KINETIKA AKTIVITAS REDUKSI NITRAT BAKTERI
NITRAT AMONIFIKASI DISIMILATIF DARI MUARA
SUNGAI PADA KONSENTRASI OKSIGEN (O
2) YANG
BERBEDA
TETI MARDIATI
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
PERNYATAAN MENGENAI TESIS DAN SUMBER INFORMASI
Dengan ini saya menyatakan bahwa tesis Kinetika Aktivitas Reduksi Nitrat Bakteri Nitrat Amonifikasi Disimilatif dari Muara Sungai pada Konsentrasi Oksigen (O2) yang Berbeda adalah benar hasil karya saya sendiri dengan arahan komisi pembimbing dan belum diajukan dalam bentuk apapun kepada perguruan tinggi mana pun. Sumber informasi yang berasal atau dikutip dari karya yang diterbitkan maupun tidak diterbitkan dari penulisan lain telah disebutkan dalam teks dan dicantumkan dalam Daftar Pustaka di bagian akhir tesis ini
Bogor, Juni 2009
Teti Mardiati
ABSTRACT
TETI MARDIATI, Nitrate Reduction Activity Kinetics of Dissimilative Nitrate Ammonification Bacteria Isolated from Estuary under Different Concentration of Oxygen. Under direction of IMAN RUSMANA and TRI WIDIYANTO.
Dissimilative Nitrate ammonification bacteria have ability to remove nitrate in aquatic environments. These groups are consist of fermentative bacteria which reduce nitrate to ammonium with nitrous oxide (N2O) as by product. Four selected isolates from tropical estuary were tested the growth and nitrate reduction activities under different concentration of oxygen (O2). The results showed that Cell density was increased by increasing of oxygen concentration. In contrast, the activities of nitrate reduction was decreased by increasing of oxygen concentration. HF7 and LF6 isolates had the highest nitrate reducing activity. The maximum specific growth of HF7 isolate with asetate as carbon source, and 49.41 mM nitrate after 96 h incubation under air saturated of 1%, 10%, 100% was 0.034 CFUh-1, 0.040 CFUh-1, and 0.070 CFUh-1 respectively. While LF6 isolate was 0.035 CFU h-1, 0.045 CFU h-1, and 0.057 CFU h-1.
Nitrate removal rate of HF7 isolate with asetate as carbon source, and 49.41 mM nitrate after 96 h incubation under air saturated of 1%, 10%, 100% was 6.3 x 10-10 mMh-1, 5.1 x 10-10 mMh-1, and 3.3 x 10-10mMh-1 respectively. While LF6 isolate was 2.0 x 10-10 mMh-1, 2,1 x 10-10 mMh-1 and 1,0 x 10-10 mMh-1 respectively.
RINGKASAN
TETI MARDIATI. Kinetika Aktivitas Reduksi Nitrat Bakteri Nitrat Amonifikasi Disimilatif dari Muara Sungai pada Konsentrasi Oksigen yang Berbeda. Dibimbing oleh IMAN RUSMANA dan TRI WIDIYANTO.
Bakteri nitrat amonifikasi disimilatif atau Dissimilatory Nitrate Reduction to Ammonium (DNRA) termasuk kedalam kelompok bakteri pereduksi nitrat. Bakteri DNRA bersifat fermentatif, dapat mendegradasi senyawa organik dalam lingkungan oksigen terbatas atau anaerob. Penggunaan nitrat sebagai akseptor elektron oleh kelompok bakteri ini, dapat mengurangi kadar nitrat di perairan. Pada sisi lain bakteri DNRA menghasilkan produk samping N2O (nitrous oksida), gas yang berpotensi meningkatkan pemanasan global.
Meskipun bakteri DNRA umumnya mereduksi nitrat menjadi amonium pada kondisi anoksik, tetapi pada beberapa bakteri ditemukan reduksi nitrat berlangsung secara aerob. Sensitivitas sistem reduktase bakteri DNRA daerah tropik terhadap oksigen belum banyak diketahui. Bertolak dari hal tersebut perlu dilakukan penelitian pengaruh oksigen terhadap kinetika aktivitas reduksi nitrat bakteri DNRA yang berasal dari muara sungai di daerah tropik.
Empat isolat terseleksi dari muara sungai daerah tropik, telah diuji pertumbuhan dan aktivitas reduksi nitratnya pada konsentrasi oksigen yang berbeda. Keempat isolat tersebut adalah isolat HF7, LF6 (berasal dari muara Sungai Cisadane) dan isolat FR1, FR2 (berasal dari muara Sungai Cimandiri). Tingkat pencemaran muara Sungai Cisadane lebih tinggi dari pada muara Sungai Cimandiri.
Metoda penelitian dilakukan melalui 3 tahap, yaitu konfirmasi kemurnian isolat dan peremajaan, seleksi aktivitas konsentrasi oksigen berbeda, dan kinetika aktivitas reduksi nitrat isolat terpilih pada konsentrasi oksigen berbeda. Pengkondisian konsentrasi oksigen dilakukan dengan mengatur volume saturasi udara pada head space botol sampel dengan penggunaan gas nitrogen murni. Aktivitas reduksi nitrat dan senyawa-senyawa yang dihasilkan diukur dengan analisis nitrat metoda Greenberg, analisis nitrit metode Sulfanilamid dan analisis amonium metoda Phenate.
Hasil penelitian tahap seleksi menunjukkan kerapatan sel cenderung meningkat sejalan dengan penambahan konsentrasi oksigen. Aktivitas reduksi nitrat cenderung menurun sejalan dengan penambahan konsentrasi oksigen. Isolat HF7 dan LF6 memiliki kemampuan mereduksi nitrat paling tinggi, dengan perkiraan gas N2O yang dihasilkan lebih sedikit dibandingkan isolat FR1 dan FR2. Kedua isolat dijadikan isolat terpilih untuk diamati laju pertumbuhan dan kinetika aktivitas reduksi nitratnya.
Kecepatan rata-rata mengurangi nitrat medium oleh setiap sel isolat HF7 dengan sumber karbon asetat, nitrat medium 49,41 mM, masa inkubasi 96 jam pada konsentrasi udara saturasi 1%, 10%, 100% secara berurut adalah 6,3 x 10-10 mMJ-1, 5,1 x 10-10 mMJ-1, dan 3,3 x 10-10mMJ-1. Sedangkan rata-rata kecepatan reduksi setiap sel isolat LF6 pada kondisi yang sama secara berurut adalah 2,0 x 10-10 mMJ-1, 2,1 x 10-10 mMJ-1 dan 1,0 x 10-10 mMJ-1.
Dari hasil penelitian keseluruhan menunjukkan bahwa oksigen berpengaruh terhadap kinetika aktivitas reduksi nitrat bakteri DNRA yang berasal dari muara sungai daerah tropik. Besarnya pengaruh oksigen terhadap kinetika aktivitas reduksi nitrat tergantung pada sensitivitas sistem nitrat reduktase yang dimiliki oleh isolat bakteri. Konsentrasi oksigen berbeda lebih besar pengaruhnya pada isolat HF7 dibandingkan terhadap isolat LF6. Perkiraan konsentrasi gas N2O tertinggi yang dihasilkan pada isolat HF7 dengan nitrat medium 49,41 mM dan sumber karbon asetat terdapat pada konsentrasi saturasi udara 100%. Sedangkan pada isolat LF6 diperkirakan menghasilkan gas N2O terbesar pada konsentrasi saturasi udara 10%.
© Hak Cipta milik IPB, tahun 2009
Hak Cipta dilindungi Undang-Undang
Dilarang mengutip sebagian atau seluruh karya tulis ini tanpa mencantumkan
atau menyebutkan sumber. Pengutipan hanya untuk kepentingan pendidikan,
penelitian, penulisan karya ilmiah, penyusunan laporan, penulisan kritik atau
tinjauan suatu masalah; dan pengutipan tersebut tidak merugikan kepentingan
yang wajar IPB.
Dilarang mengumumkan dan memperbanyak sebagian atau seluruh karya tulis
KINETIKA AKTIVITAS REDUKSI NITRAT BAKTERI
NITRAT AMONIFIKASI DISIMILATIF DARI MUARA
SUNGAI PADA KONSENTRASI OKSIGEN (O2)
YANG BERBEDA
TETI MARDIATI
Tesis
Sebagai salah satu syarat untukmemperoleh gelar Magister Sains pada Mayor Mikrobiologi
SEKOLAH PASCASARJANA
INSTITUT PERTANIAN BOGOR
Judul Tesis : Kinetika Aktivitas Reduksi Nitrat Bakteri Nitrat Amonifikasi Disimilatif dari Muara Sungai pada Konsentrasi Oksigen (O2) yang Berbeda
Nama : Teti Mardiati
NRP : G351070201
Disetujui
Komisi Pembimbing
Dr. Ir. Iman Rusmana, M. Si. Dr. Tri Widiyanto, M. Si
Ketua Anggota
Diketahui
Ketua Mayor Mikrobiologi Dekan Sekolah Pascasarjana
Dr. Ir. Gayuh Rahayu Prof. Dr. Ir. Khairil Anwar Notodiputro, M.S.
PRAKATA
Puji dan syukur penulis panjatkan kepada Allah SWT atas limpahan rahmat, karunia dan ridho-Nyalah tesis yang berjudul ”Kinetika Aktivitas Reduksi Nitrat Bakteri Nitrat Amonifikasi Disimilatif dari Muara Sungai pada Konsentrasi Oksigen (O2) yang Berbeda” dapat diselesaikan.
Pelaksanaan penelitian dan penulisan tesis ini tidak terlepas dari bantuan dan bimbingan berbagai pihak. Pada kesempatan ini penulis menyampaikan terima
kasih yang sebesar-besarnya kepada:
1. Bapak Dr. Ir. Iman Rusmana, M.Si dan Bapak Dr. Tri Widiyanto, M.Si selaku komisi pembimbing atas bantuan, bimbingan dan pengarahan yang telah diberikan selama penelitian dan penulisan tesis ini sehingga penulis dapat menyelesaikan dengan baik.
2. Departemen Agama RI yang telah memberikan beasiswa pendidikan.
3. Ketua Program Studi Biologi, Ketua Mayor Mikrobiologi, Ketua dan Staf Laboratorium Mikrobiologi Departemen Biologi, Fakultas Ilmu Pengetahuan Alam, Institut Pertanian Bogor yang telah membantu dalam menyediakan fasilitas hingga terlaksananya penelitian ini.
4. Suami, Ibunda, dan anak-anak tercinta, seluruh keluarga serta rekan-rekan di MAN Batumandi yang telah banyak memberikan dukungan, hingga penulis mampu menyelesaikan studi ini dengan baik.
5. Rekan-rekan angkatan 2007 Program studi Biologi, rekan-rekan lab
mikrobiologi, Mbak Ratna, Irul dan Ria atas segala bantuan dan kerjasamanya.
Akhirnya semoga tesis ini bermanfaat bagi semuanya. Amin.
Bogor, juni 2009
RIWAYAT HIDUP
Penulis dilahirkan di Bandung pada tanggal 19 Agustus 1967 dari ayah Ahmad Hidayat dan ibu Siti Rokilah. Penulis merupakan anak pertama dari enam
bersaudara.
Penulis menyelesaikan program sarjana (SI) Jurusan Biologi Fakultas Pendidikan Matematika dan Ilmu Pengetahuan Alam IKIP Bandung pada tahun 1990. Penulis bertugas sebagai staf Pengajar Biologi pada Madrasah Aliyah
Negeri (MAN) Batumandi Kabupaten Agam Sumatera Barat sejak tahun 1992 sampai sekarang. Pada bulan Juli 2007 penulis melanjutkan kuliah ke Sekolah Pascasarjana Institut Pertanian Bogor Program Studi Biologi Mayor Mikrobiologi melalui beasiswa pendidikan Departemen Agama RI.
Bogor, juni 2009
DAFTAR ISI
Halaman.
DAFTAR TABEL ... xiii
DAFTAR GAMBAR ... xiv
DAFTAR LAMPIRAN... xv
PENDAHULUAN ... 1
Latar Belakang ... 1
Tujuan... 3
TINJAUAN PUSTAKA... 4
Muara sungai... 4
Muara Sungai Cimandiri dan Cisadane... 6
Bakteri Pereduksi Nitrat ... 7
Enzim-Enzim yang Berperan Dalam Reduksi Nitrat ... 8
Bakteri Nitrat Amonifikasi Disimilatif (DNRA) ... 9
METODE PENELITIAN... .... 13
Waktu dan Tempat penelitian ... 13
Bahan Penelitian ... 13
Konfirmasi dan Peremajaan Isolat. ... 13
Seleksi Aktivitas Reduksi Nitrat pada Konsentrasi Oksigen (O2) yang Berbeda ... 14
Analisis nitrit. ... 15
Analisis nitrat ... 15
Analisis amonium ... 16
Perhitungan Pembentukan Gas. ... 16
Kinetika Aktivitas Reduksi Nitrat Isolat Terpilih ... 17
HASIL ... 19
yang Berbeda ... 21
Aktivitas Reduksi Nitrat Bakteri DNRA pada Konsentrasi Oksigen yang Berbeda... 22
Pertumbuhan Isolat Terpilih ... 25
Kinetika Aktivitas Reduksi Nitrat ... 26
PEMBAHASAN... 32
SIMPULAN DAN SARAN ... 41
Simpulan... 41
Saran... 41
DAFTAR PUSTAKA ... 42
DAFTAR TABEL
Halaman
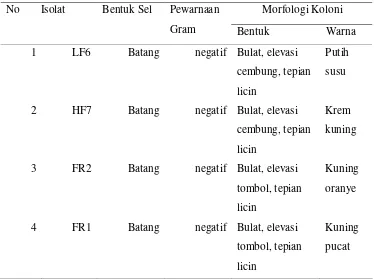
1. Karakterisasi morfologi dan fisiologi isolat LF6, HF7, FR2,dan FR1 ... 20
2. Aktivitas reduksi nitrat dan senyawa yang dihasilkan isolat HF7 pada konsentrasi saturasi udara 1%, 10% dan 100% setelah inkubasi 96
jam suhu ruang (29-31)oC ... 31
3. Aktivitas reduksi nitrat dan senyawa yang dihasilkan isolat LF6 pada konsentrasi saturasi udara 1%, 10% dan 100%, setelah inkubasi
DAFTAR GAMBAR
Halaman
1. Daur biogeokimia nitrogen di perairan...5
2. Lintasan reduksi nitrat oleh aktivitas bakteri (1) Denitrifikasi, (2).Reduksi nitrat amonifikasi disimilatif (3) Oksidasi amonia
secara anaerob....8
3. Bagan pembentukan N2O pada reduksi nitrat oleh bakteri DNRA...10
4. Enzim pereduksi nitrat bakteri nitrat amonifikasi disimilatif...11
5. Morfologi koloni dari isolat LF6, HF7, FR2 dan FR1 pada
medium denitrifikasi...19
6. Hasil uji fermentatif pada isolat FR1, FR2, HF7 dan LF6, setelah
inkubasi 48 jam...21
7. Densitas sel Isolat LF6, HF7, FR2, dan FR1 pada konsentrasi saturasi udara yang berbeda, setelah diinkubasi selama 96 jam
pada suhu ruang (29-31)0C....22
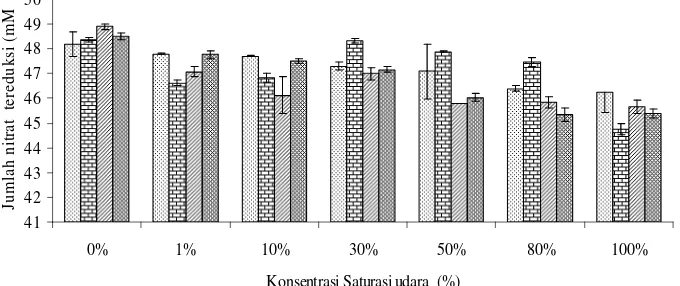
8. Jumlah nitrat tereduksi oleh isolat LF6, HF7, FR2, dan FR1 pada konsentrasi saturasi udara yang berbeda, setelah diinkubasi selama 96
jam pada suhu ruang ( 29-31)0C...22
9. Jumlah nitrit terakumulasi dari isolat LF6, HF7, FR2, dan FR1 pada konsentrasi saturasi udara yang berbeda, setelah diinkubasi
selama 96 jam pada suhu ruang (29-31)0C ...23
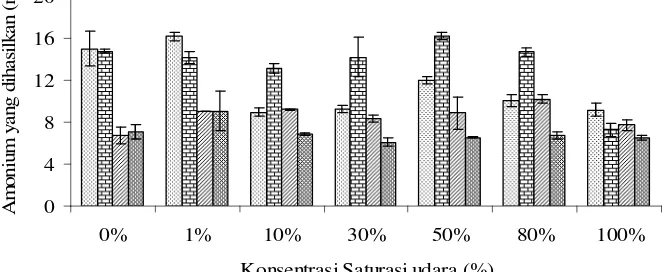
10.Jumlah amonium yang dihasilkan dari isolat LF6, HF7, FR2, dan FR1 pada konsentrasi saturasi udara yang berbeda, setelah diinkubasi
selama 96 jam pada suhu ruang ( 29-31)0C...24
11.Pola pertumbuhan isolat HF7 dan LF6 pada konsentrasi saturasi udara
Berbeda...25
12.Laju pertumbuhan maksimum isolat HF7 dan LF6 pada konsentrasi
saturasi udara berbeda...26
13. Aktivitas reduksi nitrat, pembentukan nitrit dan amonium dari isolat
HF7 dan LF6 pada konsentrasi saturasi udara 1%, 10% dan 100%....27
14. Perbandingan pertumbuhan sel isolat HF7 dan LF6 dengan penurunan
DAFTAR LAMPIRAN
Halaman
1. Pembentukan warna pada uji kadar nitrat (A), nitrit (B) dan
amonium (C)...47
2. Kurva standar konsentrasi amonium (A) dan kurva standar
konsentrasi nitrit (B) ....48
3. Jumlah CFU Isolat HF7 dan LF6 pada pengenceran 10-8...49
4. Kurva standar pertumbuhan isolat HF7 dan LF6 pada kondisi
nitrat medium 49,41 mM...50
5. Presentase nitrat tereduksi dan senyawa yang dihasilkan isolat LF6,HF7 dan FR2 pada kadar saturasi udara berbeda dengan
inkubasi selama 4 hari suhu ruang (29-31)oC...51
6. Presentase nitrat tereduksi dan senyawa yang dihasilkan isolat FR1 pada kadar saturasi udara berbeda dengan inkubasi selama
4 hari suhu ruang (29-31)oC...52
7. Perbandingan konsentrasi nitrat tereduksi, nitrit, nitrat, amonium
dan estimasi gas yang dihasilkan isolat LF6 dan HF7...53
8. Perbandingan konsentrasi nitrat tereduksi, nitrit, nitrat, amonium
PENDAHULUAN
Latar Belakang
Bakteri nitrat amonifikasi disimilatif atauDissimilatory Nitrate Reduction to
Ammonium (DNRA) termasuk ke dalam kelompok bakteri pereduksi nitrat. Kelompok bakteri pereduksi nitrat menggunakan nitrat sebagai akseptor elektron dalam respirasi anaerob (Purwoko 2007). Di alam bakteri pereduksi nitrat berperan dalam degradasi senyawa organik pada lingkungan oksigen terbatas atau anaerob. Bakteri pereduksi nitrat juga merupakan bagian dari mikroorganisma
yang terlibat dalam siklus biogeokimia nitrogen global. Kelompok bakteri ini dapat mengurangi kadar nitrogen tanah pertanian (kelompok denitrifikasi), bermanfaat dalam pengolahan air limbah, perbaikan kualitas perairan seperti tambak (Madiganet al. 2006; Maieret al. 2000; Widiyanto 2006).
Pada sisi lain kelompok bakteri ini dapat menghasilkan produk samping atau produk antara gas nitrous oksida (N2O). Gas N2O merupakan salah satu gas rumah kaca yang dapat meningkatkan pemanasan global seperti gas CO2dan CH4.
Penambahan gas N2O setiap tahunnya 0,2-0,3% (Conrad 1995) dengan kalor jenis N2O lebih besar dan waktu tinggal (residence time) di atmosfer lebih lama, maka jumlah kalor yang diserap oleh N2O tinggi. Gas N2O memiliki potensi penyerapan panas 150 kali lebih besar dari pada gas CO2. Pada stratosfer N2O dapat terfotolisis menjadi NO atau N2 yang menyebabkan terjadinya pengkisan
lapisan ozon (Rusmana 2003c). Proses Fotolisis N2O menjadi NO atau N2 dapat
mengubah senyawa ozon (O3) menjadi gas oksigen (O2). Hal tersebut
memperbesar radiasi cahaya matahari terhadap bumi.
N2O diperkirakan sebanyak 90% berasal dari proses biotik (Rusmana 2003c). Penambahan pupuk nitrogen, limbah senyawa nitrogen dari industri, dan
pembakaran bahan bakar fosil memperbesar emisi senyawa N2O. Terdapat
Proses pembentukan N2O pada bakteri DNRA bukan sebagai senyawa antara seperti yang dihasilkan oleh bakteri denitrifikasi, tetapi merupakan produk
samping melalui reaksi kimia dalam menghasilkan amonium (Rusmana 2003a). Kelompok bakteri ini memanfatkan senyawa nitrat sebagai penerima elektron alternatif untuk mendapatkan energi dibawah kondisi oksigen terbatas (Ricardson
et al. 2001). Purwoko (2007) menyatakan bahwa fermentasi sebenarnya merupakan proses yang tidak memerlukan oksigen tetapi organisma fermentatif terkadang memerlukan oksigen untuk proses metabolisma lainnya maupun
pertumbuhannya.
Walaupun nitrat amonifikasi disimilatif dan denitrifikasi termasuk proses anoksik tetapi reduksi nitrat dapat terjadi pada kondisi aerob. Jenis bakteri yang dapat melakukan reduksi nitrat pada kondisi aerob diantaranya Escherichia coli, Thiosphaera pantotropha, dan Pseudomonas aeroginosa (Carter et al. 1995).
Kemampuan bakteri tersebut mereduksi nitrat pada kondisi aerob dan anaerob berkaitan dengan enzim yang dimilikinya. Enzim nitrat reduktase pada membran sel (Nar) aktivitasnya dapat dipengaruhi oleh konsentrasi oksigen, dimana oksigen menghambat sistem transpor nitrat ke membran. Enzim Nap yang mereduksi nitrat dalam periplasma tidak sensitif terhadap oksigen. Ekspresi gen nar dipengaruhi
oleh ketersediaan oksigen dan nitrit sedangkan gennap tidak (Moreno - Vivian
et al. 1999).
Menurut Holt et al. (1994) sebagian besar bakteri fermentatif memiliki dua jalur metabolisma, yaitu respirasi dan fermentatif. Secara umum bakteri fermentatif pereduksi nitrat bersifat anaerob fakultatif, Desulfovibrio desulfuricans adalah salah satu bakteri yang dapat memanfaatkan nitrat atau sulfat sebagai akseptor elektron yang bersifat anaerob obligat. Keberadaan
oksigen berpengaruh pada aktivitas reduksi nitrat bakteri fermentatif. Terdapat hubungan antara ketersediaan oksigen dengan pembentukan amonium hasil aktivitas bakteri fermentatif. Hasil penelitian pada sedimen perairan estuari Colne di Inggris menunjukkan konsentrasi amonium bertambah dengan bertambahnya
kedalaman (Rusmana 2003a).
oksigen lebih besar dari 4,05 mg/l; 2,15 mg/l; 0,25 mg/l sepenuhnya dihambat. Demikian juga pada percobaan Kester et al. (1997) menunjukkan bahwa pada
kultur Alcaligenes eutropus, Nitrosomonas europia dan Pseudomonas stutzeri
dengan aerasi berbeda didapatkan jumlah produk yang berbeda. Pada udara saturasi 80 % produksi NO dan N2O adalah 0,87% dan 0,17%, sedangkan pada udara saturasi 1% berubah menjadi 2,32% dan 0,78%. Kemampuan mereduksi nitrat Pseudomonas stutzeri pada udara saturasi 90% setelah diinkubasi 92 jam, dengan sumber karbon suksinat adalah 99,24%. Sementara itu pada Thiosphaera pantotropha ATCC 35512 pada kondisi udara saturasi, suhu dan sumber karbon
yang sama adalah 27,29% (Suet.al 2000).
Berdasarkan beberapa penelitian tersebut yang banyak dilakukan pada kelompok bakteri denitrifikasi dan bakteri yang berasal dari daerah beriklim sedang, maka perlu dilakukan pengujian pengaruh konsentrasi oksigen terhadap
kinetika aktivitas bakteri DNRA muara sungai di daerah tropik. Pengaruh oksigen sebagai sesama akseptor elektron terhadap efektivitas reduksi nitrat menjadi amonium perlu diamati, termasuk prediksi senyawa N2O yang dihasilkan.
Hasil penelitian ini diharapkan dapat melengkapi pengetahuan fisiologi respirasi bakteri pereduksi nitrat. Langkah selanjutnya adalah kemungkinan
kelompok bakteri ini dapat dimanfaatkan dalam strategi penanggulangan masalah lingkungan seperti pengolahan limbah dan perairan tercemar, konservasi kesuburan tanah dan menjaga kualitas perairan seperti tambak, danau tanpa menimbulkan emisi Nitrous Oksida (N2O).
Tujuan
Penelitian ini bertujuan untuk mengetahui pengaruh konsentrasi oksigen
TINJAUAN PUSTAKA
Muara sungai
Muara sungai termasuk ke dalam ekosistem estuari, dimana air tawar dan air
laut bercampur. Bahan-bahan organik dan anorganik yang terdapat di muara sungai sebagian diendapkan, terlarut dan terbawa oleh arus ke laut. Salah satu proses yang mempengaruhi konsentrasi bahan-bahan organik dan anorganik pada muara sungai adalah proses biologi (Chester 1990).
Menurut Dahuri et al. (1996) kualitas air suatu perairan pesisir seperti
muara dicirikan oleh karakteristik kimianya yang mudah dipengaruhi oleh masukan air yang berasal dari daratan (melalui sungai) dan lautan sekitarnya. Proses pembuangan limbah dari daratan melalui aliran sungai atau dari lautan yang berlangsung secara terus menerus akan mengakibatkan turunnya kualitas air muara sungai. Pencemaran muara sungai oleh senyawa nitrogen lebih dominan berasal dari daratan.
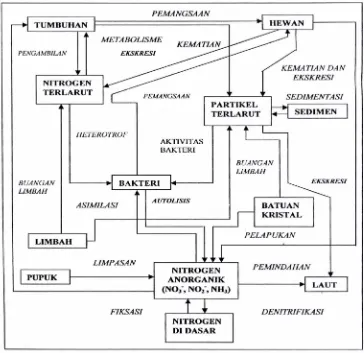
Pada muara sungai terjadi daur biogeokimia nitrogen, yaitu terjadinya
perubahan-perubahan senyawa atau unsur nitrogen dalam medium dan peristiwa yang berbeda. Beberapa proses transformasi senyawa nitrogen melibatkan berbagai jenis bakteri yaitu bakteri yang berperan dalam amonifikasi, asimilasi, fiksasi, nitrifikasi dan reduksi nitrat disimilatif.
Amonifikasi adalah proses pembentukan amonia dari materi organik
dilakukan oleh bakteri dan cendawan safrofit. Asimilasi adalah proses pemanfaatan senyawa nitrogen anorganik untuk pembentukan asam amino dalam protoplasma. Fiksasi nitrogen yaitu peristiwa pengikatan gas nitrogen (N2) umumnya terjadi di daratan, simbiosis alga di perairan dan percampuran nitrogen dari atmosfer. Nitrifikasi merupakan reaksi oksidasi pembentukan nitrat yang
bagian sedimen muara sungai adalah nitrat amonifikasi disimilatif (Rusmana 2003a).
Madiganet al. (2006) menyatakan proses penghilangan senyawa nitrogen di tanah dan perairan melalui proses denitrifikasi dan anaerob ammonia oxidation
(anammox). Kedua proses tersebut menghasilkan gas nitrogen dan nitrous oksida (denitrifikasi) sebagai produk akhir. Proses ini jika terjadi di areal pertanian dapat menurunkan produktivitas hasil pertanian.
Senyawa-senyawa nitrogen yang terdapat di muara sungai secara umum
terlibat dalam daur biogeokimia seperti yang dijelaskan oleh Kennish (1994) seperti terlihat pada Gambar 1.
Muara Sungai Cimandiri dan Cisadane
Muara Sungai Cimandiri kabupaten Sukabumi Jawa Barat, merupakan
muara sungai yang telah tercemar. Berbagai sampah rumah tangga / pemukiman, limbah areal pertanian (sawah, perkebunan) dan bahan-bahan lain sepanjang Daerah Aliran Sungai (DAS) Sungai Cimandiri terakumulasi di muara. Kualitas air pada muara sungai Cimandiri sudah mengalami penurunan (Marnis 2008).
Selain limbah yang masuk ke perairan sungai, aktivitas penambangan pasir sepanjang DAS menyebabkan warna perairan di sekitar muara menjadi coklat
sampai jarak yang cukup jauh dari pantai (Marnis 2008). Pada kondisi kimia dan fisik perairan seperti itu akan memicu pertumbuhan mikroorganisma-mikroorganisma aerob fakultatif dan anaerob disamping bakteri aerob, termasuk mikroorganisme yang memanfaatkan senyawa nitrogen baik organik (protein, asam amino) atau senyawa anorganik (nitrat, nitrit, dan amonium).
Muara Sungai Cisadane merupakan muara sungai yang tingkat pencemarannya lebih tinggi dari muara Sungai Cimandiri. Sungai Cisadane selain sebagai sumber air yang dimanfaatkan penduduk untuk keperluan sehari-hari, juga merupakan sumber perairan pertanian. Disamping itu berfungsi sebagai tempat penampungan limbah yang berasal dari limbah rumah tangga, industri,
pertanian dan perternakan (Brahmana & Suriati 2001).
Beberapa parameter kualitas air yang menunjukkan muara Sungai Cisadane tercemar adalah rendahnya kandungan oksigen terlarut, tingginya konsentrasi nitrit dan amonia serta suhu perairan yang tinggi. Kandungan oksigen terlarut berkisar antara 0,39-2,33 mg/l, dan suhu 32,0-32,60C. Kandungan senyawa nitrit 0,03-0,14 mg/l dan amonia 0,09-1,19 mg/l (Syahputra 2007). Tingkat pencemaran muara Sungai Cisadane pada musim hujan akan lebih rendah karena faktor
pengenceran oleh air hujan, termasuk air hujan dari hulu melalui aliran sungai. Muara sungai yang banyak mengandung limbah organik dan anorganik yang mengandung senyawa nitrogen diduga menghasilkan emisi Nitrous Oksida (N2O) yang lebih besar. Emisi N2O dari perairan dan muara kurang lebih 66%, produksi
Pada kedua muara sungai diatas senyawa nitrogen umumnya berasal dari limbah organik rumah tangga sepanjang daerah aliran sungai yang mengalami
dekomposisi, limbah pupuk nitrogen anorganik areal pertanian dan senyawa nitrogen anorganik tanah yang mengalami erosi tepi sungai. Sumber senyawa nitrogen lain berasal dari proses nitrifikasi, proses dekomposisi mikroorganisma dan eksresi organisma di habitat muara sungai serta gas oksida nitrogen dari udara yang terlarut melalui air hujan. Limbah industri merupakan faktor utama pencemaran yang meningkatkan senyawa nitrogen pada muara Sungai Cisadane
disamping faktor-faktor di atas.
Bakteri Pereduksi Nitrat
Kelompok bakteri pereduksi nitrat merupakan mikroorganisma yang terlibat dalam daur nitrogen di alam. Kelompok bakteri ini berperanan dalam pengubahan
senyawa nitrat menjadi produk akhir gas nitrogen atau senyawa amonium.
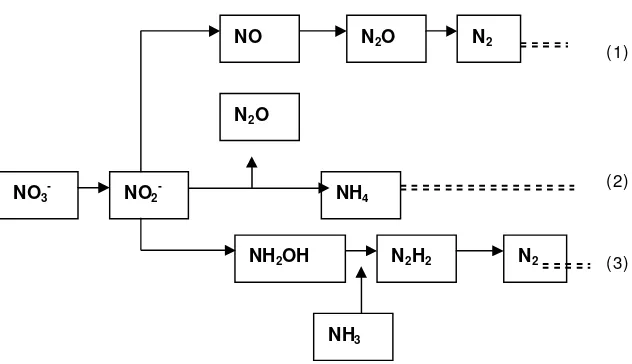
Rusmana (2003b) menyatakan bahwa terdapat tiga proses reduksi nitrat disimilatif pada bakteri yaitu denitrifikasi, reduksi nitrat menjadi amonium disimilatif dan oksidasi amonium disimilatif (anaerob ammonia oxidation,
anammox). Denitrifikasi adalah proses reduksi nitrat menjadi N2O atau N2. Pada
proses ini bakteri menggunakan nitrat sebagai penerima elektron terakhir untuk memperoleh energi pada kondisi O2 terbatas atau anaerob (Zumft 1997; Ricardson 2000; Ricardsonet al. 2001). Reduksi nitrat menjadi amonium disimilatif adalah proses untuk menghilangkan kelebihan tenaga pereduksi dan menunjang pertumbuhan bakteri pada kondisi anaerob (Cole 1996). Anamoks adalah oksidasi amonia secara anaerobik dimana terjadi pengubahan amonium dan nitrat atau nitrit menjadi gas nitrogen. Pada metabolisma ini membentuk senyawa antara
hidroksil amin dan hidrazin (Jetten et al. 2001).
(3)
Enzim-Enzim yang Berperan pada Reduksi Nitrat
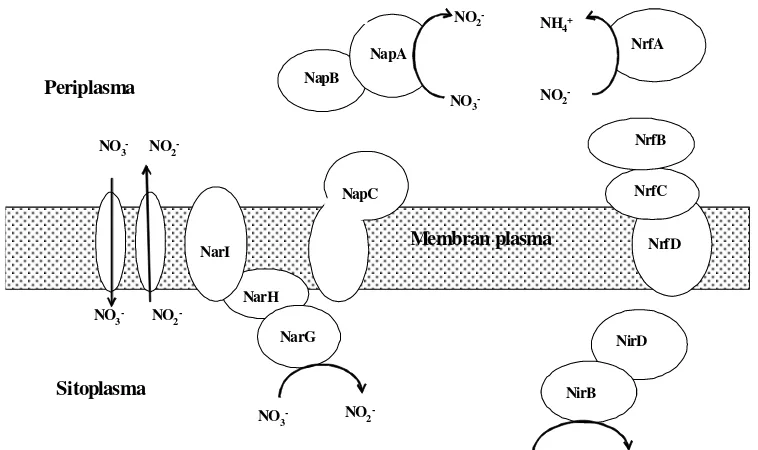
Kelompok bakteri pereduksi nitrat baik denitrifikasi, bakteri DNRA maupun anamoks memiliki enzim-enzim tertentu untuk mengkatalisasi reaksi-reaksi reduksi nitrat. Enzim pada bakteri denitrifikasi adalah nitrat reduktase (Nar dan Nap), Nitrit reduktase (Nir), Nitrit oksid reduktase (Nor), Nitrous oksid reduktase (Nos) (Moreno - Vivian et al.1999; Zumft 1997; Ricardsonet al. 2001).
Bakteri DNRA memiliki enzim nitrat reduktase (Nap dan Nar) yang mereduksi nitrat menjadi nitrit dan dua enzim yang mereduksi nitrat menjadi amonium yaitu Nir B (Harboneet al. 1992) danenzim formate dependent nitrite reduction to amonium atau Nrf (Cole 1996). Pada spesies tertentu seperti
Wollinella succinogens danCampylobacter fetus memiliki enzim nitrous oksida reduktase (Nos). Enzim ini mereduksi gas N2O menjadi gas Nitrogen (Zumft
1997).
Enzim yang terdapat pada bakteri anamoks adalah nitrit reducing enzyme
(NR) yaitu enzim yang mereduksi nitrit menjadi hidroksil amin (NH2OH). Hidrazin Hidrolase (HH) enzim yang mereduksi hidroksilamin menjadi hidrazin
(N2H4) dan Hydrazine – oxidising enzyme (HZO), enzim yang mengoksidasi hidrazin menjadi gas nitrogen (Jettenet al. 2001).
NH4
Gambar 2 Lintasan reduksi nitrat oleh aktivitas bakteri (1) Denitrifikasi,
Bakteri Nitrat Amonifikasi Disimilatif (DNRA)
Pada kondisi yang sesuai beberapa genus dapat mereduksi nitrit menjadi
amonia melalui proses reduksi nitrat disimilasif. Prosesnya berlangsung pada saat kandungan senyawa nitrit tinggi (Kelsoet al.1997). Kelompok bakteri ini bersifat fermentatif (Rusmana & Nedwell 2004), dapat memanfaatkan senyawa organik untuk pembentukan energi melalui transfer elektron di sitoplasma (Purwoko 2007).
Purwoko (2007) mengemukakan bahwa sebagian besar prokariota
fermentatif menghasilkan semua ATP melalui fosforilasi tingkat subtrat, kemudian ATP dihidrolisis oleh ATP sintetase, sehingga dapat menghasilkan perbedaan potensial yang digunakan untuk aktivitas membran. Organisma fermentatif memerlukan akseptor elektron untuk menerima elektron dari NADH. Kelompok bakteri pereduksi nitrat dapat memanfaatkan senyawa nitrat sebagai
penerima elektron alternatif untuk mendapatkan energi dibawah kondisi oksigen terbatas (Ricardsonet al. 2001).
Kelompok bakteri DNRA mereduksi senyawa nitrat sejalan dengan penambahan senyawa nitrit, ketika konsentrasi nitrat rendah terjadi penambahan senyawa amonium (Kelso et al. 1997). Pada beberapa pengamatan di lapangan
menunjukkan bahwa proses nitrat amonifikasi disimilatif banyak terjadi pada suhu tinggi (Herbert & Nedwell 1990). Bakteri DNRA lebih dominan pada lingkungan yang rasio C/N nya tinggi (Nedwell 1982). Bakteri DNRA lebih kompetitif dari pada bakteri denitrifikasi pada konsentrasi bahan organik tinggi.
Bakteri-bakteri nitrat amonifikasi disimilatif diantaranya adalah Bacillus pyocyanes, Clostridium pasteurianum, Desulfovibrio desulfuricans (Moat et al.
2002). Bacillus licheniformis, Wollinella succinogenes, Citrobacter freundii,
Klebsiella oxytoca, K. Pneumonia, Escherichia coli (Rusmana 2003a).
Rusmana (2003a) telah mengisolasi dan mengkarakterisasi Klebsiella pneumonia yang mereduksi nitrat menjadi amonium dan menghasilkan N2O sebagai produk sampingnya. Syahputra (2007) telah mengisolasi dan menyeleksi
bakteri nitrat amonifikasi disimilatif dari muara sungai Cisadane. Isolat terpilih
tingkat kemiripan 97%. Sebagian besar famili Enterobacteriaceae mereduksi nitrat dan bersifat fermentatif (Holt et al. 1994).
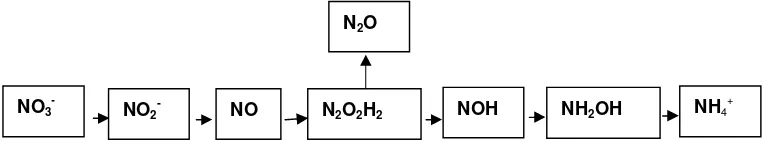
Proses pembentukan N2O pada kelompok bakteri nitrat amonifikasi disimilatif bukan sebagai senyawa antara seperti pada kelompok bakteri denitrifikasi, tetapi merupakan produk samping melalui reaksi kimia dalam menghasilkan amonium (Rusmana 2003a). Bagan proses reaksi kimia reduksi nitrat ke amonium dapat ditunjukkan pada Gambar 3.
Pada kelompok bakteri DNRA terjadi dua tahap reaksi enzimatik yaitu tahap pertama oleh enzim Nap dan Nar seperti pada bakteri dinitrifikasi. Patereu et al.
(1994) menyatakan bahwa ada dua tipe enzim yang mereduksi nitrat menjadi nitrit yaitu nitrat reduktase yang terdapat pada membran (Nar) dan nitrat reduktase yang
terdapat pada periplasmik (Nap). Enzim Nar aktivitasnya berhubungan langsung dengan proses respirasi pembentukan ATP yang dapat dipengaruhi oleh konsentrasi oksigen dimana oksigen menghambat sistem transpor nitrat. Enzim Nap yang mereduksi nitrat dalam periplasma tidak sensitif terhadap oksigen. Enzim Nar yang terdapat di membran plasma mempunyai transmembran proton motive force (PMF) tempat terjadinya sintesis ATP (Moreno- Vivianet.al 1999).
Demikian pula menurut Carteret al. (1995) reduksi nitrat dalam periplasma
tidak sensitif terhadap hambatan oksigen. Oleh karena itu, nitrat reduktase periplasmik berhubungan dengan kemampuan untuk respirasi nitrat dalam ketersediaan oksigen pada spesies laboratorium. Oksidasi quinol oleh nitrat reduktase periplasmik tidak terpusat untuk menghasilkan gradien proton
elektrokimia, tetapi cukup bertindak sebagai katup untuk membantu keseimbangan regulasi reaksi redoks dan mempertahankan jalannya rantai respirasi aerobik.
Sitoplasma
Enzim Nap merupakan komplek dari dua sub unit protein. Sub unit Nap A mengikat kofaktor Molibdenum (Mo) sebagai sisi aktif dari enzim. Sub unit Nap
B mempunyai dua situs ikatan heme C yang memiliki perbedaan potensial. Nap C merupakan sitokrom tipe tetraheme C, berperan dalam transfer elektron antara Quinol dan Nap AB (Richardson 2000).
Reaksi enzimatik tahap kedua yaitu perubahan nitrit menjadi amonium oleh enzim Nir B yang aktivitasnya bergantung pada NADH sitoplasma sebagai donor elektron (Harbone et al. 1992) dan enzim Nrf yang aktivitasnya bergantung pada
format sebagai donor elektron (Cole 1996). Dalam Richardson (2000) kinerja enzim pereduksi nitrat menjadi amonium disimilatif digambarkan sebagai berikut (Gambar 4).
Ketersediaan oksigen di lingkungan pada bakteri nitrat amonifikasi disimilatif diantaranya berkaitan erat dengan sistem sensor regulator sebagai mikroorganisme anaerob fakultatif. Kelompok bakteri DNRA memiliki kemampuan adaptasi dari aerob ke anaerob atau sebaliknya. Oksigen merepresi
kondisi jalur asam sitrat dari siklus TCA oksidatif menjadi non siklus/ reduktif (White 1995).
Pada White (1995) dijelaskan bahwa pergantian respirasi oksidatif dan reduktif Escherichia coli sebagai salah satu bakteri DNRA diatur oleh tiga sistem, yaitu sistem Arc (Aerobic respiration control), sistem Nar (Nitrat anaerobic respiration) dan sistem Fnr. Sistem Arc merupakan sistem dua komponen. Sistem ini merepresi gen-gen aerob berupa enzim-enzim siklus asam sitrat, piruvat dehidrogenase, asam lemak oksidase, dan sitokrom oksidase. Selain
itu sistem Arc menginduksi gen-gen mikroaerob (sitokrom d oksidase dan sintesis kobal alamin).
Sistem Nar terdiri dari 3 protein yaitu Nar X, Nar L dan Nar Q. Sistem ini juga merupakan sistem dua komponen, Nar X berperan sebagai histidin kinase, sedangkan Nar L sebagai protein regulator respons. Sistem ini menginduksi
sintesis transkripsi gen nitrat reduktase dan menghambat gen fumarat reduktase serta gen aerobik lainnya. Penelitian selanjutnya diketahui bahwa Nar Q juga berperan sebagai penerima sensor (White 1995 ; Purwoko 2007).
Sistem Fnr merupakan sistem tunggal yang berperanan pada kondisi anaerob. Protein Fnr berperan sebagai regulator positif untuk transkripsi gen-gen
dalam pertumbuhan anaerob dan regulator negatif untuk transkripsi gen-gen pertumbuhan aerob (White 1995 ; Purwoko 2007).
METODE PENELITIAN
Waktu dan Tempat penelitianPenelitian ini dilaksanakan pada bulan Oktober 2008 sampai Maret 2009. Tempat penelitian di Laboratorium Mikrobiologi Departemen Biologi Fakultas Matematika dan Ilmu Pengetahuan Alam, Institut Pertanian Bogor.
Bahan Penelitian
dan muara Sungai Cisadane Tangerang Banten. Kode isolat dari muara Sungai Cimandiri adalah FR1 dan FR2 (Marnis 2008). Kode isolat dari muara Sungai
Cisadane HF7 dan LF6 (Syahputra 2007).
Konfirmasi dan Peremajaan Isolat.
Peremajaan isolat murni dimulai dengan konfirmasi kemurnian isolat menggunakan media padat, dilanjutkan dengan peremajaan isolat murni pada media cair. Komposisi media cair adalah: 10 g Natrium asetat, 4,2 g NaNO3,
3,0 g yeast ekstrak, 0,5 g (NH4)2SO4,0,9 g K2HPO4.3 H2O, 0,5 g MgSO4.7H2O, 0,2 g KH2PO4, 0,1 g CaCl2.2H2O dengan salinitas 2% dan pH 7 (Rodina 1972). Media padat dibuat sama dengan media cair ditambah agar bacto 20 g per liter. Media disterilisasi pada suhu 1210C tekanan udara 1 atmosfer selama 15 menit.
Konfirmasi kemurnian isolat dilakukan dengan cara diinokulasikan
masing-masing isolat murni bakteri nitrat amonifikasi disimilatif dengan metoda cawan gores pada medium agar, kemudian diinkubasikan selama 7 hari. Karakterisasi dan morfologi sel tiap isolat dilakukan dengan teknik pewarnaan gram (Hadioetomo 1993).
Konfirmasi kemurnian isolat secara fisiologi dilakukan dengan uji
fermentatif. Keempat isolat ditumbuhkan pada medium glukosa dengan indikator
brom cressol purple. Isolat diletakkan pada tabung bertutup untuk membatasi masukan oksigen. Uji ini dilakukan untuk melihat kemampuan fermentasi dari tiap isolat. Perubahan warna ungu menjadi kuning menunjukkan isolat tersebut melakukan fermentasi, jika tidak terjadi perubahan warna bukan kelompok bakteri fermentatif. Peremajaan bakteri pada medium cair dilakukan dengan cara diinokulasikan 1 lup tiap isolat pada medium cair masing-masing sebanyak 20
Seleksi Aktivitas Reduksi Nitrat pada Konsentrasi Oksigen (O2) yang Berbeda
Pengukuran aktivitas reduksi nitrat pada konsentrasi oksigen berbeda dilakukan pada botol serum steril 75 ml yang ditutup dengan butyl rubber
septum. Pengkondisian konsentrasi oksigen dengan pengaturan volume saturasi udara padahead space botol serum.
Botol serum steril volume 75 ml diisi dengan 23 ml medium cair. Botol dibuat kondisi an aerobik dengan cara menghilangkan oksigen yang terdapat di dalam botol. Gas nitrogen bebas oksigen dialirkan ke dalam botol reaksi dengan menggunakan jarum/syringe steril selama 15 menit. Sebelum gas dimasukkan,
gas terlebih dahulu disaring dengan filter steril diameter 47 mm dengan ukuran 0,45 mikro meter.
Langkah berikutnya, saturasi udara pada masing-masing botol dikondisikan menjadi 0%, 1%, 10%, 30%, 50%, 80%, dan 100%. Saturasi udara dimasukkan
pada masing-masing botol serum yang telah diberi perlakuan pembebasan oksigen sebanyak 0,5 ml, 5 ml, 15 ml, 25 ml, dan 40 ml. Pada waktu proses pemasukan saturasi udara, gas nitrogen akan terdorong keluar melalui jarum yang dipasang pada tutup botol disampingsyringe, sehingga volume saturasi udara pada masing-masing botol menjadi 1%, 10%, 30%, 50%, 80%. Pengkondisian saturasi udara 100% dilakukan dengan tanpa memasukkan gas nitrogen murni pada botol serum.
Kondisi saturasi udara 0% dengan pengisian gas nitrogen murni tanpa memasukkan saturasi udara.
Seluruh botol serum yang telah dikondisikan saturasi udaranya dikocok untuk melarutkan gas perlakuan. Masing-masing botol serum diinokulasi dengan 1 ml isolat. Isolat diletakan pada inkubator berpenggoyang pada kecepatan 80 rpm suhu ruang (28-31)oC selama 96 jam (Suet al. 2001).
Setelah diinkubasi pada kadar oksigen yang berbeda, dilakukan pengukuran densitas sel dengan spektrofotometer pada panjang gelombang 620 nm. Kultur biakan bakteri diambil sebanyak 1ml pada tabung efendorf dan disentrifugasi dengan kecepatan 4000 rpm selama 15 menit untuk mendapatkan supernatan.
Analisis nitrit.
Analisis Nitrit menggunakan metode Sulfanilamide (Cleseri et al 1989).
Nitrit dengan amina aromatik pada sulfalinamide akan membentuk diazonium. Senyawa tersebut dengan NED (N-I-Naphtyl Ethylene Diamine Dyhidrochloride) membentuk gugus kromofor yang berwarna merah muda (Lampiran 1) dan dapat diukur dengan spektrofotometer pada panjang gelombang 540 nm. Kadar nitrit ditentukan dengan cara memasukkan 5 ml sampel yang telah diencerkan kedalam tabung reaksi, ditambahkan 0,1 sulfanilamid (C6H8N2O2S) dan 0,1 NED.
Kemudian diukur absorbansinya dengan spektrofotometer pada panjang gelombang 540 nm. Kurva standar natrium nitrit (NaNO2) dengan konsentrasi sebesar : 0 ; 0,2; 0,4; 0,6; 0,8 , dan 1,0 mg/l kemudian dikonversi ke mM digunakan untuk menentukan kadar nitrit pada sampel (Lampiran 2).
Analisis nitrat
Analisis senyawa nitrat dengan menggunakan metode Greenberg et al
(1992). Kadar nitrat ditentukan dengan cara memasukkan 20 ml sampel yang telah diencerkan ke dalam kolom reduksi yang berisi cadmium. Pada saat sampel melewati kolom reduksi, cadmium akan mereduksi nitrat menjadi nitrit. Sampel sebanyak 5 ml direaksikan dengan reagen pada analisis nitrit. Sampel akan menunjukkan perubahan warna merah muda (Lampiran 1), diukur pada panjang
gelombang 540 nm. Kurva standar natrium nitrit (NaNO2) dengan konsentrasi sebesar : 0 ; 0,2; 0,4; 0,6; 0,8; dan 1,0 mg/l, kemudian dikonversi ke mM digunakan untuk menentukan kadar nitrit pada sampel. Nilai konsentrasi nitrat adalah hasil pengurangan antara konsentrasi nitrit yang dilewatkan pada kolom reduksi dengan konsentrasi nitrit yang tidak dilewatkan.
Analisis amonium
Pengukuran konsentrasi amonium menggunakan metode phenate (Cleseriet al. 1989). Kadar amonium ditentukan dengan cara 5 ml sampel yang diencerkan ,
hipoklorit (NaOCl2) dan dibiarkan selama satu jam pada suhu ruang (28-31)o C. Sampel yang telah diberi larutan indikator menunjukkan perubahan warna hijau
sampai biru (Lampiran 1), diukur dengan spektrofotometer pada panjang gelombang 640 nm. Kurva standar amonium klorida (NH4Cl) sebesar: 0 ; 0,2; 0,4; 0,6; 0,8 ; 1,0; 2,0; 4,0; dan 8,0 mg/l kemudian dikonversi ke mM digunakan untuk menghitung konsentrasi amonia pada sampel (Lampiran 2).
Perhitungan Pembentukan Gas.
Setelah konsentrasi nitrat (NO3-), nitrit (NO2-), dan amonium (NH4+) diketahui dari masing-masing sampel, dihitung jumlah nitrat yang tereduksi dengan rumus :
[NO3-]tereduksi = [NO3-]kontrol – [NO3-]inokulasi
% [NO3-] tereduksi = [NO3-]red x 100 [NO3-]kontrol
Jumlah senyawa nitrit yang terbentuk dihitung dengan rumus sebagai berikut:
[NO2-]terbentuk = [NO2-]perlakuan – [NO2-] kontrol
%[NO2-]terbentuk = [NO2-]perlakuan – [NO2-] kontrol x 100 [NO3-]tereduksi
Jumlah senyawa amonium yang terbentuk dihitung dengan rumus sebagai berikut:
[NH4+]terbentuk = [NH4+]perlakuan – [NH4+] kontrol
% [NH4+]terbentuk = [NH4+]perlakuan – [NH4+] kontrol x 100 [NO3-]tereduksi
Estimasi produk akhir gas yang terbentuk dihitung dengan rumus [Gas ] = [NO3-]reduksi – [NO2-]terbentuk– [NH4+]terbentuk
%[ Gas] = [Gas] X 100% [NO3-]tereduksi
Kinetika Aktivitas Reduksi Nitrat Isolat Terpilih
Dua isolat terbaik dalam mereduksi nitrat dan responsif terhadap oksigen
dipilih untuk diamati kinetikanya. Botol serum steril volume 125 ml diisi dengan 50 ml medium cair. Sebanyak 1 ml isolat diinokulasikan pada tiap botol yang telah dikondisikan saturasi udaranya secara berurutan masing-masing 1 %, 10%, 100%. Kultur biakan diinkubasikan pada suhu ruang (28-31)oC selama 96 jam (Su et al 2001). Kultur biakan diamati pola pertumbuhannya dan dianalisis konsentrasi nitrat, nitrat, dan amonium dengan metoda yang telah diuraikan
sebelumnya. Analisis dilakukan setiap selang waktu 12 jam selama 96 jam (4 hari).
Hasil pertumbuhan digambarkan pada suatu grafik pertumbuhan pada kondisi oksigen berbeda. Korelasi antara konsentrasi sel dengan kerapatan optis (OD) isolat, dilakukan metoda hitungan cawan (Lampiran 3) pada konsentrasi
pengenceran 10-6, 10-7, 10-8, dan 10-9. Kultur biakan yang sama diencerkan dan OD dari beberapa pengenceran diukur. Kurva standar pertumbuhan dibuat dari nilai-nilai OD pengenceran dan konsentrasi sel sehingga didapatkan nilai regresi dan persamaan garis (Lampiran 4) untuk menentukan jumlah sel. Laju pertumbuhan spesifik maksimum didapatkan dengan mencari nilai regresi dan
persamaan garis dari korelasi waktu inkubasi dengan nilai ln jumlah sel. Laju pertumbuhan maksimum merupakan koefisien variabel x pada persamaan garis Y= ax + c (Maieret al. 2000).
Hasil uji kimia digambarkan dalam suatu grafik yang menunjukkan hubungan lama inkubasi dengan nitrat tereduksi, nitrit, dan amonium serta estimasi gas N2O yang terbentuk pada setiap konsentrasi oksigen yang berbeda. Kecepatan reduksi setiap sel masing-masing isolat dihitung pada kondisi saturasi
HASIL
Karakteristik, Morfologi dan Fisiologi Bakteri Nitrat Amonifikasi Disimilatif
Hasil konfirmasi kemurnian dari keempat isolat dengan metoda cawan gores, morfologi koloninya berbentuk bulat, elevasi tombol dan cembung, tepian
licin. Faktor pembeda keempat isolat adalah warna koloni (Gambar 5).
Secara mikroskopis sel isolat LF6, HF7, FR2 dan FR1 berupa bakteri Gram
negatif dan berbentuk batang (Tabel 1).
Tabel 1 Karakterisasi morfologi dan fisiologi isolat LF6, HF7,FR2, dan FR1
Morfologi Koloni
No sIIsolat Bentuk Sel Pewarnaan
Gram Bentuk Warna
1 LF6 Batang negatif Bulat, elevasi
cembung, tepian licin
Putih susu
2 HF7 Batang negatif Bulat, elevasi
cembung, tepian licin
Krem
kuning
3 FR2 Batang negatif Bulat, elevasi
tombol, tepian licin
Kuning oranye
4 FR1 Batang negatif Bulat, elevasi
tombol, tepian licin
Kuning pucat
Hasil uji fermentatif dengan menggunakan glukosa dan indikator brom cressol purple menunjukkan keempat isolat merupakan kelompok bakteri fermentatif (Gambar 6). Hal ini ditunjukkan dengan adanya perubahan warna
medium dari ungu menjadi kuning. Kecepatan untuk merubah warna medium dari keempat isolat berbeda. Isolat LF6 dan HF7 (asal muara Sungai Cisadane) merubah warna medium glukosa dan indikator brom cressol purple dari ungu menjadi berwarna kuning dalam waktu 24 jam. Perubahan warna medium oleh isolat FR1 dan FR2 (asal muara Sungai Cimandiri) memerlukan waktu 36 jam dan
48 jam.
Densitas Sel Isolat Bakteri DNRA pada Konsentrasi O2 yang Berbeda
Empat isolat bakteri DNRA yang ditumbuhkan pada konsentrasi oksigen
berbeda, sumber karbon asetat, masa inkubasi 96 jam menunjukkan kerapatan (densitas) sel yang berbeda (Gambar 7). Secara umum densitas sel keempat isolat lebih rendah pada konsentrasi oksigen (saturasi udara) 0%, yaitu antara OD 0,06 sampai 0,28 (620 nm). Densitas sel tertinggi pada konsentrasi saturasi udara 0 % adalah isolat FR1 yaitu dengan OD sebesar 0,28.
Pada konsentrasi saturasi udara 1-10% densitas sel cenderung lebih tinggi dari konsentrasi saturasi udara 0%. Pada konsentrasi saturasi udara 30%, 50% dan 80% kecenderungan densitas keempat isolat tidak seluruhnya lebih tinggi dari konsentrasi saturasi udara sebelumnya. Densitas tertinggi dari keempat isolat terdapat pada konsentrasi saturasi udara 100% yaitu dengan OD antara 0,27 sampai 0,42 (620 nm).
Kecenderungan peningkatan densitas sel isolat FR1 sejalan dengan
penambahan konsentrasi saturasi udara lebih nyata dari isolat lainnya. Densitas sel isolat FR1 lebih tinggi dari pada tiga isolat lainnya pada saturasi udara 0%, 30%, 50%, 80% dan 100%. Densitas sel terendah pada konsentrasi saturasi udara 100 % adalah isolat LF6 dengan OD 0,27.
Aktivitas Reduksi Nitrat Bakteri DNRA pada Konsentrasi Oksigen yang Berbeda
Berdasarkan hasil uji nitrat (NO3-) terhadap isolat FR1, FR2, HF7, dan LF6 menunjukkan kemampuan yang berbeda dalam mereduksi nitrat. Konsentrasi nitrat tereduksi oleh keempat isolat dengan nitrat awal sebanyak 49,41 mM, sumber karbon asetat setelah diinkubasi 4 hari adalah diatas 45 mM (Gambar 8).
Gambar 7 Densitas sel isolat LF6, HF7, FR2, dan FR1 pada konsentrasi saturasi udara yang berbeda, setelah diinkubasi selama 96 jam pada suhu ruang (29-31)0C. LF6 HF7 FR2 FR1
Gambar 8 Jumlah nitrat tereduksi oleh isolat LF6, HF7, FR2, dan FR1 pada konsentrasi saturasi udara yang berbeda, setelah diinkubasi selama 96 jam pada suhu ruang (29-31)0C
LF6 HF7 FR2 FR1
Konsentrasi Saturasi udara (%)
O
Konsentrasi Saturasi udara (%)
Secara umum kecenderungan konsentrasi nitrat tereduksi lebih besar pada konsentrasi saturasi udara lebih kecil. Reduksi nitrat terbesar terjadi pada
konsentrasi saturasi udara 0%. Konsentrasi nitrat tereduksi cenderung menurun seiring dengan peningkatan konsentrasi saturasi udara.
Dari hasil uji nitrit (NO2-), isolat FR1 mengakumulasi nitrit paling tinggi dibandingkan isolat lainnya. Akumulasi nitrit tertinggi terjadi pada konsentrasi saturasi udara 100% (Gambar 9), yaitu 4,95 mM . Isolat LF6, HF7 dan FR2 pada akhir masa inkubasi masih mengakumulasi nitrit. Konsentrasi nitrit tertinggi pada
isolat LF6 adalah 0,05 mM. Isolat HF7 dan FR2 mengakumulasi nitrit paling tinggi pada akhir masa inkubasi dengan konsentrasi yang sama yaitu 0,03 mM (Lampiran 5 dan 6).
Gambar 9 Jumlah nitrit terakumulasi dari isolat LF6, HF7, FR2, dan FR1 pada konsentrasi saturasi udara yang berbeda, setelah diinkubasi selama 96 jam pada suhu ruang (29-31)0C
LF6 HF7 FR2 FR1
Pada uji amonium (NH4+), isolat LF6 dan HF7 menghasilkan amonium lebih tinggi dari isolat lainnya (Gambar 10), sehingga diperkirakan gas yang terbentuk pada kedua isolat bakteri tersebut lebih kecil dibandingkan isolat FR1
dan FR2. Konsentrasi amonium tertinggi yang dihasilkan isolat LF6 adalah sebesar 15,02 mM (30,95%) pada konsentrasi saturasi udara 0%. Isolat HF7 menghasilkan amonium sebesar 14,77 mM (30,29%) pada konsentrasi saturasi udara 0% (Lampiran 5).
0.00
Gambar 10 Jumlah amonium yang dihasilkan dari isolat LF6, HF7, FR2, dan FR1 pada konsentrasi saturasi udara yang berbeda, setelah diinkubasi selama 96 jam pada suhu ruang (29-31)0C.
LF6 HF7 FR2 FR1
Perkiraan pembentukan gas N2O dari keempat isolat menunjukkan bahwa
keempat isolat tersebut dapat menghasilkan gas yang besar, pada medium dengan konsentrasi nitrat yang tinggi. Isolat yang paling sedikit menghasilkan gas N2O adalah isolat HF7 sebesar 29,62 mM (64,63%) pada konsentrasi saturasi udara 50%, dan isolat LF6 sebesar 31,61 mM (66,15 %) pada konsentrasi saturasi udara 10% (Lampiran 5).
Perbandingan konsentrasi nitrat tereduksi, nitrit terakumulasi, amonium dan
estimasi gas N2O yang dihasilkan dari setiap isolat pada kondisi saturasi udara berbeda setelah diinkubasi 96 jam pada suhu ruang (29-31)0C dapat dilihat di Lampiran 7 dan Lampiran 8.
Berdasarkan hasil uji fermentatif, analisis nitrat, nitrit, amonium dan estimasi gas N2O pada keempat isolat dipilih isolat HF7 dan LF6 untuk diamati kinetika aktivitas reduksi nitratnya. Dasar pemilihan kedua isolat adalah pada uji
fermentatif kedua isolat lebih cepat memfermentasi glukosa. Aktivitas reduksi nitrat isolat HF7 dan LF6 lebih tinggi dibandingkan isolat FR1 dan FR2. Kedua isolat menghasilkan amonium lebih banyak dengan akumulasi nitrit yang kecil. Isolat HF7 dan LF6 diduga menghasilkan gas N2O lebih kecil dari isolat FR1dan
reduksi nitrat pada konsentrasi saturasi udara yang relatif kecil (1 % dan 10%) dibandingkan dengan konsentrasi saturasi udara besar (100%).
Pertumbuhan Isolat Terpilih
Dua isolat terpilih HF7 dan LF6 yang ditumbuhkan pada konsentrasi saturasi udara 1%, 10%, dan 100%, sumber karbon asetat, suhu ruang (29-31)oC,
dengan konsentrasi nitrat 49,41mM menunjukkan pertumbuhan yang berbeda. Kerapatan sel pada awal pertumbuhan dikondisikan sama yaitu pada OD 0,03 (620 nm) dengan inokulan berumur 24 jam. Perubahan populasi sel atau pertambahan jumlah sel dari jumlah inokulan sebelumnya pada isolat HF7
dengan saturasi udara 100% lebih cepat dibandingkan dengan isolat LF6 (Gambar 11).
Kecepatan pertumbuhan kedua isolat pada konsentrasi saturasi udara 100% paling tinggi. Kecepatan pertumbuhan pada konsentrasi saturasi udara 10 % pada
kedua isolat lebih tinggi dari konsentrasi saturasi udara 1%. Fase eksponensial dimulai dari 12 jam pertama sampai jam ke 48 pada isolat HF7 saturasi udara 100%. Pada isolat HF7 kondisi saturasi udara 10% dan isolat LF6 pada kondisi saturasi udara 10% dan 100% fase eksponensial mulai jam ke 12 sampai jam ke 60. Sementara itu pada konsentrasi saturasi udara 1 % kedua isolat menunjukkan fase eksponensial sampai jam ke 72 (gambar 11).
Laju pertumbuhan maksimum kedua isolat pada konsentrasi saturasi udara 1%, 10% dan 100% dengan sumber karbon asetat dan nitrat media 49,41mM
dapat dilihat pada Gambar 12.
Laju pertumbuhan maksimum isolat HF7 dan LF6 pada konsentrasi saturasi udara 1% hampir sama. Pada konsentrasi saturasi udara 10% laju pertumbuhan
maksimum isolat HF7 lebih kecil dari isolat LF6. Pada saat konsentrasi saturasi udara 100% laju pertumbuhan maksimum isolat HF7 lebih besar dari isolat LF6.
Kinetika Aktivitas Reduksi Nitrat
Laju aktivitas reduksi nitrat pada isolat HF7 dan LF6 menunjukkan kemampuan reduksi nitrat terkecil pada konsentrasi saturasi udara 100%. Pada kondisi saturasi udara 1% jumlah nitrat tereduksi lebih besar (Gambar 13). Jumlah nitrat tereduksi pada kedua isolat bertambah seiring dengan pertambahan densitas sel. Reduksi nitrat tercepat pada umumnya terjadi dari inkubasi jam ke
12 sampai jam ke 60, dengan isolat yang diinokulasikan pada umur 24 jam. Umumnya pada fase stasioner reduksi nitrat masih berlangsung lambat, meskipun densitas sel tidak mengalami pertambahan yang signifikan.
0.034 0.035 0.
Gambar 13 Aktivitas reduksi nitrat, pembentukan nitrit dan amonium dari isolat HF7 dan LF6 pada konsentrasi saturasi udara 1%, 10% dan 100%.
nitrat tereduksi nitrit terbentuk amonium terbentuk
LF6 Saturasi udara 100%
Kemampuan mereduksi nitrat isolat HF7 konsentrasi saturasi udara 100% berbeda dengan kondisi lainnya. Isolat tersebut setelah jam ke 60 masih
mengalami kenaikan reduksi nitrat yang signifikan sampai jam ke 84(Gambar 13). Hubungan antara kecepatan reduksi nitrat dengan tingkat pertumbuhan sel pada konsentrasi saturasi udara berbeda jelas terlihat pada isolat HF7 dan LF6 (Gambar14). Pada konsentrasi saturasi udara kecil (1%), gradien penurunan konsentrasi nitrat medium oleh kedua isolat paling besar, diikuti konsentrasi saturasi udara 10%. Pada konsentrasi saturasi udara 100% penurunan konsentrasi
nitrat lebih kecil dari 10%. Penurunan konsentrasi nitrat sejalan dengan kenaikan jumlah sel. Penurunan konsentrasi nitrat yang signifikan terjadi pada saat pertumbuhan sel-sel isolat fase eksponensial.
Setelah inkubasi 96 jam kemampuan menurunkan nitrat media dari isolat HF7 pada kondisi saturasi udara 1%, 10%, dan 100% adalah masing-masing
sebesar 93,2%, 80,74% dan 77,81% (Tabel 2). Umumnya kecepatan mereduksi nitrat tinggi dari awal jam ke 12 sampai jam ke 60. Isolat HF7 lebih menunjukkan kemampuan reduksi nitrat yang berbeda pada kondisi saturasi udara berbeda dari pada isolat LF6. Isolat LF6 setelah diinkubasi dalam waktu yang sama mampu menurunkan nitrat media pada kondisi saturasi udara 1%, 10%, dan 100% adalah
Gambar 14 Perbandingan pertumbuhan sel isolat HF7 dan LF6 dengan penurunan konsentrasi nitrat pada konsentrasi saturasi udara 1%, 10% dan 100%. Konsentrasi nitrat OD sel
HF7 Saturasi udara 1%
0
HF7 Saturasi udara 10%
0
LF6 Saturasi udara 10%
0
HF7 Saturasi udara 100%
0
LF6 Saturasi udara 100 %
Kecepatan rata-rata mereduksi nitrat media oleh koloni isolat HF7 adalah 0,48 mMJ-1, 0,41 mMJ-1, 0,40 mMJ-1 pada saturasi udara 1%, 10% , 100% setelah
diinkubasi 96 jam. Selisih kecepatan reduksi nitrat isolat HF7 pada tiap konsentrasi saturasi udara lebih besar. Sementara pada isolat LF6 selisih kecepatan reduksi nitrat pada setiap konsentrasi saturasi udara berbeda tidak menunjukkan perbedaan yang nyata. Kecepatan reduksi isolat LF6 pada saturasi udara 1%, 10% , 100% setelah diinkubasi 96 jam yaitu masing-masing sebesar 0,41 mMJ-1, 0,41 mMJ-1, 0,40 mMJ- .
Perbandingan kecepatan rata-rata reduksi nitrat untuk setiap sel isolat HF7 dan LF6 hampir seluruhnya berbanding lurus dengan perbandingan rata-rata kecepatan reduksi koloni isolatnya. Kecepatan rata-rata mereduksi nitrat pada saturasi udara 1%, 10% , 100% setelah diinkubasi 96 jam untuk setiap sel isolat HF7 secara berurut adalah 6,3 x 10-10 mMJ-1, 5,1 x 10-10 mMJ-1, dan 3,3 x 10-10
mMJ-1. Sementara untuk setiap sel isolat LF6 dengan kondisi yang sama secara berurut adalah 2,0 x 10-10 mMJ-1, 2,1 x 10-10 mMJ-1 dan 1,0 x 10-10 mMJ-1. Kecepatan rata-rata reduksi nitrat pada setiap sel isolat HF7 dan isolat LF6 pada konsentrasi 1% lebih tinggi. Perbedaan kecepatan reduksi nitrat pada konsentrasi saturasi udara kecil (1%) dengan konsentrasi saturasi udara lebih
besar (100%) jelas terlihat. Selisih kecepatan setiap sel isolat HF7 adalah 3 x 10-10 mMJ-1. Sementara selisih kecepatan reduksi nitrat untuk setiap sel isolat LF6 pada konsentrasi saturasi udara 1% dengan 100% adalah 1 x 10-10 mMJ-1.
Jumlah senyawa-senyawa yang dihasilkan dari aktivitas reduksi nitrat bervariasi pada kedua isolat. Isolat HF 7 mengakumulasi nitrit pada jam ke 12 sampai jam ke 24 dan konsentrasinya menurun seiring peningkatan konsentrasi amonium. Setelah inkubasi 96 jam tidak terdapat akumulasi senyawa nitrit.
Konsentrasi senyawa nitrit sudah menurun pada inkubasi jam ke 12 (Gambar 13) Pada isolat LF6 terjadi fluktuasi akumulasi senyawa nitrit pada inkubasi jam ke12 sampai dengan jam ke 60. Setelah inkubasi 96 jam pada isolat LF6 terdapat akumulasi senyawa nitrit dalam jumlah kecil yaitu 0,32% (Tabel 3).
senyawa nitrit menurun. Konsentrasi nitrit sudah menurun pada inkubasi jam ke 12 (Gambar 13).
Amonium yang dihasilkan oleh isolat HF7 dan LF6 setelah inkubasi selama 96 jam dengan sumber karbon asetat pada konsentrasi saturasi udara 1%, 10%, 100% secara berurutan adalah 56,06%, 36,07%, 35,93% dan 41,66%, 36,75%, 38,19%. Perkiraan gas N2O yang dihasilkan HF7 dan LF6 secara berurutan adalah 53,20%, 63,93%, 64,06% dan 58,34%, 63,25%, 61,49% (Tabel 2 dan 3).
Tabel 2 Aktivitas reduksi nitrat dan senyawa yang dihasilkan isolat HF7 pada konsentrasi saturasi udara 1%, 10% dan 100% setelah inkubasi 96 jam, suhu ruang (29-31)oC
Nitrat
10% 49,41 39,90 80,74 0,00 0,00 14,39 36,07 25,51 63,93
100% 49,41 38,45 77,81 0,00 0,00 13,82 35,93 24,63 64,06
Tabel 3 Aktivitas reduksi nitrat dan senyawa yang dihasilkan isolat LF6 pada konsentrasi saturasi udara 1%, 10% dan 100%, setelah inkubasi 96 jam, suhu ruang (29-31)oC
Nitrat
10% 49,41 39,36 79,65 0,00 0,00 14,46 36,75 24,90 63,25
PEMBAHASAN
Isolat FR1, FR2, HF7 dan LF6 bersifat fermentatif, tumbuh pada medium
denitrifikasi yang mengandung nitrat dan tumbuh pada kondisi saturasi udara dari variasi 0-100%. Hal ini menunjukkan keempat isolat memiliki enzim-enzim respirasi oksidatif dan nitrat reduktase untuk proses respirasinya. Pada tahap uji seleksi menunjukkan kerapatan sel (densitas) isolat HF7 dan LF6 lebih rendah dibandingkan dengan isolat FR1 dan FR2.
Pertumbuhan FR1 paling responsif terhadap oksigen, peningkatan densitas sel sesuai dengan peningkatan kadar saturasi udara cenderung lebih tinggi dari isolat lainnya. Isolat FR1 diduga memiliki enzim nitrat reduktase periplasma (Nap) lebih dominan. Aktivitas enzim-enzim respirasi oksidatif lebih dominan dari pada enzim nitrat reduktase, karena enzim Nap berfungsi sebagai penyeimbang regulasi redoks pada transfer elektron aerobik, energi yang
dihasilkan dari aktivitas enzim Nap kecil (Ricardsonet al. 2001). Proses tersebut menyebabkan pertumbuhan FR1 meningkat seiring peningkatan konsentrasi oksigen. Hal ini sesuai dengan awal isolasi isolat FR1. Pada saat diisolasi isolat FR1 tumbuh baik pada medium rendah nitrat yaitu 0,1 mM (Marnis 2008).
Densitas sel keempat isolat pada kondisi oksigen yang berbeda, secara umum menunjukkan makin bertambah konsentrasi oksigen densitas sel makin
tinggi. Pada saturasi udara 30-80 % terdapat isolat-isolat yang densitas selnya fluktuatif, kondisi ini diduga karena sensitivitas sistem nitrat reduktase terhadap oksigen berbeda dari setiap isolat. Keempat isolat lebih cenderung memanfaatkan oksigen sebagai akseptor elektron pada saat oksigen tersedia pada medium. Disisi lain keberadaan akseptor elektron nitrat dapat menimbulkan interaksi dan
berpengaruh terhadap jalur metabolisme oksidatif. Nitrat dapat dimanfaatkan sebagai akseptor elektron alternatif pada saat oksigen di lingkungan terbatas. Selain itu reduksi nitrat oleh enzim Nap dapat berlangsung sebagai pembuangan kelebihan tenaga pereduksi pada transfer elektron aerobik (Carteret al.1995).
Pada kondisi saturasi udara 100% keempat isolat memiliki densitas sel tinggi. Enzim-enzim respirasi oksidatif lebih mendominasi, transfor nitrat ke
aktivitas enzim Nap, dimana aktivitas enzim Nap tidak dipengaruhi keberadaan oksigen (Moreno -Vivian et al. 1999). Sementara enzim Nar baru bekerja
setelah konsentrasi oksigen terbatas dalam medium perlakuan.
Oksigen merupakan akseptor elektron yang lebih potensial negatif. Sebagai perbandingan perbedaan potensial redoks dengan donor elektron asam format, akseptor elektron oksigen hasil akhir H2O adalah -1,28 (Mairer 2000). Sedangkan perbedaan potensial redoks dengan donor elektron sama akseptor elektron NO3 -hasil akhir NH4
+
memiliki nilai -0,83. Energi yang dihasilkan dari respirasi
dengan oksigen sebagai akseptor elektron lebih tinggi dari pada akseptor elektron nitrat. Energi tinggi memacu pertumbuhan sel lebih cepat, sehingga densitas sel akhir masa inkubasi tinggi.
Pada respirasi aerob bakteri DNRA, sumber karbon asetat akan diubah menjadi asetil Co A dan masuk ke siklustricarboxylic acid(TCA) sehingga akan
menghasilkan ATP, NADH dan FADH2. NADH dan FADH2 diubah menjadi
ATP melalui rangkaian transfer elektron (posforilasi oksidatif) dengan akseptor elektron terakhir oksigen (White 1995).
Pada kondisi oksigen terbatas di lingkungan, sumber karbon asetat tetap melalui jalur TCA, tetapi pada saat transfer elektron di membran digunakan
nitrat atau nitrit sebagai akseptor elektron terakhir. Akibat dari penggunaan nitrat atau nitrit sebagai akseptor elektron, nitrat tereduksi menjadi nitrit, nitrit tereduksi menjadi amonium. ATP yang dihasilkan dari respirasi anaerob lebih sedikit, menyebabkan pertumbuhan bakteri lebih lambat.
Pada sumber karbon yang lebih kompleks seperti glukosa (C6H12O6) suksinat (C4H6O4), sitrat (C6H8O7) dan gliserol (C3H8O3) perolehan energi melalui proses fermentatif diduga tetap kurang efektif. Keberadaan asam
organik atau alkohol hasil posforilasi subtrat akan mengganggu kecepatan pertumbuhan sel. Hal ini dapat diamati di alam secara langsung, pada perairan-perairan yang kadar oksigen terlarutnya kecil proses degradasi senyawa organik berlangsung lambat, meskipun tersedia akseptor elektron alternatif selain oksigen.
Pengaruh oksigen terhadap pertumbuhan isolat lebih jelas terlihat pada grafik pertumbuhan isolat terpilih HF7 dan LF6 yang diukur setiap selang waktu
12 jam. Dua isolat terpilih HF7 dan LF6 yang ditumbuhkan pada konsentrasi saturasi udara 1%, 10% dan 100%, sumber karbon asetat menunjukkan pertumbuhan yang berbeda. Pertumbuhan sel pada konsentrasi oksigen saturasi udara rendah (1% dan 10%) dari kedua isolat lebih lambat dari pertumbuhan pada konsentrasi saturasi udara 100%. Kerapatan sel pada awal pertumbuhan dikondisikan sama yaitu pada OD 0,03 (620 nm) dengan inokulan berumur 24
jam. Perubahan populasi atau pertambahan jumlah sel dari inokulan sebelumnya berbeda pada setiap perlakuan konsentrasi oksigen. Secara kuantitatif perbedaan laju pertumbuhan ditunjukkan oleh µ maks kedua isolat yang berbeda pada kondisi saturasi udara berbeda. Hal ini menunjukkan oksigen berpengaruh terhadap laju pertumbuhan bakteri DNRA.
Isolat HF7 diduga memiliki enzim Nar yang lebih dominan. Pada saat oksigen rendah kandungan nitrat medium tinggi aktivitas enzim Nar tinggi. Sistem respirasi nitrat dominan sehingga pada konsentrasi saturasi udara 1% dan 10% laju pertumbuhannya rendah, karena energi yang dihasilkan dari respirasi nitrat lebih rendah. Sebaliknya pada saat kandungan oksigen tinggi (saturasi
udara 100%) laju pertumbuhan sel HF7 lebih tinggi karena pada tahap awal inkubasi digunakan oksigen sebagai akseptor elektron, energi yang dihasilkan lebih tinggi. Pada saat oksigen pada medium terbatas, enzim Nar menggantikan proses pembentukan ATP meskipun tidak sebanyak respirasi aerob. Enzim Nar mempunyai transmembranproton motive force (PMF) tempat terjadinya sintesis ATP (Moreno- Vivianet.al 1999). Proses pergantian sistem respirasi oksidatif ke respirasi anaerob reduktif pada isolat HF7 diduga diatur oleh sistem sensor
regulator seperti pada Escherichia coli(Purwoko 2007).
Kekurangan oksigen di lingkungan diterima dan direspons oleh sistem Arc, sistem Nar dan sistem Fnr. Sistem Arc menerima stimulus kekurangan oksigen dan merespons dengan cara merepresi gen-gen yang mengekspresikan
Isolat HF7 juga diduga memiliki enzim Nap karena pada 12 jam pertama inkubasi pada konsentrasi udara 100% nitrat medium sudah tereduksi.
Sementara pada isolat LF6 diduga sistem respirasi lebih didominasi sistem respirasi oksidatif. Reduksi nitrat isolat LF6 diduga lebih didominasi enzim Nap. Pada saat konsentrasi oksigen dalam medium habis aktivitas enzim Nap dan enzim Nar yang tidak dominan menghasilkan energi kecil, tidak sebesar isolat HF7. Kurangnya ATP yang terbentuk setelah respirasi oksidatif tidak berjalan menyebabkan energi untuk pertumbuhan rendah. Pada konsentrasi
saturasi udara 10% µ maks isolat LF6 lebih tinggi dari pada isolat LF7, diduga respirasi oksidatif pada isolat LF6 dominan dengan memanfaatkan oksigen 10%.
Dari hasil uji nitrat (NO3-) tahap seleksi, keempat isolat dapat mereduksi nitrat pada kisaran konsentrasi oksigen 0% sampai 100%. Keempat isolat dapat menggunakan akseptor elektron nitrat. Penggunaan nitrat sebagai akseptor
elektron dibatasi keberadaan oksigen. Hal ini dapat terlihat dari kecenderungan reduksi nitrat berbanding terbalik dengan penambahan konsentrasi oksigen. Kultur biakan isolat dengan konsentrasi saturasi udara 100% menghasilkan reduksi nitrat lebih sedikit.
Oksigen dapat menghambat reduksi nitrat dengan 3 cara yaitu menekan
ekspresi gen yang mensintesis sistem nitrat reduktase, mengganggu enzim nitrat reduktase yang telah terbentuk, dan mengalihkan aliran elektron dari sistem respirasi nitrat ke sistem respirasi oksigen (Hernandez dan Rowe 1988). Sensitivitas sistem nitrat reduktase setiap jenis bakteri terhadap oksigen berbeda. Hasil penelitian Su et al. (2001), bakteri Pseudomonas stutzeri SU2 dapat mereduksi nitrat 99,24% pada konsentrasi oksigen tinggi (92% oksigen). Sementara itu padaThiosphaera pantotropha ATCC 35512 pada kondisi oksigen
dan suhu yang sama hanya dapat mereduksi nitrat 27-29%.
Menurut Otte et al. (1996) implikasi penghambatan oksigen tidak selalu terjadi, oleh karenanya reduksi nitrat dapat berlangsung secara simultan dibawah kondisi aerobik contohnyaAlcaligenes faecalis. Reduksi nitrat pada konsentrasi
udara 100% diduga lebih dominan dikatalisasi enzim nitrat reduktase
diperiplasma, yang merupakan ekspresi gen Nap. Gen penyandi enzim Nap
karena terletak di periplasma tidak dipengaruhi transfor nitrat ke membran plasma. Nitrat diduga dapat masuk melalui membran luar (pada bakteri gram
negatif) melalui porin dengan perbedaan konsentrasi atau pertukaran muatan ion. Perbedaan hasil uji nitrat (NO3-) pada konsentrasi oksigen rendah dan konsentrasi oksigen tinggi lebih jelas terlihat pada pengamatan kinetika aktivitas reduksi nitrat dua isolat terpilih. Pada saturasi udara kecil 1% dan 10% reduksi nitrat dikatalisasi oleh kedua enzim (Nap dan Nar). Pada kondisi ini transfor nitrat ke membran dapat berlangsung karena konsentrasi oksigen relatif kecil.
Aktivitas reduksi nitrat pada kondisi saturasi udara 1 dan 10% menjadi lebih besar, karena terjadi di periplasma dan di sitoplasma. Menurut Korner dan Zumft (1989) pada saturasi udara 17 %, keberadaan nitrat menyebabkan ekspresi optimal dari enzim nitrat reduktase.
Reduksi nitrat tercepat umumnya mulai jam ke 12 sampai jam ke 60 dengan
inokulan yang sudah dalam fase eksponensial (24 jam). Penurunan konsentrasi nitrat yang tereduksi lebih terlihat berbanding lurus dengan peningkatan jumlah sel pada konsentrasi saturasi udara rendah (1% saturasi udara), dari pada konsentrasi oksigen tinggi (100% saturasi udara). Hal ini dikarenakan pada konsentrasi oksigen rendah nitrat digunakan sebagai satu-satunya akseptor
elektron untuk menghasilkan energi. Pada kondisi oksigen di lingkungan terbatas bakteri fermentatif mengubah jalur respirasi oksidatif ke jalur reduktif (Purwoko 2007).
Enzim Nar pada membran plasma berperanan penting pada kondisi anaerob sebagai penghasil ATP. Sedangkan pada saat konsentrasi oksigen tinggi reduksi nitrat banyak terjadi pada periplasma oleh enzim Nap yang berperan sebagai pengatur keseimbangan redoks pada posforilasi oksidatif membran. Pada
konsentrasi oksigen tinggi (100%) aktivitas enzim Nar terhambat. Hal ini jelas terlihat pada isolat HF7 konsentrasi saturasi udara 100% setelah jam ke 60 masih mengalami kenaikan reduksi nitrat. Peristiwa ini diperkirakan bahwa aktivitas enzim Nar meningkat dengan habisnya oksigen pada medium.
reduksi nitrat setiap sel pada isolat LF6 terdapat perbedaan dari kecenderungan umum. Pada saturasi udara 10 % kecepatan reduksi rata-rata setiap sel isolat LF6
lebih besar dari konsentrasi saturasi udara 1%. Sementara kecepatan rata-rata reduksi nitrat pada koloninya, mengikuti kecenderungan umum yaitu reduksi nitrat makin kecil seiring dengan peningkatan konsentrasi oksigen. Hal ini dimungkinkan karena pengaruh faktor pembulatan perhitungan jumlah sel dengan penggunaan kurva standar yang regresinya tidak tepat 100%.
Kecepatan rata-rata reduksi nitrat baik koloni maupun setiap sel pada isolat
LF6 tidak terlalu berbeda pada konsentrasi oksigen yang berbeda. Hal demikian diduga enzim Nap isolat LF6 lebih dominan. Enzim Nap tetap mereduksi nitrat pada saat konsentrasi nitrat rendah atau tinggi. Isolat LF6 dapat tumbuh pada konsentrasi nitrat rendah seperti pada isolasi awal. Isolat LF6 diisolasi dari sedimen muara sungai Cisadane dengan media pengkayaan nitrat 0,1 mM
(Syahputra 2008).
Hasil uji nitrit (NO2-) pada tahap seleksi menunjukkan isolat FR1 mengakumulasi nitrit paling tinggi setelah masa inkubasi 96 jam. Isolat FR1 memiliki gen nitrit reduktase terbatas, sehingga pengubahan nitrit menjadi amonium lambat. Isolat FR1 cenderung memanfaatkan lebih banyak nitrat
sebagai akseptor elektron untuk menimbulkan energi dari pada menggunakan nitrit. Hal ini sesuai dengan perhitungan termodinamika perubahan nitrat menjadi nitrit menghasilkan energi lebih besar dari pada perubahan nitrit menjadi amonium (Rusmana 2003a). Persamaan reaksinya adalah sebagai berikut:
NO3- + H2 NO2- + H2O ( G = -163,03 KJ per mol H2) NO2- + 3H2 +2H+ NH4+ + 2 H2O ( G = -145,95 KJ per mol H2)
Akumulasi nitrit tertinggi pada FR1 terjadi pada konsentrasi udara 100%
diduga selain faktor genetik (dominan enzim Nap) aktivitas reduksi nitrit juga dipengaruhi faktor lingkungan. Aktivitas Nir B dan Nrf terbatas karena tidak tersedianya NADH dan format dalam jumlah banyak sehingga nitrit terakumulasi. Sebab aktivitas Nir B dipengaruhi NADH (Harboneet al. 1992), sedangkan Nrf
dipengaruhi format sebagai akseptor elektron (Cole 1996).