BAB II PEMBAHASAN A. DUPLIKASI GEN
Makna evolusi duplikasi gen pertama kali dicetuskan oleh Haldane (1932) dan Muller (1935), yang menyatakan bahwa duplikasi gen yang berlebihan dari gen dapat menyebabkan adanya mutasi yang berbeda dan pada akhirnya akan muncul sebagai gen baru. Duplikasi gen pertama kali di amati oleh Bridges (1936) pada Bar lokus di Drosophila, namun sebagian contoh kecil duplikasi gen ditemukan sebelum munculnya biokimia dan teknik biomolekuler. Pengembangan metode sekuensing protein pada tahun 1950an menjadi alat pertama untuk pembelajaran proses evolusi dalam jangka panjang, dan di akhir tahun 1950an rantai α dan β hemoglobin diketahui memiliki turunan dari duplikasi gen (Itano 1957; Rhinesmith et al., 1958;. Braunitzer et al., 1961).
Lebih lanjut, isozyme dan penelitian scytologi memberikan bukti seringnya terjadi duplikasi gen selama evolusi. Berdasarkan bukti berbagai jenis studi, Ohno (1970) mengemukakan bahwa duplikasi gen merupakan satu-satunya sarana yang dapat memunculkan gen baru. Selanjutnya, Gilbert (1978) mengemukakan bahwa rekombinasi dalam intron menyediakan mekanisme untuk pertukaran urutan ekson antar gen. Banyak contoh pertukaran ekson telah ditemukan, hal ini menunjukkan bahwa mekanisme ini telah memainkan peran penting dalam evolusi gen eukariot dengan fungsi yang baru.
Tipe Duplikasi Gen
Peningkatan jumlah salinan dari segmen DNA dapat dibawa oleh beberapa jenis duplikasi gen. Duplikasi gen umumnya diklasifikasikan berdasarkan luasnya wilayah genom yang terlibat. Berikut ini jenis duplikasi yang telah diketahui: 1) sebagian atau parsial atau duplikasi gen internal, 2) duplikasi gen lengkap, 3) duplikasi kromosom sebagian, 4) duplikasi kromosom lengkap, 5) poliploidi atau duplikasi genom. Empat duplikasi pertama disebut sebagai duplikasi regional, karena tidak mempengaruhi seluruh set kromosom haploid. Ohno (1970)
berpendapat bahwa duplikasi genom umumnya lebih penting dibandingakan dengan duplikasi regional, karena hanya bagian dari sistem regulasi gen struktural yang dapat di duplikasi, dan ketidakseimbangan tersebut dapat mengganggu fungsi normal duplikasi gen. Namun, duplikasi regional memainkan peran penting dalam evolusi.
Mekanisme molekuler utama yang bertanggung jawab untuk duplikasi gen yaitu unequal crossing over. Antara Unequal crossing over dengan urutan sejajar menimbulkan daerah duplikasi tandem pada satu kromosom, dan penghapusan komplemen pada sisi lain, yang panjangnya tergantung pada ukuran urutan sejajar tersebut. Unequal crossing over difasilitasi oleh kehadiran pengulangan urutan duplikasi. setelah urutan DNA mengalami duplikasi di tandem, proses duplikasi gen dapat dilanjutkan atau proses Cascading semakin dipercepat karena kesempatan unequal crossing over ditambah dengan jumlah salinan duplikasi.
Duplikasi DNA telah lama diketahui sebagai faktor penting dalam evolusi pada tingkat genom. Secara khusus, duplikasi seluruh genom (atau sebagian besar dari genom, seperti kromosom) dapat mengakibtakan subtansi genom tiba-tiba bertambah besar. Peristiwa duplikasi genom telah diketahui berulang kali selama evolusi pada berbagai kelompok organisme.
Domains dan Exons
Domain adalah wilayah dengan protein yang memiliki fungsi tertentu, seperti mengikat substrat, atau bagian yang stabil, tempat pelipatan atau pelekukan, unit struktural kompak dalam protein yang dapat dibedakan dari semua bagian. Bagian awal disebut sebagai domain fungsional, dan bagian yang terakhir disebut sebagai domain struktural atau modul (G5 dan Nosaka 1987). Batas-batas domain seringkali sulit dikenali karena banyak fungsi residu asam amino yang tersebar diseluruh polipeptida. Struktural modul, pada bagian lain adalah co-linear dengan urutan asam amino dari protein (yaitu modul yang terdiri dari urutan asam amino).
Perbedaan diatas penting ketika mempertimbangakan mekanisme evolusi yang tepat oleh multidominan protein yang ada di dalamnya. Jika domain
fungsional bertepatan dengan modul, efek duplikasi akan meningkatkan segmen fungsional. Sebaliknya, jika domain fungsional yang diberikan oleh residu asam amino tersebar di antara modul yang berbeda, efek duplikasi dari modul tunggal mungkin tidak akan fungsional. Pengulangan internal ditemukan pada banyak protein seringkali sesuai dengan modul struktural atau modul tunggal domain fungsional (Barker et al., 1978).
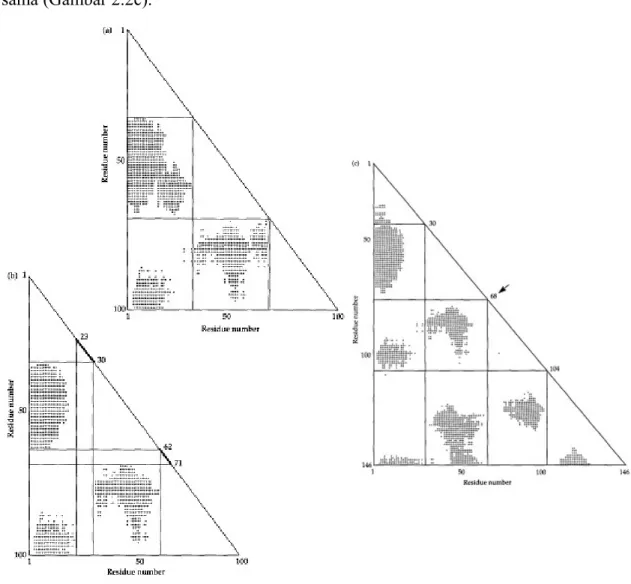
Identifikasi protein pada modul biasanya dilakukan dengan memakai metode grafis yang disebut plot Gŏ (Gŏ 1981). Pada metode ini, residu asam amino pada protein yang tercantum berturut-turut pada dua sumbu dari matriks dua dimensi. Mengingat struktur tersier dari protein, sebuah tanda plus (+) yang dimasukkan dalam matriks jika jarak antara dua residu yang sesuai lebih besar dari nilai yang sudah ditentukan (gambar 1.1). Untuk protein globular, nilai yang digunakan biasanya dalam radius lingkup yang mengandung protein globular, dilambangkan sebagai R. Dalam kasus normal, domain diidentifikasi dengan jelas sebagai daerah yang kosong, non overlapping, segitiga siku-sudut yang hypotenuses berada di diagonal plot Gŏ, dan sisi yang ditentukan oleh empat persegi panjang yang berbeda mengandung gugus tanda plus (Gambar 2.1a). Pada kasus yang tidak umum, kemungkinan terdapat tumpang tindih, di antara segitiga kosong yang berdekatan (Gambar 2.1b). Pada kehidupan nyata, situasi mungkin jauh lebih kompleks, dan metode statistik kompleks digunakan untuk mengidentifikasi lokasi batas yang paling mungkin diantara modul. Gambar 2.1c menunjukkan plot Gŏ untuk β subunit hemoglobin manusia.
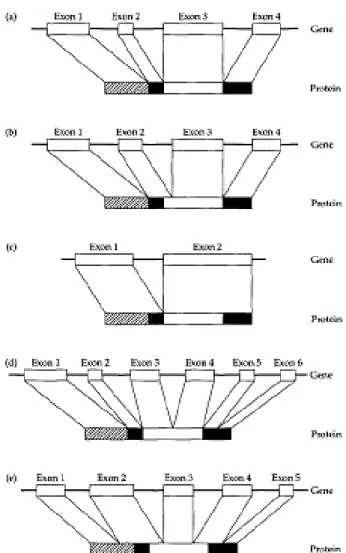
Secara teoritis, beberapa kemungkinan hubungan dapat diketahui antara domain struktural dan pengaturan ekson pada gen (Gambar 2.2). Gŏ (1981) menemukan bahwa protein globular untuk devisi modul internal telah ditentukan, lebih atau kurang ada korespondensi yang tepat antara ekson gen dan domain struktural pada protein produk (Gambar 2.2a dan b). Pada beberapa kasus, satu modul ditemukan, dikode oleh lebih dari satu ekson (Gambar 2.2d). Suatu kejanggalan lengkap antara struktur modul dari protein dan pembagian gen ke dalam ekson (Gambar 2.2e) tidak terdapat dalam penelitiannya. Dalam sejumlah
besar kasus, beberapa modul yang berdekatan ditemukan dikode oleh ekson yang sama (Gambar 2.2c).
Gambar 2.1. Plot Gŏ
(Sumber: Graur, 2000).
Gambar 2.1 diatas menunjukkan plot Gŏ yang dapat digunakan untuk mengidentifikasi protein pada modul. Sebuah tanda plus dimasukkan dalam matriks jika jarak antara dua residu asam amino yang sesuai lebih besar dari jari-jari protein. (A) Sebuah kasus umum, di mana modul dapat diidentifikasi dengan jelas, tidak mungkin untuk menarik garis horizontal atau vertikal lainnya tanpa memotong melalui blok tanda plus. (B) Sebuah kasus yang kurang umum, di mana beberapa alternatif horisontal dan garis vertikal dapat ditarik, dan batas-batas antara modul yang berdekatan diidentifikasi sebagai rentang (garis tebal).
(C) Sebuah plot Gŏ sebenarnya untuk rantai β hemoglobin dari manusia. Radius protein adalah 27Å. Posisi prediksi dari intron ditampilkan pada sisi miring. Sebuah rantai gen manusia hanya terdiri dari tiga ekson. Gŏ (1981) mendalilkan bahwa peleburan terjadi antara dua ekson sebagai hasil hilangnya intron pusat. Panah menandai posisi yang diprediksikan dari intron pusat. Prediksi ini ditanggung oleh temuan intron pada gen leghemoglobin serta di beberapa gen globin invertebrata. Dimodifikasi dari Gŏ (1981).
Gambar 2.2. Lima kemungkinan hubungan antara susunan ekson dalam gen dan domain structural proteinnya (Sumber: Graur, 2000).
Gambar 2.2 di atas menunjuukan bahwa susunan ekson dalam gen dan domain structural proteinnya ada lima kemungkinan. Untuk yang pertama, Setiap ekson sesuai persis dengan domain struktural (a); kedua, korespondensi ini hanya perkiraan (letak exon tidak secara tepat pada satu protein namun secara perkiraan)
(b); ketiga, Sebuah ekson mengkodekan dua atau lebih domain (c); keempat, Sebuah domain struktural tunggal dikodekan oleh dua atau lebih ekson (d); dan kelima, Kurangnya korespondensi antara ekson dan domain. struktural domain dari protein yang ditunjuk oleh kotak yang berbeda (arsiran, hitam, putih, dan abu-abu) (e).
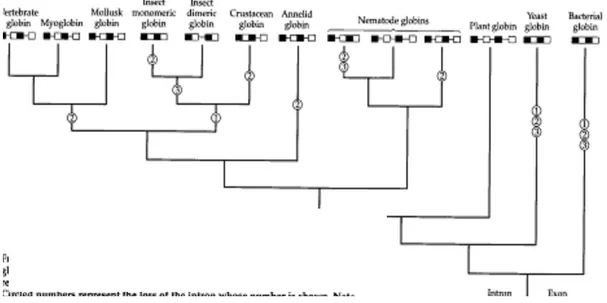
Pada hemoglobin vertebrata rantai α dan β, misalnya terdiri dari empat domain, sedangkan gen mereka hanya terdiri dari tiga ekson, yang mengkode kedua domain yang berdekatan. Gŏ (1981) mengendalikan penggabungan yang terjadi antara dua ekson sebagai akibat hilangnya intron pusat. Gen globin homolog pada tumbuhan (leghemoglobins) ditemukan mengandung intron tambahan atau sangat dekat pada posisi yang diprediksi oleh struktur domain dari globins vertebrata (Landsman et al., 1986). Intron serupa ditemukan pada gen dari nematoda Pseudoterranova decipiens (Dixon et al., 1991). Menariknya, gen globin-encoding dari nematoda Caenorhabditis elegans ditemukan mengandung intron tunggal yang sesuai di posisi intron pusat di leghemoglobins (Kloek et al., 1993). Dengan demikian, selama evolusi keluarga gen globin dari empat ekson gen leluhur, beberapa garis keturunan kehilangan beberapa atau semua tiga intron mereka, sehingga menghasilkan mutasi ekson-intron (Gambar 2.3).
Gambar 2.3. Hilangnya intron selama evolusi gen globin. Leluhur gen globin leluhur memiliki empat ekson dan tiga intron. Beberapa globins (misalnya, pada tanaman) ditahan pada struktur leluhur, sementara garis keturunan lainnya kehilangan satu atau lebih intron. Nomor yang dilingkari mewakili hilangnya intron. Perhatikan bahwa intron tidak digambar dengan skala.
Skema ini didasarkan pada pohon filogenetik tentatif oleh Goodman et al., (1988).
Dalam sebagian besar kasus, duplikasi domain di tingkat protein menunjukkan bahwa duplikasi ekson telah terjadi di tingkat DNA, oleh karena duplikasi ekson adalah salah satu jenis paling penting dari duplikasi gen internal. Gen eukariotik umumnya terdiri dari banyak ekson dan intron, dan ekson antara tetangga sering identik atau sangat mirip satu sama lain. Selain itu, banyak protein dari organisme masa kini menunjukkan pengulangam internal sekuens asam amino, dan sering mengulangi domain fungsional atau struktural dalam protein (Barker et al., 1978). Hal ini menunjukkan bahwa banyak protein yang dibentuk oleh duplikasi gen internal dan fungsi dari protein ini ditingkatkan dengan meningkatkan stabilitas mereka atau jumlah situs aktif. Duplikasi internal juga dapat memberikan segmen DNA berlebihan yang dapat memungkinkan gen untuk mengembangkan fungsi baru. Banyak gen yang kompleks dalam organisme masa kini mungkin telah berevolusi dari gen primordial yang sederhana dan kecil melalui duplikasi internal dan modifikasi berikutnya (Li 1983).
Duplikasi Domain dan ELongasi Gen
Hasil survei gen eukariotik saat ini menunjukkan bahwa duplikasi internal sering terjadi dalam evolusi. Peningkatan ukuran gen, atau gen mengalami elongasi, adalah salah satu langkah yang paling penting dalam evolusi gen kompleks dari yang sederhana. Secara teoritis, pemanjangan gen juga dapat terjadi dengan cara lain. Misalnya, perubahan mutasi mengubah kodon stop ke kodon sense juga bisa memanjangkan gen. Demikian pula, baik penyisipan dari segmen DNA asing ke dalam ekson atau terjadinya mutasi melenyapkan situs splicing akan mencapai hasil yang sama. Jenis perubahan molekul kemungkinan besar akan mengganggu fungsi pemanjangan gen, karena akan menambahkan random array asam amino pada daerah tersebut. Dalam sebagian besar kasus, seperti perubahan molekul telah ditemukan terkait dengan manifestasi patologis. Misalnya, kelainan α hemoglobin Constan spring dan Icaria mengakibatkan mutasi yang mengubah kodon stop ke kodon untuk glutamin dan lisin, sehingga
terdapat 30 residu tambahan untuk rantai α pada varian ini (Weatherall dan Clegg 1979).
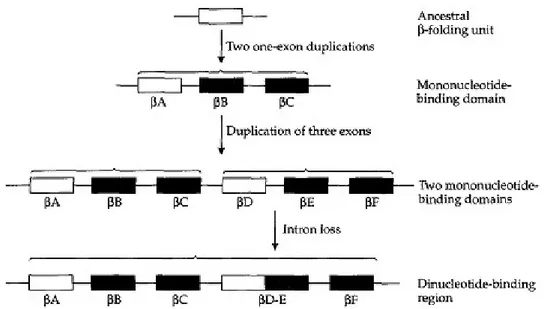
Sebaliknya, duplikasi dari struktur domain cenderung kurang bermasalah. Duplikasi tersebut dapat beberapa kali bahkan meningkatkan fungsi dari protein yang dihasilkan, misalnya dengan meningkatkan jumlah situs aktif (perubahan kuantitatif), sehingga memungkinkan gen untuk menjalankan fungsinya lebih cepat dan efisien, atau dengan memiliki efek sinergis yang menghasilkan fungsi baru (perubahan kualitatif). Salah satu contohnya adalah daerah dinukleotida mengikat dehidrogenase gliseraldehida-3-fosfat (GAPDH) dan alkohol dehidrogenase (ADH). Pada banyak spesies, wilayah-wilayah yang mengikat terdiri dari dua domain (Gambar 2.4). Masing-masing domain terdiri dari tiga homolog β-folding unit, dilambangkan βA, βB, dan βC dalam domain N-terminal dan βD, βE, dan βF dalam domain C-terminal. Seluruh wilayah pengikatan dinukleotida dikodekan oleh lima ekson, tiga untuk domain N-terminal dan dua untuk domain C-terminal. Setiap koenzim hanya mengikat domain mononukleotida. Seluruh wilayah diduplikasi, sehingga tidak hanya dapat mengikat dua mononucleotida, tetapi juga di nukleotida.
Gambar 2.4. Skenario hipotesis untuk evolusi pada pengikatan wilayah dinukleotida (Sumber: Graur, 2000).
Gambar 2.4 di atas menunjukkan gambaran hipotesis untuk menceritakan terjadinya evolusi pada pengikatan wilayah dinukleotida. Ekson ditampilkan sebagai kotak yang dihubungkan oleh intron, yang ditampilkan dalam bentuk garis. Leluhur β-folding Unit dikodekan oleh ekson mengalami dua peristiwa duplikasi untuk menghasilkana triexonic mononukleotida-binding domain. Duplikasi dari tiga ekson mengakibatkan terciptanya dua domain mononukleotida-binding. Modifikasi berikutnya dari urutan utama memunculkan daerah pengikatan dinucleotide . Susunan pentaexonic ini disebabkan oleh hilangnya intron.
Kemungkinan kedua, munculnya fungsi baru sebagai duplikasi gen parsial untuk salinan internal, hal demikian menyimpang dari urutan, akhirnya masing-masing melakukan fungsi yang berbeda. Misalnya, daerah variabel dan konstan gen immunoglobulin yang kemungkinan berasal dari sebuah domain primordial umum, tetapi memiliki sifat yang berbeda (Leder 1982). Dengan demikian, walaupun berasal dari nenek moyang molekul yang umum, wilayah variabel imunoglobulin mengikat antigen, sedangkan wilayah konstan menengahi fungsi non-antigenik. Banyak gen yang kompleks mungkin telah muncul dengan cara ini. Bahkan jika duplikasi internal yang tidak melibatkan situs aktif, mungkin masih menguntungkan. Misalnya, peristiwa duplikasi melibatkan domain struktural terlibat baik dalam pemberian stabilitas spasial untuk protein atau melindungi bagian aktif yang secara tidak langsung dapat mengubah fungsi protein atau mengakibatkan umur panjang, Sebagai contoh, yang disebut PEST polipeptida-protein kaya prolin (P), asam glutamat (E), serin (S), dan treonin (T), mereka ditemukan terdegradasi dengan cepat dalam sel eukariotik sel (Rogers et al.,. 1986).
Dalam beberapa kasus, duplikasi domain PEST ditemukan dalam protein, sehingga menyebabkan degradasi yang sangat cepat, yang penting dalam protein, seperti pada faktor nuklir, yang memiliki fungsi tertentu (Chevaillier 1993). Hal ini juga telah menyarankan bahwa, selama mereka tidak mengganggu fungsi normal, duplikasi domain yang berlebihan dapat dipertahankan tanpa batas dalam genom, dan mungkin berfungsi sebagai bahan baku untuk membuat fungsi baru.
Bagian berikut menyajikan beberapa contoh duplikasi gen internal yang untuk menggambarkan konsekuensi dari perpanjangan gen selama evolusi (Graur, 2000).
Gen Ovomucoid
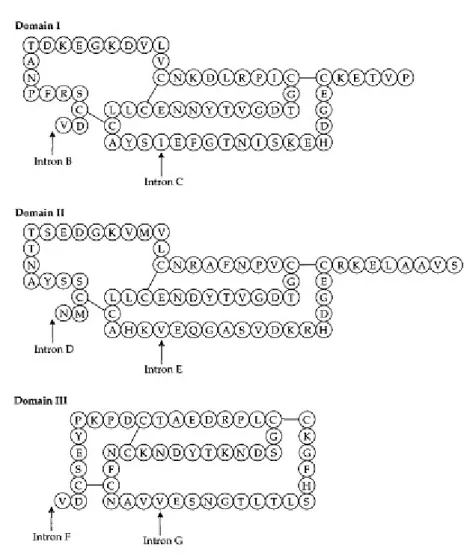
Ovomucoid adalah inhibitor tripsin, enzim yang mengkatalisis pencernaan protein. Terdapat dalam albumen (putih telur) burung. Polipeptida ovomucoid dapat dibagi menjadi tiga domain fungsional (Gambar 2.5).
Gambar 2.5 Tiga domain fungsional dari ovomucoid dari ayam dan tingkat kesamaan pada urutan antara domain asam amino dan tingkat nukleotida. Intron B-G yang ditunjukkan oleh panah. Intron A mengganggu 5' non coding region dan tidak terlihat. Data dari Stein et al., (1980) dan O'Malley et al., (1982). Setiap domain mampu mengikat satu molekul baik tripsin atau serin proteinase lain. Daerah DNA coding untuk tiga domain fungsional jelas berbagi
dari asal mula yang sama dan terpisah satu sama lain oleh intron (Stein et al., 1980). Domain I dan II, I dan III, dan II dan III pada asam amino terlihat pada 46, 33, dan 30%, dan masing-masing terdapat pada 66, 42, dan 50% pada urutan nukleotida. Masing-masing terdiri dari tiga wilayah, dua ekson dipisahkan oleh intron dan dua ekson menunjukkan ada kesamaan di antara mereka. Dengan demikian, gen ovomucoid tampaknya berasal dari satu gen domain primordial yang disebabkan dua duplikasi internal yang masing-masing melibatkan dua ekson tetangga. Karena domain I dan II lebih mirip satu sama lain daripada baik domain III, mereka mungkin berasal dari duplikasi kedua, sementara domain III adalah produk dari duplikasi pertama.
Peningkatan Fungsi Alel dalam Alel α2 pada Haptoglobin
Contoh terkenal dari peningkatan fungsi konsekuensi dari duplikasi gen internal adalah alel haptoglobin α2 pada manusia (Smithies et al., 1962). Haptoglobin adalah protein tetrameric yang terbuat dari dua rantai α dan dua rantai β. Kedua rantai diproduksi oleh gen yang sama sebagai single polipeptida, yang kemudian dibelah pada residu arginin untuk menghasilkan sununit α dan β. Haptoglobin ditemukan dalam serum darah, berfungsi mengangkut glikoprotein untuk menghilangkan hemoglobin bebas dari sirkulasi vertebrata. Pada manusia, haptoglobin α adalah polimorfik karena keberadaan tiga alel umum: slow α1 (α1S), fast α1 (α1F), dan α2.
Alel α2 kemungkinan dibuat oleh nonhomologous crossing over berbeda dengan intron dari dua alel α2 dalam membawa kedua individu heterozigot slow α1 (α1S), fast α1 (α1F) pada varian elektroforesis. Duplikasi internal sekitar 1,7 kb, dan 177 bp pada bagian exonic, hampir dua kali lipat panjang polipeptida (84-143 asam amino). Sebagai konsekuensi, stabilitas kompleks haptoglobin-hemoglobin dan efisiensi rendering kelompok heme dari haptoglobin-hemoglobin rentan terhadap degradasi yang meningkat pesat (Hitam dan Dixon 1968).
Alel α2 kemungkin berasal dari baru-baru ini, setidaknya lebih baru daripada perpecahan antara manusia-simpanse, tetapi memiliki frekuensi yang cukup tinggi (30-70%) di Eropa dan di bagian Asia (Mourant et al., 1976). Jika
memang individu yang membawa alel α2 memiliki keuntungan khusus karena membawa alel α2, ada kemungkinan bahwa di masa depan alel α2 akan menjadi tetap dalam populasi manusia dengan mengorbankan varian α1. Menariknya, bahkan lebih lama alel, α3 (atau haptoglobin Johnson), adalah ditemukan pada populasi manusia. Alel ini berisi pengulangan tandem tiga kali lipat dari segmen 1.7-kb yang sama terlibat dalam duplikasi alel α2 (Oliviero et al., 1985).
Asal Gen Antibeku Glikoprotein
Cairan tubuh yang paling teleosts (ikan pari-bersirip) membeku pada suhu berkisar dari -1.0° C hingga -0,7 ° C. Oleh karena itu, sebagian besar ikan tidak dapat bertahan hidup pada suhu pembekuan (-1,9 ° C) di Samudera Antartika. Kekuatan pembekuan ikan di Antartika ini disebabkan adanya protein di dalam darah yang menurunkan suhu pembekuan dengan menyerap kristal es kecil dan menghambat pertumbuhan mereka, yang dikenal dengan pecahnya membran sel. Ada beberapa protein seperti, protein antibeku I, II, dan III, dan antibeku glikoprotein. Kasus gen antibeku glikoprotein pada toothfish Antartika (Dissostichus mawsoni) berasal dari duplikasi gen internal.
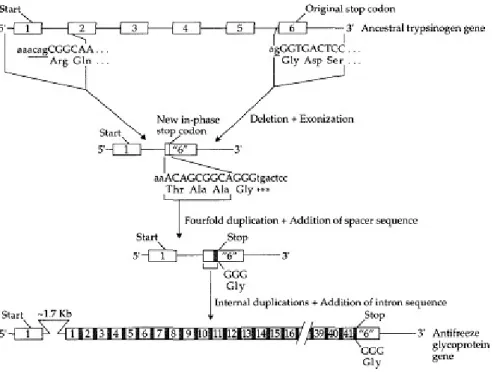
Ada banyak glikoprotein antibeku yang berbeda ikan, masing-masing sebagian besar terdiri dari dua pengulangan tripeptide sederhana: Thr-Ala-Ala dan Thr-Pro-Ala. (Perhatikan bahwa keluarga kodon prolin berbeda dari alanin yang merupakan keluarga kodon nukleotida tunggal.) Antibeku glikoprotein dikodekan oleh keluarga gen besar, di mana masing-masing gen mengkode prekursor poliprotein yang dipecah pascatranslasi untuk menghasilkan beberapa molekul antibeku glikoprotein. Chen et al., (1997) menandai satu gen antibeku glikoprotein dari toothfish Antartika dan menemukan bahwa gen itu berasal dari gen encoding tripsinogen pankreas (Gambar 2.6). Sejarah evolusi gen antibeku glikoprotein gen dapat direkonstruksi secara akurat, terutama karena gen itu disimpulkan telah muncul pada 5-14.000.000 tahun yang lalu, sebuah estimasi dengan perkiraan pembekuan Samudra Antartika (10-14.000.000 tahun yang lalu).
Gambar 2.6. jalur evolusi dari gen leluhur tripsinogen (Sumber: Graur, 2000). Gambar 2.6 di atas menunjukkan sebuah jalur evolusi, kemungkinan dimana suatu gen leluhur tripsinogen dengan enam ekson (kotak bernomor) berubah menjadi sebuah gen antibeku glikoprotein di Antartika toothfish (Dissostichus mawsoni). Berikut penghapusan dan exonization lima nukleotida intronic (digarisbawahi huruf kecil) pada gen tripsinogen leluhur, gen baru dengan dua ekson muncul (Kedua ekson ditandai "6" untuk menekankan keturunan nya, dan kodon stop baru yang dibawa ke dalam bingkai oleh exonization yang ditandai dengan tanda bintang.) Urutan encoding Thr-Ala-Ala digandakan untuk membuat pengulangan empat kali lipat, dan pengaturan urutan pendek diketahui (pada kotak) ditambahkan ke unit berulang. Beberapa duplikasi gen internal yang mengakibatkan pengulangan 41 kali. Penambahan panjang urutan -1,7 kb untuk intron yang diindikasikan sebagai lingkaran segitiga. Penambahan ini bisa terjadi pada salah satu dari langkah-langkah sebelumnya dalam evolusi gen ini, dan penambahan diakhir untuk kenyamanan grafis saja.
Sejarah evolusi gen antibeku glikoprotein dapat diringkas seperti yang ditunjukkan pada Gambar 2.6. Penghapusan awal di leluhur gen tripsinogen, dimulai dari nukleotida 6 di ekson kedua dan diakhiri pada salah satu nukleotida
sebelum dimulainya ekson 6, dan menciptakan gen baru dengan dua ekson. Empat nukleotida dari intron pertama dan satu dari intron kelima menjadi bagian dari ekson yang baru dibuat ini, yang membaca frameshifted singkat pada bingkai yang mengkodekan tetrapeptide (Thr-Ala-Ala-Gly) sebelum mencapai frame baru di kodon stop (TGA). Urutan 12-bp mengkode tiga asam amino pertama (Thr-Ala-Ala) diduplikasi dua kali untuk membuat pengulangan empat kali lipat dari tripeptide ini. Berikut ini putaran awal duplikasi, pengaturan jarak urutan asal tidak diketahui ditambahkan ke ujung 3' dari unit berulang.
Urutan yang dihasilkan kemudian digandakan beberapa kali untuk menghasilkan 41 pengulangan diikuti oleh kodon Gly dari tetrapeptide yang asli dan kodon stop. Pengaturan urutan jarak pengkodean peptida yang berfungsi sebagai sinyal untuk pembelahan antibeku glikoprotein polipeptida menjadi protein aktif. Kadang-kadang sepanjang garis dari leluhur gen tripsinogen hingga gen antibeku glikoprotein berurutan dengan panjang sekitar 1,7 Kb ditambahkan ke intron tersebut. Kemungkinan, penyisipan ini tidak memiliki konsekuensi fungsional. Kami mencatat bahwa fungsi yang baru telah dibuat oleh beberapa peristiwa mutasi (sebagian besar duplikasi gen internal) dalam kurun waktu yang sangat singkat. Dengan demikian, gen baru telah mengalami seleksi positif intens, kemungkinan besar karena pergeseran tiba-tiba dalam kondisi lingkungan (Logsdon dan Doolittle, 1997).
Prevalensi Duplikasi Domain
Perpanjangan gen selama evolusi sebagian besar bergantung pada duplikasi domain. Tabel 2.1 menyajikan daftar gen yang memiliki bukti duplikasi internal selama sejarah evolusi mereka. Semua melibatkan satu atau lebih duplikasi domain, dan beberapa urutan (misalnya, ferredoxin, albumin serum, dan tropomiosin rantai) berasal dari perkalian dari urutan primordial, sehingga menghasilkan struktur berulang yang mengambil seluruh yang panjang protein. Dalam setiap contoh, duplikasi bisa mudah disimpulkan dari kesamaan urutan protein atau DNA. Kemungkinan banyak gen kompleks lainnya yang telah berevolusi dengan duplikasi gen internal, tetapi kemungkinan adanya
penyimpangan pada daerah mereka yang digandakan sehingga urutan homologi antara mereka tidak lagi dapat dilihat. Dalam beberapa kasus, seperti daerah konstan dan variabel gen immunoglobulin, kami dapat menyimpulkan nenek moyang yang sama dengan membandingkan struktur sekunder dari domain, karena struktur sekunder telah dipertahankan lebih baik daripada urutan asam amino (Hood et al., 1975). Dengan demikian, duplikasi protein internal mungkin jauh lebih meluas daripada data empiris yang telah ada.
Pembentukan Famili Gen dan Penambahan Fungsi Baru
Sebuah duplikasi gen lengkap menghasilkan dua salinan identik. Bagaimana mereka akan berevolusi dengan bervariasi dari kasus ke kasus. Pada prinsipnya terdapat tiga kemungkinan. Salinan tersebut dapat mempertahankan fungsi asli mereka, memungkinkan suatu organisme untuk menghasilkan kuantitas yang lebih besar dari RNA atau protein. Atau, salah satu salinan mungkin lumpuh yang disebabkan terjadinya mutasi yang merusak dan menjadi pseudogene yang tidak berfungsi. Bagian yang terpenting adalah kemungkinan yang ketiga, bahwa duplikasi gen dapat mengakibatkan munculnya genetik baru atau gen baru. Hal ini akan terjadi jika salah satu duplikat mempertahankan fungsi aslinya sementara molekul yang lain terakumulasi, dan dapat melakukan tugas yang berbeda.
Pengulangan gen dapat dibagi menjadi dua jenis: varian dan invarian repeats. Pengulangan invarian merupakan hal identik atau hampir identik secara berurutan satu dengan yang lain. Dalam beberapa kasus, pengulangan urutan identik dapat ditunjukkan berkorelasi dengan peningkatan kuantitas sintesis produk gen yang diperlukan untuk fungsi normal dari suatu organisme. Pengulangan tersebut disebut sebagai dose repetitions. Dose repetitions cukup umum ketika terdapat kebutuhan metabolik untuk menghasilkan sejumlah besar dari RNA atau protein tertentu (Ohno 1970). Misalnya, duplikasi lokus asam monophosphatase pada ragi, memungkinkan untuk menghasilkan dua kali jumlah enzim, sehingga pemanfaatan fosfat yang tersedia lebih efisien bila fosfat adalah membatasi faktor pertumbuhan (Hensche 1975). Contoh perwakilan duplikasi gen termasuk gen untuk rRNA dan tRNA, yang dibutuhkan untuk penerjemahan, dan gen histon, yang merupakan komponen protein utama kromosom dan karena itu harus disintesis dalam jumlah besar terutama selama fase S dari siklus sel, ketika DNA direplikasi (Elgin dan Weintraub 1975). Namun, genom eukariota juga diketahui mengandung pengulangan invarian yang tidak memiliki fungsi apapun. Varian repeats adalah salinan dari gen itu, meskipun mirip satu sama lain, namun berbeda dalam urutan pada tingkat lebih rendah atau lebih besar. Menariknya, varian repeats kadang-kadang dapat melakukan fungsi yang sangat berbeda. Sebagai contoh, trombin, yang membelah fibrinogen selama proses pembekuan
darah, dan enzim tripsin pencernaan telah diturunkan dari duplikasi gen lengkap di masa lalu. Demikian pula, lactalbumin, subunit enzim yang mengkatalisis sintesis laktosa gula, dan lisozim, yang melarutkan bakteri tertentu dengan membelah komponen polisakarida dinding sel mereka yang terkait dengan keturunan satu dengan lain. Diferensiasi fungsi biasanya membutuhkan sejumlah besar substitusi. Namun, banyak kasus, fungsi baru mungkin dicapai melalui beberapa pergantian pemain. Misalnya, dehidrogenase laktat dapat diubah menjadi malat dehidrogenase dengan hanya mengganti satu pada 317 asam amino (Wilks et al.,. 1988).
Semua gen termasuk juga kelompok tertentu, urutan berulang dalam genom disebut sebagai gen atau multigene family. Fungsional dan nonfungsional anggota multigene family kemungkinan berada di dekat satu sama lain pada kromosom yang sama, atau mereka mungkin terletak pada kromosom yang berbeda. Anggota multigene familiy yang terletak sendirian di sebuah genom yang berbeda lokasi dari anggota multigene yang lain disebut orphon.
Ketika duplikasi gen menjadi terlalu berbeda dari satu sama lain baik fungsi atau urutan, hal tersebut mungkin tidak lagi bisa untuk menetapkan mereka dalam satu gene family. Superfamili istilah ini diciptakan oleh Dayhoff (1978) digunakan untuk membedakan protein yang terkait erat dari protein yang memiliki keterkaitan jauh. Hal demikian, menunjukkan bahwa protein setidaknya memiliki 50% kesamaan satu sama lain di tingkat asam amino dan dianggap sebagai anggota keluarga, sementara protein homolog menunjukkan kurang dari 50% kesamaan dianggap sebagai anggota dari superfamili. Sebagai contoh, α- dan β-globins diklasifikasikan menjadi dua keluarga terpisah dan bersama-sama dengan mioglobin mereka membentuk superfamili globin. Namun, dua istilah tidak selalu dapat digunakan secara ketat sesuai kriteria Dayhoff. Untuk rantai Misalnya, manusia dan ikan mas α-globin memperlihatkan urutan kesamaan hanya 46%, yang menjadi penanda dari keluarga gen yang sama. Untuk alasan ini, klasifikasi protein menjadi keluarga dan superfamilies tidak hanya ditentukan sesuai dengan urutan kemiripan, tetapi juga dengan mempertimbangkan bukti tambahan yang berkaitan dengan kesamaan fungsional, spesifisitas jaringan, atau jenis homologi.
Sebuah fitur penting terkait duplikasi gen adalah bahwa selama dua atau lebih salinan gen yang berada di dekat satu sama lain, proses duplikasi gen dapat dipercepat di wilayah ini, dan banyak salinan yang dapat dihasilkan. Salah satu hasil praktis duplikasi gen dan modifikasi berikutnya yaitu salinan yang dihasilkan banyak gen yang melakukan fungsi yang berbeda diturunkan dari leluhur gen umum dan dengan demikian homolog satu sama lain. Berdasarkan urutan data DNA, jumlah kesamaan yang tak terduga antara protein yang sebelumnya tidak diketahui berkaitan satu sama lain telah terungkap (Tabel 2.2). Salah satu contoh yaitu tripsin dan kimotripsin. Perbedaan mereka sejak sekitar 1,5 miliar tahun yang lalu, dua enzim pencernaan ini memiliki fungsi yang berbeda: tripsin berfungsi memotong rantai polipeptida di arginin dan residu lisin residu, sedangkan kimotripsin membelah polipeptida rantai di fenilalanin, triptofan, dan residu tirosin (Barker dan Dayhoff, 1980). Jumlah gen dalam keluarga gen bervariasi. Berikut ini, gen rRNA dan tRNA akan digunakan untuk menggambarkan gen invarian yang berulang-ulang. Gen yang berulang dengan tingkatan rendah, diwakili dengan isozim dehidrogenase laktat dan warna sensitif opsins.
RNA-Specifing Genes
Genom mitokondria vertebrata hanya satu salinan dari kedua gen 12S dan gen 16S rRNA. Hal ini tampaknya cukup untuk sistem terjemahan mitokondria karena genom mengandung hanya 13 gen penyandi protein. Mycoplasmas, yang merupakan replikasi diri terkecil dari prokariota, mengandung dua set gen rRNA. Genom Escherichia coli adalah 4-5 kali lebih besar dari Mycoplasma capricolum, dan berisi tujuh set gen rRNA. Jumlah gen rRNA ragi adalah sekitar 140, dan angka-angka pada lalat buah dan manusia bahkan lebih besar. Xenopus laevis memiliki genom yang lebih besar dan lebih dari gen rRNA manusia.
Demikian, korelasi positif antara jumlah gen rRNA dan ukuran genom. Hubungan antara ukuran genom juga berlaku untuk gen tRNA dan gen RNA (Tabel 2.3). Namun demikian, terdapat beberapa pengecualian. Misalnya, genom jagung (Zea mays) ukurannya sama dengan genom manusia, tapi mengandung sekitar 45 kali lebih banyak set gen rRNA. Satu aturan pengecualian yang sangat menarik, hubungan di mana jumlah gen RNA-menentukan (dosis) ukuran genom. Ciliata Tetrahymena memiliki genom yang lebih besar daripada ragi Saccharomyces cerevisiae, tetapi hanya satu set gen rRNA. Set ini, berada di inti germinal, mikronukleus. Turunan macronuclei vegetatif dari mikronukleus, jumlah salinan gen diperkuat 200-600 kali (Yao et al., 1974). Diperkirakan bahwa Tetrahymena memiliki 600 eksemplar ekstrakromosomal gen rRNA dan dekat dengan 1.500 salinan gen tRNA. Dengan demikian, sejumlah besar rRNA dapat diproduksi selama pertumbuhan vegetatif, meskipun terdapat sejumlah pengkopian kecil di set haploid.
Mungkin terdapat dua alasan untuk korelasi umum positif antara ukuran genom dan jumlah salinan gen RNA-specifying. Seberapa besar genom membutuhkan RNA, atau jumlah gen RNA- specifying hanyalah konsekuensi pembesaran genom pasif genom oleh duplikasi. Gen yang berulang-ulang, seperti gen rRNA, umumnya sangat mirip satu lainnya. Salah satu faktor yang bertanggung jawab untuk homogenitas kemungkinan memurnikan seleksi, karena gen ini harus mematuhi spesifik fungsional dan persyaratan struktural. Namun, homogenitas meluas ke daerah tanpa signifikansi fungsional atau struktural, dan
dengan demikian pemeliharaan homogenitas mengharuskan adanya mekanisme lain.
Isozymes
Selain invarian repeats, genom organisme yang lebih tinggi mengandung banyak multigene family yang anggotanya telah menyimpang ke berbagai luasan. Contohnya adalah keluarga gen coding untuk isozim, seperti laktat dehidrogenase, aldolase, creatine kinase, karbonat anhidrase, dan piruvat kinase. Isozim adalah enzim yang mengkatalisis rekasi biokimia yang sama tetapi kemungkinan berbeda satu sama lain dalam spesifisitas jaringan, perkembangan regulasi, mobilitas elektroforesis, atau properti biokimia. Isozim dikodekan oleh lokus yang berbeda, gen biasanya diduplikasi, sebagai lawan untuk isozim yang disebut allozymes, yang memiliki bentuk yang berbeda dari enzim yang sama dikodekan oleh alel yang berbeda pada lokus tunggal. Studi tentang sistem isozim multilokus meningkatkan pemahaman kita tentang bagaimana sel-sel dengan sumbangan genetik identik dapat berdiferensiasi menjadi ratusan jenis
khusus yang berbeda dari sel yang membentuk organisasi tubuh vertebrata yang kompleks. Meskipun semua anggota keluarga isozim melayani dasar fungsi katalitik yang sama, anggota yang berbeda mungkin telah berevolusi untuk jaringan atau tahap perkembangan yang berbeda sehingga meningkatkan fisiologis fine-tuning dari sel.
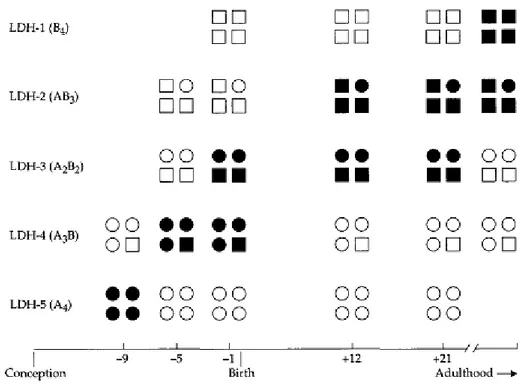
Dua gen yang mengkode subunit A dan B laktat dehidrogenase (LDH) pada mamalia (Hiraoka et al.,. 1990). Kedua subunit berbentuk lima isozim tetrameric, A4,A3B, A2B2, AB3, dan B4, semuanya mengkatalisis baik konversi laktat menjadi piruvat karena teroksidasi koenzim nikotinamida adenin dinukleotida (NAD+) atau reaksi sebaliknya karena reduksi koenzim (NADH). B4 dan isozim lain yang kaya subunit B, yang memiliki afinitas tinggi untuk NAD+, berfungsi sebagai dehidrogenase laktat dalam metabolisme aerob di jaringan seperti jantung, sedangkan A4 dan isozim kaya subunit A, yang memiliki afinitas tinggi untuk NADH, secara khusus diarahkan untuk reduktase piruvat pada metabolisme anaerob di jaringan seperti otot rangka.
Gambar 2.7. Perkembangan produksi LDH di jantung (Sumber: Graur, 2000). Gambar 2.7 menunjukkan urutan perkembangan produksi LDH di jantung. Kita melihat bahwa semakin jantung anaerobik (khusus, pada awal tahap
kehamilan), semakin tinggi proporsi LDH isozim kaya akan subunit A. Dengan demikian, dupliksi dua gen menjadi khusus untuk jaringan berbeda dan tahap perkembangan yang berbeda. Subunit yang hadir hampir di semua vertebrata dipelajari untuk saat ini, duplikasi yang menghasilkan gen untuk LDH-A dan LDH-B mungkin terjadi baik sebelum atau selama tahap awal evolusi vertebrata. Sebuah fitur menarik dari LDH adalah bahwa dua subunit dapat membentuk heteromultimeric, sehingga lebih meningkatkan fisiologis fleksibilitas enzim. Banyak contoh lain dari enzim multimeric yang terdiri dari polipeptida yang dikode oleh duplikasi gen dikenal (Harris 1979, 1980/1981).
Opsins
Penglihatan warna pada manusia, kera, dan monyet Old World dimediasi di mata oleh tiga jenis sel fotoreseptor (kerucut), yang merubah energi fotik menjadi potensial listrik. Setiap jenis warn memiliki tingkat sensitif maksimal dengan panjang gelombang tertentu, tergantung pada jenis pigmen warna-sensitif (Photopigment) yang hadir dalam sel kerucut. Pada manusia, sel kerucut merah, hijau, dan biru memiliki sensitifitas maksimal sekitar 560, 530, dan 430 nanometer. Setiap warna merangsang satu atau lebih jenis kerucut. Sebagai contoh, lampu merah merangsang hanya kerucut merah, biru merangsang kerucut biru, kuning cahaya merangsang kerucut merah dan hijau sama-sama, dan cahaya putih merangsang semua tiga jenis kerucut secara bersamaan (Carlson 1991). Setiap photopigment warna-sensitif terdiri dari dua bagian: protein yang disebut opsin, dan turunan lipid vitamin A1 disebut retina. Spesifisitas warna ditentukan oleh opsins, yang merupakan anggota dari superfamili dari G-protein coupled reseptor. Opsin biru dikodekan oleh gen autosomal, sedangkan opsins merah dan hijau dikodekan oleh gen X-linked. Setiap kromosom X hanya satu gen opsin merah, tapi mungkin berisi lebih dari satu opsin hijau gen (Nathans et al., 1986). Urutan asam amino dari opsin merah dan opsin hijau 96% identik, tetapi hanya 43% asam amino identik dengan opsin biru. Gen opsin biru dan nenek moyang dari opsin hijau dan merah gen menyimpang sekitar 500 juta tahun yang lalu (Yokoyama dan Yokoyama 1989). Sebaliknya, hubungan dekat dan kesamaan
yang tinggi antara gen opsin merah dan hijau menunjuk duplikasi gen baru-baru ini.
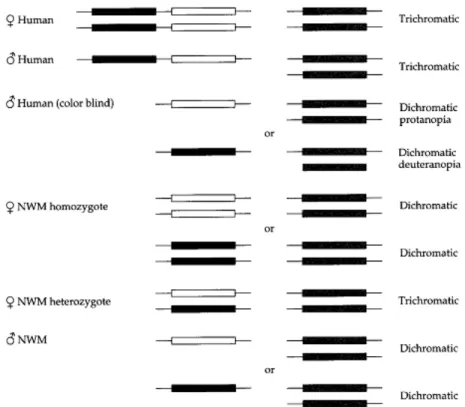
Karena sebagian besar monyet hanya memiliki satu gen pigmen X-linked (lihat di bawah), sedangkan monyet kuno (termasuk kera dan manusia) telah memiliki dua atau lebih, diasumsikan bahwa duplikasi terjadi sekitar 25-35 juta tahun yang lalu di leluhur monyet kuno setelah perbedaan mereka dengan monyet baru. Sebagai konsekuensi dari duplikasi ini, monyet kuno adalah triwarna; yaitu, setiap warna yang dirasakan oleh organisme ini dapat direproduksi dengan mencampur berbagai intensitas dari lampu merah, hijau dan biru. Dengan pengecualian dari howler monkey (genus Alouatta), yang memiliki satu autosomal dan dua X-linked gen (Jacobs et al., 1996), monyet baru memiliki hanya satu autosomal dan satu gen opsin X-linked. Namun, di banyak monyet Baru (misalnya, monyet tupai dan tamarins), opsin X-linked pada lokus yang polimorfik (Jacobs et al., 1993;. Boissinot et al., 1998). Dua alel ini memiliki puncak sensitivitas maksimal mirip dengan
opsin merah manusia dan opsin hijau, sedangkan alel ketiga memiliki puncak sensitivitas maksimal. Untuk alasan ini, seorang wanita yang heterozigot, dua dari tiga alel tersebut adalah trikromatik, sementara laki-laki dan perempuan homozigot adalah dichromatic (Gambar 2.8). Hewan dichromatic tidak bisa membedakan antara merah dan hijau, dan dalam hal ini mereka menyerupai menderita protanopia (buta warna karena kekurangan photopigment merah) atau deuteranopia (warna kebutaan akibat kekurangan photopigment hijau).
Dengan demikian, dalam kasus manusia, kera, dan monyet Afrika, penglihatan trikromatik dicapai dengan mekanisme mirip dengan isozim, (yaitu, protein yang berbeda dikodekan oleh lokus yang berbeda). Heterozigot perempuan, tupai, monyet, sebaliknya, mencapai Trichromatic melalui penggunaan dua "allozymes," (yaitu, protein yang berbeda dikodekan oleh bentuk-bentuk alel yang berbeda pada lokus tunggal). Trichromatic memberikan keuntungan selektif, beberapa alel yang sensitif warna pada lokus di monyet baru dicapai dengan mempertahankan polimorfisme tingkat tinggi pada opsin lokus X-linked. Polimorfisme tingkat tinggi dipertahankan untuk waktu yang lama seperti
dari evolusi waktu kemungkinan memerlukan bentuk seleksi overdominant. Keuntungan selektif penglihatan trichromatic dianggap kemampuan untuk mendeteksi buah matang dengan latar belakang dedaunan hijau lebat
Gambar 2.8 Dasar molekuler dari penglihatan dichromatic dan trichromatic pada laki-laki dan perempuan dari manusia dan monyet baru (NWM) (Sumber: Graur, 2000).
Dating Gene Duplications
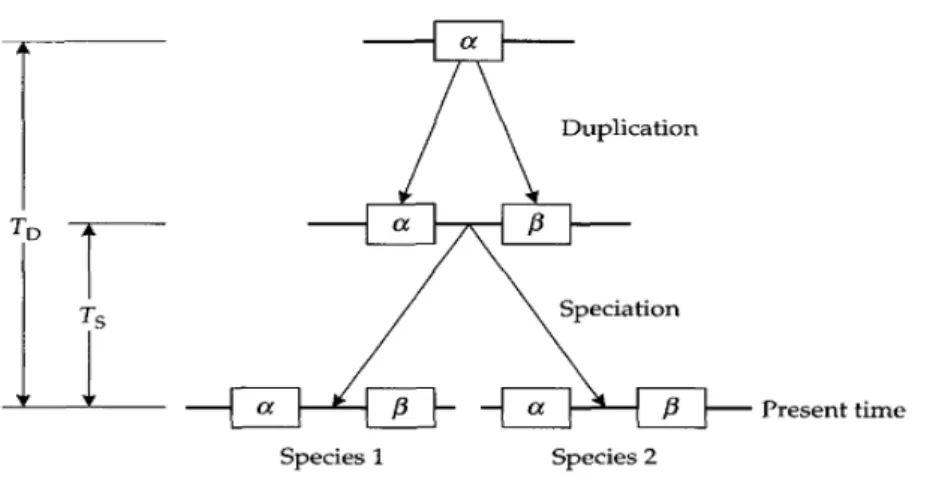
Dua gen dikatakan paralogous jika mereka berasal dari duplikasi, namun dikatakan orthologous jika mereka berasal dari spesiasi. Sebagai contoh, pada Gambar 2.9, gen α dan β berasal dari duplikasi gen leluhur dan karena itu paralogous, sementara gen α dari spesies 1 dan gen α dari spesies 2 adalah orthologous, seperti gen β dari spesies 1 dan gen β dari spesies 2.
Gambar 2.9. Model untuk memperkirakan adanya duplikasi gen (TD).
(Sumber: Graur, 2000).
Gambar 2.9 menunjukkan model yang dapat memperkirakan adanya duplikasi gen (TD). pada gambar terlihat dua gen α dan β, berasal dari waktu duplikasi unit TD pada spesies leluhur. Leluhur spesies dibagi menjadi dua spesies (1 dan 2) Ts unit waktu lalu. Gen α dalam spesies 1 dan 2 yang orthologous, seperti gen β. Gen α yang paralogous ke gen β.
Perkirakan dapat terjadi pada tanggal duplikasi TD, dari urutan data jika telah diketahui tingkat substitusi pada gen α dan β. Tingkat substitusi dapat diperkirakan dari jumlah substitusi antara gen orthologous dengan pengetahuan tentang waktu divergensi, Ts, antara spesies 1 dan 2 (Gambar 2.9). Selanjutnya dapat ditunjukkan bagaimana perkiraan TD dapat diperoleh. Untuk gen α, Kα menjadi jumlah substitusi per situs antara dua jenis. Kemudian, tingkat substitusi gen α, rα, diperkirakan dengan rumus sebagai berikut:
Sedangkan tingkat substitusi di gen β, rβ, dapat diperoleh dengan cara yang sama. Tingkat rata-rata substitusi untuk dua gen menggunakan rumus sebagai berikut:
Untuk memperkirakan TD, kita perlu mengetahui jumlah substitusi per situs antara gen α dan β (Kαβ). Jumlah ini dapat diperkirakan dari empat berpasangan
perbandingan: (1) gen α dari spesies 1 dan gen β dari spesies 2; (2) gen α dari spesies 2 dan gen β dari spesies 1; (3) gen α dan gen β dari spesies 1; dan (1) gen α dan gen β dari spesies 2. Dari empat perkiraan ini kita bisa menghitung nilai rata-rata untuk Kαβ (Ǩαβ), dari hal tersebut kita dapat memperkirakan TD dengan rumus berikut
Perhatikan bahwa dalam kasus gen penyandi protein, dengan menggunakan nomor identik dan nonsynonymous substitusi secara terpisah, kita bisa memperoleh dua perkiraan independen TD. Rata-rata dari dua perkiraan ini mungkin digunakan sebagai perkiraan akhir TD. Namun, jika jumlah substitusi per Situs identik antara gen α dan β besar, dikatakan lebih besar dari 1, maka jumlah substitusi identik tidak dapat diperkirakan secara akurat, dan substitusi identik mungkin tidak memberikan perkiraan yang dapat diandalkan dari nilai TD. Dalam kasus demikian, hanya jumlah substitusi nonsynonymous yang harus digunakan. Sebaliknya, jika jumlah substitusi per situs nonsynonymous antara gen paralogous kecil, maka perkiraan jumlah nonsynonymous substitusi mengikuti pada kesalahan sampling besar, dan dalam kasus tersebut, hanya jumlah substitusi identik harus digunakan. Dalam contoh di atas, kita telah mengasumsikan tingkat keteguhan. Asumsi ini dapat diuji oleh empat perbandingan berpasangan yang disebutkan di atas. Didalam kasus ini, perkiraan TD mungkin keliru. Adanya masalah karena peristiwa evolusi yang kompleks juga mungkin timbul dan mempersulit estimasi TD.
Metode lain untuk mengetahui tentang peristiwa duplikasi gen adalah untuk mempertimbangkan distribusi filogenetik gen dalam hubungannya dengan data paleontologis, yang berkaitan dengan tanggal perbedaan dari spesies yang bersangkutan. Sebagai contoh, semua vertebrata dengan pengecualian ikan tanpa rahang (hagfishes dan lamprey) encode rantai α- dan β-globin. Ada dua kemungkinan penjelasan untuk observasi ini. Salah satunya adalah bahwa peristiwa duplikasi memproduksi α- dan β-globins terjadi dalam nenek moyang dari semua vertebrata (Craniata), tetapi dua ikan tanpa rahang garis keturunan
(Myxini dan Cephalaspidomorphi) telah kehilangan salah satu dari dua duplikasi . Hal ini dimungkinkan tetapi sangat tidak mungkin, karena skenario seperti itu akan menyebabkan kerugian yang terjadi secara independen setidaknya dua garis keturunan evolusi. Penjelasan lain adalah bahwa peristiwa duplikasi terjadi setelah divergence ikan tanpa rahang dari nenek moyang vertebrata berahang (Gnathostomata), tapi sebelum radiasi dari vertebrata berahang.
Penjelasan terakhir ini dianggap lebih masuk akal, dan waktu duplikasi umumnya terjadi pada tahun 450-500.000.000 tahun yang lalu. Jelas, metode di atas hanya dapat memberikan perkiraan kasar dari waktu duplikasi, dan semua perkiraan harus diambil dengan hati-hati. Dalam memperkirakan waktu divergensi antara spesies, pertama dapat menggunakan data dari banyak gen milik banyak keluarga gen. Sebagai perbandingan, memperkirakan waktu duplikasi gen, salah satu yang harus ada yaitu bergantung hanya pada data dari gen milik satu keluarga gen. Karena keterbatasan yang ketat pada urutan data yang dapat digunakan, perkiraan duplikasi gen sering dikenakan standar kesalahan yang sangat besar. Gene Loss
Hampir 7.000 penyakit genetik yang telah didokumentasikan dalam literatur medis (McKusick, 1998) membuktikan fakta bahwa mutasi dapat dengan mudah menghancurkan fungsi gen (protein-coding). Sebagian besar mutasi tersebut merusak, dan dihilangkan dengan cepat dari populasi atau dipertahankan pada frekuensi sangat rendah karena pilihan overdominant atau pergeseran genetik. Namun, menurut Haldane (1932), selama ada salinan gen yang berfungsi secara normal, duplikasi gen dapat merusak mutasi dan menjadi nonfungsional tanpa merugikan kebugaran/ketahanan suatu organisme. Memang, mutasi banyak merusak daripada yang menguntungkan, duplikasi gen berlebihan lebih mungkin untuk menjadi nonfungsional daripada berkembang menjadi gen baru (Ohno 1972).
Unprocessed Pseudogenes
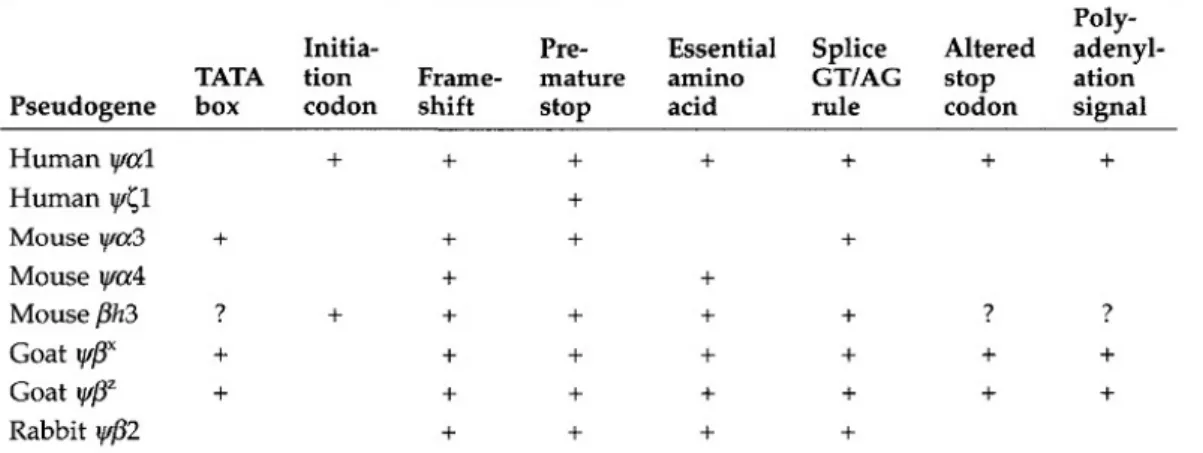
Nonfungsional atau pembungkaman gen karena mutasi yang merusak menghasilkan proses terbentuknya pseudogene, yaitu pseudogene yang belum hilang melalui pengolahan RNA. Sebagian besar pseudogenes yang belum diproses berasal dari nonfungsional yang awalnya fungsional duplikasi gen. Beberapa pseudogen diproses, seperti ΨβX dan ΨβZ di kambing β-globin multigene family, telah diturunkan dari duplikasi dari pseudogene yang sudah ada sebelumnya (Cleary et al., 1981). Jumlah yang sangat kecil yang belum diproses dari pseudogen telah diturunkan dari gen fungsional tanpa duplikasi sebelumnya.
Tabel di atas menunjukkan daftar cacat struktural ditemukan di beberapa pseudogen globin. Sebagian besar pseudogen diproses, berisi beberapa cacat seperti frameshifts, kodon berhenti prematur, dan pemusnahan situs splicing atau peraturan elemen, sehingga sulit untuk mengidentifikasi mutasi yang menyebabkan pembungkaman gen. Dalam beberapa kasus, identifikasi "pelakunya" adalah mungkin dapat diketahui. Misalnya, Ψξ1 pada manusia, hanya satu cacat utama mutasi-yang mungkin adalah penyebab langsung dari nonfungsionalnya. Dalam beberapa kasus, adalah mungkin untuk mengidentifikasi mutasi yang bertanggung jawab untuk nonfungsional gen melalui analisis filogenetik. Sebagai contoh, pseudogene Ψ pada manusia diῃ
saja cukup untuk membungkam itu. β-globin cluster di simpanse dan gorila, ditemukan mengandung
jumlah gen sama dan pseudogen seperti pada manusia, menunjukkan bahwa pseudogene yang diciptakan dan dibungkam sebelum tiga spesies ini menyimpang dari satu jenis yang lain. Tiga pseudogen ditemukan hanya memiliki tiga cacat yang sama: substitusi dalam kodon inisiasi (ATG - GTA); substitusi nonsense di kodon triptofan pada posisi 15 (TGG -> TGA); dan penghapusan di kodon 20 mengakibatkan penghentian kodon di ekson kedua (Chang dan Slightom 1984).
Dengan demikian, "daftar tersangka" dikurangi menjadi tiga. Penelitian lebih lanjut menunjukkan bahwa pseudogene sama, terdapat di semua primata. Perbandingan cacat di antara semua urutan primata menunjukkan bahwa mutasi awal yang bertanggung jawab untuk nonfungsional dari Ψ adalah satu di kodonῃ
inisiasi (Harris et al., 1984). Menariknya, mutasi yang menyebabkan nonfungsional hanya jarang terjadi mutasi missense, kemungkinan besar karena mutasi tersebut menghasilkan produksi protein cacat yang dapat dimasukkan ke produk akhir biologi dan dengan demikian memiliki efek merusak. Misalnya, ada puluhan gen korion-coding dalam genom dari ulat Bombyx mori, jika salah satu dari mereka terdapat gen nonfungsional yang disebabkan oleh mutasi missense, akibatnya seluruh kulit telur menjadi rusak (misalnya, Spoerel et al., 1989).
Karena mereka diciptakan oleh duplikasi, proses pseudogen ditemukan di lingkungan gen homolog fungsional dari yang mereka telah diturunkan. Namun demikian, kasus-kasus di mana pseudogen belum diolah menjadi tersebar sebagai penyusunan ulang genomik. Contoh, sebuah cluster α-globin di tikus terletak pada kromosom 11, namun sebuah proses pseudogene ditemukan pada kromosom 17 (Tan dan Whitney 1993).
Kesatuan Pseudogenes
Sejak pseudogene ini tidak memiliki fungsional counterpart dalam genom manusia, itu disebut Kesatuan pseudogene. Kesatuan pseudogene untuk L-gulono-y-lakton oxidase di guinea babi mengandung perbedaan cacat pada pseudogene
dari manusia, menunjukkan bahwa fungsionalisasi non gen ini terjadi secara independen dalam dua garis keturunan (Nishikimi et al. 1992). Telah dihipotesiskan bahwa guinea pig dan nenek moyang manusia hidup dari diet alami asam askorbat yang kaya, dan karena itu hilangnya enzim tidak merupakan suatu kerugian.
Waktu Pembentukan Gen Fungsional
Sejarah evolusi dari proses pseudogene diasumsikan terdiri dari dua periode yang berbeda. Periode pertama dimulai dengan adanya gen duplikasi dan berakhir ketika duplikat salinan diberikan non fungsional. Selama periode ini, calon pseudogene mungkin mempertahankan fungsi aslinya, dan tingkat substitusi diharapkan tetap kurang lebih sama seperti sebelum acara duplikasi. Setelah hilangnya fungsi, pseudogene yang dibebaskan dari semua kendala fungsional dan laju substitusi nukleotida diperkirakan akan meningkat jauh. Dari sudut pandang evolusi, menarik untuk memperkirakan berapa lama salinan berlebihan dari gen fungsional mungkin tetap fungsional setelah acara duplikasi. Untuk memperkirakan waktu fungsionalisasi non ini, metode berikut telah disarankan (Li et al 1981;. MIY-ata dan Yasunaga 1981).
Pertimbangkan pohon filogenetik pada Gambar 2.10. T menunjukkan waktu perbedaan antara spesies 1 dan 2, yaitu, waktu sejak pemisahan antara gen fungsional orthologous A dan B; TD menunjukkan waktu sejak duplikasi, perbedaan kesalahan antara gen A fungsional dan pseudogene paralogous yang ψA.; dan TN menunjukkan waktu sejak non fungsionalisasi pseudogene ψA. Jumlah substitusi nukleotida per situs pada posisi i dari kodon (i = 1, 2, atau 3) antara ψA dan A, ψA dan B, dan A dan B, dilambangkan sebagai d (ψA) i, d (ψAB) i , dan d (AB) i, dan masing-masing dapat dihitung langsung dari urutan data. Li, Mi, dan Ni menjadi nomor substitusi nukleotida per situs pada posisi kodon i antara titik 0 dan ψA, 0 dan A, dan 0 dan B, masing-masing.
Berdasarkan hal tersebut Li, Mi, dan Ni memperkirakan, sebagai berikut.
Berikut ini, kita mengasumsikan bahwa tingkat substitusi pada posisi kodon yang diberikan adalah sama dalam gen fungsional A dan B. Kami menunjukkan tarif ini dengan aj, di mana subscript i singkatan posisi kodon. Kami juga menganggap bahwa setelah ψA menjadi nonfungsional, yaitu, semua kendala fungsional dilenyapkan, tingkat substitusi nukleotida menjadi sama untuk semua tiga posisi kodon. Kami menunjukkan tingkat ini oleh b. Sebuah harapan yang masuk akal adalah bahwa b akan berubah tobe jauh lebih besar daripada, dan a2, dan mungkin sedikit lebih besar dari a3.
Dengan menetapkan waktu divergensi (T) antara tikus, kelinci, dan manusia pada sekitar 80 juta tahun yang lalu, Li et al. (1981) memperkirakan bahwa pseudogene globin dari tikus ψa3 diciptakan oleh 27 duplikasi gen ± 6 juta tahun yang lalu dan menjadi nonfungsional sekitar 4 juta tahun kemudian. Demikian pula, pseudogene globin pada manusia ψal kehilangan fungsinya sekitar
4 juta tahun setelah itu telah diciptakan oleh 49 duplikasi gen ± 8 juta tahun yang lalu.
Secara umum, tampaknya bahwa mereka mengalami duplikat berlebihan yang pada akhirnya menjadi pseudogen yang mempertahankan fungsi asli mereka untuk waktu yang sangat singkat setelah terjadi peristiwa duplikasi gen. Beberapa pseudogen diproses, seperti pseudogene globin pada kelinci ψβ2, tampaknya kemungkinan hampir kehilangan fungsinya mengikuti gen duplikasi.
Superfamily Globin
Gen superfamili globin telah diketahui memiliki kemungkinan semua jalur evolusi yang terjadi pada famili dengan sekuen yang berulang, sebagai contoh, 1) mempertahankan fungsi asli, 2) penambahan fungsi baru, dan 3) hilangnya fungsi. Pada manusia, superfamili globin terdiri dari tiga famili yang kurang lebih terdapat satu anggota fungsional: famili myoglobin, yang berada pada kromosom 22, famili globin α pada kromosom 16, famili globin β pada kromosom 11. Ketiga famili gen tersebut memproduksi 2 tipe protein fungsional, yaitu myoglobin dan hemoglobin. (Gambar 2.11)
Gambar 2.11. Susunan kromosom dari tiga gen globin pada superfamily globin manusia (Sumber: Graur, 2000).
Dinilai dari fakta mengenai protein globin yang terdapat pada berbagai makhluk hidup, globin tergolong gen yang berasal dari nenek moyang yang asli. Selama masa spesifikasi jaringan, myoglobin menjadi tempat penyimpanan oksigen pada otot, sedangkan hemoglobin menjadi pembawa oksigen dalam darah. Selama masa pembentukan struktur yang berjumlah empat, myoglobin
mempertahankan struktur monomer sedangkan hemoglobin membentuk struktur tetramer. Pada bagian fungsional, myoglobin mengembangkan afinitas yang tinggi untuk oksigen daripada hemoglobin, sedangkan fungsi hemoglobin lebih ke regulasi dan penyaringan. Hemoglobin mamalia, secara singkatnya, memperoleh beberaapa kemempuannya yang tidak ada pada myoglobin. Fungsi tersebut yakni, 1) mengikat molekul oksigen 2) merespon terhadap kadar keasaman dan kadar CO2 dalam sel darah merah dan 3) meregulasi afinitas oksigennya melalui tingkat keberadaan fosfat organik. Kenyataannya, struktur heteromik dari hemoglobin memfasiilitasi perbaikan fungsi dari hemoglobin.
Hemoglobin pada manusia dan sebagian besar vertebrata terbentuk dari dua tipe rantai, satu dikode oleh anggota famili α sedangkan yang lain dikode oleh anggota famili β. Duplikasi yang menciptakan globin α dan β umumnya terjadi secara berurutan, menghasilkan 2 hubungan gen pada satu kromosom.
Gambar 2.12 Sejarah evolusi gen globin pada manusia (Sumber: Graur, 2000). Pada manusia, famili α terdiri dari 4 gen fungsional: gen embrionik ξ, dua gen dewasa, α1 dan α2 dan banyak gen famili anggota yang belum diketahui. Itu juga terdiri dari dari 3 pseudogen yang tidak diproses, ψε, ψα1, dan ψα2. Famili β terdiri dari 5 gen fungsional: gen embrionik ε, dua gen fetal, Gγ dan Aγ, dan dua gen dewasa, β dan δ. Famili ini juga terdiri dari satu pseudogen yang tidak diproses, ψη (yang dulunya ψβ). Dua famili ini mengalami divergensi pada
fisiologisnya maupun regulasi ontologinya. Kenyataannya, hemoglobin secara jelas terlihat perbedaannya pada fase perkembangan: ζ2ε2 dan α2ε2 pada embrio, α2γ2 pada fetus dan α2β2 dan α2δ2 pada fase dewasa. Gen θ1 ditranskripsikan 5-8 minggu setelah kehamilan, namun dengan level yang rendah sehingga protein tidak terdeteksi secara in vivo. Perbedaan pada afinitas pengikatan oksigen ditingkatkan pada globin ini. Sebagai contoh, hemoglobin embronik dan fetal memiliki afinitas yang paling tinggi daripada hemoglobin dewasa, hal ini disebabkan karena mereka tidak mengikat 2,3-difosfogliserate dengan kuat saat bentuk dewasa. Hemoglobin ini juga berfungsi lebih baik pada lingkungan yang kekurangan oksigen (hipoksia) dimana embrio dan fetus berada. Contoh ini mengilustrasikan bagaimana duplikasi gen dapat menghasilkan perbaikan evolusi dari sistem fisiologi.