Symptoms: A Behavior–Genetic Perspective
H.H. Goldsmith and Kathryn S. Lemery
Research on mood and anxiety disorders has historically proceeded without sufficient reference to the growing body of work on the nature of typical emotional develop-ment and temperadevelop-ment. Reviewing data from several studies, we consider experiential, biological, and genetic factors as providing causal input to typical developmental variation in fearfulness and anxiety during infancy and early childhood. Longitudinal behavioral methods, psy-chophysiologic measures, and a behavior– genetic frame-work are used to approach these issues. Results from twin studies implicate moderately strong genetic influences on different facets of temperamental fearfulness, as well as childhood anxiety symptoms. Then, we consider the dis-tinction between normal range temperament and overt anxiety symptoms from a quantitative genetic perspective. Biological correlates (cortisol, asymmetric frontal EEG activation, cardiac reactivity) of inhibited behavior are considered as related endophenotypes for anxiety. In a nongenetic analysis, we report the prediction of internal-izing problems during kindergarten from earlier temper-ament and earlier basal cortisol measures. Our review highlights connections between behavioral indicators and various putative endophenotypes and the fuzzy boundary between normal-range temperament and anxiety disor-ders. Biol Psychiatry 2000;48:1199 –1209 © 2000 Soci-ety of Biological Psychiatry
Key Words: Anxiety, temperament, twins, genetics, fear,
childhood
Introduction
A
re individual differences in fearfulness and shyness sources of vulnerability to anxiety disorders? Focus-ing on recent work from our laboratory, we review results from a coordinated set of studies that incorporate a diverse but complementary range of research methods and assess-ment techniques.The notion of temperament incorporates the normal range of variation and is broad enough to include issues of
attention, emotion, and regulatory processes (Goldsmith et al 1987). Temperament is alternately conceived of in terms of traits (e.g., fearfulness) or types of children (the inhibited child). Temperamental concepts such as behav-ioral inhibition, shyness, and fearfulness overlap concep-tually with the anxiety domain. Fearfulness often is viewed as a family of traits, including social reticence (beginning in infancy), concern about social evaluation (beginning in toddlerhood or early childhood and becom-ing more prominent later), and distress about events in the physical world (fear of the dark, of large animals, of loud noises, etc.). This definition bears obvious similarities to anxiety symptoms, as described in DSM-IV. It is probably impossible to distinguish temperamental qualities from anxiety based solely on the observable nature of behav-ioral responses (distress, withdrawal, etc.) in the two domains. Key distinctions include the contextual appro-priateness of the response, its pervasive and unregulated quality, and its maladaptive consequences. Anxiety occurs in contexts other than those involving novelty, threat, and loss; it is more pervasive and less well modulated across occasions; and it often leads to interpersonal and school-or wschool-ork-related dysfunction. Studying the phenotypic distinctions between temperament and anxiety in develop-mental perspective is one route to better understanding these distinctions, but real progress requires moving be-yond the phenotypic level of analysis.
Several important themes have figured prominently in recent research, including the boundary between normal and disordered, the nature of affect regulation, and the importance of context in determining the significance of affective responding. These are not mutually exclusive issues. As it applies to childhood anxiety, the issue of the boundary between normal and disordered involves tem-perament and clinical diagnoses, as mentioned above. The most straightforward question is whether some forms of psychopathology represent extreme values on a normal continuum (e.g., anxious disorder as extreme temperamen-tal fearfulness) or whether forms of psychopathology are qualitative types that might simply appear phenotypically as though they are on a continuum with normal-range functioning. Given that the current nosology for childhood anxiety disorders yields a high degree of comorbidity and
From the Center for Affective Science, University of Wisconsin, Madison. Address reprint requests to H. Hill Goldsmith, UW-Madison, Department of
Psychology, 1202 West Johnson Street, Madison WI 53706. Received March 7, 2000; revised July 26, 2000; accepted July 28, 2000.
incorporates minimal consideration of developmental pro-cesses (Anxiety Disorders Association of America, 2000), few would claim that it is optimal as a starting point for discovering etiology. Perhaps studying anxiety disorders in relation to temperament might aid in clarifying nosol-ogy more generally, in addition to elucidating the border-lines between normal-range functioning and dysfunction. A second perspective that distinguishes psychopathol-ogy from extreme temperament is regulation, particularly affective regulation. This topic is complex (Fox 1994), but the kernel of the perspective is that failures of emotional regulation rather than differences in reactive properties of the nervous system account for clinical disorders.
The perspective that emphasizes context suggests that even extreme fear reactions are not indicative of presence of, or risk for, anxiety disorders as long as these reactions occur in the face of real or perceived threat (i.e., the normal context for fear). When fear reactions begin to occur out of typical context (i.e., the individual begins to appraise objectively benign contexts as threatening), psy-chopathologic implications accrue.
Clearly, distinguishing among these alternatives, or perhaps weighting the importance of these alternatives, would be problematic if research were confined to behav-ioral approaches. One framework that can aid understand-ing is the quantitative framework of behavioral genetics, which we pursue in this article.
A behavior– genetic framework includes many compo-nents. The demonstration that the temperamental and behavior problem domains are indeed phenotypically cor-related for nonartifactual reasons is an obvious prelimi-nary step. Then, genetic contributions to variation in temperament and anxiety symptom clusters need to be documented. A key follow-up to that documentation is the joint study of temperament and symptoms within a behav-ior– genetic perspective. Such joint study should establish the degree to which the association between temperamen-tal traits and anxiety problems is accounted for by over-lapping genetic factors and overover-lapping environmental factors. Then, the continuity of temperament and of anxiety problems needs to be estimated because an etio-logic role of temperament seems to imply such continuity. To expand the nomologic network and facilitate the search for candidate genes, biological correlates of fearful tem-perament and anxiety disorders need to be analyzed for common genetic underpinnings. All of these components are not yet fully established empirically, but our review will document considerable progress.
Our strategy is to treat each of the components of the approach outlined in the previous paragraph. Under each component, we first provide a selective account of the literature and then present recent findings from our re-search group to illustrate the issues in more detail.
Phenotypic Correlations between
Temperament and Anxiety Symptoms and
Disorders
Background Literature
Several studies adopt a cross-sectional, correlational ap-proach to examining the relation between temperament and anxiety in children. Fewer studies examine tempera-ment with children with diagnosed disorders or take a longitudinal approach to partially protect against situa-tional specificity. In a cross-secsitua-tional analysis, Merikan-gas and colleagues (MerikanMerikan-gas et al 1998) found that children with diagnosed anxiety disorders using DSM-III-R criteria had low scores on adaptability and approach/ withdrawal and high scores on reactivity (using the Di-mensions of Temperament Survey) compared with children without a disorder. Biederman et al (1993) followed up two existing samples with diagnostic assess-ments separated by 3 years. At the second follow-up of the combined samples, the previously inhibited children showed significantly higher rates of multiple anxiety disorders and avoidant and phobic disorder than did noninhibited children. In another longitudinal sample, Kagan et al (1999) found that infants who were highly reactive to unfamiliar stimuli at 4 months were more likely to be inhibited at 4 years (about 20% of all high reactives) and displayed more anxious symptoms at 7 years. In fact, toddler behavioral inhibition predicted generalized social anxiety at 13 years, especially for girls (Schwartz et al 1999). Similarly, adolescents’ retrospective self-reports of social avoidance and fearfulness were associated with current social phobia (Hayward et al 1998). We now turn to a more detailed account of results from our research group.
Correlations between Temperament and Anxiety Scales
For these analyses (and several others in this paper), we used Rothbart’s Children’s Behavior Questionnaire (CBQ; Rothbart et al 1994) as our measure of childhood temper-ament. Three CBQ scales are relevant for the study of the reactive aspects of temperament possibly related to anxi-ety: Fear (negative affect resulting from anticipation of threat), sadness (negative affect related to disappoint-ment), and shyness (inhibited approach in a social setting). Parents decide whether each item is “true” or “untrue” (rated on a seven-point scale with a “not applicable” option) of their child within the past 6 months.
“Overly upset when leaving someone s/he is close to.” The HBQ symptom scales are a revision of the Ontario Child Health Study Scales (OCHS-R; Offord et al 1991), includ-ing content similar to the Child Behavior Checklist (Achenbach and Edelbrock 1981). This instrument is applicable for caregivers of 4- to 16-year-olds and yields “screening-quality” diagnoses of overanxious disorder, separation anxiety, and other forms of child psychopathol-ogy. The reliability and validity of the OCHS-R is com-parable to that obtained by the more structured Diagnostic Interview for Children and Adolescents (Boyle et al 1997). Most of the results to be reported are qualified by our decision to use the HBQ anxiety measure dimensionally rather than to set thresholds to capture only clinically meaningful levels of anxiety.
If the CBQ contains items that tap anxiety symptoms, then any association between temperament and anxiety might be partially artifactual. Lemery and colleagues (Lermy et al 2000) explored this issue in detail, using both a conceptual expert rater approach and an empirical confirmatory factor analysis approach to identify con-founded items on the CBQ. For example, experts
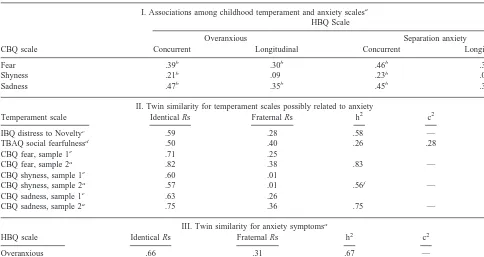
identi-fied three fear items (“Is afraid of fire,” “Is very frightened by nightmares,” and “Is afraid of the dark”) and two sadness items as confounded (“Becomes tearful when told to do something s/he does not want to do,” and “Some-times appears downcast for no reason”). After dropping these potentially problematic items from the CBQ scales, the predictive power of the purified CBQ scales for later behavior problems remained, suggesting that conceptual content contamination did not account for the association. As part of a longitudinal study of twins identified from birth records, we administered the CBQ to mothers and fathers when twins were at a mean age of 5.06 years (SD 5 1.15), ranging from 3.06 to 8.56 years. About 1 year and 3 months later, we followed-up with both the CBQ and HBQ. Written informed consent was obtained at both phases of the study (and in all other studies reported). As Table 1, part I, shows, there were moderate associa-tions, and no difference, in the magnitude of the correla-tions with temperament for the two types of anxiety. The fear and sadness scales of the CBQ correlated with concurrent anxiety more strongly than did the shyness scale. When we examined the longitudinal correlations,
Table 1. Phenotypic Correlations among temperament and Anxiety Measures and Twin Similarity for Infant, Toddler, and Childhood Temperament and Anxiety Scales
I. Associations among childhood temperament and anxiety scalesa
HBQ Scale
Overanxious Separation anxiety
CBQ scale Concurrent Longitudinal Concurrent Longitudinal
Fear .39b .30b .46b .32b
Shyness .21b .09 .23b .02
Sadness .47b .35b .45b .30b
II. Twin similarity for temperament scales possibly related to anxiety
Temperament scale Identical Rs Fraternal Rs h2 c2 e2
IBQ distress to Noveltyc .59 .28 .58 — .42
TBAQ social fearfulnessd .50 .40 .26 .28 .46
CBQ fear, sample 1e .71 .25
CBQ fear, sample 2a .82 .38 .83 — .17
CBQ shyness, sample 1e .60 .01
CBQ shyness, sample 2a .57 .01 .56f — .44
CBQ sadness, sample 1e .63 .26
CBQ sadness, sample 2a .75 .36 .75 — .25
III. Twin similarity for anxiety symptomsa
HBQ scale Identical Rs Fraternal Rs h2 c2 e2
Overanxious .66 .31 .67 — .33
Separation anxiety .76 .54 .40 .35 .25
All correlations were corrected for gender differences and for linear and quadratic effects of age. Heritability (h2), common (shared) environmental (c2), and nonshared
environmental (e2) estimates were obtained from genetic model fitting. —, the estimate was nonsignificant and dropped from the model. HBQ, Health and Behavior
Questionnaire; CBQ, Children’s Behavior Questionnaire; IBQ, Infant Behavior Questionnaire; TBAQ, Toddler Behavior Assessment Questionnaire.
aFrom Lemery (1999); N5107 identical and 163 fraternal twin pairs. Scales are composites of mother and father report. bP,.01.
the CBQ shyness scale was not predictive even of sepa-ration anxiety, suggesting that shyness is not the crucial temperamental substrate of overanxious and separation anxiety symptoms, at least according to these assessments. Of course, if we had measured avoidant disorder or social phobia, a link with temperamental shyness might have been uncovered.
Genetic Factors in Early Temperamental
Traits Possibly Related to Anxiety
Background Literature
Two longitudinal twin studies in early childhood provide evidence for genetic influences on behavioral inhibition. Robinson et al (1992) formed a composite inhibition measure coded from observed laboratory behaviors for a sample of twins seen at 14, 20, and 24 months of age. Heritability estimates clustered around .50 at all ages, with no influence of the shared environment. Genetic correla-tions indicated that the same genetic factor influenced inhibition at all ages (see also DiLalla et al 1994). Similarly, Matheny (1989) measured inhibition in a lab setting, during mental testing, and by parent report ques-tionnaire for a sample of twins assessed at 12, 18, 24, and 30 months of age. Again, behavioral inhibition was heri-table at all ages. Genetic influences also accounted for the degree of change in the trait over time. Thus, the literature suggests that behavioral inhibition is moderately heritable, the same genetic factor can account for the stability of the trait, and genes also influence the degree of change in the trait.
Twin Analyses of Temperament Questionnaire Scales
Part II of Table 1 shows twin intraclass correlations for infant (Rothbart 1981), toddler (Goldsmith 1996), and early childhood (Rothbart et al 1994) temperament scales related to anxiety. The data come from three studies (Goldsmith et al 1997a, 1999; Lemery 1999). Then, with univariate genetic model fitting, we estimated the herita-bility (h2), common or shared environment (c2), and unique environment (e2) for each scale. For CBQ shyness, we also estimated dominant or nonadditive genetic effects (d2) because of the pattern of identical twins being more than twice as similar as fraternal twins, who showed no intrapair similarity. We started with the full model (i.e., allowing both genetic and common environmental influ-ences to account for twin similarity); then we dropped parameters to consider reduced models (e.g., twin similar-ity entirely due to genetic effects or twin similarsimilar-ity entirely due to environmental influences). There were no signifi-cant gender effects, and older children tended to have
slightly lower scores on the three temperament scales, a finding consistent with other literature. Nevertheless, we regressed out the effects of gender and linear and quadratic effects of age because they can inflate estimates of twin similarity (McGue and Bouchard 1984).
Identical twin resemblance exceeded fraternal twin similarity in all cases, with the pattern generally suggest-ing moderate genetic effects. An exception was the toddler results, where the fraternal correlation approached the identical twin value, suggesting shared environmental effects during this age period. Because the first sample tested with the CBQ was small (Goldsmith et al 1997a), we provide results from a second, much larger and more representative sample (Lemery 1999); the degree of rep-lication was remarkably strong, even including the near-zero fraternal twin correlation for shyness.
Genetic Factors in Childhood Anxiety
Disorders
Background Literature
We note some highlights of the modest literature on quantitative genetic factors in anxiety, a literature that focuses on older children and adolescents. With commu-nity samples from various studies, the Child Behavior Checklist has yielded heritability estimates of 40 –58%, with a smaller significant effect of the shared environment of 22–38% on internalizing symptoms (for review, see Goldsmith et al 1997b).
Using the Revised Childrens Manifest Anxiety Scale, Thapar and McGuffin (1995) obtained a heritability esti-mate of 59% for mother report of anxious symptoms with more than 300 pairs of 8- to 17-year-old twins, with no effect of the shared environment. Similarly, with self-report at 7 years, Warren et al (1999) found heritable influences on physiologic and social anxiety symptoms. In contrast, with adolescent self-report of anxious symptoms, shared environment accounted for 55% of the variance, with no significant genetic effect (Thapar and McGuffin 1995). This finding of large effects of the shared environ-ment on self reports compared with parental reports also occurred in another project (Stevenson et al 1992), in which 49% of the variance in childhood fears was due to the shared environment. Typically, parents report fewer symptoms than do children and perhaps report on more enduring traits. Topolski et al (1997) reported that shared environmental, and not additive genetic, influences ac-counted for separation anxiety symptoms in a community-based twin study of 8- to 16-year-olds using self-report. In contrast, for overanxious disorder symptoms, the shared environmental effect was nonsignificant, and genetic ef-fects accounted for about 40% of the variance.
symptoms in childhood are moderately heritable, with separation anxiety symptoms also influenced by the shared environment. Parent report yields higher heritability esti-mates, whereas adolescent self-report yields larger influ-ences of the shared environment. In a useful developmen-tal extension of this approach to the genetics of anxiety, Gjone and colleagues (Gjone et al 1996) found the heritability of internalizing to decrease and the shared environmental influence to increase with age. Addition-ally, with younger children, the heritability of internalizing decreased and the shared environmental influence in-creased in the extremes of the distribution.
Twin Analyses of the HBQ Anxiety Scales
Part III of Table 1 shows twin analyses for the two HBQ anxiety scales from the same project that provided the part I and part II, CBQ sample 2 results. We made composites from the correlated maternal and paternal reports on the HBQ in an effort to partially protect against rater bias. We also adjusted for any age and gender effects. For the genetic model fitting, the frequency distributions of the HBQ composites were transformed to approximate nor-mality, using the equation, y5 ln (11x).
There was evidence of different patterns for the over-anxious symptoms versus separation anxiety. The three right-most columns of Table 1 display the standardized estimates obtained for the best fitting model of each scale. Similar to the temperament scales, the best fitting model for overanxious included a moderately strong genetic effect (h25 .67) with no effect of the common environ-ment. On the other hand, twin similarity for separation anxiety was accounted for by both genetic factors (h25
.40) and the common environment (c25.35), a pattern not typically found in the temperament literature.
Another question that can be addressed with the twin data on these anxiety scales is whether high anxiety is qualitatively or only quantitatively distinct from the typi-cal level of manifestation of anxiety in the nondisordered population. To explore the quantitative versus qualitative distinction in more detail, we used the DF regression approach (Cherny et al 1992; DeFries and Fulker 1985) to compare the heritability estimates for full-range variation (individual heritability) with the extreme group scoring beyond some threshold (group heritability). Differences between these types of heritability would support the hypothesis that extreme symptoms reflect a qualitative departure from normal range variation. Group heritability is the extent to which the difference between the proband mean and the mean of the unselected population is due to heritable influences. Large samples are needed for ade-quate power to detect significant differences among heri-tability estimates; therefore, we plan to expand the sample on which the following analyses are based.
In the basic model, estimates of group heritability are obtained by predicting the cotwin’s score by the proband score and the coefficient of genetic relationship (1.00 for identical twins who share 100% of their genes and 0.50 for fraternal twins who share on average 50% of their segre-gating genes). With our sample, the proband group was selected as having scores 1 SD above the mean. If both cotwins were probands, each twin was entered once as the proband and once as the cotwin, and standard errors were corrected for the appropriate sample size. We fitted the DF regression model to the HBQ overanxious and separation anxiety scales separately.
For overanxious, part III of Table 1 gives the estimate of individual heritability, h25.67. Using DF regression, we obtained an estimate of .52 for group heritability, a nonsignificant difference. This result supports continuity in the underlying etiology of normal range and extreme scores on overanxious. For separation anxiety, the indi-vidual heritability estimate given in Table 1 was .40. The estimate of group heritability from the regression model was .21, again a nonsignificant difference with our sample size. Thus, for both anxiety scales, we cannot conclude that there are differences in heritability between the extremes and the full range of scores. On the other hand, the descriptive pattern does suggest slightly lower herita-bility in the extremes. Perhaps the safest empirical con-clusion is simply that the extremes are not more heritable than the full range and that additional sources of environ-mental variance might come into play for extreme scores.
Continuity of Temperament
Much of the reasoning concerning links between temper-ament and anxiety disorders entails an assumption that temperament is stable. Perhaps surprisingly, this assump-tion only finds limited support in the literature.
Background Literature
Stability correlations in childhood are generally modest to moderate for temperament, depending on the interval of repeated measurement and the method of measurement (Guerin and Gottfried 1994; Pettit and Bates 1984; Plomin et al 1993; Wilson and Matheny 1986). The majority of these studies used inexpensive parental report methodol-ogy, and there exists concern that the stability of parental report might be due to stable parental perception of the child rather than actual stable behavior. Despite such problems, several reviews have concluded that parent report of temperament is reasonably valid (Bates 1989; Goldsmith and Rieser-Danner 1990; McDevitt 1986).
expla-nation for these moderate correlations is contextual differ-ences. Contexts such as family life and school differ in their demands on children. Contexts vary in structure, intensity of stimulation, and novelty. A third explanation for moderate stability correlations is the subsequent devel-opment of systems that influence temperament, such as attentional systems (Rothbart and Bates 1998). Fourth, stability correlations could be dampened because the middle of the distribution may be less stable than the extremes, as noted empirically (Kagan et al 1988; Sanson et al 1996), although this statistical fact does not compel strong conclusions about etiology. This fourth possibility is important for linking temperament and anxiety disorders because an assumption of temperamental continuity need only apply to those who eventually develop anxiety disorders.
Change in temperament has not been studied systemati-cally, mainly because of the difficulty in separating real change from measurement error. In addition to documenting the magnitude of change, we need to understand the behav-ioral and emotional regulatory systems that develop during infancy and childhood and then influence temperament and adjustment. Behavior genetic studies have shown strong identical twin similarity in change over time, with Matheny’s work (Matheny 1989), described above, on behavioral inhi-bition being perhaps most relevant to anxiety.
Evidence for Stability and Change in Temperament
In our own work, we have used both the dimensional and categorical approaches to the study of the continuity of childhood temperamental traits related to anxiety. For in-stance, using the CBQ temperament scales introduced above, we computed the following stability correlations from 4 years (Time 1) to 7 years (Time 2) in a sample of 58 children previously classified as inhibited, uninhibited, or intermediate (Pfeifer et al 1999): fear (r5.60), shyness (r5 .77), and sadness (r5 .64). In another much larger and more repre-sentative sample from our research program, we recomputed these three CBQ stability correlations over a 1-year interval, from age 3.5 to 4.5 years (Lemery et al 2000). The results were strikingly consistent: fear (r5.60), shyness (r5.73), and sadness (r5.62).
Next, in the smaller sample, we considered stability and change of the shy/bold classification made in toddlerhood on the basis of carefully observed and coded behavior in the laboratory. Three follow-up measures showed evi-dence of continuity. Children who had been in the inhib-ited group as toddlers were rated as significantly more shy than those who had been in the uninhibited group on 1) laboratory-based measures of inhibition at age 7 years; 2) on the CBQ shyness scale by their mothers at 4 years; 3) and on the same CBQ shyness scale by their mothers at 7
years. Parallel findings emerged for the uninhibited pole of the toddler classification; children rated as stably bold by mothers had been uninhibited as toddlers in the lab; however, considerable change was present as well. About half of the inhibited toddlers were not stably shy from 4 to 7 years, and one third of uninhibited toddlers were not later stably bold. One extreme example of change was the single previously inhibited child who was later reported as bold at both 4 and 7 years by the mother (Pfeifer et al 1999). Thus, change from one extreme to the other was very rare over the age span of this study, but change from an extreme to the broad middle range of the distribution was the norm, and this conclusion held across different modes of assessment.
Our evidence for partial continuity is consistent with recent analyses in which inhibition was first measured in infancy by Kagan et al (1998). The results invite a longitu-dinal examination of the effects of temperament on anxiety symptoms, and we did so using a twin study design.
The Joint Study of Temperament and
Anxiety Symptoms: A Behavior–Genetic
Perspective
Background Literature
To what extent is the genetic vulnerability to anxiety associated with temperament? Schmitz et al (1999) ob-tained mother report on the Colorado Childhood Temper-ament Inventory at 14, 20, 24, and 36 months. At 48 months, mothers reported internalizing symptoms on the Child Behavior Checklist. Earlier temperamental emotion-ality and shyness were positively correlated with internal-izing at 48 months. The intraclass correlations for the identical twins were modest (about .40), and for fraternal twins, they were near zero for emotionality and shyness at all ages. In contrast, internalizing showed large shared environmental influences and more twin similarity overall. Across the four ages of temperament assessment, common genetic influences explained an average of 94% (range 76 –100%) of the expected phenotypic correlation between emotionality and later internalizing and an average of 79% (range 50 –100%) of the correlation for shyness and later internalizing across age. Nonshared environment also influenced the relation between shyness rated after 2 years and later internalizing. Thus, common genetic influences generally accounted for the relation between temperament and symptoms.
Bivariate Analyses of Temperament and Anxiety Symptoms
measure, we conducted bivariate biometrical analyses (Lemery 1999). These analyses addressed genetic and environmental influences on the covariation between tem-perament and anxiety in addition to the variation of these measures themselves. Obviously, such analyses cannot be undertaken when there is no phenotypic covariation be-tween a pair of temperament and anxiety measures, and there is probably stronger motivation to study the sources of higher degrees of phenotypic covariation; however, there is no necessary link between the degree of pheno-typic covariation and the degree of genetic influence on that overlap. Thus, it is of interest to estimate the sources of even modest levels of temperament by anxiety symp-tom covariation. Thus, the phenotypic association between temperament and anxiety was decomposed into its genetic and environmental components. To partially guard against biases due to completing both temperament and anxiety questionnaires on the same occasion (e.g., mood of rater), we focused on the longitudinal relation between early temperament and anxiety symptom clusters measured over 2 years later, using a composite of mother and father report.
We first fit full models (A5additive genetic influence, C 5 common environmental influence, E 5 nonshared environmental influence; the ACE model), then dropped parameters to fit more parsimonious models (e.g., the AE or CE model). The chi-square test was used to evaluate the fit of the model to the data; thus, a small chi-square corresponded to good fit, and a large chi-square corre-sponded to bad fit. The chi-square difference test was used to compare simpler models with the full model and determine which nested model fit best. In addition, a small (negative) Akaike’s Information Criterion (AIC) value was associated with better fit (for a comparison of fit indices, see Tanaka 1993).
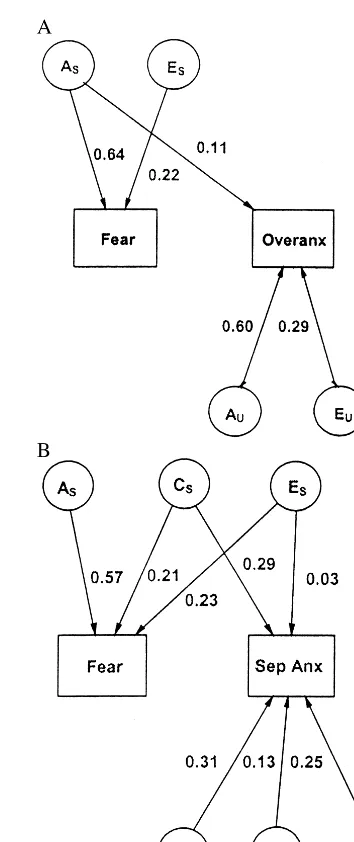
Figure 1A portrays the best fitting bivariate genetic model (the ACE model) for early fear and overanxious [x2(15)516.06, p5.38, AIC5 213.94]. The AE model fit best, with no effect of the common environment on either phenotype. In addition, the nonshared environmen-tal influences on early fear and overanxious were com-pletely independent. A similar model fit best for early sadness and overanxious [x2(14)519.15, p5.16, AIC5 28.85, with a small, but significant joint nonshared environmental influence accounting for 1% of the variance in Overanxious]. Thus, although most of the genetic influences on temperament and overanxious symptoms were independent, genes did account for their phenotypic association.
Next, we move to an examination of the biometric associations of fear and separation anxiety, which had a similar phenotypic association. Figure 1B depicts the best fitting bivariate genetic model (the ACE model) for early
fear and separation anxiety [x2(12) 5 10.79, p 5 .55, AIC 5 213.21]. In this model, 29% of the variation in separation anxiety was due to a common environmental effect that was shared with earlier fear (and separation
anxiety had an additional unique common environmental effect of 13%). A similar model fit best for early sadness and separation anxiety [x2(13)5 13.46, p5.41, AIC5 212.54]. With this model, all of the 40% of the variance in separation anxiety that was due to a common environ-mental effect was shared with earlier sadness, so the unique C could be dropped. In contrast to the results with overanxious, a shared genetic influence did not account for any of the association between temperament and separa-tion anxiety.
A Tractable Set of Biological Substrates for
Studying Inhibition in Young Children
Biological underpinnings of fear and anxiety are the subject of systematic research programs, several of which are represented in this issue. We shall not review the relevant neural circuitry, its functioning, or the strengths and limitations of that research here (see Charney et al 1998a, 1998b; LeDoux 1996). Three key aspects of the functioning of fear circuitry are 1) levels of cortisol as an index of the HPA axis functioning, 2) activation of the prefrontal cortex (and particularly asymmetry of that activation), and 3) activation and inhibition of the amyg-dala. The first two of these aspects are amenable to measurement in children.
We begin with studies that have collected saliva sam-ples for cortisol assays. Building on the work of Gunnar (in press) in human children and Kalin et al (1998) in rhesus macaques, we have measured cortisol levels in two samples for whom anxiety-related measures were also available. In a sample of children followed through nine waves of data collection from pregnancy through the first grade, we and our research collaborators studied approx-imately 500 families and their offspring living in the Milwaukee and Madison, Wisconsin, areas. Observational data, self-report data, videotaped behaviors from standard-ized emotion-eliciting episodes, and biological measures that could be collected in the field were obtained from the index children, their siblings, parents, and teachers.
With data from this large sample, we asked whether baseline cortisol would predict socioemotional adjustment after one of the most important and potentially stressful transitions of childhood, starting school (Smider et al 2000). Cortisol values were the mean of samples taken at approximately the same times on three afternoons in the home. After applying quality-control screens, useable cortisol data were available for 172 children. Kindergarten teachers answered specific questions about how the chil-dren acted in school around peers, how the chilchil-dren felt about school, and so forth. Children rated by their kinder-garten teachers as high on the correlated measures of internalizing behaviors and social inhibition had
signifi-cantly higher baseline cortisol nearly 1.5 years earlier, compared with children exhibiting few such maladaptive behaviors. That is, earlier baseline cortisol predicted sig-nificant (p , .01) increments of the variance in these teacher ratings (5– 6% more variance), after regressing out the effects of child gender, medication effects, and time of day effects (Smider et al 2000). We also collected saliva samples from parents in this sample. Mother– child (r 5
.42) and father– child (r 5 .33) resemblance for the measures of basal cortisol was high, given that the ceiling for these correlations should be .50 if basal cortisol were a completely heritable trait with no transmissible environ-mental effect. Of course, basal cortisol is unlikely to be completely heritable. Also, jointly experienced stressors might have affected the similarity of cortisol levels of some parent– offspring dyads. Nevertheless, the observed parent– offspring similarity at least alerts us that the cortisol– behavior link in this study might involve the genes.
The measure of prefrontal activation asymmetry used in much of our work is derived from extensive work by Davidson and his colleagues showing that individual differences in electroencephalographic (EEG) measures of prefrontal asymmetry are stable over time, correlated with positron emission tomographic measures of regional glu-cose metabolism from lateral prefrontal cortex, and predict various psychologic and biological measures of affect (Davidson 1998; Davidson and Irwin 1999). Subjects with greater relative right-sided prefrontal activation show ele-vated levels of dispositional negative affect, greater be-havioral inhibition (at least in children), and higher levels of cortisol. Stable individual differences in prefrontal asymmetry are also present in rhesus monkeys and corre-lated with certain fearful behaviors (Davidson and Fox 1989; Kalin et al 1998).
more distress vocalizations (r5 2.38, p5.06), and more bodily and facial sadness combined (r5 2.40, p,.05). Reactive cortisol was associated with vocal distress (cry-ing and its milder variants) dur(cry-ing this stranger approach. These data were derived from a sample comprising one twin from each pair and were replicated in a sample comprising the other twin from each pair (Buss et al 1997). Other investigators have implicated prefrontal cortex as an important level of the circuit that regulates the hypo-thalmic-pituitary-adrenal axis (Diorio et al 1993), and thus we attempted to demonstrate associations between pre-frontal activation and cortisol measures in these 6-month-olds. Higher baseline cortisol and EEG asymmetry (more right frontal activation) were correlated at .55. Thus, our two endophenotypic measures were correlated with each other and with several behavioral measures even by age 6 months, suggesting an early developmental organization of the relevant systems.
Genetic analyses have not yet been conducted, but they should eventually inform us about the common underpin-nings of these physiologic and behavioral measures.
Future Directions
Research aimed at integrating genetic, neuroscience, and behavioral findings now deserves a high priority. Given the nexus of associations among genetic factors, asymmet-ric frontal activation, reactive and basal cortisol levels (and other biological measures not treated in this paper), early temperamental traits, and later anxiety symptoms, multivariate genetic approaches are clearly needed to understand their common roots. Even at the behavioral level, these methods can elucidate the bases of the asso-ciations between temperament (e.g., negative affectivity, behavioral inhibition, fearfulness) and specific types of dysfunctional anxiety, as well as comorbidity. Candidate genes (e.g., Heils et al 1996; Lesch et al 1996) are typically identified in samples where only a subset of factors in this nexus are measured (such as a diagnosis of anxiety disorder); however, such candidate genes should be tested for contributions to presumed behavioral sub-strates such as temperament and to biological endopheno-types. One implication is that the next generation of studies should “overmeasure” the anxiety disorder pheno-type, that is, measure behavior and biology well beyond the diagnostic criteria. This overmeasurement would allow refinement of nosology, including identification of genet-ically more homogeneous subtypes. A thorough analysis of multiple, specific anxiety-relevant temperamental traits, along with multiple, specific patterns of dysfunctional anxiety, would have important implications for early identification and assessment of risk. More generally, a deeper understanding of childhood anxiety disorders is
likely to emerge synergistically with a more searching analysis of the typical patterns of the development of affective style.
Research from our laboratory reported or reviewed in this article was supported by the following National Institute of Mental Health Grants: No. R01-MH59785 (HHG, P.I.), No. R01-MH 50560 (HHG, P.I.), No. R01-MH44340 (M. Essex, P.I.), No. P50-MH52354 (R. Davidson, Center Director), and No. F31-MH11747 (HHG, P.I.). Research also was supported by the University of Wisconsin Graduate School.
The authors appreciate the collaboration of Richard Davidson, Ned Kalin, Marilyn Essex, Kristin Buss, Nazan Aksan, Nancy Smider, and Joan Bihun.
Aspects of this work were presented at the conference “Genetics and Brain Function: Implications for the Treatment of Anxiety,” March 22–23, 2000, Washington, DC. The conference was jointly sponsored by the Anxiety Disorders Association of America (ADAA), the ADAA Scientific Advisory Board, and the National Institute of Mental Health.
References
Ablow JC, Measelle JR, Kraemer HC, Harrington R, Luby J, Smider N, et al (1999): The MacArthur three-city outcome study: Evaluating multi-informant measures of young chil-dren’s symptomatology. J Am Acad Child Adolesc Psychiatry 38:1580 –1590.
Achenbach TM, Edelbrock C (1981): Behavioral problems and competencies reported by parents of normal and disturbed children aged 4 through 16. Monogr Soc Res Child Dev 46(1):1– 82.
Anxiety Disorders Association of America (2000): Conference
on Treating Anxiety Disorders in Youth: Current Problems and Future Solutions. Washington, DC: ADAA.
Bates JE (1989): Applications of temperament concepts. In: Kohnstamm GA, Bates JE, Rothbart MK, editors.
Tempera-ment in Childhood. Chichester, UK: Wiley, 321–355.
Biederman J, Rosenbaum JF, Bolduc-Murphy EA, Faraone SV, Chaloff J, Hirshfeld MA, Kagan J (1993): A 3-year follow-up of children with and without behavioral inhibition. J Am Acad
Child Adolesc Psychiatry 32:814 – 821.
Boyle MH, Offord DR, Racine YA, Szatmari P, Sanford M, Fleming JE (1997): Adequacy of interviews versus checklists for classifying childhood psychiatric disorder based on parent reports. Arch Gen Psychiatry 54:793–799.
Buss KA, Dolski I, Malmstadt J, Davidson RJ, Goldsmith HH, Kalin NA (1997): EEG asymmetry, salivary cortisol, and affect expression: An infant twin study. Psychophysiology 34(suppl 1):S25.
Charney DS, Grillon C, Bremner JD (1998a): The neurobiolog-ical basis of anxiety and fear: Circuits, mechanisms, and neurochemical interactions (part I). Neuroscientist 4:35– 44. Charney DS, Grillon C, Bremner JD (1998b): The
neurobiolog-ical basis of anxiety and fear: Circuits, mechanisms, and neurochemical interactions (part II). Neuroscientist 4:122– 132.
Davidson RJ (1998): Affective style and affective disorders: Perspectives from affective neuroscience. Cogn Emotion 12:307–330.
Davidson RJ, Fox NA (1989): Frontal brain asymmetry predicts infants’ response to maternal separation. J Abnorm Psychol 98:127–131.
Davidson RJ, Irwin W (1999): The functional neuroanatomy of emotion and affective style. Trends Cogn Sci 3:11–21. DeFries JC, Fulker DW (1985): Multiple regression analysis of
twin data. Behav Genet 15: 467– 473.
DiLalla LF, Kagan J, Reznick JS (1994): Genetic etiology of behavioral inhibition among 2-year-old children. Infant
Be-hav Dev 17:405– 412.
Diorio D, Viau V, Meany MJ (1993): The role of the medial prefrontal cortex (cingulate gyrus) in the regulation of the hypothalamic-pituitary-adrenal responses to stress. J
Neuro-sci 13:3839 –3847.
Fox NA (1994): The development of emotion regulation: Bio-logical and behavioral considerations. Monogr Soc Res Child
Dev 59(2–3).
Gjone H, Stevenson J, Sundet JM, Eilertsen DE (1996): Changes in heritability across increasing levels of behavior problems in young twins. Behav Genet 26:419 – 426.
Goldsmith HH (1996): Studying temperament via construction of the toddler behavior assessment questionnaire. Child Dev 67:218 –235.
Goldsmith HH, Buss AH, Plomin R, Rothbart MK, Thomas A, Chess S, et al (1987): Roundtable: What is temperament? Four approaches. Child Dev 58:505–529.
Goldsmith HH, Buss KA, Lemery KS (1997a): Toddler and childhood temperament: Expanded content, stronger genetic evidence, new evidence for the importance of environment.
Dev Psychol 33:891–905.
Goldsmith HH, Gottesman II, Lemery KS (1997b): Epigenetic approaches to developmental psychopathology. Dev
Psycho-pathol 9:365–387.
Goldsmith HH, Lemery KS, Buss KA, Campos JJ (1999): Genetic analyses of focal aspects of infant temperament. Dev
Psychol 35:972–985.
Goldsmith HH, Rieser-Danner L (1990): Assessing early tem-perament. In: Reynolds CR, Kamphaus R, editors. Handbook
of Psychological and Educational Assessment of Children, Vol. 2. Personality, Behavior, and Context. New York:
Guilford, 345–378.
Guerin DW, Gottfried AW (1994): Developmental stability and change in parent reports of temperament: A ten-year longitu-dinal investigation from infancy through preadolescence.
Merrill-Palmer Q 40:334 –355.
Gunnar MR (in press): The role of glucocorticoids in anxiety disorders: A critical analysis. In: Vasey MW, Dabbs MR, editors. The Developmental Psychopathology of Anxiety. New York: Oxford University Press.
Hayward C, Killen JD, Kraemer HC, Taylor CB (1998): Linking self-reported childhood behavioral inhibition to adolescent social phobia. J Am Acad Child Adolesc Psychiatry 37:1308 – 1316.
Heils A, Teufel A, Petri S, Stro¨ber G, Riederer P, Bengel D, Lesch KP (1996): Allelic variation of human serotonin transporter gene expression. J Neurochem 66:2621–2624.
Kagan J, Reznick JS, Snidman N (1988): Biological bases of childhood shyness. Science 240:167–171.
Kagan J, Snidman N, Arcus D (1998): Childhood derivatives of high and low reactivity in infancy. Child Dev 69:1483–1493. Kagan J, Snidman N, Zentner M, Peterson E (1999): Infant temperament and anxious symptoms in school age children.
Dev Psychopathol 11:209 –224.
Kalin NH, Larson C, Shelton SE, Davidson RJ (1998): Asym-metric frontal brain activity, cortisol, and behavior associated with fearful temperaments in rhesus monkey. Behav Neurosci 112:1–7.
LeDoux J (1996): The Emotional Brain. New York: Simon & Schuster.
Lemery KS (1999): Exploring the etiology of the relationship between temperament and behavior problems in children. Unpublished doctoral dissertation, University of Wisconsin, Madison.
Lemery KS, Essex M, Smider N (in submission): A multi-informant longitudinal study of measurement confounding between temperament and behavior problems in childhood. Lesch K-P, Bengel D, Heils A, Sabol SZ, Greenberg BD, Petri S,
et al (1996): Association of anxiety-related traits with a polymorphism in the serotonin transporter gene regulatory region. Science 274:1527–1531.
Matheny AP (1989): Children’s behavioral inhibition over age and across situations: Genetic similarity for a trait during change. J Pers 57:215–235.
McDevitt SC (1986): Continuity and discontinuity of tempera-ment in infancy and early childhood: A psychometric per-spective. In: Plomin R, Dunn J, editors. The Study of
Temperament: Changes, Continuities, and Challenges.
Hills-dale, NJ: Erlbaum, 27–38.
McGue M, Bouchard TJ Jr (1984): Adjustment of twin data for the effects of age and sex. Behav Genet 14:325–343. Merikangas KR, Swendsen JD, Preisig MA, Chazan RZ (1998):
Psychopathology and temperament in parents and offspring: Results of a family study. J Affect Disord 51:63–74. Offord DR, Boyle MH, Racine Y (1991): Ontario Child Health
Study: Correlates of disorder. In: Chess S, Hertzig ME, editors. Annual Progress in Child Psychiatry and Child
Development. New York: Brunner-Mazel, 194 –204.
Pettit GS, Bates JE (1984): Continuity of individual differences in the mother-infant relationship from six to thirteen months.
Child Dev 55:729 –739.
Pfeifer M, Goldsmith HH, Davidson RJ, Rickman M (in submis-sion): Elaborating and circumscribing the construct of early behavioral inhibition.
Plomin R, Emde RN, Braungart JM, Campos JJ, Corley R, Fulker DW, et al (1993): Genetic change and continuity from fourteen to twenty months: The MacArthur Longitudinal Twin Study. Child Dev 64:1354 –1376.
Robinson JL, Kagan J, Reznick JS, Corley R (1992): The heritability of inhibited and uninhibited behavior: A twin study. Dev Psychol 28:1030 –1037.
Rothbart MK (1981): Measurement of temperament in infancy.
Child Dev 52:569 –578.
Rothbart MK, Bates J (1998): Temperament. In: Eisenberg N, editor. Handbook of Child Psychology, Vol. 3. Social,
Emo-tional, and Personality Development. New York: Wiley,
105–176.
Sanson AV, Pedlow R, Cann W, Prior M, Oberklaid F (1996): Shyness ratings: Stability and correlates in early childhood.
Int J Behav Dev 19:705–724.
Schmitz S, Fulker DW, Plomin R, Zahn-Waxler C, Emde RN, DeFries JC (1999): Temperament and problem behavior during early childhood. Int J Behav Dev 23:333–355. Schwartz CE, Snidman N, Kagan J (1999): Adolescent social
anxiety as an outcome of inhibited temperament in childhood.
J Am Acad Child Adolesc Psychiatry 38:1008 –1014.
Smider NA, Essex MJ, Kalin NH, Buss KA, Davidson RJ, Goldsmith HH, Klein MH (in submission): Salivary cortisol as a predictor of internalizing and exernalizing behaviors during kindergarten: A prospective study.
Stevenson J, Batten N, Cherner M (1992): Fears and fearfulness
in children and adolescents: A genetic analysis of twin data.
J Child Psychol Psychiatry 33:977–985.
Tanaka JS (1993): Multifaceted conceptions of fit in structural equation models. In: Bollen KA, Long JS, editors. Testing
Structural Equation Models. Newbury Park, CA: Sage, 10 –
39.
Thapar A, McGuffin P (1995): Are anxiety symptoms in child-hood heritable? J Child Psychol Psychiatry 36:439 – 447. Topolski TD, Hewitt JK, Eaves LJ, Silberg JL, Meyer JM, Rutter
M, et al (1997): Genetic and environmental influences on child reports of manifest anxiety and symptoms of separation anxiety and overanxious disorders: A community-based twin study. Behav Genet 27:15–28.
Warren SL, Schmitz S, Emde RN (1999): Behavioral genetic analyses of self-reported anxiety at 7 years of age. J Am Acad
Child Adolesc Psychiatry 38:1403–1408.
Wilson RS, Matheny AP Jr (1986): Behavior-genetics research in infant temperament: The Louisville Twin Study. In: Plomin R, Dunn J, editors. The Study of Temperament: Changes,