www.elsevier.nlrlocateraqua-online
Muscle activity at slaughter: I. Changes in flesh
colour and gaping in rainbow trout
D.H.F. Robb
), S.C. Kestin, P.D. Warriss
Department of Clinical Veterinary Science, UniÕersity of Bristol, Langford, Bristol, BS40 5DU, UK
Accepted 23 July 1999
Abstract
High activity at slaughter is known to cause changes to the muscle biochemistry in terms of
Ž .
muscle pH and adenosine triphosphate ATP content. This experiment looked at how the muscle activity may affect the flesh quality of fish. Electro-stimulation of a carcass immediately after death can be used as a model for high levels of muscle activity prior to death. This experiment
Ž .
showed that electro-stimulation of 1.5 kg rainbow trout Oncorhynchus mykiss immediately after slaughter not only resulted in short times to rigor, but that the flesh colour was affected. The flesh
Ž . Ž . Ž .
became significantly lighter P-0.001 , less red P-0.001 and had higher chroma P-0.001
w
as measured by the CIE CIE, 1976. 18th Session, London, U.K., Sept. 1975. CIE Publication 36,
x U U U Ž .
Paris, France. L a b system, and the Roche colour card score was lowered P-0.01
compared to flesh from fish that had shown very low levels of activity before slaughter and were not electro-stimulated after death. The fillets were also more susceptible to gaping following the
Ž .
post-mortem stimulation P-0.001 . This indicated that electro-stimulation after death reduces the quality of the flesh.q2000 Elsevier Science B.V. All rights reserved.
Keywords: Salmonids; Slaughter; Flesh quality; Colour; Rigor
1. Introduction
There is increasing evidence that activity at slaughter affects the flesh quality of fish. Greater activity immediately prior to death results in a more rapid drop in muscle pH as
Ž the white muscle metabolism is predominantly anaerobic, producing lactic acid Lowe et
.
al., 1993 . Greater activity also results in shorter times to the onset of rigor. The strength
)Corresponding author. Tel.:q44-117-9289312; fax:q44-117-9289324; E-mail: [email protected]
0044-8486r00r$ - see front matterq2000 Elsevier Science B.V. All rights reserved. Ž .
of the muscle is affected, the muscle breaking more easily under strain after increased
Ž .
activity Jerret et al., 1996 .
Anecdotal evidence showed that there were differences in the number of fish rejected
Ž .
for poor colour from populations of Atlantic salmon Salmo salar that were harvested over several days. The flesh of all fish should have had approximately the same levels of
Ž
the pigment astaxanthin, but the Roche colour scores of some were lower A. Brown, .
pers commun., Norsk Hydro . The fish with the poor colour were harvested under the most severe conditions and could have been showing higher levels of activity prior to slaughter. It has also been observed that some fish show a greater propensity to muscle
Ž gaping than others. Gaping occurs when the tissues between the muscle blocks the
.
myosepta break and the blocks become separated. Commercially it is a problem as it makes the processing of the flesh much more difficult.
Variation in visual colour scores and colourimetric measurements between samples
Ž .
with the same pigment concentration was also reported by Little et al. 1979 . Using
Ž . U U U
CIELab CIE, 1976 , CIELUV and Hunter L a b measurements it was concluded that differences in colour measurements between samples were caused by differences in light scattering and absorption within the flesh. However, the cause of these differences was not suggested by the authors.
Previous work on the effects of slaughter treatments showed an influence on muscle
Ž . Ž .
pH Lowe et al., 1993 and the times to rigor Azam et al., 1990 . The development of rigor can be followed by measuring the flexibility of the fish. As the fish enter rigor they are able to bend less and as rigor resolves so they are more flexible. However, anecdotal evidence indicates that there are also potential effects of muscle activity on gaping and muscle colouration. Muscle gaping can be scored subjectively according to the amount of gaping and this system is used regularly commercially as it is the most convenient
Ž .
method available A. Dingwall, pers. commun., Pinneys of Scotland . Muscle coloura-Ž
tion of salmonids can be scored subjectively using the Roche colour card NS9402,
. Ž . U U U Ž
1994 and objectively using the CIE 1976 or Hunter L a b systems Skrede and .
Storebakken, 1986; Christiansen et al., 1995 . The Roche card is commonly used commercially, but for research purposes the use of a tristimulus meter is preferable, giving a repeatable methodology. The measurements of LU, aU and bU give information
Ž . Ž .
on the lightness, angle of hue colour and chroma of the flesh Warriss, 1996 . These can be used to determine differences between treatment groups or to follow changes in these parameters during storage.
Electro-stimulation of the flesh post-mortem with low frequency, pulsed direct current results in the rapid and repeated contraction of the muscle and can be used as a
Ž .
model for very high muscle activity levels Jerret et al., 1996; Fletcher et al., 1997 . This has advantages over using natural activity in that it is repeatable between fish. It is also likely that the degree of muscle activity produced is greater than normal activity levels would result in, so causing maximum difference between the treatments.
This experiment aimed to investigate the extent that activity affects flesh quality by using two extremes of muscle activity. AQUI-Sw
, an anaesthetic used commercially on fish farms in Australia and New Zealand, anaesthetises fish slowly and was used to
Ž
produce a ‘no activity’ or ‘rested’ group Jerret et al., 1996; Robb and Kestin, unpubl. .
Ž .
for rainbow trout Keene et al., 1998 , without causing swimming reactions. A model muscle high activity group was generated using electro-stimulation post-mortem. The
Ž .
fish used were rainbow trout Oncorhynchus mykiss , as they are good models for the more valuable salmon flesh, having the same muscle pigments and similar constraints of commercial flesh quality. They were used preferentially to salmon for this experiment as they were easier to obtain and hold at the experimental facilities at Langford.
2. Materials and methods
Forty fish were selected at random from a population of one thousand rainbow trout of average weight 1.5 kg. The fish had been raised together from hatching. None of the
Ž
fish selected were maturing and all had been fed a commercial trout diet Vextra Classic, .
EWOS, U.K. containing the pigment astaxanthin at 50 mgrkg of diet. All fish were placed in a 3-m diameter tank with a 1-m depth of water, fed by spring water at 168C and oxygenated to a level of 85% saturation. The fish were left to recover, unfed, from the handling for 3 days.
The water level in the tank was slowly reduced and the water supply shut off. The
w Ž .
anaesthetic AQUI-S AQUI-S New Zealand, New Zealand was slowly added to achieve a concentration of 17 ppm. This resulted in deep anaesthesia of the fish after about 30 min. The fish were then netted out by hand and killed by a blow to the head.
Ž Twenty fish were chosen at random and were electro-stimulated immediately within
.
1 min after being killed in order to model a high level of muscle activity. This was carried out by inserting a needle dorso-ventrally through the flesh just behind the head, passing close to the backbone. The needle was attached to an electrode connected to a
Ž .
control box J.S. Engineering, U.K. . A second electrode from the box was attached by a crocodile clip to the tip of the caudal peduncle, just at the end of the spine. Each fish received 94 V pulsed direct current at 14.3 Hz, causing muscular contractions along the length of the fish. After 2 min of stimulation, the fish were disconnected from the power. The remaining twenty fish were left unstimulated, resulting in the ‘rested’ group. Ten fish from each treatment group were filleted, individually tagged and a series of measurements were made on the area between the dorsal fin and the midline. The surface of the fillet was wiped with absorbent paper and then the core temperature was
Ž .
determined using a temperature probe Whatman, U.K. . The pH was measured using a
Ž .
PHM8 Portable pH Meter Radiometer, Denmark with a spear combination pH
Ž .
electrode Mettler Toledo . The colour of the fillets was determined using a CR-200
Ž .
Chromameter Minolta, U.K. with a pulsed xenon arc lamp giving diffuse illumination,
Ž .
with 08viewing angle over an 8-mm diameter area. This resulted in CIE 1976 values
Ž U. Ž y1 Ž U U.. Ž Ž U2 U2..
for lightness L , the angle of hue tan b ra and chroma 6 a qb . After measuring, the fillets were placed one layer deep on a sheet of plastic over a layer of ice in polystyrene boxes. A second layer of plastic and ice was placed above the fillets and the boxes stored in a dark chiller at 48C. Further measurements were carried out on the fillets after approximately 1, 2, 6, 8, 22, 33, 45, 57, 69 and 75 h.
At the end of the storage period the fillets were measured a final time and were Ž
.
Switzerland by four assessors. The fillets were then drawn, skin down, over a right angle using a constant force. This simulated rough handling and potentially caused the appearance of gaping along the fillet, which was then scored subjectively by four judges,
Ž . Ž . Ž
using a four point scale: 0 no gaping , 1 one or two breaks , 2 four or more small
. Ž .
breaks , or 3 large breaks .
The remaining 10 fish from each group were used to follow the progression of rigor.
Ž .
A modified version of the apparatus used by Azam et al. 1990 was set up. The fish were placed on a flat table so that the body behind the posterior end of the dorsal fin was hanging over the edge, unsupported. The angle between the horizontal and the line from the corner of the table to the tip of the tail was then measured to the nearest 58 Žtermed the angle of droop . The fish were then turned over and the measurement. repeated for the other side of the fish. The fish were measured at 0, 4, 8, 12, 20, 24, 34, 48, 56, 72 and 96 h after slaughter. In between measurements the fish were placed on ice in polystyrene boxes which were stored in a chiller at 48C.
The significance of the differences between the groups in temperature, pH, lightness, angle of hue, chroma and rigor development were tested at each time post-slaughter
Ž .
using a repeated measures analysis of variance ANOVA . In order to determine the rigor changes, the mean angle of droop for each fish was calculated from the two angles measured and then the ANOVA performed. This compensated for the fish setting at an angle during rigor. The mean Roche colour card scores for each fish were compared using Student’s t-test. A Mann–Whitney U-test was applied to the non-parametric data from the gaping scores.
3. Results
During anaesthesia the fish showed very little movement. Some slow swimming around the tank was observed, but many fish just held their position until unable to move. At no point were any rapid movements observed. During the electro-stimulation, however, rapid contractions of the muscles resulted in a large degree of movement of the whole fish. By the end of the 2-min stimulation the gross movements had almost stopped, but the muscles were still contracting as the fish obviously relaxed after the cessation of stimulation.
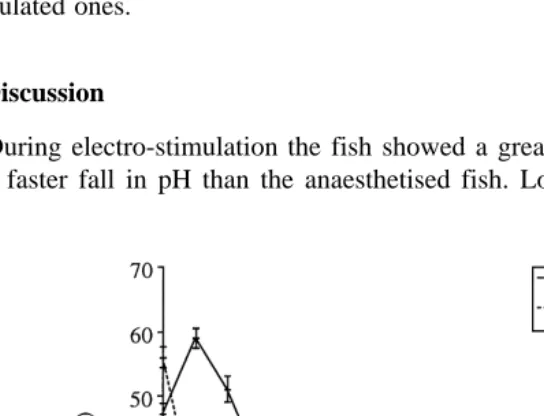
Ž .
There was no difference in temperature between the groups at any point P)0.05 . However, the pH differed significantly between the groups until 56 h after slaughter ŽP-0.01 . Fig. 1 shows the changes in pH with time after slaughter. The anaesthetised. fish showed a much slower rate of fall in pH, starting at 7.8"0.31 and dropping below pH 7.0 after 22 h. By 45 h after slaughter the pH was below 6.6. In contrast, the electro-stimulated fish showed a very rapid drop in pH. Immediately after slaughter the pH was 6.7"0.03 and after 2.5 h it had dropped below 6.6. The final pH of the flesh of the anaesthetised group was significantly higher than that of the electro-stimulated group ŽP-0.001 although the difference between the means was only 0.07..
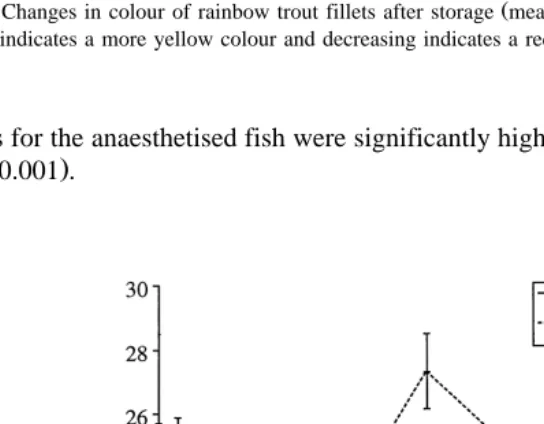
There were differences in flesh colour between the anaesthetised and the
electro-Ž . Ž . Ž
stimulated fish as measured by lightness Fig. 2 , angle of hue Fig. 3 and chroma Fig. .
Ž .
Fig. 1. Changes in muscle pH of rainbow trout fillets with time after slaughter mean values"S.E., ns10 .
Ž U. Ž .
significantly darker lower L and redder lower angle of hue flesh, which had lower
Ž .
chroma than the electro-stimulated fish P-0.01 . This was confirmed by the
differ-Ž .
ence in the Roche colour card scores at the end of the storage period Table 1 . The
Ž .
Ž .
Fig. 3. Changes in colour of rainbow trout fillets after storage mean values"S.E., ns10 . Increasing angle of hue indicates a more yellow colour and decreasing indicates a redder colour in the range 0–908.
scores for the anaesthetised fish were significantly higher than the electro-stimulated fish ŽP-0.001 ..
Ž .
Table 1
Mean Roche colour card and gaping scores for flesh of rainbow trout at the end of the 75-h storage period
Ž"S.E.M..
A higher gaping score indicates greater gaping of the flesh. A higher colour card score indicates a darker red fillet colour.
Anaesthetised Electro-stimulated Significance of difference between means
Roche colour card score 15.0"0.18 14.2"0.23 P-0.01
Gaping score 1.1"0.21 2.3"0.25 P-0.01
The degree of gaping of the flesh was significantly affected by the treatment ŽP-0.001 , with the anaesthetised fish showing less gaping after simulated rough.
Ž . Ž .
handling Table 1 . The time to the onset of rigor was delayed in these fish Fig. 5 . At each time there was a significant difference between the two groups, with the
anaes-Ž .
thetised fish showing resolution of rigor later than the electro-stimulated fish P-0.001 . They also showed a greater angle of droop when in full rigor than the electro-stimulated
Ž .
group P-0.001 , which indicated that these fish were more flexible than the electro-stimulated ones.
4. Discussion
During electro-stimulation the fish showed a great deal of muscle activity, resulting
Ž .
in a faster fall in pH than the anaesthetised fish. Lowe et al. 1993 showed that fish
Ž .
exhibiting more activity at slaughter had a faster drop in pH. The results of this paper
Ž . Ž .
support the views of Jerret et al. 1996 and Fletcher et al. 1997 that electro-stimula-tion of the whole body is a suitable model for high levels of activity.
The rapid onset of rigor in the electro-stimulated fish would have occurred when the
Ž .
fish muscles ceased to resynthesise adenosine triphosphate ATP . The good relationship between measured ATP concentration and rigor index in fish has been reviewed by
Ž .
Erikson 1997 . The onset of rigor will occur when the concentration of ATP in the cells is exhausted. Increasing exercise immediately prior to death, or immediately post-mortem,
Ž
depletes energy reserves in the muscles Lowe et al., 1993; Berg et al., 1997; Sigholt et .
al., 1997 . Thus, a faster onset of rigor is observed in active or electro-stimulated fish. The technique used to follow rigor involved repeated measures on the same fish, as in
Ž .
Azam et al. 1990 . This will have affected the muscle properties of the fish as the bending will have put stresses on the muscles. However, the results indicate that the fish
Ž
followed a normal pattern of onset and resolution of rigor as observed by Berg et al., .
1997, for example . Further, the difference between the two treatment groups was highly significant, despite the low numbers of fish investigated. This suggests that although the method used obviously affected the muscle it was still valid for following rigor development.
The electro-stimulated fish became more rigid when in rigor than the ‘rested’ fish. This was shown by them having a lower minimum angle of droop when in maximum
Ž .
rigor. Berg et al. 1997 suggested that this effect may be due to cells in ‘rested’ fish entering rigor at different times, resulting in the more flexible carcass. They supported this hypothesis by showing that when at peak rigor, fish that had shown low levels of activity had much higher ATP concentrations than fish that had been active, indicating that not all of the cells entered rigor at the same time in the low activity group.
The gaping scores show that the electro-stimulated fish were more susceptible to gaping within the fillets than the ‘rested’ fish. This could be caused by the electro-stimulated fish having the greater rigidity when in rigor discussed above, resulting in a
Ž .
greater force on the muscles while in rigor. However, Jerret et al. 1996 also showed that, after rigor, flesh from active fish was more susceptible to the myosepta breaking under strain than in flesh from anaesthetised fish. The gaping scores from this experi-ment show that this difference exists on a whole fillet scale. The change in flesh strength
Ž .
could reflect changes caused to the protein structure of the muscle Sigholt et al., 1997 . The idea that changes in the structure of the protein in the muscle occur following the increase in muscle activity is supported by the changes in the colour of the muscle found in this experiment.
Changes in muscle colour resulting in the lightening and yellowing of the flesh
Ž U .
increased L and angle of hue, respectively and increased chroma have been observed Ž
in red meat animals subjected to high levels of activity prior to slaughter Warriss, .
1996 . These changes are caused by a decreased level of soluble muscle proteins in the flesh of stressed animals compared with the level in unstressed animals. The proteins denature, becoming insoluble and causing a loss of water from the flesh, and resulting in changes in the reflection of light from the surface; hence the changes in lightness, hue and chroma. The higher level of insoluble protein is caused by the rapid drop in pH
Ž .
The cause of the changes in lightness, hue and chroma values throughout the experiment are not known, which is a reflection on the scarcity of information in this area of research. The drop in lightness and angle of hue in the first 4 h and the changes in chroma in the first day post-mortem observed in both groups appear to be a real effect, but later fluctuations may just be random, as most of these changes are smaller than the standard errors of the means. The differences in lightness, hue and chroma in red meat have been shown to be caused by the increased level of insoluble muscle protein in the flesh of active animals post-mortem relative to that in rested ones ŽWarriss, 1996 . The differences in lightness, hue and chroma between the electro-. stimulated and ‘rested’ fish flesh are very similar to those observed in red meat, leading to the hypothesis that the changes are caused by differences between the two groups in the levels of insoluble protein. Further work being carried out in our laboratory will determine whether there are indeed changes in the amount of soluble muscle protein in the flesh with the different levels of muscle activity.
References
Azam, K., Strachan, N.J.C., Mackie, I.M., Smith, J., Nesvadba, P., 1990. Effect of slaughter method on the
Ž .
progress of rigor of rainbow trout Salmo gairdneri as measured by an image processing system. Int. J. Food Sci. Technol. 25, 477–482.
Ž .
Berg, T., Erikson, U., Nordvedt, T.S., 1997. Rigor mortis assessment of Atlantic salmon Salmo salar and effects of stress. J. Food Sci. 62, 439–446.
Christiansen, R., Struksnaes, G., Esterman, A., Torrissen, O.J., 1995. Assessment of flesh colour in Atlantic salmon Salmo salar L. Aquacult. Res. 26, 311–321.
CIE, 1976. 18th Session, London, U.K., Sept. 1975. CIE Publication 36, Paris, France.
Ž .
Erikson, U., 1997. Muscle quality of Atlantic salmon Salmo salar as affected by handling stress. PhD Thesis, Norwegian University of Science and Technology, Trondheim, Norway.
Fletcher, G.C., Hallett, I.C., Jerrett, A.R., Holland, A.J., 1997. Changes in the fine structure of the myocommata–muscle fibre junction related to gaping in rested and exercised muscle from king salmon
ŽOncorhynchus tshawytscha . Lebensm.-Wiss. Technol. 30, 246–252..
Jerret, A.J., Stevens, J., Holland, A.J., 1996. Tensile properties of white muscle in rested and exhausted king
Ž .
salmon Oncorhynchus tshawytscha . J. Food Sci. 61, 527–532.
Keene, J.L., Noakes, D.L.G., Moccia, R.D., Soto, C.G., 1998. The efficacy of clove oil as an anaesthetic for
Ž .
rainbow trout Oncorhynchus mykiss Walbaum . Aquacult. Res. 29, 89–101.
Little, A.C., Martinsen, C., Sceurman, L., 1979. Color assessment of experimentally pigmented rainbow trout. Color Res. Appl. 4, 92–95.
Lowe, T.E., Ryder, J.M., Carragher, J.F., Wells, R.M.G., 1993. Flesh quality in snapper, Pagrus auratus, affected by capture stress. J. Food Sci., 58, 770–773, 796.
NS9402, 1994. Atlantic salmon. Colour and fat measurement. Norges Stanardiseringsforbund, Oslo, Norway. Translated from Norwegian.
Sigholt, T., Erikson, U., Rustad, T., Johansen, S., Nordtvedt, T., Seland, A., 1997. Handling stress and storage
Ž .
temperature affect meat quality of farm-raised Atlantic salmon Salmo salar . J. Food Sci. 62, 898–905. Skrede, G., Storebakken, T., 1986. Instrumental colour analysis of farmed and wild Atlantic salmon when raw,
baked and smoked. Aquaculture 53, 279–286.
Warriss, P.D., 1996. Instrumental Measurement of Colour. In: Taylor, S.A., Raimundo, A., Severini, M.,
Ž .
Smulders, F.J.M. Eds. , Meat Quality and Meat Packaging. ECCEAMST, Utrecht, The Netherlands, pp. 221–231.