KINETIKA
FERMENTASI PRODUKSI
SELULASE
DARI
ISCLAT
ACTINOMYCETES ACP-7 PADA MEDIA PADAT JERAMI
PADI
(FERMENTATION KINETIC
OF
CELLULASE PRODIJCTIONBY
ACTINOMYCETESACP.
7 lsoLATE rN
SOLID STATE R|CE STRAW MEDTUM)Heri Satria, Dian Herasari,
Suripto
Dwi
Yuwono
Jurusan Kimia FMIPA Universitas LampungJl. Prof. Dr. Soemantri Brodjonegoro No 1, Gedung Meneng gandarLampung 35145
E-mal : satria chemistrv@unila.ac.id
Accepted: 23 Agustus ; Received: 3 Oktober 2011
ABSTRAK
Biokonversi selulosa yang terkandung di dalam jerami paci memerlukan sumber enzim selulase yang
dapat dihasilkan oleh mikroorganisme anta:'a lain actinomycetes. lsolat AcP-7 merupakan salah satu actinomycetes yang telah berhasil disolasi dari jerami padi yang memiliki kernampuan setutotitit cukup baik dengan indek
seiulolitik sebesar 3,36. Penelitian ini memiliki tujuan untuk mengetahui kondisi optimum fermeniisi padat menggunakan inokulum actinomycetes isolat AcP-7 pada media jerami padi. Tiga perlakuan pH pada level 6,00;7,00;dan 8,00 diujikan bersamaan dengan perlakuan perbandingan substrat :-ntoisture masing-masing 1:1;
1:2:1:3: dan 1:4. Setelah kondisi optimum diperoleh enzim selulase diproduksi
r"nggrniLrn
kondisi tersebutdan beberapa parameter kinetika fermentasi dievaluasi. Kondisi optimum fermentasi padat isolat
Acp-7 pada substrat jerami padi berada pacia pH 8,00 dengan perbandingan substrat
:
moisture 1:4. Fase eksponensiall:::1?]
!?d? fgtTgniasi hari ke-9 sampai hari ke-21 dengan laju pertumt,uhan spesifik biomass sebesar 1,394 ctumL
per-hari. Koefisien rasio produk terhadap substrat (Yp/s) sebesar 0,32g,rasio produk terhadap biomass sel (Yp/x) sebesar 15,160, dan rasio biomass terhadap subitiat
(ws;
sebesar o,ozo. 'r-a;u maksimum produksiglukosa (p maks) sebesar 3'1,25 g per hari dengan Konstanta Monod (Ks) sebesar
ii,ii;,
sedangkan laju spesifik pembentukan procuk maksimum (qu) sebesar 2,06I
glg biomass per-hari darr periggL,naan substrat maksimum (q') dengan sistem fermentasi yang diterapkan dapaimencapat 1557,5 g.Kata kunci
:
selulosa, selulase, Jerami padi, Actinomycetes, Kinetika fermentasi ABSTRACTBioconversion of cellulose contained in the rice straw requires a source
of
cellulase enzymesthat can be produced by microorgant'srns such as actinomycefes. /so/ates AcP-7 is one of the actinomycites
that have been successfully isolated from rice stra.
lt
has goood cellulotic ability with cellulolytic index ofi.36.
Objective of this research is to determine the optimum conditions of solid fermentation using'inoculums acitinomycete.s lso/afes AcP-7in
the rice straw media. Three treatmentsat
the level ofpH
6.00; 7.00,;;i
g.00 were tested simultaneously with the treatment ratio of substrate to moisture each 1':1; 1:2;iS,
anat:1.
Once the optimum conditions obtained by cellulase enzymes using these conditions and some fermentation kinetics parameterswereevaluated.Optimumconditionsof solidfermentationlsolafes AcP-TinthericeslrawsubslratewasatpH 8'00 with a ratio of substrate to moisture 1:4. Exponentia! phase of fermentation
is
reached on day 9 to day 21 with the specific biomass growth rate of 1.394 cfu mL-i per day. Tlte coefficientof
the ratioof
product to subsfrafe (Yp/s) of 0.329, the ratic of products to cell biomass (Yp/x)of
15.160, and the ratio of blomass to subsfrafe (Yx/s) of 0.020. The maximum rate of g-lucose production'(p max)is
31.2sg
peitr day with Monod constant (Ks) of 74'34 g, while the maximum rate of specific product foination' (qr)is
z.o"oi g t g of biomass per day. The maximum subsfrale (q) are applied on fermentation system to achievi'|'SSZ,S g.Key words: cellulose, cellulases, Rice straw, Actinomycetes, Krnetrbs of fermentation
PENDAHULUAN
Jerami padi merupakan salah satu bagian
dari
limbah
biomassa berlignoseilulosa yangdihasilkan dari kegiatan pertanian
di
lndonesia.Menurut data BPS pada tahun 2010, produksi
padi
di
lndonesia mengalami
peni:gkatan sebesar 65,15 juta ton per tahun. Sebagai salahsatu
produsenpadi
nasronal angka kenaikanproduksi padi
di
propinsi Lampung pada tahun2007 sampai 2008 mencapai 0,77%, pada tahun
2008 sampai 2009 mencapai (),85%, dan pada
tahun 2009
sampai
2010
mencapai 0,2%.Peningkatan
ini
secara
tidak
langsungmenyebabkan peningkatan volume jerami padi
sebagai
hasil
samping produksipadi.
Potensilimbah
berlignoselulosauntuk
menglrasilkanproduk
yang
bermanfaat
sangat
besar.Biokonvcrsi lignoselurosa
dapat
rnenghasilkanbeberapa produk yang memiliki
nilai
ekonomiseperti sumber gula yang dapat dimanfaatkan
untuk industri bioetanol, asam sitrat, furfutal dan
xiiitol, atau dapat dimanfaatkan sebagai sumber
pakan ternak
bahk;anpakan
bagi
manusia(Howard et a\.2003),
Jerami padi dikctahui memiliki kandungan
selulosa
yang
tirrggi, mencapai 34,2Y0 beratkering,
24,5%
hemi:elulosadan
kandunganlignin
hingga
23,40/o(V/yman
et
al.
2002). Selurlosa merupakan komponen utama penyusundinding
sel
tanaman yang merupakan polimerglukosa
yang
tersusun
dari
unit-unitB-1,4-glukosa yang drnubungkan dengan ikatan
8. 1,4-D-glikosida (llan et al, 1995). Jika ikatan ini
diputus
baik
secara hld.oltsisasarn
maupLrnenzinratis, akan dihasilkan komponen penyusun
setulosa berupa oligosakarida bahkan grukosa.
Secara
biokimia pemutusanikatan
glikosidikpada selulosa memerlukan enzim yang dtkenal
sebagai
enzinr
selulase.Enzim
ini
berperandalam
hidroiisis seluiosa
dengan
nremecahikalan
B-1,4-D-glikosrdauntuk
menghasilkanoligosakarida maupurr glukosa. Selulase banyak
dihasilkan
oleh
ritikroba pengurai
sepertimikrofungi, actinomycetes, dan beberapa bakteri
yang memiliki kemampuan mensekresikan enzim
untuk mendapatkan sunrber karbon dalam siklus Iridupnya.
Kendala dalam pendegradasian
lignoselulosa untuk n-endapatkan gula-gula yang
dapat
difermentasi
lebih
lanjut
adalahkeberadaan lignin. Lignoselulosa tersusun atas
li1lrirr, lrotttisclulor;rt, rl ttt r;clrtlrlr;n.
l.irlril
lcril<;rtpada
hemiselulosa
dan
selulosa
yangmembentuk
struktur berlapis. Struktur
inir,renghalangi
larutan
alau enzim yang
akandigunakan
untuk
menguraikan selulosa danhemiselulosa menjadi gula-gula penyusunnya (Howard et a\.2003). Oleh sebab itu diperlukan
proses
delignifikasi
sebelum
sakarifikasi(Tahezadeh
and
Karimi 2008).Proses
hidrolisis lignoselulosa secaralengkap
memerlukansistem
enzim
yangsimultan untuk memperoleh hasil akhir yang
diinginkan. Penggunaan actinomycetes dalam
pendegradasian
limbah
berlignoselulosamemiliki
nilai
strategis
jika
dibandingkandengan mikroba lainnya karena mikroba ini
memiliki sistem enzim hidrolitik yang lengkap.
Actinomycetes merupakan bakterial tanah yang
memiliki
kemampuan
untuk
menguraikanlignoselulosa secara lengkap, karena genus ini
memiliki
kemampuan
untuk
menghasilkanenzim-enzinr hidrolitik ekstraselular (Wendish
and Kutzner 1992). Kemampuan actino:'nycetes
dalam mendegradasi iignoselulosa bukan saja
dapat dimanfaatkan untuk proses delignifikasi tetapi juga sekaligus proses sakarafikasi karena
kebanyakan
dari
genus
ini
mampumenghasilkan enzim ligninase, seiulase, dan
hemiselulase
secara
bersamaan.
Mikro-organisme yang selama rni diketahui memiliki
kemampuan
menghidrolisisselulosa
danhemiselulosa
seperti
Trichoderma
reseei(Howard et a|.2003) tidak memiliki kemampuan
menghidrolisis lignin. Sebaliknya jamur pelapuk
akar
dari
kelas
Basidiomycetes mampumendegradasi
lignin tetapi
tidak
memilikikernampuan
menqhidrolisisselulosa
danhemiselulosa (Gold dan Alic 1993).
Penelitian ini bertujuan untuk mengetahui
kemampuan actinomycetes isolat AcP-7 yang
telah
diperoleh
sebelumnyacjan
memilikikemampuan selulolitik dengan
indeks
3,36pada media
yeast-maltosa
agar
yangdisuplementasi 0,5% CMC dalam menghasilkan
enzim
selulase
pada
koncjisi
perlakuanoptimum,
dan
tinjauan kinetika
fernrentasimenggunakan substrat
jerami padi
dengan menggunakan teknik fermentasi padat.BAHAN DAN METODE Bahan
Jerami yang
digunakanberasal
dariPringsewu, Lampung varietas crherang, isolat rrctirrotnyr;r:tcs rrtorttp;rkrrrt kolrrk:'i l-lr:rl [i;r1ria
(Laboratorium
Biokirnia
Fl/lfrA
UniversitasLampung)
yang
ditumbuhkanpada
mediadigunakan
berkualifikasi
pro-analis
kecualidisebutkan khusus.
Optimasi Fermentasi Padat
lsolat
AcP-7
ditumbuhkanpada
mediainokulum yang dibuat dengan mengkultur dua
loop isolat pada media Yeast-Maltosa caii' yang
terdiri dari (g/L) ekstrak khamir
4;
ekstrak malt10;
glukosa14.
Sebanyak50
ml
inokulumditambahkan kepada
media
fermentasi yangtelah
disterilkanyang berisi
500
gr
jerami berukuran 60 mesh, 0,6% ekstrak khamir dalamlarutan trace elemen. Moisture yang digunakan
adalah buffer fosfat dengan variasi perbandingan
substrat:moisfure sebesar 1:1, 1:2, 1:3, dan 1:4
dan
variasi
pH
6,0;7,0;8,0.Kultur
dilakukandengan
mengunakanwadah
kaca
denganmemperhatikan
sisa
ruang
diatasnya
untuknremherikan suasana fakultatif aerobik selama
21
hsr'i.
Parameter
yang diarlati
dalamfermentasi adalah dekomposisi substrat jerami dengan mengukur kandungan total glukosa yang
dibebaskan dengan menggunakan metode DNS,
dan aktivitas enzim selulase yang disekresikan.
Untuk memperoleh kondisi optimum fermentasi
digunakan perhitungan ANOVA
dan
uji
bedanyata terkecil Duncan (BNT-Duncan) pada taraf
5% menggunakan progam SPSS14.
Kurva Pertumbuhan lsolat Acp-7
Kurva pertumbuhan isolat AcP-7 diperoleh
dengan mengkultur isolat
AcP-7 pada
mediafermentasi
jerami padi pada
kondisi optimumyang
diperoleh
pada
prosedur
optimasi.Sebanyak 50 ml inokulum ditambahkan kepada
media fermentasi yang
telah
disterilkan yangberisi 500 gr jerami berukuran
60
mesh, 0,6%ekstrak khamir dalam
larutantrace
elemen.Pengukuran jumlah sel actinomycetes dilakukan secara tidak langsung pada permentasi hari ke 0,
3, 6, 9, 12,15, 18,21,24,27 dan
30.
Sebanyak1
gram
media yang sudah
diinokulasikandimasukkan
ke
dalam100 mL
larutan garamfisiologis lalu di homogenisasikan hingga merata.
Pengenceran dilakukan dengan menggunakan
larutan yang sama sampai tingkat pengenceran
optimum agar sel dapat dihitung.
Sebanyak '100 pL campuran biakan dalam
larutan garam
fisiologis
di
kultur
dengan menggunakan teknik pour plate pada media YMAyang terdiri dari (g/L) ekstrak khamir
4;
ekstrakmalt 10;
glukosa
14,
agar-agar15.
Biakandiinkubasi pada suhu
37
oC selama14
hari,koloni
actinomycetesyang tumbuh
dihitungsebagai jumlah sel per-ml (cfu mL-'). Data yang
diperoleh diplotkan sebagai kurva pertumbuhan
AcP-7 pada media fermentasi. Untuk melihat
kurva aktivitas enzim selulase pada periode
yang sama
dilakukan
pemanenan enzimdengan menambahkan
200
ml
0,2M
bufferfosfat
pH
optimum, kemudiandiaduk
dandisaring dengan menggunakan glass wool dan
kertas saring. Filtrat yang diperoleh digunakan
untuk
mengkuantifikasi
aktivitas
enzim (Ramachandra ef al. 1987).Pengukuran
Aktivitas Enzim
dan
Kadar Glukosa.Aktivitas selulase diujikan dari aktivitas
endoglukanase dengan pengujian enzim pada
substrat CMC. Sebanyak 500 prl substrat 0,5% CMC ditambahkan 450 pl 0,2 M buffer fosfat pH
optimum. Kemudian larutan
ini
ditambahkan50
pl
ekstrakkasar
enzim dihomogenisasi,diinkubasi pacia suhu 37oC selama
30
menit lalu ditambahkan 1000pl
pereaksi DNS dansegera dipanaskan pada suhu 100oC selama
15
menitdan
didinginkan pada suhu ruang.Pengukuran absorbansi masing-masing larutan
dilakukan pada
l.
540 nm (Miller, 1959). Kadarglukosa
yang
terbentuk ditentukan denganmenggunakan kurva standar glukosa. Satu unlt
aktivitas seiulase didefinisikarr sebagai jumlah pmol glukosa yang dihasilkan per menit untuk setiap mlenzim pada kondisi optimumnya.
Pengukuran Kadar Selulosa Jerami Padi
Kandungan selulosa pada jerami padi
diukur berdasarkan metode yang
dikembang-kan oleh Van Soest dan Wine (1967) dengan
sedikit modifikasi. Sebanyak
1
gram jeranrikering
dimasukkanke
dalam
labu
bundar,tambahkan 15 ml asam asetat B0% dan 1,5 ml
asam nitrat pekat, kemudian direfiuks selama
20
menit. Selanjutnya dilakukan penyaringandengan menggunakan kertas saring Whatman
No.
91
yang
telah
diketahui bobotnya (A).Setelah dilakukan penyaringan, padatan hasil
dari refluks dicuci dengan etanol. Erlenmeyer
dan corong dibilas dengan air suling sebanyak
3 kali. Kertas saring beserta residu dikeringkan
pada oven dengan suhu 'i00 sampai 105 oC selama 1-2 jam. Kertas saring didinginkan dan
ditimbang bobotnya (B). Kertas saring dengan
residu diabukan
pada suhu
540"C,
laludidinginkan ditimbang bobotnya
(C).
Kadarselulosa
di
hitung
menggunakan rumus persamaan (1)sebagai berikut :lte,l'r
Se]#os*{yal}=
#*
x 100cr,,i ....(1)Pengukuran Kinetika Fermentasi
Laju pertumbuhan spesifik actinomycetes
dihitung secara
tidak
langsung
. denganmenghitung konsentrasi
sel
(cfu
mL')
pada media dan diplotkan pada kurva regresi ln jumlahsel
terhadapberat
biomassayang
dihitungberdasarkan
berat kering
sel
(Panda
andRameshaiah,
2009). Rasio
produk
tei-hadapsubstrat (Yp/s), rasio produk terhadap biomass
sel (Yp/x), dan rasio biomass terhadap substrat (Yx/s) dihiiung berdasarkan data biomass sel (x),
selulosa
pada
jerami
terfermentasi(s),
cianglukosa
total
(p)
selama
fermentasi. Untukrnengetahui nilai Ks (Konstanta Monod (g) pada saal
'/,
lalu
pertumbuhan spesifik maksirnum)dan p*"1. (laju pertumbuhan spesifik maksimum)
ciigunakan persamaan (2) yang dikenal dengan
persamaan Monod (Viccini
et
al., 2001) sebagaiberikut :
i/ ncks -(
u=
_
...(2)
' Xs-S
spesifik
pembentukan
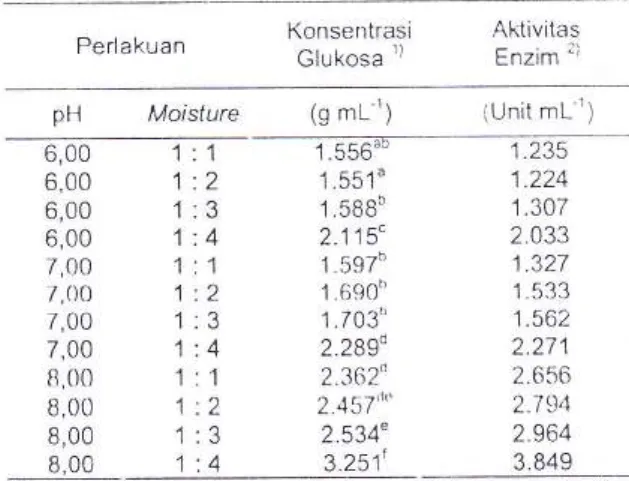
ProdukTabel 1. Hasil optimasi fermentasi padat terhadap perlakuan pH dan tingkat motsiure
Perlakuan Konsentrast Glukosa 1) Aktivitas ;'j tnztm pH Mc:isture (g mL"') iUnit mL'1;
maksimum (qu) clilriturrg dengan rnenggunakan
persamaan
(3)
dan
penggunaan
substrat maksimum (q.)dihitung dengan persamaan (4).g nak:
r?
yipIxi;l:ncks
Ys
yi-x I s)HASIL DAN PEMBAHASAN Optimasi Fermentasi Padat
Pada
penelitian
ini dikembangkan fermentasi padat dengan tujuan untuk melakukanefesiensi pengolahan terhadap limb'ah jerami
padi,
karena
fermentasipadat lebih
sedikitmenggunakan
energi,
bahan
kimia
danpenanganan substrat jerami padi pada langkah
perlakuan
awal tidak
begiturumit.
Selain ituactrnomycetes
yang
digunakan
merupakankelompok bakteri yang memiliki hyfa sehingga memungkinkan untuk melakukan kolonisasi pada
substrat padat jerami Padi.
Pengaruh pH dan moisture yang diujikan
dilihat dari dua aspek yaitu konsentrasi glukosa sebagai produk akhir hidrolisis selulosa yang
dilakukan secara enzimatis oleh actinomyceies,
dan
aktivitas enzimnya
sendiri pada
akhirfermentasi
hari
kt-?-1.
Jika
ditrnjau
dari kandungan glukosa pada media'pengaruh pl-'ldan moisture memiliki interaksi yang signifikan, Itirl irri
tli
lil,at r.letri trar;rl arralisis s{-'cnra slitlislik;t rjimana nilai F hitung lebihbesar
dariF
tabel'Tetapi hasil perhitungan untuk aktivitas enzim
'. 1 .z .a '.4 '. 1 '.2
1)
lnteraksi perlakuan pH dan moisture memberikansignifikansi pada uji F pada laral 5o/o. Arrgka yang diikuti
huruf yang sarr'la dalam satu kolom tidak berbeda nyata
pada taraf 5% paCa uji BNT-Duncan.
2)
lnteraksi perlakrran pH dan moisture tidak signitikan pada uji F padataraf 5%.
menunjukkan interaksi antara pH dan moislure
tidak
memberikan pengaruh yarrg signifikan.Dari hasil yang diperoleit fermentasi pada pH B
dan perbarrdingan moisture '1
:
4
rrerrberikanhasil
terbaik
dan
memiliki perbedaan yangnyata
pada
uji
BNT-Duncan.Data
hasilpengukuran kandr.rngan glukosa dan aktivitas
enzim seluiase pada akhir fermentasi disajikan pada Tabel 1.
Pada fermentasi padat pH yang terukur
adalah pH awal atau yang lebih dikerlal sebagai
pH
inisrasi
yang
dtkondisikan
dertganmemberikan buffer fosfat. Chellapandi and Jairi
(2008) :Tlerrgatakan untuk mengkontrol pH pada
saat fermentasi padat berjalan tidak semudah
pada fermentasi kultur cair. Hal ini disebabkan
partikel
padat
menyerap
metabolit
yangdihasilkan mikroba tidak homogen tergantung
dari
konsentrasi kolonisasi mikroba tersebut.Sehingga
homogenisasi
moisture
diawalfermentasi dengan menggunakan buffer yang
sesuai sangat nrenentukan jalannva fermentasi.
Jika
dilihatdari
hasil yang ditunjukkan padapenelitian
ini
selulase dihasilkanpada
pHinisiasi yang cenderung basa.
Kisaran
PH
untuk
selulase
bakterikhususnya
actinomycetesmemiliki
rentangyang
cukup
luas
dari
4,5
samPai
9'0(Subramaniyan and Prema, 2002). Solingen et
al.
(2001)
memperolehisolat
Streptomycesyang berasal dari danalu di Afrika l"irnur y;rrtr.;
menghasilkan selulase optimum pada
pH
B.Dari fenomena ini asal isolat diperoleh sangat 6,00 6,00 6,00 6,00 7,00 7,00 7,00 7,00 8,00 8,00 8,00 8,00 1.556"" 1.551" 1,sBBb 2.115' 1,597b 1.690" 1.703" 2.2}gd 2.362n 2.457d" 2.534" 3.251t t-zJ3 1.224 1.307 2.033 1.327 1 aa) 1.562 2.271 2.656 2.794 2.964 3.849 4 1 2 3 4 Lajt.r (3) (4)
menentukan pada pH inisiasi berapa isolat dapat menghasilkan selulase secara optimal.
Kurva Pertumbuhan lsolat AcP-7 pada Media
Fermentasi
Kurva
pertumbuhan rsolatAcP-7
padamedia fermentasi dilihat
dari
dua data
yaitujumlah sel actinomycetes dalam satuan cfu mL-'
dan aktivitas enzim yang dihasilkan dalam satuan
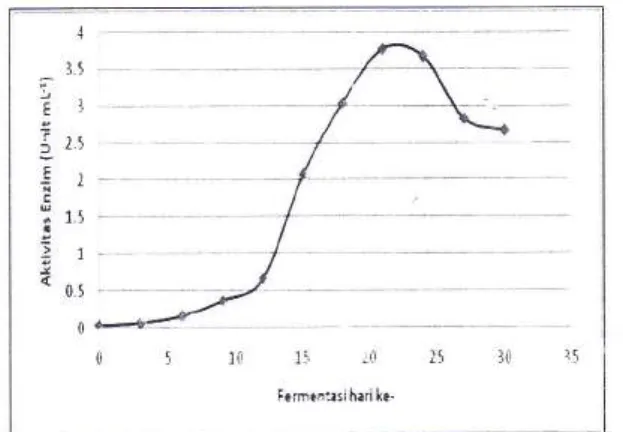
unit mL-1. Dari grafik yang diperoleh (Gambar 1
dan Gambar 2) fase eksponensial tercapai pada
hari
ke-9
sampai
hari
ke-21
fermentasi. Kepadatan sel tertinggi tercapai pada hari ke-21dengan kuantitas senesar 2,6 x10e cfu mL-1 dan
aktivitas enzim selulase juga_tercapai pada hari
ke-21 sebesar 3,765 unit mL''.
Hasil
yang
diperoleh
menunjukkanfenomena yang sama dengan apa yang telah
diteliti oleh Yamac dan Tamer (2C08), dimana
mereka menrpelajzrri tentang penguraian iignin
jerami gandum oleh isolat Streptomyces terpilih. Fase eksponensial yang diperoleh mulai tercapai
pada
fermentasiminggu
ke-2
hingga
ke-3.Sebagaimana dijelaskan oleh Hamelinck
et
ai.(2005) bahwa pada lign<;selulosa struktur ikatan hemiselulosa dan lignin terbentuk secara kovaien
dan
n,elapisi selulosa.
Struktur
ini
harusdimodifikasi dengan menghilangKan lignin untuk
menghasilkan
hidrolisis selrtlosa
danhemiselulosa lebih efisien. Dari fenomena vang
diperoleh
selulosa
terdegradasi
untukmenghasilkan
glukcsa
mulai
mencapai faseeksponensial
pacia
hari
ke-9,
hal
inimemungkinkan dicapai setelah lignin pada jerami
padi terdegradasi lebih awal. Pola pertumbuhan
sel
actinomycetes memiliki kesamaan dengan pola aktivitas enzim selulase yang dihasilkan.Laju pertumbuhan spesifik biomass isolat
AcP-7
diperoleh
dengan
memperhatikan pencapaian fase eksponensial yaitu pada harike-9
sampai hari ke-21. Gambar3.
menunjukkangrafik iinier yang
menggambar-kan lajupertumbuhan spesifik isolat AcP-7 sebesar 1,394
cfu
mL-'
per-hari. Pertumbuhan actinomycetesdibandingkan dengan bakteri
pada
umumnyarelative
lebih
lambat (Pauland Clark
1996)terlebih dalam penelitian ini digunakan substrat
jerami padi yang merupakan substrat kompleks
yang
harus diuraikan terlebih dahulu menjadisenyawaan
yang lebih
sederhana sebelumdimanfaatkan
dalam
metabolisme
selactinomyceies.
Kinetika Fermentasi
Beberapa parameter fermentasi yaitu
koefisien rendemen produk (Yp/s)
yangAcP-7 pada media padat jerami padi
Garnbar 2. Aktivitas enzinr selulase selama periode fermentasr 50
:
:40 g !rn a Ezo E 10 r=119fi-8018 ,.-' Rt-0919 -.' "t'.-{
0 05101:2025ieffi.niali h.ri ke.
Gambar 3. Grafik linier untuk mencari laju spesifik pertumbuhan aclinomycetes isolat AcP-7 pada media jerami padi
menggambarkan jumlah produk yang dihasilkan
(g)
dalamhal
ini glukcsa berbanding denganjumlah
selulosa padajerami padi
(g)
yangdikonsumsi, koefisien rendemen
biomassa(Yx/s) yang menggambarkan jumlah biomassa
sel yang
dihasilkan(g)
berbanding denganl0
-t Eq i. j E o 6 0169111s1821741110 F.m.dr3ih:rikeGambar 1. Kurva pertumbuhan actinomycetes isolat
4 t) z ) L) t c ! 0) 0
jumlah
selulosapada
jerami padi
(g)
yangdikonsumsi,
dan
koefisicn rendemenpr"oduk-biomass (Yp/x)
yang
menggambarkan jumlahproduk yarrg drhasilkan (g) berbanding biomass
yang
dihasilkan
(g)
dievaluasi
denganmenggunakan
teknik
linierisasi.Data
produk,biomass, dan substrat yang dihasilkan selama
fermentasi diplotkarr
untuk
mendapatkankoefisien yang merupakan harga dari intersept
grafik yang diperoleh. Gambar 4 sampai Gambar
6
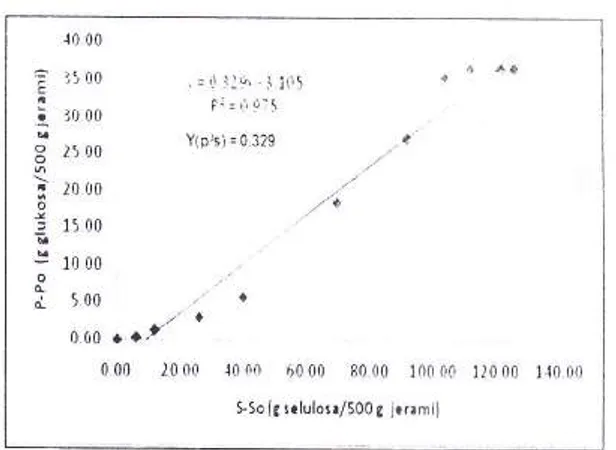
menunjukkan hasil linierisasi untuk mendapat-kan nilai-nilaiY(p/s), Y(x/s), dan Y(p/x).Koefisien rendernen produk yang diperoleh
sebesar 0,329 g glukosa per-gram selulosa yang
dikonsumsi. Pada perlakuan ini kadar selulosa di
hitung
dari jerami
yang terfermentasi derrganmetode Van Soest dan Wine (1967), sehingga
konsentrasi selulosa
yang terukur
nreruprakannilai real
yarrg dapat menggambarkan berapabesar
selulosayang
telah
terciegradasi olelrenzim selulase yang dihasilkan actinomycetes
isolat
AcP-7.
Selisih kandungan selulosa padasaat
pengukuran dibandingkan
dengankandungan selulosa awal pada jerami padi yang
digunakan dihitung sebagai nilai S-So yang akan
menjadi n;lai pembanding teriradap glukosa yang dihasilkan pada saat fermentasi.
Koefisien rendemen
biontass
yangdihasilkan sebesar 0,02 g berat sel kering AcP-7
per-gram selulosa yang dikonsumsi. Jika dilihat
dari angka yang diperoieh perturnbuhan biomass
menunjukkan hubungan tidak langsung dengan konsumsi selulosa. Hal ini dapat dipahami bahwa
sumber karbcn yang dapat digunakan langsung
untux pertumbuhan sel adalah gula sederhana
scperti
glukosa (Jaradatet al.
2008),
yangdihasilkan setelah actinonrycetes menghidrolisis selulosa.
Hasil
penelitian Chellapandiand
Jani(2008) menyatakan penambahan glukosa diawal
fermentasi dapat meningkatkan aktivitas selulase
yang
dihasilkanoleh
Streptomyces mencapai 8,05 unit mLldibandingkan dengan penainbahanselu_losa murni yang hanya mencapai 4,00 unit
mL-'.
Sedangkan rendemen produk-biomassmenunjukkan 15,16 g glukosa mampu dihasilkan
setiap '1 gr kering biomass AcP-7 .
Permodelan
kineiika
MonoC
yangdigunakarr untuk mengevaluasi
laju
maksimumproduksi glukosa (p maks) dan laju penggunaan
substrat selulosa
yang
digambarkan dengankonstanta Monod (Ks) menunjukkan bahwa nilai
p maks sebesar 31,25 dan konsentarasi substat pada saat setengah dari laju maksimum produksi gtukosa (Ks) sebesar 74,34 g.
Gambar 4. Graflk linier antara P-Po dan S-So untuk mencari nilai Y(p/s)
Gambar 5. Grafik iinier antara X-Xo dan S-So uniuk mencari nilai Y(x/s)
l.xc 1€ biofirit/500 I j.rrfli1
Gambar 6. Grafik linier antara P-Po dan X-Xo untuk mencari nilai Y(p/x)
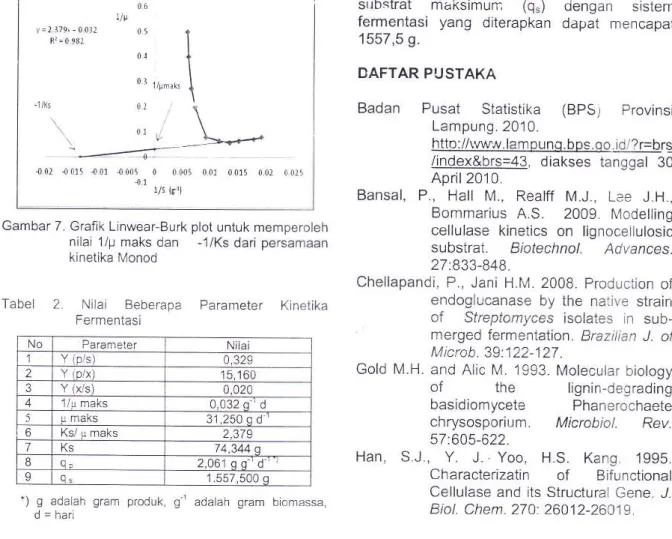
Nilai
p maksdan
Ks diperoleh denganmemplotkan nilai 1ip yaitu invers dari laju reaksi
produksi glukosa
dan 1/S
yaitu
invers darikonsentrasi
selulosa
yang
dikenal
denganLinewear-Burk
plot.
Garislinier
ditarik untukmendapatkan
nilai 1lV
maks
dan
-1lKst0 00 - ?,too -1., 16,.r1n ' : ll - (r 1).'l : 1o u{l 3 1 5.00 y{p rr : ,5.16 I
)
zooo o'o ! l\oo /. 3 to.oo .z' i loo ,,u t ooo 'l '' ll 00 {1 \ir I Lrrr I i0 I 0il 10 00 i .tll rr,q rr,,q r ' rr . c.-,r.r-( a 1000 ; yloJs::03?9 . 3 )500 /i
zooo " .'4 r 15 00 - 1000 o ,' d 500 t /a lto ...J 0 00 20 00 l0 00 60 00 80 00 100 00 lto 00 1.10 00 !!o I1 rrlulo:r/500 j lcrrml) 3.00 E tsn g j loo i tt0 E 5 l$(t:
,: o5n ./ *z' 0u0 -o i ]:i10:0\-0f5.1 Pr.0 9!9 Yi,tJs) =6.92 I 000 2000 ri0{ 6000 3000 100trri llrtq t{00,,sehingga nilai-nilai
p
maksdan Ks
diketahui (Gambar 7).Model
ini
mengikuti
kinetika
yangdijelaskan
oleh
Michaelis-Menten, walaupununtuk
diterapkanpada kultur padat
modetkinetika ini memiliki kelemahan terutama dalam
hal
homogenitas dan deaktivasi enzim karena adsorbsi enzim pada material padatan (Bansal elal.
2009),
tetapi
penggunaannya pernah dilakukan oleh Movagharnejad ef a/. (2005) untukmengevaluasi kinetika selulase komersial (Novo,
Denmark) dengan hasil konversi selulosa diatas
70%
meskipun menghadapikendala
dalamkemampuan mengakses
sisi
aktif
enzim.Beberapa nilai parameter kinetika disajikan pada Tabel 2.
Dari
Tabel2
dapat ditihat bahwa tajuspesifik pembentukan produk maksimum (qo)
sebesar 2,061 glg biomassa
per-hari.
Hasil inimenunjukkan bahwa
laju
degradasi selulosa pacja jerami pacii oleh selulase yang dinasilkanactinomycetes
isolat AcP-7 cukup
tinggi.Sedangkan penggunaan substrat maksimum (q.)
dengan sistem fermentasi yang diterapkan dapat
mencapat 1557,5 g.
Gambar 7. Grafik Linwear-Burk plot untuk memperoleh nilai 1/p maks
dan
-1lKs dari persamaan kinetika MonodTabel
2.
Nilai
Beberapa Parameter Kinetika FermentasiNo Parameter Niiai
1 Y /n/s) 0,329
2 Y (o/x) 1 5,1 60
J Y (x/s) 0,020
4 '1lu maks 0.032 o-' d
5 u maks 31,250 q d-' 6 Ks/ u maks 2,375 7 Ks 74 244 a o a" 2.061 o o'' d-'
I
a. 1.557,500 qBelum
banyak
hasil
penelitian yangmengeksplorasi kinetika penguraian selulosa
dari
jerami
padi
menggunakan
isolatactinomycetes, sehingga data yang disajikan
masih
sulit
untuk
dibandingkan dengan pencapaian peneliti lainnya.KESIMPULAN
Dari
penelitianyang telah
dilakukandapat
disimpulkanbahwa kondisi
optimumfermentasi padat isolat AcP-7 pada substrat
jerami padi pada pH 8,00 dan perbandingan
substrat:moisfure
1:4.
Fase
eksponensialtercapai pada fermentasi hari ke-9 sampai hari
ke-21
dengan
laju
pertumbuhan spesifikbiomass
sebesar 1,394
cfu
mL-1 per-hari.Kcefisien rasio produk terhadap substrai (yp/s)
sebesar 0,329, rasio produk terhadap biomass
sel (Yp/x) sebesar 15,160, dan rasio biomass
terl'radap substrat (Yx/s) sebesar 0,020. Laju
maksimum prociuksi glukosa (p.maks) sebesar 31,25 g per hari dengan Konstanta Monod (Ks)
sebesar
74,34
g,
sedangkanlaju
spesifikpembentukan produk maksimum (qo) sebesar
2,061
glg
biomass per-haridan
penggunaansubstrat
maksimurn
(q.)
dengan
sistemfermentasi
yang
diterapkan dapat mencapat 1557,5 g.DAFTAR PUSTAKA
Badan
Pusat
Statistika
(BPSl
Provinsi Lampung. 2010.htto :/iivww. lam punq. bps.qo. id/?r=brq
/index&brs=43, diakses tanggal 30
April 2010.
Bansal,
P.,
Hall
M.,
RealffM.J., Lee
J.H.,Bommarius
A.S.
2009. Modellingcellulase kinetics
on
lignocellulosicsubstrat.
Biotechnol.
Advances.27:833-848.
Chellapandi. P., Jani H.M. 2008, Production of endoglucanase by the native strain
of
Streptomyces isolatesin
sub-merged fermentation. Brazilian J. of Microb. 39:122-127 .
Gold M.H. and Alic M. 1993. Molecutar biology
of
thebasidiomycete
lignin-degrading Phanerochaete
chrysosporium.
Microbiol.
Rev.57:605-622.
Han, S.J.,
Y.
J."
Yoo, H.S.
Kang.
1995.Characterizatin
of
BifunctionalCellulase and its Structural Gene. J.
Biol. Chem. 27 O: 26012-260'1 9.
-) g adalah
d = hari
gram produk, g-1 adalah gram biomassa,
0.6 rh y = 2.179t - 0.012 0.5 Rr=0981 0..t 0t., I /pmaxs -1lKs o.z ,, \r'
\
or
'\v
{.02 {.015 {.01 { 005 0 0.005 0 01 0.015 0.02 0.025 {.1 l/s {r'l\
\_*
Howard, R.1., Abotsi E., Jansen van Rensburg, E.L. Howard
S.
2003. Lignocellulosebiotechnology:
issues
ofbioconversion
and
enzyme production. Afr. J. Biotechnol.2:
602-01 9.Hamelinck, C.N., Geertje
van
Hooijdonk, FaaijA.P.C. 2005.
Ethanol
fromlignocellulosic
biomass:
techno-economic
performancein
shcrt-,middle- and long-term. Biomass and
Bioe n e rg v. 28: 384-41 O.
Jaredat, 2., Dawagreh A., Ababneh Q., Saadourn
l.
2008.
lnflerrce
of
culture
oncelluiose production Slrepfomyces Sp.
(strain J2). Jorcian
J.
of
Biol. Sci. 4:141-146.
Miller Gl. 1959. Dinitrosalisilic assay. Anal Chem 31:426-428
Movagharnejad, K., Sohrabi M.
2003.
A
modelfor the rate cf enzymatic hydrolysis of
some
cellulosicwaste
materials inhetcrogenous
solid-liquid
system. Biochem Eng J.14:1-BPanda, T., Rameshaiah G.N. 2009. Application of
actinobacterial and fungal morphtlogy on the design of operating strategis in
bioprocess developmenl. The Open
Biotechnol../. 3:31 -39
Paul EA, Clark
FE.
'1996. Soi/ Microbiotogy and Biochemistry. 2no ediiion. Caiifornia:A.cacjemic Press; 30:69-99
Ranrachandra,
M.,
Crawfo;'dD.L.,
Pametto lllA.L.
1987.
Extracellulai
enzymeactivities
during
l!gnoceilulosedegradation by Streptomyces spp.j a
.
comparative
study
of
wild-typegenetically'nanipulated strains. App. and Env. Microb. 53:2754-2760.
Solingen, V.P., Meijer D., Kleij W.A., Branett C.,
Bolle
R.,
PowerS.D.,
.Jones B.E.2C01. Cioning and expression
of
anendocellulase
gene
from
novelstreptomycet isolated
fronr
an
EastAfrican soda
lake.
Extremophiles.tr.aa') a / I
J.JJJ-JN I.
Subramanryan S, Prerna
P.2002.
Biotechnoiogy of microbial xylanases: enzymology,molecular biclogy, and application, Criticai Rev Brotechnol 22:33-54.
Taherzadeh,
M., and
Karimi
K,
2008.Preti'eatment
of
lignoceilulosicwaste
to
irnprove
ethanol
andbiogas production:
a
review. 1nl. J.lvlol. Sci. 9'.1621-1 65'1.
Van Soest, P.J. and R.H. Wine. 1g67. Use of
detergents
in
the
analysis
offibrous feed. lV. Determination of
plant
cell-wall
constituents. J, Assoc. Anal. Chem. 50: 50-55.Viccini, G., Mitchell D.A., Boit S.D., Gern J.C.,
da
Rosa
A.S.,
Costa
R.M.,Dalsenter F.D.H., von Meren O.F.,
Krieger
N.
2001.
Analysis ofgroMh
kinetic
profilesin
solid-state fermentation. Food Technot. Biotech nol. 39:27 1 -294.
Wendisch, F.K., Kurtzner
H.J.
1992. Role ofStreptomycetaceae
inbiodegradation
in:
Balows,
A.,Truper,
H.,
Dworkin,M.
Harder,W.,
Schleifer,K.H. editor.
TheProcaryotes, A Handbook On The Bacteria : Ecophysiology, Isdation, ldentification Apptication.
/d
e,1.vol 1. Springer-Verlag. New york.
Wynan, C.E.
2002. Potential synergies andchallenges
in
refining cellulosicbiomass
to
fuels.
BtotechnolProgress.
ig
.. 254-262.Yarnac,
l/.,
and
Tamer
AU.
20Cg. Lignindegradation and acid p:ecipitable