TINJAUAN PUSTAKA
Sagu
Tanaman sagu (Gambar 1) termasuk tumbuhan monokotil, keluarga Palmae, Marga Metroxylon, ordo Spadisiflorae. Nama Metroxylon berasal dari bahasa Yunani yang terdiri dari kata ”Metra” dan ”Xylon”. Metra berarti isi batang, sedang Xylon berarti kayu (Flach, 1983).
Gambar 1. Tanaman Sagu
Tanaman sagu digolongkan secara garis besar menjadi dua golongan yaitu golongan yang berbunga/berbuah satu kali dan golongan yang berbunga atau berbuah dua kali atau lebih (Soerjono, 1980). Golongan sagu yang berbunga atau berbuah satu kali bernilai ekonomis lebih tinggi karena memiliki kandungan pati yang tinggi. Jenis sagu yang termasuk dalam golongan tersebut adalah Metroxylon rumphii Martius, Metroxylon sagus Rottboll, Metroxylon sylvester Martius, Metroxylon micracantum Martius, Metroxylon longispinum Martius. Sedangkan golongan yang berbunga atau berbuah dua kali atau lebih rendah mengandung karbohidrat, tumbuh di daerah dataran tinggi yang terdiri dari Metroxylon filare dan Metroxylon elanum.
Bagian terpenting dari tanaman sagu adalah batang sagu, karena merupakan tempat penyimpanan cadangan makanan (karbohidrat) yang dapat menghasilkan pati sagu. Tinggi pohon sagu dapat mencapai 15 meter (Ruddle et al., 1978). Ukuran dari batang sagu dan kandungan patinya tergantung pada jenis sagu, umur dan habitatnya. Pada bagian dalam batang pohon sagu terdapat gumbar (empulur) yang mengandung karbohidrat. Pada umur panen sekitar 11 tahun ke atas empelur sagu mengandung pati sekitar 15-20 persen (Rumalu, 1981 dalam Haryanto dan Pangloli, 1992). Setiap pohon sagu dapat menghasilkan sagu berkisar antara 50-500 kg tepung sagu basah (Sastrapradja et al., 1980).
Kandungan pati maksimal ada pada saat sagu sebelum berbunga. Munculnya primordia bunga biasanya menunjukkan kandungan pati menurun. Kandungan pati menurun karena digunakan sebagai energi untuk pembentukan bunga dan buah. Setelah pembungaan dan pembentukan buah, batang akan menjadi kosong dan tanaman sagu mati. Keadaan tersebut mempermudah petani untuk mengetahui kandungan pati sagu secara maksimum (Haryanto dan Pangloli, 1992).
Sagu (Metroxylon sp.) merupakan tumbuhan asli Indonesia. Selain di Indonesia sagu ditemukan hanya dibeberapa negara lain, seperti Papua New Guinea, Malaysia, Thailand, dan Filipina (Ruddle et al., 1978). Indonesia memiliki areal sagu terbesar dengan luas sekitar 1,5 juta hektar atau 60% dari 2,5 juta hektar sagu dunia. Peta populasi sagu dunia disajikan pada Gambar 2.
Sagu merupakan salah satu sumber karbohidrat potensial disamping beras, khususnya bagi sebagian masyarakat di kawasan Timur Indonesia seperti Papua dan Maluku. Beberapa produk olahan pati sagu antara lain papeda, sinoli, soun, ongol-ongol, dan sebagainya. Diperkirakan sekitar 90% areal sagu Indonesia berada di kawasan Timur Indonesia. Lima daerah potensial sagu terbesar di Indonesia, yaitu untuk Papua 600.000 hektar; Maluku 94.949 hektar; riau 51.250 hektar; Sulawesi Utara 23.400 hektar dan Sulawesi Tenggara 13.706 hektar (Chiljo,2009).
Umumnya daerah pertumbuhan sagu pada daerah terisolasi dan jauh dari perkotaan. Sagu banyak tumbuh di daerah-daerah marjinal, dimana tumbuhan penghasil karbohidrat lainnya lebih sulit untuk tumbuh. Pemanfaatan sagu di Indonesia belum maksimal dibandingkan dengan Malaysia dan Thailand yang hanya memiliki luas areal sagu seluas 1,5% dan 0,2% dari total luas areal sagu dunia. Sistem pengolahan sagu di Indonesia masih sangat rendah yang ditandai dengan kapasitas dan produktivitas pengolahan yang masih rendah. Hal ini disebabkan oleh sebagian besar pengolahan sagu ditujukan hanya untuk memenuhi kebutuhan keluarga. Cara sederhana tersebut menghasilkan rendemen yang rendah dan kurang efisien.
Pati sagu di pasar internasional digunakan sebagai bahan subsitusi tepung terigu untuk pembuatan biskuit, mie, sirup berkadar fruktosa tinggi, industri perekat, dan industri farmasi. Pemanfaatan dan nilai tambah sagu ditingkat petani masih sangat sederhana. Sagu memiliki kandungan karbohidrat, protein, lemak, kalsium, dan zat besi yang tinggi. Dengan kandungan tersebut, sagu berpotensi sebagai bahan baku untuk membuat bioenergi.
Batang merupakan komponen hasil utama pada tanaman sagu. Tepung sagu diperoleh dari empulur, sehingga pengolahan hasilnya cukup berat dan memerlukan alat yang khusus pula. Batang sagu yang sudah ditebang selanjutnya dikuliti untuk mendapatkan empulur yang mengandung tepung. Selanjutnya, empulur yang dihasilkan diparut agar memudahkan peremasan (pengepresan). Peremasan dilakukan dengan menggunakan alat pres untuk mengeluarkan pati dari parutan empulur. Setelah selesai peremasan, dilakukan penyaringan untuk membuang serat-serat kasar dari empulur. Suspensi pati yang didapatkan
kemudian diendapkan untuk memisahkan pati sagu dari air. Langkah selanjutnya adalah pengeringan, pengepakan dan penyimpanan atau distribusi ke konsumen.
Peralatan pengolahan sagu sudah tersedia atau mudah disediakan kecuali untuk kegiatan pemarutan. Pemarutan empulur dapat dilakukan dengan menggunakan alat berupa silinder berpaku yang digerakkan oleh generator. Silinder berpaku mudah direkayasa di pedesaan, sementara generator membutuhkan biaya yang cukup mahal. Kondisi seperti ini tidak mudah bagi kebanyakan masyarakat Papua yang rata-rata berpendapatan sangat rendah. Oleh karena itu, umumnya masyarakat Papua menggunakan teknik pengolahan dan peralatan tradisional.
Untuk meningkatkan efisiensi pengolahan sagu, Balai Pengkajian Teknologi Pertanian (BPTP) Papua telah merakit alat pangkur sagu dan alat peremas spesifik lokasi, yaitu pangkur rantai, pangkur tali, dan pangkur gendong. Pangkur merupakan alat pemarut empulur untuk mempermudah peremasan, sedangkan alat peremas digunakan untuk memisahkan tepung sagu dari empulur. Pangkur dirakit dengan menggunakan bahanbahan yang tersedia di pedesaan. Konstruksi pangkur terdiri atas silinder kayu berpaku, gir sepeda roda belakang, pegas, dan rantai atau tali yang berfungsi sebagai belt. Penggunaan pangkur lebih efisien dibandingkan dengan pengolahan cara tradisional.
Ekstraksi Pati Sagu
Cara dan alat pengambilan pati sagu dapat dibedakan antara lain (1) cara tradisional; (2) tradisional yang disempurnakan; dan (3) cara modern. Pengambilan pati sagu pada umumnya terdiri dari dua tahap pekerjaan, yaitu penghancuran ”ela” atau bagian lunak dari batang sagu setelah ditebang serta pemerasan ”ela” yang telah dihancurkan. Proses penghancuran ”ela” bertujuan untuk menghancurkan serat yang ada pada batang sagu, sehingga tepung yang terkandung dalam serat dapat diambil semaksimal mungkin.
Pati sagu perlu dipisahkan dari serbuk batang yang tercampur. Pemisahan dilakukan dengan pemerasan dan pengendapan. Serbuk batang sagu yang telah dikeruk dimasukkan ke dalam keranjang, lalu diangkut ke tempat pemerasan, dan dituang ke dalam bak pemerasan. Pemerasan dilakukan dengan tangan atau
menggunakan kaki. Untuk memudahkn pemisahan pati sagu, serat-serat batang tersebut diberi air. Pemerasan dihentikan setelah air yang menetes jernih.
Sifat Fisika Kimia Pati
Pati merupakan salah satu bentuk karbohidrat tanaman yang terbentuk melalui proses fotosintesis. Pati terdapat dalam butir-butir kecil atau granula yang terkumpul dalam biji, umbi dan bagian dalam batang. Adapun sumber pati yang telah banyak digunakan antara lain ubi kayu, ubi jalar, jagung, beras, sagu, kentang, dan barley (Brautlecht, 1953).
Bentuk dan ukuran granula khas untuk setiap jenis pati, sehingga sifat ini dapat digunakan untuk menentukan sumbernya (Hill dan Kelly, 1942). Granula pati terdiri atas lapisan tipis yang merupakan susunan melingkar dari molekul-molekul pati dengan bentuk kristal kecil yang disebut micella. Granula pati dalam keadaan murni berwarna putih, mengkilat, tidak berbau dan tidak berasa (Brautlecht, 1953). Granula pati sagu berbentuk ellips dengan ukuran 20 -60 µ. Sifat fisika kimia pati sagu sebagai berikut :
Gelatinisasi
Bila granula pati dimasukkan ke dalam air dingin maka akan terjadi pengembangan granula yang dapat bersifat bolak-balik (reversible). Besarnya pengembangan tergantung pada jenis pati dan perlakuan sebelumnya (Williams, 1968). Pengembangan terjadi karena sejumlah kecil air menyerap ke bagian amorf dari granula pati, yang memiliki ikatan intermolekuler kurang kuat. Bila suspensi pati dipanaskan maka air akan lebih mudah masuk ke dalam granula. Mula-mula terjadi sedikit pengembangan sampai tercapai suhu 60oC akan terjadi pengembangan granula yang sangat cepat dengan meningkatnya suhu (Radley, 1968).
Pengembangan granula pati karena pemanasan ini disebut suhu gelatinisasi pati dan proses ini tidak kembali (irreversible). Suhu pada saat terjadinya pengembangan granula pati dengan membentuk gel disebut suhu gelatinisasi. Pengembangan granula pati akan menyebabkan terjadinya beberapa perubahan diantaranya hilangnya sifat birefringent granula, peningkatan viscositas dan kecerahan suspensi (Radley, 1968).
Menurut Brautlecht (1953), ukuran granula pati sagu berkisar antara 10-70 µm, sebagian besar terdiri dari granula berukuran besar (50-70 µm), dan hanya sedikit yang berukuran kecil (10-20 µm). Umumnya butiran pati yang berukuran kecil akan menggelatinisasi lebih lambat dan pada suhu yang lebih tinggi dibandingkan dengan butiran pati yang berukuran besar, karena setiap granula akan mengembang pada saat tercapainya suhu gelatinisasi masing-masing. Oleh karena itu, adanya variasi dari ukuran gelatinisasi dan merupakan ciri khas dari masing-masing jenis pati. Suhu gelatinisasi pati sagu berkisar pada 60oC (Jarowenko, 1976).
Amilosa dan Amilopektin
Amilosa adalah komponen berantai lurus dengan ikatan α-(1,4) D-glukosa. Tiap polimer pati mengandung 200-2000 unit D-glukosa. Amilosa bersifat hidrofilik, karena banyak mengandung gugus hidroksil pada molekulnya yang bersifat polar. Rantai lurus amilosa cenderung membentuk susunan paralel satu sama lain dan saling berkaitan melalui ikatan hidrogen. Jika hal ini terjadi maka afinitas amilosa terhadap air akan menurun karena adanya ikatan antara molekul. Kumpulan molekul amilosa akan meningkat sampai mencapai suatu titik dimana terjadi pengendapan bila konsentrasinya rendah, dan akan terbentuk gel bila konsentrasinya tinggi.
Amilopektin adalah polimer berantai cabang, yang setiap cabang terdiri 15-25 unit D-glukosa dengan ikatan α-(1,4) glikosidik dan pada tempat sambungan percabangannya dengan ikatan α-(1,6) glikosidik. Perbandingan antara jumlah amilosa dan amilopektin dari tiap jenis pati berlainan, pada umumnya pati mengandung amilosa sekitar 22-26 % dengan amilopektin 74-78 % (Whistler, 1977). Khususnya pati sagu menurut Johnson dan Peterson (1974) perbandingan kandungannya adalah 17-27 % amilosa dan 73-83 % amilopektin.
Komposisi Kimia Pati Sagu
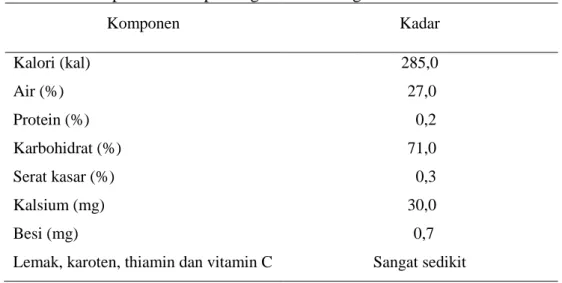
Komposisi kimia pati sagu sangat bervariasi. Variasi tersebut tidak banyak dipengaruhi oleh perbedaan spesies, umur, dan habitat di mana pohon sagu tumbuh. Faktor utama yang mempengaruhi variasi tersebut adalah faktor pengolahanannya. Komposisi kimia pati sagu dapat dilihat pada Tabel 1.
Tabel 1. Komposisi kimia pati sagu dalam 100 gram bahan Komponen Kadar Kalori (kal) 285,0 Air (%) 27,0 Protein (%) 0,2 Karbohidrat (%) 71,0 Serat kasar (%) 0,3 Kalsium (mg) 30,0 Besi (mg) 0,7
Lemak, karoten, thiamin dan vitamin C Sangat sedikit Sumber: Ruddle et al. (1978).
Sirup Glukosa Pati Sagu
Sirup glukosa pati sagu berupa cairan jernih dan kental dengan komponen utamanya glukosa yang diperoleh dari hidrolisis pati. Hidrolisis pati menjadi glukosa dapat dilakukan dengan menggunakan bantuan enzim atau asam pada waktu, suhu, dan pH tertentu. Berbagai metode hidrolisis pati telah banyak dikembangkan, diantaranya hidrolisis asam, hidrolisis enzim dan kombinasi antara asam dan enzim.
Hidrolisis pati menjadi sirup glukosa dengan asam telah lama digunakan pada pembuatan sirup glukosa dan menghasilkan sirup glukosa dengan DE-42 yang banyak digunakan pada industri permen. Hidrolisis secara asam memiliki kekurangan dibandingkan dengan hidrolisis enzim, yaitu timbulnya warna dan flavor yang tidak diinginkan, sehingga dapat menurunkan mutu produk, serta memerlukan kondisi proses yang ekstrim. Berbeda dengan hidrolisis enzimatis, selain kondisi proses yang tidak ekstrim, pemakaian enzim dapat menghasilkan rendemen dan mutu larutan glukosa yang lebih tinggi dibandingkan dengan hidrolisis secara asam. Pada hidrolisis secara enzimatis ikatan pati terpotong secara teratur atau sesuai dengan jenis enzim yang digunakan, sedangkan hidrolisis secara asam pemotongan terjadi secara acak.
Terdapat tiga tahapan dalam mengkonversi pati pada proses hidrolisis pati sagu, yaitu tahap gelatinisasi, likuifikasi, dan sakarifikasi. Tahap gelatinisasi merupakan tahapan pembentukan suspensi kental dari granula pati, tahap
likuifikasi berupa proses hidrolisis pati parsial yang yang ditandai dengan menurunnya viscositas, dan tahap sakarifikasi adalah proses lebih lanjut dari hidrolisis untuk menghasilkan glukosa (Chaplin dan Buckle, 1990).
Pada tahap likuifikasi terjadi pemecahan ikatan α-1,4 glikosidik oleh enzim α-amilase pada bagian dalam rantai polisakarida secara acak, sehingga dihasilkan glukosa, maltosa, maltodekstrin dan α-limit dekstrin. Enzim α-amilase merupakan enzim yang menghidrolisis secara khas melalui bagian dalam dengan memproduksi oligosakarida dari konfigurasi alfa yang memutus α-(1,4) glikosidik dan amilosa, amilopektin, dan glikogen. Ikatan α-(1,6) glikosidik tidak dapat diputus oleh α-amilase, tetapi dapat dibuat menjadi cabang-cabang yang lebih pendek (Nikolov dan Reilly, 1991). Enzim α-amilase umumnya diisolasi dari Bacillus amyloquefaciens, B. licheniformis, Aspergillus oryzae, dan A. niger. Nilai pH optimum untuk aktivitas enzim ini sekitar 6 dengan suhu optimum 60oC. Apabila suhu dinaikkan, maka pH optimum pun semakin meningkat sampai kisaran tujuh (Tjokroadikoesomo, 1986).
Pada likuifikasi pati biasanya α-amilase yang digunakan memiliki aktivitas yang tinggi, sehingga dosis enzim yang digunakan sekitar 0,5-0,6 kg/ton pati atau 1500 U/kg substrat kering (Chaplin dan Bucke, 1990). Enzim α-amilase komersial dibuat oleh Novo Industry AS antara lain dengan nama Termamyl yang mempunyai ketahanan terhadap suhu sekitar 95-110oC. Stabilitas Termamyl tergantung pada suhu, konsentrasi Ca2+, kandungan ion dan ekuivalen dekstrosa. Dosis α-amilase yang biasa digunakan antara 0,5-0,6 kg Termamyl 120 L per ton pati kering. Satu kNU (kilo Novo α-amilase Unit) adalah jumlah enzim yang dapat menghidrolisis 5,26 pati (gram standar) per jam dengan suhu 37oC, pH 5,6 pada kondisi standar.
Proses selanjutnya adalah sakarifikasi oleh enzim amiloglukosidase. Amiloglukosidase merupakan eksoenzim yang terutama memecah ikatan α-(1,4) dengan melepaskan unit-unit glukosa dari ujung non reduksi molekul amilosa dan amilopektin untuk memproduksi β-D-Glukosa. Nama trivial yang sering digunakan pada enzim ini adalah amiloglukosidase (AMG), glukomilase, dan gamma-amilase (Kulp, 1975). Amiloglukosidase ditemukan pada tahun 1950-an dan digunakan secara luas pada teknologi bioproses pati dan industri makanan.
Kegunaan yang luas dan spesifik menyebabkan amiloglukosidase digunakan pada produksi gula cair.
Amiloglukosidase diproduksi pada skala besar dari kapang dan khamir, tetapi hanya Aspergillus dan Rhizopus yang digunakan secara komersial. Suhu optimum untuk amiloglukosidase berkisar 40-60oC dengan pH optimum 3-8. amiloglukosidase yang umum digunakan pada tahap likuifikasi berasal dari Aspergillus niger. Pada kondisi yang sesuai, amiloglukosidase ditambahkan dengan dosis berkisar 1,65-0,80 liter enzim per ton pati dengan dosis sebesar 200 U/kg pati (Chaplin dan Buckle, 1990). Amiloglukosidase yang berasal dari Novo yaitu AMG yang tersedia dalam bentuk cairan dengan aktivitas 200, 300 atau 400 AGU g-1. satu AGU (Amiloglukosidase Unit) adalah jumlah enzim yang menghidrolisis 1 µmol maltosa per menit pada suhu 25oC dan kondisi standar (Kearsley dan Dziedzic, 1995).
Kultivasi Etanol
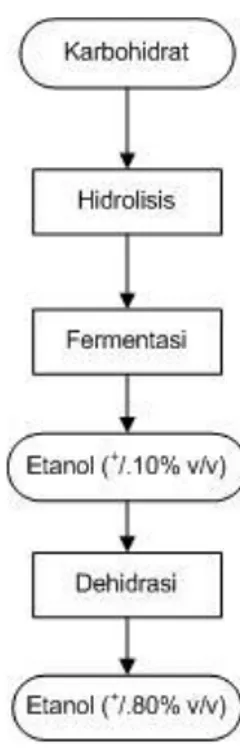
Menurut Prescot dan Dunn (1959), etanol dapat diproduksi dari gula melalui kultivasi pada kondisi tertentu. Sedangkan pati dan karbohidrat lainnya dapat dihidrolisis menjadi gula kemudian dikultivasi untuk membentuk etanol. Etanol merupakan nama kimia untuk alkohol dengan rumus kimia C2H5OH. Etanol adalah suatu cairan tak berwarna dengan bau yang khas. Berat jenis secara spesifikasi cairan ini pada 15oC sebesar 0,7937 dan mulai mendidih pada suhu 78,32oC (760 mm air raksa). Bioetanol adalah cairan biokimia dari proses kultivasi gula dari sumber karbohidrat dengan menggunakan bantuan mikroorganisme. Proses produksi etanol berbahan baku karbohidrat dapat dilihat pada Gambar 4.
Mikroba yang digunakan dalam kultivasi etanol adalah khamir. Khamir yang biasa digunakan untuk menghasilkan etanol adalah Saccharomyces cerevisiae var. ellipsoides. Khamir ini sering digunakan pada kultivasi etanol karena menghasilkan etanol yang tinggi, toleran terhadap kadar etanol yang tinggi, mampu hidup pada suhu yang tinggi, tetap stabil selama kondisi kultivasi dan dapat bertahan hidup pada pH rendah (Rehm dan Reed, 1981).
Gambar 4. Diagram alir proses produksi etanol (Wyman, 2001)
Saccharomyces cerevisiae var. ellipsoides dapat diperoleh dalam bentuk kultur murni biasanya digunakan dalam pembuatan minuman beralkohol (brewing yeast dan wine yeast). Khamir memerlukan medium dan lingkungan yang sesuai untuk pertumbuhan dan perkembang-biakannya. Unsur-unsur dasar yang dibutuhkan berupa karbon, hidrogen, oksigen, fosfor, zat besi, dan magnesium. Unsur karbon banyak diperoleh dari gula dan sumber nitrogen didapatkan dari amonia, asam amino, peptida, pepton, nitrit atau urea tergantung pada jenis khamir. Fosfor merupakan unsur penting dalam kehidupan khamir terutama dari pembentukan alkohol dari gula.
Pada awal proses kultivasi, khamir memerlukan oksigen untuk pertumbuhannya, sehingga kultivasi berlangsung secara aerob. Selanjutnya setelah terbentuk CO2, reaksi akan berubah secara anaerob. Alkohol yang terbentuk akan menghalangi kultivasi lebih lanjut setelah tercapai konsentrasi antara 13-15% volume. Konsentrasi alkohol akan menghalangi kultivasi, tergantung pada suhu dan jenis khamir yang digunakan (Prescot dan Dunn, 1959).
Khamir tumbuh terbaik pada kondisi aerobik, walaupun demikian beberapa khamir dapat tumbuh pada kondisi anaerobik. Proses respirasi pada kondisi aerobik selanjutnya digantikan dengan proses kultivasi pada kondisi anaerobik
karena tidak tersedia lagi oksigen. Khamir akan selalu berespirasi pada setiap keadaan yang memungkinkan karena energi yang dihasilkan pada respirasi jauh lebih besar dari pada energi yang dihasilkan pada kultivasi (Barnett et al., 2000). Apabila terdapat udara pada proses kultivasi maka etanol yang dihasilkan lebih sedikit, karena terjadi respirasi yang mengakibatkan terjadinya konversi gula menjadi karbondioksida dan air.
Suhu optimum pertumbuhan khamir adalah pada suhu 25-30 oC dan maksimum pada 35-47 oC. Sedangkan pH optimum adalah 4-5. Batas minimal aw untuk khamir biasanya adalah 0,88-0,94 sedangkan untuk khamir osmofilik dapat tumbuh pada aw yang lebih rendah yaitu sekitar 0,32-0,65. Namun banyak juga khamir osmofilik yang pertumbuhannya terhenti pada aw 0,78 seperti pada larutan garam atau sirup (Frazier dan Westhoff, 1978).
Menurut Casida (1968), pH pertumbuhan khamir yang baik adalah pada rentang antara 3-6. Perubahan pH dapat mempengaruhi pembentukan hasil samping kultivasi. Nilai pH pertumbuhan berhubungan positif dengan pembentukan asam piruvat. Pada pH tinggi, maka lag fase akan lebih singkat dan aktifitas kultivasi akan meningkat. Pengaruh pH pada pertumbuhan khamir juga tergantung pada konsentrasi gula dan etanol. Nilai pH dapat dipertahankan dalam medium konstan dengan menambahkan buffer (larutan penyangga).
Paturau (1981) menyatakan bahwa kultivasi etanol memerlukan waktu 30-72 jam. Prescott dan Dunn (1959) menyatakan waktu kultivasi etanol adalah 3-7 hari. Amerine dan Cruess (1960) menyatakan bahwa proses konversi gula menjadi etanol dan CO2 dilakukan oleh sel khamir Pada kondisi anaerob, metabolisme glukosa menjadi etanol melalui jalur Embden Meyerhoff-Parnas yang merupakan reaksi-reaksi fosforilasi dan defosforilasi dengan ATP dan ADP sebagai donor aseptor fosfat, reaksi pemecahan C6 menjadi 2 molekul C3 yang terforforilasi, reaksi oksidasi-reduksi dan reaksi dekarboksilasi. Jalur Embden Meyerhoff-Parnas dapat dilihat pada Gambar 5.
Keterangan : ATP = Adenosin Trifosfat ADP = Adenin Difosfat
NAD = Nikotinamida Adenin Dinukleotida NADP = Nikotinamida Adenin Dinukleotida Fosfat NADPH = Nikotinamida Adenin Dinukleotida Tereduksi
Gambar 5. Skema Embden Meyerhoff-Parnas Pathway (Prescott dan Dunn, 1959)
Enzim yang berperan dalam pembuatan etanol dari glukosa adalah heksosinase, fosfoheksoisomerase., fosfofruktokinase, aldose, triosefosfat isomerase, 3-fosfat gliseraldehid dehidrogenase, fosfogliserokinase, piruvat karboksilase, dan alkohol dehidrogenase.
Secara teoritis konversi molekul gula menjadi 2 molekul etanol dan 2 molekul CO2 menurut persamaan Gay Lussac sebagai berikut:
Berdasarkan persamaan di atas dapat dijelaskan bahwa secara teoritis 51,1 % gula diubah menjadi etanol dan 48,9 % diubah menjadi karbondioksida. Akan tetapi hasil ini tidak sepenuhnya dapat tercapai diakibatkan karena adanya hasil samping. Pada kenyataannya hanya 90-95 % dari nilai tersebut yang dapat tercapai. Konsentrasi alkohol yang dihasilkan dalam kultivasi tergantung pada jenis khamir yang digunakan dan kadar gula, sedangkan konsentrasi produk samping dipengaruhi oleh suhu, aerasi, kadar gula, dan keasaman (Underkofler dan Hickey,1954). Produk samping yang dihasilkan berupa asam piruvat dan asam laktat.
Glukosa mengalami fosforilasi menjadi glukosa-6-P dan fruktosa-6-P dengan ATP sebagai donor fosfat. Fruktosa-6-P kemudian diubah menjadi fruktosa 1,6-di-P menggunakan ATP sebagai donor fosfat. Fruktosa-1,6-di-P selanjutnya dipecah menjadi dua molekul C3 yang terfosforilasi yaitu dihidroksiaseton fosfat dan gliseridehida-3-P. Dihidroksi aseton fosfat selanjutnya teroksidasi menjadi gliserolfosfat, kemudian diubah menjadi gliserol yang merupakan metabolit sekunder. Gliseraldehid-3-P tereduksi membentuk asam 1,3-di-fosfogliserat kemudian mengalami defosforilasi menjadi 3-P-asam gliserat dengan melepaskan fosfat dan aseptor fosfat ADP membentuk ATP. Selanjutnya, 3-P-asam gliserat membentuk 2-P-asam gliserat kemudian terbentuk asam fosfoetanol piruvat dengan menghasilkan ATP. Melalui reaksi dekarboksilasi, asam piruvat akan membentuk asetaldehid dan CO2 yang berikutnya akan mengalami reaksi oksidasi membentuk etanol.
Penambahan inokulum khamir dapat dilakukan dalam berbagai bentuk, diantaranya dalam bentuk suspensi atau dalam bentuk kering. Banyaknya suspensi khamir yang ditambahkan dalam kultivasi besar adalah sekitar 1-3 % (Prescott dan Dunn,1959). Menurut Undekofler dan Hickey (1954) paling sedikit penambahan inokulum aktif pada pembuatan wine adalah sekitar 1 % apabila substrat yang dipergunakan bersih dan bebas dari khamir yang tidak diinginkan. Sementara itu, Rinaldy (1987) menggunakan konsentrasi inokulum sebesar 10 % (v/v).
Komposisi media untuk setiap mikroba berbeda satu dengan yang lainnya. Zat makanan utama bagi pertumbuhan mikroba adalah sumber karbon, nitrogen, mineral terutama fosfat (Casida, 1968). Pertumbuhan mikrobial dipengaruhi oleh
konsentrasi komponen penyusun media pertumbuhannya. Pasokan sumber karbon merupakan faktor yang sangat berpengaruh pada pertumbuhan optimal, akan tetapi pada kenyataannya konsentrasi sumber karbon mempunyai batas maksimum. Apabila konsentrasi sumber karbon melampaui batas maka laju pertumbuhan akan terhambat (Sa’id, 1987).
Sumber karbon yang digunakan pada kultivasi etanol skala industri adalah karbohidrat yang dapat diperoleh dari berbagai jenis pati seperti jagung, serelia, kentang, sagu, ubi kayu, dan lain-lain. Sumber nitrogen yang dapat dipergunakan dalam proses kultivasi diantaranya corn steep liquor, ekstrak gandum atau tauge, hidrolisat kasein, dan ekstrak khamir. Vogel (1983) membedakan sumber nitrogen menjadi sumber organik dan anorganik, yang termasuk sumber organik adalah corn steep liquor, urea, protein, ekstrak khamir dan tepung ikan, sedangkan sumber nitrogen anorganik adalah gas amonia, amonium hidroksida dan amonium sulfat.
Menurut Hartoto (1992) sumber nitrogen yang biasa digunakan untuk kultivasi skala besar adalah garam amonium, urea atau amonia. Pemilihan garam ammonium sebagai sumber nitrogen disebabkan oleh faktor ekonomis yaitu harga yang relatif murah dan mudah didapat. Selain itu NPK dan ZA juga harga ekonomis dan mudah didapat.
Kinetika Fermentasi
Pertumbuhan mikrobial ditandai dengan peningkatan jumlah dan massa sel. Sedangkan kecepatan pertumbuhan tergantung pada lingkungan fisik dan kimianya (Reed dan Rehm, 1983). Kinetika kultivasi mempelajari perkembang-biakan mikroba yang dilanjutkan oleh kenaikan konsentrasi biomassaa karena konsumsi substrat. Pada saat yang bersamaan dihasilkan produk baik metabolit primer maupun metabolit skunder (Mangunwidjaja dan Suryani, 1994).
Menurut Bailey dan Olis (1991) kultivasi media cair dapat dilakukan dengan tiga cara, yaitu kultivasi sistem nir-sinambung (batch), kultivasi semi sinambung (fed-batch), dan sistem sinambung (continuous). Pada kultivasi nir-sinambung, pemanenan produk dilakukan setelah kultivasi berakhir dan tidak dilakukan lagi penambahan komponen substrat selama kultivasi berlangsung (Rachman, 1989).
Proses kultivasi untuk pertumbuhan mikroba secara umum mengikuti pola pertumbuhannya. Fase-fase pertumbuhan mikroba secara umum yaitu fase permulaan (initial phase), fase pertumbuhan dipercepat (phase of accelerated growth), fase logaritma atau eksponensial (logaritmic phase atau exponential phase), fase pertumbuhan yang mulai terhambat (phase of neganitive accelerated growth), fase kematian yang dipercepat (phase of accelerated death), fase kematian logaritma (logaritmic death phase) (Hidayat et al., 2006). Fase eksponensial terjadi pertumbuhan secara cepat dimana jumlah sel bertambah secara eksponensial terhadap waktu. Menurut Rehm dan Reed (1981) pada fase eksponensial kondisi lingkungan berubah karena substrat dan nutrien dikonsumsi, sementara dihasilkan metabolit.
Pada saat substrat mendekati habis dan terjadi penumpukan produk-produk penghambat maka terjadi penurunan laju pertumbuhan. Pada fase stasioner konsentrasi biomassa mencapai maksimum. Setelah fase tersebut terjadi fase kematian yang ditandai dengan penurunan jumlah individu sel yang hidup (Bailey dan Olis, 1991). Pertumbuhan mikroba pada proses kultivasi secara umum mengikuti pola seperti disajikan pada Gambar 6.
Gambar 6. Kurva pertumbuhan mikroba.
Pada keadaan lingkungan tertutup/nir-sinambung (batch) pertumbuhan mikrobial dapat dinyatakan dengan persamaan berikut:
Fase stasioner Fase adaptasi Fase log/eksponensial Fase p er tum buhan l am b at Ln konse ntra si bi omassa Waktu
x x dt dx
... (1) keterangann : x : Konsentrasi sel t : waktu kultivasiµ : laju pertumbuhan spesifik
α : laju lisis sel yang menghambat pertumbuhan
Pada kondisi yang sesuai maka penurunan massa sel sangat kecil, sehingga α dapat diabaikan, maka persamaan 1 menjadi:
x dt dx
... (2)
Integrasi dari persamaan 2 untuk menghasilkan nilai peningkatan massa sel pada suatu selang waktu tertentu adalah:
dt dx dx t t x X
2 1 2 1
... (3) akan diperoleh persamaan:t x x
1 2 ln atau lnx2lnx1
t ... (4)Laju pertumbuhan spesifik (µ) bersifat tidak konstan tergantung pada kondisi lingkungan fisik dan kimianya. Nilai laju pertumbuhan maksimum (µmaks) dicapai pada kondisi pasokan substrat dan nutrien masih berlebih serta konsentrasi zat-zat metabolik yang menghambat pertumbuhan masih rendah.
Menurut Wang et al. (1979), koefisien hasil sel hidup terhadap sumber karbon dinyatakan sebagai Yx/s, koefisien konversi nutrien dalam substrat menjadi produk pada periode tertentu dinyatakan sebagai Yp/s, sedangkan koefisien produk terhadap jumlah sel hidup dinyatakan sebagai Yp/x. Perhitungan yang biasa dilakukan untuk proses pembentukan produk yang berasosiasi dengan pertumbuhan sel adalah sebagai berikut:
X X
Y
S S
S S X X S Y o x s o o o s x / / ... (5)
S S
P P
Y
S S
P P S P Y o p s o o o s p / / ... (6)
P P
Y
X X
X X P P X P Y o p x o o o x p / / ... (7)Parameter-parameter di atas perlu diketahui agar pada kultivasi skala yang lebih besar dapat ditentukan jumlah substrat yang diperlukan untuk menghasilkan jumlah produk dan biomassa yang tertentu. Informasi tersebut digunakan untuk meningkatkan efisiensi kultivasi.
Kultur fed-batch merupakan kultur batch dengan pemasukan secara sinambung atau secara teratur dengan medium tanpa pengeluaran cairan kultur, sehingga volume kultur bertambah dengan bertambahnya waktu. Pembentukan biomassa pada sistem ini dapat ditulis dalam persamaan di bawah ini (Hidayat, 2006). ) (Sr S Y Xo Xt ... (8) Konsentrasi biomassa akhir dihasilkan pada saat S = 0 yang dapat dijelaskan sebagai Xmaks, dan X0 relatif kecil terhadap Xmaks,sehingga :
Sr Y
Xmaks . ... (9) Jika pada waktu X = Xmaks mulai dimasukkan substrat baru dengan laju umpan pengenceran yang kurang/lebih kecil dari µmaks, maka semua substrat akan dikonsumsi sama cepatnya dengan kultur awal, sehingga :
Y X
FSr. / ... (10) X = total biomassa dalam kultur, diuraikan oleh X = µV dimana V merupakan volume fermentor pada waktu t. Dari persamaan 10 dapat disimpulkan bahwa substrat yang masuk setara dengan substrat yang dikonsumsi oleh sel. Jadi dS/dt = 0. Meskipun total gula dalam kultur (X) bertambah dengan waktu, konsentrasi sel (x) sebenarnya tetap konstan, dan dx/dt = 0, oleh karena itu µ = D. Laju pengenceran akan berkurang dengan bertambahnya volume dan D (laju dilusi) akan memberikan bentuk:
) (V0 Ft F D ... (11) Menurut kinetika Monod, sisa substrat akan berkurang dengan semakin rendah D dan menghasilkan peningkatan konsentrasi sel. Quasi – steady state dapat tercapai pada D < µmaks dan Ks < Sr.
Penggunaan kultur fed-batch dalam industri kultivasi mampu membuat sisa konsentrasi substrat menjadi sangat rendah. Rendahnya sisa substrat memberikan keuntungan dalam:
1. Mengurangi pengaruh penekanan (represi) langsung dari sumber karbon yang digunakan dan membuat kondisi kultur dalam kapasitas aerasi fermentor. 2. Menghindari efek toksik dari komponen medium.