. Ander Acanthaceae √
132 Pecah beling Strobilanthes crispus Acanthaceae √ √ √
133 Poh-pohan Buchanania arborescens (Bl.) Bl. Anacardiaceae √
134 Kicarirang Cladium bicolor Araceae √
135 Talas-talasan Colocasia esculenta (L) Shott Araceae √
136 Talas hutan Homalomena cordata Schott. Araceae √
137 Binbin Areca pumida Arecaceae √ √
138 Kawung Arenga pinnata (Wurmb) Merr. Arecaceae √ √
139 Kawung monyet Arenga sp. Arecaceae √
140 Bubuai Pinanga coronata Arecaceae √ √
141 Kirinyuh Eupatorium odoratum L.f. Asteraccae √ √ √
142 Babadotan Ageratum conyzoides L. Asteraceae √ √ √
143 Kispong Emilia sonchifolia (L.) DC. ex Wight Asteraceae √
144 Begonia Begonia laciniata Roxb Begoniaceae √
145 Paku tihang Cyathea contaminans Ceatheaceae √
146 Seruni Wedelia biflora (L.) DC. Compositae √
147 Paku-pakuan Alsophila glauca J.Sm. Cyatheaceae √ √ √ √
148 Teki Cyperus compactus Cyperaceae √
149 Karas tulang Euphorbia teruccali Euphorbiaceae √ √
150 Singkong Manihot esculenta Crautz Euphorbiaceae √
151 Meniran Phyllanthus urinaria Linn Euphorbiaceae √
152 Orok-orok Clotaria striata Fabaceae √
153 Babakoan Scaevola frutescens Goodenaeaceae √
154 Harendong Melastoma sylvaticum Blume Melastomataceae √ √ √
155 Kisuruh Nyctanthes arboristis L. Oleaceae √
Lampiran 1. Lanjutan
No. Nama Lokal Spesies Famili Formasi
HPn HDR HPg HSA 157 Kacang-kacangan Desmodium heterophyllum D.C. Papilionaceae √
158 Sengserehan Piper aduncumLinn. Piperaceae √
159 Seruh Piper betle Piperaceae √
160 Sereh gunung Piper caducibrateum C.DC Piperaceae √
161 Jukut embun Bothriochloa pertusa Poaceae √
162 Ilat (Ilalang) Imperata cylindrica (L.) Beauv. Poaceae √
163 Rumput gajah Pennisetum purpureum Poaceae √
164 Seta Setaria sphacelata Poaceae √
165 Pakis hatta Acrostichum aureum L. Polypodiaceae √
166 Pakis Pityrogramma calomelanos (L.) Link Polypodiaceae √ √ √ 167 Jukut bau Paederia scandens (Lour.) Merr. Rubiaceae √
168 Cabe hutan Capsicum annuum L. Solanaceae √
169 Hurip cai Pteudoran themum Sterculiaceae √ √
170 Lantanan Centella asiatica (L.) Urb. Umbelliferae √
171 Pulus hutan Laportea stimulans Urticaceae √
172 Tembelekan Lantana camara L. Verbenaceae √
173 Tepus Achasma megalocheilos Griff. Zingiberaceae √ √
174 Laja gowa Alpinia malaccensis Zingiberaceae √
175 Laos Languas sp. Zingiberaceae √
176 Lampuyan Zingiber americans Bl. Zingiberaceae √
177 Bokoran UK UK √ 178 Bunga sagu UK UK √ 179 Bunga talas UK UK √ 180 Jukut galunggung UK UK √ √ 181 Jukut kilaki UK UK √ 182 Jukut rambat UK UK √ √ 183 Kembang kingkong UK UK √ 184 Oyeh UK UK √ 185 Pokpongsueng UK UK √ 186 Rerambatan UK UK √ √ √ 187 Rinai UK UK √ √ 188 Rumput rambat UK UK √ 189 Rumput-rumputan UK UK √