Summary Net photosynthesis and stomatal conductance were measured in ten red spruce (Picea rubens Sarg.) saplings, growing near Ithaca, New York, throughout the early spring and late-fall growing periods. Gas exchange and daily minimum and maximum soil and air temperatures were also measured. Linear regression analysis showed that rates of net photosyn-thesis were positively correlated with both minimum daily soil and air temperatures but that minimum soil temperature was a better predictor of net photosynthesis. Moreover, net photosyn-thesis was more sensitive to changes in soil temperature than to changes in air temperature, and photosynthesis was approxi-mately twice as sensitive to temperature changes during the fall than during the spring.
Keywords: air temperature, climatic change, gas exchange, seasonal effects, temperature sensitivity.
Introduction
Evergreen coniferous trees growing in temperate climates be-come frost hardened and dormant during part of the winter (Levitt 1980, Kozlowski et al. 1991, Havranek and Tranquillini 1995). While dormant, they may show an almost total cessa-tion of gas exchange (but see Schaberg et al. 1995). However, the evergreen habit affords the potential for year-round net photosynthesis. Furthermore, climatic change resulting in warmer winters may permit temperate conifers to continue photosynthesizing later in the fall and to begin photosynthesiz-ing earlier in the sprphotosynthesiz-ing, thereby extendphotosynthesiz-ing their growphotosynthesiz-ing season. In the northeastern United States and eastern Canada, red spruce (Picea rubens Sarg.) is an important evergreen coniferous species in many montane and low-elevation forests (Blum 1990). The present study was undertaken to assess the possible influence of climatic warming on winter photosynthe-sis in red spruce by documenting the relationships between photosynthetic rates and soil and air temperatures during the spring and fall.
Previous studies of conifers native to western North America have demonstrated significant reductions in gas exchange rates on days following nights with near-freezing or sub-freezing air temperatures (e.g., Fahey 1979, Kaufmann 1982, Smith et al. 1984), and other field and laboratory studies have shown that
low air temperatures can significantly limit gas exchange in conifers (e.g., Smith 1985, DeLucia 1987, DeLucia and Smith 1987, Carter et al. 1987). In addition, several studies have demonstrated the role of low soil temperatures in limiting gas exchange in conifers (DeLucia 1986, DeLucia and Smith 1987, Carter et al. 1987, Day et al. 1989, 1990, 1991, Lippu and Puttonen 1991).
Although recent observations of gas exchange in red spruce at low temperatures have been conducted in conjunction with studies of the species’ cold tolerance and susceptibility to winter injury (Fincher et al. 1989, DeHayes 1992, Fincher and Alscher 1992, L’Hirondelle et al. 1992, Amundson et al. 1993, Eamus 1993, Hadley et al. 1993), no systematic examination of the relative importance of air and soil temperatures in influencing photosynthesis of red spruce during spring and fall has been reported. Moreover, most studies of conifer gas ex-change have focused on a single period during the growing season. In this study, we examined both the beginning and the end of the growing season. We hypothesized that gas exchange responses of red spruce to temperature differs between the fall and spring growing periods. In fall, air and soil temperatures decline in tandem, and subsequently trees undergo winter hardening. In spring, the presence of a snowpack may effec-tively decouple air and soil temperatures resulting in a more complex suite of cues leading to springtime recovery of gas exchange. The nature of these different temperature responses will be important in determining the effects of climatic change on a species’ performance.
Materials and methods
Site description and plant material
In April 1992, ten 7-year-old red spruce saplings were ran-domly selected from an even-aged plantation growing near Cornell University, Ithaca, New York. The stand was estab-lished in 1987 with 2-year old, greenhouse-grown seedlings from a South Range, Nova Scotia provenance collected in 1985 (Alscher et al. 1989). The plantation contained approxi-mately 100 saplings planted in three rows in an east--west orientation. Rows were approximately 2.1 m apart, and
spac-Seasonal air and soil temperature effects on photosynthesis in red
spruce (Picea rubens) saplings
PAUL A. SCHWARZ,
1TIMOTHY J. FAHEY
1and TODD E. DAWSON
21
Department of Natural Resources, Fernow Hall, Cornell University, Ithaca, New York 14853, USA 2 Section of Ecology and Systematics, Corson Hall, Cornell University, Ithaca, New York 14853, USA
Received January 23, 1996
ing within rows was approximately 0.9 m. Saplings were between one and two meters tall.
Temperature measurements
For the first two sampling periods of the study (spring and fall 1992), calibrated thermistors were installed at four depths in the soil profile (1, 5, 10, and 20 cm) beneath the canopy of three of the saplings. Soil temperature measurements were made between 0800 and 0900 h on each day of gas exchange sampling, and it was assumed that these measurements ap-proximated the minimum overnight soil temperatures. Mini-mum and maxiMini-mum air temperatures for the preceding 24-h period were also recorded each morning with a Taylor min--max thermometer located in a weather shed at the site.
During the second year of the study (spring and fall 1993), a Campbell CR10 data logger (Campbell Scientific, Inc., Logan, UT) was installed to automate the collection of air and soil temperatures. Copper-constantan thermocouples were in-stalled at soil depths of 1, 5, and 10 cm beneath the canopy of the same saplings measured during the previous year. An additional sensor measured air temperature and relative hu-midity. The data logger was programed to scan each sensor every 60 s, to calculate sample averages every 15 min, and to calculate 24-h minimum and maximum temperatures for each of the sensors.
Gas exchange measurements
Gas exchange was measured in situ on a single shoot selected from the south side of each of the ten saplings. The measure-ments were made with an LI-6200 Portable Gas Exchange System (Li-Cor, Inc., Lincoln, NE) equipped with a 0.25-liter cuvette. The rate of CO2 depletion was measured over a 20-s interval after orienting the shoot normal to the direction of the sun. The same shoot from each of the saplings was used for all of the measurements during the sampling period. In a few cases, however, a shoot was accidentally damaged or broken, in which case a nearby shoot (usually on the same branch) was selected to take its place.
Two gas exchange sampling schedules were followed. Diur-nal sampling sessions were scheduled periodically to measure net photosynthetic rate and stomatal conductance at hourly intervals on each of the three trees where the thermistors or thermocouples were located. These sampling sessions were used to characterize the diurnal pattern of gas exchange in the saplings. A total of six diurnal sessions during the two spring sampling periods and five sessions during the fall periods suggested that daily maximum photosynthesis and stomatal conductance occurred in the afternoon over a broad time pe-riod (1300--1600 h). Subsequent sampling sessions, during which all ten saplings were measured, were scheduled during these times. Sampling sessions were scheduled on sunny or partly cloudy days in an attempt to minimize the variation in irradiance. However, on four occasions over the two years, the sky became cloudy during a sampling session and irradiances were substantially reduced. In addition, late in the fall 1993 sampling period irradiances were reduced because of low sun angles.
Leaf areas for individual shoots were measured with a Li-Cor LI-3100 portable area meter. The shoot from each sapling was harvested at the end of the sampling period and then brought to the laboratory where the needles were stripped off and the projected leaf area measured. Three measurements were made for each shoot and a sample mean was used to calculate net photosynthetic rate and stomatal conductance on a projected needle area basis.
Data analyses
Sample means were computed for each of the following gas-exchange-related variables for each sampling session: daily maximum net photosynthesis (Anet), stomatal conductance (gs), photosynthetically active radiation (PAR), and intercellu-lar CO2 concentration (Ci). Sample means were also computed for 24-h minimum and maximum soil temperatures at each of the sampled depths. Atmospheric vapor pressure deficit (VPD) at the time of the 24-h maximum air temperature was calcu-lated for the two sampling periods (spring and fall 1993) for which the necessary relative humidity data were available. Pearson product-moment correlation coefficients were com-puted between average daily maximum Anet and average PAR, 24-h maximum and minimum air temperatures, and average 24-h maximum and minimum soil temperatures at each soil depth. Minimum air temperature (min Tair), average minimum soil temperature measured at 5 cm depth (min5Tsoil), and aver-age minimum soil temperature measured at 10 cm depth
Results
Air and soil temperature
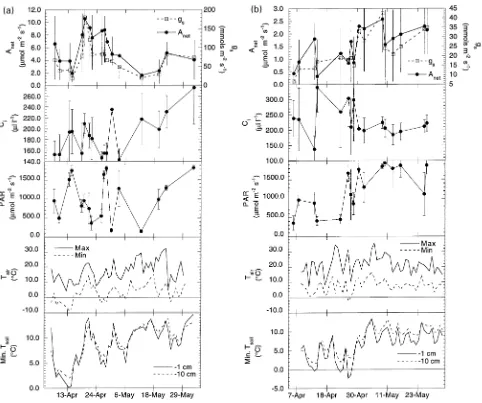
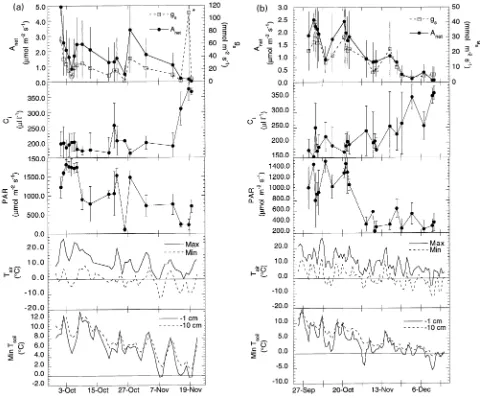
There was considerable day-to-day fluctuation in both 24-h minimum and maximum air temperatures (Figures 1a, 1b, 2a, and 2b) and in 24-h minimum soil temperature. The time series showed trends of gradually increasing temperature during the two spring sampling periods and gradually decreasing tem-perature during the two fall sampling periods. Exceptions to this general pattern were the air temperature data for the spring 1993 sampling, which showed little overall trend. Subfreezing nights were common throughout the spring of 1992 but oc-curred only once during spring 1993. There were frequent overnight frosts and steadily declining daytime temperatures during the fall sampling periods of both years.
Soil temperature tracked changes in air temperature, but the day-to-day fluctuation in soil temperature measured within the
rooting zone (at a depth of 10 cm) was less than the fluctuation at the soil surface (at a depth of 1 cm). In spring 1992, soil temperature remained above freezing throughout the measure-ment period, whereas in 1993, soil surface temperatures dropped below 0 °C on several occasions during April before warming significantly in May. Throughout the four sampling periods, soil temperatures in the rooting zone were usually 1 to 2 °C higher than surface temperatures, and minimum surface soil temperatures were usually 3 to 6 °C higher than minimum air temperatures, except for spring 1993 when the average temperature difference was less than 1 °C .
Net photosynthesis
Seasonal patterns in peak, daily gas exchange rates during the four sampling periods are shown in the top half of Figures 1a, 1b, 2a, and 2b. Gas exchange rates were measured between early April and late May on 17 days during spring 1992 and 16
Figure 1. Average daily maximum net photosynthesis (Anet), stomatal conductance (gs), intercellular CO2 concentration (Ci), photosynthetically active radiation (PAR), daily minimum and maximum air temperature (Tair), and daily minimum soil temperature at 1 cm and 10 cm depths
days during spring 1993. Fall measurements were begun in late September. In 1992, measurements ended in mid-November when average daily maximum net photosynthesis (Anet) reached approximately 0 µmol m−2 s−1 (a total of 19 days of measurements), whereas in 1993, measurements were made on 22 days because Anet was positive until well into December. During the two fall periods, peak rates of photosynthesis oc-curred early in the sampling period (late September and early October), whereas the pattern was less clear during the spring periods. In spring 1992, the peak occurred midway through the sampling period in mid-April, whereas in 1993 the peak oc-curred toward the end of the sampling period (mid- to late-May). During the spring 1992 sampling period, average daily maximum Anet was consistently higher than the rates during the spring 1993 sampling period and both fall periods. In both fall periods, Anet declined steadily and there were short-term
reduc-tions in gas exchange that corresponded with overnight frost events.
Overall, average daily maximum Anet ranged from nearly 0 µmol m−2 s−1 in late fall to over 10 µmol m−2 s−1 in mid-April 1992. The patterns of Anet, stomatal conductance (gs), and intercellular CO2 concentration (Ci) varied considerably among trees. This variation was consistent within sampling periods and among sampling periods.
Functional relationship between temperature and Anet
Daily maximum Anet and the various temperature variables were highly correlated. The association between average daily maximum Anet and average PAR was not as strong as the association between Anet and temperature (Table 1); however, the spring 1992 coefficient for PAR appears to be anomalous. The fall data displayed stronger correlations than the spring Figure 2. Average daily maximum net photosynthesis (Anet), stomatal conductance (gs), intercellular CO2 concentration (Ci), photosynthetically active radiation (PAR), daily minimum and maximum air temperature (Tair), and daily minimum soil temperature at 1 cm and 10 cm depths
data, and the 1993 data displayed stronger linear relationships than the 1992 data. This latter observation may be explained in part by the improved accuracy of the environmental data that were collected in 1993 compared with 1992. In particular, the continuous monitoring of temperatures by the data logger may have resulted in more reliable measurements of minimum and maximum temperatures, especially at the various soil depths.
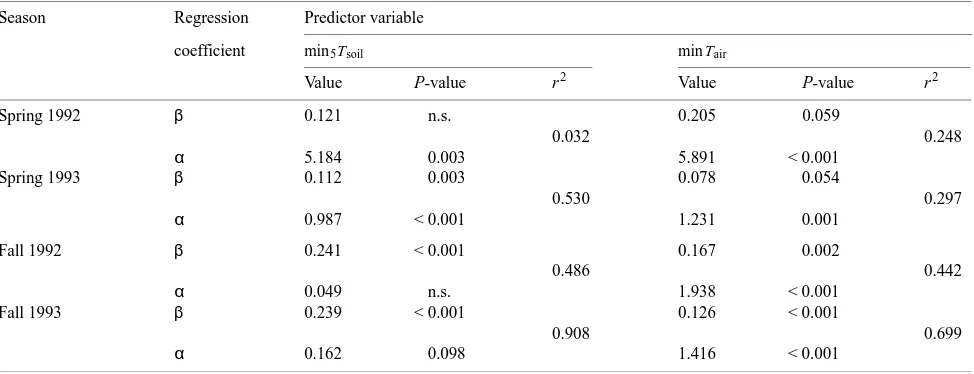
Summaries of the regressions for each of the four sampling periods (Table 2) illustrate that min5Tsoil was a significant predictor of daily maximum Anet (P < 0.005) in all but the spring 1992 sampling period, explaining 50 to over 90% of the variation in average daily maximum net photosynthesis in the other sampling periods.
Minimum Tair was a significant predictor of Anet (P < 0.005) only during the two fall sampling periods, and it explained only 30 to 70% of the variation. The regression summaries indicate that the least squares regression model fits the fall data better than the spring data and that the model fits the 1993 data better than the 1992 data. None of the environmental variables provided a significant linear relationship for the spring 1992 data, and scatter plots of average daily maximum Anet and other variables did not suggest other functional relationships.
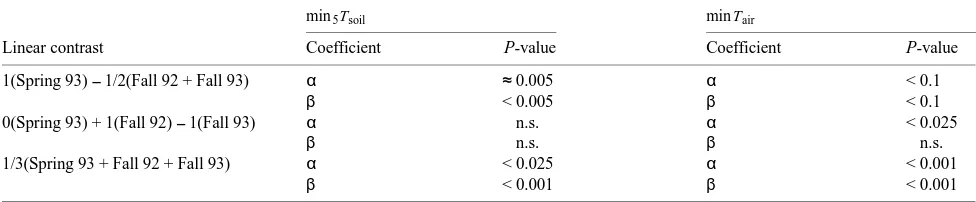
Although the regression slope coefficient for soil in spring 1992 (Table 2) was similar to the one in 1993, the regression itself was not significant. Moreover, the magnitude and range of measurements in 1992 were considerably higher than those during the same period in 1993. The measurements were also considerably higher than published values for many conifers (Teskey et al. 1995). This discrepancy suggested a potential calibration problem with the LI-6200, and the unit was sub-sequently re-calibrated by Li-Cor, Inc. Given these uncertain-ties, the spring 1992 data were removed from further analyses. There was a significant, positive linear relationship between average minimum Tsoil measured at 5 cm depth and average daily maximum Anet and between minimum Tair and average daily maximum Anet (Table 3). Furthermore, the results suggest that the linear relationship with minimum Tsoil was signifi-cantly different between the spring 1993 sampling period and the two fall periods, and that the relationship between the two fall periods for both predictors was not significantly different. In contrast, no seasonal differences were observed for the relationship between Anet and minimum Tair.
Discussion
Both minimum air and soil temperatures appeared to influence the pattern of photosynthesis during the spring and fall. During fall, short-term reductions in daily maximum Anet that were superimposed on a trend of steadily declining gas exchange (Figures 2a and 2b) were associated with overnight frost events and are consistent with the results of others (Fahey 1979, Table 1. Pearson product-moment correlation coefficients calculated
between average daily maximum Anet and various environmental fac-tors: PAR, photosynthetically active radiation; max Tair/min Tair, maxi-mum/minimum daily air temperature during the preceding 24 h; VPD, atmospheric vapor pressure deficit at time of max Tair; max1Tsoil/min1Tsoil, maximum/minimum daily soil temperature measured at a soil depth of 1 cm during the preceding 24 h; max5Tsoil/min5Tsoil, maximum/minimum daily soil temperature measure at a soil depth of 5 cm during the preceding 24 h; max10Tsoil/min10Tsoil, maximum/minimum daily soil temperature measured at a soil depth of 10 cm during the preceding 24 h.
Variable Spring Fall
1992 1993 1992 1993
PAR −0.065 0.570 0.487 0.746
max Tair −0.145 0.115 0.704 0.834
min Tair 0.498 0.545 0.665 0.836
VPD n.a. 0.315 n.a. 0.474
max1Tsoil n.a. 0.335 n.a. 0.861
min1Tsoil 0.205 0.696 0.719 0.902
max5Tsoil n.a. 0.548 n.a. 0.928
min5Tsoil 0.180 0.728 0.697 0.953
max10Tsoil n.a. 0.603 n.a. 0.938
min10Tsoil 0.137 0.734 0.680 0.956
Table 2. Linear, least squares regression summary for: daily max Anet = intercept (α) + slope (β) (minT).
Season Regression Predictor variable
coefficient min5Tsoil min Tair
Value P-value r2 Value P-value r2
Spring 1992 β 0.121 n.s. 0.205 0.059
0.032 0.248
α 5.184 0.003 5.891 < 0.001
Spring 1993 β 0.112 0.003 0.078 0.054
0.530 0.297
α 0.987 < 0.001 1.231 0.001
Fall 1992 β 0.241 < 0.001 0.167 0.002
0.486 0.442
α 0.049 n.s. 1.938 < 0.001
Fall 1993 β 0.239 < 0.001 0.126 < 0.001
0.908 0.699
Smith et al. 1984, DeLucia 1987). Together with the strong relationships between Anet and Tsoil in the fall sampling periods, these results suggest a simple model of environmental regula-tion of photosynthesis: the gradual decline in Anet may be driven by root-zone soil temperature, whereas short-term re-ductions are associated with overnight freezing. Such a model is consistent with the observations of DeLucia and Smith (1987) and Carter et al. (1987) who indicated that, for Rocky Mountain Engelmann spruce (Picea engelmannii (Parry)), low soil temperatures are more limiting to net photosynthesis and stomatal conductance than low air temperatures only when air temperatures are above freezing. However, maximum Tsoil was also highly correlated with fall Anet (Table 1), suggesting that a model for red spruce photosynthetic performance under declining temperatures may be more complex than the simple model proposed above. In addition, maximum Tair was strongly correlated with Anet during the two fall sampling periods (Table 1). This is likely to be related to the fact that maximum daily air temperatures during the fall measurement periods (15--30 °C) (Figures 2a and 2b) were in the optimal range for photosynthesis in conifers (Belous 1986, as cited in Teskey et al. 1995, p 114).
The regulation of Anet in red spruce during the spring appears to be more complex than during the fall, and the results of the present study do not provide a clear picture of environmental controls. Although air and soil temperatures were significant predictors of Anet, in spring 1993, possible effects of sub-freez-ing nights were not demonstrated because only one overnight frost occurred during the sampling period. This may explain, in part, the weaker relationship between minimum Tair and Anet that was observed during the spring 1993 sampling period compared with the spring 1992 sampling period (Figure 1b). Moreover, in spring 1992, the pattern of daily maximum Anet differed greatly from the pattern in 1993 with the highest photosynthetic rates observed early in the spring (Figure 1a).
In a laboratory study, DeLucia (1986) reported sharp de-clines in photosynthesis in Engelmann spruce seedlings after soil temperatures dropped below 8 °C. In another laboratory study, Day et al. (1991) reported sharp reductions in Anet in loblolly pine (Pinus taeda L.) seedlings when soil tempera-tures dropped below 10 °C, and Lippu and Puttonen (1991) reported significant reductions in Anet in Scots pine (Pinus
sylvestris L.) seedlings in an 8 °C soil temperature treatment
relative to a 12 °C treatment. Moreover, both DeLucia (1986) and Day et al. (1991) reported an overall curvilinear response in Anet to changes in soil temperature, but the response was linear when soil temperature fell below 10 °C. In the present study, minimum subsurface soil temperatures during the two fall sampling periods and the spring 1993 period were gener-ally below 10 °C, and the data for these three periods suggest a linear response of Anet to minimum soil temperature.
The high degree of covariance between air and soil tempera-tures in our field study makes it difficult to separate the effects of these factors on gas exchange in spring and fall. In general, minimum soil temperature explained more of the variation in average daily maximum Anet than did minimum Tair (Table 2). In addition, the steeper slope coefficients (β) in the regression equations with average minimum soil temperature suggest that Anet was more sensitive to changes in minimum soil tempera-ture than to changes in minimum air temperatempera-ture. A unit change in minimum soil temperature resulted in an Anet re-sponse that was 50 to 90% greater than the photosynthetic response from a similar change in minimum air temperature. This sensitivity suggests that, during both the fall cooling period and the spring warming period (1993), minimum soil temperature may not only be a better predictor of daily maxi-mum Anet than minimum Tair, but that photosynthesis may be more limited by minimum soil temperature than by minimum air temperature. Jurik et al. (1988) also concluded that soil temperature was a better predictor of Anet than air temperature during the springtime recovery of photosynthesis in white pine (Pinus strobus L.). Thus, after accounting for the effects of sub-freezing nights, it would seem that models of environ-mental control of fall (and probably spring) net photosynthesis in conifers in cold temperate regions should focus on soil temperatures.
We found a significant difference in the photosynthetic response of red spruce to changes in temperature between the spring and fall periods. The linear contrasts in Table 3 indicate that the difference between the spring 1993 slope estimate for minimum Tsoil (the predictor variable) and the average of the two fall estimates was highly significant (P < 0.005). However, the difference between the spring 1993 slope estimate for minimum Tair and the average of the two fall estimates was not significant (P < 0.1). Furthermore, although the regression equation between Anet and minimum Tsoil for spring 1992 was Table 3. Linear contrasts among regression slope and intercept coefficients. Coefficients were compared based on the t-test, computed using unequal variance assumptions and degrees of freedom calculated by Satterthwaite’s approximation.
min5Tsoil min Tair
Linear contrast Coefficient P-value Coefficient P-value
not significant, the slope estimate for spring 1992 was similar to the estimate for spring 1993 (Table 2). Because of the uncertainties in the spring data, these results are tentative; however, they suggest that the sensitivity of net photosynthesis to changes in soil temperature during the spring was approxi-mately half of that during the fall. This result is consistent with the hypothesis that the photosynthetic response of red spruce to temperature differs between the fall and spring.
The simplest regression-type model, based solely on Tair and Tsoil, may be inadequate for predicting the photosynthetic re-sponse of red spruce to changes in the spring warming and fall cooling periods of the growing season. In particular, the sensi-tivity of Anet to changes in temperature depends on the particu-lar physiological mechanisms that may constrain Anet during these periods, and these constraints probably differ between spring and fall. Changes in chloroplast thylakoid structure, chlorophyll content, photosynthetic enzyme activity, and elec-tron transport (see reviews by Öquist 1983, Havranek and Tranquillini 1995, Teskey et al. 1995 and references therein) are associated with winter hardening, and although the rela-tionships among these physiological changes is unclear, it is believed that these changes in combination with stomatal clo-sure are largely responsible for the winter decline in net pho-tosynthesis (Öquist 1983, Havranek and Tranquillini 1995, Teskey et al. 1995). Limitations to the springtime recovery of photosynthesis, on the other hand, have been associated with photoinhibition (Lundmark et al. 1988, Ottander and Öquist 1991, Strand and Lundmark 1995), insufficient soil water associated with soil thawing (Fahey 1979, Smith 1985, Jurik et al. 1988, Day et al. 1990), root hormones and mycorrhizal activity (Havranek and Tranquillini 1995). Thus, an improved model of photosynthetic sensitivity to temperature will depend on experimental work to isolate the relevant mechanisms and to describe their seasonal dependence on air and soil tempera-tures. Finally, although warmer air and soil may delay the onset of winter hardening and hasten springtime recovery, the sig-nificance of extending the growing season must be assessed in terms of whole-plant responses and overall carbon balance.
Acknowledgments
The senior author gratefully acknowledges the support and assistance of the staff at the Boyce Thompson Institute Research Field, especially C. Tutton. In addition, the author thanks M. Stover, C. Williams, R. Sladek, and S. Schwarz for their help in the field and also C. McCul-loch for his advice on statistical analysis.
References
Alscher, R.G., R.G. Amundson, J.R. Cumming, S. Fellows, J. Fincher, G. Rubin, P. Van Leuken and L.H. Weinstein. 1989. Seasonal changes in the pigments, carbohydrates and growth of red spruce as affected by ozone. New Phytol. 113:211--224.
Amundson, R.G., R.J. Kohut, J.A. Laurence, S. Fellows and L.J. Colavito. 1993. Moderate water stress alters carbohydrate content and cold tolerance of red spruce foliage. Environ. Exp. Bot. 33:383--390.
Blum, B.M. 1990. Picea rubens Sarg. red spruce. In Silvics of North America: 1. Conifers. Eds. R.H. Burns and B.H. Honkala. Agric. Handbook 654. U.S. Department of Agriculture, Forest Service, Washington, DC, pp 250--259.
Carter, G.A., W.K. Smith and J.L. Hadley. 1987. Stomatal conduc-tance in three conifer species at different elevations during summer in Wyoming. Can. J. For. Res. 18:242--246.
Day, T.A., E.H. DeLucia and W.K. Smith. 1989. Influence of cold soil and snowcover on photosynthesis and leaf conductance in two Rocky Mountain conifers. Oecologia 80:546--552.
Day, T.A., E.H. DeLucia and W.K. Smith. 1990. Effect of soil tem-perature on stem sap flow, shoot gas exchange and water potential of Picea engelmannii Parry during snowmelt. Oecologia 84:474--481.
Day, T.A., S.A. Heckathorn and E.H. DeLucia. 1991. Limitations of photosynthesis in Pinus taeda L. (loblolly pine) at low soil tempera-tures. Plant Physiol. 96:1246--1254.
DeHayes, D.H. 1992. Winter injury and developmental cold tolerance in red spruce. In The Ecology and Decline of Red Spruce in the Eastern United States. Eds. C. Eager and M.B. Adams. Springer-Verlag, New York, pp 296--337.
DeLucia, E.H. 1986. Effect of low root temperature on net photosyn-thesis, stomatal conductance and carbohydrate concentration in Engelmann spruce (Picea engelmannii Parry ex Engelm.) seed-lings. Tree Physiol. 2:143--154.
DeLucia, E.H. 1987. The effect of freezing nights on photosynthesis, stomatal conductance, and internal CO2 concentration in seedlings of Engelmann spruce (Picea engelmannii Parry). Plant Cell Envi-ron. 10:333--338.
DeLucia, E.H. and W.K. Smith. 1987. Air and soil temperature limita-tions on photosynthesis in Engelmann spruce during summer. Can. J. For. Res. 17:527--533.
Eamus, D. 1993. Assimilation and stomatal conductance responses of red spruce to midwinter frosts and the constituent ions of acid mist. Tree Physiol. 13:145--155.
Fahey, T.J. 1979. The effect of night frost on the transpiration of Pinus contorta ssp. latifolia. Oecol. Plant. 14:483--490.
Fincher, J. and R.G. Alscher. 1992. The effect of long-term ozone exposure on injury in seedlings of red spruce (Picea rubens Sarg.). New Phytol. 120:49--59.
Fincher, J., J.R. Cumming, R.G. Alscher, G. Rubin and L. Weinstein. 1989. Long-term ozone exposure affects winter hardiness of red spruce (Picea rubens Sarg.) seedlings. New Phytol. 113:85--96. Hadley, J.L., R.G. Amundson, J.A. Laurence and R.J. Kohut. 1993.
Physiological response to controlled freezing of attached red spruce branches. Environ. Exp. Bot. 33:591--609.
Havranek, W.M. and W. Tranquillini. 1995. Physiological processes during winter dormancy and their ecological significance. In Eco-physiology of Coniferous Forests. Eds. W.K. Smith and T.M. Hinckley. Academic Press, New York, pp 95--124.
Jurik, T.W., G.M. Briggs and D.M. Gates. 1988. Springtime recovery of photosynthetic activity of white pine in Michigan. Can. J. Bot. 66:138--141.
Kaufmann, M.R. 1982. Evaluation of season, temperature and water stress effects on stomata using a leaf conductance model. Plant Physiol. 69:1023--1026.
Kozlowski, T.T., P.J. Kramer and S.G. Pallardy. 1991. The physiologi-cal ecology of woody plants. Academic Press, New York, 657 p. Levitt, J.D. 1980. Responses of plants to environmental stresses, 2nd
L’Hirondelle, S.J., J.S. Jacobson and J.P. Lassoie. 1992. Acidic mist and nitrogen fertilization effects on growth nitrate and reductase activity and frost hardiness of red spruce seedlings. New Phytol. 121:611--622.
Lippu, J. and P. Puttonen. 1991. Soil temperature limitations on gas exchange in 1-year-old Pinus sylvestris (L.) seedlings. Scand. J. For. Res. 6:73--78.
Lundmark, T., J.-E. Hällgren and J. Hedén. 1988. Recovery from winter depression of photosynthesis in pine and spruce. Trees 2:110--114.
Öquist, G. 1983. Effects of low temperature on photosynthesis. Plant Cell Environ. 6:281--300.
Ottander, C. and G. Öquist. 1991. Recovery of photosynthesis in winter-stressed Scots pine. Plant Cell Environ. 14:345--349. Schaberg, P.G., R.C. Wilkinson, J.B. Shane, J.R. Donnelly and P.F.
Cali. 1995. Winter photosynthesis of red spruce from three Vermont seed sources. Tree Physiol. 15:345--350.
Smith, W.K., D.R. Young, G.A. Carter, J.L. Hadley and G.M. McNaughton. 1984. Autumnal stomatal closure in six conifer spe-cies of the Central Rocky Mountains. Oeologia 63:237--242.
Smith, W.K. 1985. Environmental limitations on leaf conductance in Central Rocky Mountain conifers, USA. In Establishment and Tending of Subalpine Forests: Research and Management. Eds. H. Turner and W. Tranquillini. Proc. 3rd IUFRO Workshop. Eidg. Anst. Forstel Versuch. 270:95--101.
Snedecor, G.W. and W.G. Cochran. 1980. Statistical methods, 7th Edn. Iowa State University Press, Ames, Iowa, 507 p.
Strand, M. and T. Lundmark. 1995. Recovery of photosynthesis in 1-year-old needles of unfertilized and fertilized Norway spruce (Picea abies (L.) Karst.) during spring. Tree Physiol. 15:151--158. Teskey, R.O., D.W. Sheriff, D.Y. Hollinger and R.B. Thomas. 1995.
external and internal factors regulating photosynthesis. In Resource Physiology of Conifers. Eds. W.K. Smith and T.M. Hinckley. Aca-demic Press, New York, pp 105--140.