Bio-ekologi Burung Beo
Burung yang mampu menirukan suara-suara yang didengarnya, terdiri dari beberapa ordo, salah satu diantaranya adalah Passeriformes. Burung-burung yang termasuk ordo Passeriformes, subordo Passeres merupakan burung penyanyi (Songbird) yang terdiri dari 52 famili. Namun demikian, hanya tiga spesies dari ordo ini yang mampu menirukan suara yaitu magpies (Corvus corax, Corvidae), mockingbirds (Mimus polyglottos, Mimidae) (Lampiran 4) dan burung beo (Gracula religiosa) yang termasuk famili Sturnidae (Rutgers & Norris 1977; Storer et al. 1979). Selain burung beo, burung nuri dan kakatua juga mampu menirukan suara yang didengarnya (Forshow 1977; Sudaryanti 1984), walaupun perbendaharaan kata yang diikuti tidak sebanyak burung beo. Hal ini merupakan daya tarik tersendiri bagi penggemar burung untuk menjadikan burung beo sebagai hewan kesayangan.
Taksonomi
Menurut Peters (1937) dan Monroe dan Sibley (1993) burung beo diklasifikasikan ke dalam : Kingdom : Animalia Filum : Chordata SubFilum : Vertebrata Kelas : Aves Ordo : Passeriformes SubOrdo : Passeres Famili : Sturnidae Genus : Gracula
Orang yang pertama kali memberi nama beo dengan nama latin Gracula religiosa adalah Linnaeus pada tahun 1758 (Peters 1937; Monroe dan Sibley 1993). Burung beo banyak disukai oleh masyarakat karena kepandaiannya di dalam menirukan suara yang didengarnya. Burung beo di Jawa dikenal dengan sebutan menco, sedangkan di Sumatera Utara dinamai tiung atau tiong (Prijono dan Waluyo 1996), di Pulau Nias dikenal dengan mageao (Sitepu 1997) dan dalam bahasa Inggris disebut Mynah (Campbell dan Lack 1985).
Morfologi
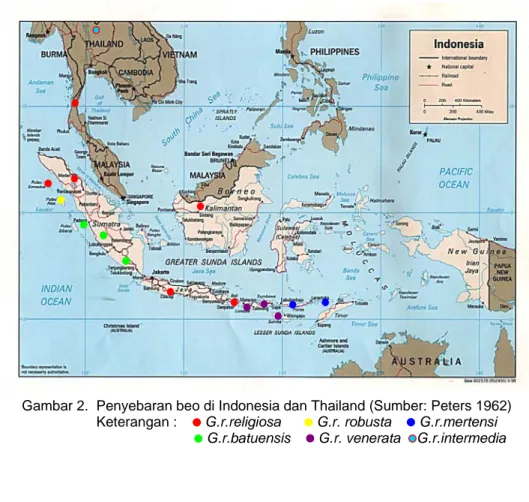
Berdasarkan cuping kuduknya, beo di Indonesia dibedakan atas lima subspesies (Peters 1962; Prijono dan Waluyo 1996, Gambar 1)yaitu: 1. G. r. religiosa Linnaeus 1758, mempunyai pangkal cuping kuduk
terpisah. Subspesies ini tersebar di Tanah Genting Kra Selatan, Sumatera, Simalue, Enggano, Pulau Dua, Bangka, Belitung, Kalimantan, Natuna, Karimata, Jawa, Bawean, Bali dan Kangean. Dalam dunia perdagangan burung dikenal dengan nama beo medan. 2. G. r. robusta Salvadori 1887, memiliki cuping kuduk bersatu antara kiri
dan kanan. Daerah sebarannya meliputi Nias, Pulau Babi, Tuangku dan Bangkaru dan dikenal dengan sebutan beo nias.
3. G. r. batuensis Finsch 1899, mempunyai ujung cuping kuduk membelok ke atas dan pangkalnya terpisah. Penyebarannya meliputi Tello, Siberut, Sipora, Pagi, Anambas, Tambelan dan Tioman, sering disebut beo lampung.
4. G. r. mertensi Rensch 1928, memiliki ujung cuping kuduk yang membelok ke atas, pangkalnya terpisah dan juga membelok ke atas. Pangkal cuping kuduk jika dilihat dari atas berbentuk seperti dua garis kuning memanjang ke arah paruh. Mempunyai daerah penyebaran di Flores, Pulau Pantai dan Alor.
5. G. r. venerata Bonaparte 1851, hampir sama dengan G. r. mertensi tapi ukurannya lebih kecil. Anak jenis beo ini tersebar di Sumbawa dan kepulauan Sunda Kecil.
Gambar 1. Beberapa spesimen beo yang diawetkan (Sumber : Prijono & Waluyo 1996 (a,c,d,e) dan dokumentasi pribadi (b))
a. Beo medan (G.r.religiosa) d. Beo nias (G.r.robusta) b. Beo kalimantan (G.religiosa spp) e. Beo flores (G.r.mertensi)
c. Beo lampung (G.r.batuensis)
Pada umumnya, beo mempunyai ukuran panjang tubuh sekitar 300-380 mm. Warna bulunya hitam mengkilap dengan refleksi ungu dan hijau (Direktorat PPA 1981; Prijono & Waluyo 1996). Beo dewasa mempunyai noda putih di ujung bulu primer, sedangkan beo muda memiliki bulu yang kurang mengkilap. Paruh berwarna kuning, oranye sampai merah dan di bawahnya terdapat cuping kuduk berwarna kuning (Maradjo 1977). Beo memiliki leher yang pendek, berpial kuning di bawah mata sampai belakang kepala (Direktorat PPA 1981). Kulmen (paruh beo berukuran 30 mm, ulnar (sayap) 15 mm, ekor 69 mm dan tarso-metatarsus (kaki) 30 mm (Direktorat PPA 1981). Karakter morfologi beo
(a) (b) (c)
thailand utara (n=206) memiliki ukuran bobot badan 189,37±15,94 g, panjang badan 185,20± 7,40 mm, panjang cuping kuduk 28,60±5,60 mm, panjang tarsus 32,80±2,10 mm dan panjang ekor 75,10±5,10 mm (Archawaranon & Mevatee 2002, Lampiran 2).
Telur burung beo memiliki lebar yang bervariasi antara 22,90-33,00 mm (Whistler 1949), 22,50-25,00 mm (Noerdjito 1987), sedangkan panjangnya berkisar antara 33,00-38,00 mm (Noerdjito 1987). Telur berwarna biru kehijauan atau kecoklatan (Whistler 1949), kebiruan (Direktorat PPA 1980a), berbintik biru kotor atau merah coklat (Direktorat PPA 1981). Menurut Hoogerwerf (1949) ukuran telur beo kalimantan 35,10x26,50 mm, telur beo dari daerah Sumatera berukuran 37x26,5 mm.
Habitat dan Penyebaran
Daerah penyebaran subspesies beo di dunia meliputi India, Cina, Daratan Asia Tenggara, Thailand, Palawan, Kalimantan, Sumatera, Jawa, Bali, Pulau Enggano, Bawean, Sumbawa, Flores, Alor dan Pulau Aru (MacKinnon 1993; Direktorat PPA 1980a). Beo dapat hidup di dataran rendah, bukit hutan primer, hutan rawa, hutan sekunder dan hutan persawahan sampai ketinggian 1000 m di atas permukaan laut (m dpl) (Marlee & Voous 1988; Noerdjito 1987). MacKinnon (1993) menyebutkan bahwa pada umumnya beo terdapat di beberapa tempat dataran rendah Sumatera (termasuk pulau-pulau di sekitarnya) dan Kalimantan (termasuk pulau-pulau di sekitarnya), sampai ketinggian 1000 m dpl, sedangkan di Pulau Jawa dan Bali, dahulu umum ditemukan di pinggir hutan, tetapi sekarang sudah jarang karena adanya penangkapan dan kerusakan habitat.
Gambar 2. Penyebaran beo di Indonesia dan Thailand (Sumber: Peters 1962) Keterangan : G.r.religiosa G.r. robusta G.r.mertensi G.r.batuensis G.r. venerata G.r.intermedia
Perilaku
Perilaku satwa dapat diartikan sebagai ekspresi suatu satwa yang disebabkan oleh semua faktor yang mempengaruhinya. Pola-pola perilaku yang ditampilkan oleh satwa dapat digunakan untuk menggolongkan satwa secara sistematis berdasarkan persamaan dan perbedaan pola perilaku pada masing-masing satwa tersebut (Drickamer & Vessey 1982). Menurut Pettingil (1969) dan Lehner (1996), faktor-faktor yang mempengaruhi perilaku satwa dinamakan rangsangan, stimuli atau agents, sedangkan aktivitas yang ditimbulkan dinamakan respon.
Pola perilaku merupakan segmen perilaku yang mempunyai fungsi adaptasi, untuk menjamin kelangsungan hidup dan keberadaan makhluk hidup pada suatu kawasan tertentu yang sesuai dengannya (Scott 1969). Perilaku terdiri atas campuran komponen-komponen yang diwariskan, dibawa dari lahir (insting) dan diperoleh dari lingkungan selama hidup yaitu berupa pengalaman (Tanudimadja dan Kusumamihardja 1985). Kebanyakan studi tentang perilaku satwa dimulai dengan penyusunan etogram, yaitu suatu uraian lengkap dari perilaku yang ditampilkan oleh satwa (Drickamer & Vessey 1982). Dalam mempelajari perilaku satwa pola-pola perilaku dapat dikelompokkan ke dalam sistem-sistem perilaku, yaitu sekumpulan pola perilaku yang memiliki fungsi yang sama (Lehner 1979), yaitu:
1. Perilaku makan (Ingestive behavior)
2. Perilaku mencari tempat bernaung (Shelter seeking) 3. Perilaku bertentangan (Agonistic behavior)
4. Perilaku seksual (Sexual behavior)
5. Perilaku memelihara (Epilemetic behavior)
6. Perilaku mendekati yang memelihara (Et-epilemetic behavior) 7. Perilaku meniru (Allelomimetic behavior)
8. Perilaku membuang kotoran (Eliminative behavior) 9. Perilaku memeriksa (Investigative/Exploratory behavior)
Perilaku Beo di Alam
Beo merupakan burung terbang yang tangkas dan sering terlihat terbang berkelompok (20-30 ekor), di hutan yang tidak terlalu lebat (Rutgers & Norris 1997). Mereka mempunyai kebiasaan bersuara pada pagi hari dan sore hari dengan suaranya yang bervariasi, kadang lembut dan riang, kadang serak dan kasar, bahkan dengan nada yang tinggi seperti seruling. Pada saat terbang tinggi sering dilakukan komunikasi dengan mengeluarkan suara-suara siulan keras atau seperti suara tangisan (Rutgers & Norris 1997).
Di alam burung beo nias memulai aktifitasnya pada pagi hari, dengan meninggalkan pohon sekitar pukul 05.00 untuk mencari makan di Hililego (Bukit Lelego, Kabupaten Nias) dan akan kembali lagi sekitar pukul 17.00 (Direktorat PPA 1981; Sitepu 1997). Makanan yang disukai beo adalah buah berdaging dari liana dan buah dari Dipterocarpaceae, yang mempunyai kulit buah tidak keras. Selain buah, beo menyukai serangga seperti capung, belalang, jangkrik dan telur semut (Direktorat PPA 1981, Prijono & Waluyo 1996).
MacKinnon & Phillips (1993) menyebutkan burung beo sering berada di pohon yang tinggi dan hidup berpasangan, kadang-kadang berkumpul dalam kelompok. Mereka mengunjungi daerah terbuka dan pohon berbuah, bercak putih pada sayapnya terlihat jelas saat terbang. Direktorat PPA (1980) menyebutkan mereka terbang bergerombol. Ketika terbang, warna kuning dari paruh serta kulit kepala yang tidak berbulu dan noda putih besar di sayapnya nampak jelas sekali. Sedangkan Noerdjito (1987) menambahkan bahwa beo nias sering berada di puncak-puncak pohon, kadang-kadang hinggap di tanah dan suka sekali bertengger di ranting pohon yang telah mati.
Perilaku Bersarang. Burung beo termasuk burung yang bersarang dalam lubang (hole nesting species). Di alam, beo membuat sarang di dalam lubang batang pohon yang sudah mati, atau bekas sarang burung rangkong juga pada tanaman pakis (yang terdapat di atas dahan) atau
dengan merebut sarang burung pelatuk. Burung beo hampir tidak memiliki persaingan dengan burung lainnya, kecuali dengan burung gagak. Kadang-kadang beo ditemukan bersama-sama dengan burung rangkong kecil (Direktorat PPA 1981). Beo di alam menyukai tidur di sarang dalam lubang pohon walaupun tidak sedang bertelur (Frisch 1986).
Perilaku burung beo yang bersarang dalam lubang pohon saat mencari sarang menurut Stokes dan Stokes (1990) adalah sebagai berikut: satu atau sepasang burung mendatangi sarang, memeriksa di luar dan kemudian masuk ke dalam sarang. Hal ini dapat dilakukan berhari-hari atau berminggu-minggu dengan mendatangi beberapa sarang sebelum memilih satu diantaranya. Pada hari berikutnya burung kembali dengan betinanya dan mencoba menarik perhatian betinanya.
Menurut Whistler (1949) musim kawin burung beo berlangsung pada bulan Pebruari sampai Mei, sedangkan di Malaysia pada bulan Desember sampai Mei (Noerdjito 1987). Di Sumatera, musim kawin terjadi pada bulan Pebruari dan di Kalimantan pada bulan Pebruari dan Maret.
Perilaku membuat sarang sering berhubungan dengan proses bertelur. Jumlah telur burung beo dalam satu kali musim bertelur ialah dua sampai dengan tiga butir dengan bentuk oval (Whistler 1949), tetapi ada yang berpendapat lain dalam satu sarang terdapat empat butir telur (Direktorat PPA 1980a). Pengeraman dilakukan secara bergantian antara beo jantan dan beo betina, dengan lama pengeraman selama 12 hari (Stokes & Stokes 1990), sedangkan menurut Frisch (1986) lama pengeraman 14 sampai 18 hari. Telur beo yang baru menetas akan mengeluarkan anakan beo yang tidak berbulu (mulai hari pertama sampai dengan hari ke tiga) dan mulai hari ke empat sudah terlihat adanya pertumbuhan bulu seperti yang terlihat pada Lampiran 3 (http://www.mynahbird.com/articles).
Lamanya anak burung berada di sarang sampai bisa terbang berkisar antara 20-23 hari (Frisch 1986). Anak burung memerlukan waktu 4-8 hari untuk belajar terbang, sehingga ada yang berpendapat bahwa
anak burung beo meninggalkan sarang kurang lebih berumur 31 hari (Stokes & Stokes 1990).
Perilaku Beo Nias di Penangkaran. Di penangkaran, beo nias lebih banyak mengalokasikan waktu hariannya untuk aktivitas menjelajah (49,55%). Interaksi sosial lebih banyak terjadi di siang hari (Muallivah 1997). Perilaku agonistik pada kelompok beo nias terjadi pada saat aktivitas makan, perebutan tempat bertengger dan aktivitas saling menyelisik bulu. Periode keaktifan interaksi sosial terjadi pada siang hari, dimana aktivitas yang lain cenderung menurun (Muallivah 1997).
Gambaran Darah Burung Beo
Darah terdiri atas dua komponen yaitu 30-40% komponen sel, yang terdiri atas eritrosit dan leukosit, serta 60-70% merupakan komponen plasma yang terdiri atas air, elektrolit, metabolit, zat makanan, protein dan hormon (Swenson 1977). Darah berfungsi sebagai : 1) media transportasi, antara lain mengangkut oksigen dari paru-paru ke sel-sel jaringan tubuh dan karbondioksida dari jaringan tubuh ke paru-paru, mengangkut bahan makanan dari usus ke sel-sel tubuh, mengangkut air, elektrolit, enzim dan hormon; 2) homeostasis, dan 3) pertahanan tubuh terhadap mikroorganisme (Swenson 1977; Widjajakusuma & Sikar 1986).
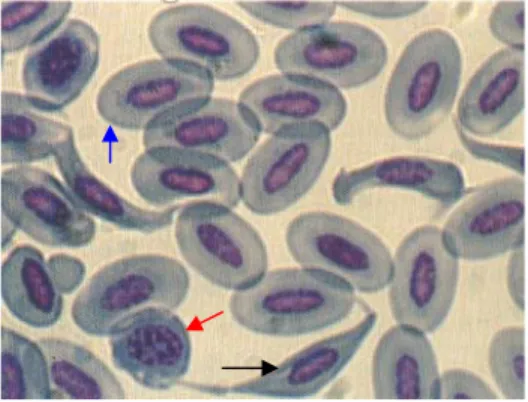
Eritrosit mempunyai fungsi pasif di dalam pembuluh darah, sedangkan leukosit mampu keluar dari pembuluh darah menuju jaringan (Brown 1989). Eritrosit bangsa burung berbentuk lonjong dan memiliki inti. Pada umumnya jumlah eritrosit dalam sirkulasi darah dipengaruhi oleh umur, jenis kelamin, hormon, keadaan hipoksia, faktor musim, waktu harian, suhu lingkungan, status reproduksi, nutrisi dan aktivitas (Sturkie 1976; Hodges 1977 & Mitruka et al. 1977). Eritrosit beo disajikan pada Gambar 4 (Hawkey & Dennet 1989).
Hemoglobin merupakan pigmen darah pada vertebrata yang terdiri atas protein kompleks terkonjugasi yang mengandung besi. Semua faktor
yang berpengaruh terhadap proses eritropoiesis dan jumlah eritrosit akan berpengaruh pula terhadap kadar hemoglobin darah. Hal ini terjadi karena hemoglobin terdapat di dalam eritrosit dan dibentuk saat proses pematangan eritrosit. Dengan demikian maka secara normal kadar hemoglobin di dalam darah berbanding lurus dengan jumlah eritrositnya. Pada burung jantan dewasa kadar hemoglobin lebih tinggi daripada burung betina dewasa (Schalm et al. 1975; Hodges 1977).
Nilai hematokrit atau Packed Cell Volume (PCV) adalah persentase volume eritrosit di dalam 100 ml darah. Pada hewan normal nilai hematokrit berbanding lurus dengan jumlah eritrosit dan kadar hemoglobin. Semua faktor yang mempengaruhi proses produksi dan jumlah eritrosit berpengaruh pula terhadap nilai hematokrit (Sturkie 1976).
Leukosit berperan sebagai unit pertahanan tubuh (Guyton 1982). Pada umumnya jumlah total leukosit cenderung lebih tinggi pada burung betina dewasa daripada jantan (Hodges 1977). Jumlah leukosit dipengaruhi oleh umur, jenis kelamin, kondisi stres, aktivitas fisiologis yang tinggi, gizi, lingkungan, efek hormon, obat dan sinar x (Brown 1989; Sturkie 1976).
Gambar 4. Eritrosit burung beo jawa (Hawkey & Dennet 1989) Keterangan : Tanda panah hitam: sel sabit darah merah Tanda panah biru : eritrosit
Tanda panah merah : limfosit
Sel sabit darah merah sering ditemui pada burung beo (G.r.religiosa dari Jawa) dan ini merupakan fenomena invitro yaitu sampel darah yang
(a) (b)
(c) (d)
mengandung oksigen dan tidak berhubungan dengan tanda klinik yang abnormal. Sebagaimana juga ditemukan pada beberapa spesies rusa dewasa (swamp deer), sapi, kambing dan beberapa carnivora kecil (Hawkey & Dennet 1989). Gambaran darah pada beberapa burung dapat dilihat pada Gambar 5 (Hawkey & Dennet 1989)
Gambar 5. Gambaran darah pada beberapa jenis burung (Hawkey & Dennet 1989) (a) Eosinofil (tanda panah hitam) pada burung lappet faced yang terinfestasi parasit
(b) Monosit (tanda panah hijau) pada burung flamingo normal (c) Heterofil (tanda panah hitam) dan trombosit (tanda panah merah) pada burung merpati normal
(d) Limfosit besar (tanda panah biru) pada burung elang
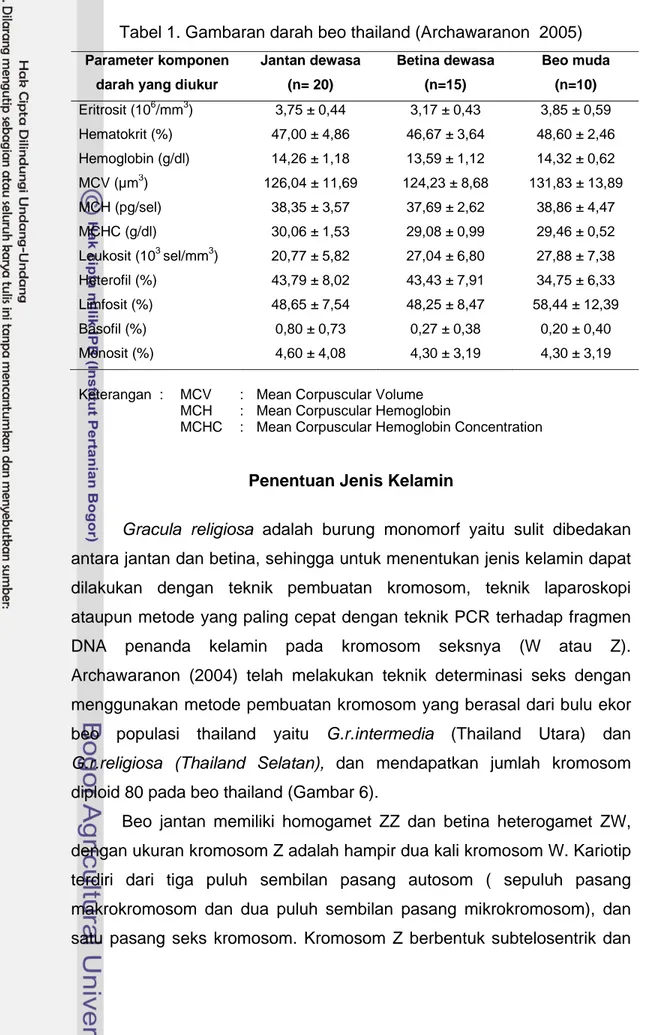
Penelitian tentang gambaran hematologis pada burung beo thailand telah dilakukan oleh Archawaranon (2005) seperti yang disajikan pada Tabel 1. Beo thailand muda memiliki jumlah eritrosit yang paling tinggi (3,85±0,59 x 106/mm3), menyusul beo jantan dewasa (3,75±0,44 x 106/mm3) dan beo betina dewasa (3,17±0,43 x 106/mm3).
Tabel 1. Gambaran darah beo thailand (Archawaranon 2005) Parameter komponen
darah yang diukur
Jantan dewasa (n= 20) Betina dewasa (n=15) Beo muda (n=10) Eritrosit (106/mm3) 3,75 ± 0,44 3,17 ± 0,43 3,85 ± 0,59 Hematokrit (%) 47,00 ± 4,86 46,67 ± 3,64 48,60 ± 2,46 Hemoglobin (g/dl) 14,26 ± 1,18 13,59 ± 1,12 14,32 ± 0,62 MCV (µm3) 126,04 ± 11,69 124,23 ± 8,68 131,83 ± 13,89 MCH (pg/sel) 38,35 ± 3,57 37,69 ± 2,62 38,86 ± 4,47 MCHC (g/dl) 30,06 ± 1,53 29,08 ± 0,99 29,46 ± 0,52 Leukosit (103 sel/mm3) 20,77 ± 5,82 27,04 ± 6,80 27,88 ± 7,38 Heterofil (%) 43,79 ± 8,02 43,43 ± 7,91 34,75 ± 6,33 Limfosit (%) 48,65 ± 7,54 48,25 ± 8,47 58,44 ± 12,39 Basofil (%) 0,80 ± 0,73 0,27 ± 0,38 0,20 ± 0,40 Monosit (%) 4,60 ± 4,08 4,30 ± 3,19 4,30 ± 3,19
Keterangan : MCV : Mean Corpuscular Volume
MCH : Mean Corpuscular Hemoglobin
MCHC : Mean Corpuscular Hemoglobin Concentration
Penentuan Jenis Kelamin
Gracula religiosa adalah burung monomorf yaitu sulit dibedakan antara jantan dan betina, sehingga untuk menentukan jenis kelamin dapat dilakukan dengan teknik pembuatan kromosom, teknik laparoskopi ataupun metode yang paling cepat dengan teknik PCR terhadap fragmen DNA penanda kelamin pada kromosom seksnya (W atau Z). Archawaranon (2004) telah melakukan teknik determinasi seks dengan menggunakan metode pembuatan kromosom yang berasal dari bulu ekor beo populasi thailand yaitu G.r.intermedia (Thailand Utara) dan G.r.religiosa (Thailand Selatan), dan mendapatkan jumlah kromosom diploid 80 pada beo thailand (Gambar 6).
Beo jantan memiliki homogamet ZZ dan betina heterogamet ZW, dengan ukuran kromosom Z adalah hampir dua kali kromosom W. Kariotip terdiri dari tiga puluh sembilan pasang autosom ( sepuluh pasang makrokromosom dan dua puluh sembilan pasang mikrokromosom), dan satu pasang seks kromosom. Kromosom Z berbentuk subtelosentrik dan
makrokromosom, sedangkan kromosom W berbentuk submetasentrik dan berukuran lebih kecil (Archawaranon 2004). Kebanyakan kariotip unggas terdiri atas sejumlah sedikit makrokromosom dan sejumlah besar mikrokromosom ( Hammer 1970; Tegelstrom H & Ryttman H 1981). Pada Gambar 6 terlihat perbedaan cuping kuduk beo populasi thailand utara dan thailand selatan, yaitu beo populasi thailand utara memiliki cuping kuduk bersatu dengan cuping di pipi membentuk segitiga, sedangkan beo populasi thailand selatan memiliki cuping kuduk terpisah dengan cuping di pipi.
Gambar 6. Kariotip burung beo thailand (G.r.religiosa dan G.r.intermedia), MN: Modified Northern, S: Southern, N: Northern I: Intermediate, MS: Modified Southern (Archawaranon 2004)
Genetika Konservasi
Masalah genetika akhir-akhir ini banyak mendapat sorotan dunia dalam hubungannya dengan masalah konservasi. Tolok ukur keberhasilan kegiatan konservasi didasarkan pada keanekaragaman genetiknya, sehingga keanekaragaman genetik menjadi indikator kunci yang penting. Keadaan alam yang berubah dari waktu ke waktu menyebabkan setiap spesies harus mampu beradaptasi dengan lingkungan yang berubah, untuk bertahan hidup. Variasi genetik menentukan kemampuan individu untuk beradaptasi terhadap perubahan (Haig & Nordstrom 1991; Masy’ud 1992).
Kepunahan suatu spesies sebenarnya fenomena yang telah ada sejak dulu. Kepunahan suatu spesies diawali dengan terpecahnya populasi besar menjadi populasi kecil, yang mengakibatkan kehilangan atau berkurangnya variasi genetik. Pada akhirnya terjadi pengikisan kelenturan evolusi sehingga suatu spesies tidak mampu beradaptasi terhadap lingkungan yang berubah (Nilsson 1983; Haig & Nordstrom 1991).
Keanekaragaman genetik identik dengan keanekaragaman alela dan heterozigositas alela. Hilangnya keanekaragaman hayati dapat menyebabkan 1) penyimpangan genetik secara acak, 2) derajat silang dalam (inbreeding) yang tinggi dan 3) pengurangan populasi efektif dari suatu spesies (Lacy 1987; Haig & Nordstrom 1991). Menurut Haig dan Nordstrom (1991), strategi yang dapat diaplikasikan pada populasi yang kecil yaitu dengan memasukkan individu-individu baru dari populasi yang lain secara teratur, sehingga akan memperbesar populasinya secara cepat.
Sampai saat ini masih sedikit sekali informasi mengenai variasi genotip pada burung beo di Indonesia. Studi yang sudah dilakukan adalah analisis polimorfisme protein darah pada burung beo nias, medan, irian (Siregar 1997). Dengan menggunakan rumus jarak genetik dan kesamaan genetik Nei (1987) menunjukkan bahwa beo nias dan beo medan memiliki jarak genetik yang dekat (D=0,0014) dan kesamaan genetik yang tinggi
(I=0,9969). Beo irian (Mino dumontii) merupakan spesies berbeda dengan beo medan dan beo nias, tapi masih berkerabat dekat. Antara beo nias dan beo irian mempunyai jarak genetik sebesar I=0,6022 dan D=0,2203, sedangkan antara beo medan dan beo irian memiliki jarak genetik sebesar I=0,6425 dan D=0,1921. Dari analisis polimorfisme protein darah, menunjukkan bahwa beo nias dan beo medan mempunyai kekerabatan yang lebih dekat dibandingkan dengan beo irian (Siregar 1997). Sedangkan di Thailand, Kudan et al. (2002 unpublished) telah melakukan perunutan DNA sebagian daerah D-loop mtDNA burung beo G.r.intermedia dan G.r.religiosa masing-masing dari populasi Thailand Utara dan Thailand Selatan. Primer PCR yang digunakan untuk amplifikasi sebagian daerah D-loop adalah Primer F (NDGE) 5’-CCA TAA CCA ACA ACC TGT CAAT-3’ dan Primer R (H417) 5’-AGT AGC TCG GTT CTC GTG AG-3’ (Kudan S 20 Juli 2004, komunikasi pribadi).
Genotip, Fenotip, Mutasi Gen, dan Evolusi Burung
Istilah genotip dan fenotip muncul untuk memahami interaksi antara gen dan lingkungan terhadap organisme. Suatu organisme dikatakan mirip atau menyerupai induknya karena ada beberapa persamaan ciri-ciri yang dapat dilihat. Dalam hal ini ciri-ciri tersebut diwariskan. Ditinjau dari sudut ilmu keturunan, ciri-ciri yang diwariskan tadi disimpan sebagai informasi genetik dalam gen-gen yang secara molekular tersusun atas asam nukleat DNA (Burns & Bottino 1988; Sofro 1994).
Informasi genetik yang disimpan dalam DNA selamanya akan tetap tersimpan dalam runutan DNA kodon-kodon apabila tidak diekspresikan dalam molekul protein. Molekul protein yang terbentuk untuk selanjutnya akan menjadi ciri-ciri yang dapat dilihat, baik dalam bentuk sebagai protein itu sendiri atau alternatifnya bersama faktor-faktor lain merupakan suatu gambaran bentuk anatomi, fisiologis, biokimia atau bahkan aktivitas tertentu. Dalam kaitan ekspresi ciri-ciri ini, bila suatu ciri masih dinyatakan dalam perangkat genetiknya, maka ciri-ciri ini disebut genotip. Tapi apabila genotip ini sudah diekspresikan dalam bentuk suatu molekul protein dan
dipengaruhi oleh faktor lingkungan, maka ciri-ciri ini menjadi suatu penampilan atau fenotip. Organisme termasuk genotip yang sama bila mereka memiliki seperangkat gen yang sama. Sementara itu organisme termasuk dalam fenotip yang sama bila mereka memiliki beberapa persamaan ciri-ciri yang ditampilkan. Genotip memberikan seperangkat komplit gen yang diwariskan oleh suatu individu, sedangkan fenotip merupakan ciri-ciri lahiriah organisme yang dihasilkan karena interaksi antara gen dan lingkungan. Sebenarnya tidak ada dua individu yang memiliki fenotip sama karena pasti ada perbedaan walaupun sedikit (Singer & Berg 1981; Sofro 1994).
Mutasi terdiri atas dua jenis yaitu mutasi kromosom dan mutasi gen. Mutasi kromosom adalah perubahan yang terjadi pada kromosom, meliputi: a) duplikasi (penambahan), b) delesi (lenyapnya gen dari suatu kromosom), c) inversi (perubahan letak segmen kromosom), d) pindah silang (pertukaran gen dari kromosom homolog), e) translokasi (pertukaran dari kromosom (nonhomolog) (Sugiri 1988). Mutasi gen secara umum adalah perubahan apapun dalam bahan genetik. Mutasi gen tidak dapat dilihat dengan mikroskop. Perubahan bahan genetik ini pada dasarnya disebabkan oleh lima hal, yaitu : a) mutasi noktah yang meliputi perubahan pada kodon-kodon tunggal, b) transisi, c) transversi, d) insersi dan e) delesi. Namun secara lebih terbatas mutasi lebih menunjuk ke perubahan dalam gen (intragenik). Dalam mutasi semacam ini ada dua kelompok besar, yaitu penggantian basa dan mutasi pergeseran rangka.
Mutasi gen merupakan faktor penentu timbulnya keanekaragaman genetik yang berakibat pada timbulnya keanekaragaman dalam kehidupan. Pada mutasi penggantian basa dapat terjadi apa yang disebut transisi dan tranversi. Mutasi transisi adalah penggantian purin yang satu dengan purin yang lain atau pirimidin yang satu dengan pirimidin yang lain, dan umumnya terjadi selama replikasi DNA. Pada peristiwa ini terjadi pergeseran elektron yang menyebabkan bentuk molekul menjadi sedikit berubah. Pergeseran tautomer pada basa DNA mengubah sifat pasangan basa sehingga A dapat berpasangan dengan C, dan T dengan G. Mutasi
transversi yaitu penggantian basa purin dengan pirimidin atau sebaliknya. Umumnya penyebab transversi berbeda dengan penyebab transisi karena tranversi tidak terjadi selama replikasi DNA, melainkan terkait dengan sistem reparasi DNA yang rentan terhadap kesalahan (Burns & Bottino 1988; Sofro 1994).
Akibat adanya mutasi transisi dan transversi dapat terjadi mutasi misens atau nonsens. Pada mutasi misens, asam amino pada rantai polipeptida yang disintesis digantikan dengan asam amino lain. Pada mutasi nonsens, kodon yang menyandi suatu asam amino berubah menjadi kodon henti sehingga sintesis rantai polipeptida berhenti sebelum waktunya (Burns & Bottino 1988; Sofro 1994).
Pada mutasi pergeseran rangka, mutasi terjadi apabila satu pasangan basa atau lebih ditambahkan (insersi) atau terhapus (delesi) dari molekul DNA. Hal ini dapat dimengerti karena kodon dibaca berurutan untuk tiap triplet nukleotida. Dengan adanya tambahan atau pengurangan basa, maka pembacaan triplet nukleotida menjadi bergeser. Akibat terjadinya geseran pembacaan kodon, maka asam amino yang disandi juga mengalami perubahan. Gen mutan semacam ini akan bertahan bila protein yang terbentuk dapat bertahan (Burns & Bottino 1988; Sofro 1994).
Dari kenyataan adanya mutasi, tampak bahwa meskipun proses replikasi DNA merupakan proses yang tepat, tetapi kadang-kadang juga terjadi kesalahan. Seringkali kesalahan ini dibetulkan dalam sel. Tetapi kadang-kadang kesalahan ini tidak diperbaiki dan terbawa terus pada proses replikasi selanjutnya. Kesalahan yang tidak diperbaiki semacam ini dapat bersifat merusak, tetapi tidak jarang pula tidak menimbulkan kerugian. Justru adanya mutasi semacam inilah yang menyebabkan timbulnya keanekaragaman kehidupan di dunia (Burns & Bottino 1988; Sofro 1994).
Burung termasuk salah satu kelompok vertebrata yang paling umum dijumpai. Ciri utama kelompok burung adalah adanya bulu yang menutupi tubuhnya. Secara evolusi, burung berkembang dari reptilia yang
berkaki dua (Storer et al. 1979; Welty 1982). Evolusi terdiri dari dua jenis yaitu mikroevolusi dan makroevolusi, dimana mikroevolusi ialah evolusi yang berhubungan dengan gene pool pada populasi tunggal dan makroevolusi adalah evolusi yang termasuk asal usul spesies baru dan pengelompokan taksonomi tingkat tinggi ( Campbell et al. 2006).
Teknik DNA telah memungkinkan penelusuran sejarah evolusi organisme, termasuk burung. Menurut Sibley dan Ahlquist (1991) perbandingan DNA menunjukkan bahwa ada hubungan kekerabatan yang dekat antara dua kelompok passerine, yaitu antara kelompok jalak dan beo (starling) di Dunia Lama dengan kelompok mockingbird di Dunia Baru yang ternyata baru terpisah 25 juta tahun yang lalu. Berdasarkan penelitian-penelitian tentang anatomi kepala, syrinx dan protein otot yang pernah dilakukan oleh beberapa peneliti sebelumnya Sibley dan Ahlquist (1991) juga menyimpulkan bahwa terdapat kesamaan yang signifikan antara beo dengan mockingbird, yaitu memiliki kepandaian meniru suara.
Dibandingkan dengan vertebrata terestrial lainnya burung adalah satu-satunya yang memiliki kemampuan terbang yang sangat kuat (Welty 1982). Kemampuan terbang burung inilah yang menurut Darlington (1966) merupakan faktor utama dalam penyebaran burung, atau dengan kata lain kondisi penyebaran burung di dunia saat ini adalah hasil dari pergerakan berpindah burung, dan bukan akibat dari pergeseran benua.
Daerah Kontrol D-loop Mitokondrion DNA
Mitokondrion merupakan organel berupa kantung yang diselaputi oleh dua membran yaitu membran luar, dan membran dalam, sehingga memiliki dua kompartemen yaitu matriks mitokondrion yang diselimuti langsung oleh membran dalam dan ruang antar membran, seperti terlihat pada Gambar 7. Membran luar dapat dilalui oleh ion dan molekul berukuran kecil, sedangkan membran dalam bersifat impermiabel. Pada membran dalam terdapat kompleks protein rantai respirasi, ATP sintase dan berbagai transporter membran. Ruang matriks mengandung berbagai jenis enzim yang antara lain terlibat dalam oksidase piruvat, asam lemak
dan asam amino serta enzim yang terlibat dalam siklus asam sitrat (Lodish et al. 1995, Artika 2003).
Membran dalam memiliki struktur melekuk, melipat kedalam ke bagian matriks mitokondrion, dikenal sebagai krista. Struktur melekuk-lekuk ini sangat membantu dalam meningkatkan luas permukaan membran dalam, sehingga meningkatkan kemampuannya dalam memproduksi ATP (Lodish et al. 1995, Artika 2003).
Matriks mitokondrion berupa cairan kental serupa gel, dengan campuran ratusan jenis enzim dengan konsentrasi yang sangat tinggi, untuk proses oksidasi piruvat, oksidasi asam lemak dan untuk menjalankan siklus asam trikarboksilat. Matriks mitokondrion juga mengandung salinan identik DNA genom mitokondrion, ribosom mitokondrion, tRNA dan berbagai jenis enzim yang diperlukan untuk ekspresi gen mitokondrion (Lodish et al. 1995, Artika 2003).
Molekul DNA mitokondrion mempunyai banyak kelebihan sebagai penanda molekular dalam mempelajari hubungan evolusi hewan pada berbagai tingkatan. Hal ini disebabkan ukuran DNA mitokondrion vertebrata relatif kecil (±16 kb) yang mengandung 13 gen menyandikan protein, 22 gen menyandikan tRNA (transfer RNA), 2 gen menyandikan rRNA (ribosomal RNA) dan satu ruas DNA berukuran besar yang tidak menyandikan protein. DNA mitokondrion tidak memiliki intron, kodon stop beberapa gennya tidak sempurna dan ujung 3’-CCA dari gen-gen tRNA-nya tidak ada.
Pola pewarisan mitokondrion melalui garis ibu yang menyebabkan tidak ada rekombinasi dan laju mutasinya tinggi, sehingga mempunyai keunggulan tersendiri sebagai penanda molekular tingkat intraspesies pada sebagian besar vertebrata (Avise 1994). Ekspresi genom mitokondrion berlangsung di mitokondrion. Berbeda dengan mRNA nukleus, mRNA mitokondrion tidak mengandung intron (Lodish et al. 1995).
Gambar 7. Diagram struktur mitokondrion manusia (Lodish et al. 1995) Keterangan: (a) Struktur tiga dimensi mitokondrion
(b) Diagram penampang melintang mitokondrion (c) Diagram satu dimensi mitokondrion
Genom mitokondrion mempunyai variasi runutan DNA yang tinggi, beberapa kali lebih tinggi dibandingkan DNA inti dan kecepatan evolusi 5-10 kali lebih cepat daripada genom inti dan di daerah D-loop mengalami evolusi lebih cepat lagi (Brown et al. 1982, Majerus 1996). Genom mitokondrion ayam leghorn putih berukuran 16775 nukleotida. Genom mitokondrion unggas menyandikan set gen yang sama sebagaimana genom vertebrata lainnya (13 penyandi protein, 2 rRNA dan 22 tRNA) dan mempunyai model yang sangat ekonomis (Desjardins & Morais 1990).
Genom mtDNA ayam memperlihatkan dua karakter berbeda yaitu : menunjukkan novel gene order, ND6 dan tRNA Glu mempunyai lokasi berdekatan dengan daerah kontrol D-loop, di depannya terdapat tRNA Phe, 12sRNA, 16sRNA, ND1, ND2, COI, COII, ATP6, COIII, ND3, ND4, ND5 dan gen penyandi sitokrom b dan pada ayam tidak ada daerah yang setara dengan light-strand replication origin yang merupakan runutan DNA yang kekal, yang ditemukan diantara tRNA Cys dan tRNA Asn pada semua genom mitokondrion vertebrata. Hal ini menunjukkan bahwa genom
16569 pb
c b
16775 pb
mitokondrion galliformes terpisah dengan mamalia dan amfibia selama evolusi spesies vertebrata (Desjardins & Morais 1990).
Dalam sel vertebrata yang sedang aktif, sebagian dupleks DNA mitokondrion mengandung struktur pendek berutas tiga yang disebut D-loop (Displacement Loop). D-D-loop tidak memiliki penyandi untuk gen-gen struktural tetapi kebanyakan mengandung elemen pengatur replikasi mt DNA, transkripsi, dan pengatur ekspresi genetik mtDNA serta memiliki laju mutasi yang tinggi, bahkan tertinggi di seluruh genom sehingga mempunyai nilai informasi yang tinggi dalam menguji kekerabatan di dalam satu spesies (Brown et al. 1982; Aquadro & Greenberg 1983; Lodish et al. 1995; Vigilant et al. 1989).
D-loop bangsa burung diapit oleh gen tRNAGlu dan tRNAPhe, tidak seperti vertebrata lainnya diapit oleh tRNAPro dan tRNAPhe (Desjardins dan Morais 1990, Gambar 8). Daerah D-loop mtDNA telah dirunut secara lengkap pada tujuh spesies ayam hutan Alectoris. Panjang D-loop sangat kekal (1155 ± 2 nukleotida) dan laju substitusi lebih rendah dibandingkan dengan gen sitokrom b pada spesies yang sama.
Analisis komparatif menunjukkan adanya pembagian daerah D-loop ke dalam tiga domain dengan bagian sisi domain I dan domain III mengandung variabilitas nukleotida yang tinggi, sedangkan domain II sebagai pusat yang sangat kekal pada vertebrata (Baker & Marshall, 1997 seperti yang diacu dalam Randi & Lucchini 1998). Walaupun demikian, 161 nukleotida pertama dari domain I D-loop Alectoris pada sisi setelah tRNAGlu, berevolusi pada laju yang lebih lambat dan memperlihatkan motif yang serupa dengan runutan DNA pada mamalia yang disebut ETAS (Extended Termination-Associated Sequences) I dan ETAS II (Sbisa et al. 1997) yang dapat membentuk struktur sekunder yang stabil. Bagian ke dua dari domain I mengandung daerah hipervariabel dengan dua kopi runutan DNA berulang ganda (tandemly repeated sequence) pada spesies lain dari anseriformes dan galliformes (Quinn & Wilson 1993; Fumihito et al.1995). Beberapa blok daerah kekal pada mamalia dapat dipetakan dalam domain pusat Allectoris. Domain III mempunyai variabilitas yang tinggi dan runutan DNA serupa dengan CSBI (Conserved Sequence Block I) mamalia (Gambar 9).