1
AKTIVITAS OKSIDASI AMONIUM DAN REDUKSI NITRAT
Pseudomonas stutzeri
ASLT2 PADA SUMBER KARBON YANG BERBEDA
Oleh :
Lena Novita
G34102005
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
BOGOR
2006
AKTIVITAS OKSIDASI AMONIUM DAN REDUKSI NITRAT
Pseudomonas stutzeri
ASLT2 PADA SUMBER KARBON YANG BERBEDA
Skripsi
Sebagai Salah Satu Syarat untuk Memperoleh Gelar Sarjana Sains
pada Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
Oleh :
Lena Novita
G34102005
DEPARTEMEN BIOLOGI
FAKULTAS MATEMATIKA DAN ILMU PENGETAHUAN ALAM
INSTITUT PERTANIAN BOGOR
3
Judul
: Aktivitas Oksidasi Amonium dan Reduksi Nitrat
Pseudomonas
stutzeriASLT2 pada Sumber Karbon yang Berbeda
Nama : Lena Novita
NRP
: G34102005
Menyetujui :
Pembimbing I,
Pembimbing II,
Dr. Ir. Iman Rusmana, M.Si.
Dr. Tri Widiyanto, M.Si.
NIP 131956713 NIP 320005966
Mengetahui :
Dekan Fakultas Matematika dan Ilmu Pengetahuan Alam
Institut Pertanian Bogor
Dr. Ir. Yonny Koesmaryono, M.S.
NIP 131473999
RIWAYAT HIDUP
Penulis dilahirkan di Sumedang pada tanggal 7 Nopember 1983 sebagai anak kedua dari tiga bersaudara, dari pasangan Sukardi, SPd. dan Rohmah, AmdPd..
Pada tahun 2002 penulis lulus dari SMU Negeri 1 Sumedang dan pada tahun yang sama diterima di Departemen Biologi, Fakultas Matematika dan Ilmu Pengetahuan Alam (FMIPA), Institut Pertanian Bogor (IPB) melalui jalur Undangan Seleksi Masuk IPB (USMI).
Selama kuliah di Departemen Biologi FMIPA IPB, penulis pernah menjadi asisten praktikum mata kuliah Mikrobiologi Dasar pada tahun ajaran 2004/2005 dan 2005/2006, asisten praktikum mata kuliah Biologi Dasar pada tahun ajaran 2005/2006. Penulis juga pernah aktif di BIOWORLD periode 2003/2004 dan Himpunan Mahasiswa Biologi (Himabio) periode 2004/2005.
Pada bulan Juli-Agustus 2004 penulis melaksanakan praktik magang di Laboratorium Hama dan Penyakit Tanaman, Balai Penelitian Tanaman Obat dan Rempah (Balittro), Bogor. Pada bulan Juni-Juli 2005 penulis melaksanakan Praktik Lapangan di Research and Development Syngenta
stasiun Cikampek dengan topik Pengujian Suatu Bahan Aktif Fungisida di Research and
5
PRAKATA
Segala puji dan syukur penulis panjatkan kepada Allah SWT atas rahmat dan karuniaNya sehingga karya ilmiah ini dapat diselesaikan. Karya ilmiah dengan judul Aktivitas Oksidasi Amonium dan Reduksi Nitrat Pseudomonas stutzeri ASLT2 pada Sumber Karbon yang Berbeda ini disusun berdasarkan hasil penelitian dari bulan Desember 2005 sampai April 2006.
Penulis mengucapkan terima kasih kepada semua pihak yang telah membantu penyusunan karya ilmiah ini, terutama kepada Dr. Ir. Iman Rusmana, MSi. dan Dr. Tri Widiyanto, MSi. selaku pembimbing yang telah membantu dalam penelitian dan penyusunan karya ilmiah serta Dr. Ir. Juliarni MAgr. yang telah memberikan saran dan petunjuknya. Selain itu, penulis mengucapkan terima kasih kepada seluruh staf Laboratorium Mikrobiota, Pusat Penelitian Limnologi, Lembaga Ilmu Pengetahuan Indonesia (LIPI) Cibinong, Bogor atas bantuan dan kerjasamanya selama pelaksanaan penelitian.
Ungkapan terima kasih untuk sahabat-sahabat Ghazali Kost dan teman-teman Biologi 39 atas kebersamaannya. Terima kasih yang tidak terhingga kepada mama, papa, A Yudi dan Leni atas segala do’a, dukungan, serta kasih sayangnya.
Semoga karya ilmiah ini bermanfaat.
Bogor, Juli 2006
ABSTRAK
LENA NOVITA. Aktivitas Oksidasi Amonium dan Reduksi Nitrat Pseudomonas stutzeri ASLT2 pada Sumber C yang Berbeda. Dibimbing oleh IMAN RUSMANA dan TRI WIDIYANTO.
Isolat ASLT2 merupakan bakteri pengoksidasi amonium yang diisolasi dari perairan tambak udang. Berdasarkan analisis molekuler 16S rRNA, isolat ini memiliki kemiripan sebesar 99 % dengan bakteri pereduksi nitrat P. stutzeri.
Hasil pengamatan menunjukkan bahwa isolat ASLT2 dapat mengoksidasi amonium pada kondisi autotrof dengan karbonat sebagai sumber karbon dan kondisi heterotrof dengan asetat, suksinat, glukosa, dan gliserol sebagai sumber C. Konsentrasi amonium menurun hinggá tujuh hari inkubasi pada semua perlakuan sumber C. Penurunan konsentrasi amonium yang terjadi disebabkan penggunaan amonium sebagai sumber nitrogen dan sumber energi melalui proses nitrifikasi. Isolat ini memiliki aktivitas oksidasi amonium paling tinggi pada medium dengan menggunakan suksinat sebagai sumber C. Isolat ASLT2 dapat mereduksi nitrat pada kondisi aerob dan anaerob dengan asetat, suksinat, glukosa dan gliserol sebagai sumber C. Aktivitas reduksi nitrat secara aerob paling tinggi terjadi pada medium dengan menggunakan glukosa sebagai sumber C dan secara anaerob aktivitas paling tinggi terjadi pada medium dengan menggunakan suksinat sebagai sumber C. Perbedaan aktivitas reduksi nitrat pada masing-masing perlakuan sumber C disebabkan oleh perbedaan banyaknya elektron yang dihasilkan dari proses oksidasi sumber C yang berbeda.
ABSTRACT
LENA NOVITA. Activity of Ammonium Oxidation and Nitrate Reduction by Pseudomonas
stutzeri ASLT2 on Different Carbon Sources. Supervised by IMAN RUSMANA and TRI WIDIYANTO.
Isolate ASLT2 was an ammonium oxidizing bacterium isolated from shrimp pond. Based on mollecular analysis of 16S rRNA sequences, the isolate was 99 % closely related to nitrate reducing bacterium P. stutzeri.
The result showed that isolate ASLT2 was able to oxidize ammonium autothrophically with carbonate as a carbon source and heterotrophycally with acetate, succinate, glucose, and glycerol as C sources. Ammonium concentration was decreased up to seven days of incubation in all C source treatments. The declining of ammonium concentration was due to the utilization of amonium as nitrogen and energy source through nitrification process. The highest activity of ammonium oxidation was found in the medium using succinate as C source. Isolate ASLT2 was able to reduce nitrate aerobically and anaerobically with acetate, succinate, glucose, and glycerol as C sources . The highest activity of aerobic nitrate reduction was found in the medium using glucose as C source and anaerobically was found in the medium using succinate as C source. The different of nitrate reduction activity in each C sources was due to the different amount of electron generated by oxidation process using different C sources.
7
DAFTAR ISI
Halaman
DAFTAR TABEL ... vii
DAFTAR GAMBAR... vii
DAFTAR LAMPIRAN... vii
PENDAHULUAN Latar Belakang... 1
Tujuan ... 1
Waktu dan Tempat Penelitian ... 1
BAHAN DAN METODE Bahan... 1
Metode Penyiapan Medium ... 1
Pemurnian dan Peremajaan Isolat ... 1
Uji Aktivitas Oksidasi Amonium... 2
Kinetika Oksidasi Amonium... 2
Uji Aktivitas Reduksi Nitrat... 2
Kinetika Reduksi Nitrat... 2
Analisis Kadar Amonium... 3
Analisis Kadar Nitrit ... 3
Analisis Kadar Nitrat ... 3
HASIL DAN PEMBAHASAN Pemurnian dan Peremajaan Isolat ... 3
Aktivitas Oksidasi Amonium... 3
Aktivitas Reduksi Nitrat... 5
SIMPULAN ... 10
SARAN ... 10
DAFTAR PUSTAKA... 10
DAFTAR TABEL
Halaman 1 Kemampuan P. stutzeri ASLT2 untuk mengoksidasi amonium dengan menghasilkan
senyawa nitrit d an gas setelah 7 hari inkubasi... 5 2 Laju oksidasi amonium (µNH3) dan laju pertumbuhan spesifik (µsel) isolat ASLT2 pada
medium nitrifikasi dengan sumber C yang berbeda ... 5 3 Kemampuan isolat ASLT2 dalam mereduksi nitrat dan menghasilkan senyawa nitrit
dan gas pada kondisi aerob dengan sumber C yang berbeda... 8 4 Kemampuan isolat ASLT2 dalam mereduksi nitrat dan menghasilkan senyawa nitrit
dan gas pada kondisi anaerob dengan sumber C yang berbeda... 8
DAFTAR GAMBAR
Halaman 1 Profil penurunan senyawa amonium (a) oleh isolat ASLT2 pada sumber C yang
berbeda dengan senyawa nitrit (b) dan nitrat (c) yang dihasilkan... 4 2 Pertumbuhan isolat ASLT2 pada medium nitrifikasi dengan sumber C yang berbeda... 4 3 Profil penurunan senyawa nitrat oleh isolat ASLT2 pada kondisi aerob (a) dan anaerob (b) dengan sumber C yang berbeda... 7 4 Profil akumulasi senyawa nitrit medium selama proses reduksi nitrat oleh isolat ASLT2 pada kondisi aerob (a) dan anaerob (b)... 7 5 Pertumbuhan isolat ASLT2 pada kondisi aerob (a) dan anaerob (b) dengan sumber C
yang berbeda ... 7
DAFTAR LAMPIRAN
Halaman 1 Profil senyawa amonium pada medium kontrol pengujian aktivitas oksidasi
amonium... 13 2 Profil senyawa nitrat pada medium kontrol pengujian aktivitas reduksi nitrat secara
aerob (a) dan anaerob (b) ... 13 5 Laju reduksi nitrat (µN03) dan laju pertumbuhan spesifik (µsel) isolat ASLT2 dengan
1
PENDAHULUAN
Latar Belakang
Isolat ASLT2 merupakan hasil penelitian sebelumnya yang diisolasi pada medium nitrifikasi dan telah diuji kemampuan nitrifikasinya. Isolat ini diisolasi dari lingkungan perairan tambak udang di Kendari (Widiyanto 2006). Nitrifikasi merupakan proses oksidasi senyawa amonium menjadi nitrit dan nitrat. Proses ini dapat terjadi pada kondisi pertumbuhan autotrof maupun heterotrof (Bock et al. 1992, Atlas & Bartha 1998). Isolat ASLT2 memiliki kemampuan oksidasi amonium yang paling tinggi dibandingkan isolat-isolat bakteri nitrifikasi yang telah berhasil diisolasi. Hasil analisis molekuler 16S rRNA menunjukkan bahwa isolat ASLT2 memiliki kemiripan sebesar 99 % dengan Pseudomonas stutzeri (Widiyanto 2006).
P. stutzeri merupakan bakteri gram negatif yang berbentuk batang, bersifat nonfermentatif dan motil (Madigan et al. 2000, Rius et al. 2001). Bakteri ini dapat ditemukan di tanah dan lingkungan perairan. P. stutzeri termasuk bakteri denitrifikasi. Denitrifikasi ialah proses reduksi nitrat menjadi nitrit, nitrit menjadi nitrik oksida, nitrik oksida menjadi gas nitrous oksida hingga pada akhirnya dihasilkan gas nitrogen (Atlas & Bartha 1998, Richardson 2000). Richardson (2000) mengemukakan bahwa denitrifikasi merupakan proses reduksi nitrat yang berhubungan langsung dengan proses transfer elektron dan senyawa organik berperan sebagai donor elektron. P. stutzeri merupakan bakteri denitrifikasi yang mampu mereduksi nitrat dengan menghasilkan gas nitrogen (Korner & Zumft 1989, Zumft 1997, Rius et al. 2001).
Laju oksidasi amonium dan reduksi nitrat pada bakteri dipengaruhi oleh ketersediaan sumber karbon sebagai sumber utama untuk memperoleh energi bagi pertumbuhan (Kelso et al. 1999, Firth & Edwards 2000). Bakteri pereduksi nitrat merupakan mikroorganisme heterotrof yang memerlukan sumber karbon organik seperti asam asetat, asam propionat, asam benzoat, gliserol, metanol dan glukosa untuk pertumbuhannya (Teixeira & Oliveira 2002).
Kemampuan P. stutzeri dalam
mengoksidasi amonium hingga saat ini belum pernah dilaporkan. Isolat ASLT2 yang
merupakan P. stutzeri telah diketahui
memiliki kemampuan untuk mengoksidasi amonium tetapi belum pernah diuji
kemampuan reduksi nitratnya. Oleh karena itu, diperlukan pengujian aktivitas oksidasi amonium dan reduksi nitrat dari isolat bakteri tersebut.
Tujuan
Penelitian ini bertujuan untuk menelaah pengaruh berbagai jenis sumber karbon terhadap aktivitas oksidasi amonium dan reduksi nitrat oleh P. stutzeri ASLT2.
Waktu dan Tempat Penelitian
Penelitian ini dilaksanakan dari bulan Desember 2005 sampai dengan April 2006 di Laboratorium Mikrobiota, Pusat Penelitian Limnologi, LIPI Cibinong, Bogor.
BAHAN DAN METODE
Bahan
Bahan yang digunakan ialah isolat bakteri pengoksidasi amonium asal tambak udang P.
stutzeri ASLT2 koleksi Laboratorium
Mikrobiota, Pusat Penelitian Limnologi, LIPI Cibinong, Bogor.
Metode
Penyiapan Medium
Medium cair nitrifikasi yang digunakan memiliki komposisi (g/l) sebagai berikut 13.5 KH2PO4, 0.7 K2HPO4, 0.1 MgCl2.6H2O, 0.5
NaHCO3, 0.014 FeCl3.6H2O, 0.18
CaCl2.2H2O, 0.1 NH4Cl, 0.2 EDTA, dan 0.5 glukosa (Rodina 1972) dengan salinitas 2% dan pH 7.
Medium cair denitrifikasi yang digunakan memiliki komposisi (g/l) sebagai berikut 10 Na-asetat, 5 KNO3, 0.5 (NH4)2SO4, 0.2 KH2PO4, 0.9 K2HPO4, 0.1 CaCl2.2H2O, 0.5 MgSO4.7H2O, dan 0.2 EDTA (Rodina 1972) dengan salinitas 2% dan pH 7. Kondisi anaerob dibuat dengan metode Oxygen-Free Nitrogen (OFN). Gas N2 dialirkan ke dalam medium dengan menggunakan syringe steril selama 10 menit.
Medium padat nitrifikasi dan denitrifikasi dibuat dengan menambahkan 20 g/ l bacto agar.
Pemurnian dan Peremajaan Isolat
Pemurnian isolat dilakukan dengan menumbuhkan isolat bakteri pada medium agar nitrifikasi melalui metode kuadran dan diinkubasi pada suhu ruang (28-31 °C) selama
tujuh hari. Koloni murni yang diperoleh ditumbuhkan kembali pada medium agar nitrifikasi dan denitrifikasi, kemudian diinkubasi pada suhu ruang selama 3-4 hari. Uji Aktivitas Oksidasi Amonium
Inokulum untuk uji aktivitas disiapkan dengan menumbuhkan isolat pada 50 ml medium cair nitrifikasi, kemudian diinkubasi di atas inkubator berpenggoyang pada kecepatan 80 rpm dan suhu ruang (28-31 °C) selama tiga hari. Sebanyak 1 ml kultur isolat ditumbuhkan pada 50 ml medium nitrifikasi dengan sumber karbon dari karbonat, glukosa, asetat, suksinat, dan gliserol dengan konsentrasi amonium ± 5000 µM. Kemudian diinkubasi selama tujuh hari di atas inkubator berpenggoyang pada kecepatan 80 rpm dan suhu ruang (28-31 °C). Masing-masing perlakuan sumber karbon diulang tiga kali. Kontrol masing-masing perlakuan dibuat tanpa diinokulasi kultur bakteri.
Pengukuran pertumbuhan sel dan aktivitas oksidasi amonium dilakukan pada hari ke-0, 2, 3, 5, dan 7. Pengukuran pertumbuhan sel dilakukan melalui analisis kerapatan optik (OD) dengan menggunakan spektrofotometer pada panjang gelombang 550 nm. Pengukuran aktivitas oksidasi amonium dilakukan melalui analisis kadar amonium, nitrit, dan nitrat (Greenberg et al. 1992). Filtrat yang digunakan untuk analisis kadar amonium, nitrit, dan nitrat diperoleh melalui teknik penyaringan dengan menggunakan membran nitroselulosa berdiameter pori 0.45 µm (Whatman).
Jumlah amonium yang dioksidasi dari medium diperoleh dengan menghitung selisih kadar amonium awal dengan kadar amonium saat analisis. Laju pertumbuhan spesifik (µsel) bakteri dalam masing-masing medium perlakuan dan laju oksidasi amonium (µNH3) ditentukan berdasarkan hasil pengukuran tersebut (White 2000).
Kinetika Oksidasi Amonium
Inokulum untuk pengujian disiapkan dengan menumbuhkan isolat pada 50 ml medium cair nitrifikasi yang menggunakan suksinat sebagai sumber C, kemudian diinkubasi di atas inkubator berpenggoyang pada kecepatan 80 rpm dan suhu ruang (28-31 °C) selama tiga hari. Kultur bakteri dibuat pelet dengan cara sentrifugasi pada kecepatan 5000 rpm selama 15 menit. Pelet dipisahkan dan diresuspensi dengan medium nitrifikasi
diinokulasikan ke dalam 50 ml medium nitrifikasi dan suksinat sebagai sumber C dengan konsentrasi amonium 0, 500, 1000, 1500, dan 2000 µM. Kemudian diinkubasi selama 20 jam di atas inkubator berpenggoyang pada kecepatan 80 rpm dan suhu ruang (28-31 °C). Masing-masing perlakuan diulang tiga kali. Kontrol masing-masing perlakuan dibuat tanpa diinokulasi kultur bakteri.
Pengukuran pertumbuhan sel dan pengukuran aktivitas oksidasi amonium dilakukan setiap 4 jam. Analisis kinetika oksidasi amonium dilakukan dengan menggunakan persamaan kinetika Michaelis -Menten (White 2000).
Uji Aktivitas Reduksi Nitrat
Pengujian aktivitas reduksi nitrat dilakukan pada kondisi aerob dan anaerob. Inokulum untuk pengujian disiapkan dengan menumbuhkan isolat pada 50 ml medium cair denitrifikasi, kemudian diinkubasi di atas inkubator berpenggoyang pada kecepatan 80 rpm dan suhu ruang (28-31 °C) selama 3 hari. Sebanyak 1 ml kultur isolat ditumbuhkan pada 50 ml medium denitrifikasi dengan sumber karbon dari glukosa, asetat, suksinat, dan gliserol dengan konsentrasi nitrat ± 5000 µM kemudian diinkubasi selama tujuh hari di atas inkubator berpenggoyang pada kecepatan 80 rpm dan suhu ruang (28-31 °C). Masing-masing perlakuan sumber karbon diulang tiga kali dan kontrol masing-masing perlakuan dibuat tanpa diinokulasi kultur bakteri. Pengujian aktivitas reduksi nitrat pada kondisi anaerob dilakukan dengan menggunakan medium denitrifikasi anaerob dan tanpa aerasi.
Pengukuran pertumbuhan sel dan pengukuran aktivitas reduksi nitrat dilakukan pada hari ke-0, 2, 3, 5, dan 7. Pengukuran pertumbuhan sel dilakukan melalui analisis OD pada panjang gelombang 550 nm. Pengukuran aktivitas reduksi nitrat dilakukan melalui analisis kadar nitrat, nitrit, dan amonium (Greenberg et al. 1992).
Jumlah nitrat yang direduksi dari medium diperoleh dengan menghitung selisih kadar nitrat awal dengan kadar nitrat yang diperoleh saat analisis. Laju pertumbuhan spesifik (µsel) bakteri dalam masing-masing medium perlakuan dan laju reduksi nitrat (µNO3) ditentukan berdasarkan hasil pengukuran tersebut (White 2000).
3
Kinetika Reduksi Nitrat
Inokulum untuk pengujian disiapkan dengan menumbuhkan isolat pada 50 ml medium cair denitrifikasi dengan glukosa sebagai sumber C, kemudian diinkubasi di atas inkubator berpenggoyang pada kecepatan 80 rpm dan suhu ruang (28-31 °C) selama tiga hari. Kultur bakteri dibuat pelet dengan cara sentrifugasi pada kecepatan 5000 rpm selama 15 menit. Pelet dipisahkan dan diresuspensi dengan medium denitrifikasi tanpa nitrat. Sebanyak 2 ml kultur bakteri diinokulasikan ke dalam 50 ml medium denitrifikasi dan glukosa sebagai sumber C dengan konsentrasi nitrat 0, 500, 1000, 1500, dan 2000 µM kemudian diinkubasi selama 20 jam di atas inkubator berpenggoyang pada kecepatan 80 rpm dan suhu ruang (28-31 °C). Masing-masing perlakuan diulang tiga kali. Kontrol masing-masing perlakuan dibuat tanpa diinokulasi kultur bakteri.
Pengukuran pertumbuhan sel dan pengukuran aktivitas reduksi nitrat dilakukan setiap 4 jam. Analisis kinetika reduksi nitrat dilakukan dengan menggunakan persamaan kinetika Michaelis -Menten (White 2000). Analisis Kadar Amonium
Kadar amonium ditentukan dengan metode spektrofotometri. Sebanyak 5 ml sampel yang telah disaring ditambah dengan 0.2 ml fenol alkohol 10 %, 0.2 ml nitroprusid 0.5 %, dan 0.5 ml campuran hipoklorit teknis dan asam sitrat 20 % (1:4), kemudian didiamkan selama 1 jam. Setiap setelah pemberian pereaksi dilakukan pengadukan. Pemberian pereaksi akan menghasilkan warna biru. Warna yang terbentuk dibaca serapannya pada panjang gelombang 640 nm (Greenberg et al. 1992). Nilai absorbans yang diperoleh dikonversi menggunakan standar hingga diperoleh satuan dalam konsentrasi (µM). Analisis Kadar Nitrit
Sebanyak 5 ml sampel yang telah disaring ditambah dengan 0.1 ml sulfanilamid 1 % dan 0.1 ml NED (Naftalena Etilena Diamina) 0.1 %, kemudian didiamkan selama 10 menit. Setiap setelah pemberian pereaksi dilakukan pengadukan. Pemberian pereaksi akan menghasilkan warna merah muda hingga keunguan. Warna yang terbentuk dibaca serapannya pada panjang gelombang 540 nm (Greenberg et al. 1992). Nilai absorbans yang diperoleh dikonversi menggunakan standar hingga diperoleh satuan dalam konsentrasi (µM).
Analisis Kadar Nitrat
Sebanyak 5 ml sampel yang telah disaring ditambah dengan 0.2 ml brusin 0.5 % dan 4 ml asam sulfat pekat, kemudian didiamkan selama 0.5 jam. Setiap setelah pemberian pereaksi dilakukan pengadukan. Pemberian pereaksi akan menghasilkan warna kuning. Warna yang terbentuk dibaca serapannya pada panjang gelombang 420 nm (Greenberg et al. 1992). Nilai absorbans yang diperoleh dikonversi menggunakan standar hingga diperoleh satuan dalam konsentrasi (µM).
HASIL DAN PEMBAHASAN
Pemurnian dan Peremajaan Isolat
Isolat murni ASLT2 diperoleh pada medium nitrifikasi setelah 7-10 hari inkubasi dengan bentuk koloni bulat, berwarna putih
dengan diameter koloni ± 1 mm. ASLT2
diisolasi pada medium nitrifikasi autotrof (Widiyanto 2006). Isolat ASLT2 dapat tumbuh pada medium agar nitrifikasi dan denitrifikasi setelah 2-3 hari inkubasi.
Aktivitas Oksidasi Amonium
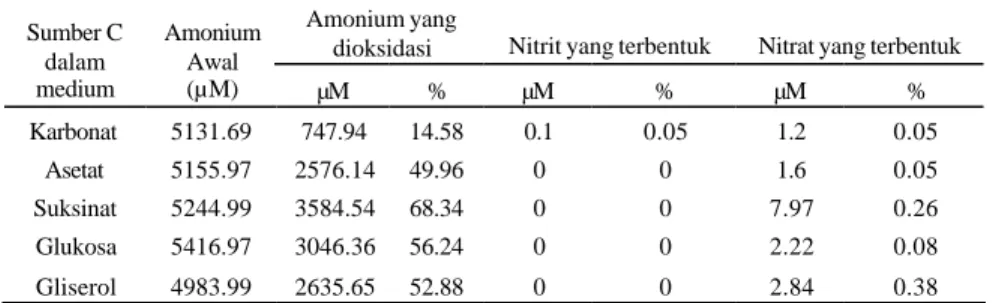
P. stutzeri ASLT2 dapat mengoksidasi amonium baik pada medium autotrof dengan pemberian karbonat sebagai sumber C maupun pada medium heterotrof dengan asetat, suksinat, glukosa, dan gliserol sebagai sumber C. Kemampuan isolat dalam mengoksidasi amonium ditunjukkan oleh penurunan konsentrasi amonium pada medium pertumbuhannya (Gambar 1(a)). Konsentrasi senyawa amonium tidak mengalami perubahan pada medium kontrol (Lampiran 1).
Konsentrasi senyawa amonium menurun hingga tujuh hari inkubasi pada setiap perlakuan sumber C. Penurunan konsentrasi senyawa amonium diduga disebabkan penggunaan amonium oleh isolat ASLT2 sebagai sumber nitrogen dan sumber energi. Penggunaan amonium sebagai sumber N yaitu melalui proses asimilasi, dimana amonium diubah menjadi senyawa nitrogen organik untuk sintesis material sel. Penggunaan senyawa amonium sebagai sumber energi yaitu melalui proses nitrifikasi, dimana amonium dioksidasi menjadi senyawa nitrit dan nitrat (Bock et al. 1992, Arp & Stein 2003). Proses ini diduga terjadi pada kultur isolat dengan adanya akumulasi senyawa nitrat dalam medium (Gambar 1 (c)).
Penurunan senyawa amonium tidak diikuti oleh akumulasi senyawa nitrit dalam médium dengan asetat, suksinat, glukosa, dan gliserol sebagai sumber C (Gambar 1 (b)). Senyawa nitrit hanya dihasilkan pada medium dengan karbonat sebagai sumber C (Gambar 1 (b), Tabel 1). 0 1000 2000 3000 4000 5000 6000 0 1 2 3 4 5 6 7 Hari ke-Amonium (µM)
Asetat Suksinat Glukosa Gliserol Karbonat
(a) 0.00 0.02 0.04 0.06 0.08 0.10 0.12 0 1 2 3 4 5 6 7 Hari ke-Nitrit (µM)
Asetat Suksinat Glukosa Gliserol Karbonat
(b) -2 0 2 4 6 8 10 0 1 2 3 4 5 6 7 Hari ke-Nitrat (µM)
Asetat Suksinat Glukosa Gliserol Karbonat
Gambar 1 Profil penurunan senyawa
amonium (a) oleh isolat
ASLT2 pada sumber C yang berbeda dengan senyawa nitrit (b) dan nitrat (c) yang
Penurunan senyawa amonium pada masing-masing perlakuan sebanding dengan peningkatan biomassa sel bakteri dalam mediu m pertumbuhan (Gambar 2). Kemampuan isolat ASLT2 untuk mengoksidasi amonium pada medium heterotrof lebih tinggi dibandingkan autotrof. Isolat ASLT2 memiliki aktivitas oksidasi amonium paling tinggi pada medium dengan suksinat sebagai sumber C, walaupun pertumbuhan selnya cenderung lebih rendah (Gambar 2). Dengan demikian, dapat diduga bahwa aktivitas oksidasi amonium dengan suksinat sebagai sumber C per sel bakteri lebih tinggi. Isolat ini pada medium dengan suksinat sebagai sumber C dapat mereduksi nitrat hingga 68.34% setelah tujuh hari inkubasi (Tabel 1). Pertumbuhan isolat lebih tinggi pada medium dengan glukosa sebagai sumber C dibandingkan dengan sumber C yang lainnya (Gambar 2) tetapi aktivitas
oksidasi amoniumnya lebih rendah
dibandingkan dengan suksinat sebagai sumber C. -0.05 0.00 0.05 0.10 0.15 0.20 0.25 0 1 2 3 4 5 6 7 Hari ke- OD Sel (550 nm)
Asetat Suksinat Glukosa Gliserol Karbonat
Gambar 2 Pertumbuhan isolat ASLT2 pada medium nitrifikasi dengan sumber C yang berbeda.
Penurunan senyawa amonium pada masing-masing perlakuan sumber C oleh isolat ASLT2 mempunyai laju yang berbeda (Tabel 2). Nilai laju penurunan senyawa amonium yang tinggi pada medium dengan suksinat sebagai sumber C sebanding dengan jumlah amonium yang dapat dioksidasi oleh isolat pada medium dengan sumber C tersebut. Berdasarkan analisis kinetika diperoleh nilai laju penurunan senyawa
amonium maksimum (Vmax) oleh isolat
ASLT2 sebesar 39.216 µM/jam dengan KM
sebesar 1.604 µM.
5
Nitrifikasi merupakan proses oksidasi amonium menjadi hidroksilamin, oksidasi hidroksilamin menjadi nitrit, dan oksidasi senyawa nitrit menjadi nitrat dengan produk samping gas NO dan N2O (Paul & Clark 1996 diacu dalam Rusmana 2003). Tahapan reaksi dalam proses ini melibatkan enzim yang berbeda, yaitu amonia monooksigenase (AMO) yang mengubah amonium menjadi hidroksilamin, hidroksilamin oksidoreduktase (HAO) yang mengubah hidroksilamin menjadi nitrit, dan nitrit oksidoreduktase (NO) yang mengubah nitrit menjadi nitrat (Bock et a1 1992, Atlas & Bartha 1998).
Proses nitrifikasi dapat terjadi pada kondisi pertumbuhan autotrof maupun heterotrof. Perbedaan oksidasi amonium pada masing-masing perlakuan sumber C diduga dipengaruhi oleh kemampuan bakteri untuk memperoleh energi dari sumber C yang tersedia. Bakteri nitrifikasi autotrofik memanfaatkan CO2 sebagai sumber C dan memperoleh energi dari oksidasi amonium atau nitrit. Sedangkan bakteri nitrifikasi heterotrofik memanfaatkan senyawa organik sebagai sumber C dan memperoleh energi dari oksidasi senyawa tersebut (Atlas & Bartha 1998).
Kemampuan mikroorganisme untuk memperoleh energi pada kondisi heterotrof bergantung pada kemampuan metabolismenya untuk mengoksidasi senyawa karbon (bahan organik) sebagai sumber energi utama. Senyawa karbon dalam metabolisme berperan penting yaitu menghasilkan energi melalui oksidasi senyawa tersebut dan menyediakan unsur C untuk pembentukan material sel (Prescott et al. 2000).
Oksidasi suksinat dalam sistem
metabolisme di dalam sel berlangsung melalui siklus asam sitrat (TCA). Oksidasi senyawa suksinat menjadi fumarat dikatalisis oleh enzim suksinat dehidrogenase (Prescott et al. 2000, White 2000). Suksinat dehidrogenase merupakan enzim flavin yang terikat pada protein membran dan secara langsung mentransfer elektron dari matriks ke koenzim Q pada rantai respirasi. Potensial redoks suksinat/fumarat memiliki nilai yang hampir
sama dengan potensial redoks QH2/Q
sehingga perubahan energi bebas yang terjadi pada tahap ini tidak cukup untuk memompa elektron dari matriks ke ruang intermembran (White 2000). Hal ini diduga menyebabkan bakteri kesulitan untuk mendapatkan energi sehingga pertumbuhan selnya cenderung lebih rendah dibandingkan pada medium heterotrof
Amonium yang
dioksidasi Nitrit yang terbentuk Nitrat yang terbentuk Sumber C dalam medium Amonium Awal (µM) µM % µM % µM % Karbonat 5131.69 747.94 14.58 0.1 0.05 1.2 0.05 Asetat 5155.97 2576.14 49.96 0 0 1.6 0.05 Suksinat 5244.99 3584.54 68.34 0 0 7.97 0.26 Glukosa 5416.97 3046.36 56.24 0 0 2.22 0.08 Gliserol 4983.99 2635.65 52.88 0 0 2.84 0.38 Sumber C
dalam medium µNH3 (µM/ hari) R-sq (µNH3) µsel(sel/hari) R-sq (µsel)
Karbonat 0.0871 0.999 0.1940 0.919
Asetat 0.2250 0.845 0.1955 0.817
Suksinat 0.3564 0.975 0.2021 0.974
Glukosa 0.2373 0.868 0.1477 0.622
Gliserol 0.1267 0.803 0.2511 0.725
Tabel 1 Kemampuan P. stutzeri ASLT2 untuk mengoksidasi amonium dengan menghasilkan senyawa nitrit dan nitrat setelah 7 hari inkubasi
Tabel 2 Laju penurunan senyawa amonium (µNH3) dan laju pertumbuhan spesifik (µsel) isolat ASLT2 pada medium nitrifikasi dengan sumber C yang berbeda
lainnya (Gambar 2). Aliran elektron pada ruang intermembran melalui rantai respirasi akan menyebabkan sintesis energi (Prescott et al. 2000, White 2000). Oleh karena itu, diduga oksidasi amonium merupakan proses utama untuk memperoleh energi bagi pertumbuhan sel bakteri pada saat suksinat sebagai satu-satunya sumber C yang tersedia sehingga kadar amonium yang dioksidasi lebih tinggi. Aktivitas oksidasi amonium yang tinggi ditunjukkan oleh laju penurunan senyawa amonium yang tinggi pada medium dengan sumber C tersebut (Tabel 2). Oksidasi amonium menjadi nitrit dan nitrat merupakan proses metabolisme mikroorganisme untuk memperoleh energi (Atlas & Bartha 1998, Madigan et al. 2000).
Kelompok bakteri Pseudomonas dapat memanfaatkan glukosa sebagai sumber C dengan lintasan oksidasi Entner-Doudoroff (Prescott et al. 2000). Oksidasi 1 molekul glukosa secara total melalui rantai respirasi dengan oksigen sebagai akseptor elektron menghasilkan 38 ATP (Prescott et al. 2000, White 2000). Perolehan energi ini lebih besar dibandingkan hasil oksidasi satu molekul asetat, suksinat dan gliserol sehingga dapat mendukung pertumbuhan sel bakteri dengan baik. Proses oksidasi amonium menjadi nitrit dan nitrat diduga dapat berperan sebagai lintasan untuk memperoleh energi ketika bakteri tidak memperoleh energi yang cukup dari oksidasi senyawa karbon. Pertumbuhan bakteri heterotrofik sangat bergantung pada oksidasi senyawa karbon untuk memperoleh energi (Bock et al. 1992).
Pertumbuhan sel isolat ASLT2 (Gambar 2) dan aktivitas oksidasi amonium pada medium autotrof sangat rendah dibandingkan pada kondisi heterotrof (Gambar 1). Bakteri
autotrofik harus mampu memfiksasi CO2
untuk membentuk material sel. Energi untuk fiksasi tersebut dapat diperoleh dari oksidasi senyawa anorganik. Bakteri pengoksidasi amonium autotrofik memperoleh energi melalui proses oksidasi amonium menjadi nitrit dan nitrat (Bock et al. 1992, Arp & Stein 2003).
Senyawa nitrit yang tidak diperoleh saat analisis dan nitrat yang rendah (Tabel 1) diduga senyawa amonium dioksidasi menjadi senyawa nitrogen organik. Selain itu, amonium diduga dioksidasi menjadi
hidroksilamin. Bock et al. (1992)
mengemukakan bahwa senyawa nitrit dapat diperoleh pada kultur bakteri nitrifikasi
reduktase dihambat oleh kadar oksigen yang rendah.
Aktivitas Reduksi Nitrat
P. stutzeri ASLT2 dapat mereduksi nitrat pada kondisi aerob maupun anaerob. Kemampuan isolat dalam mereduksi nitrat ditunjukkan oleh penurunan konsentrasi senyawa nitrat dalam médium pertumbuhan (Gambar 3). Perubahan konsentrasi senyawa nitrat tidak terjadi pada medium kontrol (Lampiran 2). Penurunan senyawa nitrat diduga disebabkan penggunaan nitrat sebagai akseptor elektron alternatif pengganti oksigen dalam rantai respirasi. Peningkatan kadar nitrat yang direduksi sebanding dengan peningkatan biomassa sel dalam medium pertumbuhan (Gambar 5).
Penurunan senyawa nitrat oleh isolat ASLT2 pada medium diikuti oleh akumulasi senyawa nitrit (Gambar 4). Isolat ASLT2 dapat mereduksi senyawa nitrat menjadi nitrit dan diduga dapat menghasilkan gas baik pada kondisi aerob (Tabel 3) maupun anaerob (Tabel 4) dengan sumber C asetat, glukosa, dan gliserol. Sedangkan dengan suksinat sebagai sumber C pada kondisi aerob tidak menghasilkan senyawa nitrit (Tabel 3). Senyawa nitrit yang terakumulasi dalam medium pada kondisi pertumbuhan anaerob relatif lebih tinggi dibandingkan kondisi aerob. Penurunan senyawa nitrit dalam medium diduga disebabkan penggunaan nitrit oleh isolat ASLT2 sebagai akseptor elektron alternatif pengganti oksigen dalam rantai respirasi.
Pemberian sumber C yang berbeda pada medium memberikan pengaruh yang berbeda terhadap aktivitas dan laju reduksi nitrat. Isolat ASLT2 memiliki aktivitas reduksi nitrat dan pertumbuhan sel yang lebih tinggi pada medium dengan glukosa dibandingkan gliserol dan asetat sebagai sumber C baik pada kondisi aerob maupun anaerob. Aktivitas reduksi nitrat paling tinggi pada kondisi aerob terjadi dengan glukosa sebagai sumber C dan paling rendah dengan suksinat sebagai sumber C. Aktivitas reduksi nitrat (Gambar 3) dan pertumbuhan isolat (Gambar 5) paling tinggi pada kondisi anaerob terjadi dengan suksinat sebagai sumber C.
Laju reduksi senyawa nitrat pada masing-masing perlakuan sumber C dapat dilihat pada
Tabel 5. Berdasarkan analisis kinetika
diperoleh nilai laju reduksi nitrat maksimum (Vmax) oleh isolat ASLT2 sebesar 40.650
7
0 1000 2000 3000 4000 5000 6000 0 1 2 3 4 5 6 7 Hari ke-Nitrat (µM)Asetat Suksinat Glukosa Gliserol
(a) -500 0 500 1000 1500 2000 2500 3000 0 1 2 3 4 5 6 7 Hari ke-Nitrit (µM)
Asetat Suksinat Glukosa Gliserol
(a) -0.3 0.0 0.3 0.6 0.9 1.2 1.5 0 1 2 3 4 5 6 7 Hari ke- OD sel (550 nm)
Asetat Suksinat Glukosa Gliserol
(a) (b) 0 1000 2000 3000 4000 5000 6000 0 1 2 3 4 5 6 7 Hari ke-Nitrat (µM)
Asetat Suksinat Glukosa Gliserol (b)
0 500 1000 1500 2000 2500 0 1 2 3 4 5 6 7 Hari ke-Nitrit (µM)
Asetat Suksinat Glukosa Gliserol
(b) 0.00 0.03 0.06 0.09 0.12 0.15 0.18 0 1 2 3 4 5 6 7 Hari ke-OD (550 nm)
Asetat Suksinat Glukosa Gliserol
(b) .
Gambar 3 Profil penurunan senyawa nitrat oleh isolat ASLT2 pada kondisi aerob (a) dan anaerob (b) dengan sumber C yang berbeda.
Gambar 4 Profil akumulasi senyawa nitrit dalam medium selama proses reduksi nitrat oleh isolat ASLT2 pada kondisi aerob (a) dan anaerob (b).
Gambar 5 Pertumbuhan isolat ASLT2 pada kondisi aerob (a) dan anaerob (b) dengan sumber C yang berbeda.
Perbedaan aktivitas reduksi nitrat pada masing-masing perlakuan diduga berhubungan dengan jalur metabolisme sumber C yang merupakan senyawa organik yang sangat diperlukan bagi bakteri pereduksi nitrat (Kelso et al. 1999) baik pada kondisi aerob maupun anaerob. Richardson (2000) mengemukakan bahwa denitrifikasi merupakan proses reduksi nitrat yang berhubungan langsung dengan proses transfer elektron, dimana senyawa organik berperan sebagai donor elektron dan nitrat sebagai akseptor elektron terakhir.
Perbedaan kadar nitrat yang direduksi pada masing-masing perlakuan diduga dipengaruhi oleh banyaknya elektron yang dapat diperoleh dari oksidasi senyawa karbon
pan pada molekul NADH dan FADH2 yang
akan berperan sebagai donor elektron antara pada rantai respirasi (Madigan et al. 2000, White 2000). Richardson (2000) mengemukakan bahwa bakteri denitrifikasi dapat memanfaatkan nitrat, nitrit, nitrik oksida, atau nitrous oksida sebagai akseptor elektron terakhir pengganti oksigen dalam rantai respirasi.
Oksidasi molekul glukosa pada sistem metabolisme aerob akan berlangsung melalui lintasan glikolisis yang dilanjutkan dengan oksidasi piruvat menjadi CO2 melalui siklus TCA. Gliserol akan masuk ke dalam lintasan glikolisis melalui pembentukan gliseraldehida-3-fosfat sedangkan asetat akan masuk ke dalam siklus TCA setelah diubah
Nitrat yang direduksi Nitrit yang terbentuk Gas yang terbentuk Sumber C dalam medium Nitrat Awal (µM) µM % µM % µM % Asetat 5256.67 4325.08 82.28 527.97 12.21 3797.11 87.79 Suksinat 5282.13 651.13 12.33 0 0 651.13 100 Glukosa 5271.22 4304.79 81.67 1763.34 40.96 2541.43 59.04 Gliserol 5285.77 2331.69 44.11 52.9 2.27 2278.79 97.73
Nitrat yang direduksi Nitrit yang terbentuk Gas yang terbentuk Sumber C dalam medium Nitrat Awal (µM) µM % µM % µM % Asetat 4805.97 1728.22 35.96 811.3 46.95 916.91 53.06 Suksinat 5105.54 2449.74 47.98 1250.73 51.06 1199.01 48.94 Glukosa 5116.24 2082.123 40.7 1944.69 93.4 137.44 6.6 Gliserol 4805.97 1160.75 24.15 1004.49 86.54 156.27 13.46 Aerob Anaerob Sumber C dalam medium µN03 (µM/ hari) R-sq (µN03) µsel (sel/hari) R-sq (µsel) µN03 (µM/ hari) R-sq (µN03) µsel (sel/hari) R-sq (µsel) Asetat 0.4571 0.979 0.9769 0.951 0.3944 0.903 0.422 0.892 Suksinat 0.0527 0.535 0.5639 0.662 0.3657 0.737 0.4629 0.897 Glukosa 0.5111 0.905 1.0875 0.944 0.4392 0.902 0.6892 0.982 Gliserol 0.3059 0.736 0.6589 0.988 0.4364 0.845 0.3888 0.959
Tabel 4 Kemampuan isolat ASLT2 dalam mereduksi nitrat dan menghasilkan senyawa nitrit dan gas pada kondisi anaerob dengan sumber C yang berbeda
Tabel 5 Laju reduksi nitrat (µN03) dan laju pertumbuhan spesifik (µsel) isolat ASLT2 dengan sumber C yang berbeda pada kondisi aerob dan anaerob
Tabel 3 Kemampuan isolat ASLT2 dalam mereduksi nitrat dan menghasilkan senyawa nitrit dan gas pada kondisi aerob dengan sumber C yang berbeda
9
lintasan glikolisis dan siklus TCA akan menghasilkan ATP, NADH, dan FADH2 yang lebih banyak dibandingkan dari hasil oksidasi satu molekul asetat, suksinat, dan gliserol. Perolehan energi yang lebih banyak akan
mendukung untuk pertumbuhan sel
sedangkan perolehan molekul NADH dan
FADH2 yang lebih banyak sebagai donor
elektron yang akan masuk ke rantai respirasi memungkinkan aktivitas reduksi nitrat yang lebih tinggi.
P. stutzeri merupakan bakteri non fermentatif (Madigan et al. 2000). Dengan demikian, pada kondisi anaerob molekul
NADH dan FADH2 yang diperoleh dari
oksidasi sumber C direoksidasi melalui rantai respirasi. Dalam hal ini, isolat ASLT2 diduga menggunakan nitrat, nitrit, nitrik oksida, atau nitrous oksida sebagai akseptor elektron terakhir untuk memperoleh energi.
Pertumbuhan isolat ASLT2 (Gambar 5 (a)) dan aktivitas reduksi nitrat pada kondisi aerob yang sangat rendah dengan suksinat sebagai sumber C (Gambar 3 (a)) diduga
berhubungan dengan kesulitan untuk
mendapatkan elektron dari oksidasi senyawa
tersebut. Asam-asam organik termasuk
suksinat pada kondisi anaerob akan diubah menjadi piruvat dan diokidasi lebih lanjut setelah diubah menjadi asetil-CoA (Madigan et al. 2000). Dari metabolisme tersebut dihasilkan molekul NADH yang diduga oleh isolat ASLT2 direoksidasi melalui rantai respirasi dengan memanfaatkan nitrat, nitrit, nitrik oksida atau nitrous oksida sebagai akseptor elektron. Firth dan Edwards (2000) melaporkan bahwa P. stutzeri dapat tumbuh dan mereduksi nitrat dengan maksimal pada medium mikroaerofil yang mengandung nitrat dengan penambahan suksinat dibandingkan penambahan glukosa, gliserol, dan piruvat.
P. stutzeri merupakan bakteri
denitrifikasi yang mampu mereduksi nitrat dengan menghasilkan gas nitrogen (Korner & Zumft 1989, Zumft 1997, Rius et al. 2001).
Denitrifikasi merupakan proses reduksi
senyawa nitrat menjadi nitrit, nitrit menjadi nitrik oksida, nitrik oksida menjadi gas
nitrous oksida hingga pada akhirnya
dihasilkan gas nitrogen (Richardson 2000). Setiap tahapan reaksi dikatalisis oleh enzim yang berbeda. Reduksi nitrat menjadi nitrit dikatalisis oleh enzim nitrat reduktase. Ada dua macam nitrat reduktase yaitu nitrat reduktase terikat membran (NAR) dan nitrat reduktase pada periplasmik (NAP). Aktivitas enzim NAP terjadi pada kondisi aerob dan anaerob, sedangkan aktivitas enzim NAR
diduga hanya terjadi pada kondisi anaerob. Hal ini disebabkan adanya penghambatan sistem transfer nitrat ke dalam sel oleh oksigen (Moreno-Vivian et al. 1991, Zumft 1997). Richardson (2000) mengemukakan bahwa proses reduksi nitrat oleh enzim NA R berhubungan dengan konservasi energi yaitu sebagai akseptor elektron terakhir dalam rantai respirasi pada kondisi anaerob, sedangkan aktivitas enzim NAP cenderung untuk mengontrol keseimbangan energi pereduksi.
Bakteri heterotrofik memperoleh energi dari serangkaian proses oksidasi sumber C organik. Hasil oksidasi sumber C yang lebih tereduksi pada kondisi aerob sangat memungkinkan terjadinya kelebihan energi pereduksi. Oleh karena itu, untuk menjaga keseimbangan energi pereduksi diperlukan lintasan untuk mengalirkan elektron-elektron hasil oksidasi tersebut tetapi dengan tujuan bukan untuk konservasi energi yaitu melalui reduksi nitrat oleh enzim NAP (Richardson 2000).
Peranan enzim NAP dalam mengontrol keseimbangan energi pereduksi selama metabolisme oksidatif senyawa karbon telah banyak dilaporkan. Sears et al. (1997) dalam Ellington et al. (2002) menunjukkan bahwa jenis sumber C mempengaruhi aktivitas enzim NAP. Pada kondisi aerob, aktivitas enzim NAP Paracoccus denitrificans Pd22 lebih rendah dengan sumber C yang lebih teroksidasi (malat dan suksinat) dibandingkan dengan sumber C yang lebih tereduksi (kaproat dan butirat). Selain itu, Ellington et al. (2002) melaporkan bahwa ekspresi nap dan aktivitas enzim NAP pada Paracoccus pantotrophus lebih tinggi pada kondisi pertumbuhan aerob dengan sumber C yang lebih tereduksi dibandingkan dengan sumber C yang lebih teroksidasi (suksinat < asetat < butirat).
Reduksi nitrit menjadi nitrik oksida dikatalisis oleh enzim nitrit reduktase (NIR). Nitrik oksida diubah menjadi gas nitrous oksida dengan melibatkan enzim nitrik oksidoreduktase (NOR) sedangkan nitrous oksida diubah menjadi gas nitrogen dengan melibatkan enzim nitrous oksidoreduktase (NOS) (Zumft 1997, Richardson 2000). Akumulasi senyawa nitrit pada kultur bakteri pereduksi nitrat menurut Blaszczyk (1993) dapat disebabkan oleh beberapa faktor, antara lain adanya penghambatan nitrit reduktase oleh nitrat karena nitrat lebih kompetitif sebagai akseptor elektron dibandingkan nitrit, ketidakseimbangan reaksi reduksi nitrat atau
nitrit oleh masing-masing reduktase, dan induksi nitrit reduktase yang lebih lambat daripada nitrat reduktase.
SIMPULAN
Sumber C dapat mempengaruhi aktivitas oksidasi amonium dan reduksi nitrat. P. stutzeri ASLT2 dapat mengoksidasi amonium baik pada medium autotrof dengan karbonat sebagai sumber C maupun pada medium heterotrof dengan asetat, suksinat, glukosa, dan gliserol sebagai sumber C. Aktivitas oksidasi amonium isolat ASLT2 yang paling tinggi terjadi pada medium dengan suksinat sebagai sumber C.
Isolat ASLT2 dapat mereduksi nitrat pada medium dengan asetat, suksinat, glukosa dan gliserol sebagai sumber C baik pada kondisi aerob maupun anaerob. Aktivitas reduksi nitrat yang paling tinggi pada kondisi aerob terjadi dengan glukosa sebagai sumber C sedangkan pada kondisi anaerob aktivitas paling tinggi terjadi dengan suksinat sebagai sumber C.
SARAN
Diperlukan pengujian lebih lanjut dari isolat ASLT2 melalui deteksi gen-gen yang terlibat dalam proses oksidasi amonium dan reduksi nitrat untuk memperkuat hasil analisis
bahwa P. stutzeri ASLT2 memiliki
kemampuan dalam melakukan proses tersebut.
DAFTAR PUSTAKA
Arp DJ, Stein LY. 2003. Metabolism of inorganic N compound by ammonia-oxidizing bacteria. Critical rev in Biochemist and Mol Biol 38: 471-495.
Atlas RM, Bartha R. 1998. Microbial
Ecology: Fundamentals and Applications. 4th Edition. California: Benjamin/Cumming Sciences Publishing. Blaszczyk M. 1993. Effect of medium composition on the denitrification of nitrate by Paracoccus denitrificans. Appl Environ Microbiol 59: 3951-3953 Bock E, Koops HP, Ahlers B, Harms H. 1992.
Oxidation of Inorganic Compounds as Energy Sources. Di dalam Balows A, Truper GH, Dworkin M, Harder W,
Hand Book on the Biology of Bacteria. New York: Springer – Verlag.
Ellington MJK, Bhakoo KK, Sawers G, Richardson DJ, Ferquson SJ. 2002. Hierarchy of carbon sources selection in Paracoccus pantothrophus: strict correlation between reduction state of the carbon substrate and aerobic expression of the nap operon. J of Bacteriol 184: 4767-4774.
Firth JR, Edwards C. 2000. Analysis of denitrification by Pseudomonas stutzeri under nutrient-limited condition using membran inlet mass spectrometry. J of Appl Microbiol 88: 853-859.
Greenberg AE, Clesceri LS, Eaton AD. 1992. Standard Methods for Examination of Water and Wastewater. 18th Edition. Washington DC: Publication Office American Public Health Association. Kelso BHL, Smith RV, Laughlin RJ. 1999.
Effect of carbon substrates on nitrite accumulation in freshwater sediment. Appl Environ Microbiol 65: 61-66. Korner H, Zumft WG. 1989. Expression of
denitrification enzymes in response to the dissolved oxygen level and respiratory substrate in continuos culture of Pseudomonas stutzeri. Appl Environ Microbiol 55: 1670-1676.
Madigan MT, Martinko JM, Parker J. 2000. Biology of Microorganism. 9th edition. New Jersey: Prentice Hall.
Moreno-Vivian C, Cabello P, Martinez-Luque M, Blasco R, Castillo F. 1991. Procaryotic nitrate reduction; Molecular properties and functional distinction among bacterial nitrate reductase. J of Bacteriol 181: 6573-6484.
Prescott LM, Harley JP, Klein DA. 2000. Microbiology. 5th Edition. USA: McGraw-Hill Companie.
Richardson DJ. 2000. Bacterial respiration: a flexible process for a changing environment. Microbiology 146: 551-571.
Rius N, Fuste MC, Guasp C, Lalucat J, Loren JG. 2001. Clonal population structure of Pseudomonas stutzeri, a species with exc eptional diversity. J of Bacteriol 183: 736-744.
Rodina GA. 1972. Methods in Aquatic
Microbiology. Rita RC, Machael S, editor. USA: University Park Press Baltimore.
11
Teixe ira P, Oliveira R. 2002. Metabolism of Alcaligenes denitrificans in bio film vs planktonic cells. J Of Appl Microbiol 92: 256-260.
White D. 2000. The Physiology and
Biochemistry of Procaryotes. 2nd Edition. USA: Oxford University Press.
Widiyanto T. 2006. Seleksi Bakteri Nitrifikasi dan Denitrifikasi untuk Bioremediasi di
Tambak Udang [Disertasi]. Bogor:
Institut Pertanian Bogor.
Zumft WG. 1997. Cell biology and molecular basic of denitrification. Microbiol and Mol Biol Rev:533-616.
13
Lampiran 1 Profil senyawa ammonium pada medium kontrol pengujian aktivitas oksidasi amonium 0 1000 2000 3000 4000 5000 0 1 2 3 4 5 6 7 Hari ke-Amonium (µM)
Asetat Suksinat Glukosa Gliserol Karbonat
Lampiran 2 Profil senyawa nitrat pada medium kontrol pengujian aktivitas reduksi nitrat secara aerob (a) dan anaerob (b)
0 1000 2000 3000 4000 5000 6000 0 1 2 3 4 5 6 7 Hari ke- Nitrat (µM)
Asetat Suksinat Glukosa Gliserol
(a) 0 1000 2000 3000 4000 5000 6000 0 1 2 3 4 5 6 7 Hari ke-Nitrat (µM)
Asetat Suksinat Glukosa Gliserol