POPULASI PROTOZOA DAN KARAKTERISTIK FERMENTASI
RUMEN DENGAN PEMBERIAN DAUN KERSEN
(Muntingia calabura) SECARA IN VITRO
SKRIPSI
IDE RISENTITO PUSPITANING
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2012
RINGKASAN
IDE RISENTITO PUSPITANING. D24080097. 2012. Populasi Protozoa dan Karakteristik Fermentasi Rumen dengan Pemberian Daun Kersen (Muntingia calabura) secara In Vitro. Skripsi. Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor.
Pembimbing Utama : Dr. Sri Suharti, S.Pt. M.Si.
Pembinmbing Anggota : Dr. Anuraga Jayanegara, S.Pt. M.Sc.
Daun kersen (Muntingia calabura) memiliki kandungan saponin yang cukup tinggi sehingga dapat dimanfaatkan sebagai agen defaunasi protozoa yang dapat meningkatkan efisiensi fermentasi rumen. Penyebarannya yang luas yang mudah ditemukan pada berbagai kondisi lahan termasuk lahan marginal, menjadikan tumbuhan ini sangat potensial untuk dimanfaatkan sebagai pakan ternak. Penelitian ini bertujuan untuk menganalisis kandungan fitokimia daun kersen yang meliputi senyawa tanin dan saponin serta mengevaluasi dosis daun kersen yang tepat dan sesuai untuk mengoptimalkan fermentasi rumen.
Rancangan yang digunakan pada penelitian ini adalah Rancangan Acak Kelompok (RAK) dengan 5 taraf perlakuan dan 4 kelompok sebagai ulangan. Perlakuan berupa ransum komplit terdiri dari R0 = 30% konsentrat + 70% rumput lapang; R1 = 30% konsentrat + 65% rumput lapang + 5% daun kersen; R2 = 30% konsentrat + 60% rumput lapang + 10% daun kersen; R3 = 30% konsentrat + 55% rumput lapang + 15% daun kersen; R4 = 30% konsentrat + 50% rumput lapang + 20% daun kersen. Peubah yang diamati adalah nilai pH rumen, populasi protozoa, konsentrasi amonia (NH3), produksi Volatile Fatty Acid (VFA) , Kecernaan Bahan
Kering (KCBK), dan Kecernaan Bahan Organik (KCBO). Data hasil penelitian dianalisis sidik ragam (ANOVA) jika hasilnya berbeda nyata akan diuji lanjut dengan Uji Jarak Duncan. Analisis data dilakukan menggunakan software statistik SPSS versi 16.
Hasil analisis fitokimia menunjukkan bahwa daun kersen memiliki kandungan saponin 10,28% dan tanin 1,41%. Hasil analisis ragam menunjukkan bahwa penggunaan daun kersen pada ransum tidak nyata (P>0,05) mempengaruhi nilai pH rumen dan populasi protozoa. Konsentrasi NH3 menurun sangat nyata
(P<0,01), namun produksi VFA meningkat sangat nyata (P<0,01). Kecernaan bahan kering dan bahan organik menurun secara nyata (P<0,05). Penggunaan tepung daun kersen sampai level 20% dapat menurunkan kecernaan bahan kering, kecernaan bahan organik, dan konsentrasi NH3 serta mampu meningkatkan produksi VFA total,
namun belum efektif dalam penekanan populasi protozoa dengan waktu inkubasi 4 jam secara in vitro.
ABSTRACT
Protozoal Population and Rumen Fermentation Characteristics of Diet with Muntingia calabura Leaves Addition In Vitro
Puspitaning, I. R., S. Suharti, and A. Jayanegara
The exotic tree Muntingia calabura L. (MCL), is a fast-growing tree that contains secondary metabolite compounds such as saponins and tannins. Saponins and tannins has potency to improve rumen fermentation. An in vitro study was conducted to determine the effect of MCL supplementation on protozoal population and fermentation characteristics. Randomized complete block design was applied to this research by using 5 treatments with 4 replications for each treatment. The treatments were: R0: 30% concentrate (C) + 70% forage (F); R1: 30% C + 65% F + 5% MCL; R2: 30% C + 60% F + 10% MCL; R3: 30% C + 55% F + 15% MCL; R4: 30% C + 50% F + 20% MCL. Variables observed were pH, protozoal population, N-NH3 and
total volatile fatty acid (VFA) production, dry matter and organic matter digestibility. Data were analyzed using analysis of variance (ANOVA). The results showed that the addition of MCL up to 20% in the ration did not affect (P>0.05) pH and protozoal population. Amonia-N concentration decreased (P<0.01) and total VFA increased (P<0.01) when MCL was included. In vitro dry matter and organic matter digestibility decreased (P<0.05) with MCL addition. In conclusion, addition of MCL powder up to 20% from total diet could decrease in vitro dry matter and organic matter digestibility, amonia-N concentration, and increase total VFA production, but did not effective to decrease protozoal population in 4 h incubation in vitro.
POPULASI PROTOZOA DAN KARAKTERISTIK FERMENTASI
RUMEN DENGAN PEMBERIAN DAUN KERSEN
(Muntingia calabura) SECARA IN VITRO
IDE RISENTITO PUSPITANING D24080097
Skripsi ini merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada
Fakultas Peternakan Institut Pertanian Bogor
DEPARTEMEN ILMU NUTRISI DAN TEKNOLOGI PAKAN FAKULTAS PETERNAKAN
INSTITUT PERTANIAN BOGOR 2012
Judul : Populasi Protozoa dan Karakteristik Fermentasi Rumen dengan Pemberian Daun Kersen (Muntingia calabura) secara In Vitro
Nama : Ide Risentito Puspitaning NIM : D24080097
Menyetujui, Pembimbing Utama,
(Dr. Sri Suharti, S.Pt.M.Si) NIP: 19741012 200501 2 002 Pembimbing Anggota, (Dr. Anuraga Jayanegara, S.Pt.M.Sc) NIP: 19830602 200501 1 001 Mengetahui Ketua Departemen,
Ilmu Nutrisi dan Teknologi Pakan
(Dr. Ir. Idat Galih Permana, M.Sc.Agr) NIP: 19670506 199103 1 001
RIWAYAT HIDUP
Penulis dilahirkan pada tanggal 26 September 1990 di Pati, Jawa Tengah. Penulis adalah anak pertama dari dua bersaudara dari pasangan Bapak Lis Indrianto dan Ibu Eri Lestari Muryani.
Penulis mengawali pendidikan dasar pada tahun 1996 di Sekolah Dasar Negeri Pati Lor 04 dan diselesaikan pada tahun 2002. Pendidikan lanjutan tingkat pertama dimulai pada tahun 2002 dan diselesaikan pada tahun 2005 di Sekolah Lanjutan Tingkat Pertama Negeri 2 Pati.
Penulis melanjutkan pendidikan di Sekolah Menengah Atas Negeri 3 Pati pada tahun 2005 dan diselesaikan pada tahun 2008.
Penulis diterima sebagai mahasiswa Departemen Ilmu Nutrisi dan Teknologi Pakan, Fakultas Peternakan, Institut Pertanian Bogor pada tahun 2008 melalui jalur Undangan Seleksi Masuk IPB (USMI). Penulis aktif dalam Unit Kegiatan Mahasiswa Uni Konservasi Fauna (UKF) sebagai staf Konservasi Eksitu (periode 2009-2011) dan sekretaris bidang Sosial Lingkungan (periode 2010-2011). Penulis juga aktif dalam organisasi Himpunan Mahasiswa Nutrisi dan Makanan Ternak (HIMASITER) sebagai staf Kewirausahaan (periode 2009-2011), Kelompok Pecinta Alam Fakultas Peternakan (KEPAL-D) dan Organisasi Mahasiswa Daerah Ikatan Keluarga Mahasiswa Pati (IKMP). Penulis pernah mengikuti kegiatan magang di Internasional Animal Rescue (IAR) Bogor dan Taman Margasatwa Ragunan Jakarta pada tahun 2010, serta mengikuti kegiatan magang di Balai Embrio Ternak (BET) Bogor pada tahun 2011. Karya tulis dari penulis yang pernah dipublikasikan adalah Prosiding Seminar Nasional Uni Konservasi Fauna 2011 dengan judul Konservasi Raptor sebagai Salah Satu Potensi Ekowisata.
Bogor, Juli 2012 Ide Risentito Puspitaning D24080097
KATA PENGANTAR
Bissmillahirrahmannirrahim.
Puji syukur penulis panjatkan kehadirat Allah SWT karena atas rahmat, karunia serta ridho-Nya penyusunan skripsi ini dapat terselesaikan dengan baik. Skripsi yang berjudul “Populasi Protozoa dan Karakteristik Fermentasi Rumen dengan Pemberian Daun Kersen (Muntingia calabura) secara In Vitro” merupakan salah satu syarat untuk memperoleh gelar Sarjana Peternakan pada Fakultas Peternakan, Institut Pertanian Bogor.
Tujuan penulisan skripsi ini yaitu untuk menganalisis kandungan fitokimia daun kersen yang meliputi senyawa tanin dan saponin serta mengevaluasi dosis daun kersen yang tepat dan sesuai untuk mengoptimalkan fermentasi rumen secara in vitro. Penelitian ini dilaksanakan dari bulan Maret sampai Mei 2012. Lokasi Penelitian bertempat di Laboratorium Biokimia, Fisiologi, dan Mikrobiologi Nutrisi, Laboratorium Nutrisi Perah, dan Laboratorium Pusat Penelitian Sumberdaya Hayati dan Bioteknologi Institut Pertanian Bogor serta Laboratorium Balai Penelitian Ternak (Balitnak) Ciawi, Bogor.
Penulis mengucapkan terima kasih atas kritik dan saran yang membangun oleh berbagai pihak demi kesempurnaan skripsi ini. Penulis berharap semoga skripsi ini dapat bermanfaat terutama dalam dunia peternakan.
Bogor, Juli 2012
11
DAFTAR ISI
Halaman RINGKASAN ... ii ABSTRACT ... iii LEMBAR PERNYATAAN ... iv LEMBAR PENGESAHAN ... v RIWAYAT HIDUP ... viKATA PENGANTAR ... vii
DAFTAR ISI ... viii
DAFTAR TABEL ... x
DAFTAR GAMBAR ... xi
DAFTAR LAMPIRAN ... xii
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan Penelitian ... 2
TINJAUAN PUSTAKA ... 3
Daun Kersen (Muntingia calabura) ... 3
Saponin ... 4
Tanin ... 5
Mikroba Rumen ... 6
Amonia (NH3) ... 7
Volatile Fatty Acid (VFA) ... 9
Kecernaan Bahan Kering dan Bahan Organik ... 10
MATERI DAN METODE ... 11
Lokasi dan Waktu ... 11
Materi ... 11
Alat ... 11
Bahan ... 11
Prosedur ... 12
Pembuatan Tepung Daun Kersen ... 12
Analisis Proksimat dan Fitokimia ... 13
Pembuatan Larutan McDougall ... 13
Pengambilan Cairan Rumen ... 13
Fermentasi In Vitro ... 13
Nilai pH ... 14
Populasi Protozoa ... 14
Konsentrasi NH3 ... 14
KCBK dan KCBO ... 15
Perlakuan ... 16
Peubah ... 16
Model ... 16
Analisis Data ... 16
HASIL DAN PEMBAHASAN ... 17
Daun Kersen sebagai Pakan ... 17
Nilai pH Rumen ... 18
Populasi Protozoa ... 18
Konsentrasi Amonia (NH3) ... 20
Produksi VFA Total ... 21
Kecernaan Bahan Kering dan Bahan Organik ... 22
KESIMPULAN DAN SARAN ... 23
Kesimpulan ... 23
Saran ... 23
UCAPAN TERIMA KASIH ... 24
DAFTAR PUSTAKA ... 25
DAFTAR TABEL
Nomor Halaman
1. Komposisi Konsentrat ... 12 2. Kandungan Zat Makanan Ransum dalam Bahan Kering ... 12 3. Kandungan Fitokimia Daun Kersen ... 17 4. Pengaruh Perlakuan terhadap Populasi Protozoa dan Karakteristik
DAFTAR GAMBAR
Nomor Halaman
1. Daun Kersen (Muntingia calabura) ... 3 2. Struktur Saponin Steroid ... 4 3. Struktur Saponin Triterpenoid ... 4 4. Unit Monomer Tanin Terkondensasi (Catechin dan Gallocatechin dan
Terhidrolisis (Asam Gallic dan Ellagic) ... 5 5. Proses Metabolisme Protein dalam Rumen Ternak Ruminansia ... 8 6. Proses Metabolisme Karbohidrat dalam Rumen Ternak Ruminansia 9
DAFTAR LAMPIRAN
Nomor Halaman
1. Hasil Analisa Pengaruh Perlakuan terhadap Nilai pH ... 29 2. Hasil Analisa Pengaruh Perlakuan terhadap Populasi Protozoa ... 29 3. Hasil Analisa Pengaruh Perlakuan terhadap Konsentrasi NH3 ... 29
4. Hasil Analisa Pengaruh Perlakuan terhadap Produksi VFA Total ... 30 5. Hasil Analisa Pengaruh Perlakuan terhadap Nilai KCBK ... 31 6. Hasil Analisa Pengaruh Perlakuan terhadap Nilai KCBO ... 31
PENDAHULUAN
Latar Belakang
Kebutuhan impor daging belum dapat dihindarkan karena tingkat kebutuhan daging di Indonesia lebih banyak daripada produksinya. Badan Pusat Statistik (2012) mencatat terdapat 14.805.053 ekor sapi potong pada masa sensus Juni 2011, namun hasil sensus tersebut belum dapat mengurangi kebutuhan impor karena hanya 1,425 juta ekor sapi yang siap potong. Laporan tahunan tahun 2011 oleh Badan Penelitian dan Pengembangan Pertanian (2012) menyatakan bahwa Indonesia diperkirakan mengimpor sapi hidup sebanyak 650 ribu ekor dari Australia dan 72 ribu ton daging sapi beku (setara dengan 220 ribu ekor sapi). Penekanan jumlah impor daging dapat dilakukan dengan meningkatkan produksi daging lokal.
Peternakan rakyat menjadi kunci keberhasilan program kecukupan daging karena sistem peternakan sapi potong di Indonesia masih mengandalkan peternakan rakyat (Mayulu et al., 2010). Sementara peternakan rakyat menggunakan ransum hijauan dalam jumlah banyak (hampir 100%) yang sering menyebabkan defisiensi nutrien sehingga mengakibatkan produktivitas ternak rendah. Peningkatan efisiensi fermentasi rumen perlu dilakukan sebagai upaya untuk meningkatkan produktivitas ternak yang sangat ditentukan oleh aktifitas mikroba rumen.
Ekosistem rumen ternak ruminansia termasuk sapi potong dihuni oleh bakteri, protozoa, fungi, dan arkae yang berperan dalam pencernaan pakan. Populasi protozoa yang tinggi dalam rumen ruminansia kurang menguntungkan karena sifatnya yang memangsa (predator) bakteri untuk memenuhi kebutuhan proteinnya. Akibatnya biomassa bakteri akan berkurang sehingga laju degradasi pakan di dalam rumen dan suplai protein mikroba akan berkurang pula (Soetanto, 2004). Sementara itu pada ternak yang mendapat pakan tinggi hijauan, perkembangan populasi mikroba rumen terutama bakteri pencerna serat sangat menentukan fermentasi pakan di dalam rumen.
Salah satu strategi untuk meningkatkan efisiensi fermentasi rumen pada ternak yang mendapat ransum hijauan tinggi adalah menghambat pertumbuhan protozoa secara parsial (defaunasi parsial). Defaunasi parsial pada protozoa tersebut diharapkan dapat mengakibatkan peningkatan biomassa bakteri, sehingga terjadi peningkatan aktifitas fermentasi rumen. Beberapa penelitian telah dilakukan untuk
2 mengontrol populasi protozoa dengan berbagai agen defaunasi, salah satunya menggunakan senyawa saponin asal tanaman, yaitu tanaman teh (Hu et al., 2005), akasia (Santoso et al., 2007), dan lerak (Suharti et al., 2011).
Muntingia calabura atau yang biasa disebut dengan kersen (talok) merupakan tumbuhan yang mengandung saponin, tanin, dan flavonoid (Zakaria et al., 2010). Kandungan saponin dalam daun kersen tersebut diharapkan mampu menekan populasi protozoa. Pohon kersen sangat mudah ditemukan di Indonesia termasuk di kota-kota besar dan di tempat yang kering berkepanjangan. Menurut Figueiredo et al. (2008) pohon kersen merupakan tumbuhan yang mampu tumbuh cepat di lahan marginal sehingga mudah dikembangkan dan sangat berpotensi sebagai pakan ternak.
Sejauh ini penelitian tentang pemanfaatan daun kersen sebagai sumber pakan ruminansia belum ada. Oleh karena itu dilakukan analisis in vitro untuk mengetahui pengaruh pemberian daun kersen terhadap ternak.
Tujuan
Penelitian ini bertujuan untuk menganalisis kandungan fitokimia daun kersen yang meliputi senyawa tanin dan saponin serta mengevaluasi dosis penggunaan daun kersen yang tepat dan sesuai untuk mengoptimalkan kecernaan bahan kering, kecernaan bahan organik, produksi VFA, konsentrasi NH3, dan populasi protozoa.
3 TINJAUAN PUSTAKA
Kersen (Muntingia calabura)
Kersen berasal dari Amerika tropis dan banyak ditanam di kebun sebagai pohon peneduh. Kersen memiliki pohon yang kecil dengan tinggi 2-10 m. Rantingnya diselimuti rapat oleh rambut biasa yang halus dan oleh rambut kelenjar. Daunnya berseling, helaian daun tidak sama sisi, bulat telur bentuk lanset dengan ujung runcing bergerigi, berambut rapat terutama di bawah daun, lebarnya 4,5-14 kali 1,5-4 cm, tangkai daun pendek dan berambut seperti wol. Bunga berjumlah 1-3 menjadi satu di ketiak daun, berbilangan 5 dan berkelamin 2. Mahkota bunganya berbentuk bulat telur terbalik dan berwarna putih. Buahnya buni berwarna merah (Steenis, 2006). Taksonomi tumbuhan Muntingia calabura adalah sebagai berikut:
Kingdom : Plantae (Tumbuhan)
Subkingdom : Tracheobionta (Tumbuhan berpembuluh) Superdivisi : Spermatophyta (Menghasilkan biji) Divisi : Magnoliophyta (Tumbuhan berbunga) Kelas : Magnoliopsida (Dikotil/berkeping dua) Ordo : Malvales
Famili : Muntingiaceae Genus : Muntingia L. Spesies : M. calabura
Pohon kersen termasuk pohon yang tumbuh cepat dan dapat tumbuh subur di lahan marginal. Tumbuhan ini mampu melakukan penyerbukan sendiri secara spontan, mampu memproduksi buah dan biji dalam jumlah besar sepanjang tahun serta memiliki laju perkecambahan biji yang tinggi (Figueiredo et al., 2008). Daun kersen mengandung flavonoid, tanin, dan saponin (Zakaria et al., 2010).
Gambar 1. Daun Kersen (Muntingia calabura)
4 Saponin
Saponin termasuk ke dalam golongan glikosida yang terdapat pada tanaman tinggi dan dapat menimbulkan buih bila dikocok. Glikosida adalah suatu senyawa yang bila dihidrolisis akan terurai menjadi gula (glikon) dan senyawa lain (aglikon atau genin). Saponin memiliki rasa pahit atau getir dan dapat membentuk senyawa kompleks dengan kolesterol. Sebagian besar saponin bereaksi netral (larut dalam air), beberapa ada yang bereaksi asam (sukar larut dalam air), sebagian kecil ada yang bereaksi basa. Saponin berdasarkan struktur aglikonnya dapat dikelompokkan menjadi dua yaitu saponin sterol (steroid) dan saponin triterpen (triterpenoid). Saponin sterol bila dihidrolisis akan membentuk senyawa sterol, sedangkan saponin triperten bila dihidrolisis akan membentuk senyawa triterpen (Sirait, 2007).
Saponin mempunyai pengaruh yang lebih menguntungkan pada ternak ruminansia dibandingkan pada ternak non ruminansia. Pemberian bahan yang mengandung saponin dapat meningkatkan pertumbuhan, efisiensi pakan, dan kesehatan ternak dengan kemampuannya sebagai agen defaunasi. Kemampuan saponin sebagai agen defaunasi dapat menyebabkan penurunan total populasi protozoa rumen (Suparjo, 2008). Hal ini sesuai dengan penelitian sebelumnya yang menyatakan bahwa senyawa saponin asal tanaman yaitu teh (Hu et al., 2005) dan lerak (Suharti et al., 2010) dapat menekan pertumbuhan protozoa.
Gambar 2. Struktur Saponin Steroid Gambar 3. Struktur Saponin Triterpenoid
Sumber: Sirait (2007) Sumber: Sirait (2007)
Tanin
Tanin disebut juga zat samak yang memiliki sifat dapat menciutkan dan mengendapkan protein dari larutan dengan membentuk senyawa yang tidak larut (Sirait, 2007). Tanin merupakan polimer polifenolik yang dapat larut dalam air dengan berat molekuler yang relatif tinggi dan memiliki kemampuan untuk
HO 3 17 H3C CH3 CH3 HO 17 3
5 membentuk senyawa kompleks dengan protein membentuk kelompok fenolik hidroksil yang besar. Tanin banyak terdapat pada hijauan pohon yang memiliki nutrisi baik, semak belukar, dan kacang-kacangan, buah-buahan serta biji-bijian. Tanin dapat dikelompokkan menjadi dua yaitu tanin terhidrolisa dan tanin terkondensasi. Tanin terhidrolisa merupakan molekul kompleks dengan polyol sebagai intinya seperti glukosa, glusitol, asam quinic, quersitol, dan asam shikimic yang sebagian atau seluruhnya teresterifikasi dengan kelompok fenolik. Tanin terkondensasi merupakan sebagian besar dari polimer flavan-3-ol unit (epi)catechin dan (epi)gallocatechin yang berikatan dengan hubungan C4-C8 dan C4-C6 interflavoniod (Patra dan Saxena, 2010).
Gambar 4. Unit Monomer Tanin Terkondensasi (Catechin dan Gallocatechin) dan Terhidrolisis (Asam Gallic dan Ellagic)
Sumber: Patra dan Saxena (2010)
Pengaruh umum tanin terhadap ruminansia yaitu dapat menurunkan pemanfaatan nutrien secara in vivo terutama dalam pemanfaatan protein oleh ternak, menurunkan tingkat pertumbuhan ternak, menurunkan palatabilitas dan konsumsi atau menurunkan berbagai aktivitas enzim. Tanin dapat bersifat racun dan dapat mengakibatkan kematian jika dikonsumsi dalam jumlah yang besar (Makkar, 2003).
Catechin Asam Gallic
6 Mikroba Rumen
Perut ruminansia terdiri dari 4 bagian yaitu rumen, retikulum, omasum, dan abomasum. Rumen merupakan saluran fermentasi terbesar pada ruminansia yang di dalamnya terdapat beberapa jenis mikroba. Mikroba tersebut memiliki hubungan simbiosis mutualisme (saling menguntungkan) dengan tubuh inangnya (ruminansia). Mikroba rumen mendapatkan makanan dari pakan yang masuk ke saluran pencernaan inangnya, sementara mikroba tersebut membantu dalam mencerna pakan inangnya (Dehority, 2004). Mikroba rumen dibagi menjadi 4 kelompok utama yaitu bakteri, protozoa, jamur, dan bakteriophage atau virus. Disamping itu terdapat amoeba yang populasinya belum diketahui secara pasti (Soetanto, 2004). Faktor utama yang mempengaruhi pertumbuhan dan aktifitas populasi mikroba rumen adalah temperatur, pH, kapasitas buffer, tekanan osmotik, kandungan bahan kering, dan potensial oksidasi reduksi. Temperatur di dalam rumen relatif konstan yaitu 38-40oC sedangkan pH normal rumen sekitar 5,5-7,0 dengan komposisisi rasio pakan normal (Dehority, 2004).
Bakteri merupakan biomasa terbesar di dalam rumen, terdapat sekitar 50% dari total bakteri hidup bebas dalam cairan rumen serta sekitar 30%-40% menempel pada partikel makanan. Bakteri rumen punya peran yang besar dalam membantu mencerna pakan (Soetanto, 2004). Spesies-spesies bakteri dan protozoa yang berbeda saling berinteraksi di dalam rumen melalui hubungan simbiosis dan menghasilkan produk-produk yang khas seperti selulosa, hemiselulosa, dan pati melalui pencernaan polimer tumbuhan. Bakteri-bakteri tertentu yang bertanggung jawab terhadap proses fermentasi pregastric membentuk asetat, propionat, butirat, CO2, dan H2. Metanogen
mempergunakan CO2, H2, dan format untuk membentuk metana. Beberapa spesies
memproduksi amonia dan asam lemak terbang berantai cabang dari asam-asam amino tertentu. Beberapa organisme mengeluarkan urease untuk memecah urea sehingga menjadi amonia dan CO. Bakteri juga mensintesis vitamin-vitamin golongan B kompleks (Arora, 1989).
Protozoa bersifat anaerob. Apabila kadar oksigen maupun nilai pH isi rumen tinggi, maka protozoa tidak dapat membentuk cyste untuk mempertahankan diri dari lingkungan yang jelek, sehingga dengan cepat akan mati (Arora, 1989). Protozoa memiliki jumlah yang lebih sedikit daripada bakteri. Protozoa memiliki ukuran tubuh
7 lebih besar sehingga total biomasanya hampir sama dengan bakteri (McDonald et al., 2002). Jumlah protozoa dalam rumen sangat beragam menurut jenis makanan, umur, dan jenis hewan yang menjadi inangnya. Biasanya jumlah protozoa ciliata adalah 105 per ml pada makanan berserat kasar tinggi, namun jumlah ini meningkat menjadi 106 per ml pada adaptasi terhadap gula-gula terlarut (Arora, 1989).
Banyak bukti yang menunjukkan bahwa interaksi antara bakteri dan protozoa di dalam rumen lebih bersifat kompetitif. Protozoa memangsa bakteri yang terdapat pada cairan rumen dan mencernanya sebagai sumber asam amino bagi pertumbuhannya. Akibatnya populasi bakteri akan berkurang sehingga laju kolonisasi partikel makanan di dalam rumen akan berkurang juga (Soetanto, 2004). Setiap individu protozoa dapat memangsa bakteri dengan kecepatan antara 60-700 bakteri/protozoa/jam pada kondisi kepadatan bakteri 109 sel/ml (Arora, 1989). Pencernaan bakteri dalam sel protozoa berkisar antara 345–1200 bakteri/protozoa/jam. Populasi jamur rumen (zoospores) telah dilaporkan meningkat setelah defaunasi (menghilangkan protozoa dari rumen) yang dapat meningkatkan daya cerna serat kasar secara nyata 6-10 unit/24 jam. Disamping itu jumlah bakteri juga meningkat apabila protozoa dihilangkan dari rumen (Soetanto, 2004).
Amonia (NH3)
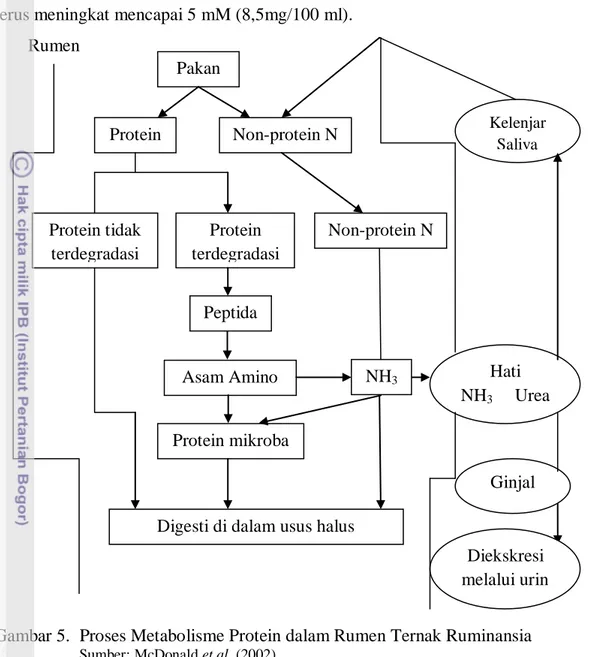
Seluruh protein yang terkandung dalam pakan akan dihidrolisa oleh mikroba rumen menjadi asam amino. Hidrolisa protein menjadi asam amino tersebut diikuti oleh proses deaminasi untuk membebaskan amonia. Sumber lain amonia adalah dari hidrolisa urea atau garam-garam amonium lain yang dapat berasal dari saliva maupun pakan (Arora, 1989). Proses metabolisme protein pada rumen ruminansia dapat dilihat pada Gambar 5.
Amonia merupakan sumber nitrogen utama yang penting untuk sintesis protein mikroba. Bakteri yang mampu memanfaatkan amonia sebagai sumber energi disebut bakteri penitrifikasi (Dehorty, 2004). Menurut Arora (1989), amonia yang dibebaskan dari urea atau garam-garam amonium tersebut dapat menggantikan protein makanan sampai 50%. Amonia dibebaskan di dalam rumen selama proses fermentasi dalam bentuk ion NH4 maupun dalam bentuk tak terion sebagai NH3.
Sebanyak 82% mikroba rumen dapat tumbuh dengan NH4+ sebagai sumber N dan
8 amonia dalam cairan rumen adalah penting karena pemakaian amonia oleh mikroba terus meningkat mencapai 5 mM (8,5mg/100 ml).
Gambar 5. Proses Metabolisme Protein dalam Rumen Ternak Ruminansia
Sumber: McDonald et al. (2002)
Amonia yang dihasilkan dari proses hidrolisa protein tidak semuanya disintesis menjadi protein mikroba, namun sebagian akan diserap ke dalam darah. Amonia yang tidak terpakai akan dibawa ke hati yang diubah menjadi urea, sebagian dikeluarkan melalui urin serta yang lain dibawa ke kelenjar saliva. Konsentrasi amonia yang optimum dalam menunjang proses sintesis mikroba sangat bervariasi berkisar antara 6-21 mM. Faktor utama yang mempengaruhi penggunaan amonia yaitu ketersediaan karbohidrat dalam ransum yang berfungsi sebagai energi untuk pembentukan protein mikroba (McDonald et al., 2002).
Kadar amonia dalam rumen merupakan petunjuk antara proses degradasi dan sintesis protein oleh mikroba di dalam rumen. Pakan yang defisien akan protein atau
Protein mikroba Non-protein N Protein terdegradasi Peptida NH3 Non-protein N Asam Amino Kelenjar Saliva Hati NH3 Urea Ginjal Diekskresi melalui urin Pakan Protein Protein tidak terdegradasi
Digesti di dalam usus halus Rumen
9 proteinnya tahan degradasi memiliki konsentrasi amonia yang rendah dalam rumen serta pertumbuhan mikroba rumen akan lambat yang menyebabkan turunnya kecernaan pakan (McDonald et al., 2002).
Volatile Fatty Acid (VFA)
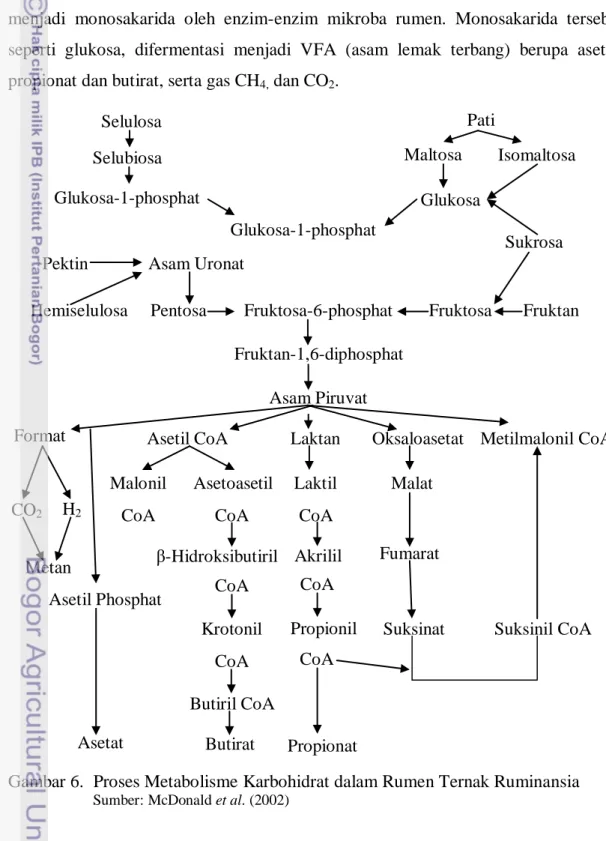
Karbohidrat yang terkandung dalam ransum pakan akan dipecah di dalam rumen ruminansia. Karbohidrat yang berbentuk polisakarida akan dihidrolisa menjadi monosakarida oleh enzim-enzim mikroba rumen. Monosakarida tersebut seperti glukosa, difermentasi menjadi VFA (asam lemak terbang) berupa asetat, propionat dan butirat, serta gas CH4, dan CO2.
Gambar 6. Proses Metabolisme Karbohidrat dalam Rumen Ternak Ruminansia
Sumber: McDonald et al. (2002)
Suksinil CoA Laktil Laktan CoA Asetil CoA Asetoasetil Malonil CoA CoA CoA Krotonil CoA Butiril CoA Butirat Selulosa Selubiosa Glukosa Glukosa-1-phosphat Glukosa-1-phosphat Pati Maltosa Isomaltosa Sukrosa Pektin Asam Uronat
Pentosa
Hemiselulosa Fruktosa-6-phosphat Fruktosa Fruktan Fruktan-1,6-diphosphat Asam Piruvat Format CO2 H2 Metan Asetil Phosphat Asetat Akrilil β-Hidroksibutiril CoA Propionil CoA Propionat Oksaloasetat Malat Fumarat Suksinat Metilmalonil CoA
10 VFA akan diserap melalui dinding rumen, sementara gas CH4 dan CO2 akan
hilang melalui proses eruktasi. Sekitar 75% dari produksi total VFA yang akan diserap langsung oleh retikulo-rumen yang masuk ke darah, sekitar 20% VFA diserap di abomasum dan omasum serta sekitar 5% diserap oleh usus halus (McDonald et al., 2002).
Arora (1989) menyatakan bahwa VFA merupakan sumber energi utama bagi ruminansia. Glukosa pada ruminansia diabsorpsi dari saluran pencernaan dalam jumlah kecil, dan kadarnya di dalam darah dipertahankan melalui sintesa endogenous untuk keperluan fungsi-fungsi esensial jaringan tubuh. Produksi VFA cairan rumen berkisar antara 70-150 mM. Konsentrasi VFA yang dihasilkan tergantung dari jenis pakan yang dikonsumsi ternak (McDonald et al., 2002). Proses fermentasi karbohidrat pada rumen ternak ruminansia dapat dilihat pada Gambar 6.
Kecernaan Bahan Kering dan Bahan Organik
Kecernaan pakan dapat didefinisikan sebagai bagian pakan yang tidak diekskresikan dalam feses sehingga diasumsikan bagian tersebut diserap oleh tubuh ternak. Kecernaan dinyatakan dengan dasar bahan kering (McDonald et al., 2002). Tingkat pencernaan merupakan fungsi waktu pencernaan dan waktu yang tersedia bagi pakan untuk dicerna. Absorpsi total neto pakan ditentukan oleh selisih antara yang masuk (inflow) dan yang keluar (outflow) pada setiap seksi saluran pencernaan. Kecernaan dapat dihitung dengan berbagai teknik. Salah satunya adalah teknik in vitro yang harus mirip dengan in vivo agar sedapat mungkin menghasilkan pola yang sama. Kadar substrat harus berkisar antara 0,5-0,15 gram per pakan hijauan sehingga cukup untuk menjalankan kecepatan fermentasi yang normal di dalam sistem pencernaan. Media harus mengandung sumber energi, namun di dalam evaluasi makanan hijauan sumber energi tersebut dihindari dan tidak ditambahkan ke dalam sistem. Substrat penting lainnya yang diperlukan dalam sistem in vitro ini adalah sumber nitrogen (Arora, 1989).
Tilley dan Terry (1963) memperkenalkan metode two stage yang paling banyak digunakan untuk mengukur kecernaan secara in vitro. Tahap pertama dari metode ini adalah inkubasi dalam larutan buffer cairan rumen selama 48 jam dalam kondisi anaerob. Tahap kedua adalah pemberian pepsin dan inkubasi selama 48 jam.
11 MATERI DAN METODE
Lokasi dan Waktu
Penelitian ini dilaksanakan dari bulan Maret sampai Mei 2012. Analisis fitokimia dilakukan di Laboratorium Balai Penelitian Ternak Ciawi, Bogor. Analisis proksimat dilakukan di Laboratorium Pusat Penelitian Sumberdaya Hayati dan Bioteknologi, Institut Pertanian Bogor. Analisis konsentrasi amonia, produksi Volatile Fatty Acid (VFA) total, dan nilai kecernaan dilakukan di Laboratorium Nutrisi Ternak Perah Fakultas Peternakan, Institut Pertanian Bogor. Analisis populasi protozoa dilakukan di Laboratorium Biokimia, Fisiologi, dan Mikrobiologi Nutrisi Fakultas Peternakan, Institut Pertanian Bogor.
Materi
Alat
Peralatan yang digunakan dalam penelitian ini antara lain oven 60oC, grinder, timbangan digital, sudib, termos, kain penyaring, corong, tabung fermentor, tutup karet, pipet volumetrik, labu Erlenmeyer, bulp, botol film, freezer, kulkas, spoit, jarum suntik, shaker water bath, tabung gas CO2, selang, pH meter, timbangan
analitik, sentrifuge, alat destilasi VFA, magnetic stirer, buret, pipet mikro, cawan Conway, ruang asam, oven 105oC, tanur, eksikator, gegep, cawan porselen, pompa vakum, mikroskop, dan Fuchs Rosenthal Counting Chamber.
Bahan
Bahan yang digunakan dalam penelitian antara lain cairan rumen yang diambil dari Rumah Pemotongan Hewan (RPH) Bubulak Bogor, aquades, larutan McDougall, Trifanblue Formalsalin (TBFS), kertas saring, gas CO2, HgCl2 jenuh,
larutan Na2CO3 jenuh, asam borat, vaselin, larutan H2SO4 0,005 N, larutan H2SO4
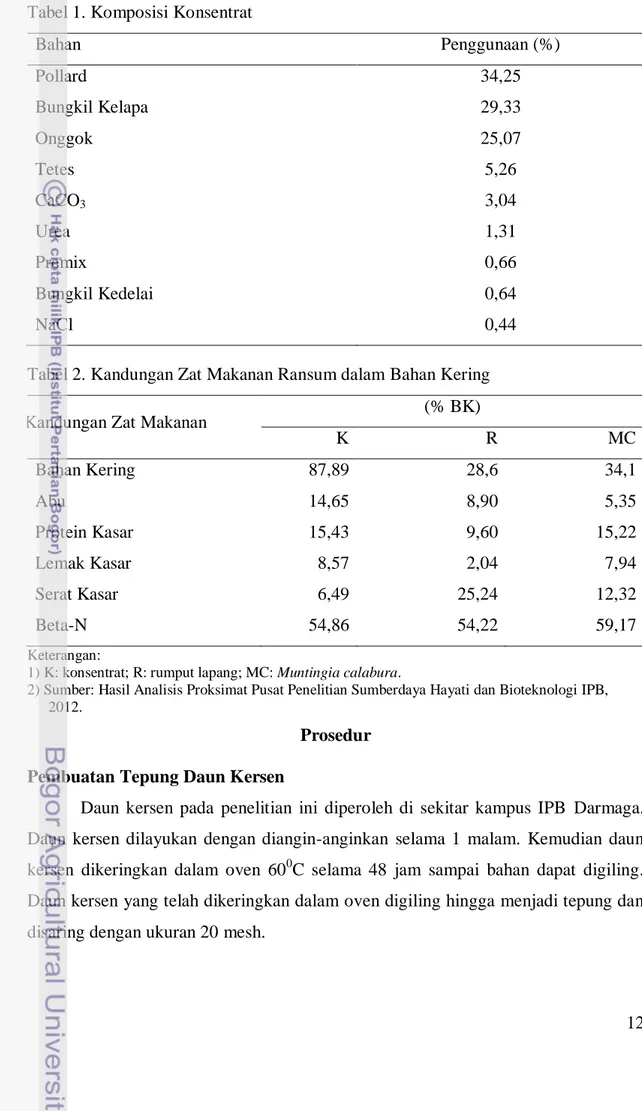
15%, larutan NaOH 0,5 N, indikator phenolphthalein, larutan HCl 0,5 N, dan pepsin-HCl 0,2%. Bahan yang digunakan dalam pembuatan larutan McDougall dan TBFS serta cara pembuatannya dicantumkan dalam prosedur penelitian. Bahan pakan yang digunakan berupa ransum komplit berbentuk mash dengan perbandingan hijauan dan konsentrat 70% : 30%. Hijauan yang digunakan dalam penelitian ini terdiri dari rumput lapang dan daun kersen. Komposisi dan kandungan zat makanan ransum dapat dilihat pada Tabel 1 dan Tabel 2.
12 Tabel 1. Komposisi Konsentrat
Bahan Penggunaan (%) Pollard 34,25 Bungkil Kelapa 29,33 Onggok 25,07 Tetes 5,26 CaCO3 3,04 Urea 1,31 Premix 0,66 Bungkil Kedelai 0,64 NaCl 0,44
Tabel 2. Kandungan Zat Makanan Ransum dalam Bahan Kering
Kandungan Zat Makanan (% BK)
K R MC Bahan Kering 87,89 28,6 34,1 Abu 14,65 8,90 5,35 Protein Kasar 15,43 9,60 15,22 Lemak Kasar 8,57 2,04 7,94 Serat Kasar 6,49 25,24 12,32 Beta-N 54,86 54,22 59,17 Keterangan:
1) K: konsentrat; R: rumput lapang; MC: Muntingia calabura.
2) Sumber: Hasil Analisis Proksimat Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB, 2012.
Prosedur
Pembuatan Tepung Daun Kersen
Daun kersen pada penelitian ini diperoleh di sekitar kampus IPB Darmaga. Daun kersen dilayukan dengan diangin-anginkan selama 1 malam. Kemudian daun kersen dikeringkan dalam oven 600C selama 48 jam sampai bahan dapat digiling. Daun kersen yang telah dikeringkan dalam oven digiling hingga menjadi tepung dan disaring dengan ukuran 20 mesh.
13 Analisis Proksimat dan Fitokimia
Analisis proksimat dilakukan di Pusat Penelitian Sumberdaya Hayati dan Bioteknologi IPB. Analisis fitokimia dilakukan di Laboratorium Balai Penelitian Ternak Ciawi, Bogor. Analisis saponin menggunakan metode uji spektrofotometri. Analisis tanin berdasarkan metode Folin-Ciocalteu (Makkar, 2003).
Pembuatan Larutan McDougall
Pembuatan larutan McDougall berdasarkan Tilley dan Terry (1963). Bahan yang digunakan untuk pembuatan larutan McDougall sebanyak 1 liter ditimbang dan dimasukkan ke dalam labu Erlenmeyer. Bahan-bahan tersebut adalah NaHCO3 (9,8
gram), Na2HPO4.7H2O (7 gram), KCl (0,57 gram), NaCl (0,47 gram), dan
MgSO4.7H2O (0,12 gram). Kemudian ditambahkan aquades sampai kira-kira 900 ml
dan diaduk dengan magnetic stirer. Setelah larut ditambahkan CaCl2 (0,04 gram),
ditambahkan aquades sampai mencapai tanda tera 1 liter, dan dilarutkan kembali sampai semua bahan larut.
Pengambilan Cairan Rumen
Cairan rumen diambil dari Rumah Pemotongan Hewan Bubulak Bogor. Cairan rumen tersebut diambil dari dua ekor sapi yang dimasukkan dalam satu termos. Menurut Mauricio (2001), cairan rumen diambil dari 2 ekor ternak. Suhu di dalam termos harus disesuaikan suhu rumen dengan diisi oleh air panas sebelum diisi cairan rumen. Kemudian air panas dalam termos dibuang terlebih dahulu sebelum diisi cairan rumen. Isi rumen diperas dan disaring lalu dimasukkan ke dalam termos dan segera dibawa ke laboratorium untuk dilakukan fermentasi in vitro. Pengambilan cairan rumen dilakukan sebanyak 4 kali.
Fermentasi In Vitro
Metode in vitro yang digunakan adalah metode dua tahap (two-stage method) (Tilley dan Terry, 1963). Sebanyak 0,5 gram sampel sesuai perlakuan dimasukkan dalam tabung fermentor, ditambahkan 40 ml larutan McDougall dan 10 ml cairan rumen. Kemudian dikocok dengan gas CO2 selama 30 detik untuk menciptakan
suasana anaerob dan ditutup dengan tutup karet berventilasi. Tabung dimasukan ke dalam shaker water bath dengan suhu 39oC. Proses fermentasi dihentikan setelah inkubasi selama 4 jam dan 48 jam. Tutup tabung dibuka kemudian ditambahkan 2
14 tetes HgCl2 jenuh untuk menghentikan aktivitas mikroba. Sampel disentrifuge
dengan kecepatan 3000 rpm selama 15 menit. Supernatan yang diperoleh pada tabung inkubasi 4 jam diambil untuk analisis produksi VFA total dan konsentrasi NH3. Supernatan yang diperoleh pada tabung inkubasi 48 jam dibuang dan diambil
endapannya. Endapan tersebut yang akan digunakan dalam analisis KCBK dan KCBO.
Nilai pH
Nilai pH rumen diukur pada sampel yang telah diinkubasi selama 4 jam. Nilai pH tersebut diukur dengan menggunakan pH meter sesudah tabung diangkat dari shaker waterbath dan sebelum ditetesi dengan HgCl2.
Populasi Protozoa
Cairan rumen yang telah diinkubasi selama 4 jam, sebelum diangkat dari shaker waterbath dan sebelum ditetesi HgCl2 diambil sebanyak 1 ml. Kemudian
dimasukkan ke dalam botol film yang berisi Trifanblue Formalsalin (TBFS) sebanyak 1 ml. TBFS dibuat dari campuran 4% formalin dengan NaCl Fisiologis 0,9% dalam 100 ml larutan. Perhitungan populasi protozoa dilakukan menurut metode Ogimoto dan Imai (1981) dengan menggunakan Fuchs Rosenthal Counting Chamber. Populasi protozoa diamati dengan mikroskop lensa obyektif dengan pembesaran 40x dan okuler 10x. Populasi protozoa dihitung dengan rumus :
Populasi protozoa = 1
0,2 x 0,0625 x 16 x 16 x 1000 x C x Fp Keterangan : C = jumlah koloni yang dihitung
Fp = faktor pengencer ( 2 ) Konsentrasi NH3
Konsentrasi NH3 (amonia) diukur dengan metode mikrodifusi Conway
(Conway, 1962). Sebanyak 1 ml supernatan dipipet dan dibubuhkan disebelah kanan sekat dan 1 ml larutan Na2CO3 jenuh disebelah kiri cawan Conway, sedangkan
cawan kecil ditengah diisi dengan 1 ml asam borat berindikator. Cawan Conway tersebut ditutup rapat dengan tutup yang telah diolesi vaselin, kemudian digerakkan perlahan sehingga supernatan dan larutan Na2CO3 bercampur. Cawan Conway
dibiarkan selama 24 jam dalam suhu kamar. Amonia yang dihasilkan dari reaksi antara supernatan dan larutan Na2CO3 diikat oleh asam borat sehingga warna merah
15 asam borat berubah menjadi ungu kebiruan. Kemudian dititrasi dengan larutan H2SO4 0,005 N. Titrasi dihentikan setelah terjadi perubahan warna dari ungu
kebiruan menjadi merah kembali. Konsentrasi NH3 dihitung dengan rumus:
NH3 (mM) =Volume H2SO4 x N. H2SO4 x 1000 Berat sampel x BK sampel Produksi VFA Total
Produksi VFA total diukur dengan menggunakan metode destilasi uap (General Laboratory Procedures, 1966). Sebanyak 5 ml supernatan dipipet kedalam tabung penyulingan khusus, kemudian ditambahkan 1 ml larutan H2SO4 15%. Hasil
penyulingan ditampung dengan labu Erlenmeyer yang berisi 5 ml larutan NaOH 0,5 N, sampai kira-kira tertampung sebanyak 250 ml. Kemudian ditambah 2-3 tetes indikator phenolphthalein dan dititrasi dengan larutan HCl 0,5 N. Titrasi dihentikan setelah terjadi perubahan warna dari merah jambu menjadi jernih. Produksi VFA total yang dihasilkan, dihitung dengan rumus :
Produksi VFA (mM) =[(a − b) x N HCl x 1000/5] Berat sampel x BK sampel a = ml titran blanko
b = ml titran sampel KCBK dan KCBO
Supernatan yang diperoleh pada tabung inkubasi 48 jam dibuang dan diambil endapannya. Endapan tersebut ditambahkan dengan larutan pepsin-HCl 0,2% sebanyak 50 ml dan diinkubasi lagi selama 48 jam tanpa tutup karet. Kemudian endapan disaring dengan kertas saring Whatman No.41 dengan bantuan pompa vakum. Hasil saringan di oven 105oC selama 24 jam sehingga diperoleh bahan kering. Selanjutnya dimasukkan ke dalam tanur 600oC selama 6 jam sehingga diperoleh bahan organik (Tilley & Terry, 1963). Kecernaan bahan kering dan bahan organik diperoleh dengan rumus:
% KCBK =BK asal − BK residu − BK blanko
BK asal x 100%
% KCBO =BO asal − (BO residu – BO blanko)
16 Rancangan
Perlakuan
Penelitian ini menggunakan 5 taraf perlakuan ransum dan 4 kelompok. Perlakuan yang digunakan adalah :
R0 : 30% konsentrat : 70% rumput lapang : 0% daun kersen R1 : 30% konsentrat : 65% rumput lapang : 5% daun kersen R2 : 30% konsentrat : 60% rumput lapang : 10% daun kersen R3 : 30% konsentrat : 55% rumput lapang : 15% daun kersen R4 : 30% konsentrat : 50% rumput lapang : 20% daun kersen Peubah
Peubah yang diamati pada penelitian ini adalah nilai pH, populasi protozoa, konsentrasi NH3 (Amonia), produksi VFA total, KCBK (Kecernaan Bahan Kering),
dan KCBO (Kecernaan Bahan Organik). Model
Rancangan percobaan yang digunakan adalah Rancangan Acak Kelompok (RAK) dengan 5 taraf perlakuan ransum dan 4 kelompok sebagai ulangan. Pengelompokan berdasarkan waktu pengambilan cairan rumen. Model matematika yang digunakan adalah sebagai berikut :
Yij = µ + βi + τj + εij
Keterangan :
Yij : nilai pengamatan perlakuan ke-i, blok ke-j
µ : rataan umum βi : efek perlakuan ke-i
τj : efek blok ke-j
εij : galat perlakuan ke-i dan blok ke-j
Analisis Data
Data yang diperoleh dianalisis dengan sidik ragam (ANOVA) dan jika berbeda nyata pada level P<0,05 dilakukan Uji Jarak Duncan. Analisis data dilakukan menggunakan software statistik SPSS versi 16.
17 HASIL DAN PEMBAHASAN
Daun Kersen sebagai Pakan
Peningkatan produksi daging lokal dengan mengandalkan peternakan rakyat menghadapi permasalahan dalam hal pakan. Pakan yang digunakan oleh peternak rakyat memiliki kualitas yang rendah karena menggunakan hijauan lebih banyak (hampir 100%), sehingga dapat mengakibatkan defisiensi nutrien. Salah satu upaya untuk mengatasi permasalahan tersebut adalah dengan mencari bahan pakan yang mudah didapat dan memiliki kemampuan meningkatkan efisiensi fermentasi pakan. Muntingia calabura (kersen) merupakan salah satu tumbuhan yang belum digunakan sebagai pakan dan memiliki kemungkinan dapat dijadikan sebagai sumber pakan. Hal tersebut karena daun kersen mengandung saponin, tanin, dan flavonoid (Zakaria et al., 2010). Kandungan saponin dalam daun kersen diduga dapat meningkatkan efisiensi fermentasi rumen pada ruminansia melalui defaunasi parsial pada protozoa. Selain itu daun kersen memiliki kandungan protein yang lebih tinggi (15,22%) daripada rumput lapang (9,60%). Kandungan fitokimia daun kersen yang meliputi saponin dan tanin dapat dilihat pada Tabel 3, sementara analisis proksimat daun kersen terdapat pada Tabel 2.
Tabel 3. Kandungan Fitokimia Daun Kersen
Sampel Tanin (%) Saponin (%)
Daun kersen 1,41 10,28
Keterangan: Analisis di Laboratorium Balai Penelitian Ternak, Ciawi Bogor (2012)
Daun kersen sangat aplikatif, karena dapat diberikan dalam bentuk segar sehingga tidak memerlukan pengolahan dan memudahkan peternak rakyat. Daun kersen juga mudah ditemukan pada berbagai kondisi lahan. Menurut Figueiredo et al. (2008) pohon kersen merupakan tumbuhan yang mampu tumbuh cepat di lahan marginal sehingga disebut sebagai tanaman perintis. Hal tersebut membuat daun kersen mudah untuk dikembangbiakkan. Data hasil analisis populasi protozoa dan karakteristik fermentasi rumen secara in vitro dapat dilihat pada Tabel 4.
18 Tabel 4. Pengaruh Perlakuan terhadap Populasi Protozoa dan Karakteristik
Fermentasi
Keterangan:
1) Superskrip berbeda pada baris yang sama menunjukkan berbeda nyata (P<0,05) 2) R0: 30% K + 70% R
R1: 30% K + 65% R + 5% MC R2: 30% K + 60% R + 10% MC R3: 30% K + 55% R + 15% MC R4: 30% K + 50% R + 20% MC
3) K: konsentrat; R: rumput lapang; MC: Muntingia calabura
Nilai pH Rumen
Nilai pH rumen yang dihasilkan menunjukkan kondisi rumen yang sesuai atau tidak sesuai bagi pertumbuhan mikroba rumen. Hasil analisis ragam menunjukkan bahwa penggunaan daun kersen tidak mempengaruhi nilai pH rumen (Tabel 4). Nilai pH yang sama dengan kontrol menunjukkan bahwa penggunaan daun kersen tidak mengganggu aktivitas fermentasi rumen. Rumen pada kondisi normal memiliki pH 6,8 (Arora, 1989). Menurut Dehority (2004), pH normal rumen sekitar 5,5-7,0 dengan pemberian rasio pakan normal.
Nilai pH sebenarnya menggambarkan jumlah asam yang diproduksi oleh mikroflora yang ada di dalam rumen. Nilai pH minimum umumnya dicapai sekitar 2-6 jam setelah makan, sesuai dengan produksi asam yang maksimum. Perubahan pH dipengaruhi oleh waktu setelah makan, sifat pakan, dan frekuensi makan ternak (Dehority, 2004).
Populasi Protozoa
Populasi protozoa yang bersilia lebih dominan di dalam rumen, sementara populasi protozoa yang berflagel hanya sedikit (Dehority, 2004). Protozoa yang bersilia berkembang di dalam rumen ternak ruminansia secara alami, dan membantu pencernaan zat-zat makanan dari rumput-rumputan yang kaya akan serat kasar
Peubah Perlakuan R0 R1 R2 R3 R4 pH 6,86+0,09 6,84+0,05 6,85+0,07 6,81+0,06 6,83+0,07 Protozoa (Log 10/ml) 3,97+ 0,49 3,94+ 0,69 3,68 + 0,33 3,72 + 0,22 3,57 + 0,39 N-NH3 (mM) 6,93+ 1,05c 6,35+ 1,75bc 5,52+ 1,09bc 5,37+ 0,95ab 5,25+ 0,41a VFA Total (mM) 89,98+ 30,35a 104,07+ 33,55b 109,01+ 27,94b 112,41+ 24,44b 150,62+ 33,51c KCBK (%) 63,72+ 5,47b 57,84+ 2,22ab 54,28+ 9,93a 50,57+ 4,14a 53,69+ 6,85a KCBO (%) 65,07+ 5,11b 58,21+ 3,70ab 56,06+ 8,62a 50,62+ 5,85a 54,72+ 4,99a
19 (Arora, 1989). Namun protozoa juga bersifat merugikan karena sifatnya yang memangsa bakteri, akibatnya biomassa bakteri akan berkurang sehingga laju degradasi pakan dan suplai protein mikroba akan berkurang pula (Soetanto, 2004).
Hasil analisis ragam menunjukkan bahwa penggunaan daun kersen hingga taraf 20% tidak nyata mempengaruhi populasi protozoa. Hasil tersebut tidak sama dengan penelitian sebelumnya yang menyatakan bahwa saponin asal tanaman dalam bentuk ekstrak yaitu Yucca schidigera (Pen et al., 2006) dan lerak (Suharti et al., 2011) dapat menekan populasi protozoa dalam rumen. Hal ini diduga karena penelitian ini menggunakan sumber saponin yang masih dalam bentuk tepung, sehingga banyak sanyawa lain yang mengikat saponin dan mengakibatkan belum efektif menekan protozoa. Disamping itu perbedaan tersebut diduga pula disebabkan oleh adanya perbedaan tipe dan asal saponin yang terdapat pada masing-masing tanaman, seperti yang dilaporkan oleh Pen et al. (2006).
Pengambilan sampel pada inkubasi 4 jam diduga juga dapat mempengaruhi keefektifan saponin dalam daun kersen untuk menekan populasi protozoa. Penelitian Suharti et al. (2011) memperlihatkan adanya perbedaan populasi protozoa antara waktu inkubasi 4 jam dan 24 jam. Waktu inkubasi 24 jam memperlihatkan penurunan yang signifikan. Protozoa akan melalui fase cair yang mengalami pergantian setiap 24 jam sekali, sehingga organisme yang ada di dalam rumen harus memiliki kecepatan pertumbuhan 16,6 jam atau lebih kecil untuk mempertahankan diri. Kecepatan pertumbuhan yang dibutuhkan protozoa antara 7-11 jam untuk mempertahankan diri di dalam rumen (Dehority, 2004).
Konsentrasi protozoa dalam rumen sapi maupun domba pada kondisi normal sekitar 1x106/ml (Dehority, 2004). Jumlah tersebut berbeda dengan populasi protozoa yang dihasilkan dalam penelitian ini yaitu sekitar 103/ml. Hal tersebut karena populasi protozoa pada penelitian in vitro lebih sedikit jika dibandingkan dengan penelitian in vivo, karena pada penelitian in vitro terdapat pengenceran oleh larutan McDougall. Jumlah tersebut masih dikatakan normal karena ransum hijauan yang digunakan mencapai 70%. Hasil tersebut hampir sama dengan penelitian Suharti et al. (2011) yang menggunakan ransum hijauan 70% dengan jumlah protozoa 104/ml serta penelitian Hess et al. (2003) yang menggunakan ransum hijauan 100% dengan jumlah protozoa 103/ml. Populasi protozoa dipengaruhi oleh
20 waktu setelah makan (waktu inkubasi) dan rasio pakan hijauan dan konsentrat (Dehority, 2004). Perbedaan rasio antara hijauan dan konsentrat menghasilkan populasi protozoa yang berbeda dalam penelitian Suharti et al. (2011).
Kandungan tanin dalam daun kersen diduga juga dapat mempengaruhi populasi protozoa dalam rumen. Tan et al. (2011) menyatakan bahwa penggunaan ekstrak tanin terkondensasi dari tanaman Leucaena mampu menurunkan populasi protozoa secara in vitro. Sementara pengaruh senyawa flavonoid dalam daun kersen terhadap populasi protozoa rumen belum diketahui, seperti yang telah dilaporkan oleh Patra dan Saxena (2010).
Konsentrasi Amonia (NH3)
Konsentrasi amonia dalam rumen menunjukkan banyaknya kandungan protein kasar yang dirombak oleh mikroba. Perubahan konsentrasi amonia menggambarkan efektivitas proses fermentasi. Syahrir et al. (2008) menyatakan bahwa konsentrasi amonia yang rendah dalam cairan rumen dapat menggambarkan proses fermentasi yang berjalan baik sehingga amonia dimanfaatkan dengan baik, protein ransum yang sulit didegradasi atau kandungan protein ransum yang rendah.
Penggunaan daun kersen sampai dengan level 20% dalam ransum sangat nyata (P<0,01) menurunkan konsentrasi amonia (Tabel 4). Pemberian daun kersen pada level yang semakin tinggi akan semakin menurunkan konsentrasi amonia. Kandungan protein daun kersen lebih tinggi jika dibandingkan dengan rumput lapang, maka seharusnya semakin tinggi penggunaan daun kersen dan semakin rendah penggunaan rumput lapang akan meningkatkan konsentrasi amonia. Hal tersebut diduga karena pengaruh tanin yang terkandung dalam daun kersen. Telah diketahui bahwa protein dan tanin dapat membentuk ikatan kompleks yang tidak dapat dihidrolisa di dalam sistem pencernaan fermentatif. Menurut Arora (1989) tanin pada hijauan biasanya dari jenis terkondensasi sehingga resisten terhadap hidrolisa. Hasil tersebut sama dengan penelitian Santoso et al. (2007) yang menggunakan Acacia mangium dengan kandungan tanin 4,51% mampu menurunkan konsentrasi amonia rumen. Kandungan tanin dalam daun kersen yang hanya 1,41% bisa jadi sama efektifnya dengan tanin dari Acacia mangium dalam membentuk ikatan kompleks dengan protein. Menurut Jayanegara dan Sofyan (2008) hubungan
21 antara kandungan tanin hijauan (total fenol, total tanin, dan tanin terkondensasi) belum tentu linier dengan aktivitas biologis tanin.
Rataan konsentrasi amonia yang dihasilkan (Tabel 4) menunjukkan bahwa ransum kontrol dan ransum penggunaan daun kersen pada level 5% memiliki nilai konsentrasi amonia yang optimal bagi pertumbuhan mikroba rumen. Hal ini sesuai dengan McDonald et al. (2002) yang menyatakan bahwa konsentrasi amonia yang optimal untuk menunjukkan sintesis protein mikroba berkisar 6-21 mM. Konsentrasi amonia yang dihasilkan oleh penggunaan daun kersen pada level 10%, 15%, dan 20% masih dapat menunjang pertumbuhan mikroba rumen. Menurut Sutardi (1980) konsentrasi amonia yang dibutuhkan untuk menunjang pertumbuhan mikroba rumen berkisar antara 4-12 mM.
Produksi VFA Total
VFA merupakan produk akhir fermentasi karbohidrat oleh mikroba rumen serta sebagai sumber energi utama bagi ternak ruminansia. Produksi VFA yang utama yaitu asam asetat, propionat, dan butirat yang dianggap sebagai faktor utama dalam mempengaruhi produksi ternak ruminansia (McDonald et al., 2002). Peningkatan produksi VFA menunjukkan mudah atau tidaknya pakan difermentasi oleh mikroba rumen.
Penggunaan daun kersen sampai level 20% sangat nyata (P<0,01) meningkatkan produksi VFA (Tabel 4). Hal ini menunjukkan bahwa semakin tinggi level daun kersen yang diberikan maka semakin tinggi pula produksi VFA total. Produksi VFA total berkaitan dengan lama inkubasi yang dilakukan yaitu selama 4 jam. Telah diketahui bahwa pada awal fermentasi, mikroba rumen memfermentasi bahan yang paling mudah tercerna. Peningkatan produksi VFA ini diduga karena proporsi karbohidrat yang mudah tercerna pada daun kersen lebih tinggi daripada rumput lapang (Tabel 2). Hal tersebut dapat dilihat dari kandungan Beta-N (karbohidrat mudah tercerna) dalam daun kersen yang lebih tinggi bila dibandingkan dengan rumput lapang. Selain itu peningkatan produksi VFA total yang diikuti dengan rendahnya konsentrasi amonia merupakan gambaran efisiensi penggunaan amonia oleh bakteri untuk mensintesis protein mikroba. Selanjutnya bakteri tersebut akan mencerna pakan untuk memproduksi VFA yang dapat dimanfaatkan sebagai
22 sumber energi bagi induk semang dan sumber karbon bakteri itu sendiri (Syahrir et al., 2009).
Rataan nilai produksi VFA yang dihasilkan meningkat dari 89,98 (ransum kontrol) sampai 150,62 (level daun kersen 20%). Nilai konsentrasi tersebut berada dalam kisaran normal dalam mendukung sintesis protein mikroba. McDonald et al. (2002) menyatakan bahwa produksi VFA total yang dapat mendukung proses sintesis protein mikroba adalah 70-150 mM.
Kecernaan Bahan Kering dan Bahan Organik
Kecernaan pakan dapat didefinisikan sebagai bagian pakan yang tidak diekskresikan dalam feses sehingga diasumsikan bagian tersebut diserap oleh tubuh ternak. Kecernaan dinyatakan dengan dasar bahan kering (McDonald et al., 2002). Kecernaan zat makanan dari suatu pakan menunjukkan kualitas pakan.
Penggunaan daun kersen nyata (P<0,05) menurunkan kecernaan bahan kering dan kecernaan bahan organik (Tabel 4). Nilai kecernaan yang dihasilkan merupakan nilai kecernaan yang rendah. Sutardi (1980) menyatakan bahwa nilai kecernaan bahan pakan >60% dikatakan memiliki nilai kecernaan yang tinggi. Rendahnya nilai kecernaan yang dihasilkan dan penurunan nilai kecernaan diduga karena pengaruh tanin dalam daun kersen yang mampu berikatan dengan protein pakan. Penambahan tanin murni pada level 0,5 mg/ml dalam ransum secara nyata menurunkan kecernaan bahan organik (Jayanegara et al., 2009). Ikatan yang terbentuk antara tanin dan protein akan berpengaruh terhadap kecernaan pakan (Mueller, 2006). Menurut Arora (1989) tanin dalam hijauan mampu memberikan perlindungan secara alami terhadap protein pakan. Tanin dari hijauan biasanya dari jenis terkondensasi yang resisten terhadap hidrolisa, sehingga dapat menurunkan kecernaan pakan dalam rumen. Sementara kandungan saponin yang cukup tinggi (10,28%) dalam daun kersen belum dapat dibuktikan mempengaruhi kecernaan pakan.
11 KESIMPULAN DAN SARAN
Kesimpulan
Penggunaan tepung daun kersen sampai level 20% dalam ransum menghasilkan nilai pH yang stabil, mampu meningkatkan produksi VFA total, menurunkan kecernaan bahan kering, kecernaan bahan organik, dan konsentrasi NH3
serta belum efektif dalam penekanan populasi protozoa dengan waktu inkubasi 4 jam secara in vitro.
Saran
Perlu dilakukan penelitian in vitro dengan waktu inkubasi 24 jam untuk mengetahui tingkat keefektifan daun kersen dalam penekanan populasi protozoa. Perlu dilakukan pula penelitian secara in vivo agar efek penggunaan daun kersen dapat langsung terlihat pada ternak.
24 UCAPAN TERIMA KASIH
Ahamdulillahirobbil’alamin, puji syukur penulis panjatkan kehadirat Allah SWT atas segala limpahan rahmat dan karunia-Nya serta atas ridho-Nya sehingga penyusunan skripsi ini dapat terselesaikan dengan baik. Shalawat serta salam tercurah kepada Nabi Besar Muhammad SAW.
Kesempatan kali ini penulis mengucapkan terima kasih banyak kepada Dr. Sri Suharti, S.Pt.M.Si selaku dosen pembimbing akademik sekaligus pembimbing utama yang telah memberikan bantuan finansial, bimbingan, dan arahan selama penelitian sampai selesainya skripsi ini dan kepada Dr. Anuraga Jayanegara, S.Pt.M.Sc selaku dosen pembimbing anggota atas kesediaan waktunya dalam mengarahkan, memotivasi, dan membimbing penulis selama penelitian sampai selesainya skripsi ini. Terima kasih kepada Dr. Ir. Didid Diapari, M.Si selaku penguji seminar, Ir. Anita S. Tjakradidjaja, M.Rur.Sc dan Ir. Komariah, M.Si selaku penguji sidang serta Ir. Widya Hermana, M.Si selaku panitia sidang atas kritik dan saran yang menjadikan skripsi ini lebih baik. Terima kasih penulis ucapkan kepada Bu Dian dan Bu Adriani yang telah bersedia membantu penulis selama penelitian.
Terima kasih yang sebesar-besarnya penulis ucapkan kepada ibunda Eri Lestari Muryani dan ayahanda Lis Indrianto serta adik Rizky Mahardhika atas kasih sayang, dukungan, dan doanya. Terima kasih kepada sahabat penulis (Chandra) atas motivasinya, partner selama penelitian (Dea) atas kerjasamanya, kepada Keluarga 8 (Habibah, Dea, Ponam, Apdila, Mutia, Pratita, Liza) atas bantuan, dukungan, dan semangatnya, kepada keluarga besar GENETIC 45 yang tidak dapat disebutkan satu-persatu atas kebersamaan dan kekompakannya serta kepada anak-anak wisma Pondok Indah (Ponam, Riska, Dian, Enda, Anjani) atas keceriaan, semangat, dan motivasinya.
Penulis mengucapkan terima kasih kepada semua pihak yang tidak dapat penulis sebutkan satu-persatu yang telah menjadi bagian dari perjalanan penulis dengan memberikan dukungan dan pengalaman yang berharga sampai terselesaikannya skripsi ini. Penulis berharap semoga skripsi ini dapat bermanfaaat terutama dalam dunia peternakan.
Bogor, Juli 2012 Penulis
25 DAFTAR PUSTAKA
Arora, S. P. 1989. Pencernaan Mikroba pada Ruminansia. Edisi Indonesia. Gajah Mada University Press, Yogyakarta.
Badan Penelitian dan Pengembangan Pertanian. 2012. Inovasi Teknologi untuk Kesejahteraan Petani. Laporan Tahunan 2011. Kementrian Pertanian, Jakarta. Badan Pusat Statistik. 2012. Populasi sapi potong Indonesia masa sensus Juni 2011.
http://www.bps.go.id. [23 Februari 2012].
Conway, E. J. 1962. Microdiffusion Analysis and Volumetric Error. 5th ed. Crosby Lockwood, London.
Dehority, B. A. 2004. Rumen Microbiology. Nottingham University Press, Nottingham.
Figueiredo, R. A., A. A. Oliveira, M. A. Zacharias, S. M. Barbosa, F. F. Pereira, G. N. Cazela, J. P. Viana, & R. A. Camargo. 2008. Reproductive ecology of the exotic tree Muntingia calabura L. (Muntingiaceae) in Southeastern Brazil. J. R. Arvore. 32: 993-999.
General Laboratory Procedure. 1966. Report of Dairy Science. University of Wisconsin Madison, USA.
Hess, H. D., M. Kreuzer, T. E. Diaz, C. E. Lascano, J. E. Carulla, C. R. Soliva, & A. Machmuller. 2003. Saponin rich tropical fruits affect fermentation and methanogenesis in faunated and defaunated rumen fluid. J. Anim. Feed. Sci. 109: 79-94.
Hu, W. L., J. X. Liu, J. A. Ye, Y. M. Wu, & Y. Q. Guo. 2005. Effect of tea saponin on rumen fermentation in vitro. J. Anim. Feed. Sci. 120: 333-339.
Jayanegara, A. & A. Sofyan. 2008. Penentuan aktivitas biologis tanin beberapa hijauan secara in vitro menggunakan Hohenheim Gas Test dengan polietilen glikol sebagai determinan. Med. Pet. 31(1): 44-52.
Jayanegara, A., H. P. S. Makkar, & K. Becker. 2009. Emisi metana dan fermentasi rumen in vitro ransum hay yang mengandung tanin murni pada konsentrasi rendah. Med. Pet. 32(3): 185-195.
Makkar, H. P. S. 2003. Quantification of Tannins in Tree and Shrub Foliage. Kluwer Academic Publishers, Netherlands.
Mayulu, H., Sunarso, C. I. Sutrisno, & Sumarsono. 2010. Kebijakan Pengembangan Peternakan Sapi Potong di Indonesia. J. Litbang Pertanian. 29(1):34-41.
26 McDonald, P., R. A. Edward, J. F. D. Greenhalgh, & C. A. Morgan. 2002. Animal Nutrition. 6th Edition. Scientific and Tech John Willey & Sons. Inc, New York.
Mueller, H. I. 2006. Unrevelling the conundrum of tannins in animal nutrition and health. J. Sci. Food Agric. 86: 2010-2037.
Ogimoto, K. & S. Imai, 1981. Atlas of Rumen Microbiology. Japan Scientific Society Press, Tokyo.
Patra, A. K. & J. Saxena. 2010. A new perspective on the use of plant secondary metabolites to inhibit methanogenesis in the rumen. J. Phytochemistry. 71: 1198-1222.
Pen, B., C. Sar, B. Mwenya, K. Kuwaki, R. Morikawa, & J. Takahashi. 2006. Effects of Yucca schidigera and Quillaja saponaria extracts on in vitro ruminal fermentation and methane emission. J. Anim. Feed. Sci. 129:175-186.
Santoso, B. & B. Tj. Hariadi. 2007. Pengaruh suplementasi Accacia mangium Willd pada Pennisetum purpureum terhadap karakteristik fermentasi dan produksi gas metana in vitro. Med. Pet. 30(2): 106-113.
Sirait, M. 2007. Penuntun Fitokimia dalam Farmasi. Penerbit ITB (Institut Teknologi Bandung), Bandung.
Smithsonian Tropical Research Institute. 2003. Muntingia calabura immature-fruit plant. http://www.discoverlife.org/mp/20q?search=Muntingia+calabura. [23 Februari 2012].
Soetanto, H. 2004. Mikrobiologi Rumen. Fakultas Peternakan. Universitas Brawijaya, Malang. http://images.hendrawansoetanto.multiplycontent.com. [24 Februari 2012].
Steenis, J. 2006. Flora: Untuk Sekolah di Indonesia. PT Pradnya Paramita, Jakarta. Suharti, S., D. A. Astuti, E. Wina, & T. Toharmat. 2011. Rumen microbial
population in the in vitro fermentation of different rations of forage and concentrate in the presence of whole lerak (Sapindus rarak) fruit extract. Asian-Aust. J. Anim. Sci. 24(8):1086-1091.
Suparjo. 2008. Saponin, peran dan pengaruhnya bagi ternak dan manusia. Laboratorium Makanan Ternak. Fakultas Peternakan. Universitas Jambi. http://jojo66.wordpress.com. [23 Februari 2012].
Sutardi, T. 1980. Landasan Ilmu Nutrisi. Departemen Ilmu Makanan Ternak. Fakultas Peternakan. Institut Pertanian Bogor, Bogor.
27 Syahrir, S., K. G. Wiryawan, A. Parakkasi, M. Winugroho, & O. N. P. Sari. 2009. The effectivity of Mulberry leaves to substitute concentrate in the in vitro ruminal system. Med. Pet. 32(2): 112-119.
Tan, H. Y., C. C. Sieo, N. Abdullah, J. B. Liang, X. D. Huang, & Y. W. Ho. 2011. Effects of condensed tannins from Leucaena on methane production, rumen fermentation and populations of methanogens and protozoa in vitro. J. Anim. Feed. Sci. 169: 185-193.
Tilley, J. M. A & R. A. Terry. 1963. A two stage technique for the in vitro digestion of forage. J. British Grassland Society. 18: 104-111.
Mauricio, R. M., E. Owen, F. L. Mould, I. Givens, M. K. Theodorou, J. France, D. R. Davies, & M. S. Dhanoa. 2001. Comparison of bovine rumen liquor and bovine faeces as inoculum for an in vitro gas production technique for evaluating forages. J. Anim. Feed. Sci. 89: 33-48.
Zakaria, Z. A., A. S. Sufian, K. Ramasamy, N. Ahmat, M. R. Sulaiman, A. K. Arifah, A. Zuraini, & M. N. Somchit. 2010. In vitro antimicrobial activity of Muntingia calabura extracts and fractions. Afr. J. Microbiol. Res. 4: 304-308.
28 LAMPIRAN
29 Lampiran 1. Hasil Analisa Pengaruh Perlakuan terhadap Nilai pH
Sumber Keragaman (SK) Jenis Keragaman (JK) Derajat Bebas (db) Kuadrat Tengah (KT) F. hit Signifikansi (Sig.) Level kersen 0,006 4 0,001 1,656 0,224 Kelompok 0,062 3 0,021 24,345 0,000 Galat 0,010 12 0,001 Total 935,995 20
Lampiran 2. Hasil Analisa Pengaruh Perlakuan terhadap Populasi Protozoa (Log 10/ml) Sumber Keragaman (SK) Jenis Keragaman (JK) Derajat Bebas (db) Kuadrat Tengah (KT) F. hit Signifikansi (Sig.) Level kersen 0,479 4 0,120 0,849 0,521 Kelompok 1,416 3 0,472 3,346 0,056 Galat 1,692 12 0,141 Total 288,881 20
Lampiran 3. Hasil Analisa Pengaruh Perlakuan terhadap Konsentrasi NH3
Sumber Keragaman (SK) Jenis Keragaman (JK) Derajat Bebas (db) Kuadrat Tengah (KT) F. hit Signifikansi (Sig.) Level kersen 8,433 4 2,108 6,571 0,005 Kelompok 15,430 3 5,143 16,031 0,000 Galat 3,850 12 0,321 Total 720,532 20
30 Uji Lanjut Duncan Pengaruh Level Daun Kersen terhadap Nilai NH3 (P<0,05)
Level Kersen N Subset 1 2 3 20 4 5,25036 10 4 5,37551 15 4 5,51818 5,51818 5 4 6,35503 6,35503 0 4 6,92919
Lampiran 4. Hasil Analisa Pengaruh Perlakuan terhadap Produksi VFA Total Sumber Keragaman (SK) Jenis Keragaman (JK) Derajat Bebas (db) Kuadrat Tengah (KT) F. hit Signifikansi (Sig.) Level kersen 8163,850 4 2040,962 27,151 0,000 Kelompok 12744,035 3 4248,012 56,512 0,000 Galat 902,046 12 75,171 Total 278188,102 20
Uji Lanjut Duncan Pengaruh Level Daun Kersen terhadap Produksi VFA (P<0,05) Level Kersen N Subset 1 2 3 0 4 89,98 5 4 104,07 10 4 109,01 15 4 112,41 20 4 150,62
31 Lampiran 5. Hasil Analisa Pengaruh Perlakuan terhadap Nilai KCBK
Sumber Keragaman (SK) Jenis Keragaman (JK) Derajat Bebas (db) Kuadrat Tengah (KT) F. hit Signifikansi (Sig.) Level kersen 402,762 4 100,690 3,773 0,033 Kelompok 272,232 3 90,744 3,401 0,053 Galat 320,210 12 26,684 Total 63763,969 20
Uji Lanjut Duncan Pengaruh Level Daun Kersen terhadap Nilai KCBK (P<0,05) Level Kersen N Subset 1 2 15 4 50,57 20 4 53,69 10 4 54,28 5 4 57,84 57,84 0 4 63,72
Lampiran 6. Hasil Analisa Pengaruh Perlakuan terhadap Nilai KCBO Sumber Keragaman (SK) Jenis Keragaman (JK) Derajat Bebas (db) Kuadrat Tengah (KT) F. hit Signifikansi (Sig.) Level kersen 453,804 4 113,451 4,947 0,014 Kelompok 245,062 3 81,687 3,562 0,047 Galat 275,175 12 22,931 Total 65811,166 20
32 Uji Lanjut Duncan Pengaruh Level Daun Kersen terhadap Nilai KCBO (P<0,05)
Level Kersen N Subset 1 2 15 4 50,62 20 4 54,72 10 4 56,06 5 4 58,21 58,21 0 4 65,07