Characterization of the low affinity transport system for NO
3−uptake by
Citrus
roots
M. Cerezo
a, V. Flors
a, F. Legaz
b, Pilar Garcı´a-Agustı´n
a,*
aDepartamento de Ciencias Experimentales,Unidad de Biotecnologı´a Vegetal,ESTCE,Campus Riu Sec,Uni6ersidad Jaume I,
Castellon12071,Spain
bDepartamento de Citricultura,IVIA,Apartado Oficial46113,Moncada,Valencia,Spain
Received 5 June 2000; received in revised form 10 August 2000; accepted 10 August 2000
Abstract
Three-month old citrange Troyer (hybrid ofCitrus sinensis x Poncirus trifoliata) seedlings were grown hydroponically and, after a period of NO3− starvation, plants were transferred to solutions enriched with K15NO3 (96% atoms 15N excess) to measure 15NO
3
−uptake rates as a function of external15NO 3
−concentrations. Two different NO
3
−uptake systems were found. Between 1
and 50 mM15NO 3
−in the uptake solution medium, the uptake rate increased linearly due to the low affinity transport system
(LATS). Nitrate reductase activity showed the same response to external [NO3−], and also appears to be regulated by the rate of
nitrate uptake. Nitrate pre-treatments had a represive effect on NO3−uptake rate measured at 5 or 30 mM external [15NO3−]. The
extent of the inhibition depended on the [NO3−] during the pre-treatment and in the uptake solution. These results suggest that
the LATS ofCitrusseedlings is under feedback control by the N status of the plant. Accordingly, addition of amino acids (Glu, Asp, Asn, Gln) to the uptake solution resulted in a decrease in15NO
3
−uptake rate. However, the inactivation of nitrate reductase
activity after treatment of the seedlings with either 100 or 500mM WO42− did not affect the activity of the LATS. Metabolic
uncouplers, 2,4-DNP and KCN, reduced the uptake rate by 43.3% and 41.4% respectively at 5mM external [15NO 3
−]. However,
these compounds had little effect when 15NO 3
− uptake was assayed at 30 mM external concentration. The ATPase inhibitors
DCCD and DES reduced15NO 3
−uptake by 68.8% – 35.6%, at both external [15NO 3
−]. Nitrate uptake by the LATS declined with
the increase of the solution pH beyond pH 4. The data presented are discussed in the context of the kinetics, energy dependence and regulation of NO3−uptake. © 2000 Elsevier Science Ireland Ltd. All rights reserved.
Keywords:Citrus; H+-ATPase; Inhibitors; LATS; Nitrate uptake
www.elsevier.com/locate/plantsci
1. Introduction
Absorption of nitrate by roots provides the predominant source of N for the growth and yield of most crop species.
In several higher plants it was found that nitrate uptake rate follows a biphasic relationship with external nitrate concentration, suggesting the exis-tence of at least two different uptake systems. A high affinity transport system (HATS) which shows Michaelis – Menten kinetics operates at [NO3−] lower than 1 mM. The HATS seems to be substrate inducible (IHATS) since NO3− influx as well as net uptake rate are considerably enhanced when NO3− is supplied after a period of NO3− starvation. Uninduced plants also exhibited a con-stitutive high affinity system (CHATS) which op-erates concurrently with the IHATS in the induced state. A constitutive low affinity transport system (LATS), which shows linear kinetics, contributes
Abbre6iations: DC, cell membrane electrical potential difference;
DCCD, N,N’-dicyclohexyl-carbodiimide; DES, diethylstilbestrol; DTT, dithiothreitol; EDTA, ethylennediaminetetraacetic acid; FAD, flavin adenine dinucleotide; 2,4-DNP, 2,4-dinitrophenol; HATS or LATS, high-affinity or low-affinity transport systems, respectively; Mes, 2(N-morpholino)-ethanesulfonic acid; NADH, b-nicotinamide adenine dinucleotide, reduced form; NR(A), nitrate reductase (activ-ity); PVPP, polyvinylpyrrolidone K 15; Tris, Tris (hydroxymethyl)-aminomethane.
* Corresponding author. Tel.: +34-96472-8042; fax: + 34-96472-8066.
E-mail address:[email protected] (P. Garcı´a-Agustı´n).
significantly to the uptake rate at external [NO3−] higher than 1 mM. It is known that the linear LATS responds markedly to tissue N status and that its slope is significantly depressed in barley [1] or Spruce plants [2] grown at high compared to low external NO3−. This dual-phase pattern for NO3− uptake has been found in several plant spe-cies, including Hordeum 6ulgare [1,3], Arabidopsis thaliana [4], maize [5], Triticum aesti6um [6],Picea glauca [2] or Citrus [7].
The HATS for NO3− uptake is sensitive to metabolic inhibitors and appears to be an active transport system [1,8 – 10]. The mechanism of en-ergy coupling for active NO3− transport by HATS has been investigated in a limited number of spe-cies through electrophysiological studies [11 – 13]. Nitrate absorption was associated with depolariza-tion of the root cell plasma membrane. The de-crease in the electrical potential difference (DC) observed after NO3−addition, is inducible by NO3− and saturable with respect to exogenous [NO3−] [13]. These observations are consistent with a mechanism for NO3− uptake by the HATS involv-ing a 2H+:1NO
3
− symport moved by the energy
derived from the proton gradient generated by the plasma membrane H+-ATPase [11 – 14].
However, very little information has been re-ported about the characteristics of the LATS for NO3−, especially in fruit trees in which NO3− up-take by this system would play a preponderant role after mineral fertilization, when NO3−levels in soil are raised for a limited period of time.
The purpose of the present study was to charac-terize the LATS for NO3− in Citrus roots. There-fore, net uptake rates were measured using 15N. The regulation of the LATS activity was investi-gated through the effects of NO3− pre-treatments, pH and specific inhibitory substances.
2. Material and methods
2.1. Plant material
Citrange Troyer (Citrus sinensis L. Osbeck x Poncirus trifoliata Blanco) seedlings were grown under glasshouse conditions in 0.5 L pots filled with fine sand. Temperature ranged between 16 – 18°C at night and 25 – 27°C during the day.
Rela-tive humidity was maintained at 80%
approximately. The plants were irrigated twice
weekly until the beginning of the experiments, with the following nutrient solution: 3 mM Ca(NO3)2, 3 mM KNO3, 2 mM MgSO4, 3.4 mM H3PO4 and 17.9 mM Fe-EDTA. Trace elements were present as prescribed by Hoagland and Arnon (1950) [15]. The pH of the nutrient solution was adjusted to 6.0 with 1 M KOH. Three-month old plants growing in a single shoot were selected for uniformity of size at the beginning of the experimental treatments.
The seedlings were then gently removed from the sand and transferred to a 0.2 mM CaSO4 solution, in a controlled environment chamber at 25°C and 60 – 65% RH in the dark. After 6 days, seedlings were transferred to the aerated complete nutrient solution described above lacking nitrogen
(Ca2+ and K+ were supplied as 3 mM
CaSO42H2O and 1.5 mM K2SO4) and were kept in continuous light. The PPFD at the seedlings canopy was 1200 mmol m−2s−1. Continuous light was used to eliminate diurnal variations in the uptake rates. The seedlings were maintained in the nitrogen-free solution for 3 days (control seedlings).
2.2. NO−3 uptake
Nitrate uptake was determined by placing for 24 h, 10 seedlings or 10 detached roots in a 200 cm3 glass flask containing 1 mM MES (pH 6.0), CaSO4 0.2 mM and 1 – 50 mM K15NO
3 (96% atoms 15N excess). In these conditions, the uptake is negligi-ble compared to total nitrate in the solution. Opti-mum nitrate levels (150 – 200 mmol) were maintained to observe a linear response after transferring 10 seedlings to 200 ml containing 1 mM MES (pH 6.0), CaSO40.2 mM and 1 – 50 mM K15NO
3 during 24 h.
The environmental conditions during the la-belling period (24 h) were 27°C, 65% RH and 1200
mmol m−2 s−1 continuous light intensity. Solu-tions were aerated vigorously and water losses by transpiration and evaporation were compensated by frequent additions of water, to maintain con-stant the solution volume.
Nitrogen determinations were made on roots, stems and leaves by a semi-micro Kjeldahl method [16]. For the emission spectrometer deter-mination of 15N, 0.32 N H
2SO4 was added to distillates, to prevent NH4+ evaporation. These solutions were then dried at 70°C for 48 h and the precipitated ammonium salt was re-dissolved to 1 mg nitrogen per 2 ml H2O. An appropriate amount (200 mg nitrogen) of concentrated digest was transferred to Rittenberg tubes. After de-gassing under high vacuum, the ammonia was oxidised to N2 gas with excess sodium hypo-bromite solution. The gas samples were analyzed by emission spectrometry with a JASCO-15 ana-lyzer.
15N enrichment was determined from peaks 28,
29 and 30 [17]. Nitrate uptake was calculated by addition of 15N contents of roots, stems and leaves and was expressed by g dry weigh root h.
2.3. Kinetics of net 15NO 3
− uptake by the low
affinity transport system
The kinetics of net 15NO 3
− uptake by the LATS
as a function of external [NO3−] was either mea-sured in control seedlings (deprived with N for 9 days) either on intact seedlings or in detached roots after transfer to various 15NO
3
− solutions
(1 – 50 mM). Nitrate uptake and nitrogen determi-nations were performed as described above.
2.4. Effect of NO3− pre-treatments
After 6 days in the dark, seedlings were trans-ferred to in the nitrogen-free solution for 2 days and then pre-treated with 3 or 10 mM KNO3 for 1 day (pre-treated seedlings). Nitrate uptake was either measured at 5 or 30 mM K15NO
3 (96% atoms 15N excess) and nitrate uptake and nitro-gen determinations were carried as above. Based on our results of the kinetic studies on Citrus [7] above 1 mM external 14NO
3
−, uptake appeared to
result from the participation of both transport systems (HATS and LATS). Therefore, the satu-rated values of the HATS represent only 16% or 4% when uptake was measured at 5 or 30 mM external [14NO
3
−], respectively. Thus, the measure
uptake at 5 and 30 mM external [15NO 3
−]
were taken to represent uptake rates due to the LATS.
2.5. Nitrate reductase assays
Nitrate reductase activity was measured in roots of seedlings exposed for 24 h at 1 – 50 mM KNO3, as described above for the determinations of kinetics of the LATS. Roots (2 g fr. wt.) were homogenized with pestle and mortar at 0°C in 6 ml 0.1 M Tris– HCl (pH 8.5), 20 mM FAD, 2mM Na2MoO4, 2 mM EDTA, 1 mM DTT, 0.01 mM leupeptin and 2.5% PVPP (w/v). The extracts were centrifuged at 16,000 rpm for 15 min. Sam-ples of 0.2 ml were incubated for 15 min at 27°C in 0.8 ml 50 mM K3PO4, 20 mM KNO3 and 3 mM NADH, pH 7.5. The amount of nitrite formed was measured spectrophotometrically at 540 nm after the addition of 0.5 ml sulphanil-amide and 0.5 ml N-(1-naphthyl)-ethylenediamine dihydrochloride [18]. Soluble protein content of roots (mg of protein per FW from two replicates and two independent samples) was estimated in centrifuged extracts (10 min at 4000g) using the Bradford method [19].
2.6. Effect of WO42− on nitrate reductase acti6ity and nitrate uptake
Nitrate reductase activity was measured in roots of control seedlings transferred to uptake solutions containing 1 mM MES (pH 6.0), CaSO4 0.2 mM, and the either 5 or 30 mM 15KNO
3. Tungstate was added to the nitrate solutions at 100 or 500 mM. The nitrate reductase activity and uptake measurements were carried as above.
2.7. Effect of amino acids on nitrate uptake
The amino acids (Glu, Asp, Asn and Gln) were added to the uptake solution at either 3 or 10 mM concentrations. The nitrate uptake was mea-sured at 5 and 30 mM. The uptake measurements were carried as above.
2.8. Effect of pH on nitrate uptake
The effect of pH on 15NO 3
− uptake was studied
in control seedlings, as described above. Uptake solutions contained 0.2 mM CaSO4, 1 mM MES-TRIS (pH 4, 5, 6, 7, 8 or 9) and either 5 or 30 mM K15NO
2.9. Inhibitor assays
The effect of 2,4-DNP, KCN (metabolic uncou-plers), DCCD and DES (plasmalemma H+
-AT-Pase inhibitors) on 15NO 3
− uptake was studied in
non NO3− pre-treated seedlings. The compounds 2,4-DNP and KCN were added to the uptake solutions at concentrations of 20mM and 0.1 mM respectively. For the plasmalemma H+-ATPase
inhibitors assays, the seedlings were pre-incubated for 1 h with 0.1 mM of either DCCD or DES supplied in the nutrient solution these compounds were also added, at the same concentration, to the uptake solutions. For experiments with DES or DCCD, all solutions (including control solutions) contained 0.25% (v/v) ethanol.
Nitrate uptake was measured at 5 and 30 mM K15NO
3
− (96% atoms 15N excess), as described above.
3. Results
Rates of 15NO 3
− uptake by roots of nitrate
starved seedlings were measured over a wide range of external [15NO
3
−] (1 – 50 mM). Cerezo et al.,
(1997) [7] found that nitrate uptake showed typical Michaelis – Menten curve up to 1 mM [14NO
3
−].
The apparent Km value by the HATS was 276mM and the correspondingVmax value was 0.46mmol/g DW root h. Km and Vmax were calculated from regression equations in the concentration range of 0.04 – 1 mM external [15NO
3
−] [20] (Fig. 1A).
Above 1 mM external [15NO 3
−] the concentration
response was linear, showing a slope which was more pronounced in roots from intact plants than in detached roots, with uptake rate constants of 0.14 and 0.078 mmol/g h mM, respectively (Fig. 1A).
Nevertheless, 15N concentration was similar in roots from intact plants than in detached roots (Fig. 1B), indicating that the differences in 15NO
3
−
uptake between both were due to the fraction of 15N transported to shoots.
To establish a possible relationship between ni-trate uptake and reduction, the activity of nini-trate reductase in roots was determined after exposure of the seedlings to different external [NO3−]. The enzyme activity increased linearly with a slope of 0.0369mmol NO2−/mg protein h as nitrate levels in the external solution were raised from 1 to 50 mM in intact plants (Fig. 1C).
Fig. 1. Effects of increasing [15NO 3
−] in the uptake solution on
net15NO 3
−uptake rates by roots (A: seedlings;detached
roots); 15N accumulation in roots (B: roots from intact
Table 1
Effect of nitrate pre-treatments on net15NO 3
−uptake rate (mmol/g DW root h) byCitrusroots on both HATS and LATS
NO3−pre-treatment LATS
5 mM 15NO
Control 1.5990.04a
E79.3 E2.0990.15a
E0.339 0.02a E58.2
3 mM KNO3
E100
10 mM KNO3 E0.009 0.00a E0.9390.04a E81.4
aSignificant at 5% level respect to the control. NS not significant. The results are the means (9S.D.) of three replicate
experiments.
Table 1 shows the effect of pre-treatments with NO3− solutions (3 or 10 mM) on NO3− uptake measured at 5 and 30 mM external [15NO
3
−].
Pre-treatments on HATS are not tested, but uptake rate of HATS represents 16% of LATS at 5 mM and 4% at 30 mM external [14NO
3
−], respectively;
so any reduction or not on uptake rate by HATS are a little effect on LATS. Furthermore, the strong decrease in absorption by LATS after treat-ments can not be explained by the sole contribu-tion of HATS variacontribu-tion. Thus, uptake rate at 5 and 30 mM external [15NO
3
−], are considered to be
the combined contributions of both HATS and LATS. A previous exposure to NO3− inhibited net 15NO
3
− uptake rate and this reduction was more
pronounced when the concentration of NO3− in the pre-treatment was increased. For example, the uptake rate at 5 mM of external [15NO
3
−] was
reduced by 69% and 93.3% by the pre-treatments with 3 and 10 mM KNO3, respectively. Also, the inhibition seemed to be dependent on [15NO
3
−] in
the uptake solution, being lower at 30 mM than at 5 mM of external [15NO
3
−]. Thus, the inhibition
due to the 10 mM KNO3 pre-treatment decreased from 93.3% to 78.5% when [15NO
3
−] in the uptake
solution was increased from 5 to 30 mM.
Table 2 shows the effect of WO42− on nitrate uptake and activity nitrate reductase. Nitrate up-take rate either 5 or 30 mM [15NO
3
−] was not
affected by WO42−. However, the nitrate reductase activity was inhibited with 100 or 500 mM WO42− after transfer of the seedlings on 5 or 30 mM external [NO3−]. Nitrate reductase activity was in-hibited by 50 – 83% when plants were treated with 500 mM WO42− and transferred to 5 or 30 mM, respectively. To determinate the effect of amino acids on the nitrate uptake rate, different amino acids such as Glu, Asn, Asp, Gln were added to
the uptake solutions containing either 5 or 30 mM [15NO
3
−] (Table 3). The amino acids Glu, Asn,
Asp, Gln at 3mM reduced the uptake rate at 5 mM external concentration. The inhibitory effect of amino acids ranged from 23 to 73%. Similar results were obtained at higher concentrations of amino acids (10 mM) and NO3− in the uptake solution (30 mM) (Table 3).
Net nitrate uptake by roots displayed a marked dependence upon the pH of external solution (Fig. 2). Uptake was drastically inhibited as the solution pH raised from pH 4 to pH 7. The pH-dependence of nitrate uptake showed the same pattern at 5 and 30 mM external [15NO
3
−] (Fig. 2) as well as 0.2
mM external [14NO 3
−] (data not shown). Values
obtained at 0.2 mM external [14NO 3
−] indicate that
HATS contribution to total absorption is not sig-nificant and that the pattern obtained at 5 and 30 mM [15NO
3
−] is representative of LATS behaviour
upon pH variation.
Assays with the same inhibitors used in this work were tested when the external [14NO
3
−] was
0.2 mM (HATS). In all cases, a reduction on 14NO
3
− uptake was observed between 50 and 80%
(data not show), so contribution from HATS to reduction on HATS + LATS (5 or 30 mM exter-nal [15NO
3
−]) are insignificant and we can take
these concentrations as representative for uptake rates due to the LATS only. Metabolic uncouplers such as 2,4-DNP or KCN reduced nitrate uptake by 43.4% and 41.4% respectively at 5 mM external 15NO
3
−, but had negligible effects at 30 mM
exter-nal [15NO 3
−]. Treatment of roots with either
DCCD or DES, which have been shown to inhibit the plasmalemma H+-ATPase, prevented nitrate
absorption on both [15NO 3
−], with an inhibition
4. Discussion
In a previous report, Cerezo et al. (1997) [7] reported the presence of both systems (HATS and LATS) in Citrus roots, in a narrow range of external NO3− concentrations (0.02 – 10 mM), and only with depletion measurements for NO3− up-take rate. The results presented here confirm that the LATS activity remained linear over a wider range of external [NO3−] (1 – 50 mM), and provide much more relaible measurements based on 15N labelling. Also inCitrus roots, a LATS for nitrate uptake operates at elevated external [NO3−], showing a linear kinetics. Similarly, a LATS for NO3− uptake with linear kinetics up to 50 mM was reported by Siddiqi et al. (1990) [1] in barley roots. The LATS was also found in other species, such as Arabidopsis, maize or spruce [21 – 25].
Nitrate uptake rate was significantly depressed in plants pre-treated with NO3− solutions com-pared with N-starved plants. This observation agree with previous reports indicating that N-starvation results in an accelerated net uptake of nitrate when the supply is restored [21 – 25].
It is therefore reasonable to deduce that nitrate transport could be affected by a product of ni-trate assimilation or by nini-trate itself. Moreover, it is known that NO3− absorption can be pro-foundly influenced by amino acids [26 – 28]. Also, [NO3−] inside the root cell determines in part the concentration gradient across the plasmalemma, and therefore could affect the rate of NO3− up-take.
The levels of nitrate and amino acids in the roots will depend on a balance between NO3− uptake, amino acid synthesis from incoming inor-ganic N, amino acid import via the phloem and
export of NO3− and amino acids via xylem. Cy-cling of amino acids between shoots and roots has been considered as a possible mechanism to regulate NO3− absorption and a way to transmit the information on the nitrogen status of the shoot into the roots [29,30]. The mechanism for the regulation of NO3− uptake by the ‘‘N de-mand’’ is still unknown, but appears to be related with feedback effects on NO3− influx, caused by NO3− itself or by amino acids [27,31 – 34]. On the other hand, translocation of NO3− and amino acids from root to shoot appears to be a means to reduce the possible inhibitory effects of these compounds on NO3− uptake by roots.
In agreement with this, the data of Fig. 1B and Fig. 1C indicate that, in short term experiments with N-starved plants, the export of some part of N from roots to shoots is necessary to maintain a high rate of NO3− uptake by the LATS in Citrus roots. Since it has been reported [35 – 37] that NO3− and amino acids are the main N-com-pounds present in the xylem sap of Citrus, it is reasonable to consider that these are involved in the regulation of the LATS. Accordingly, an inhi-bition of nitrate uptake occurred when amino acids were added in the uptake solution. This would favour the hypothesis that the NO3− assim-ilation products play a major role on the repres-sion of the LATS activity. In support to this hypothesis, it was reported that in transgenic Nicotiana plants constitutively overexpressing NR, the NO3− reduction efficiency was offset by a decline in NO3− uptake. This appeared to be the result of a negative feedback regulation of NO3− influx, and was interpreted as repression of NO3− uptake by amino acids to prevent excessive or-ganic N synthesis [38].
Table 2
Effect of WO42−concentration on nitrate reductase activity at 5 and 30 mM external [15NO3−]
LATS
5 mM 15NO 3
− 30 mM 15NO
3
−
Nitrate uptake NRAa Nitrate uptake NRA
5.6090.23 4.969 0.32 Control 2.4290.15 1.5590.11
5.669 0.33 NS 1.3090.11b
1.4690.12 NS 1.4090.12b
100mM WO42−
1.2090.12b 1.5490.13 NS 0.9090.12b 5.389 0.23 NS
500mM WO42−
aNRA is expresed asmmol NO 2
−/mg protein h. Nitrate uptake rate is expressed asmmol/g DW root h.
bSignificant at 5% level respect to the control. NS not significant. The results are the means (9S.D.) of three replicate
Table 3
Effect of amino acids on net15NO 3
−uptake rate at 5 and 30 mM of external concentrations [15NO 3
−]a
5 mM 15NO 3
− % inhibition 30 mM 15NO 3
− % inhibition
1.2090.22
Control 4.8090.67
Glu 3 mM 0.5190.11b E57 2.3890.25b E50
0.4990.03b E59
Glu 10 mM 1.8090.13b E62
Asn 3 mM 0.3390.03b E73 2.0090.11b E58
Asn 10 mM 0.4390.05b E64 2.3090.22b E52 0.5590.02b E54
Asp 3 mM 2.2090.43b E54
Asp 10 mM 0.4690.06b E61 1.7590.35b E63
0.9390.10b E23
Gln 3 mM 2.1590.76b E55
0.2590.01b E79 1.1590.09b E76
Gln 10 mM
aUptake rate at 5 and 30 mM external [15NO 3
−] are considered to be the combined contributions of both HATS and LATS.
Effect of amino acids on HATS are not tested, but uptake rate of HATS represents 16% of LATS at 5 mM and 4% at 30 mM external [15NO
3
−], respectively; so any reduction or not on uptake rate by HATS (E) are a little effect on LATS. Uptake rate is
expresed asmmol/g DW root h.
bSignificant at 5% level respect to the control. The results are the means (9S.D.) of three replicate experiments.
This supports the ‘‘N demand’’ theory, consid-ering that over short periods of pre-treatment, growth of Citrus plants was not significant, and therefore a great fraction of the N compounds would be retained in roots (Fig. 1).
Also, the inhibition of NO3− uptake rate by NO3−pre-treatments was lower when the [NO3−] in the uptake solution was high (Table 2). This would seem to indicate that the [NO3−] gradient across the plasma membrane may have an influ-ence on the net NO3− uptake by the LATS. Alter-natively, this could result from the occurrence of two separate transport system with different affini-ties and specific regulations. In this way two CHL 1 (AtNRT1) homologs identified from tomato, LeNRT 1-2 and LeNRT 1-1, are nitrate-induced and constitutively expresed, respectively [39].
Reduction of NO3− to NO2−, catalyzed by ni-trate reductase (NR), is considered the biochemi-cal rate-limiting step in the NO3− assimilation pathway [40]. Thus, NR could play an indirect role in the absorption of nitrate, regulating the levels of NO3− and amino acids in root cells. Nitrate reductase is induced by nitrate [41,42] and it is believed that the rate of nitrate influx to the site of induction is the main controlling factor for the levels of NR activity [41,43,44]. Our results confirm that in Citrus roots the pattern of induc-tion of nitrate reductase was coincident with the pattern of nitrate uptake by the LATS as a func-tion of external nitrate concentrafunc-tions. However, when the nitrate reductase activity was inhibited
by WO4
2− supply, the nitrate uptake was not
affected. This observation had been made already in NR deficient plants. In Tobacco genotypes un-derexpressing NR, net uptake of NO3−was slightly decreased or remained similar to that in the wild type control [38]. Similar effects were observed with NR-deficient mutants of Arabidopsis thaliana [4,45] or barley [46 – 48]. Nitrate concentration was higher in low-NR genotypes than in wild types, indicating that NO3− itself may be involved in the control of the absorption, and its accumulation would cause a repression of NO3− influx or a stimulation of NO3− efflux [38]. This indicates that there is no direct correlation between nitrate up-take and nitrate reductase, although nitrate reduc-tase activity could be implied in regulating the nitrate uptake system through synthesis of amino acids.
trans-porting NO3− against its electrochemical potential gradient. These authors also presented evidence indicating that the LATS for NO3− uptake is probably mediated by an electrogenic proton co-transport system.
The linear high concentration system for NO3− uptake in Citrus roots appeared to be sensitive to inhibitors of H+ translocating ATP-ases (DCCD
and DES). Also, at moderate [NO3−] (near 5 mM) in the uptake solution, the presence of metabolic uncouplers (KCN or 2,4-DNP) de-creased nitrate absorption.
These results indicate that the LATS can act as an active transport system, linked to the H+
-ATPase. Probably, at relatively low external
[NO3−], the transport of this ion from the solu-tion to the cytoplasm of root cortex cells is ther-modynamically disfavoured in terms of both electrical potential gradient – (negative inside the cell) – and chemical potential gradient, since in-ternal nitrate concentration in root cells usually shows values of over 5 mM [32,49]. It is there-fore clear that in these conditions NO3− uptake requires energy, presumably as a proton driving force.
The results support a model for nitrate uptake by the LATS, which includes a NO3−/H+ sym-port coupled to an ATP dependent proton extru-sion pump which generates the H+ gradient
required for NO3− transport.
However, when nitrate concentration in the culture medium was higher (30 mM), the activity of the LATS was not affected by KCN or 2,4-DNP. These results suggest that at very elevated external [NO3−], nitrate influx in the cell is prob-ably moved by a favourable concentration gradi-ent, without requiring energy from ATP. Also, the hypothesis mentioned previously of the exis-tence of at least two different systems for the LATS, can also explain these differences at 5 and 30 mM.
We observed that net nitrate uptake on Citrus seedling roots was progressively inhibited by in-creases in pH from 4 to 9. At pH 7 and above, inhibition of nitrate uptake was very pro-nounced. These observations are in agreement with previous reports of acidic pH optima for nitrate uptake by the HATS [9,12,50,51].
According to the proposed NO3−/H+ symport mechanism for the LATS, a high H+
concentra-tion in the external medium would generate the proton electrochemical gradient across the plas-malemma (membrane) and, therefore, the driving force for NO3− uptake.
In conclusion, the LATS for NO3− uptake by Citrus roots shows a linear kinetics over a wide range of external [NO3−], is H+ -ATPase-depen-dent and, at least at moderate NO3− levels in the external solution (5mM), requires metabolic en-ergy to surpass the NO3− electrochemical poten-tial gradient and requires an acidic pH to reach maximum activity. In addition, the LATS at high NO3− levels seems to be regulated by amino acids synthesis and a favourable nitrate concentration gradient.
Fig. 2. Effect of uptake solution pH on net 15NO 3
− uptake
rate byCitrusroots at 5 mM (A) and 30 mM (B) of external [15NO
3
−]. The data are the means (9S.D.) of 3 replicate
Table 4
Reduction of net uptake15NO 3
−inCitrusroots by various inhibitors
LATS
5 mM 15NO 3
− % reduction
Treatment 30 mM 15NO
3
− % reduction
1.7590.11
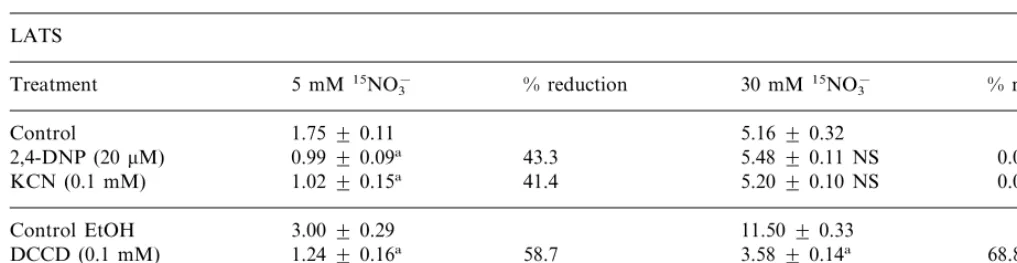
Control 5.1690.32
43.3 0.9990.09a
2,4-DNP (20mM) 5.4890.11 NS 0.0 1.0290.15a 41.4 5.2090.10 NS 0.0
KCN (0.1 mM)
11.5090.33 Control EtOH 3.0090.29
DCCD (0.1 mM) 1.2490.16a 58.7 3.5890.14a 68.8 49.3
1.5290.12a 7.4090.12a 35.6
DES (0.1 mM)
aSignificant at 5% level respect to the control; NS Not significant. The results are the means (9S.D.) of three replicate
experiments.
Acknowledgements
Research was supported by The Consellerı´a de Cultura, Educacio´n y Ciencia (Generalitat Valen-ciana) Grant no. GV-3240/97 and Plan de Promo-cio´n de la InvestigaPromo-cio´n Fundacio´ Caixa Castello´ 1997 P1B97-09. We thank Dr. Alain Gojon of Biochimie et Physiologie Mole´culaire des Plantes AGRO-M/CNRS-UMR 5004/INRA/ UM2-Mont-pellier (France) for their critical review of the manuscript.
References
[1] M.Y. Siddiqi, A.D.M. Glass, T.J. Ruth, T.W. Rufty, Jr., Studies of the uptake of nitrate in barley. Kinetics of
13NO 3
−influx, Plant Physiol. 93 (1990) 1426 – 1432.
[2] H.J. Kronzuker, M.Y. Siddiqi, A.D.M. Glass, Kinetics of NO3− influx in spruce, Plant Physiol. 109 (1995)
319 – 326.
[3] M. Aslam, R.L. Travis, R.C. Huffaker, Comparative kinetics and reciprocal inhibition of nitrate and nitrite uptake in roots of uninduced and induced barley (Hordeum6ulgareL) seedlings, Plant Physiol. 99 (1992) 1124 – 1133.
[4] H. Doddema, G.P. Telkamp, Uptake of nitrate by mu-tants of Arabidopsis thaliana, disturbed in uptake by reduction of nitrate. II. Kinetics, Physiologia Plantarum 45 (1979) 332 – 338.
[5] G.M. Pace, P.R. McClure, Comparison of nitrate up-take kinetic parameters across maize inbred lines, J. Plant Nutrition 9 (1986) 1095 – 1111.
[6] S.S. Goyal, R.C. Huffaker, The uptake of NO3−, NO2−
and NH4+by intact wheat (Triticum aesti6um) seedlings,
Plant Physiol. 82 (1986) 1051 – 1056.
[7] M. Cerezo, P. Garcia-Agustin, M.D. Serna, E. Primo-Millo, Kinetics of nitrate uptake by Citrus seedlings and inhibitory effects of salinity, Plant Science 126 (1997) 105 – 112.
[8] W.A. Jackson, D. Flesher, R.H. Hageman, Nitrate up-take by dark-grown corn seedlings. Some characteristics of apparent induction, Plant Physiol. 51 (1973) 120 – 127. [9] K.P. Rao, D.W. Rains, Nitrate absorption by barley. I. Kinetics and energetics, Plant Physiol. 57 (1976) 55 – 58. [10] A.D.M. Glass, M.Y. Siddiqi, T.J. Ruth, T.W. Rufty, Studies on the uptake of nitrate in barley. II. Energetics, Plant Physiol. 93 (1990) 1585 – 1589.
[11] W.R. Ulrich, A. Novacky, Nitrate-dependent membrane potential changes and their induction in lemna gibba, Plant Science Letters 22 (1981) 211 – 217.
[12] P.R. McClure, L.V. Kochian, R.M. Spanswick, J.E. Shaff, Evidence for cotransport of nitrate and protons in maize roots. Measurement on NO3−and H+fluxes with
ion-selective microelectrodes, Plant Physiol. 93 (1990) 290 – 294.
[13] A.D.M. Glass, J.E. Shaff, L.V. Kochian, Studies on the uptake of nitrate in barley. IV. Electrophysiology, Plant Physiol. 99 (1992) 456 – 463.
[14] S. Santi, G. Locci, R. Pinton, S. Cesco, Z. Varanini, Plasma membrane H+-ATPase in maize roots induced
for NO3−uptake, Plant Physiol. 109 (1995) 1277 – 1283.
[15] D.R. Hoagland, D.J. Arnon, The water culture method for growing plants without soil, California Agricultural Experimental Station Cir. 347 (1950) 1 – 32.
[16] J.M. Bremmer, Total nitrogen, in: C.A. Black (Ed.), Methods of soil analysis, part 2. An. Soc. Agrom., Madison, Wisconsin, 1965, pp. 1149 – 1178.
[17] R.D. Hauck, J.M. Bremmer, Use of tracers for soil and fertilizer nitrogen research, Adv. Agron. 26 (1976) 219 – 266.
[18] P.A. Meyerhoff, P. Garcia-Agustı´n, R.C. Huffaker, Pre-cipitation of barley soluble nitrate reductase, Supple-ment to Plant Physiol. 105 (1994) 40.
[19] M. Bradford, A rapid and sensitive method for the quantitation of microgram quantities of proteins utiliz-ing the principal of protein-dye bindutiliz-ing, Anal. Biochem-istry 72 (1976) 249 – 254.
[20] H. Segel, Behavior and analysis of rapid equilibrium and steady state enzyme systems, in: Enzyme kinetics, Wiley, New York, 1975, pp. 100 – 160.
[22] R.B. Lee, M.C. Drew, Nitrogen-13 studies of nitrate fluxes in barley roots. II. Effect on plant N-status on the kinetic parameters of nitrate influx, J. Exp. Bot. 37 (1986) 1768 – 1779.
[23] P. Laine´, A. Ourry, J. Macduff, J. Boucaud, J. Salette, Kinetic parameters of nitrate uptake by different catch crop species. Effects of low temperatures or previous nitrate starvation, Physiol. Plant 88 (1993) 85 – 92. [24] H. Hasegawa, M. Ichii, Variation in Michaelis-Menten
kinetic parameters for nitrate uptake by the young seedlings in rice (Oryza sati6aL.), Breed. Sci. 44 (1994) 383 – 386.
[25] R.B. Lee, K.A. Rudge, Effects of nitrogen deficiency on the absorption of nitrate and ammonium by barley plants, Ann. Bot. 57 (1986) 471 – 486.
[26] H. Breteler, P.A. Arnozis, Effect of amino compounds on nitrate utilization by roots of dwarf bean, Phyto-chemistry 24 (1985) 653 – 657.
[27] R.B. Lee, J.V. Purves, R.G. Ratcliffe, L.R. Saker, Nitro-gen assimilation and the control of ammonium and nitrate absorption by maize roots, J. Exp. Bot. 43 (1992) 1385 – 1396.
[28] A. Oaks, I. Stulen, I. Boesel, Influence of amino acids and ammonium on nitrate reduction in corn seedlings, Canadian J. Bot. 57 (1977) 1824 – 1829.
[29] H.D. Cooper, D.T. Clarkson, Cycling of amino-nitrogen and other nutrients between shoots and roots in cereals: a possible mechanism integrating shoot and root in the regulation of nutrient uptake, J. Exp. Bot. 40 (1989) 753 – 762.
[30] C.M. Larsson, M. Larsson, J.V. Purves, D.T. Clarkson, Translocation and cycling through roots of recently absorbed nitrogen and sulphur in wheat (Triticum aes
-ti6um) during vegetative and generative growth, Physi-ologia Plantarum 82 (1991) 345 – 352.
[31] D.T. Clarkson, U. Lu¨ttge, II. Mineral nutrition: in-ducible and repressible nutrient transport systems, Prog. Bot. 52 (1991) 61 – 83.
[32] B.J. King, M.Y. Siddiqi, A.D.M. Glass, Studies of the uptake of nitrate in barley. V. Estimation of root cyto-plasmic nitrate concentration using nitrate reductase ac-tivity – Implications for nitrate influx, Plant Physiol. 99 (1992) 1582 – 1589.
[33] T.W. Rufty, D.W. Israel, R.J. Volk, J. Qiu, T. Sa, Phosphate regulation of nitrate assimilation in soybean, J. Exp. Bot. 44 (1993) 879 – 891.
[34] J. Insande, B. Touraine, N demand and the regulation of nitrate uptake, Plant Physiol. 105 (1994) 3 – 7.
[35] T. Kato, Major nitrogen compounds transported in xylem vessels from roots to top in citrus tree, Physiolo-gia Plantarum 52 (1981) 275 – 279.
[36] J. Moreno, J.L. Garcı´a-Martı´nez, Extraction of tracheal sap from citrus and analysis of its nitrogenous com-pounds, Physiologia Plantarum 50 (1980) 298 – 303. [37] J. Moreno, J.L. Garcı´a-Martı´nez, Seasonal variation of
nitrogenous compounds in the xylem sap of citrus, Phys-iologia Plantarum 59 (1983) 669 – 675.
[38] A. Gojon, L. Dapoigny, L. Lejay, P. Tillard, T.W.
Rufty, Effects of genetic modification of nitrate reduc-tase expression on15NO
3
−uptake and reduction in
Nico-tiana plants, Plant Cell and Environonment 21 (1998) 43 – 53.
[39] F.R. Lauter, O. Ninnemann, M. Bucher, J.W. Ries-meier, W.B. Frommer, Preferential expression of an ammonium transporter and two putative nitrate trans-porters in root hairs of tomato, Proc. Natl. Acad. Sci. USA 93 (1996) 8139 – 8144.
[40] L. Beevers, R.H. Hageman, Nitrate and nitrite reduc-tion, in: B.J. Miflin (Ed.), The Biochemistry of Plants, Amino Acids and Derivatives, vol. 5, Academic Press, New York, 1980, pp. 115 – 168.
[41] L. Beevers, L.E. Schraeder, D. Flesher, R.H. Hageman, The role of light and nitrate in the induction of nitrate reductase in radish cotyledons and maize seedlings, Plant Physiol. 40 (1965) 691 – 698.
[42] R.H. Hageman, D. Flesher, Nitrate reductase activity in corn seedlings as affected by light and nitrate content of nutrient media, Plant Physiol. 35 (1960) 700 – 708. [43] G.B. Meeker, A.C. Purvis, C.A. Neyra, R.H. Hageman,
Uptake and accumulation of nitrate as a major factor in the regulation of nitrate reductase activity in corn (Zea mays L.) leaves: effects of high ambient CO2 and malate, in: R.L. Bieleski, A.R. Ferguson, M.M. Creswell (Eds.), Mechanisms of Regulation of Plant Growth, Royal Society Wellington, New Zealand, 1974, pp. 40 – 58.
[44] C.A. Neyra, R.H. Hageman, Nitrate uptake and induc-tion of nitrate reductase in excised corn roots, Plant Physiol. 56 (1975) 692 – 695.
[45] L. Lejay, P. Tillard, M. Lepetit, F.D. Olive, S. Filleur, F. Daniel-Vedele, A. Gojon, Molecular and functional regulation of two NO3− uptake systems by N- and
C-status of Arabidopsis plants, The Plant J. 18 (1999) 1 – 11.
[46] R.L. Warner, R.C. Huffaker, Nitrate transport is inde-pendent of NADH and NAD(P)H nitrate reductases in barley seedlings, Plant Physiol. 91 (1989) 947 – 953. [47] B.J. King, M.Y. Siddiqi, T. Ruth, R.L. Warner, A.D.M.
Glass, Feedback regulation of nitrate influx in barley roots by nitrate, nitrite, and ammonium, Plant Physiol. 102 (1993) 1279 – 1286.
[48] J. Wieneke, Nitrate (13NO 3
−) flux studies and response to
tungstate treatments in wild-type barley and in an NR-deficient mutant, J. Plant Nutrition 17 (1994) 127 – 146. [49] R.G. Zhen, H.W. Koyro, R.A. Leigh, A.D. Tomos, A.J. Miller, Compartmental nitrate concentrations in barley root cells measured with nitrate-selective microelectrodes and by single-cell sap sampling, Planta 185 (1991) 356 – 361.
[50] P.R. McClure, T.E. Omholt, G.M. Pace, Anion uptake in maize roots: Interactions between chlorate and ni-trate, Physiologia Plantarum 68 (1986) 107 – 112. [51] E. Agu¨era, P. De la Haba, A.G. Fontes, J.M.
Maldon-ado, Nitrate and nitrite uptake and reduction by intact sunflower plants, Planta 182 (1990) 149 – 154.
![Fig. 1. Effects of increasing [15roots);seedlings;activity (C) in roots ofNO−3] in the uptake solution onnet 15NO−3 uptake rates by roots (A: � seedlings; � detached 15N accumulation in roots (B: � roots from intact � detached roots) and levels of nitrate](https://thumb-ap.123doks.com/thumbv2/123dok/1031995.928273/4.612.311.511.68.637/increasing-seedlings-activity-solution-seedlings-detached-accumulation-detached.webp)



![Fig. 2. Effect of uptake solution pH on net [rate by15NO−3 uptake Citrus roots at 5 mM (A) and 30 mM (B) of external15NO−3]](https://thumb-ap.123doks.com/thumbv2/123dok/1031995.928273/8.612.46.249.254.653/fig-effect-uptake-solution-uptake-citrus-roots-external.webp)