Botani dan Morfologi Tanaman Kakao
Tanaman kakao (Theobroma cacao L.) termasuk tanaman kaulifloral, yaitu jenis tanaman yang membentuk bunga dan buah pada batang dan cabang yang tua. Tanaman kakao dapat tumbuh pada dataran rendah tropis dengan ketinggian hingga 1.000 m dpl, dimana temperatur rendah merupakan faktor pembatas. Tanaman kakao juga memerlukan curah hujan yang merata sepanjang tahun dengan minimal 90-100 mm per bulan. Kakao dapat tumbuh dengan baik pada tanah dengan kisaran pH 6.0-7.0 (Pusat Penelitian Kopi dan Kakao Indonesia 2004).
Kakao merupakan tanaman dikotil tahunan dari famili Sterculiaceae dan mempunyai jumlah kromosom 2n = 20. Tanaman dewasa dapat mencapai tinggi 6-9 m. Bunga kakao berwarna merah muda sampai putih, reguler, hermafrodit dan memiliki lima sepal, lima petal, 10 stamen yang tersusun dalam dua lingkaran, dan sebuah ovari superior yang merupakan gabungan dari lima carpel. Bunga kakao muncul secara bergerombol pada bantalan bunga, yaitu jaringan yang menebal yang terbentuk pada ketiak bekas menempelnya tangkai daun. Sejak bakal bunga muncul pada bantalan tersebut sampai bunga mekar diperlukan waktu sekitar 30 hari. Bila pada saat mekar bunga tidak mengalami penyerbukan, maka bunga akan segera gugur (Tjasadihardja 1987).
Iklim merupakan salah satu faktor yang berpengaruh terhadap pola pembentukan bunga. Pola curah hujan di suatu daerah dapat menentukan pola pertumbuhan vegetatif dan generatif tanaman kakao. Bunga kakao dapat terbentuk sepanjang tahun tetapi intensitas pembentukannya beragam dari waktu ke waktu. Bunga banyak terbentuk selama musim hujan dan bulan-bulan lembab
berikutnya. Di Pulau Jawa, musim pembungaan besar biasanya dimulai pada bulan Nopember (awal musim hujan) dan berlangsung terus sampai bulan Juni, dengan puncak pada bulan Desember-Januari. Pembungaan besar tersebut akan menghasilkan panen besar pada bulan Mei-Juni. Peningkatan pembentukan bunga selama musim hujan diduga berkaitan juga dengan pergantian dari lingkungan kering selama musim kemarau ke lingkungan basah selama musim hujan (Tjasadihardja 1987).
Meskipun tanaman kakao dapat berbunga berkali-kali sepanjang tahun, tetapi pembungaan yang utama terjadi pada saat hujan pertama setelah musim kering. Tanaman mulai berbunga setelah berumur dua tahun sejak tanam, dan untuk mencapai buah matang diperlukan waktu 5-6 bulan setelah polinasi. Buah yang telah masak dinamakan pod dan ukurannya dapat mencapai diameter 10 cm dan panjang 30 cm. Biji kakao kaya akan karbohidrat dan lemak, sehingga merupakan sumber energi yang baik. Kakao juga mengandung senyawa theobromin yang secara kimia dapat dikonversi menjadi kafein yang dapat digunakan untuk minuman maupun keperluan kesehatan. Selain itu kakao juga mengandung senyawa antioksidan (Mims 1998). Karena itu pemanfaatan biji kakao tidak hanya untuk memenuhi bahan makanan, tetapi juga untuk keperluan farmasi (Potts 2002).
Faktor musim terutama musim kemarau mempengaruhi perkembangan dan produksi tanaman kakao. Pada musim kemarau intensitas cahaya matahari sangat tinggi karena derajat awan rendah, kelembaban udara sangat rendah, sebaliknya temperatur lingkungan meningkat sangat tajam, sehingga tanaman kakao mengalami cekaman air (Suhadi 2002). Dalam kondisi seperti ini tanaman kakao menjadi sulit berbunga dan akibatnya produksinya juga tertunda.
Fisiologi Pembungaan
Pembungaan tanaman merupakan kejadian fisiologis yang kompleks yang secara morfologi terjadi perubahan dari fase vegetatif menuju terbentuknya organ-organ bunga. Proses pembentukan bunga tersebut secara garis besar terdiri atas empat tahap yaitu inisiasi bunga, diferensiasi bunga, pendewasaan bunga dan anthesis. Dari keempat tahap tersebut, fase inisiasi bunga merupakan fase yang paling kritis dari pembungaan yang merupakan proses biokimia dari fase vegetatif ke arah reproduktif, namun secara morfologi tidak nampak (Ryugo 1988).
Terdapat beberapa konsep yang mendasari para peneliti mempelajari proses pembungaan. Konsep pertama yaitu konsep nutrisi yang menerangkan bahwa pembungaan dikontrol oleh keseimbangan karbohidrat dan nitrogen atau nisbah C/N (konsep Kraus and Kraibill), dan konsep yang kedua yaitu konsep hormonal yang menjelaskan bahwa proses pembungaan pada tanaman tertentu diatur oleh zat pendorong pembungaan (florigen). Hormon ini diproduksi oleh daun kemudian ditranslokasi ke sebagian tunas yang akan memproduksi organ generatif (Ryugo 1988). Analisis nisbah C/N pada Sinapsis alba dan Arabidopsis yang sedang diinduksi bunganya, menunjukkan bahwa nisbah C/N pada cairan floem meningkat dengan nyata dan cepat, yang mengindikasikan bahwa penurunan suplai N dan peningkatan C organik ke meristem apikal merupakan kejadian penting pada transisi pembungaan (Corbesier et al. 2002).
Dalam pembungaan tanaman, transisi dari fase pertumbuhan vegetatif ke fase reproduktif merupakan proses perkembangan yang kritis, yang ditandai oleh sejumlah perubahan pada apeks tunas pada tingkat molekuler, fisiologi dan morfologi. Dengan kondisi lingkungan yang dapat menginduksi dan kemampuan tanaman untuk merespon faktor eksternal ini, meristem vegetatif akan
berkembang menjadi meristem reproduktif, yang mana dapat secara langsung menjadi infloresen dan kemudian menghasilkan banyak bunga (Immink et al. 1999). Pada kebanyakan tanaman, faktor lingkungan (terutama panjang hari dan temperatur) berperan mengendalikan transisi dari meristem vegetatif ke meristem bunga. Meristem vegetatif menjadi meristem infloresen yang selanjutnya akan membentuk satu atau lebih meristem bunga (Staveley 2005).
Dalam kondisi yang menginduksi pembungaan, meristem tunas utama dari tanaman Arabidopsis menghasilkan meristem bunga lebih cepat daripada meristem daun yang mengapitnya. Selama fase vegetatif, meristem tunas utama dari Arabidopsis menghasilkan daun roset dengan jarak yang sangat dekat. Transisi ke fase reproduksi, yang dikontrol dengan ketat oleh jaringan kompleks dari gen pengontrol waktu pembungaan, dipengaruhi oleh sinyal lingkungan, seperti panjang hari, kualitas cahaya dan temperatur, sebagaimana faktor internal seperti umur tanaman.
Pada beberapa spesies, waktu pembungaan terutama dipengaruhi oleh faktor lingkungan yang memberikan kondisi pertumbuhan yang baik untuk reproduksi seksual dan pemasakan biji. Faktor ini meliputi fotoperiod atau panjang hari, kualitas cahaya, kuantitas cahaya, vernalisasi, dan ketersediaan nutrisi dan air. Pembungaan dapat juga diinduksi oleh stres seperti defisiensi nutrisi, kekeringan dan kerapatan tanaman (Levy dan Dean 1998). Sementara itu Ratcliffe dan Riechmann (2002), mengatakan bahwa transisi pembungaan merupakan proses yang plastis, yang selain dipengaruhi oleh faktor endogen juga faktor lingkungan, yang meliputi panjang hari, kualitas cahaya, temperatur, stres biotik dan stres abiotik.
Aplikasi GA jarang efektif untuk menginduksi pembungaan pada tanaman short-day. Pada umumnya GA menghambat pembungaan pada tanaman berkayu angiosperm, meskipun dapat memacu pembungaan pada conifer
(Pharis dan King 1985). Pada satsuma mandarin (Citrus unshiu Marc.), GA1+3
dapat menghambat pembentukan kuncup bunga. GA eksogen memperlihatkan penghambatan terhadap pembentukan kuncup bunga tidak hanya pada citrus, tetapi juga pada apel, pear, cherrie dan peach. Bahkan tidak hanya GA eksogen yang menghambat, tetapi GA endogen juga menurunkan pembentukan kuncup bunga (Koshita et al. 1999).
Berdasarkan pada penelitian-penelitian di atas, maka dalam pengaturan pembungaan diperlukan zat yang dapat menghambat biosintesis giberelin yang kemudian disebut retardan. Namun demikian, pemberian retardan selain mendorong pembungaan juga dapat mengakibatkan dormansi tunas. Untuk itu diperlukan usaha pemecahan dormansi pada tunas yang sudah terinduksi bunganya.
Senyawa Penginduksi Pembungaan
Zat penghambat tumbuh (retardan) merupakan zat yang mempunyai efek fisiologis memperlambat pertumbuhan vegetatif dan dapat mendorong pembungaan pada tanaman tertentu yaitu dengan menghambat pembelahan dan pembesaran sel subapikal. Retardan ini menghambat sintesis giberelin, sehingga akan menghambat pembelahan dan pembesaran sel (pertumbuhan vegetatif) dan mendorong pertumbuhan generatif.
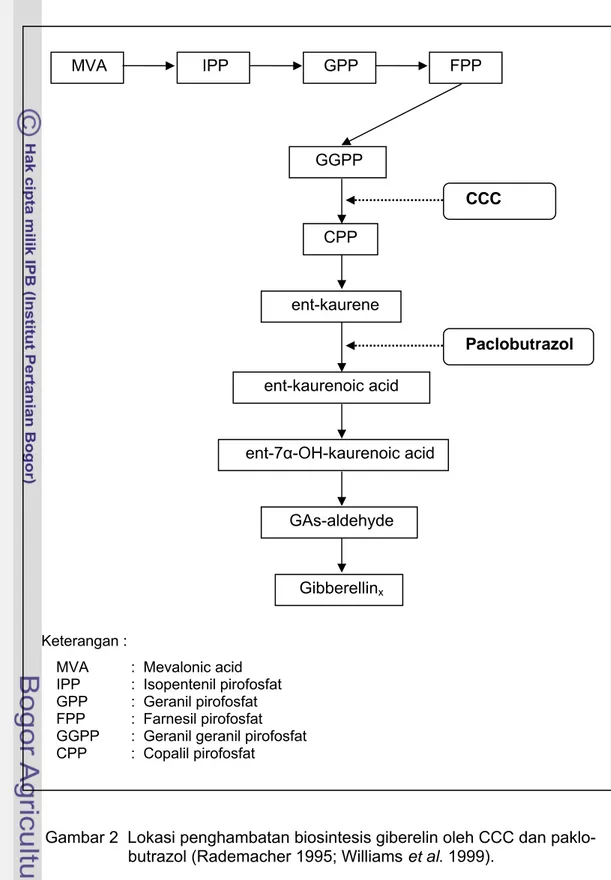
Dari kelompok senyawa triazol yang digunakan untuk meginduksi pembungaan pada tanaman berkayu, paklobutrazol adalah yang paling umum digunakan (Meilan 1997). Dasar teori penggunaan paklobutrazol adalah bahwa senyawa ini dapat menghambat biosintesis giberelin. Paklobutrazol menghambat biosintesis giberelin pada oksidasi ent-kaurene menjadi ent-kaurenoic acid (Gambar 2). Hasil penelitian yang dilakukan oleh William dan Edgerton (1983) menunjukkan bahwa paklobutrazol yang diaplikasikan pada pohon apel berumur
25 tahun dengan dosis 2 g/m2 melalui penyiraman akar dapat mengendalikan
pertumbuhan vegetatif dan meningkatkan hasil.
Hasil serupa terjadi pada tanaman Eucalyptus nitens dan E. globulus, suatu tanaman berkayu yang berbunga secara biennial atau sporadis, dengan aplikasi paklobutrazol melaui penyiraman, penyemprotan lewat daun, atau injeksi batang ternyata dapat menginduksi inisiasi kuncup bunga. Paklobutrazol juga diketahui dapat menurunkan level giberelin endogen pada beberapa spesies, meningkatkan aktivitas reproduktif, menekan pertambahan tinggi dan produksi daun (Hasan 1993; Moncur dan Hasan 1994). Karena itu induksi pembungaan dengan paklobutrazol merupakan metode praktis dalam memacu pembungaan untuk keperluan pemuliaan dan produksi biji.
Perlakuan paklobutrazol dapat menyebabkan pembungaan pada pohon durian. Aplikasi senyawa paklobutrazol dengan dosis 15 g/pohon dapat menghasilkan pembungaan secara nyata pada pohon durian (Utama 2003). Aplikasi paklobutrazol juga dapat meningkatkan kandungan karbohidrat dalam jaringan kayu. Kandungan karbohidrat ini merupakan sumber energi untuk pembentukan bunga. Namun secara tidak langsung aplikasi paklobutrazol dapat meningkatkan biosintesis asam absisat (ABA), sehingga mengakibatkan terjadinya dormansi tunas. Untuk itu diperlukan penyemprotan dengan zat pemecah dormansi sesudah aplikasi paklobutrazol (Usman 1997).
Pada tanaman mangga, perlakuan paklobutrazol mampu menginduksi pembungaan di luar musim dengan cara menghambat biosintesis giberelin sehingga dapat menstimulir pembungaan dan meningkatkan munculnya tunas reproduktif. Perlakuan paklobutrazol dengan dosis 1.0 g bahan aktif/pohon dan waktu pemberian ethephon 60 hari sesudah aplikasi paklobutrazol mempunyai potensi pembungaan terbaik dengan mempercepat saat berbunga, meningkatkan
jumlah ranting berbunga, meningkatkan jumlah malai normal dan meningkatkan jumlah bunga sempurna (Usman 1997).
Keterangan :
MVA : Mevalonic acid IPP : Isopentenil pirofosfat GPP : Geranil pirofosfat FPP : Farnesil pirofosfat GGPP : Geranil geranil pirofosfat CPP : Copalil pirofosfat MVA IPP GPP FPP GGPP CCC CPP ent-kaurene Paclobutrazol ent-kaurenoic acid ent-7α-OH-kaurenoic acid GAs-aldehyde Gibberellinx
Gambar 2 Lokasi penghambatan biosintesis giberelin oleh CCC dan paklo- butrazol (Rademacher 1995; Williams et al. 1999).
Aplikasi paklobutrazol pada tanaman mangga juga dapat menyebabkan dormansi mata tunas yang berkepanjangan, sehingga mata tunas yang mungkin sudah terinduksi menjadi bakal bunga tersebut tidak segera muncul. Karena itu, penyemprotan dengan zat pemecah dormansi setelah aplikasi paklobutrazol diharapkan dapat mempercepat munculnya bunga dan meningkatkan jumlah bunga. Terdapat beberapa bahan kimia yang dapat memecahkan dormansi pada pohon buah-buahan, diantaranya yang telah berhasil mempercepat munculnya bunga dan meningkatkan jumlah bunga pada tanaman jeruk siem, yaitu ethephon 200 ppm, KNO3 20 g/l atau BA 100 ppm, yang disemprotkan pada
1-2 bulan setelah aplikasi paklobutrazol (Poerwanto dan Susanto 1996).
Aplikasi paklobutrazol didasarkan pada ukuran kanopi pohon dan biasanya dengan kisaran dosis 1.0-1.5 g bahan aktif per pohon (Nartvaranant et al. 2000). Pada pohon lychee, aplikasi paklobutrazol melalui penyiraman tanah dengan dosis 1.0 atau 1.5 g b.a./m2 dapat mengurangi terjadinya flush
selama periode pembungaan dibandingkan dengan kontrol (Chaitrakulsub et al. 1992). Pada pohon E. nitens, paklobutrazol dan juga CCC dapat mengontrol pertumbuhan vegetatif dan merangsang pembungaan, serta menurunkan level GA1 dan GA20 pada apeks sebagai akibat terjadinya penghambatan biosintesis
GA pada daerah tersebut (Williams et al. 1999).
Chlormequat chloride (CCC) juga merupakan senyawa yang bersifat anti giberelin, yang menghambat biosintesis giberelin pada tahap pembentukan copalil pirofosfat dari geranil geranil pirofosfat (Gambar 2). Pemberian CCC dapat menurunkan pertumbuhan batang, daun dan stolon, tetapi dapat memacu pengumbian kentang (Sharma et al. 1998). Penyemprotan CCC juga dapat meningkatkan kandungan gula pereduksi, pati dan sukrosa pada batang. Sukrosa merupakan salah satu produk akhir dari proses fotosintesis dan merupakan bentuk utama dari gula yang ditranslokasikan pada kebanyakan
tanaman. Menurut Latimer et al. (2001), CCC biasanya diaplikasikan melalui foliar-spray dengan konsentrasi 1.500 sampai 3.000 ppm. Pada Hibiscus, aplikasi CCC dengan konsentrasi 1.000 ppm dapat memacu pembungaan lebih awal dan menghasilkan jumlah bunga yang lebih banyak.
CCC secara signifikan menghambat pertumbuhan tunas pada mangga, baik pada tanaman muda maupun pada tanaman dewasa. Pengaruh perlakuan CCC terhadap penurunan pertumbuhan terjadi lebih nyata pada tanaman dewasa daripada pada tanaman yang masih muda. Lebih lanjut, penghambatan pemanjangan tunas secara linier terjadi pada pohon dewasa dan muda dengan konsentrasi CCC yang lebih tinggi. Penurunan pemanjangan secara linier pada tanaman dewasa terjadi dengan perlakuan CCC 2.000 ppm, sedangkan pada tanaman muda dengan konsentrasi CCC 4.000 ppm. CCC menyebabkan penurunan produksi giberelin pada daun muda yang pada akhirnya akan menurunkan produksi auksin pada meristem apikal (Maiti et al. 1972).
Pemberian CCC pada berbagai tanaman dapat memperbaiki pem-bungaan dan membuat tanaman lebih kompak, dengan pertumbuhan tunas yang seragam. Pada tanaman jeruk, ketika senyawa penghambat tumbuh tersebut digunakan untuk menstimulasi hasil pada pohon muda, maka pengaruhnya terhadap pemanjangan dan vigor menjadi kurang efektif dan sebaliknya akan memacu pembungaan dan pembentukan buah. Aplikasi CCC dengan konsentrasi 500, 1.500, dan 3.000 ppm pada pohon jeruk muda yang berumur 11 bulan dapat menginduksi saat pembungaan yang lebih cepat dan memperbaiki pembentukan buah. Penurunan pemanjangan tunas pada tanaman yang diperlakukan dengan CCC dapat menghasilkan penampilan pohon yang kompak. Dengan perlakuan CCC, pembentukan buah diinisiasi lebih awal dan hasil akhir juga meningkat. Buah dari tanaman yang diperlakukan dengan CCC mempunyai jumlah biji yang lebih banyak daripada buah yang berasal dari
tanaman kontrol. Kandungan bahan kering buah sebagai bagian dari bahan kering total menjadi meningkat, dan hanya terjadi penurunan yang relatif kecil pada daun dan akar. Penurunan total bahan kering yang dihasilkan per tanaman secara keseluruhan hanya terjadi pada konsentrasi CCC yang paling tinggi (3.000 ppm), tetapi distribusi relatif dari bahan kering ke berbagai bagian tanaman dipengaruhi oleh CCC pada semua level (Salomon 1981).
Studi fisiologi menunjukkan bahwa terdapat biomolekul kecil yang terlibat dalam transisi pembungaan. Molekul tersebut meliputi gula, sitokinin dan giberelin (GA). Studi pada Sinapsis alba, setelah induksi pembungaan, konsentrasi molekul tersebut pada apeks meningkat dengan cepat dan nyata (Bernier et al. 1993). Hal ini membuktikan bahwa secara genetik pembungaan dikontrol oleh gula (Levy dan Dean 1998). Pada Arabidopsis, GA dapat memacu pembungaan paling tidak dalam mengaktifkan ekspresi LFY. Blazquez et al. (1998) juga menganalisis pengaruh langsung dari GA dengan dan tanpa sukrosa terhadap aktivitas promoter LFY. Perlakuan GA saja tidak memberikan pengaruh, perlakuan sukrosa saja menghasilkan sedikit peningkatan, sedangkan jika keduanya diberikan secara bersamaan dapat memberikan pengaruh sinergis (Levy dan Dean 1998). Meilan (1997) juga melaporkan bahwa karbohidrat endogen memainkan peranan dalam mengontrol induksi pembungaan pada pohon buah-buahan. Karbohidrat dapat merefleksikan status metabolit dari suatu tanaman. Hubungannya dengan proses pembungaan, bahwa sukrosa ini akan menstimulasi ekspresi LFY yang selanjutnya akan menstimulasi pembungaan (Zufall 2002).
Walaupun gula telah diketahui dapat memacu transisi pembungaan pada beberapa spesies tanaman, tetapi dalam konsentrasi yang tinggi (5%) sukrosa dapat menunda waktu pembungaan secara nyata pada Arabidopsis dan menyebabkan peningkatan jumlah daun pada waktu berbunga. Tertundanya
transisi pembungaan disebabkan oleh bertambahnya fase vegetatif, yang diakibatkan oleh lambatnya aktivasi ekspresi LFY. Konsentrasi sukrosa 1% dapat memacu transisi pembungaan dari mutan yang lambat berbunga. Hasil ini menunjukkan bahwa gula dapat mempengaruhi transisi pembungaan dengan mengaktifkan gen yang berperan mengontrol transisi pembungaan, bergantung pada konsentrasi gula, latar belakang genetik tanaman dan kapan gula tersebut diberikan (Ohto et al. 2001).
Perubahan Zat Endogen selama Induksi Pembungaan
Pada spesies tertentu pertumbuhan batang dan daun terhenti pada saat terjadi pembungaan. Alokasi asimilat pada tanaman yaitu untuk pertumbuhan vegetatif yang meliputi pertumbuhan akar, batang dan daun, dan pertumbuhan reproduktif yang meliputi pembentukan bunga, buah dan biji. Fase induksi dalam proses pembungaan merupakan fase paling penting yang menentukan apakah tanaman tersebut akan berbuah atau tidak. Pada fase ini terjadi perubahan fisiologis atau biokimia pada mata tunas dari pertumbuhan vegetatif menuju pertumbuhan generatif. Perubahan fisiologis atau biokimia yang terjadi pada fase induksi pembungaan tersebut antara lain meliputi perubahan kandungan karbohidrat, nitrogen, asam amino dan hormon (Poerwanto 2003).
Giberelin merupakan salah satu hormon yang berperanan penting pada proses pembungaan tanaman. Giberelin adalah faktor endogen yang dapat menghambat pembungaan jeruk disamping beberapa pohon buah-buahan lainnya, dan induksi bunga tersebut memerlukan penurunan aktivitas hormon giberelin (Krajewski dan Rabe 1995). Aktivitas zat mirip giberelin pada daun jeruk Satsuma yang terinduksi bunganya lebih rendah daripada yang tidak terinduksi (Poerwanto dan Inoue 1990). Sejalan dengan hal tersebut, muncul pemikiran
bahwa perangsangan pembungaan dapat dilakukan dengan perlakuan yang menghambat biosintesis giberelin (Koshita et al. 1999).
Krajewski dan Rabe (1995) menyebutkan bahwa pembungaan pada jeruk meliputi banyak proses fisiologi. Salah satu proses fisiologi tersebut adalah yang berkaitan dengan hormon giberelin. Giberelin secara endogen dapat menjadi penghambat pembungaan bagi tanaman. Berdasarkan studi pembungaan pada jeruk, ditemukan senyawa GA1 dan GA4 terdapat dalam konsentrasi yang lebih
tinggi pada pucuk vegetatif dibandingkan pada pucuk berbunga. Disamping itu kedua jenis giberelin tersebut ditemukan pada konsentrasi yang rendah pada saat terjadinya pembungaan. Salah satu fungsi fisiologis giberelin adalah pemanjangan batang dan meningkatkan pertumbuhan daun-daun muda dengan meningkatkan aktivitas pemanjangan dan pembelahan sel. Kandungan giberelin yang tinggi dapat meningkatkan aktivitas pertumbuhan vegetatif berupa pemanjangan tunas dan pertumbuhan sel pada jaringan meristem (Hooley 1994). Pada manggis, tanaman asal sambungan memiliki kandungan GA3, GA5 dan GA7
lebih rendah dibandingkan dengan tanaman asal biji (Rai 2004).
Pada kebanyakan tanaman buah-buahan, pucuk dapat terinduksi atau tidak terinduksi untuk berbunga erat kaitannya dengan perbedaan kandungan hormon tumbuh (Krajewski dan Rabe 1995; Koshita et al. 1999), perbedaan keseimbangan karbohidrat dan nitrogen serta kondisi nutrisi yang optimum bersamaan dengan perubahan-perubahan dalam tunas pucuk (Lyndon 1990; Hempel et al. 2000). Pada manggis induksi bunga juga ditandai dengan penurunan tajam kandungan giberelin dan peningkatan tajam gula total dan nisbah C/N dibandingkan dengan sebelum induksi. Pada stadium induksi terjadi peningkatan kandungan gula total dan penurunan nitrogen dibandingkan dengan sebelum induksi, baik pada tanaman asal biji maupun pada tanaman asal sambungan (Rai 2004).
Proses pembungaan dipengaruhi oleh kandungan gula total daun, nisbah C/N daun dan gula total kulit ranting. Peningkatan gula total dan penurunan nitrogen bermakna cukup penting dalam pembungaan, karena penurunan kandungan nitrogen disertai dengan peningkatan gula total dapat meningkatkan nisbah C/N, yang berperanan sangat penting dalam menginduksi bunga. Kandungan gula yang tinggi pada pucuk diperlukan sebagai sumber energi awal bagi proses induksi bunga, proses perkembangan daerah meristem dan bagian-bagian bunga. Pada lengkeng, kandungan gula terlarut meningkat tajam pada meristem pucuk yang terinduksi untuk berbunga (Prawitasari 2001).
Pentingnya peranan gula total dan nisbah C/N pada proses induksi bunga terlihat pula pada perbedaan kandungan gula total dan nisbah C/N antara pucuk berbunga dan pucuk tidak berbunga pada tanaman manggis. Pada stadium induksi, kandungan gula total dan nisbah C/N pucuk berbunga nyata lebih tinggi dibandingkan dengan pucuk tidak berbunga. Hal tersebut menunjukkan bahwa pada saat terjadi induksi bunga gula total dan nisbah C/N di pucuk meningkat dan peningkatan yang lebih besar terjadi pada pucuk-pucuk yang akan menginduksi bunga. Pucuk yang tidak mengalami peningkatan gula total secara tajam tidak mampu menginduksi bunga sehingga pucuk-pucuk tersebut tidak berbunga (Rai 2004).
Studi Molekuler Pembungaan
Pembungaan merupakan proses yang memerlukan banyak faktor yang dikontrol oleh integrasi sinyal endogen, yang berhubungan dengan faktor seperti umur tanaman dan status metabolit, dengan sinyal lingkungan seperti panjang hari, status nutrien dan temperatur. Gen yang berintegrasi dengan sinyal ini, gen pengontrol waktu pembungaan, bertindak sebagai inducer (penginduksi) atau repressor dari pembungaan. Pada Arabidopsis, gen LEAFY (LFY) mempunyai
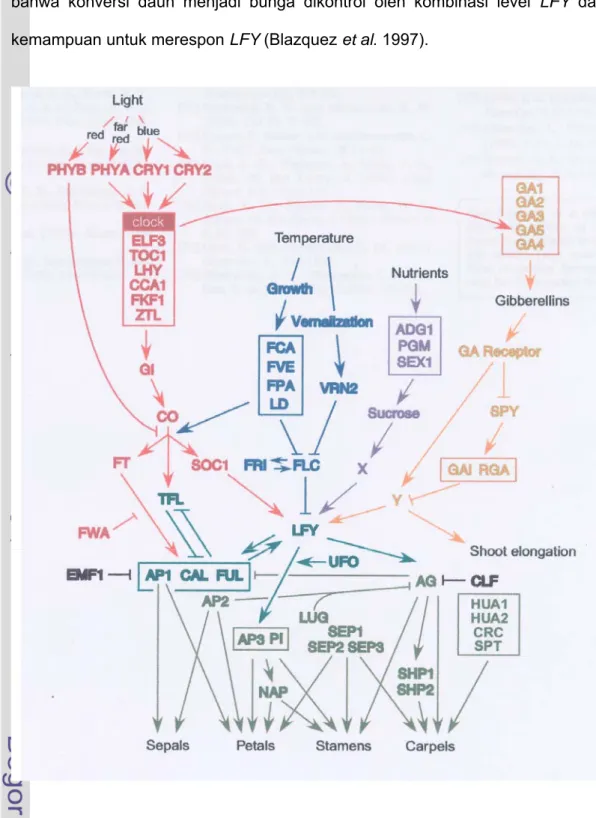
posisi sentral dan merupakan gen yang terekspresi secara absolut untuk keperluan inisiasi bunga secara normal. Gen-gen yang terlibat dalam metabolisme senyawa dapat memainkan sebagian sinyal tanaman endogen, seperti hormon tanaman giberelin dan sukrosa (status metabolit) (Nilsson 2002). Fungsi gen-gen tersebut telah banyak dilaporkan, dan dapat dikelompokkan kedalam alur genetik yang berbeda dan saling berinteraksi dalam pengaturan pembungaan (Simpson et al. 1999). Menurut Blazquez (2000) dan Zufall (2002), paling tidak terdapat empat alur atau lintasan yang mempengaruhi aktivitas gen-gen pembungaan yaitu lintasan photoperiodic, lintasan autonomous, lintasan sucrose dan lintasan gibberellin (Gambar 3).
Tahap pertama dalam perkembangan bunga adalah terjadinya transisi dari meristem vegetatif menjadi meristem bunga. Setiap meristem bunga didiferensiasi menjadi bunga yang mengandung empat tipe organ yang menempati posisi secara tepat dalam empat lingkaran konsentris. Studi genetik pada Arabidopsis thaliana dan Antirrhinum majus telah berhasil diidentifikasi gen-gen yang bertindak lebih awal yang menentukan penanda meristem bunga dan gen-gen yang bertindak pada tahapan berikutnya yang menentukan penanda organ bunga (Mandel et al. 1992). Pada Arabidopsis, perubahan perkembangan dari vegetatif ke reproduktif dikontrol oleh beberapa faktor, dan sebagai konsekuensinya muncullah beberapa kelompok gen yang terlibat dalam pengaturan tersebut. LFY merupakan gen pertama yang diekspresikan pada primordia yang mengapit meristem tunas apikal yang akan menjadi bunga. Peningkatan jumlah kopi LFY akan mempercepat waktu pembungaan, sedangkan penurunan level LFY akan memperlambat waktu pembungaan. Hasil ini menunjukkan bahwa LFY bertindak sebagai gen pengatur waktu pembungaan, yang mengindikasikan bahwa LFY merupakan switch yang menentukan antara proses induksi pembungaan dan inisiasi pembungaan, dan
bahwa konversi daun menjadi bunga dikontrol oleh kombinasi level LFY dan kemampuan untuk merespon LFY (Blazquez et al. 1997).
Gambar 3 Alur genetik perkembangan bunga pada Arabidopsis (Blazquez 2000).
Pembungaan melibatkan serangkaian aktivitas dari dua grup gen, yaitu yang mengontrol transisi dari meristem vegetatif ke bunga (gen penanda meristem bunga), dan yang secara langsung berhubungan dengan pembentukan
berbagai bagian bunga (gen penanda organ bunga) (Levy dan Dean 1998). Sejumlah gen penanda meristem bunga telah diisolasi dari Arabidopsis thaliana dan spesies tanaman model lainnya. Salah satu gen tersebut, LEAFY (LFY), terlibat dalam mengontrol transisi dari meristem vegetatif menjadi meristem bunga. Gen yang lain, APETALA1 (AP1), selain terlibat dalam mengontrol transisi dari meristem vegetatif ke meristem bunga juga diperlukan untuk perkembangan sepal dan petal. Weigel et al. (1992) memperlihatkan bahwa LEAFY berinteraksi dengan gen pengontrol pembungaan yang lain, yaitu APETALA1, untuk memacu transisi dari meristem vegetatif ke meristem bunga. Dalam alur genetik perkembangan bunga pada Arabidopsis tersebut, meristem vegetatif berkembang menjadi meristem bunga, dan kemudian menghasilkan primordia organ bunga. Pada proses ini, produk gen LFY dan AP1 berinteraksi secara sinergis untuk memacu perkembangan meristem bunga. Peranan utama dari LEAFY adalah secara langsung menekan gen penanda tunas vegetatif atau menekan faktor intermediet yang mengaktifkan gen penanda tunas vegetatif (Parcy et al. 2002). Ketika AP1 atau LFY tersebut diekspresikan secara konstitutif pada Arabidopsis transgenik secara in vitro, tanaman akan berbunga hanya dalam 10 hari (Mandel dan Yanofsky 1995; Weigel dan Nilsson 1995).
Perubahan bentuk dari meristem vegetatif menjadi meristem bunga mengakibatkan roset, hingga meristem infloresen menghasilkan daun dan bunga. Hal ini tergantung pada aktivitas dari LFY dan AP1 sebagai gen penanda meristem bunga (Bowman et al. 1993; Mandel dan Yanofsky 1995). Kedua gen tersebut diekspresikan pada level yang tinggi pada primordia bunga yang muncul, tetapi hanya LFY yang diekspresikan pada primordia daun sebelum transisi ke pembungaan. Tingkat ekspresi LFY pada primordia lateral meningkat sesuai dengan umur tanaman sampai mencapai batas ambang tertentu. Segera
setelah batas ambang ini tercapai, primordia tersebut akan berkembang dari primordia daun menjadi primordia bunga (Blazquez et al. 1997).
Pembentukan meristem bunga Arabidopsis terutama dikontrol oleh gen penanda meristem bunga yaitu LFY, AP1 dan CAULIFLOWER (CAL). Liljegren et al. (1999) dan Burnham (2000) juga menjelaskan bahwa interaksi antara LFY, AP1, dan CAL secara positif memacu fase transisi dari produksi tunas vegetatif ke pembentukan bunga. Di samping menentukan pembentukan bunga, aktivitas bersama antara LFY, AP1 dan CAL dapat menghambat ekspresi gen penanda meristem tunas vegetatif TERMINAL FLOWER1 (TFL1) pada meristem lateral, sehingga mencegah pembentukan tunas vegetatif. Dalam membentuk meristem bunga, LFY dapat mengaktifkan AP1 secara langsung, sedangkan LFY, AP1 dan CAL masing-masing diregulasi secara tidak langsung melalui regulasi negatif oleh TFL1 (Liljegren et al. 1999). Pada tanaman tingkat tinggi, perubahan fase perkembangan diregulasi melalui alur gen yang kompleks. Hilangnya fungsi gen akibat mutasi pada gen EMBRYONIC FLOWER (EMF1 dan EMF2) mengakibat-kan Arabidopsis langsung berbunga, menghindari pertumbuhan tunas vegetatif. Fenotip ini mendukung pendapat bahwa gen EMF berperan utama dalam menekan program reproduktif (Yoshida et al. 2001; Moon et al. 2003).
Ekspresi AP1 pada meristem lateral salah satunya adalah diregulasi oleh LFY. LFY mempercepat ekspresi AP1 pada meristem lateral wild-type di bawah kondisi yang menginduksi pembungaan, sehingga pada mutan lfy ekspresi AP1 menjadi tertunda. Hal ini menunjukkan bahwa LFY merupakan regulator positif bagi aktivitas AP1. Namun pada gilirannya, AP1 juga dapat meregulasi LFY secara positif, karena pada tanaman yang mengekspresikan AP1 secara konstitutif, LFY diekspresikan lebih awal pada meristem bunga yang telah mengalami perubahan tersebut (Liljegren et al. 1999). Mutasi LFY tidak mempunyai pengaruh yang signifikan terhadap fenotip yang diberikan oleh
tanaman 35S::AP1, dan konversi tunas ke bunga pada tanaman 35S::LFY terutama ditekan oleh mutasi AP1 (Mandel dan Yanofsky 1995; Weigel dan Nilsson 1995). Gen AP1 telah berhasil diklon dan memperlihatkan bahwa gen tersebut mengkode putative faktor transkripsi yang mengandung MADS-domain (DNA-binding domain). RNA AP1 diekspresikan secara seragam pada primordia bunga muda, dan selanjutnya terlokalisir pada sepal dan petal. Hal ini menunjukkan bahwa AP1 bertindak secara lokal untuk pembentukan penanda meristem bunga dan untuk menentukan perkembangan sepal dan petal (Mandel et al. 1992).
Fakta bahwa mutasi LFY hanya berpengaruh kecil terhadap pembungaan dan transformasi dari tunas ke bunga yang lebih cepat pada tanaman yang mengekspresikan AP1 secara konstitutif, menunjukkan bahwa AP1 bertindak downstream dari LFY dalam membentuk penanda meristem bunga (Liljegren et al. 1999). Namun demikian, berbagai fakta membuktikan bahwa aktivitas gabungan dari LFY dan AP1 adalah lebih efektif daripada aktivitasnya secara sendiri-sendiri. Pada kondisi yang menginduksi pembungaan, tanaman yang membawa mutasi LFY dan AP1 menunjukkan transformasi yang hampir lengkap dari semua bunga menjadi cabang-cabang tunas aksilar (Huala dan Sussex 1992; Weigel et al. 1992). Analisis genetik terhadap tanaman yang mengekspresikan LFY secara konstitutif menunjukkan bahwa tunas lateral akan menjadi penanda bunga ketika LFY diekspresikan secara konstitutif, dan transformasi ini akan kembali lagi jika tidak ada aktivitas AP1 (Weigel dan Nilsson 1995). Lebih lanjut dijelaskan bahwa meskipun aktivitas AP1 konstitutif tersebut cukup untuk merubah tunas lateral menjadi bunga, namun bunga yang dihasilkan memperlihatkan beberapa karakteristik seperti tunas jika aktivitas LFY tidak ada.
Gen yang berhubungan dekat dengan AP1, yaitu CAULIFLOWER (CAL) juga mempunyai fungsi yang terlibat dalam penentuan penanda meristem bunga (Rounsley et al. 1995). Fenotip mutan ap1 juga dapat ditingkatkan lebih lanjut oleh mutasi CAL yaitu dengan tidak terjadinya konversi secara lengkap dari meristem vegetatif menjadi meristem bunga pada double mutans ap1 cal. Karena meristem bunga tidak dihasilkan oleh tunas utama pada tanaman ap1 cal di dalam kondisi tumbuh normal, maka tunas tersebut tidak pernah menghasilkan transisi secara lengkap dari fase vegetatif ke fase reproduktif (Bowman et al. 1993).
Studi gain-of-function dari tanaman transgenik yang mengekspresikan LFY, AP1 atau CAL secara konstitutif di bawah kendali promoter Cauliflower Mosaic Virus 35S (CaMV 35S) memperkuat kesimpulan hasil studi berdasarkan loss-of-function dan menunjukkan bahwa aktivitas AP1 adalah downstream dan diregulasi oleh LFY. Tanaman yang mengekspresikan LFY, AP1 atau CAL secara konstitutif berbunga lebih cepat dan mengalami transformasi dari meristem tunas primer dan sekunder menjadi meristem bunga, meskipun fenotip yang dihasilkan oleh 35S::CAL lebih lemah daripada 35S::LFY atau 35S::AP1 (Mandel dan Yanofsky 1995; Weigel dan Nilsson 1995). Jadi sesuai dengan pendapat Liljegren et al. (1999), bahwa LFY, AP1 dan CAL bertindak bersama-sama untuk memacu transisi dari fase produksi tunas dan daun (fase vegetatif) ke pembentukan meristem bunga (fase reproduktif).
Ekspresi AP1 dibatasi secara spatial untuk meristem bunga oleh aksi gen penanda meristem vegetatif TERMINAL FLOWER1 (TFL1) (Bowman et al. 1993). Pada tanaman wild-type, AP1 dan TFL1 diekspresikan pada tempat yang berbeda, dimana TFL1 diekspresikan pada daerah subapikal meristem tunas vegetatif, sedangkan ekspresi AP1 terbatas untuk perkembangan bunga (Mandel et al. 1992). Di samping berperan dalam meregulasi perubahan fase
pertumbuhan, TFL1 juga mempunyai pengaruh antagonis terhadap AP1 dalam pembentukan penanda meristem, karena TFL1 memacu penanda meristem vegetatif dan AP1 menentukan meristem bunga. Fenotip tanaman yang mengekspresikan AP1 secara konstitutif mencerminkan fenotip tanaman yang mengalami mutasi TFL1. Tanaman 35S::AP1 dan mutan tfl1 memperpendek fase pertumbuhan vegetatifnya dan mengalami transformasi dari tunas vegetatif menjadi bunga (Mandel dan Yanofsky 1995). TFL1 diekspresikan pada level yang rendah dalam meristem vegetatif dan tampaknya berperan untuk mencegah pembungaan sebelum waktunya. Pada stadia lebih lanjut, TFL1 mengatur dan berperan dalam menekan ekspresi gen penanda meristem bunga seperti LFY dan AP1 pada meristem vegetatif (Jack 2004).
Selain sebagai gen penanda meristem bunga, AP1 juga bertindak sebagai gen kelas A yang diperlukan untuk membentuk whorl pertama dan kedua dari penanda organ bunga (Bowman et al. 1993). Sesuai dengan kedua peranannya dalam menentukan penanda meristem dan penanda organ, AP1 pada awalnya diekspresikan pada meristem bunga muda dan kemudian menjadi terbatas pada daerah dimana akan dibentuk primordia sepal dan petal (Mandel et al. 1992; Gustafson-Brown et al. 1994). Karena gen AP1 berperan dalam mengontrol pembentukan primordia bunga dan organ bunga, maka mutasi gen AP1 dapat mengganggu kedua fase perkembangan bunga tersebut. Pengaruh ini ditunjukkan dengan terjadinya konversi sebagian bunga menjadi infloresen tunas dan gangguan terhadap perkembangan sepal dan petal. Tanaman mutan ap1 akan mengalami kegagalan pembentukan sepal dan petal pada whorl pertama dan whorl ke dua dari bunga, sedangkan pengaruhnya terhadap pembentukan stamen dan carpel pada whorl ke tiga dan ke empat dari bunga adalah kecil (Bowman et al. 1993).
Pada tanaman tomat transgenik, pengaruh utama dari ekspresi AP1 adalah mempercepat saat pembungaan. Tanaman transgenik rata-rata berbunga setelah memproduksi enam daun, sedangkan pada tanaman kontrol (wild-type) setelah memproduksi 11 daun baru berbunga. Tanaman tomat yang meng-ekspresikan AP1 juga mengalami konversi lebih awal dari meristem vegetatif menjadi bunga. Setelah 18 minggu pada kondisi greenhouse, tanaman transgenik menghasilkan konversi bunga dari meristem vegetatif dua kali lipat lebih tinggi dibandingkan tanaman kontrol (Ellul et al. 2004). Selain tidak mempengaruhi fertilitas bunga, tanaman tomat transgenik yang mengekspresi-kan AP1 juga tidak mengakibatmengekspresi-kan abnormalitas bentuk dan ukuran buah yang dihasilkan. Ekspresi konstitutif AP1 pada tanaman tomat transgenik tersebut dapat memperpendek fase vegetatifnya secara signifikan, tanpa menyebabkan abnormalitas infloresen, perkembangan bunga dan produksi buah. Hasil ini sesuai dengan ekspresi AP1 yang telah diteliti pada jeruk (Pena et al. 2001).
Pada tanaman jeruk, yang mempunyai fase juvenil yang panjang dengan menunda perkembangan reproduktifnya antara 6-20 tahun, juga dapat dipercepat pembungaannya dengan mentransformasikan gen AP1 atau LFY. Bibit tanaman jeruk yang mengekspresikan secara konstitutif gen AP1 atau LFY dari Arabidopsis, dapat memproduksi bunga dan buah pada tahun pertama, sehingga dapat memperpendek fase juvenilnya. Selanjutnya dapat dijelaskan bahwa ekspresi AP1 adalah sama efisiennya dengan LFY di dalam menginisiasi pembungaan yang lebih awal, dan tidak menghasilkan perkembangan yang abnormal. Kedua tipe tanaman jeruk transgenik tersebut bunganya normal dan fertil, serta berbunga dalam tahun-tahun berikutnya (Pena et al. 2001).
Gen penanda meristem bunga seperti AP1 dan LFY memang sangat diperlukan untuk memacu pembungaan (Mandel dan Yanofsky 1995; Weigel dan Nilsson 1995). Ekspresi AP1 atau LFY pada meristem tunas apikal
Arabidopsis menyebabkan produksi bunga terminal sebagaimana fenotip yang diakibatkan oleh mutan pada gen TFL1 (Bradley et al. 1997). Ekspresi konstitutif dari AP1 atau LFY pada herbaceous maupun spesies berkayu tersebut telah terbukti dapat mempercepat inisiasi pembungaan melalui penekanan yang kuat terhadap fase juvenil (Weigel dan Nilsson 1995; Pena et al. 2001). Hasil ini telah membuka kemungkinan baru untuk program domestikasi, perbaikan genetik, dan penelitian lebih lanjut, terutama pada pohon buah-buahan dan spesies berkayu lainnya.
Berdasarkan pada pernyataan di atas, pengaturan ekspresi gen oleh perkembangan, lingkungan dan signal metabolit, adalah sangat mungkin terjadi pula dalam pengaturan inisiasi pembungaan pada tanaman kakao dengan mengikuti jalur yang sama, dimana hal tersebut ditandai oleh adanya signal penginduksi, protein pengatur transduksi signal, dan interaksi gen-gen yang terlibat dalam pengaturan pembungaan.